 Zhiqiang Lei1†
Zhiqiang Lei1† Songqing Lai
Songqing Lai Jing Lin
Jing Lin Yanni Lv
Yanni Lv
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Physiol., 27 April 2022
Sec. Vascular Physiology
Volume 13 - 2022 | https://doi.org/10.3389/fphys.2022.863265
This article is part of the Research TopicVascular Endothelial Niche: Endothelial Cells Mutually Interact With Neighboring and Circulating Cells in Complex Multicellular Networks or Vascular NichesView all 4 articles
Vascular endothelial cells, forming the inner wall of the blood vessels, participate in the body’s pathological and physiological processes of immunity, tumors, and infection. In response to an external stimulus or internal pathological changes, vascular endothelial cells can reshape their microenvironment, forming a “niche”. Current research on the vascular endothelial niche is a rapidly growing field in vascular biology. Endothelial niches not only respond to stimulation by external information but are also decisive factors that act on neighboring tissues and circulating cells. Intervention through the vascular niche is meaningful for improving the treatment of several diseases. This review aimed to summarize reported diseases affected by endothelial niches and signal molecular alterations or release within endothelial niches. We look forward to contributing knowledge to increase the understanding the signaling and mechanisms of the vascular endothelial niche in multiple diseases.
The definition of niche originated from the ecological environment, representing the environment and living habits for each species. In recent years, the concept of a “niche” has been introduced into the medical field, especially the field of oncology (Rafii et al., 2016). Metastatic tumors prefer certain tissue locations or niches for their growth and metastasis. The microenvironment can be identified as a ‘niche’ as well. It is believed that the microenvironment, namely, the “niche”, promotes the metastasis of cancer cells to specific nonadjacent organs through the circulatory system. The microenvironmental “niche” is characterized by abnormal glucose metabolism, disturbance of lipid metabolism, abnormal protein signaling molecule secretion, and so on, providing a supportive microenvironment for inflammation, immunosuppression, angiogenesis, vascular permeability, and lymphangiogenesis (Géraud et al., 2014). These characteristics of niche comprise the preferred condition for tumor metastasis. For example, exosomes derived from brain metastatic breast cancer and lung tumors were rich in hyaluronic acid binding protein, which formed the premetastatic ecological niche of the cerebrovascular endothelium and promoted tumor exosomes to brain metastasis (Rodrigues et al., 2019). Colorectal cancer exosomal miR-25-3p promoted niche formation of vascular endothelial to facilitate breast cancer metastasis by inducing vascular permeability and angiogenesis (Zeng et al., 2018a). Interference in the tumor niche might represent a potential therapy to make the niche uninhabitable for tumor cells (Friedman-Levi et al., 2021). In addition to oncology, in recent years, the idea of the niche has attracted attention in central nervous, cardiovascular, lung, inflammatory, and liver diseases.
A niche can be formed by any organ, tissue or cells, among which the role of the vascular endothelial cell niche is the most extensive. Blood vessels are formed by vascular endothelial cells, mural cells, perithelial cells, and other cells that form protective barriers and provide molecular nutrition (Psaltis et al., 2011). Current findings indicate that vascular endothelial cells play a special niche role that expand their original roles. For example, under normal conditions, the vascular endothelial microenvironment is a “harmful environment” for neutrophil migration in the brain tissue. Even if a small amount of harmful molecules stick to the vascular endothelial wall, it would not penetrate the vascular endothelium, resulting in low efficiency of recruitment and migration of harmful molecules (Nikolova et al., 2007). In response to an external stimulus or internal pathological changes, vascular endothelial cells reshape their microenvironment, forming a “niche”, which support and promote harmful cell recruitment to specific tissues from the circulatory system. Thus, the vascular endothelial niche can only be fully interpreted when its specific niche components and their functions are well understood. Therefore, this review explores the changes that occur in the vascular endothelial niche responsible for pathologic changes. As a result, new vascular endothelial intervention molecular therapies could be explored as potential therapies for more diseases.
In nature, amount species tend to occupy areas with certain characteristics, such as the specific area composed of plant, soil, and germs. Ecologists have developed the concept of a “niche” to delineate the way that organisms adapt to their environment. A “niche” is described as a constantly changing set within environmental factors, which keep a specie long lives in survival (Zhang C. D. et al., 2022). The internal essence for “niche” might be addressed as a physical factor limit for the adaptive environment, determining the extent of its geography or habitat in general, whereas survival within this range depending on competing resources with other species living there (Rubalcaba et al., 2020). The concept of “niche” goes to the heart of ecology--two critical components of “niche”: organisms and environment. When the resources and environmental conditions were changed by organisms, the organisms themselves also underwent the changes (Hoshizaki et al., 2022).
The conception of “niche” is introduced from ecology to medical field. The term “vascular niche” refers to the established microenvironment around blood vessels, in which endothelial cells, hemodynamic mechanical forces, stromal cells, parenchymal cells, extracellular matrix molecules, or pericytes, and smooth muscle cells contribute to the formation of blood vessels. Thus, a mature blood vessel contains several cell populations, all of which could contribute to the formation of vascular niche. The environment of blood vessels affect its differentiation, survival and proliferation, and even exert on the adjacent cells or tissues or circulating cells (Ribatti et al., 2021). Blood vessel endothelium composed of a thin layer of epithelial cells, characteristic with flattened, polygonal, serrated edges. Vascular endothelial cells form the lining of blood vessels and also act as the interface between blood and other vascular walls. Along the entire circulatory system, vascular endothelial cells exist from the heart down to the smallest blood vessels (Ferentinos et al., 2022). Vascular endothelial cells located between plasma and vascular tissues, it could not only complete the metabolism exchange between plasma and tissue fluid, but also synthesize and secrete a variety of biological active substances in order to ensure the normal contraction of blood vessels and maintain the normal flow of blood.
