
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Plant Sci., 02 August 2012
Sec. Plant Development and EvoDevo
Volume 3 - 2012 | https://doi.org/10.3389/fpls.2012.00167
This article is part of the Research TopicSelaginellaView all 7 articles
 Judith Lucia Gomez-Porras1†
Judith Lucia Gomez-Porras1† Diego Mauricio Riaño-Pachón2† Begoña Benito1† Rosario Haro1† Kamil Sklodowski3,4
Diego Mauricio Riaño-Pachón2† Begoña Benito1† Rosario Haro1† Kamil Sklodowski3,4 Alonso Rodríguez-Navarro1
Alonso Rodríguez-Navarro1 Ingo Dreyer1,3*
Ingo Dreyer1,3*As heritage from early evolution, potassium (K+) is absolutely necessary for all living cells. It plays significant roles as stabilizer in metabolism and is important for enzyme activation, stabilization of protein synthesis, and neutralization of negative charges on cellular molecules as proteins and nucleic acids. Land plants even enlarged this spectrum of K+ utilization after having gone ashore, despite the fact that K+ is far less available in their new oligotrophic habitats than in sea water. Inevitably, plant cells had to improve and to develop unique transport systems for K+ accumulation and distribution. In the past two decades a manifold of K+ transporters from flowering plants has been identified at the molecular level. The recently published genome of the fern ally Selaginella moellendorffii now helps in providing a better understanding on the molecular changes involved in the colonization of land and the development of the vasculature and the seeds. In this article we present an inventory of K+ transporters of this lycophyte and pigeonhole them together with their relatives from the moss Physcomitrella patens, the monocotyledon Oryza sativa, and two dicotyledonous species, the herbaceous plant Arabidopsis thaliana, and the tree Populus trichocarpa. Interestingly, the transition of green plants from an aqueous to a dry environment coincides with a dramatic reduction in the diversity of voltage-gated potassium channels followed by a diversification on the basis of one surviving K+ channel class. The first appearance of K+ release (Kout) channels in S. moellendorffii that were shown in Arabidopsis to be involved in xylem loading and guard cell closure coincides with the specialization of vascular plants and may indicate an important adaptive step.
The absolute requirement for K+in all living cells was already fixed from the cradle of evolution in the sea. Among all the cations that were present in the marine environment K+ was utilized by cells as the major cation for essential functions as maintaining electroneutrality and osmotic equilibrium. Further evolutionary steps in the cellular K+-rich environment then employed K+ as regulator of protein activities being essential for several biochemical processes. The interactions of potassium with these proteins depend on the unique electrochemical properties of K+ ions, i.e., the topology of their electrical charge-density. These features cannot or only incompletely be mimicked by Na+ or by any other cation because they all differ from K+ in their electron shell configuration and consequently also in the arrangement of the surrounding hydration shell. K+ thus became indispensably necessary for living cells; a dependency also inherited to Embryophyta, where K+ can contribute up to 10% of the dry mass (Leigh and Wyn Jones, 1984). Terrestrial plants even developed new functions for K+ such as turgor-driven processes like stomatal movement, phototropism, gravitropism, and cell elongation (Ashley et al., 2006; Rodriguez-Navarro and Rubio, 2006; Amtmann and Armengaud, 2009; Amtmann and Blatt, 2009; Maathuis, 2009; Szczerba et al., 2009). Embryophyta need to survive in oligotrophic environments where K+ is present at much lower concentrations than in sea water; the potassium concentration in normal soil solution (10–100 μM) is considerably variable and about three to four orders of magnitude lower than in the plant. Therefore, not only for potassium homeostasis (the maintenance of a dynamic equilibrium in the cellular K+ concentration) but also for K+ uptake from the environment and its distribution throughout the organism, plants have to invest energy and need a set of specialized transporter proteins.
Pioneering work by Epstein et al. (1963) proposed that K+ uptake from soil into plant cells is mediated by two mechanisms that take advantage of the electrical gradient and/or the proton motive force established by H+-ATPases. One was characterized as a high-affinity system (mechanism I), showing apparent affinities in the range of ∼20 μM, that can transport also Na+ when K+ is not present. The other one (mechanism II) showed a much lower affinity and provided an increasing contribution from >200 μM to mM external K+ concentrations. During the last two decades a variety of potassium-permeable transmembrane transport systems – potentially underlying these two components – were identified at the molecular level. They were classified into five major gene families (Maser et al., 2001, 2002b; Véry and Sentenac, 2002, 2003; Lebaudy et al., 2007): (i) voltage-gated K+ channels, (ii) non-voltage-gated (tandem-pore) K +(TPK) channels, (iii) high-affinity K + transporters of the HAK type, (iv) high-affinity K + transporters of the HKT type, and (v) cation-proton antiporters (CPAs). K+ channels likely underlie the experimentally observed low affinity component in plants, whereas HAK transporters contribute to the high-affinity K+ uptake component. HKT transporters are responsible for a K+-dependent Na+ component (Rodriguez-Navarro and Rubio, 2006). However, this is not a generalized strict separation. Channels contribute to the high-affinity K+ uptake component and transporters might contribute under certain conditions also to low affinity transport.
Here we took advantage from the recently published genome of Selaginella moellendorffii (Banks et al., 2011) and prepared an inventory of K+ transporters in this lycophyte. We focused especially on transporters of the HAK and HKT type and on K+ channels. For information on other potentially K+-permeable transporters such as KEA or CHX belonging to the class of monovalent CPAs we refer to a recent excellent review especially dedicated to these proteins (Chanroj et al., 2012). We are comparing the results from S. moellendorffii with those from the moss Physcomitrella patens, the monocotyledon Oryza sativa, as well as with those from two dicotyledonous species, the Brassicaceae Arabidopsis thaliana, and the tree Populus trichocarpa. Voltage-gated K+ channels are also compared with those from Chlorophyta.
The high-affinity K+ (HAK) transporter gene family – also called KT or KUP transporter family – is an ancient large family with members in Bacteria, Archaea, Fungi, Amoebozoa, and probably also in some species of Animalia (Grabov, 2007; Benito et al., 2011). Initially HAK genes have been deduced from plants by their similarity to K+ uptake permeases (KUP) from E. coli (Schleyer and Bakker, 1993) and high-affinity K+ transporters (HAK) from fungi (Banuelos et al., 1995; Quintero and Blatt, 1997; Santa-Maria et al., 1997; Fu and Luan, 1998; Kim et al., 1998). Several members of this family were shown to function as K+ uptake transporters in plants especially when the external potassium concentration was in the low μM range (Gierth et al., 2005; Aleman et al., 2011) indicating that HAK transporters are involved in high-affinity K+ uptake. Interestingly, all plant genomes analyzed so far contain genes encoding HAK transporters, while in Bacteria, Archaea, and Fungi they were found only in a subset of species (Grabov, 2007; Benito et al., 2011). The HAK family is the largest family of potential K+ transporters in plants and members of this family are expressed in nearly all tested plant tissues suggesting that HAK transporters have a general function in K+ supply (Banuelos et al., 2002).
To date, the topology of HAK transporters has neither been determined experimentally nor by in silico predictions; nevertheless, hydropathy profiles of these proteins suggest about 12 putative transmembrane segments and a long hydrophilic COOH-terminal region. In genome-wide screenings, HAK transporter proteins can be pinpointed by the presence of several conserved consensus motifs (see Materials and Methods). Our screenings identified 13 HAKs in Arabidopsis, 27 in rice, 22 in poplar, 18 in P. patens, and 11 in S. moellendorffii (Table 1; see Yang et al., 2009, for comparison). It is likely that the genome of P. trichocarpa contains more genes coding for HAK transporters, because especially in the screening of poplar we discarded partial sequences resulting from pre-mature gene annotation. Phylogenetic analyses allowed subdividing them into six independent groups (Figure 1A; see Rubio et al., 2000, for initial grouping-into Groups I–IV) and revealed that the last recent common ancestor of all embryophytes had two HAK transporters (Figure 1B). One of these diverged into current Group II, whereas the other was duplicated at least three times before the origin of tracheophytes. Two early duplication events got lost in the lineage leading to tracheophytes and led to P. patens-specific gene family amplifications (Groups V and VI). HAK transporters in S. moellendorffii spread over the other clades (Groups I–IV).

Table 1. Transporters of the HAK type presented in this study.
