 Lluc Montull
Lluc Montull Alex Borrallo
Alex Borrallo Maricarmen Almarcha
Maricarmen Almarcha Natàlia Balagué
Natàlia Balagué
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Physiol. , 20 January 2023
Sec. Exercise Physiology
Volume 14 - 2023 | https://doi.org/10.3389/fphys.2023.1112902
This article is part of the Research Topic Sensor Technologies and Biosignal Processing Methods to Explore the Physiological Functions of Proprioception View all 8 articles
Proprioception is a crucial property for movement stability and balance, but its current assessment, based on clinical testing, lacks precision and adequacy in real contexts. This study proposes assessing proprioception and its sensitivity to training effects through acceleration time series recorded during two slackline experiments. In the first experiment, slackliners of different expertise (highly and poorly trained) had to walk on a slackline for 30 s. In the second, twelve beginners had to balance up on the slackline for at least 11 s before and after a training process. Acceleration time series were recorded in body components (legs and centre of mass) and the slackline. The acceleration fluctuations were analysed through Detrended Fluctuation Analysis. The obtained Hurst (H)-exponents were compared between both groups (first experiment) and before and after training (second experiment) using Whitney and Wilcoxon tests, respectively. The values of H-exponents were lower in the highly trained group (Z = −2.15, p = 0.03) (first experiment), and in the post-training conditions (Z = −2.35, p = 0.02) (second experiment). These results suggest better motor and proprioceptive control with training status. Hence, the time-variability structure of acceleration in real contexts, like slackline tasks, is proposed as an objective measure of proprioception and its training effects.
Proprioception, the sensation of body position and movement, often described as the “sixth sense”, plays a crucial role in human motor control, performance, and injury prevention (Tuthill and Azim, 2018). Together with ocular, vestibular, or haptic sensitivity, it is essential in movement stability and balance tasks with dominant proprioceptive feedback (Ogard, 2011). Its objective assessment is difficult because the available methods, protocols, and analysis techniques, mainly applied to clinical contexts, lack precision and adequacy (Hillier et al., 2015).
Proprioception has been mostly assessed in the scope of a general clinical environment for “screening” the influence of impaired proprioception and guiding rehabilitation interventions (Hillier et al., 2015; Röijezon et al., 2015). Capturing the error or differences between the targeted and the performed movements, force sense tests, or threshold detection of passive motion stand as widely accepted examples of proprioception assessment (Lephart et al., 2002; Weerakkody et al., 2008; Han et al., 2016).
The proprioceptive tests used in clinical practice, based on conscious proprioception and decontextualized movements requiring reproduction or even verbalization of simple motions, are inappropriate to be applied in real settings (Ager et al., 2017; Muñoz-Jiménez et al., 2021). Unlike in clinical contexts, in daily activities, the sub-conscious proprioception is dominantly relevant (Hillier et al., 2015). For instance, in sports and exercise, proprioception contributes to the coordination and fine adjustments of body movements to maintain dynamic stability in safe conditions (Balagué et al., 2014; Vázquez et al., 2016; 2021). Such fine adjustments emerging from perception-action cycles, acting at very short timescales (i.e., actions instantaneously create new perceptions for future actions), cannot be consciously captured (Shaw and Kinsella-Shaw, 2007; Montull et al., 2020).
In addition, proprioceptive standard measurements, often contrasted with theoretical group-pooled data to categorize a prototypic “healthy” state, neglect the uniqueness and richness of individual adaptive responses under changing real constraints (Hillier et al., 2015). Then, the use of activities like slacklining, skiing, or acrobatics, which require balance and stability, can be useful in assessing individual proprioceptive properties in real settings. The registration of time series of kinematic variables extracted from body components and interacting instruments (e.g., slackline) during these activities may be seen as promising measures to capture such properties.