The specific term “vascular endothelial niche” was defined as a microenvironment that is generated by vascular endothelial cells affects the behavior of adjacent cells or the internal circulatory system like blood, circulating cells, etc. Since the vascular vessels composed of variety cellular populations, like hemodynamic mechanical forces, stromal cells, parenchymal cells, extracellular matrix molecules, or pericytes, and smooth muscle cells, there is relatively straightforward interaction between vascular endothelial cells and adjacent cells of vascular vessels. Stromal cells are the cells to assist parenchymal cells to perform organ functions in specific organ. Zhang et al.(2022) demonstrated that circANKRD36 regulated miR-599 and TGF-β signaling pathway to promote endothelial mesenchymal transition in aortic valve stromal cells. In Jackson’ paper (Jackson, 2022), endothelial cells activated channels as transient receptor vanilloid family member 4 (TRPV4) channels, promoting intermediate and small conductance Ca2+-activated K+ (IKCa and SKCa) to transmit signaling molecules to smooth muscle via intercellular junctions, which controlling the activity of smooth muscle or pericyte contraction (Jackson, 2022). Therefore, endothelial ion channels made a microcirculation and in body balance by participating in cell-cell communication (Jackson, 2022). Macromolecules secreted by cells into the extracellular stroma form a complex network that supported tissue structure, called extracellular matrix molecules, which regulated tissue genesis and cellular physiological activities. Not just adjacent cells, vascular endothelial cells had the functions on hemodynamic mechanical forces. Embryo restricted transcription factor variant 2 (ETV2) was instantaneously reactivated in mature human endothelial cells, forming a perfusion plastic vascular plexus to facilitate organ development and tumorigenesis (Palikuqi et al., 2020).
The term “niche” also own the characteristics of stem cells (Li and Xie, 2005). The term vascular endothelial stem indicated the physical and biochemical microenvironment around blood vessel where endothelial cells, pericytes, and smooth muscle cells organize themselves to form blood vessels and release molecules involved in the recruitment of hematopoietic stem cells, endothelial progenitor cells, and mesenchymal stem cells (Abkowitz et al., 2003; Kopp et al., 2005). Vascular endothelial stem form a reticular network that supports the formation and the interaction within hematopoietic stem cells, endothelial progenitor cells, and mesenchymal stem cells. Transmembrane adhesion glycoproteins and chemokines participated in the migration of hematopoietic stem cells and endothelial progenitor cells from endothelial cells within specific niche in bone marrow (Willert et al., 2003). The vascular endothelial stem niche within bone marrow regulated stem cell mobilization, proliferation, and differentiation by cell-cell communication through the secretion of vascular endothelial growth factor, fibroblast growth factor, interleukins, transforming growth factor, platelet-derived growth factor, and nitric oxide (Pasquier et al., 2020).