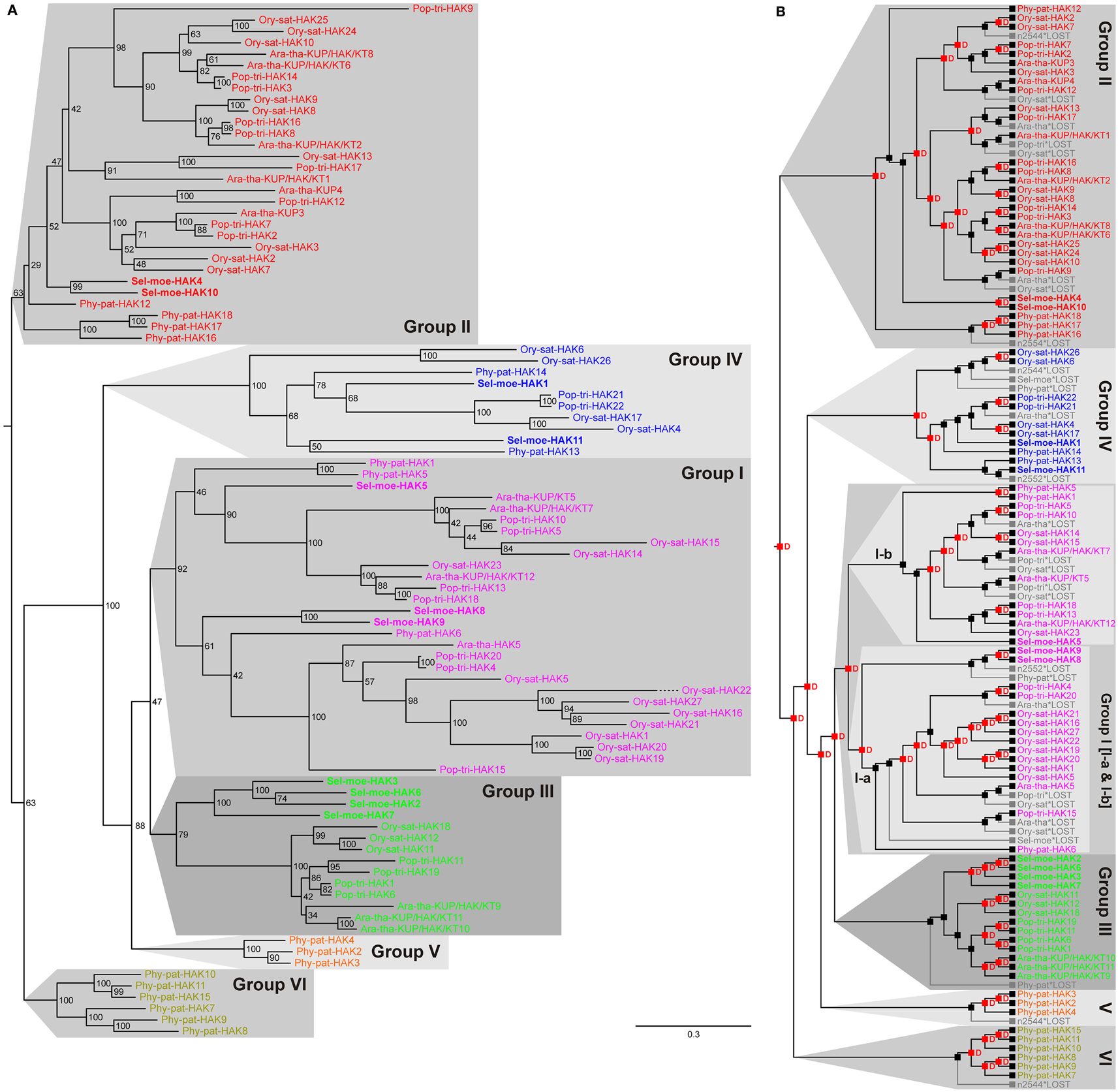
Figure 1. Evolutionary relationships among HAK transporters in land plants. (A) There are six clearly distinguished clades of HAK transporters in extant land plants, i.e., Groups I, II, III, IV, V, and VI. Each group represents an independent group of orthologs. Groups V and VI are P. patens-specific gene family amplifications. Evolutionary relationships among HAK transporters in land plants. (B) Reconciliation analysis of HAK transporters. The last recent common ancestor of all embryophytes had two HAK transporters. One of these diverged into current Group II, whereas the other was duplicated at least three times before the origin of tracheophytes. Two of these duplications got lost in the lineage leading to tracheophytes forming P. patens-specific groups. Red “D”s at branching points indicate predicted gene duplications. Gray branches indicate gene losses.
Functional information on HAK transporters is unfortunately still scarce. Most data are available for HAK transporters belonging to Group I. Ara-tha-HAK5, Ory-sat-HAK1, Ory-sat-HAK5, and Phy-pat-HAK1 were characterized as high-affinity K+ transporters (Rubio et al., 2000; Banuelos et al., 2002; Gierth et al., 2005; Garciadeblas et al., 2007; Qi et al., 2008; Horie et al., 2011b). It might thus be an educative guess to propose high-affinity K+-uptake properties also for the S. moellendorffii orthologs in the same clade, i.e., Sel-moe-HAK5, Sel-moe-HAK8, and Sel-moe-HAK9.
HAK transporters may not only mediate transport across the plasma membrane. Transient expression of the Ory-sat-HAK10::GFP fusion protein in living onion epidermal cells targeted this protein (from Group II) to the tonoplast (Banuelos et al., 2002); and Ara-tha-KUP/HAK/KT12 (from Group I) was found in the chloroplast proteome (Kleffmann et al., 2004; Peltier et al., 2004). Additionally, HAK transporters may not exclusively transport K+. Phy-pat-HAK1 and Ara-tha-HAK5, for instance, were reported to be permeable also for Cs+ (Garciadeblas et al., 2007; Qi et al., 2008); and Phy-pat-HAK13 belonging to Group IV was recently characterized as a high-affinity Na+ uptake transporter (Benito et al., 2012). We therefore propose for the closely related Sel-moe-HAK1 and Sel-moe-HAK11 from S. moellendorffii similar sodium-transport features. This phylogenetic divergence may indicate a – so far underexplored – diversity of HAK transporters in fine-tuned function of K+ uptake and re-distribution, cellular expression, and/or sub-cellular targeting.
HKTs in plants belong to a family of monovalent cation transporters comprising also the fungal TRKs (K+ transporters) and bacterial KtrABs (Na+-dependent K+ transporter), for instance (Corratge-Faillie et al., 2010). Proteins of this family share a common structure of four TM-P-TM motifs (every two transmembrane α-helices are connected by ∼30 aa-long pore-forming P segments), which might have evolved from an ancestor related to the bacterial KscA K+ channel of Streptomyces lividans (Durell and Guy, 1999; Figure 2A). The plant HKT family comprises transporters that mediate Na+ uptake in roots or in other plant organs. They accumulate Na+ from the soil and recirculate it throughout the plant. There are two types of plant HKT transporters that can be distinguished by the amino acid sequence of the selectivity filter (the narrowest part of the permeation pathways that selects one ion species over others) of the first TM-P-TM motif: (i) S-S-M and (ii) [T,S,I]-G-L. Two HKTs from O. sativa and A. thaliana belonging to the first type have been well characterized in planta as Na+ uptake transporters (Uozumi et al., 2000; Rus et al., 2001; Maser et al., 2002a,c; Berthomieu et al., 2003; Garciadeblas et al., 2003; Sunarpi et al., 2005; Horie et al., 2007; Xue et al., 2011). The function of the second type has not been studied in plants. Nonetheless, two members of this group, from barley and wheat, mediate K+ or Na+uniport or Na+-K+symport – depending on the protein expression level – when heterologously expressed in yeast cells (Haro et al., 2005; Banuelos et al., 2008). Functional expression of this type of transporters in Xenopus oocytes produced similar results with slight variations regarding K+ versus Na+ permeability, symport activity, and permeability to divalent cations (Rubio et al., 1995; Gassman et al., 1996; Jabnoune et al., 2009; Lan et al., 2010; Horie et al., 2011a; Oomen et al., 2012).
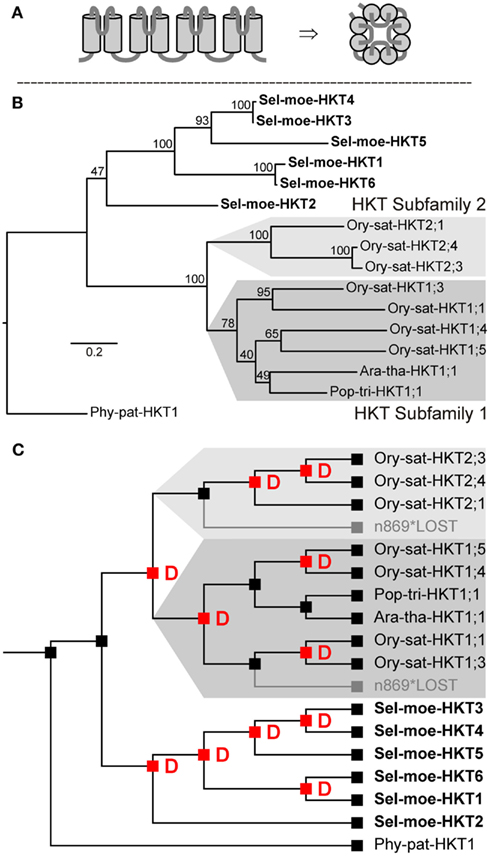
Figure 2. Transporters of the HKT type in land plants. (A) Predicted structure with a fourfold TM-P-TM motif in side view (left) and assembled as functional transporter in top view (right). (B) Evolutionary relationships among HKTs in land plants. This family is represented by a single group of orthologs that includes all considered extant land plants. (C) Reconciliation analysis of transporters of the HKT type. The last recent common ancestor of embryophytes had a single HKT-coding gene. Successively this family has undergone independent gene amplifications in different lineages, i.e., lycophytes and angiosperms. Red “D”s at branching points indicate predicted gene duplications. Gray branches indicate gene losses.
