 Meizhi Wang
Meizhi Wang Hongmei Li
Hongmei Li Wei Zhang4
Wei Zhang4 Fuyan Zhuo
Fuyan Zhuo Aihuan Zhang
Aihuan Zhang
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Physiol., 07 March 2023
Sec. Invertebrate Physiology
Volume 14 - 2023 | https://doi.org/10.3389/fphys.2023.1136559
This article is part of the Research TopicThe Strong Adaptability of Insects Shaped by PlasticityView all 13 articles
Research on the ovarian development of insect pests helps provide key information for predicting pest occurrences, and currently, there is very limited information about the reproductive system of Ceracris kiangsu Tsai. This study aimed to assess the reproductive fitness of 321 adult female insects by using traditional methods to dissect female adults, measure female ovaries, and assess the process of egg formation. The phenotypic traits including body weight and body length were also measured and used to estimate the model of ovarian developmental stages. Four ovarian developmental stages before the oviposition were identified, and the fundamental ovarian structure of C. kiangsu displayed red dots on the matured eggs inside the calyx at ovarian developmental stage V. The accessory glands of C. kiangsu had the deepest folds at stage Ⅲ. Redundancy analysis (RDA) was used to explore the correlation between ovarian development, body weight, and body length. A significant positive correlation was observed for body weight (p = 0.001) and body length (p = 0.009), which varied with the grade of ovarian development evaluated by the ovarian developmental stage, ovarian length, ovarian width, and ovarian cross-sectional area. A partial least square (PLS) regression was used to model the ovarian developmental stage, with a stage-based PLS being identified as the more effective method, which was y = 1.509x1 + 0.114x2. The model provides a potentially rapid way to identify the population source as either “native” or “immigrant” from the phenotypic traits without dissection. The aforementioned model may be used to estimate adult emergence periods and identify migratory populations from their ovarian development, potentially aiding in implementing proper prevention measures.
The monitoring and prediction of insect pests is fundamental to conducting effective pest management. Current measures and approaches for the prediction of pest occurrences include, but is not limited to, light traps, sex pheromone traps, and field monitoring. Another technique that has been used to predict pest population dynamics and occurrences in migrating species is the dissection of adult female insects (Qi et al., 2011). This technique has been shown to provide information on not only the ovarian development but also the peak oviposition period, which can then be used to estimate the reproductive capacity of the population and the migration potential of the pest (Liu and Huang, 2018). Research studies have been conducted on ovarian structures (Hodge, 1943; Liu and Lu, 1959; Snodgrass, 1993), ovarian developmental stages (Wu and Zhang, 1997; Chen et al., 2005; Ren et al., 2014; Zhang et al., 2020), and the effects of insect hormones and miRNAs on ovarian development. A good example of this is the juvenile hormones of Locusta migratoria manilensis (Meyen) in which Let-7 and miRNA-278 have been shown to play a vital role in regulating its reproductive system (Song et al., 2013; Song et al., 2019). However, external factors have also been shown to affect ovarian development; Bhalerao et al. (1987) found that the feeding reduction had a negative effect on the vitellogenesis of Euscelimena harpago (Serville) and Potua sabulosa Hancock female locusts’ ovaries. Chen et al. (2008) found that the host plants had a positive effect on ovarian development and fecundity of Fruhstorferiola tonkinensis Willemse. Additionally, mating has also been shown to exert a positive effect on the developmental phenotype of Eriopis connexa (Germar) offspring and on the increase in the proportion of mature eggs in female Aedes taeniorhynchus (Wiedemann). Mating was also found to accelerate both ovarian development and reproduction abilities of female Apolygus lucorum (Meyer-Dür) (Meara and Evans, 1977; Colares et al., 2015; Li et al., 2017). Whilst the extreme temperature and short photoperiod reduced the fecundity of Drosophila suzukii (Matsumura) and Athetis lepigone Möschler (Everman et al., 2018; Liang et al., 2021), the ovarian development of Spodoptera mauritia (Boisduval) has been retarded after being treated with fenoxycarb (Banu and Manogem, 2022). In essence, the study of ovarian development and the effect of biological factors and abiotic factors has become a core component of research around pest prediction and reproductive regulation.
Despite much research on insect ovarian development, relatively little is known about the reproduction system development of the yellow-spined bamboo locust, Ceracris kiangsu Tsai (Orthoptera: Arcypteridae). This locust has historically been regarded as the second worst pest species affecting Phyllostachys heterocycla (Carr.) and Indocalamus tessellatus (Munro). C. kiangsu Tsai has been recorded in East Asian and Southeast Asian countries, such as China, Laos, and Vietnam, with the scales and frequency of C. kiangsu population outbreaks increasing gradually, attacking agricultural crops including Zea mays L., Sorghum bicolor (L.), Oryza sativa L., and Musa basjoo Siebold (Liu et al., 2021; Li et al., 2022). Since 2014, there have been a large number of C. kiangsu outbreaks in Laos and subsequent outbreaks from Vietnam to South China in 2019 (Zhuo et al., 2020). Liu et al. (2021) revealed that during this immigration, C. kiangsu had characteristics similar to Schistocerca gregaria (Forskål), whose recurrent outbreaks have affected farming systems throughout Northern Africa. Currently, high-resolution fully polarimetric insect radars are being used to accurately assess and forecast the migration patterns of C. kiangsu; however, this is a costly process and equipment-intensive management strategy. This study aims to investigate the potential of using ovarian development monitoring in order to predict the reproduction ability and the population dynamics of C. kiangsu.
To fully understand the ovarian structure and stages of C. kiangsu, adult female insects were dissected during different developmental stages after emergence. These adult female insects were collected from both the laboratory and field populations. The rearing temperatures were kept constant (30°C), while the field population was kept at a variable temperature. The laboratory-rearing population may provide relative complete series development information of adult female insects. Meanwhile, adult female insects from the field population may ensure the enough dissection replication as the beneficial complement. Hence, there may be differences in ovarian development between the two populations. Moreover, if we just dissected the field population in this study, the whole ovarian developmental process may be missed. To rapidly get the immigration information of S. frugiperda, the ovarian length and width were used to predict the ovarian developmental stages (Zhao et al., 2019). Therefore, it is vital to infer the relationship between the phenotypic traits and the stages of oviposition and pre-oviposition. Thus, the information will contribute to easily mastering phenotypic fitness estimating the ovarian development to further provide the migratory and prediction information of C. kiangsu.