The role of vascular endothelial niches is quite extensive. Vascular endothelial niches can reshape their microenvironment, playing a role in various diseases. In addition to tumors, which are the most frequently reported niches in the literature, vascular endothelial niches provide a supportive microenvironment for other diseases, such as blood diseases, cardiovascular disease, central nervous diseases, pulmonary pathologies, liver disease, and orthopedic diseases. The changes of vascular endothelial niches in specific diseases could be seen in Table 1; Figure 1.
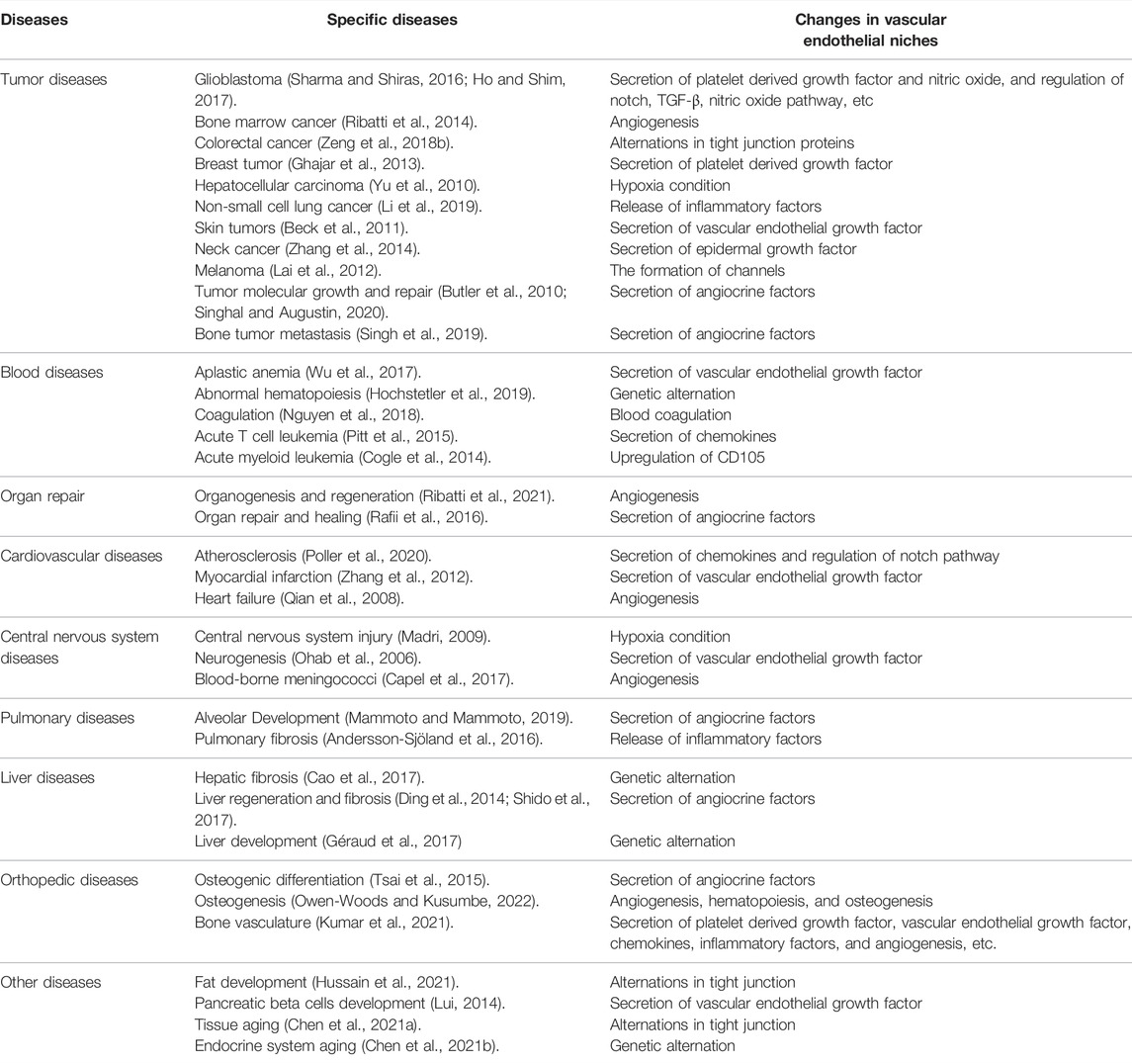
TABLE 1. The changes of vascular endothelial niches in specific diseases.
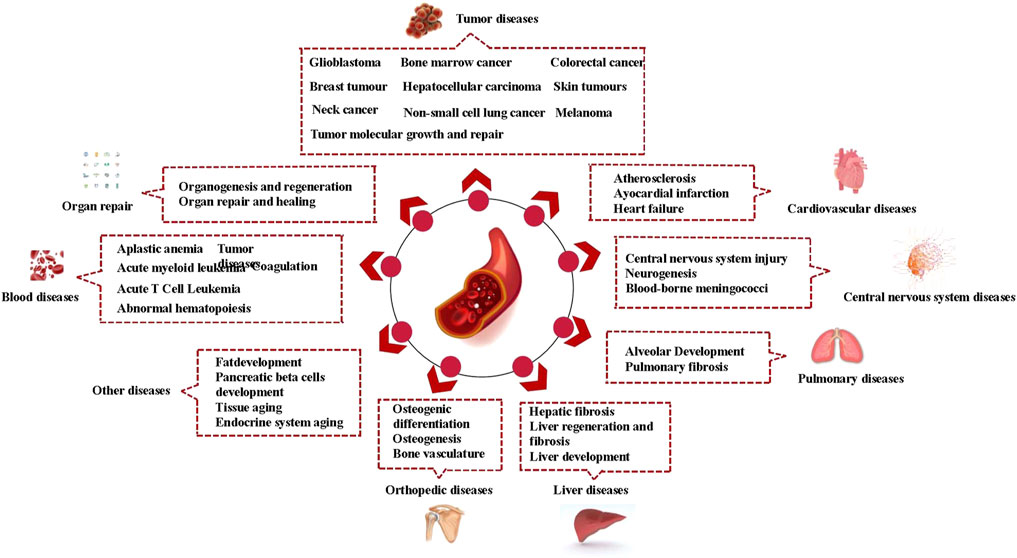
FIGURE 1. Vascular endothelial niches and diseases.
Tight junctions exist between vascular endothelial cells and epithelial cells often present as a continuous belt connecting adjacent cells closely together, forming a natural physical barrier between cells. Tight junctions are categorized as physical barriers to the vascular endothelial niche they surround. Linear shape along the junctions was reduced with less distinct connections in response to external and internal stimulation, with weaker tight junction protein signal intensity and volume (Lv and Fu, 2018). Colorectal cancer cells secreted exosome miR-25-3p, which targeted vascular endothelial growth factor (VEGF) receptor 2, zonula occludens 1, occludin, and claudin five on endothelial cells to induce vascular permeability and angiogenesis, forming a premetastatic niche for colorectal cancer metastasis (Zeng et al., 2018a). Additionally, in Lyl1−/− mice, adipose stem cell vascular niche impairment led to decreased pericyte coverage with loss of vascular endothelial (VE)-cadherin and zonula occludens 1, resulting in premature adipose stem cell vascular niche depletion (Hussain et al., 2021).
Factors expressed by endothelial cells mediate the effects of vascular endothelial niches. Cellular adhesion molecules mediate contact and binding between cells and the extracellular matrix. Adhesion molecule function results in cell-to-cell adhesion, cell-to-matrix adhesion, or cell-to-matrix adhesion by virtue of receptor–ligand binding. Adhesion molecules expressed within the vascular endothelial niche are involved in defense functions, inflammation, and tumor metastasis (Cheng et al., 2021). Adhesion molecules are divided into the integrin family, selectin family, immunoglobulin superfamily, and cadherin family according to their structural characteristics. In addition, some adhesion molecules have not yet been classified. Cadherin, selectin, and other adhesion families have been primarily reported to be involved in vascular endothelial niches (Cheng et al., 2021).