In genome-wide screenings, proteins of the HKT type can be pinpointed by the presence of several conserved consensus motifs (see Materials and Methods). Our screenings identified one HKT-coding gene in Arabidopsis, seven in rice, one in poplar, one in P. patens, and six in S. moellendorffii (Table 2). Phylogenetic analyses grouped all of them into a single group of orthologs (Figure 2B) indicating that the most recent common ancestor of all embryophytes comprised a single protein of the HKT type. P. patens has a single extant representative (Phy-pat-HKT1), whereas in tracheophytes several duplication events occurred in different lineages (Figure 2C). The obviously independent multiplication of HKT-coding genes in rice and S. moellendorffii may be correlated with the affinity of these vascular plants to moisture environments. Probably, a larger variety of Na+/K+ transporters provides some advantage for better adaptation. Initially, the HKT family has been partitioned into the two subfamilies one and two, and transporter nomenclature was adjusted accordingly of the type “species HKT subfamily; No” (Platten et al., 2006). Subfamily one gathers transporters with the S-S-M signature in the selectivity filter of the first TM-P-TM motif. Our analysis now reveals that this subfamily division emerged in land plants only after the separation of Lycopodiophyta. Thus, the proposed unified nomenclature cannot be applied to all plant HKT genes. The rules fail, for instance, for HKTs from S. moellendorffii and P. patens (see also Haro et al., 2010).
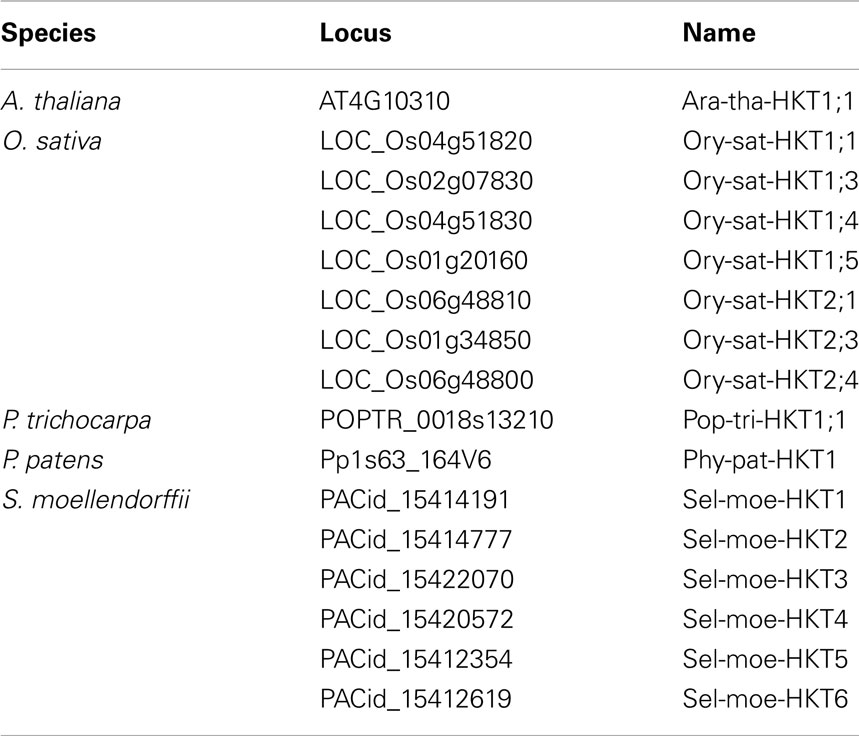
Table 2. Transporters of the HKT type presented in this study.
At the functional level, the six HKTs of S. moellendorffii very likely share properties of the orthologs from other species. They may thus be implicated in K+/Na+ recirculation in this vascular plant and could contribute not only to K+ transport but in first line to desalination and Na+ detoxification.
Potassium channels play important roles in many physiological aspects of higher plants such as osmoregulation, turgor-driven movements, and ion uptake. It is estimated that K+ channels can contribute to more than 50% of the nutritional K+ uptake under most field conditions (Spalding et al., 1999; Amtmann and Blatt, 2009). In angiosperms there are two large groups of K+ channels: voltage-gated channels, the activity of which is regulated by the transmembrane voltage (Dreyer and Blatt, 2009), and non-voltage-gated K+ channels. Non-voltage-gated K+ channels form the class of tandem-pore K+ (TPK) channels. Functional TPK channels are proposed to form dimers consisting of two identical subunits (Maitrejean et al., 2011). Each subunit is characterized by a structure with four transmembrane domains and two pore-forming loops between the first and second and the third and forth membrane-spanning domain, respectively (Figure 3A; Voelker et al., 2010).
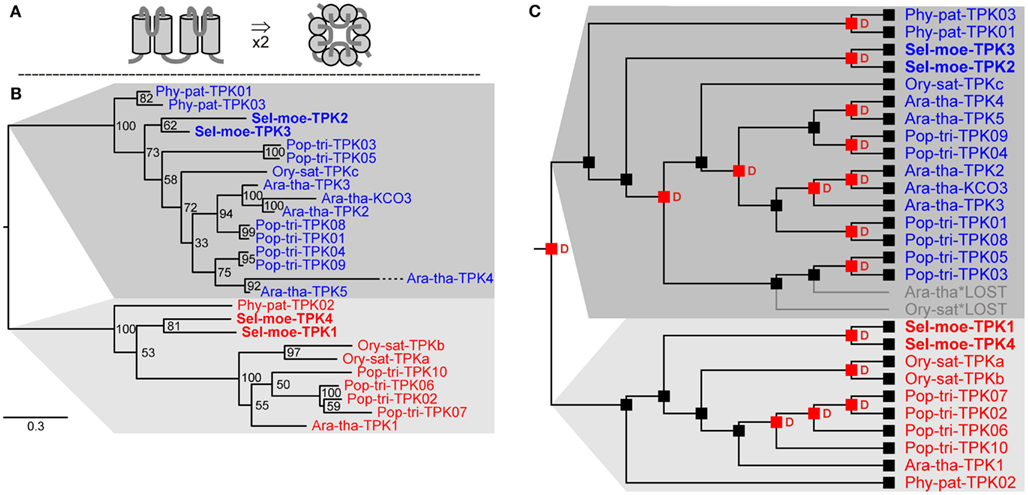
Figure 3. Tandem-pore K+ (TPK) channels in land plants. (A) Predicted structure of one subunit with a two-fold TM-P-TM motif in side view (left) and assembled functional channel dimer in top view (right). (B) Evolutionary relationships among TPK channels in land plants. There are two clear groups of TPK orthologs in extant land plants. (C) Reconciliation analysis of TPK channels. The last recent common ancestor of embryophytes had two genes coding for TPK channel subunits. Red “D”s at branching points indicate predicted gene duplications. Gray branches indicate gene losses.
Searching for proteins with the characteristic pore-forming region, in the genome of S. moellendorffii four genes coding for TPK channel subunits could be identified (Table 3). Together with the six TPKs from Arabidopsis (Ara-tha-TPK1–5 and Ara-tha-KCO3), three from rice, ten from poplar, and three from P. patens they could be classed into two groups of orthologs (Figure 3B). This implies that the ancestor of land plants had already two of these genes. A deeper phylogenetic analysis revealed several duplication events in the two groups, both species-specific and at higher levels (Figure 3C). A remarkable example in this context is KCO3 from A. thaliana. This subunit lacks the first of the two pore loops and was originally considered as founder of a separate channel family with structural features (TM-P-TM; two transmembrane α-helices, and pore-forming P segment) similar to the simplest class of K+ channels from bacteria and animals. It became evident, however, that Ara-tha-KCO3 developed through a very recent evolutionary event involving gene duplication of the Ara-tha-TPK2 gene followed by partial deletion (Marcel et al., 2010; Voelker et al., 2010). And indeed, in line with this concept, neither the genome of S. moellendorffii nor that of P. patens appears to contain genes coding for K+ channels of the TM-P-TM type.