Time series analyses allow detecting the dynamics of variables representing the coordinated behaviour of the system (Haken, 1987; Balagué et al., 2014; Balagué et al., 2020; Fonseca et al., 2020). Different kinematic or physiological variables have been studied with such purpose. For instance, Vázquez et al. (2016, 2021) applied Detrended Fluctuation Analysis (DFA) to the series of elbow joint angle while performing a quasi-isometric exercise until exhaustion. In this work, the elbow angle (a kinematic variable) integrated several processes (e.g., metabolic, contractile, reflex and volitional, among others). The authors evaluated the autocorrelation of the series through the Hurst (H) exponents which informed about the adaptive properties of the performer to effort accumulation.
Time series-based analyses, such as fractal or entropy analyses of heart rate, have also been used to detect the adaptability to workloads of the cardiovascular system (Gronwald et al., 2020) or pathological states (Dutta et al., 2013; Kirchner et al., 2014). According to Kelso (1997), it is challenging to adequately set the collective variables that reduce the system dimensionality and represent the innumerable physiological degrees of freedom that control every human movement.
Applying a DFA to acceleration time series of body components during slackline tasks, Montull et al. (2020; 2021) extracted information about the motor control of slackliners, notably modulated by proprioceptive properties. Consequently, the time series of acceleration, an increasingly common monitored variable in sporting contexts (Scott et al., 2016; Simperingham et al., 2016), has the potential of being an adequate kinematic variable to capture proprioception in stability and balance tasks (Montull et al., 2021).
Slacklining is a challenging task that relies on a tight physical coupling of the performer with the environment, represented by the slackline (Montull et al., 2020). The slackline is an unstable and narrow pendulum tensioned between two anchors that requires continuous fast adjustments of body components to stand up in balance (see Figure 1) (Paoletti and Mahadevan, 2012).

FIGURE 1. Representation of the task performed in each experiment: walking (left) and standing up balancing (right). Abbreviations of the monitored acceleration points: Dom = dominant leg; SubDom = subdominant leg; CoM = centre of mass; Sup = support leg; Free = free leg; Slack = slackline.
H-exponents have been used to evaluate the time-variability structure of acceleration of body components of slackliners Montull et al. (2020; 2021). Ranging from 0 < H < 0.5, the fluctuations are featured by an anti-persistent character and reflect a tight and fast control related to a more adaptive behaviour (Terrier and Dériaz, 2012; Vázquez et al., 2016). In contrast, in the range of 0.5 < H < 1, the fluctuations are featured by a persistent temporal structure and relate to a more rigid motor control that may lead to the consequent impending instability (Scheffer et al., 2009; Delignières et al., 2011). These acceleration fluctuations of body components change synergistically (Haken, 1987) for stabilizing the slackline (the performance variable) (Montull et al., 2021). Thus, the slackline integrates and compresses the fluctuations of the whole performer-environment system.
This exploratory study aimed to evaluate the potential of the time-variability properties of acceleration during slackline tasks to assess proprioception and its sensitivity to training effects. We hypothesized that the H-exponent would be lower 1) in highly trained compared to poorly trained slackliners, and 2) in beginners after a slackline training process. Accordingly, we also hypothesized that slackline, the performance variable, could be more sensitive to training process than body components.
Nineteen slackliners (17 males, two females, 25.3 ± 4.9 years, 70.21 ± 8.79 kg, 1.79 ± 0.05 m) of different levels (training frequency = 2.29 ± 2.98 h/week; training age = 3.05 ± 2.87 years), from a faculty of Sport Sciences and a Slackline Club, were recruited. Two training groups were formed: highly trained (n = 11, ≥ 2 h/week and ≥1 year) and poorly trained (n = 8, < 2 h/week and <1 year). As there are no specific categories to define the performance levels of slackliners, the criterion for classifying them on highly and poorly trained was based on the opinion of two expert slackliners.
To be included in the sample, the slackliners had to be able to perform the proposed slackline walking task (see procedures). The experiment was approved by the local Research Ethics Committee (072015CEICEGC) and was performed according to the ethical standards of the Helsinki Declaration. Participants were informed about the experimental procedure and signed an informed consent before the intervention.