Cadherins comprise a family of calcium-dependent adhesion molecules. Endothelial cadherin is a major protein involved in vascular remodeling and vascular integrity. Expression of various adhesion molecules at high levels on the endothelium during inflammation established a premetastatic niche that lead to the recruitment of bone marrow precursors (Grigoryeva et al., 2020). Sphingosine-1-phosphate induced key adhesion proteins of VE-cadherin in pericytes that offset the drastic increase in vascular permeability, highlighting the importance of pericyte-endothelial interactions for vascular stabilization (AbdelRahman et al., 2021). Activation of high p53 expression by VE-cadherin in vascular endothelial cells induced a reduction in the number of perivascular mesenchymal stromal cells around hematopoietic stem cells, promoting hematopoietic stem cell aging and transformation (Si et al., 2018). Brain endothelial cells directly interacted with matriptase in the neural stem cell vascular niche to induce endothelial signaling that is sensitive to cholera toxin (Tung and Lee, 2017). Members of the selectin family recognized and selectively bound specific glycosylates, primarily mediating recognition and adhesion between leukocytes and vascular endothelial cells. The selectin family consisted of L-selectin, P-selectin, and E-selectin, which were mostly expressed on the surface of leukocytes, endothelial cells, and some tumor cells. Combination therapy with the selectin inhibitor GMI-1271 and imatinib prolonged survival in chronic myelocytic leukemia mice by reducing the contact time between leukemia cells and bone marrow endothelial cells (Godavarthy et al., 2020). Blockade of the adhesion molecule E-selectin, which was expressed exclusively by bone marrow endothelial cells, inhibited hematopoietic stem cell proliferation, self-renewal, and chemoresistance (Winkler et al., 2012). In addition to classic adhesion molecules, other adhesion molecules, such as junctional adhesion molecules, mediated endothelial progenitor cells during tube formation in the perivascular niche to reduce their adhesion to the tumor endothelium in vivo (Czabanka et al., 2020).
The critical function of chemokines is to manage the migration of hemameba to their specific position during equilibrium process when the process goes disease disorders. Chemokines directionally induce chemotaxis of nearby reactive cells through promoting proliferation of vascular endothelial cells and angiogenesis (Li et al., 2022). Vascular endothelial cells, chemokine (C-X-C motif) ligand 12 (CXCL12)-rich reticular cells, and mesenchymal stromal cell-regulated chemokines, cytokines, and cell surface adhesion molecules have been identified as microenvironmental cells in neoplastic hematopoiesis (Kaushansky and Zhan, 2018). CXCL12, localized within endothelial cells in the spleen niche, contributed to attracting differentiating hematopoietic cells to facilitate erythroblastic accumulation (Miwa et al., 2013). CXCLL2 deletion from the vascular endothelial vascular niche in T cell acute lymphoblastic leukemia impeded tumor growth (Pitt et al., 2015). The chemokine CXCL6 promoted angiogenesis to remodel the arteriolar niche in acute myeloid leukemia (Li et al., 2021).
a) Vascular endothelial cells can form channels in the blood vessel niche to facilitate invasion of molecules into specific tissues or organs. Under normal circumstances, vascular endothelium has its specific morphology and process unabnormal hyperplasia (Tracy et al., 2022). The research has uncovered that when body undergone disruption with tumor or inflammation, vascular endothelial cells proliferated to form a new vascular channel morphology. Endothelial cells in contact with glioblastoma stem cells form channels in the vessel niche to facilitate macrophage invasion into glioblastoma (Schiffer et al., 2018). RNAi-mediated attenuation of CD133 weakens CD144 (VE-cadherin) (+) melanoma cells, resulting in the formation of vessel-like channels and comprising the driving force for melanoma tumor growth (Lai et al., 2012).
b) Platelet derived growth factor (PDGF) is an alkaline protein stored in platelet α particles under normal physiological condition, while released and activated by disintegrating platelets during blood coagulation. It has the biological activity of stimulating chemotaxis and growth of specific cells (Luo et al., 2022). When suffered a severe injury of tissues, macrophage, vascular smooth muscle cells, fibroblasts, endothelial cells, or embryonic stem cells could also synthesis and release PDGF, exerting the function of chemotactic activity, division, phosphate enzyme activation, or prostaglandin metabolism. In a strong adhesion state, endothelial derived thrombospondin 1 gave rise to integrin aggregation and ligand integrin affinity modification. Endothelial derived thrombospondin 1 induced the activation of transforming growth factor β1, and periostin sustained a dormant niche for breast cancer cell quiescence (Ghajar et al., 2013). Thrombospondin 1 combound with hepatocyte growth factor, and laminins derived from islet endothelial cells played a critical role in the vascular endothelial niche in maintaining β-cell function and growth (Olerud et al., 2009).
c) VEGF is a highly specific growth factor involved in promoting vascular permeability, extracellular matrix degeneration, vascular endothelial cell migration, proliferation, and vascular formation (Lai et al., 2022). Given its critical role in mediating angiogenesis, VEGF is involved in the pathogenesis and progression of many angiogenesis dependent diseases, including cancer, inflammatory diseases, and diabetic retinopathy. Inhibition of VEGF stem cells residing in vascular endothelial niches might effectively prolong the survival period in ependymoma cancer (Nambirajan et al., 2014). Tumor-derived VEGF created a perivascular niche for stimulating cancer stemness and renewal through angiogenesis in a paracrine manner (Beck et al., 2011). VEGF activated endothelial cells to create the vascular niche, which enabled leukemic cells to proliferate at a higher rate and increased leukemic adherence to endothelial cells (Poulos et al., 2014). Angiogenesis and the expression of Akt-mediated angiogenic cytokines in R2 porcine hearts induce niches beneficial to cardiac repair (Zhang et al., 2012). VEGF influenced the vascular niche by regulating angiogenic factors that promoted the development of pancreatic beta cells (Lui, 2014). VEGF- and brain-derived neurotrophic factor mediated crosstalk between neural stem cells and endothelial cells to mold the neurovascular niche causing powerful blood vessel formation and maintenance (Li et al., 2006). Helium preconditioning promoted angiogenesis by elevating the mRNA and protein expression of VEGF and angiopoietin 1 to improve the focal neurovascular niche in a neonatal rat hypoxia/ischemia brain injury model (Li et al., 2016). Other growth factors were more involved in central nervous system diseases. Growth differentiation factor 11 enhanced neurogenesis, maintaining the integrity of the cerebrovascular niche in response to the changes that occurred during aging (Katsimpardi et al., 2014). The interaction between neurogenesis and angiogenesis was related to angiogenic matrix derived factor 1 and angiopoietin 1, which promoted neuroblast migration and behavioral recovery after stroke (Ohab et al., 2006).