C. kiangsu adult female insects were collected from both the laboratory population and natural bamboo forests. The laboratory population was reared in the Institute of Plant Protection, Chinese Academy of Agricultural Sciences (IPPCAAS), Beijing, China. The egg pods were collected from a natural bamboo habitat, Anhua County, Hunan Province, China (28°62′N; 111°35′E), and then placed in vermiculite. Using the method of Fang et al. (2022), the egg pods were incubated in a chamber (MGC-1000HP-2, Shanghai Yiheng Scientific Instruments Co., Ltd., Shanghai, China) at 30°C ± 1°C and 65 ± 5% RH under a 16:8 h L:D photoperiod. Nymphs were transferred gently into nylon mesh cages (35 cm × 35 cm × 35 cm) with a maximum of approximately 300 nymphs each. Then, one female and two male adults were transferred from the aforementioned nylon mesh cages to a plastic cylinder (φ = 9 cm; height = 17.8 cm) covered with a nylon mesh for air ventilation. The nymphs and adults were reared under the same condition as the eggs’ hatch and were fed with enough fresh wheat and rice leaves.
The field population of C. kiangsu was collected from a bamboo forest in Cha’anpu Town, Taoyuan County, Hunan Province, China (28°42′N; 111°12′E), in late July and early August 2022.
For the laboratory population, after adult emergence, approximately 1–3 healthy female adults were measured and dissected every 3 days until all female adults were dead (the 81st day after adult emergence). However, for the field population, approximately 10–30 healthy female adults were measured and dissected every day from 27 July 2022 to 6 August 2022. The peak data on adult emergence were recorded in advance. In total, 51 and 270 female adults were dissected from the laboratory and field populations, respectively.
The dissection process began with the measurement of the body length and body weight of the female adults by using electronic Vernier calipers (0–150 mm, Shanghai Shenhan Measuring Tools Co., Ltd., Shanghai, China) and the electronic balance (PL203, Mettler-Toledo Instruments Shanghai Co., Ltd., Shanghai, China).
Then, the adult female insects were dissected according to Han et al. (2018). The wings and legs were removed, and the dead locust was placed on a dissecting tray with wax and fixed by means of needles through the thorax. An incision was then made on the middle of sternites using surgical scissors. Approximately 6–8 insect needles were placed through the exoskeleton on the left and right sides of the body and into the wax. Care was taken not to damage the internal organs. Finally, the target organ, ovary, was identified under a stereomicroscope (SZ2-ILST, Olympus Co., Tokyo, Japan) and carefully placed in a Petri dish with saline.
The ovaries were photographed using a super-depth-of-field three-dimensional microscopy system (VHX-2000), and the length, width, and cross-sectional area of the ovaries were also measured. The fundamental ovarian structure was similar to that shown in Liu and Lu (1959), and the ovarian development classification methods were according to Han et al. (2018).
The phenotypic traits and ovarian developmental parameters were evaluated using a one-way ANOVA (Duncan’s test; p < 0.05; SPSS 22.0).
Then, detrended correspondence analysis (DCA; Hill and Gauch, 1980) was used to select the proper constrained ordination method [e.g., redundancy analysis (RDA) and canonical correspondence analysis (CCA)]. Once the axil length of the dataset after DCA is less than 2, RDA can be selected for further analyses according to Nasser et al. (2019). RDA and Pearson’s correlation analysis were used to assess and quantify the relationship between the development duration, days after emergence, body length, body weight of female adults, and ovarian development for the optimization of variables. The variance inflation factor (VIF) value indicates the collinearity among every two independent variables. Four independent variables in this study were less than 10, and independent variables with p < 0.05 were regarded as significantly contributed to the variance in the process of ovarian development (Grech et al., 2019).
The partial least squares (PLS; Wold, 1975) regression is a powerful method that can be used to extract a few latent variables with information content, which explain as much of the covariance as possible between the dependent variables and independent variables. The optimal number of latent variables (ONLVs) was determined according to the lowest value of the root mean square error of the cross-validation (RMSECV) via the leave-one-out cross-validation (LOOCV) method, to avoid overfitting or underfitting. The coefficient of multiple determinations (R2), the root mean squared error of prediction (RMSEP), and the variance explained (%) and variable importance in the projection (VIP) were used to evaluate PLS models. The higher R2, variance explained, and VIP means the PLS model fits better; among them, the threshold score of a VIP is 1.0 (Porker et al., 2020; Wise and Dolan, 2020).
RDA (Oksanen et al., 2022), Pearson’s correlation analysis (R core team, 2022), and PLS (Liland et al., 2022) analysis were carried out using R 4.1.3.
In total, 321 adult female insects were dissected over the course of this study. Figure 1 illustrates the development of ovaries with increasing female age. Ovaries shown in Figure 1 were all collected from the laboratory population.