Slackliners were cited individually to the mobile lab, placed in a quiet location to avoid disturbing stimuli. When arriving they were first monitored and afterwards instructed about the task as follows: “You have to walk without shoes along the slackline during 30 s at a freely chosen velocity starting close to the anchor (see Figure 1). We will notify you when the time is over. You have a maximum of 3 attempts, separated by a maximum of 5 min resting time, to accomplish the task. Any question?” The number of attempts and the resting times were fixed according to the results of a pilot study performed on a similar population.
The slackline (Gibbon Slackline TM, ID Sports, Stuttgart, Germany) was 15 m long and 5 cm wide. The tension (T) of the slackline’s anchors (5.28 ± 0.65 kN), placed at 0.85 m from the ground, was calculated through the following formula:
where M is the mass of the participants, L is the length of the slackline (15 m), and S is the sag under load (ensuring at least 0.5 m in the center) (Conley, 2006; Montull et al., 2020).
Accelerometer devices WIMU PRO™ (Real Track Systems, Almería, Spain) (Gómez-Carmona et al., 2019) were placed in the legs and in the lumbar region close to the center of mass (CoM) of slackliners (see Figure 1). Through a questionnaire, slackliners reported their dominant (Dom) and subdominant (SubDom) leg. For the legs, the accelerometers were fixed on the outside part above the lateral malleolus (Mannini et al., 2013), while for the CoM on the zone of L3 (Moe-Nilssen and Helbostad, 2004; Schütte et al., 2016). The acceleration was recorded at a sampling frequency of 100 Hz. Before the measurements, the calibration of such IMUs was performed on a flat, even surface with the Z-axis perpendicular to the surface, according to the manufacturer’s specifications.
DFA was applied in all acceleration time series to analyze their time-variability properties. This analysis was carried out as follows (according to Ihlen, 2012; Peng et al., 1994; 1995): Firstly, the total length of the acceleration time series (N = 3,072 data points) was integrated with the following equation:
Then, the local trend was calculated to fit the acceleration time series using a quadratic polynomial function (Ihlen, 2012). The resulting time series were divided into different window scales n of equal length, subtracting the local trend in each window.
For all studies, root mean square (RMS) fluctuation was calculated using the following equation:
The H-exponent, obtained as the value of the linear regression slope between the scale and the local fluctuations in a log-log diffusion plot, was used to determine the character of fluctuations for each component. Matlab© R2016b was used for this analysis.
The obtained H-exponents (HDom, HSubDom, HCoM) were compared between highly and poorly trained groups. Mann-Whitney test and Cohen’s d were computed for this purpose. According to Cohen (1988), d < 0.2 means no effect, 0.2 ≤ d ≤ 0.5 means small effect, 0.5 ≤ d ≤ 0.8 means intermediate effect, and 0.8 < d means large effect. The level of significance was set at p ≤ 0.05 throughout the study. Statistical analyses were performed with SPSS v.15 (SPSS Inc., Chicago, United States).
Twelve volunteer beginners (five males, seven females, 20.6 ± 2.27 years, 65.70 ± 8.89 kg, 1.71 ± 0.08 m) with no experience at all or just introduced (less than 3 slacklining lessons or practices) were recruited. They had to be committed to following the training process from the beginning to the end (see procedures). Participants were informed about the experimental procedure and signed informed consent before the intervention.
As in the first experiment, all slackliners were cited individually to the mobile lab, placed in a quiet location to avoid disturbing stimuli. When arriving they were first monitored and afterwards instructed about the testing task as follows: “You have to stand up, without shoes, balancing on a slackline for at least 11 s (Figure 1). You must choose one leg as the supporting leg (Sup) and the other as the free leg (Free) (no contact with the slackline). The tiptoe of the Sup must be in the sign of 1 m from the anchor. We will notify you when the time is over. You have a maximum of 5 attempts, separated by a maximum of 5 min resting time, to accomplish the task. Any question?” The number of attempts and the resting times were fixed according to the results of a pilot study performed on a similar population. Slackline tension was calculated in the same way as in the first experiment.