Dysfunctional vascular endothelial cells could promote the aggregation of inflammatory factors (Yamagata, 2019), while endothelial oxidative stress could also cause inflammatory responses to vascular endothelial cells (Marchio et al., 2019), the inflammatory factors and oxidative stress reinforcing each other. The protein kinase CK2 was involved in the perivascular resistant niche stimulation under ionizing radiation, while secreting the cytokines interleukin 8 (IL-8) and IL-6, leading to resistance to radiation in non-small cell lung cancer cells (Li et al., 2019). Endothelial cell derived IL-6 activated the IL-6 receptor and signal transducer and activator of transcription 3, promoting self-renewal of dental pulp stem cells (Oh et al., 2020). A three dimensional model of endothelial cells and cancer stem like cells generated the conditional niche through elevated levels of the IL-8 and IL-8 homologous receptors CXCR1/2, which enhanced the migration, growth, and dryness characteristics of cancer stem-like cells (Infanger et al., 2013). Extracellular proteins from endothelial cells, such as IL-23, IL-17ɑ, dipeptidyl peptidase-4, and recombinant cystatin 3, organized the prometastatic molecular response in regeneration potentiated melanoma (Prakash et al., 2019). Pericytes could increase adenosine, nitric oxide, IL-10, TGF-β1 (Transforming growth factor beta 1), and MHC-II (Major histocompatibility complex-II) levels, which participated in melanoma cell extravasation (Caporarello et al., 2019). The reactive oxygen species-induced endothelial niche might play an important role in the development of pulmonary fibrosis via the regulation of pericytes and Wnt signaling (Andersson-Sjöland et al., 2016). High nitric oxide activity was observed in the tumor vascular endothelium adjacent to perivascular glioma cells and promoted stem-like characteristics in glioma cells (Charles et al., 2010).
Several hypoxia genes stabilize the internal environment of cells under hypoxia conditions to adapt to hypoxia condition (Jing et al., 2022), while they are involved in many physiological and pathological environments, such as placental development, tumor development, and metastasis. Hypoxia directly damages endothelial cells by destroying the cytoskeleton and intercellular connections, as well as disrupting metabolic and synthetic function, increasing the permeability of endothelial cells. The hypoxic niche of vascular endothelial cells promoted stemness maintenance and tumor propagation in cancer stem cells (Turpin et al., 2015). HIF-1 alpha (Hypoxia inducible factor 1 subunit alpha) mediated responses to hypoxia in central nervous system neurovascular niches inducing several signaling molecules, including BDNF (Brain derived neurotrophic factor), VEGF, and stromal cell-derived factor 1, which were involved in orchestrated angiogenesis and neurogenesis (Madri, 2009). Hypoxic and highly angiogenic areas formed a resulting niche of endothelial progenitor cells, while this particular niche might be related to liver cirrhosis (Yu et al., 2010).
Several gene alterations in vascular endothelial genes also affect the function of vascular endothelial cells as an ecological niche. Jagged2 (Jag2) was identified as a ligand of Notch receptor in multiple myeloma patient origin specimens. A lack of Jag2 in the perisinusoidal endothelial niche accelerated aging of hematopoietic stem cells (Saçma et al., 2019). JAK2V617F have shown high mutation rate in myeloproliferative disorders. Endothelial cells with the JAK2V617F mutation was an essential component of the hematopoietic vascular niche and was involved in the pathogenesis of myeloproliferative neoplasms (Lin et al., 2018). Endothelial cells with the kinase mutation JAK2V617F exhibited upregulated expression of CXCL12, and stem cell factors promoted clonal expansion in myeloproliferative neoplasms (Zhan et al., 2018b). Among the subtypes of gene KRAS, KRAS G12D is a common submutation, found in colorectal cancer, pancreatic cancer, and non-small cell lung cancer. Specifically, oncogenic KRas G12D mutations in adult endothelial cells significantly increased leukocytes and bone marrow cells in the blood of mice (Hochstetler et al., 2019). Ephrin receptors make up the largest subgroup of the receptor tyrosine kinase family, the protein encoded by ephrin type-B receptor 4 (EPHB4) plays an essential role in vascular development. EphB4 in the forebrain neurogenic niche participated in the process of vascular, molecular and structural remodeling (Colín-Castelán et al., 2011). Hepatocyte growth factor (HGF) was discovered as a substance that stimulated the proliferation of liver cells. HGF knockout mice displayed abnormally upregulated perivascular NOX4 expression near endothelial cell-induced niches, which promoted regeneration of mouse and human parenchymal cells in damaged organs (Cao et al., 2017). GATA Binding Protein 4 (GATA4) is an important transcription factor in the regulation of gene expression. Deletion of the transcription factor GATA4 in liver sinusoidal endothelial niches caused liver hypoplasia, fibrosis, and impaired colonization by hematopoietic progenitor cells through angiocrine factors (Géraud C. et al., 2017). Receptor activity modifying proteins serve as oligomeric modulators for numerous G-protein coupled receptors. Conditioned knockout of mouse endothelial receptor activity modifying protein 2 facilitated pulmonary endothelial cellular deformation and inflammatory infiltration to mediate the formation of premetastatic niches that ultimately promoted tumor metastasis (Tanaka et al., 2016).