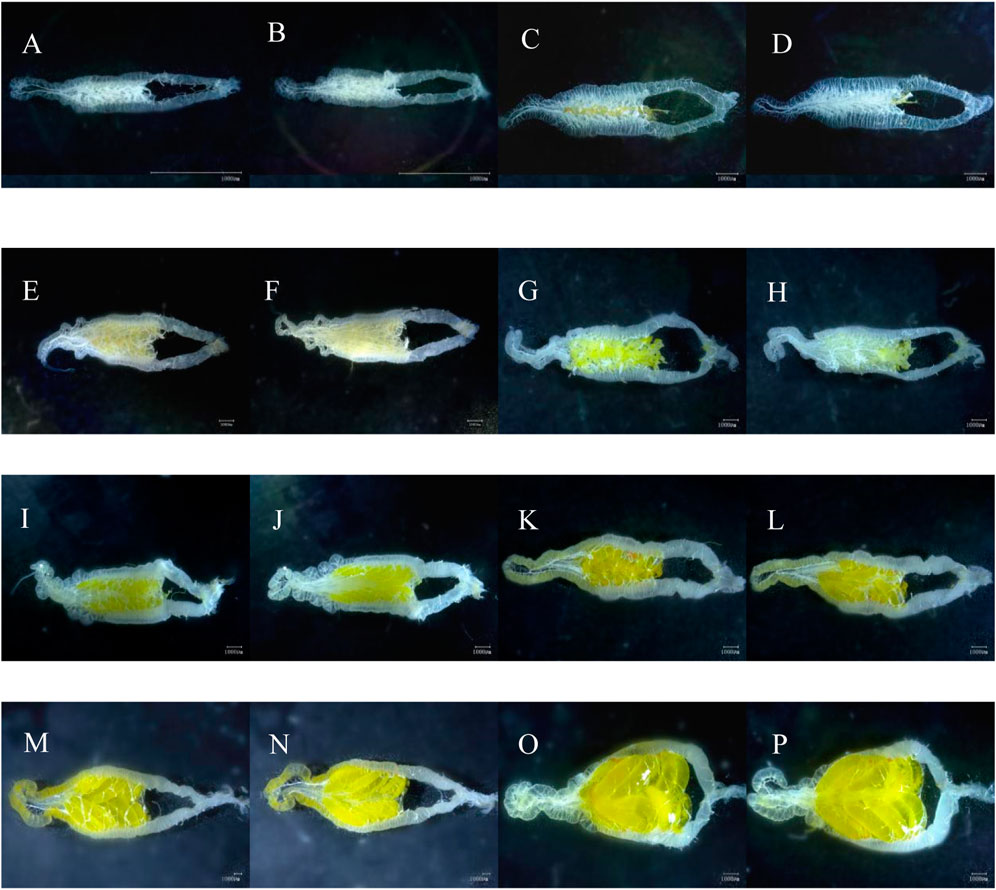
FIGURE 1. Visual view of C. kiangsu ovarian morphology. (A) Ventral view at stage Ⅰ, (B) dorsal view at stage Ⅰ, (C) ventral view at early-stage Ⅱ, (D) dorsal view at early-stage Ⅱ, (E) ventral view at end-stage Ⅱ, (F) dorsal view at end-stage Ⅱ, (G) ventral view at early-stage Ⅲ, (H) dorsal view at early-stage Ⅲ, (I) ventral view at end-stage Ⅲ, (J) dorsal view at end-stage Ⅲ, (K) ventral view at early-stage Ⅳ, (L) dorsal view at early-stage Ⅳ, (M) ventral view at end-stage Ⅳ, (N) dorsal view at end-stage Ⅳ, (O) ventral view at stage Ⅴ, and (P) dorsal view at stage Ⅴ (scale bars = 1,000 μm).
The ovarian development was divided into five stages according to characteristics of ovarioles and oviducts, accessory gland, the number of yolks and eggs, and body fat (Figure 1; Table 1). The main characteristic of ovarian developmental stage I was transparent and thin ovarioles; Figures 1A, B show an ovary from a three-day-old female adult. The main characteristic of ovarian developmental stage II was the appearance of the pale-yellow fat body and yolk, which gradually increased; Figures 1C, D and Figures 1E, F show ovaries from 12-day-old and 18-day-old female adults, respectively. At ovarian developmental stage III, the shape and number of immature eggs appeared and increased in the ovary gradually; Figures 1G, H and Figures 1I, J show ovaries from 27-day-old and 36-day-old female adults, respectively. The main characteristics of stage Ⅳ were the proper arrangement of eggs in the calyx and thickening of the accessory gland. The abdominal morphology showed that the intersegmental membranes between the fifth and sixth segments were folded (Figure 2A); Figures 1K–N show ovaries from 27-day-old and 36-day-old female adults, respectively. The eggs matured, the egg membranes were transparent, and dark red spots appeared in the calyx at stage Ⅴ; Figures 1O, P show the ovary from an 81-day-old female adult. There was an ovipositor located between the eighth and ninth segments. At stage Ⅴ, the valvulae protruded from the end of the abdomen, and the epiproct lifted up (Figure 2B).
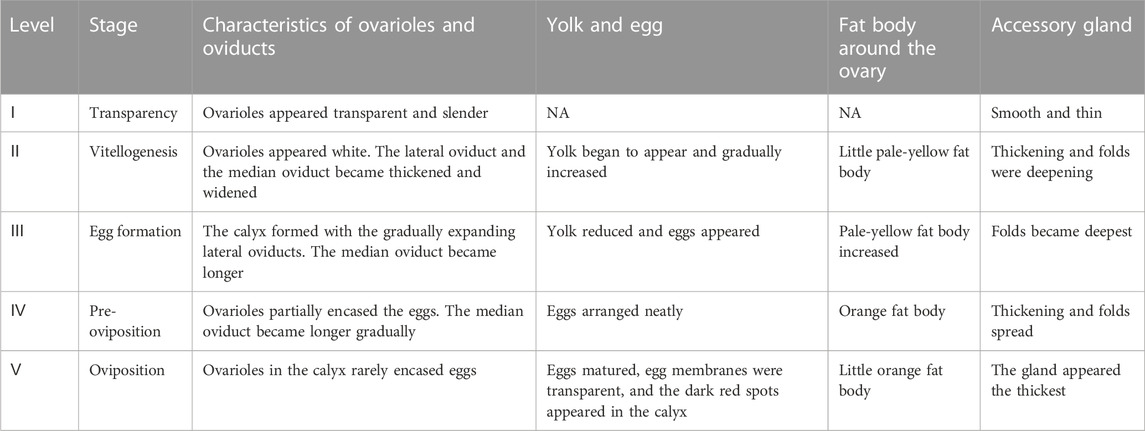
TABLE 1. Ovarian development characteristics of C. kiangsu at different stage standards.

FIGURE 2. Phenotypic morphology of the C. kiangsu abdomen at (A) stage Ⅳ and (B) stage Ⅴ.
The C. kiangsu adult female essential reproductive system (Figure 3) consisted of a pair of ovaries. There was a pair of accessory glands connected to a pair of calyxes and lateral oviducts, and the oviducts jointly form the median oviduct, which led to the vagina. The spermatheca connected with the median oviduct and vagina by the spermatheca duct. There was a suspensory ligament located in the anterior end of the accessory gland, and the suspensory ligament was generally thin and transparent and not shown in Figure 3.