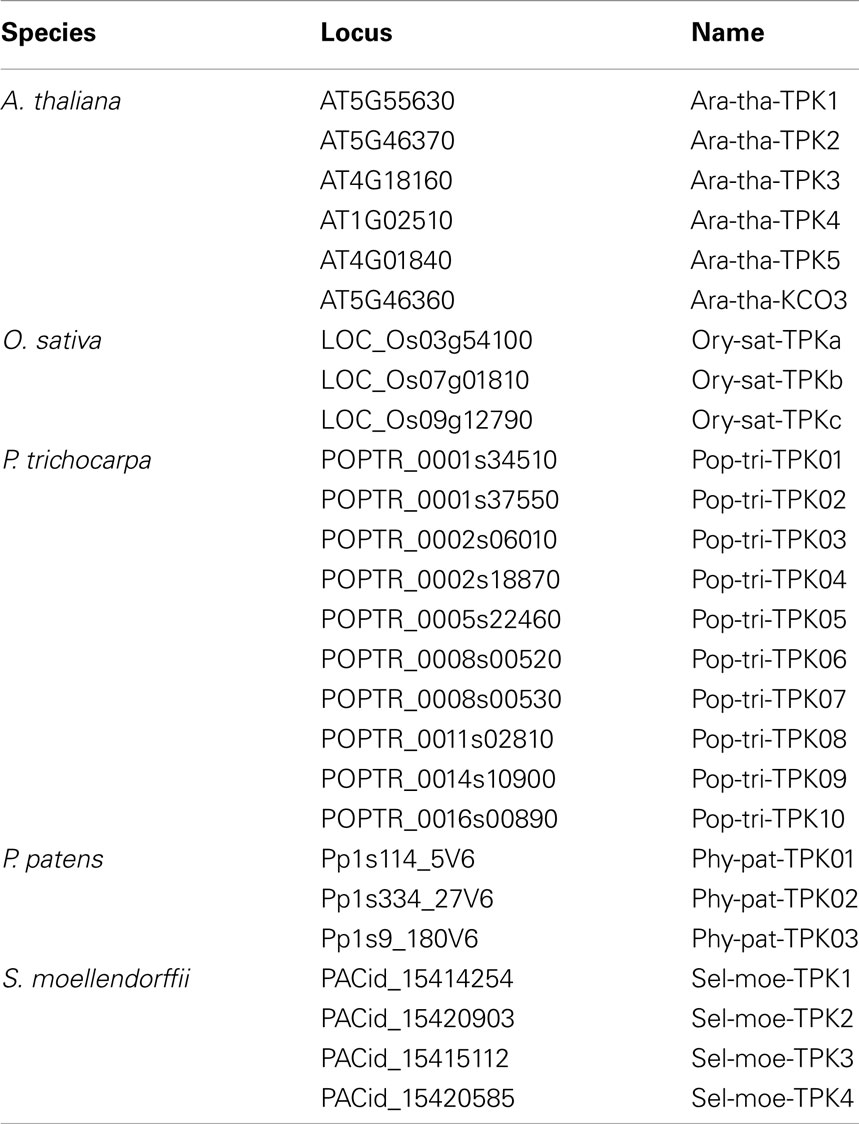
Table 3. Two pore K+ (TPK) channels presented in this study.
TPK channels in plants were reported to be targeted to the vacuolar membrane (Czempinski et al., 2002; Voelker et al., 2006; Latz et al., 2007; Dunkel et al., 2008; Isayenkov et al., 2011a,b). The exception is Ara-tha-TPK4 which has been reported to be targeted also to the plasma membrane (Becker et al., 2004). However, orthologs of Ara-tha-TPK4 were only found in the genus Arabidopsis so far (i.e., A. thaliana and A. lyrata; Voelker et al., 2010) but not in other plant species indicating a rather recent evolutionary event in channel specialization. Therefore, we have justified reasons to hypothesize that TPKs in bryophytes and lycophytes are vacuolar K+ channels. Their physiological role, however, remains as speculative as that of TPKs in other plants (Voelker et al., 2010).
Voltage-gated K+ channels are tetrameric proteins built of four α-subunits. One subunit shows usually a structure with six transmembrane domains and one pore loop (5TM-P-TM). The first four transmembrane domains fold into the voltage-sensor module, and the pore loop together with the fifth and sixth transmembrane domains establishes the permeation pathway module (Figure 4A; Dreyer and Blatt, 2009).
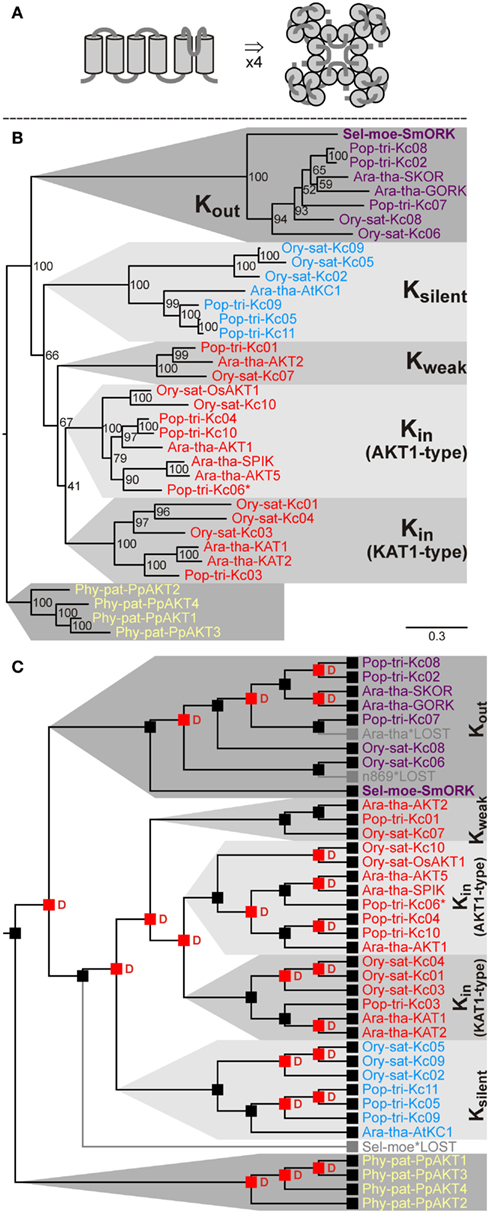
Figure 4. Evolutionary relationships among voltage-gated Shaker-like K+ channels in land plants. (A) Predicted structure of one subunit with a 5TM-P-TM motif in side view (left) and assembled functional channel tetramer in top view (right). (B) Evolutionary relationships among Shaker-like channels in land plants. Extensive functional analyses identified inward-rectifying (Kin) channels, outward-rectifying (Kout) channels, weakly rectifying (Kweak) channels, and silent (Ksilent) channel subunits that assemble with Kin subunits and modulate K+ uptake channel properties. (C) Reconciliation analysis of Shaker-like K+ channels. The common ancestor of land plants had a single Shaker-like K+ channel. Since then several amplifications have occurred: a bryophyte specific amplification following the split between the lineage leading to P. patens and the tracheophytes, and duplications in the tracheophyte lineage. The common ancestor of tracheophytes had two genes coding for Shaker-like K+ channel subunits. One of these got lost in the lineage leading to S. moellendorffii after the split of angiosperms. Red “D”s at branching points indicate predicted gene duplications. Gray branches indicate gene losses. For Pop-tri-Kc06 only partial sequence information was available (indicated by an asterisk).
Voltage-gated potassium channels in angiosperms are targeted to the plasma membrane and could normally be grouped into the class of Shaker-like K+ channels that subdivides into four functional subgroups: (a) Inward-rectifying (Kin) channels open at membrane hyperpolarization and are responsible for K+ uptake. (b) Silent (Ksilent) channel subunits assemble with Kin subunits and modulate K+ uptake channel properties. (c) Weakly rectifying (Kweak) channels are specialized Kin channels that show a bi-modal gating behavior. They appear to play a special role in the energy household of vascular tissues. (d) Outward-rectifying (Kout) channel subunits open at depolarizing voltages and mediate K+ release, e.g., during xylem loading or stomata closure (see Dreyer and Uozumi, 2011, for a contemporary review). Our screening strategy based on the characteristic pore-forming region identified nine – already known – genes coding for Shaker-like channels in Arabidopsis, eleven in rice, and eleven in poplar. Genes coding for Shaker-like K+ channels were also identified in the moss P. patens and in the fern ally S. moellendorffii (Table 4). However, whereas four Kin-like channels were identified in P. patens, despite a very careful screening strategy no such gene could be found in S. moellendorffii. Instead there, a gene coding for a Kout channel subunit was discovered as the only Shaker-like channel (Figure 4). This channel comprises all the essential structural features that were shown in the Ara-tha-SKOR Kout channel to be responsible for a unique K+ sensing property (Johansson et al., 2006). Kout channels open upon depolarization but additionally adjust their gating to the prevailing concentration of K+ outside. As a consequence, they open only at voltages positive of the K+ equilibrium voltage, EK, when the electrochemical driving force is directed outward and so ensure K+ efflux regardless of the extracellular K+ concentration. This ability to adapt channel gating to the cation concentration outside guarantees an efficient K+ release during xylem loading and stomatal closure, for instance, even under varying external K+ (from 10 nM to 100 mM; Blatt, 1988; Schroeder, 1988; Wegner and de Boer, 1997; Gaymard et al., 1998; Ache et al., 2000). From analogy we may postulate that the presence of a Kout channel in the vascular plant S. moellendorffii and its absence in the non-vascular plant P. patens is correlated with the important evolutionary step of vascularization. In contrast, it is rather difficult to find an explanation for the loss of the Kin/Kweak/Ksilent channel branch in S. moellendorffii.

Table 4. Voltage-gated Shaker-like K+ channels presented in this study.
In addition to Shaker-like channels the genomes of both, S. moellendorffii and P. patens, contain members of another class of putatively voltage-gated potassium channels (Table 5). These channels show some similarity with large conductance Ca2+-activated K+ channels (“big K” = BK channels), a channel type that is widely present in animals (including humans) but absent in flowering plants, for instance. BK channels activate in response to membrane depolarization and binding of intracellular Ca2+ and Mg2+ (Latorre et al., 2010). These channels are built of α- and β-subunits, where – as in Shaker-like channels – four α-subunits form the per se functional permeation pathway-establishing unit and the β-subunits just modulate and fine-tune channel properties. In contrast to Shaker-like channels the BK channel protein consists of seven (instead of six) transmembrane domains (6TM-P-TM structure) that lead to an exoplasmic N-terminus. Also BK-like channel α-subunits from P. patens and S. moellendorffii show a larger N-terminal region compared to plant Shaker-like channels. It might thus be speculated that also these proteins fold into a 6TM-P-TM structure instead of the 5TM-P-TM Shaker-like topology.