The platelet reactive protein family is a group of structurally related secreted proteins that are widely distributed in the extracellular matrix of various tissues and inhibit angiogenesis. The β mural PDGF receptor was subsequently reprogrammed into NeuN+ local interneurons to hasten neurogenesis (Farahani et al., 2019). Notch signaling activated PDGF receptor-β positive perivascular cells, leading to the expansion of hematopoietic stem cell niches in bone (Kusumbe et al., 2016). Several signaling or bioactive molecules were secreted by endothelial cells in peripheral or local tissues. Capillary endothelial cells supplied paracrine factors, called angiocrine factors, to adjacent cells in the niche to orchestrate these processes. Endothelial cells regulated hematopoietic stem cell maintenance and regeneration of organ-specific stem cells through endothelial cell-derived paracrine factors (Sasine et al., 2017). Additionally, activation of endothelial cells or endoglin indicated a change in the vascular endothelial niche. Endothelial progenitor cells secreted proangiogenic factors to participate in angiogenesis, particularly under ischemic conditions (Yan et al., 2017). Upregulation of CD105 was associated with activated endothelium, affecting vascular tissue associated acute myelocytic leukemia cells and interactively affecting each other (Cogle et al., 2014). Runt-related genes in the vascular niche revealed that the vascular niche regulated the engraftment of acute myeloid leukemia cells in the bone marrow, influencing the overall survival of leukemic mice (Morita et al., 2018). Soluble amyloid precursor protein from endothelial cells triggered vascular niche functions that negatively regulated growth and restricted the number of neural stem cells in the subventricular zone (Sato et al., 2017).
The vascular endothelial niche is a potential decisive and limiting factor for many pathological processes. Vascular endothelial niches have experienced dynamic changes during the psychological process between health and disease condition. In the health condition, vascular endothelial cells maintained the basic function of blood vessels. Although vascular endothelial cells were considered as passive conduits, progress in vascular endothelial niches research now suggested that vascular endothelial cells were actively involved in the pathophysiological processes of body. The cells of the vascular endothelial cells, are rather active multifunctional team players that could mutually interact with neighboring and circulating cells in complex disease condition. However, there have been few reports exploring the development of diseases from the perspective of the vascular endothelial niche. In the past, a variety of techniques and models have been used to explore the influence of the vascular endothelium on tissues and their related diseases. Using the co-culture system, isolated endothelial cells or blood vessels were found to affect the proliferation and differentiation of other cells and tissues (Shen et al., 2004; Wurmser et al., 2004). Alternatively, vascular endothelial conventional conditional knockdown mice could be used as facilitating devices (Singhal et al., 2021a). Tumor-derived exosomes were additionally added to endothelial cells to investigate their functions in branching and inflammation within the perivascular niche (Rodrigues et al., 2019). Single-cell transcriptomics has been recently applied to trace cell–cell communication within the vascular endothelial niche (Singhal et al., 2021b). Additionally, 3D microfluidic chips have been employed to simulate the microenvironmental characteristics of niches (Zheng et al., 2021).
Vascular endothelial cells are not simply a large group of identical cells, and the phenotypes of endothelial cells exhibit obvious heterogeneity. Vascular endothelial cells from different species and with diverse diameters have disparate phenotypes with respect to their structure, function, and surface molecules. The heterogeneity of endothelial cells is manifested in cell morphology, function, gene expression, antigen synthesis, and niches. The mechanism of heterogeneity might be related to internal factors of cellular gene modification or external factors induced by the extracellular microenvironment. It has been found that the intrinsic factors of endothelial cell heterogeneity are related to the modification of site-specific genes, which regulate the lineage differentiation and formation of endothelial cells. Stolz and Sims-Lucus (Stolz and Sims-Lucas, 2015) believed that endothelial cells in different arteries, or even endothelial cells in different positions within the same artery, exhibiting both partially overlapping and distinct molecular profiles. Chi (Chi et al., 2003) indicated that 14 different loci of arteriovenous endothelial cells and microvascular endothelial cells were found to exhibit heterogeneity in the expression of various genes related to cell function using DNA microarray technology. Microenvironmental factors are external factors regulating the maintenance of endothelial cell specificity, and when the cell microenvironment changes, the cell phenotype also changes accordingly. Microenvironmental factors include both biomechanical signals and biochemical signals. Biomechanical signals comprise shear force and tension of blood flow, while biochemical signals include growth factors, cytokines, hormones, nucleosides, complement, lipoproteins, and extracellular matrix components.
Because vascular tissues are distributed throughout the body, the vascular endothelium acts as the lining of the vascular niche of the body. Vascular endothelial cells can sense and respond to external environmental signals, triggering a series of nonlinear response processes that ultimately lead to posttranscriptional modification or expression changes in genes (Figure 2). With respect to vascular endothelial cells in tissues that cannot be easily obtained, similar cells or tissues could be obtained through heterogeneity stimulation experiments, which is conducive to understanding and diagnosing the pathological mechanisms of various diseases.