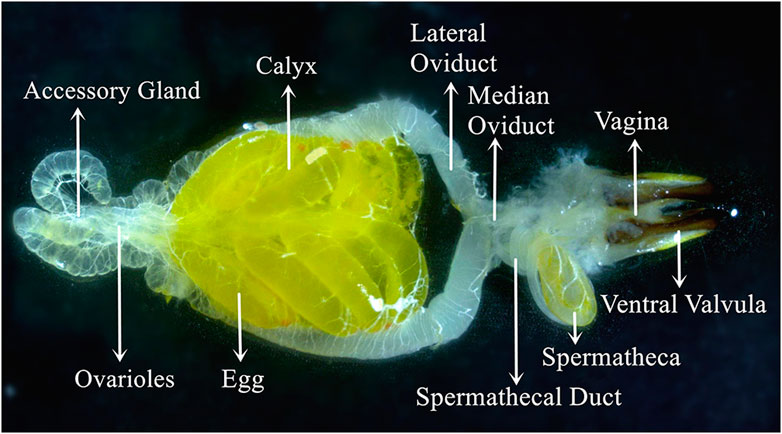
FIGURE 3. Visual view of the C. kiangsu ovary at stage Ⅴ.
The analysis indicated that there was an increasing tendency of both ovarian and phenotype characteristics with the progression of ovarian development stages as C. kiangsu matured (Figure 4). The ovarian width (7.90 ± 0.47 mm) and cross-sectional area (92.25 ± 10.24 mm2) at ovarian development stage V were significantly wider and bigger than those at the other four stages, respectively, but there was no significant difference found among the ovarian lengths of ovarian development stages from II to IV. At stage V, the ovarian length was significantly longer than that at stage I (18.95 ± 1.72 mm versus 13.47 ± 1.56 mm).
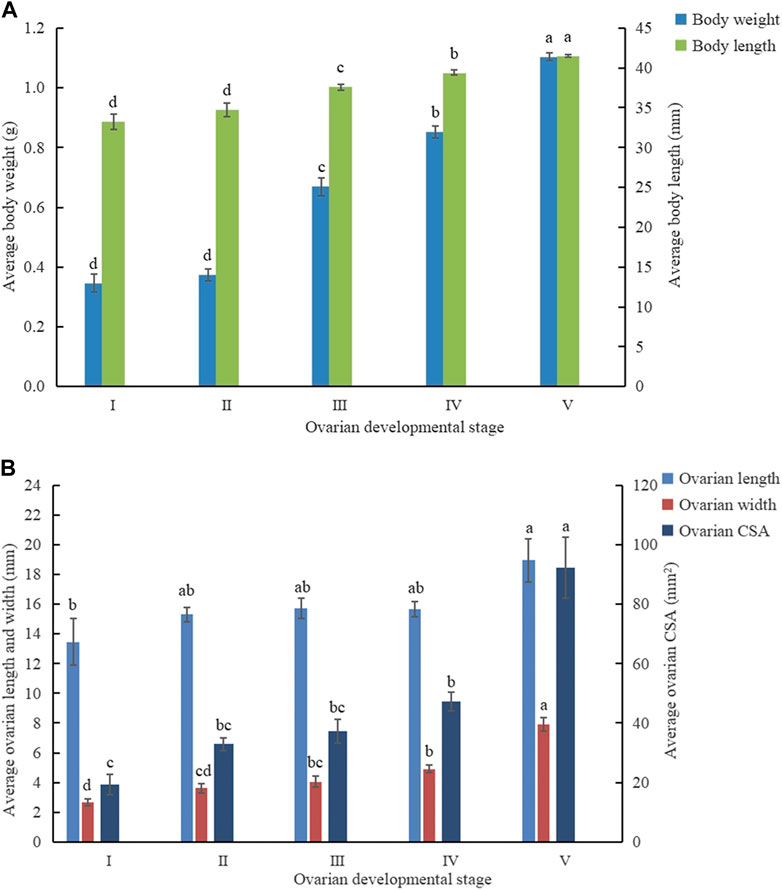
FIGURE 4. Mean value (±SE) at different ovarian developmental stages of (A) phenotypic traits and (B) ovarian developmental parameters. CSA, cross-sectional area.
Similarly, the average body weight and average body length increased proportionally to the ovarian development stage. The average body weight (1.11 ± 0.01 g) and body length (41.49 ± 0.15 mm) at stage V was significantly heavier and longer than those at the other four stages, respectively.
Pearson’s correlation coefficient analysis and RDA were used to investigate the relationship between body weight and body length during the ovarian development of C. kiangsu. Four ovarian developmental parameters were used to evaluate the degree of ovarian development in RDA (stage, ovarian length, ovarian width, and ovarian cross-sectional area).
Pearson’s correlation coefficient matrix (Figure 5) showed that body weight and body length were significantly positively correlated with every ovarian development parameter. Furthermore, RDA (Table 2; Figure 6) showed that both body length and body weight were significantly positively correlated with the degree of ovarian development.
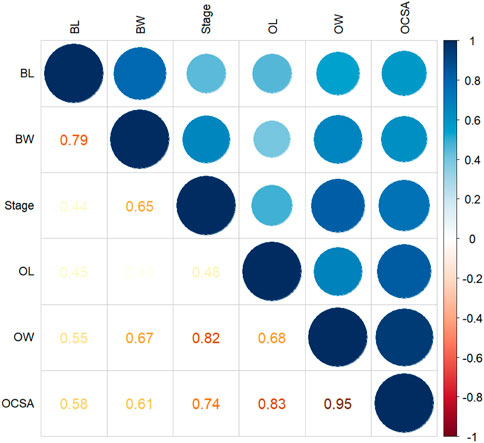
FIGURE 5. Correlation matrix of all variables. The larger the absolute value and the darker the color, the stronger the Pearson’s correlation (positive or negative). Circles in different colors represent significant correlation; p < 0.05. BL, body length; BW, body weight; OL, ovarian length; OW, ovarian width; OCSA, ovarian cross-sectional area.