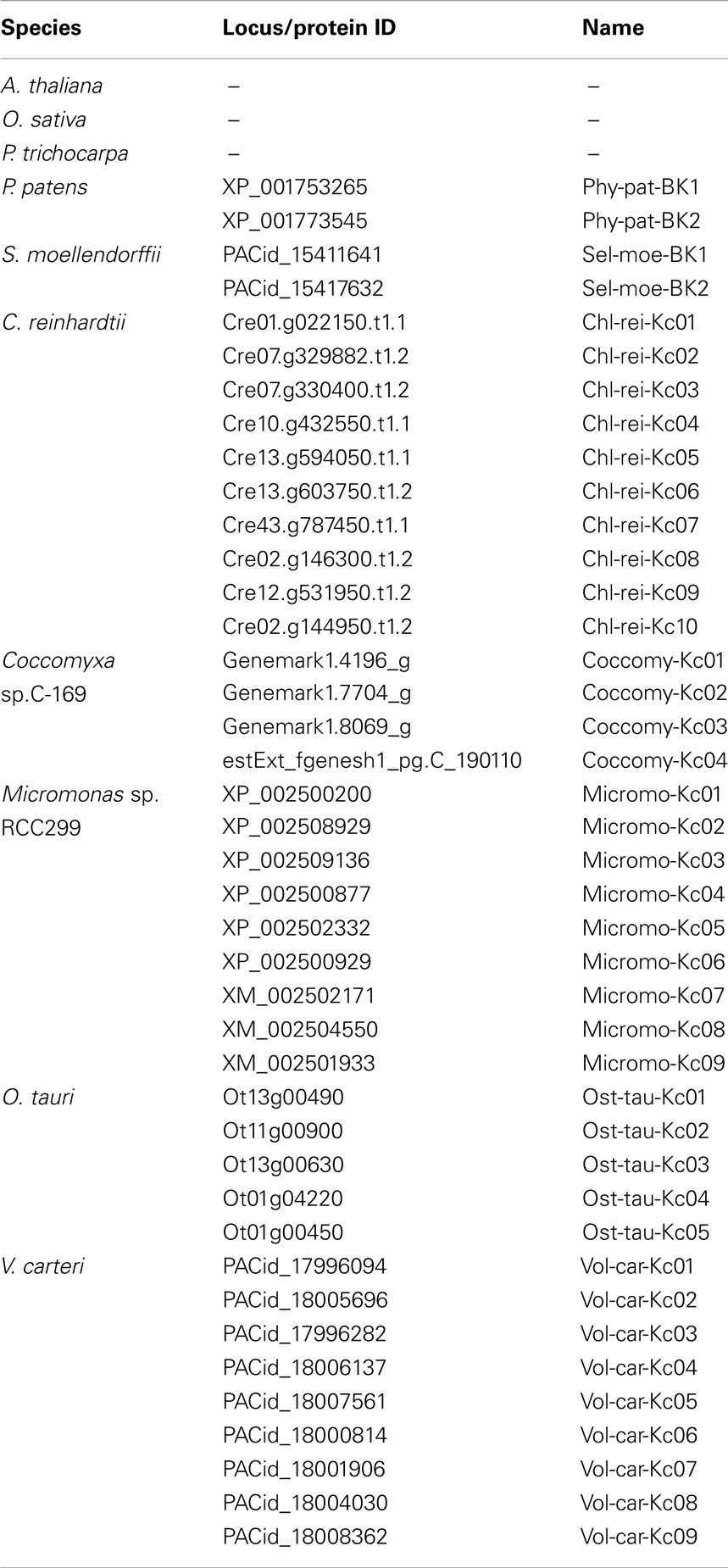
Table 5. Voltage-gated K+ channels of other types presented in this study.
The functional properties and the physiological roles of plant BK-like channels are unknown. In mammalian tissues, BK channels serve as a negative-feedback mechanism for excitatory events that lead to increases in calcium concentration or membrane depolarization. In this way, they play a key role, for instance, in regulating the contractile tone in vascular smooth muscle cells or help to terminate the action potential and thus modulate secretion in chromaffin cells. It might be speculated that – at least in S. moellendorffii – the two BK-like channels could compensate for the absent Shaker-like Kin channels in carrying out functions in K+ uptake and distribution.
To assess the evolutionary origin of the non-Shaker-like channels we screened the genomes of the green algae Chlamydomonas reinhardtii, Coccomyxa sp.C-169, Micromonas sp. RCC299, Ostreococcus tauri, and Volvox carteri for voltage-gated K+ channels. Despite the fact that this transporter class in algae exhibits a huge structural diversity comprising also homologs of plant Shaker-like channels (Figure 5), a clear trace leading to BK-like channels in S. moellendorffii or P. patens could not be identified. The two most similar channels from Volvox and Chlamydomonas share an identity of 16–19% over a stretch of ∼500 amino acids. In comparison, a BLAST search at NCBI1 limited to a query coverage of >45% resulted as best hit outside Animalia, S. moellendorffii, or P. patens in a voltage-gated K+ channel from Phytophthora infestans (XM_002998337) with ∼28% identity over a stretch of ∼500 amino acids. Unfortunately, from all these results we cannot resolve unequivocally the origin of BK-like channels in S. moellendorffii and P. patens. Neither can we exclude the possibility that bryophytes and lycophytes may have acquired these K+ channel genes from Fungi or Protozoa. However, our data (Figure 5) clearly indicate that the diversity of voltage-gated K+ channels observable in Chlorophyta collapsed contemporaneously with the transition of green plants from an aqueous to a dry environment. In higher plants the subsequent functional diversification into Kin/Kout/Kweak/Ksilent (Figure 4) took place on the basis of only one surviving channel class.

Figure 5. Voltage-gated K+ channels in Chlorophyta, Bryophyta, and Lycophyta. In contrast to land plants, voltage-gated K+ channels in algae show a large structural diversity. The functional variety of K+ channels in higher plants (Figure 4; Shaker-like K+ channels) developed from only one of these channel types.
In the haploid genome of the spike moss S. moellendorffii we identified 1 homolog of voltage-gated outward-rectifying K+ release channels, 4 homologs of voltage-independent tandem pore K+ channels, 2 homologs with some similarity to large conductance Ca2+-activated K+ (BK) channels, 11 homologs of transporters of the HAK type, as well as 6 homologs of the HKT type (Table 6). On the basis of phylogenetic analyses, detailed functional properties can be predicted for a few of them. Most probable is that Sel-moe-SmORK forms voltage-gated K+ release channels involved in stomatal closure and/or in K+ loading into the vascular bundles.

Table 6. Summary – Molecular toolkit for K+ uptake and re-distribution in S. moellendorffii.
Putative K+ transporters were identified using the conceptual proteomes of A. thaliana (TAIR10 Genome release), O. sativa spp. Indica, P. trichocarpa, P. patens, and S. moellendorffii (Phytozome v6.0) and the algae genomes Coccomyxa_C169, Micromonas RCC299, O. tauri (v2, v3, and v4 respectively2), V. carteri, and C. reinhardtii (Phytozome v8.0) by screening with different transporter class-specific protein motifs: three motifs for K+ channels ((1) [S,T]-x-x-T-x-G-[Y,F,L]-G-[D,E], (2) R-[L,F]-x-R-[L,V,I,A,G]-x-[R,C,K]-[V,A,L,M], (3) [A,V,S]-Y-[L,I]-[I,L]-G-[N,I]-[M,I]-T-[N,A]-L-[V,I]); two motifs for HKTs ((4) [S,T,A]-x-[F,Y,V,L,C]-x-[D,N,S]-G, (5) [G,A]-[Y,F]-[G,A]-x-[V,A,I]-G-[L,M,Y,F]-[S,T]); and five motifs for HAK transporters ((6) [A,G]-[D,S,G]-[V,L,I,M]-x-x-[S,A]-P-L-Y; (7) [A,G]-[N,D,H,S]-[D,N]-x-G-[E,Q,D,N]-[A,G]; (8) [A,G,S]-[D,N]-[G,S,A,C]-x-[L,I,V,F]-x-P-x-[V,I,L,M]-[A,S]; (9) G-[S,A,T,C]-E-[A,G]-x-[F,Y]-A-[D,N,E]-[L,I,V]-[G,C,S,A]-x-F; (10) [Y,F]-x-x-x-x-x-[H,F,Y]-G-Y-x-[E,D]) using the FUZZNUC program from EMBOSS (Rice et al., 2000). Additionally, results were checked against BLAST searches in the five genomes using known transporters of different classes from Arabidopsis and rice as templates. In order to eliminate false-positives the resulting raw-data were curated in a semi-automatic way. In a first step sequences with a length <70% of the average length between the outermost motifs in the corresponding Arabidopsis transporters were discarded. Subsequently, the remaining n protein sequences of each transporter type of each species were pairwise aligned using ClustalW23. From the resulting n(n−1)/2 pairs those with a score of <20 and of 100 (identical sequences) were removed. The residual pairs fragmented the sequences into distinct groups. That group with the highest similarity to the corresponding Arabidopsis transporters was selected for further analyses.