The testing task was repeated before and after an individualized training process consisting of surpassing 50 challenging slackline tasks of progressive difficulty (e.g., two steps forward and standing up balancing at least 3 s, standing for 5 s together with a pair while holding the hands, walking all the slackline without falling, etc.). The testing task was not included as a challenging task. During the training process participants were requested to keep their training habits and avoid other type of proprioceptive training (stability or balance exercises). Participants could practice every challenge on their own and as many times they needed until succeeding. The success was supervised once a week. The whole training process lasted from one to 3 months (depending on the individual learning abilities) with a median (IQR) training time of 90 h (47.5 h). After surpassing the 50 challenges, they performed the post-training testing within the next 7 days.
Acceleration was captured in the same way as in the first experiment for both legs (Sup and Free) and CoM (Figure 1). The acceleration of the slackline (Slack), the performance variable (Montull et al., 2021), fixed at 0.5 m from the toe of Sup, was also registered and included in the analysis. The acceleration was recorded at a sampling frequency of 100 Hz to ensure enough data points for the DFA. The total length of the acceleration time series ranged from N = 1,024 to 2,048 data points. DFA was calculated as explained in the first experiment, also obtaining H-exponents as an outcome.
The sensitivity of H-exponents to training was tested by comparing the H values of each component pre- and post-training. Wilcoxon test and Cohen’s d were used for this purpose. Previously, Kolgomorov-Smirnov test was applied to demonstrate a non-normal distribution of all variables.
As illustrated in Table 1, depending on body components, the fluctuations profile changed. Differently than others, the HCoM revealed, on average, a weakly anti-persistent structure of fluctuations in both experiments (H close to 0.5). In contrast, both legs and slackline had persistent structures: HDom, HSup and HSlack with more moderate persistent dynamics, while HSubDom and HFree legs tended to have higher persistent dynamics (H close to 1).
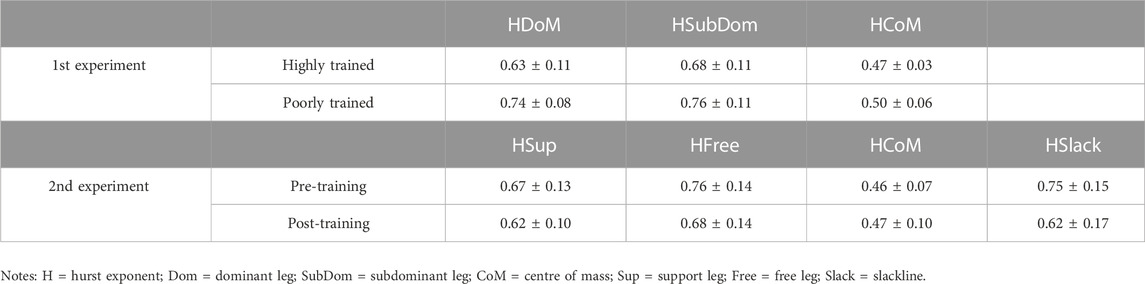
TABLE 1. Mean ± SD of the Hurst exponents of body and slackline acceleration from the two experiments.
In the first experiment, the highly trained group showed lower persistent dynamics of acceleration in all body components compared with the poorly trained group (see Table 1). In particular, displaying considerably lower values of HDom (Z = −2.15; p = 0.03; d = −1.12), and intermediately lower values of HSubDom (d = −.63) and HCoM (d = −.57).
In the second experiment, post-training values reduced the persistency of fluctuations compared with pre-training (see Table 1). HSlack was significantly lower (Z = −2.35, p = .02; d = −.81), tending towards a moderate anti-persistent profile. Also, both legs tended towards lower persistency: an intermediate reduction was found in HFree (d = −.6) while small effect size was shown in HSup (d = −.43).
This study found that H-exponents, evaluating the time variability of acceleration during slackline tasks, captured body and slackline fluctuations and were sensitive to training effects. The motor control of the different body components, favored by proprioceptive properties, showed that: CoM featured rapid and frequent fluctuations (weakly anti-persistency) for maintaining the postural control, Sup/Dom featured persistent moderate fluctuations for rapidly adjusting to slackline fluctuations, and Free/SubDom featured high persistent fluctuations for compensating through large and rigid movements the impending instability (Montull et al., 2020; 2021). All components dynamically interacted and spontaneously reorganized to accomplish the task goal (i.e., stabilize the slackline).