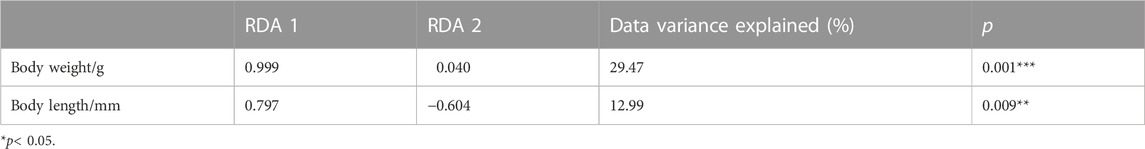
TABLE 2. Forward selection results with the test of variable significance.
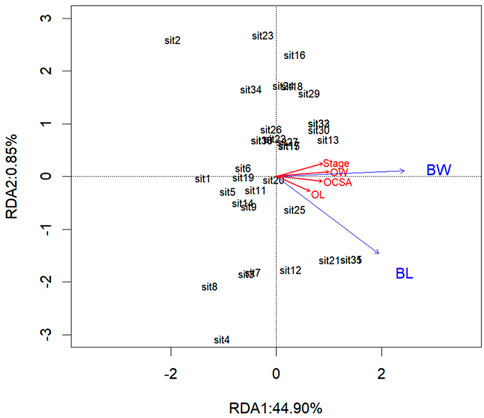
FIGURE 6. Redundancy analysis diagram of correlations between independent variables and the ovarian developmental parameters. The ovarian developmental parameters are indicated by red color arrows: OL, ovarian length; OW, ovarian width; OCSA, ovarian cross-sectional area. The independent variables are indicated by blue color arrows: BL, body length; BW, body weight.
RDA showed that body weight and body length accounted for 45.75% of the total variance in the degree of ovarian development, suggesting that the two independent variables had a significant impact on the degree of ovarian development, with Monte Carlo permutation tests with 999 unrestricted permutations (Table 2). The interpretation of the two axes was 44.90% and 0.85%, respectively (Table 3). Therefore, the two axes were selected as the main component axis (Tables 2, 3; Figure 6). From the two independent variables analyzed, the percentage of the total variance explained was the highest for the body weight with 29.47%, which disclosed that body weight had a better explanation of ovarian development than body length (Table 2; Figure 6).

TABLE 3. Summary of RDA for trapped samples.
All four ovarian developmental parameters were used to model dependent variables. The ONLV first determined the number of latent variables according to the lowest value of the RMSECV. The first latent variable was selected and then fitted in PLS models again. The optimal models based on each dependent variable and their visualization are shown in Figure 7. The latent variable elucidated 92.41% of the stage-based PLS model. R2, RMSEP, and VIP of the stage-based PLS model were 0.56, 0.67, and 1.00, respectively, which suggested that the stage-based PLS model fitted better (Figure 7).
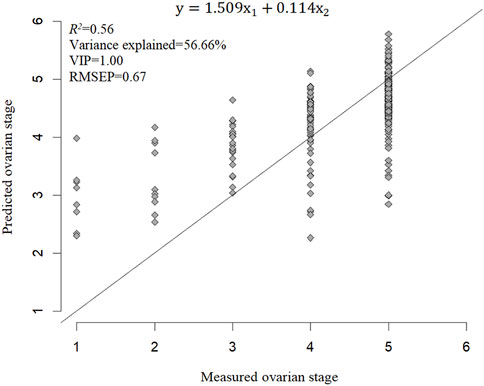
FIGURE 7. PLS prediction of ovarian developmental parameters for Stage. In this model, y was stage, x1, x2 were body weight, body length, respectively.
Furthermore, the VIP of ovarian length-based, ovarian width-based, and ovarian cross-sectional area-based PLS models were all less than 1. Body weight and body length accounted for 56.66% of the total variance explained. The VIP of the stage-based PLS model was greater than 1, which suggests that this PLS model has a relative application in stages’ prediction in the ovarian development of C. kiangsu. The PLS model was subjected to statistical validation tests, and the SE, t-value, and p-value of the PLS model are shown in Table 4, which revealed that body weight had a significant effect on the ovarian developmental stage, but there was no significant effect found among body lengths of ovarian development stages by modeling.
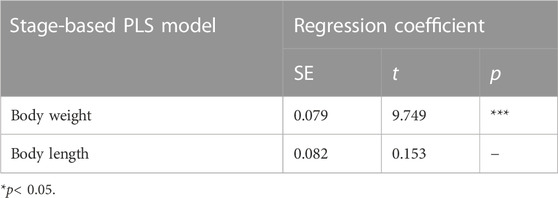
TABLE 4. PLS regression coefficient significance test.
Ovarian development significantly influences the reproduction and expansion capabilities of insect pest populations, and it should be considered when developing sustainable pest management strategies, especially for migratory species (Zhang et al., 2016; Roy et al., 2018). For example, when Masaaki and Seiji (2009) disclosed that the population of L. m. manilensis overwintered as adults or eggs by observing the ovarian development on Iheya Island, Japan, it was then this overwinter behavior that provided essential information to better control this population. Similarly, when Elamin et al. (2014) revealed that the ovarian developmental stages and the subsequent fecundity of Oedaleus senegalensis (Krauss) depended upon the number and function of the ovarioles, this helped improve the pest management operation in North Kordofan State, Sudan. Whilst this study focused on the ovarian development and grade of C. kiangsu, it may also help fill the gap by improving basic ovarian development information and thus the pest management strategies for this species. The relationship between the phenotypic traits and ovarian grade can shed light on the population development of C. kiangsu.