To verify whether the screening for K+ channels in S. moellendorffii was exhaustive, its genome was screened in the six-frame translations using the program SIXPACK from EMBOSS (Rice et al., 2000) for the presence of the K+-selectivity filter motif G-Y-G in ORFs. Following a positive hit, the closer environment of the GYG was inspected manually for further characteristic sequence features allowing categorizing the peptide to be part of a K+ channel. As a result – besides the K+ channels obtained already in the first screening – only the Kout channel SmORK could be identified in addition.
Sequences from each family were aligned using MAFFT (Katoh and Toh, 2010), and alignments were filtered using GBlocks (Castresana, 2000) in order to eliminate regions of low quality. Briefly, the minimum number of sequences for a conserved position was half the number of sequences, the minimum number of sequences for a flanking position was half the number of sequences, the maximum length of contiguous non-conserved positions was 20, and the minimum length of a block was two, positions with gaps were not treated differently from other position. Evolutionary relationships were inferred by Maximum Likelihood using RAxML and 1000 bootstrap replicates (Stamatakis, 2006). The evolutionary model used for phylogenetic analyses was inferred using ProtTest (Darriba et al., 2011). For two pore channels and HAK transporters the model was LG + γ, for HKTs and Shaker-like channels it was JTT + γ. In order to root and resolve the gene trees we performed a gene tree-species tree reconciliation analysis using the species tree from Lang et al. (2010; TreeBase 10409). Reconciliation analysis was carried out in Notung 2.6 (Chen et al., 2000; Vernot et al., 2007). To get an idea of the phylogenetic structure of the other voltage-gated K+ channels displayed in Figure 5, the sequences were hierarchically clustered based on pairwise identities between two sequences using UPGMA (Unweighted Pair Group Method with Arithmetic Mean). UPGMA analyses were carried out in MAFFT4.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
This work was supported by grants from the Spanish Ministerio de Economía y Competitividad to Ingo Dreyer and Alonso Rodríguez-Navarro (BFU2011-28815; AGL2007-61705), a Marie Curie Career Integration Grant to Ingo Dreyer (FP7-PEOPLE-2011-CIG No. 303674 – Regopoc), as well as by a Marie-Curie Cofund fellowship to Judith Lucia Gomez-Porras. Kamil Sklodowski is a recipient of a doctoral fellowship from the Max-Planck Research School “Primary Metabolism and Plant Growth.”
The alignments used for generating the phylogenetic trees presented in this study are available online as Supplementary Material.
The Supplementary Material for this article can be found online at http://www.frontiersin.org/Plant_Evolution_and_Development/10.3389/fpls.2012.00167/abstract
Ache, P., Becker, D., Ivashikina, N., Dietrich, P., Roelfsema, M. R., and Hedrich, R. (2000). GORK, a delayed outward rectifier expressed in guard cells of Arabidopsis thaliana, is a K+-selective, K+-sensing ion channel. FEBS Lett. 486, 93–98.
Aleman, F., Nieves-Cordones, M., Martinez, V., and Rubio, F. (2011). Root K+ acquisition in plants: the Arabidopsis thaliana model. Plant Cell Physiol. 52, 1603–1612.
Amtmann, A., and Armengaud, P. (2009). Effects of N, P, K and S on metabolism: new knowledge gained from multi-level analysis. Curr. Opin. Plant Biol. 12, 275–283.
Amtmann, A., and Blatt, M. R. (2009). Regulation of macronutrient transport. New Phytol. 181, 35–52.
Ashley, M. K., Grant, M., and Grabov, A. (2006). Plant responses to potassium deficiencies: a role for potassium transport proteins. J. Exp. Bot. 57, 425–436.
Banks, J. A., Nishiyama, T., Hasebe, M., Bowman, J. L., Gribskov, M., Depamphilis, C., Albert, V. A., Aono, N., Aoyama, T., Ambrose, B. A., Ashton, N. W., Axtell, M. J., Barker, E., Barker, M. S., Bennetzen, J. L., Bonawitz, N. D., Chapple, C., Cheng, C., Correa, L. G., Dacre, M., DeBarry, J., Dreyer, I., Elias, M., Engstrom, E. M., Estelle, M., Feng, L., Finet, C., Floyd, S. K., Frommer, W. B., Fujita, T., Gramzow, L., Gutensohn, M., Harholt, J., Hattori, M., Heyl, A., Hirai, T., Hiwatashi, Y., Ishikawa, M., Iwata, M., Karol, K. G., Koehler, B., Kolukisaoglu, U., Kubo, M., Kurata, T., Lalonde, S., Li, K., Li, Y., Litt, A., Lyons, E., Manning, G., Maruyama, T., Michael, T. P., Mikami, K., Miyazaki, S., Morinaga, S., Murata, T., Mueller-Roeber, B., Nelson, D. R., Obara, M., Oguri, Y., Olmstead, R. G., Onodera, N., Petersen, B. L., Pils, B., Prigge, M., Rensing, S. A., Riano-Pachon, D. M., Roberts, A. W., Sato, Y., Scheller, H. V., Schulz, B., Schulz, C., Shakirov, E. V., Shibagaki, N., Shinohara, N., Shippen, D. E., Sorensen, I., Sotooka, R., Sugimoto, N., Sugita, M., Sumikawa, N., Tanurdzic, M., Theissen, G., Ulvskov, P., Wakazuki, S., Weng, J. K., Willats, W. W., Wipf, D., Wolf, P. G., Yang, L., Zimmer, A. D., Zhu, Q., Mitros, T., Hellsten, U., Loque, D., Otillar, R., Salamov, A., Schmutz, J., Shapiro, H., Lindquist, E., Lucas, S., Rokhsar, D., and Grigoriev, I. V. (2011). The Selaginella genome identifies genetic changes associated with the evolution of vascular plants. Science 332, 960–963.
Banuelos, M. A., Garciadeblas, B., Cubero, B., and Rodriguez-Navarro, A. (2002). Inventory and functional characterization of the HAK potassium transporters of rice. Plant Physiol. 130, 784–795.
Banuelos, M. A., Haro, R., Fraile-Escanciano, A., and Rodriguez-Navarro, A. (2008). Effects of polylinker uATGs on the function of grass HKT1 transporters expressed in yeast cells. Plant Cell Physiol. 49, 1128–1132.
Banuelos, M. A., Klein, R. D., Alexander-Bowman, S. J., and Rodriguez-Navarro, A. (1995). A potassium transporter of the yeast Schwanniomyces occidentalis homologous to the Kup system of Escherichia coli has a high concentrative capacity. EMBO J. 14, 3021–3027.
Becker, D., Geiger, D., Dunkel, M., Roller, A., Bertl, A., Latz, A., Carpaneto, A., Dietrich, P., Roelfsema, M. R., Voelker, C., Schmidt, D., Mueller-Roeber, B., Czempinski, K., and Hedrich, R. (2004). AtTPK4, an Arabidopsis tandem-pore K+ channel, poised to control the pollen membrane voltage in a pH- and Ca2+-dependent manner. Proc. Natl. Acad. Sci. U.S.A. 101, 15621–15626.
Benito, B., Garciadeblas, B., Fraile-Escanciano, A., and Rodriguez-Navarro, A. (2011). Potassium and sodium uptake systems in fungi. The transporter diversity of Magnaporthe oryzae. Fungal Genet. Biol. 48, 812–822.
Benito, B., Garciadeblas, B., and Rodriguez-Navarro, A. (2012). HAK Transporters from Physcomitrella patens and Yarrowia lipolytica Mediate Sodium Uptake. Plant Cell Physiol. 53, 1117–1123.
Berthomieu, P., Conejero, G., Nublat, A., Brackenbury, W. J., Lambert, C., Savio, C., Uozumi, N., Oiki, S., Yamada, K., Cellier, F., Gosti, F., Simonneau, T., Essah, P. A., Tester, M., Very, A. A., Sentenac, H., and Casse, F. (2003). Functional analysis of AtHKT1 in Arabidopsis shows that Na+ recirculation by the phloem is crucial for salt tolerance. EMBO J. 22, 2004–2014.
Blatt, M. R. (1988). Potassium-dependent bipolar gating of potassium channels in guard cells. J. Membr. Biol. 102, 235–246.
Castresana, J. (2000). Selection of conserved blocks from multiple alignments for their use in phylogenetic analysis. Mol. Biol. Evol. 17, 540–552.
Chanroj, S., Wang, G., Venema, K., Zhang, M. W., Delwiche, C. F., and Sze, H. (2012). Conserved and diversified gene families of monovalent cation/H+ antiporters from algae to flowering plants. Front. Plant Sci. 3:25. doi:10.3389/fpls.2012.00025
Chen, K., Durand, D., and Farach-Colton, M. (2000). NOTUNG: a program for dating gene duplications and optimizing gene family trees. J. Comput. Biol. 7, 429–447.