The time-variability properties of acceleration showed sensitivity to training effects. In the first experiment, the highly trained group tended towards lower persistent profiles (H closer to 0.5) of all body components and showed better control of both legs during slackline walking. The dominant leg showed the highest differences, probably due to its essential role in keeping the stability while walking. Such improvement in the motor control has been related to longer stability performance (Montull et al., 2021). These findings agree with previous work studying other sports activities, such as running, in which trained runners, compared to non-runners, showed lower persistency of gait cycle fluctuations (Nakayama et al., 2010).
In the second experiment, legs and slackline tended towards lower persistent dynamics after training. The slackline, representing the performance variable and compressing the proprioception of the whole performer-slackline system, reflected the largest improvement. In contrast, the time-variability of CoM’s acceleration did not show significant changes. Compared to legs, this lower sensitivity to training of CoM is probably due to its longer distance from the slackline (Singh et al., 2020).
Considering the stabilizing synergies between the body components, training should probably induce improvements at different levels and timescales, including microscopic (e.g., cell mechanics), mesoscopic (e.g., reflex processes) and macroscopic processes (e.g., attention, concentration) in a correlated way. This enhancement of motor and proprioceptive control is linked to the self-development of balance-retention and fatigue-reduction (Gabel et al., 2016).
It is plausible to suggest how the multiple embedded physiological components of slackliners interact to permit motor-proprioceptive control (Singh et al., 2020; Gabel et al., 2021; Seidel-Marzi et al., 2021). Accordingly, acceleration seems an adequate variable to describe and compress performers’ behaviour in stability and balance exercises. This type of kinematic variables, capturing the action level, is promoted by the Network Physiology of Exercise because they provide integrated information about the synchronization and coordination of processes interacting horizontally (e.g., within the same level) and vertically (among levels, e.g., cells, tissues, organs, etc.) in the organism (Balagué et al., 2020; 2022). Therefore, the proposed proprioceptive assessment might be particularly helpful to avoid fragmented, oversimplified, passive and decontextualized proprioception measures capturing this property in a reasonably integrative and applied manner (Ager et al., 2017; Muñoz-Jiménez et al., 2021). In addition, it may provide sensitive information about the rapid, sub-conscious, and often imperceptible motor readjustments that, modulated by proprioception, operate in real exercise contexts (Hillier et al., 2015).
Diverse balance and coordinative activities, dependent on proprioceptive abilities, might also be assessed objectively by analysing the variability properties of acceleration. This opens an exciting research line to approach different phenomena related to health and performance. For example, to assess the state of neurodegenerative disorders (Lheureux et al., 2020), the acute effects of fatigue (Montull et al., 2019), the risks of sports injuries (Pol et al., 2019) or the rehabilitation progress (Lheureux et al., 2020).
This is an exploratory study and further research is needed to robustly claim that proprioception can be assessed through time-variability properties of acceleration. In particular, exploring the validity and reliability of acceleration time series and DFA in other types of exercise is warranted. Synergies analysis, such as Uncontrolled Manifold, should also be considered as a complement to understand better the proprioceptive communication between the studied components in balance exercises (Montull et al., 2021).
This study suggests that the time-variability structure of acceleration in real contexts, like slackline tasks, may objectively measure proprioception and be sensitive to its training effects. From a practical point of view, the monitorization of the slackline itself, compressing the motor synergies in this type of task, seems to be sufficient to inform about the proprioceptive control.
The raw data supporting the conclusion of this article will be made available by the authors, without undue reservation.
The studies involving human participants were reviewed and approved by 072015CEICEGC. The patients/participants provided their written informed consent to participate in this study.
LM and NB designed the study and wrote the manuscript. LM, AB, and MA recruited volunteers, and collected and analyzed data. AB and MA assisted with manuscript preparation.