Overall, the ovarian structure of C. kiangsu can be described as similar to Omocestus viridulus (L.), Euchorthippus unicolor (Ikonn.), Euchorthippus vittatus Zheng, etc. (Zhao and Xi, 2005). With five ovarian developmental stages, C. kiangsu was found to be similar to O. decorus asiaticus (Bey-Bienko). The ovarian developmental duration of C. kiangsu was longer than both gregarious and solitary phases of O. decorus asiaticus at stages Ⅲ, Ⅳ, and Ⅴ (Han et al., 2018). The changing trend of the accessory glands was consistent between C. kiangsu and Calliptamus italicus (L.), whilst the accessory glands folded from stage Ⅱ to stage Ⅳ, and the folds became deepest at stage Ⅲ (Ren et al., 2014). However, there were unique ovarian features specific to C. kiangsu that set it apart from other species. The median oviduct of C. kiangsu did not fold during ovarian development, which was different from O. viridulus, E. unicolor, and E. vittatus (Zhao and Xi, 2005). There were also dark red dots observed on the matured eggs inside the calyx at stage Ⅴ of a C. kiangsu ovary, which is not observed in the other species.
In this study, both the ovarian length and width were positively correlated with the ovarian developmental stages, which is similar to Tomicus yunnanensis Kirkendall & Faccoli (Liu et al., 2019). In contrast, during the process of ovarian development, the ovarian length of Telenomus theophilae Wu et Chen, L. m. manilensis, and Bactrocera cucurbitae (Coquillett) increased first and then reduced after they had finished laying eggs (Liu et al., 2006; Ouyang et al., 2014; Kwak et al., 2021). After stage Ⅳ, the body length rapidly increased with the ovarian development for the C. kiangsu adult female insects, as the ovipositor reached out from the abdomen. Furthermore, there was a positive correlation between the body weight and ovarian developmental stage, due to both the yolk continually being transformed into eggs and the increased number of eggs during stages Ⅱ–Ⅴ, and this change trend of phenotypic traits on ovarian development was in consistent with Oxya chinensis Thunberg and O. japonica Thunberg (Wu and Zhang, 1997). The RDA results showed that body weight and body length accounted for 45.75% of the total variance explained in ovarian development, with each factor significantly positively correlating with the ovarian development of C. kiangsu (p < 0.05). A similar trend has been observed in Bombus terrestris L. workers, in which the body weight positively correlated with the ovarian development. (Gosterit et al., 2016).
Generally, during the development of migrating insects, they have been shown to exhibit a delayed ovarian development during migration, which has been classified as reproductive diapause (Qi et al., 2011). However, once they arrived at their destination, the ovaries continued to develop, allowing for successful reproduction (Johnson, 1963). Thus, exploring the ovarian development and mating behaviors of migratory pests has become important in discovering their potential source areas and migratory paths. In order to better implement pest management strategies for C. kiangsu, a rapid method to obtain the ovarian developmental stage in the field is fundamental. Previously, PLS was used to predict either continuous or discrete variables (Elsherif et al., 2018); however, it can also be applied to pest management. For example, Xu et al. (2020) used this approach to produce a predictive diagnostic model for cotton aphids based on leaf textural features; the predictive accuracy of up to 91.49% was obtained for this model. In this study, in order to rapidly obtain the degree of ovarian development of C. kiangsu without dissection, body weight and body length were used as predictors. The model parameter standards showed that stage-based PLS modeling was the preferred approach, which can be used in a predictive capacity, and thus, it could potentially be a reference model to identify whether the field populations are “native” or “migratory.”
The ovarian development process is highly dynamic (Papaj, 2000). Berger et al. (2008) found that a heavier female insect usually had a higher potential fecundity, and different sizes of female insects indicated different allocation trade-offs between the self-condition and their egg production. However, this phenomenon is found in reproductive systems of not only female insects but also male insects (Leopold, 1976). Chen et al. (2019) showed that the mating rate in the current generation can be reduced through sex pheromone-trapped male adults, which is then reflected by the size of the Spodoptera litura male testis. Hence, the observation of the reproductive system of migratory insects is a determining factor in determining the origin and degree of development of migratory insects. The studies can provide information on integrated pest management (IPM) of migrants, but these aforementioned studies of C. kiangsu were much less explored.
This study identified the ovarian development process of C. kiangsu adult female insects. By grading the ovarian developmental stages, it is easier to identify the transition and critical time between different stages, thus improving the prediction accuracy of the population development of C. kiangsu. Moreover, it could be helpful in the study of migratory pathways and predicting the occurrence of pests. It also provides additional information about the further exploration of C. kiangsu reproductive systems.
The raw data supporting the conclusion of this article will be made available by the authors, without undue reservation.
All authors contributed to the manuscript and its final submission. MW and WZ contributed to the field data collection. MW and FZ analyzed the data. MW, HL, and TL contributed to the writing of the manuscript. HL and AZ contributed to the conception and field work design. AL edited and improved the manuscript language.