Corratge-Faillie, C., Jabnoune, M., Zimmermann, S., Very, A. A., Fizames, C., and Sentenac, H. (2010). Potassium and sodium transport in non-animal cells: the Trk/Ktr/HKT transporter family. Cell. Mol. Life Sci. 67, 2511–2532.
Czempinski, K., Frachisse, J. M., Maurel, C., Barbier-Brygoo, H., and Mueller-Roeber, B. (2002). Vacuolar membrane localization of the Arabidopsis “two-pore” K+ channel KCO1. Plant J. 29, 809–820.
Darriba, D., Taboada, G. L., Doallo, R., and Posada, D. (2011). ProtTest 3: fast selection of best-fit models of protein evolution. Bioinformatics 27, 1164–1165.
Dreyer, I., and Blatt, M. R. (2009). What makes a gate? The ins and outs of Kv-like K+ channels in plants. Trends Plant Sci. 14, 383–390.
Dunkel, M., Latz, A., Schumacher, K., Müller, T., Becker, D., and Hedrich, R. (2008). Targeting of vacuolar membrane localized members of the TPK channel family. Mol. Plant 1, 938–949.
Durell, S. R., and Guy, H. R. (1999). Structural models of the KtrB, TrkH, and Trk1,2 symporters based on the structure of the KcsA K(+) channel. Biophys. J. 77, 789–807.
Epstein, E., Rains, D. W., and Elzam, O. E. (1963). Resolution of dual mechanisms of potassium absorption by barley roots. Proc. Natl. Acad. Sci. U.S.A. 49, 684–692.
Fu, H. H., and Luan, S. (1998). AtKuP1: a dual-affinity K+ transporter from Arabidopsis. Plant Cell 10, 63–73.
Garciadeblas, B., Barrero-Gil, J., Benito, B., and Rodriguez-Navarro, A. (2007). Potassium transport systems in the moss Physcomitrella patens: pphak1 plants reveal the complexity of potassium uptake. Plant J. 52, 1080–1093.
Garciadeblas, B., Senn, M. E., Banuelos, M. A., and Rodriguez-Navarro, A. (2003). Sodium transport and HKT transporters: the rice model. Plant J. 34, 788–801.
Gassman, W., Rubio, F., and Schroeder, J. I. (1996). Alkali cation selectivity of the wheat root high-affinity potassium transporter HKT1. Plant J. 10, 869–852.
Gaymard, F., Pilot, G., Lacombe, B., Bouchez, D., Bruneau, D., Boucherez, J., Michaux-Ferriere, N., Thibaud, J. B., and Sentenac, H. (1998). Identification and disruption of a plant shaker-like outward channel involved in K+ release into the xylem sap. Cell 94, 647–655.
Gierth, M., Maser, P., and Schroeder, J. I. (2005). The potassium transporter AtHAK5 functions in K(+) deprivation-induced high-affinity K(+) uptake and AKT1 K(+) channel contribution to K(+) uptake kinetics in Arabidopsis roots. Plant Physiol. 137, 1105–1114.
Grabov, A. (2007). Plant KT/KUP/HAK potassium transporters: single family – multiple functions. Ann. Bot. 99, 1035–1041.
Haro, R., Banuelos, M. A., and Rodriguez-Navarro, A. (2010). High-affinity sodium uptake in land plants. Plant Cell Physiol. 51, 68–79.
Haro, R., Banuelos, M. A., Senn, M. E., Barrero-Gil, J., and Rodriguez-Navarro, A. (2005). HKT1 mediates sodium uniport in roots. Pitfalls in the expression of HKT1 in yeast. Plant Physiol. 139, 1495–1506.
Horie, T., Brodsky, D. E., Costa, A., Kaneko, T., Lo, S. F., Katsuhara, M., and Schroeder, J. I. (2011a). K+ transport by the OsHKT2;4 transporter from rice with atypical Na+ transport properties and competition in permeation of K+ over Mg2+ and Ca2+ ions. Plant Physiol. 156, 1493–1507.
Horie, T., Sugawara, M., Okada, T., Taira, K., Kaothien-Nakayama, P., Katsuhara, M., Shinmyo, A., and Nakayama, H. (2011b). Rice sodium-insensitive potassium transporter, OsHAK5, confers increased salt tolerance in tobacco BY2 cells. J. Biosci. Bioeng. 111, 346–356.
Horie, T., Costa, A., Kim, T. H., Han, M. J., Horie, R., Leung, H. Y., Miyao, A., Hirochika, H., An, G., and Schroeder, J. I. (2007). Rice OsHKT2;1 transporter mediates large Na+ influx component into K+-starved roots for growth. EMBO J. 26, 3003–3014.
Isayenkov, S., Isner, J. C., and Maathuis, F. J. (2011a). Membrane localisation diversity of TPK channels and their physiological role. Plant Signal. Behav. 6, 1201–1204.
Isayenkov, S., Isner, J. C., and Maathuis, F. J. (2011b). Rice two-pore K+ channels are expressed in different types of vacuoles. Plant Cell 23, 756–768.
Jabnoune, M., Espeout, S., Mieulet, D., Fizames, C., Verdeil, J. L., Conejero, G., Rodriguez-Navarro, A., Sentenac, H., Guiderdoni, E., Abdelly, C., and Very, A. A. (2009). Diversity in expression patterns and functional properties in the rice HKT transporter family. Plant Physiol. 150, 1955–1971.
Johansson, I., Wulfetange, K., Poree, F., Michard, E., Gajdanowicz, P., Lacombe, B., Sentenac, H., Thibaud, J. B., Mueller-Roeber, B., Blatt, M. R., and Dreyer, I. (2006). External K modulates the activity of the Arabidopsis potassium channel SKOR via an unusual mechanism. Plant J. 46, 269–281.
Katoh, K., and Toh, H. (2010). Parallelization of the MAFFT multiple sequence alignment program. Bioinformatics 26, 1899–1900.
Kim, E. J., Kwak, J. M., Uozumi, N., and Schroeder, J. I. (1998). AtKUP1: an Arabidopsis gene encoding high-affinity potassium transport activity. Plant Cell 10, 51–62.
Kleffmann, T., Russenberger, D., von, Z. A., Christopher, W., Sjolander, K., Gruissem, W., and Baginsky, S. (2004). The Arabidopsis thaliana chloroplast proteome reveals pathway abundance and novel protein functions. Curr. Biol. 14, 354–362.
Lan, W. Z., Wang, W., Wang, S. M., Li, L. G., Buchanan, B. B., Lin, H. X., Gao, J. P., and Luan, S. (2010). A rice high-affinity potassium transporter (HKT) conceals a calcium-permeable cation channel. Proc. Natl. Acad. Sci. U.S.A. 107, 7089–7094.
Lang, D., Weiche, B., Timmerhaus, G., Richardt, S., Riano-Pachon, D. M., Correa, L. G., Reski, R., Mueller-Roeber, B., and Rensing, S. A. (2010). Genome-wide phylogenetic comparative analysis of plant transcriptional regulation: a timeline of loss, gain, expansion, and correlation with complexity. Genome Biol. Evol. 2, 488–503.
Latorre, R., Morera, F. J., and Zaelzer, C. (2010). Allosteric interactions and the modular nature of the voltage- and Ca2+-activated (BK) channel. J. Physiol. (Lond.) 588, 3141–3148.
Latz, A., Becker, D., Hekman, M., Muller, T., Beyhl, D., Marten, I., Eing, C., Fischer, A., Dunkel, M., Bertl, A., Rapp, U. R., and Hedrich, R. (2007). TPK1, a Ca(2+)-regulated Arabidopsis vacuole two-pore K(+) channel is activated by 14-3-3 proteins. Plant J. 52, 449–459.
Lebaudy, A., Very, A. A., and Sentenac, H. (2007). K+ channel activity in plants: genes, regulations and functions. FEBS Lett. 581, 2357–2366.
Leigh, R. A., and Wyn Jones, R. G. (1984). A hypothesis relating critical potassium concentrations for growth to the distribution and functions of this ion in the plant cell. New Phytol. 97, 1–13.
Maathuis, F. J. (2009). Physiological functions of mineral macronutrients. Curr. Opin. Plant Biol. 12, 250–258.
Maitrejean, M., Wudick, M. M., Voelker, C., Prinsi, B., Mueller-Roeber, B., Czempinski, K., Pedrazzini, E., and Vitale, A. (2011). Assembly and sorting of the tonoplast potassium channel AtTPK1 and its turnover by internalization into the vacuole. Plant Physiol. 156, 1783–1796.
Marcel, D., Muller, T., Hedrich, R., and Geiger, D. (2010). K+ transport characteristics of the plasma membrane tandem-pore channel TPK4 and pore chimeras with its vacuolar homologs. FEBS Lett. 584, 2433–2439.
Maser, P., Eckelman, B., Vaidyanathan, R., Horie, T., Fairbairn, D. J., Kubo, M., Yamagami, M., Yamaguchi, K., Nishimura, M., Uozumi, N., Robertson, W. R., Sussman, M. R., and Schroeder, J. I. (2002a). Altered shoot/root Na+ distribution and bifurcating salt sensitivity in Arabidopsis by genetic disruption of the Na+ transporter AtHKT1. FEBS Lett. 531, 157–161.