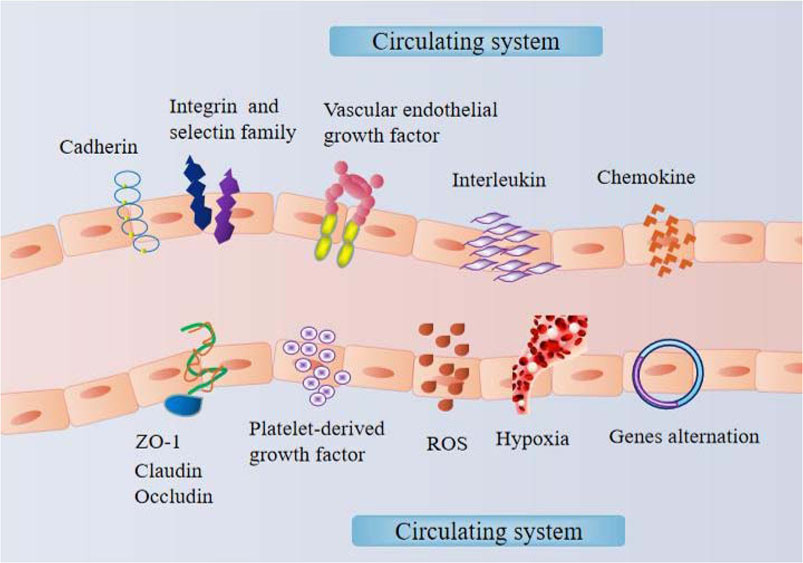
FIGURE 2. The mechanism within the vascular endothelial niche.
Given that the vascular niche is involved in multiple diseases, biomarkers in the vascular endothelial niche may serve as targets for therapy in various diseases. By predicting biomarkers in the vascular endothelial niche in advance, the threshold of disease intervention could be moved forward, providing targeted prevention and treatment for disease intervention. Vascular endothelial cells might represent a new breakthrough point for diseases, especially drug therapy targeting specific sites of blood vessels, exhibiting good clinical prospect applications.
All authors made the equal contribution to data collection, data collation, drafting, editing.
This work was supported by the Foundation Project: National Natural Science Foundation of China (No: 82100417, 81760094); The Foundation of Jiangxi Provincial Department of Science and Technology Youth Key Project (No. 20202ACBL206001, No. 20212BAB206022); Youth Project of Jiangxi Provincial Department of Education (No: GJJ200217).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Abdel Rahman F., d'Almeida S., Zhang T., Asadi M., Bozoglu T., Bongiovanni D., et al. (2021). Sphingosine-1-Phosphate Attenuates Lipopolysaccharide-Induced Pericyte Loss via Activation of Rho-A and MRTF-A. Thromb. Haemost. 121 (3), 341–350. doi:10.1055/s-0040-1716844
Abkowitz J. L., Robinson A. E., Kale S., Long M. W., Chen J. (2003). Mobilization of Hematopoietic Stem Cells during Homeostasis and after Cytokine Exposure. Blood 102 (4), 1249–1253. doi:10.1182/blood-2003-01-0318
Andersson-Sjöland A., Karlsson J. C., Rydell-Törmänen K. (2016). ROS-induced Endothelial Stress Contributes to Pulmonary Fibrosis through Pericytes and Wnt Signaling. Lab. Invest. 96 (2), 206–217. doi:10.1038/labinvest.2015.100
Beck B., Driessens G., Goossens S., Youssef K. K., Kuchnio A., Caauwe A., et al. (2011). A Vascular Niche and a VEGF-Nrp1 Loop Regulate the Initiation and Stemness of Skin Tumours. Nature 478 (7369), 399–403. doi:10.1038/nature10525
Butler J. M., Kobayashi H., Rafii S. (2010). Instructive Role of the Vascular Niche in Promoting Tumour Growth and Tissue Repair by Angiocrine Factors. Nat. Rev. Cancer 10 (2), 138–146. doi:10.1038/nrc2791
Cao Z., Ye T., Sun Y., Ji G., Shido K., Chen Y., et al. (2017). Targeting the Vascular and Perivascular Niches as a Regenerative Therapy for Lung and Liver Fibrosis. Sci. Transl. Med. 9 (405), eaai8710. doi:10.1126/scitranslmed.aai8710
Capel E., Barnier J.-P., Zomer A. L., Bole-Feysot C., Nussbaumer T., Jamet A., et al. (2017). Peripheral Blood Vessels Are a Niche for Blood-Borne Meningococci. Virulence 8 (8), 1808–1819. doi:10.1080/21505594.2017.1391446
Caporarello N., D’Angeli F., Cambria M. T., Candido S., Giallongo C., Salmeri M., et al. (2019). Pericytes in Microvessels: From "Mural" Function to Brain and Retina Regeneration. Ijms 20 (24), 6351. doi:10.3390/ijms20246351
Charles N., Ozawa T., Squatrito M., Bleau A.-M., Brennan C. W., Hambardzumyan D., et al. (2010). Perivascular Nitric Oxide Activates Notch Signaling and Promotes Stem-like Character in PDGF-Induced Glioma Cells. Cell Stem Cell 6 (2), 141–152. doi:10.1016/j.stem.2010.01.001
Chen J., Lippo L., Labella R., Tan S. L., Marsden B. D., Dustin M. L., et al. (2021a). Decreased Blood Vessel Density and Endothelial Cell Subset Dynamics during Ageing of the Endocrine System. EMBO J. 40 (1), e105242. doi:10.15252/embj.2020105242
Chen J., Sivan U., Tan S. L., Lippo L., De Angelis J., Labella R., et al. (2021b). High-resolution 3D Imaging Uncovers Organ-specific Vascular Control of Tissue Aging. Sci. Adv. 7 (6), eabd7819. doi:10.1126/sciadv.abd7819
Cheng P., Cao T., Zhao X., Lu W., Miao S., Ning F., et al. (2022). Nidogen1-enriched Extracellular Vesicles Accelerate Angiogenesis and Bone Regeneration by Targeting Myosin-10 to Regulate Endothelial Cell Adhesion. Bioactive Mater. 12, 185–197. doi:10.1016/j.bioactmat.2021.10.021