The authors acknowledge the financial support from China’s donation to the CABI Development Fund (IVM10051) and the National Key R and D Program of China (2021YFE0194800).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors, and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Banu, C. A., and Manogem, E. M. (2022). Development and characterization of Spodoptera mauritia ovarian primary cell culture and evaluation of fenoxycarb toxicity. Vitro Cell. Dev.-An. 58, 788–797. doi:10.1007/S11626-022-00728-0
Berger, D., Walters, R., and Gotthard, K. (2008). What limits insect fecundity? Body size- and temperature-dependent egg maturation and oviposition in a butterfly. Funct. Ecol. 22, 523–529. doi:10.1111/J.1365-2435.2008.01392.X
Bhalerao, A. M., Naidu, N. M., and Paranjape, S. Y. (1987). Some observations on the nutrition-reproduction correlation in grouse locusts (Orthoptera: Tetrigidae). Proc. Indian Anim. Sci. 96, 323–327. doi:10.1007/BF03180016
Chen, Q., Zeng, J., Yang, L., Jia, Y., Li, Q., Feng, C., et al. (2019). The structure of day-age of male Spodoptera litura moths by sex pheromone trapping in the field. Sci. China Agricul. 52, 3819–3827. doi:10.3864/j.issn.0578-1752.2019.21.010
Chen, W., Chen, W., and Wu, W. (2005). Grading ovarian developments of Fruhstorferiola tonkinensis. Chin. Plant Prot. 25, 5–6. doi:10.3969/j.issn.1672-6820.2005.05.001
Chen, W., Fu, Y., and Wu, W. (2008). Effects of adult feeding on ovarian development and fecundity of Fruhstorferiola tonkinensis (Orthoptera: Catantopidae). Chin. J. Trop. Crops 29, 89–92. doi:10.3969/j.issn.1000-2561.2008.01.018
Colares, F., Michaud, J. P., Torres, J. B., and Silva-Torres, C. S. A. (2015). Polyandry and male mating history affect the reproductive performance of Eriopis connexa (Coleoptera: Coccinellidae). Ann. Entomol. Soc. Am. 108, 736–742. doi:10.1093/aesa/sav056
Elamin, A. E. H., Abdalla, A. M., and Naim, A. M. E. (2014). The biology of Senegalese grasshopper (Oedaleus senegalensis, Krauss, 1877) (Orthoptera: Acrididae). Inter. J. Adv. Life Sci. Technol. 1, 6–15. doi:10.18488/journal.72/2014.1.1/72.1.6.15
Elsherif, L., Heimlich, J. B., Kamthunzi, P., Mafunga, P., Gopal, S., Key, N. S., et al. (2018). Exploratory study of urine metabolomics in sickle cell disease children with albuminuria in Malawi. Blood 132 (1), 3680. doi:10.1182/blood-2018-99-120233
Everman, E. R., Freda, P. J., Brown, M., Schieferecke, A. J., Ragland, G. J., and Morgan, T. J. (2018). Ovary development and cold tolerance of the invasive pest Drosophila suzukii (Matsumura) in the central plains of Kansas, United States. Environ. Entomol. 47, 1013–1023. doi:10.1093/ee/nvy074
Fang, L., Li, Z., Zhang, S., Zhang, W., Shu, J., and Wang, H. (2022). Identification and selection of the internal reference genes of Ceracris kiangsu (Orthoptera: Arcypteridae) by RT-PCR. Sci. Silvae Sin. 58, 70–77. doi:10.11707/j.1001-7488.20220108
Gosterit, A., Koskan, O., and Gurel, F. (2016). The relationship of weight and ovarian development in Bombus terrestris L. workers under different social conditions. J. Apic. Sci. 60, 51–58. doi:10.1515/jas-2016-0016
Grech, M. M., Manzo, L. M., Epele, L. B., Laurito, M., Claverie, A. Ñ., Ludueña-Almeida, F., et al. (2019). Mosquito (Diptera: Culicidae) larval ecology in natural habitats in the cold temperate patagonia region of Argentina. Parasite 12, 214. doi:10.1186/s13071-019-3459-y
Han, H., Wang, N., Xu, L., Gao, S., Liu, A., and De, Q. (2018). Grading criteria for the ovarian development of Oedaleus decorus asiaticus and differences between two types of ovarian development. Plant Prot. 44, 135–138. doi:10.16688/j.zwbh.2017094
Hill, M. O., and Gauch, H. G. J. (1980). Detrended correspondence analysis: An improved ordination technique. Vegetatio 42, 47–58. doi:10.1007/BF00048870
Hodge, C. (1943). The internal anatomy of Leptysma marginicollis (Serv.) and Opshomala vitreipennis (Marsch.) (Orthoptera: Acrididae). J. Morph. 72, 87–124. doi:10.1002/jmor.1050720104
Johnson, C. G. (1963). Physiological factors in insect migration by flight. Nature 198, 423–427. doi:10.1038/198423a0
Kwak, K., Ko, H., Kim, S. Y., Lee, K. Y., and Yoon, H. J. (2021). Developmental characteristics of ovary and egg of migratory locust, Locusta migratoria (Orthoptera: Acrididae). Korean J. Appl. Entomol. 60, 175–183. doi:10.5656/KSAE.2021.01.1.088
Leopold, R. A. (1976). The role of male accessory glands in insect reproduction. Ann. Rev. Entomol. 21, 199–221. doi:10.1146/annurev.en.21.010176.001215
Li, H., Wang, J., Zhuo, F., Zhu, J., Tu, X., Zhang, G., et al. (2022). Review on the occurrence and management technology of Ceracris kiangsu in China. Chin. J. Biol. Control 38, 531–536. doi:10.16409/j.cnki.2095-039x.2021.05.001
Li, W., Yuan, W., Zhao, X., Li, Y., and Wu, K. (2017). Effects of mating on ovarian development and oviposition of Apolygus lucorum. J. Asia-Pac. Entomol. 20, 1442–1446. doi:10.1016/j.aspen.2017.10.013
Liang, F., Wang, Z., He, K., Bai, S., and Zhang, T. (2021). Effects of brief exposure to high temperatures on the survival and fecundity of Athetis lepigone (Lepidoptera: Noctuidae). J. Therm. Biol. 100, 103066. doi:10.1016/J.JTHERBIO.2021.103066
Liland, K. H., Mevik, B. H., and Wehrens, R. (2022). pls: Partial least squares and principal component regression. Vienna, Austria: R Foundation for Statistical Computing.
Liu, D., Zhao, S., Yang, X., Wang, R., Cang, X., Zhang, H., et al. (2021). Radar monitoring unveils migration dynamics of the yellow-spined bamboo locust (Orthoptera: Arcypteridae). Comput. Electron. Agr. 187, 106306. doi:10.1016/J.COMPAG.2021.106306
Liu, J., Zhao, N., Yang, B., and Li, Z. (2019). Processing morphology change and grading criteria for ovarian development in Tomicus yunnanensis (Coleoptera, Scolytidae). For. Res. 32, 74–80. doi:10.13275/j.cnki.lykxyj.2019.01.010
Liu, W., and Huang, C. (2018). Advances in modern crop disease and pest forecast in China. Plant Prot. 44, 159–167. doi:10.16688/j.zwbh.2018248
Liu, Y., and Lu, B. (1959). Anatomy and organization of the reproductive system of Locusta migratoria manilensis Meyen. Acta Entomol. Sin. 9, 1–20. doi:10.16380/j.kcxb.1959.01.001
Liu, Y., Ye, G., and Li, X. (2006). Effects of temperature on the development of ovary and egg of Bombyx mandarina Bulter. Nat. Enemies Insects 28, 126–131. doi:10.3969/j.issn.1674-0858.2006.03.006
Masaaki, Y., and Seiji, T. (2009). Overwintering biology and morphological characteristics of the migratory locust, Locusta migratoria after outbreaks on Iheya Island, Japan. Appl. Entomol. Zool. 44, 165–174. doi:10.1303/aez.2009.165
Meara, G. F., and Evans, D. G. (1977). Autogeny in saltmarsh mosquitoes induced by a substance from the male accessory gland. Nature 167, 342–344. doi:10.1038/267342a0
Nasser, N. A., Cullen, J., Patterson, C. W., Patterson, R. T., Roe, H. M., and Galloway, J. M. (2019). Inter-annual Arcellinida (testate lobose amoebae) assemblage dynamics within lacustrine environments. Limnologica 76, 60–71. doi:10.1016/j.limno.2019.03.006
Oksanen, J., Simpson, G. L., Blanchet, G. F., Kindt, R., Legendre, P., Minchin, P. R., et al. (2022). vegan: Community ecology package. Vienna, Austria: R Foundation for Statistical Computing.