Maser, P., Gierth, M., and Schroeder, J. I. (2002b). Molecular mechanisms of potassium and sodium uptake in plants. Plant Soil 247, 43–54.
Maser, P., Hosoo, Y., Goshima, S., Horie, T., Eckelman, B., Yamada, K., Yoshida, K., Bakker, E. P., Shinmyo, A., Oiki, S., Schroeder, J. I., and Uozumi, N. (2002c). Glycine residues in potassium channel-like selectivity filters determine potassium selectivity in four-loop-per-subunit HKT transporters from plants. Proc. Natl. Acad. Sci. U.S.A. 99, 6428–6433.
Maser, P., Thomine, S., Schroeder, J. I., Ward, J. M., Hirschi, K., Sze, H., Talke, I. N., Amtmann, A., Maathuis, F. J., Sanders, D., Harper, J. F., Tchieu, J., Gribskov, M., Persans, M. W., Salt, D. E., Kim, S. A., and Guerinot, M. L. (2001). Phylogenetic relationships within cation transporter families of Arabidopsis. Plant Physiol. 126, 1646–1667.
Oomen, R. J., Benito, B., Sentenac, H., Rodriguez-Navarro, A., Talon, M., Very, A. A., and Domingo, C. (2012). HKT2;2/1, a K+-permeable transporter identified in a salt tolerant rice cultivar through surveys of natural genetic polymorphism. Plant J. doi:10.1111/j.1365-313X.2012.05031.x
Peltier, J. B., Ytterberg, A. J., Sun, Q., and van Wijk, K. J. (2004). New functions of the thylakoid membrane proteome of Arabidopsis thaliana revealed by a simple, fast, and versatile fractionation strategy. J. Biol. Chem. 279, 49367–49383.
Platten, J. D., Cotsaftis, O., Berthomieu, P., Bohnert, H., Davenport, R. J., Fairbairn, D. J., Horie, T., Leigh, R. A., Lin, H. X., Luan, S., Maser, P., Pantoja, O., Rodriguez-Navarro, A., Schachtman, D. P., Schroeder, J. I., Sentenac, H., Uozumi, N., Very, A. A., Zhu, J. K., Dennis, E. S., and Tester, M. (2006). Nomenclature for HKT transporters, key determinants of plant salinity tolerance. Trends Plant Sci. 11, 372–374.
Qi, Z., Hampton, C. R., Shin, R., Barkla, B. J., White, P. J., and Schachtman, D. P. (2008). The high affinity K+ transporter AtHAK5 plays a physiological role in planta at very low K+ concentrations and provides a caesium uptake pathway in Arabidopsis. J. Exp. Bot. 59, 595–607.
Quintero, F. J., and Blatt, M. R. (1997). A new family of K+ transporters from Arabidopsis that are conserved across phyla. FEBS Lett. 415, 206–211.
Rice, P., Longden, I., and Bleasby, A. (2000). EMBOSS: the European Molecular Biology Open Software Suite. Trends Genet. 16, 276–277.
Rodriguez-Navarro, A., and Rubio, F. (2006). High-affinity potassium and sodium transport systems in plants. J. Exp. Bot. 57, 1149–1160.
Rubio, F., Gassmann, W., and Schroeder, J. I. (1995). Sodium-driven potassium uptake by the plant potassium transporter HKT1 and mutations conferring salt tolerance. Science 270, 1660–1663.
Rubio, F., Santa-Maria, G. E., and Rodriguez-Navarro, A. (2000). Cloning of Arabidopsis and barley cDNAs encoding HAK potassium transporters in root and shoot cells. Physiol. Plant. 109, 34–43.
Rus, A., Yokoi, S., Sharkhuu, A., Reddy, M., Lee, B. H., Matsumoto, T. K., Koiwa, H., Zhu, J. K., Bressan, R. A., and Hasegawa, P. M. (2001). AtHKT1 is a salt tolerance determinant that controls Na+ entry into plant roots. Proc. Natl. Acad. Sci. U.S.A. 98, 14150–14155.
Santa-Maria, G. E., Rubio, F., Dubcovsky, J., and Rodriguez-Navarro, A. (1997). The HAK1 gene of barley is a member of a large gene family and encodes a high-affinity potassium transporter. Plant Cell 9, 2281–2289.
Schleyer, M., and Bakker, E. P. (1993). Nucleotide sequence and 3′-end deletion studies indicate that the K+-uptake protein kup from Escherichia coli is composed of a hydrophobic core linked to a large and partially essential hydrophilic C terminus. J. Bacteriol. 175, 6925–6931.
Schroeder, J. I. (1988). K+ transport properties of K+ channels in the plasma membrane of Vicia faba guard cells. J. Gen. Physiol. 92, 667–683.
Spalding, E. P., Hirsch, R. E., Lewis, D. R., Qi, Z., Sussman, M. R., and Lewis, B. D. (1999). Potassium uptake supporting plant growth in the absence of AKT1 channel activity: Inhibition by ammonium and stimulation by sodium. J. Gen. Physiol. 113, 909–918.
Stamatakis, A. (2006). RAxML-VI-HPC: maximum likelihood-based phylogenetic analyses with thousands of taxa and mixed models. Bioinformatics 22, 2688–2690.
Sunarpi, Horie, T., Motoda, J., Kubo, M., Yang, H., Yoda, K., Horie, R., Chan, W. Y., Leung, H. Y., Hattori, K., Konomi, M., Osumi, M., Yamagami, M., Schroeder, J. I., and Uozumi, N. (2005). Enhanced salt tolerance mediated by AtHKT1 transporter-induced Na unloading from xylem vessels to xylem parenchyma cells. Plant J. 44, 928–938.
Szczerba, M. W., Britto, D. T., and Kronzucker, H. J. (2009). K+ transport in plants: physiology and molecular biology. J. Plant Physiol. 166, 447–466.
Uozumi, N., Kim, E. J., Rubio, F., Yamaguchi, T., Muto, S., Tsuboi, A., Bakker, E. P., Nakamura, T., and Schroeder, J. I. (2000). The Arabidopsis HKT1 gene homolog mediates inward Na+ currents in Xenopus laevis oocytes and Na+ uptake in Saccharomyces cerevisiae. Plant Physiol. 122, 1249–1259.
Vernot, B., Stolzer, M., Goldman, A., and Durand, D. (2007). Reconciliation with non-binary species trees. Comput. Syst. Bioinformatics Conf. 6, 441–452.
Véry, A. A., and Sentenac, H. (2002). Cation channels in the Arabidopsis plasma membrane. Trends Plant Sci. 7, 168–175.
Véry, A. A., and Sentenac, H. (2003). Molecular mechanisms and regulation of K+ transport in higher plants. Annu. Rev. Plant Biol. 54, 575–603.
Voelker, C., Gomez-Porras, J. L., Becker, D., Hamamoto, S., Uozumi, N., Gambale, F., Mueller-Roeber, B., Czempinski, K., and Dreyer, I. (2010). Roles of tandem-pore K+ channels in plants – a puzzle still to be solved. Plant Biol. (Stuttg.) 12(Suppl. 1), 56–63.
Voelker, C., Schmidt, D., Mueller-Roeber, B., and Czempinski, K. (2006). Members of the Arabidopsis AtTPK/KCO family form homomeric vacuolar channels in planta. Plant J. 48, 296–306.
Wegner, L. H., and de Boer, A. H. (1997). Properties of two outward-rectifying channels in root xylem parenchyma cells suggest a role in K+ homeostasis and long-distance signaling. Plant Physiol. 115, 1707–1719.
Keywords: potassium, transport, channel, voltage-dependent, voltage-independent, high-affinity, Selaginella
Citation: Gomez-Porras JL, Riaño-Pachón DM, Benito B, Haro R, Sklodowski K, Rodríguez-Navarro A and Dreyer I (2012) Phylogenetic analysis of K+ transporters in bryophytes, lycophytes, and flowering plants indicates a specialization of vascular plants. Front. Plant Sci. 3:167. doi: 10.3389/fpls.2012.00167
Received: 23 April 2012; Accepted: 05 July 2012;
Published online: 02 August 2012.
Edited by:
Tomoaki Nishiyama, Kanazawa University, JapanReviewed by:
Moritz Karl Nowack, Flanders Institute for Biotechnology, BelgiumCopyright: © 2012 Gomez-Porras, Riaño-Pachón, Benito, Haro, Sklodowski, Rodríguez-Navarro and Dreyer. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits use, distribution and reproduction in other forums, provided the original authors and source are credited and subject to any copyright notices concerning any third-party graphics etc.
*Correspondence: Ingo Dreyer, Plant Biophysics, Centro de Biotecnología y Genómica de Plantas, Universidad Politécnica de Madrid, Campus de Montegancedo, Carretera M-40, km 37.7, E-28223-Pozuelo de Alarcón (Madrid), Spain. e-mail:aW5nby5kcmV5ZXJAdXBtLmVz
†Judith Lucia Gomez-Porras and Diego Mauricio Riaño-Pachón and Begoña Benito and Rosario Haro have contributed equally to this work.
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.