The authors would like to thank the participants, the National Institute of Physical Education of Catalonia (INEFC) and the Complex Systems in Sport research group for their support. Also, to Robert Hristovski for his support in statistical and theoretical issues.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
CoM, Center of mass; DFA, Detrended Fluctuation Analysis; Dom, Dominant leg; Free, Free leg; H-exponent, Hurst exponent; Slack, Slackline; SubDom, Subdominant leg; Sup, Support leg.
Ager A. L., Roy J. S., Roos M., Belley A. F., Cools A., Hébert L. J. (2017). Shoulder proprioception: How is it measured, and is it reliable? A systematic review. J. Hand Ther. 30 (2), 221–231. doi:10.1016/j.jht.2017.05.003
Balagué N., Hristovski R., Almarcha M., Garcia-Retortillo S., Ivanov P. C. (2022). Network Physiology of exercise: Beyond molecular and omics perspectives. Sports Med.-Open 8 (119), 119. doi:10.1186/s40798-022-00512-0
Balagué N., Hristovski R., Almarcha M., Garcia-Retortillo S., Ivanov P. C. (2020). Network Physiology of exercise: Vision and perspectives. Front. Physiol. 11–611550. doi:10.3389/fphys.2020.611550
Balagué N., Hristovski R., Vainoras A., Vázquez P., Aragonés D. (2014). “Psychobiological integration during exercise,” in Complex systems in sport. Editors K. Davids, R. Hristovski, D. Araújo, N. Balagué, C. Y. Button, and P. Passos (London: Routledge), 82–102.
Cohen J. (1988). Statistical power analysis for the behavioral Sciences. 2nd. Hillsdale, NJ: Lawrence Erlbaum Associates.
Conley W. (2006). A practical analysis of slackline forces. https://ww3.cad.de/foren/ubb/uploads/Andre+Furter/SlacklineAnalysis.pdf (Accessed July 10, 2022).
Delignières D., Torre K., Bernard P. L. (2011). Transition from persistent to antipersistent correlations in postural sway indicates velocity based control. PLoS Comput. Biol. 7 (2), e1001089–9. doi:10.1371/journal.pcbi.1001089
Dutta S., Ghosh D., Chatterjee S. (2013). Multifractal detrended fluctuation analysis of human gait diseases. Front. Physiol. 4, 274. doi:10.3389/fphys.2013.00274
Fonseca S. T., Souza T. R., Verhagen E., van Emmerik R., Bittencourt N., Mendonça L., et al. (2020). Sports injury forecasting and complexity: A synergetic approach. Sports Med. 50 (10), 1757–1770. doi:10.1007/s40279-020-01326-4
Gabel C. P., Guy B., Mokhtarinia H. R., Melloh M. (2021). Slacklining: A narrative review on the origins, neuromechanical models and therapeutic use. World J. Orthop. 12 (6), 360–375. doi:10.5312/wjo.v12.i6.360
Gabel C. P., Rando N., Melloh M. (2016). Slacklining and stroke: A rehabilitation case study considering balance and lower limb weakness. World J. Orthop. 7 (8), 513–518. doi:10.5312/wjo.v7.i8.513
Gómez-Carmona C. D., Bastida-Castillo A., González-Custodio A., Olcina G., Pino-Ortega J. (2019). Using an inertial device (WIMU PRO) to quantify neuromuscular load in running: Reliability, convergent validity, and influence of type of surface and device location. J. Strength Cond. Res. 34 (2), 365–373. doi:10.1519/JSC.0000000000003106
Gronwald T., Rogers B., Hoos O. (2020). Fractal correlation properties of heart rate variability: A new biomarker for intensity distribution in endurance exercise and training prescription? Front. Physiol. 11–550572. doi:10.3389/fphys.2020.550572
Haken H. (1987). “Synergetics: An approach to self-organization,” in self-organizing systems: The emergence of order. Editor F. E. Yates (New York: Plenum Press), 417–434.