Ouyang, Q., Mo, R., and Wu, W. (2014). Classification of ovarian stages of the melon fly, Bactrocera cucurbitae (Coquillett) (Diptera: Tephritidae). J. Biosaf. 23, 24–29. doi:10.3969/j.issn.2095-1787.2014.01.005
Papaj, D. R. (2000). Ovarian dynamics and host use. Annu. Rev. Entomol. 45, 423–448. doi:10.1146/annurev.ento.45.1.423
Porker, K., Coventry, S., Fettell, N. A., Cozzolino, D., and Eglinton, J. (2020). Using a novel PLS approach for envirotyping of barley phenology and adaptation. Field Crops Res. 246, 107697. doi:10.1016/j.fcr.2019.107697
Qi, G., Lu, F., Hu, G., Wang, F., Gao, Y., and Lv, L. (2011). The application of ovarian dissection in the research on migratory insects in China. Chin. Plant Prot. 31, 18–22. doi:10.3969/j.issn.1672-6820.2011.07.004
Ren, J., Zhao, L., and Ge, Q. (2014). Ovarian development in Calliptamus italicus (L.) (Orthoptera: Catantopidae). Chin. J. Appl. Entomol. 51, 1280–1288. doi:10.7679/j.issn.2095−1353.2014.152
Roy, S., Saha, T., Zou, Z., and Raikhel, A. S. (2018). Regulatory pathways controlling female insect reproduction. Annu. Rev. Entomol. 63, 489–511. doi:10.1146/annurev-ento-020117-043258
Song, J., Guo, W., Jiang, F., and Zhou, S. (2013). Argonaute 1 is indispensable for juvenile hormone mediated oogenesis in the migratory locust, Locusta migratoria. Insect biochem. Mol. 43, 879–887. doi:10.1016/j.ibmb.2013.06.004
Song, J., Li, W., Zhao, H., and Zhou, S. (2019). Clustered miR-2, miR-13a, miR-13b and miR-71 coordinately target Notch gene to regulate oogenesis of the migratory locust Locusta migratoria. Insect biochem. Mol. Biol. 106, 39–46. doi:10.1016/j.ibmb.2018.11.004
Wise, T., and Dolan, R. J. (2020). Associations between aversive learning processes and transdiagnostic psychiatric symptoms in a general population sample. Nat. Commun. 11, 4179. doi:10.1038/s41467-020-17977-w
Wold, H. (1975). Soft modelling by latent variables: The non-linear iterative partial least squares (NIPALS) approach. J. Appl. Probab. 12, 117–142. doi:10.1017/S0021900200047604
Wu, M., and Zhang, J. (1997). Study on the relationship between ovarian development level and body weight and its application in rice grasshoppers. Entomol. Knowl. 34, 257–258.
Xu, J., Lv, X., Lin, J., Zhang, Z., Yao, Q., Fan, X., et al. (2020). The diagnostic model of cotton aphids based on leaf textural features. Cott. Sci. 32, 133–142. doi:10.11963/1002-7807.xjclx.20200220
Zhang, L., Cheng, L., Chapman, J. W., Sappington, T. W., Liu, J., Cheng, Y., et al. (2020). Juvenile hormone regulates the shift from migrants to residents in adult oriental armyworm, Mythimna separata. Sci. Rep. 10, 11626. doi:10.1038/s41598-020-66973-z
Zhang, Y., Zhao, B., Roy, S., Saha, T. T., Kokoza, V. A., Li, M., et al. (2016). MicroRNA-309 targets the Homeobox gene SIX4 and controls ovarian development in the mosquito Aedes aegypti. Proc. Natl. Acad. Sci. USA. 113, E4828–E4836. doi:10.1073/pnas.1609792113
Zhao, S., Yang, X., He, W., Zhang, H., Jiang, Y., and Wu, K. (2019). Ovarian development gradation and reproduction potential prediction in Spodoptera frugiperda. Plant Prot. 45, 28–34. doi:10.16688/j.zwbh.2019413
Zhao, Z., and Xi, G. (2005). The seasonal dynamics of ovarian development of six dominant species of locusts in Siping area of Jilin. Chin. Bull. Entomol. 42, 524–527. doi:10.3969/j.issn.0452-8255.2005.05.011
Keywords: yellow-spined bamboo locust, phenotypic traits, ovary, grade, migratory pest, dissection
Citation: Wang M, Li H, Zhang W, Zhuo F, Li T, Lowry A and Zhang A (2023) Reproduction system development of Ceracris kiangsu Tsai female adults and its relationship with fitness characteristics. Front. Physiol. 14:1136559. doi: 10.3389/fphys.2023.1136559
Received: 03 January 2023; Accepted: 14 February 2023;
Published: 07 March 2023.
Edited by:
Jia Fan, State Key Laboratory for Biology of Plant Diseases and Insect Pests, Institute of Plant Protection (CAAS), ChinaReviewed by:
Huai-Jun Xue, Nankai University, ChinaCopyright © 2023 Wang, Li, Zhang, Zhuo, Li, Lowry and Zhang. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Hongmei Li, aC5saUBjYWJpLm9yZw==; Aihuan Zhang, emhhbmdhaWh1YW5AMTI2LmNvbQ==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.