Han J., Waddington G., Adams R., Anson J., Liu Y. (2016). Assessing proprioception: A critical review of methods. J. Sport Health Sci. 5 (1), 80–90. doi:10.1016/j.jshs.2014.10.004
Hillier S., Immink M., Thewlis D. (2015). Assessing proprioception: A systematic review of possibilities. Neurorehabil. Neural Repair 29 (10), 933–949. doi:10.1177/1545968315573055
Ihlen E. A. F. (2012). Introduction to multifractal detrended fluctuation analysis in matlab. Front. Physiol. 3, 141. doi:10.3389/fphys.2012.00141
Kelso J. A. S. (1997). Dynamic patterns: The self-organization of brain and behaviour. Cambridge: Bradford Books.
Kirchner M., Schubert P., Liebherr M., Haas C. T. (2014). Detrended fluctuation analysis and adaptive fractal analysis of stride time data in Parkinson's disease: Stitching together short gait trials. PLoS One 9 (1)–e85787. doi:10.1371/journal.pone.0085787
Lephart S. M., Myers J. B., Bradley J. P., Fu F. H. (2002). Shoulder proprioception and function following thermal capsulorraphy. Arthrosc.- J. Arthrosc. Relat. Surg. 18 (7), 770–778. doi:10.1053/jars.2002.32843
Lheureux A., Warlop T., Cambier C., Chemin B., Stoquart G., Detrembleur C., et al. (2020). Influence of autocorrelated rhythmic auditory stimulations on Parkinson's disease gait variability: Comparison with other auditory rhythm variabilities and perspectives. Front. Physiol. 11–572063. doi:10.3389/fphys.2020.572063
Mannini A., Intille S. S., Rosenberger M., Sabatini A. M., Haskell W. (2013). Activity recognition using a single accelerometer placed at the wrist or ankle. Med. Sci. Sports Exerc. 45, 2193–2203. doi:10.1249/MSS.0b013e31829736d6
Moe-Nilssen R., Helbostad J. L. (2004). Estimation of gait cycle characteristics by trunk accelerometry. J. Biomech. 37, 121–126. doi:10.1016/s0021-9290(03)00233-1
Montull L., Vázquez P., Hristovski R., Balagué N. (2019). Monitoring the dynamics of exhausting exercises: The time-variability of acceleration. Vibroeng. Proced. 26, 64–67. doi:10.21595/vp.2019.20980
Montull L., Vázquez P., Rocas L., Hristovski R., Balagué N. (2020). Flow as an embodied state. Informed awareness of slackline walking. Front. Psychol. 10, 29931–30011. doi:10.3389/fpsyg.2019.02993
Montull L., Passos P., Rocas L., Milho J., Balagué N. (2021). Proprioceptive dialogue-interpersonal synergies during a cooperative slackline task. Nonlinear Dyn. Psychol. Life Sci. 25 (2), 157–177. doi:10.13140/RG.2.2.35466.70087
Muñoz-Jiménez J., Rojas-Valverde D., Leon K. (2021). Future challenges in the assessment of proprioception in exercise Sciences: Is imitation an alternative? Front. Hum. Neurosci. 15–664667. doi:10.3389/fnhum.2021.664667
Nakayama Y., Kudo K., Ohtsuki T. (2010). Variability and fluctuation in running gait cycle of trained runners and non-runners. Gait Posture 31 (3), 331–335. doi:10.1016/J.GAITPOST.2009.12.003
Ogard W. K. (2011). Proprioception in sports medicine and athletic conditioning. Strength condit. J. 33 (3), 111–118. doi:10.1519/SSC.0B013E31821BF3AE
Paoletti P., Mahadevan L. (2012). Balancing on tightropes and slacklines. J. R. Soc. Interface 9, 2097–2108. doi:10.1098/rsif.2012.0077
Peng C. K., Buldyrev S. V., Havlin S., Simons M., Stanley H. E., Goldberger A. L. (1994). Mosaic organization of DNA nucleotides. Phys. Rev. E Stat. Nonlin. Soft Matter Phys. 49, 1685–1689. doi:10.1103/physreve.49.1685
Peng C. K., Havlin S., Stanley H. E., Goldberger A. L. (1995). Quantification of scaling exponents and crossover phenomena in nonstationary heartbeat time series. Chaos 5, 82–87. doi:10.1063/1.166141
Pol R., Hristovski R., Medina D., Balagué N. (2019). From microscopic to macroscopic sports injuries. Applying the Complex dynamic systems approach to sports medicine: A narrative review. Br. J. Sports Med. 53 (19), 1214–1220. doi:10.1136/bjsports-2016-097395
Röijezon U., Clark N. C., Treleaven J. (2015). Proprioception in musculoskeletal rehabilitation. Part 1: Basic science and principles of assessment and clinical interventions. Man. Ther. 20 (3), 368–377. doi:10.1016/j.math.2015.01.008
Scheffer M., Bascompte J., Brock W. A., Brovkin V., Carpenter S. R., Dakos V., et al. (2009). Early-warning signals for critical transitions. Nature 461, 53–59. doi:10.1038/nature08227
Schütte K. H., Aeles J., De Beéck T. O., van der Zwaard B. C., Venter R., Vanwanseele B. (2016). Surface effects on dynamic stability and loading during outdoor running using wireless trunk accelerometry. Gait Posture 48, 220–225. doi:10.1016/j.gaitpost.2016.05.017
Scott M. T., Scott T. J., Kelly V. G. (2016). The validity and reliability of global positioning systems in team sport: A brief review. J. Strength Cond. Res. 30 (5), 1470–1490. doi:10.1519/JSC.0000000000001221
Seidel-Marzi O., Hähner S., Ragert P., Carius D. (2021). Task-related hemodynamic response alterations during slacklining: An FNIRS study in advanced slackliners. Front. Neuroergonomics 2–644490. doi:10.3389/FNRGO.2021.644490
Shaw R. E., Kinsella-Shaw J. (2007). The survival value of informed awareness. J. Conscious. Stud. 14, 137–154.
Simperingham K. D., Cronin J. B., Ross A. (2016). Advances in sprint acceleration profiling for field-based team-sport athletes: Utility, reliability, validity and limitations. Sports Med. 46 (11), 1619–1645. doi:10.1007/s40279-016-0508-y
Singh R. E., Iqbal K., White G. (2020). Proficiency-based recruitment of muscle synergies in a highly perturbed walking task (slackline). Eng. Rep. 2–e12253. doi:10.1002/eng2.12253
Terrier P., Dériaz O. (2012). Persistent and anti-persistent pattern in stride to-stride variability of treadmill walking: Influence of rhythmic auditory cueing. Hum. Mov. Sci. 31, 1585–1597. doi:10.1016/j.humov.2012.05.004
Tuthill J. C., Azim E. (2018). Proprioception. Curr. Biol. 28 (5), R194-R203–R203. doi:10.1016/j.cub.2018.01.064
Vázquez P., Hristovski R., Balagué N. (2016). The path to exhaustion: Time-variability properties of coordinative variables during continuous exercise. Front. Physiol. 7 (37), 37–38. doi:10.3389/fphys.2016.00037
Vázquez P., Petelczyc M., Hristovski R., Balagué N. (2021). Interlimb coordination: A new order parameter and a marker of fatigue during quasi-isometric exercise? Front. Physiol. 11–612709. doi:10.3389/fphys.2020.612709
Keywords: detrended fluctuation analysis, kinematic variable, motor control, slackline, movement stability, balance, training
Citation: Montull L, Borrallo A, Almarcha M and Balagué N (2023) Assessing proprioception through time-variability properties of acceleration. Front. Physiol. 14:1112902. doi: 10.3389/fphys.2023.1112902
Received: 30 November 2022; Accepted: 05 January 2023;
Published: 20 January 2023.
Edited by:
Ahsan H. Khandoker, Khalifa University, United Arab EmiratesReviewed by:
Duarte Araújo, University of Lisbon, PortugalCopyright © 2023 Montull, Borrallo, Almarcha and Balagué. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Natàlia Balagué, bmF0YWxpYWJhbGFndWVAZ21haWwuY29t
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.