Chi J.-T., Chang H. Y., Haraldsen G., Jahnsen F. L., Troyanskaya O. G., Chang D. S., et al. (2003). Endothelial Cell Diversity Revealed by Global Expression Profiling. Proc. Natl. Acad. Sci. U.S.A. 100 (19), 10623–10628. doi:10.1073/pnas.1434429100
Cogle C. R., Goldman D. C., Madlambayan G. J., Leon R. P., Al Masri A., Clark H. A., et al. (2014). Functional Integration of Acute Myeloid Leukemia into the Vascular Niche. Leukemia 28 (10), 1978–1987. doi:10.1038/leu.2014.109
Colín-Castelán D., Phillips-Farfán B. V., Gutiérrez-Ospina G., Fuentes-Farias A. L., Báez-Saldaña A., Padilla-Cortés P., et al. (2011). EphB4 Is Developmentally and Differentially Regulated in Blood Vessels throughout the Forebrain Neurogenic Niche in the Mouse Brain: Implications for Vascular Remodeling. Brain Res. 1383, 90–98. doi:10.1016/j.brainres.2011.01.110
Czabanka M., Petrilli L. L., Elvers-Hornung S., Bieback K., Albert Imhof B., Vajkoczy P., et al. (2020). Junctional Adhesion Molecule-C Mediates the Recruitment of Embryonic-Endothelial Progenitor Cells to the Perivascular Niche during Tumor Angiogenesis. Ijms 21 (4), 1209. doi:10.3390/ijms21041209
Ding B.-S., Cao Z., Lis R., Nolan D. J., Guo P., Simons M., et al. (2014). Divergent Angiocrine Signals from Vascular Niche Balance Liver Regeneration and Fibrosis. Nature 505 (7481), 97–102. doi:10.1038/nature12681
Farahani R. M., Rezaei‐Lotfi S., Simonian M., Xaymardan M., Hunter N. (2019). Neural Microvascular Pericytes Contribute to Human Adult Neurogenesis. J. Comp. Neurol. 527 (4), 780–796. doi:10.1002/cne.24565
Ferentinos P., Tsakirides C., Swainson M., Davison A., Martyn-St James M., Ispoglou T. (2022). The Impact of Different Forms of Exercise on Circulating Endothelial Progenitor Cells in Cardiovascular and Metabolic Disease. Eur. J. Appl. Physiol. 122 (4), 815–860. doi:10.1007/s00421-021-04876-1
Friedman-Levi Y., Liraz-Zaltsman S., Shemesh C., Rosenblatt K., Kesner E. L., Gincberg G., et al. (2021). Pharmacological Blockers of CCR5 and CXCR4 Improve Recovery after Traumatic Brain Injury. Exp. Neurol. 338, 113604. doi:10.1016/j.expneurol.2021.113604
Géraud C., Koch P.-S., Goerdt S. (2014). Vascular Niches: Endothelial Cells as Tissue- and Site-specific Multifunctional Team Players in Health and Disease. JDDG: J. der Deutschen Dermatologischen Gesellschaft 12 (8), 685–689. doi:10.1111/ddg.12402
Géraud C., Koch P.-S., Zierow J., Klapproth K., Busch K., Olsavszky V., et al. (2017). GATA4-dependent Organ-specific Endothelial Differentiation Controls Liver Development and Embryonic Hematopoiesis. J. Clin. Invest. 127 (3), 1099–1114. doi:10.1172/JCI90086
Ghajar C. M., Peinado H., Mori H., Matei I. R., Evason K. J., Brazier H., et al. (2013). The Perivascular Niche Regulates Breast Tumour Dormancy. Nat. Cel Biol 15 (7), 807–817. doi:10.1038/ncb2767
Godavarthy P. S., Kumar R., Herkt S. C., Pereira R. S., Hayduk N., Weissenberger E. S., et al. (2020). The Vascular Bone Marrow Niche Influences Outcome in Chronic Myeloid Leukemia via the E-Selectin - SCL/TAL1 - CD44 axis. Haematologica 105 (1), 136–147. doi:10.3324/haematol.2018.212365
Grigoryeva E. S., Savelieva O. E., Popova N. O., Cherdyntseva N. V., Perelmuter V. M. (2020). Do tumor Exosome Integrins Alone Determine Organotropic Metastasis? Mol. Biol. Rep. 47 (10), 8145–8157. doi:10.1007/s11033-020-05826-4
Ho I. A. W., Shim W. S. N. (2017). Contribution of the Microenvironmental Niche to Glioblastoma Heterogeneity. Biomed. Res. Int. 2017, 1–13. doi:10.1155/2017/9634172
Hochstetler C. L., Feng Y., Sacma M., Davis A. K., Rao M., Kuan C.-Y., et al. (2019). KRasG12D Expression in the Bone Marrow Vascular Niche Affects Hematopoiesis with Inflammatory Signals. Exp. Hematol. 79, 3–15. e4. doi:10.1016/j.exphem.2019.10.003
Hoshizaki K., Takahashi S., Tanaka H., Oki S., Matsushita M. (2022). Stochasticity of Individual Competition and Local Matchup Inequality for Saplings in a Niche‐structured forest. Ecology 103 (4), e3624. doi:10.1002/ecy.3624
Hussain A., Deleuze V., El Kebriti L., Turali H., Pirot N., Glasson Y., et al. (2021). In Lyl1 −/− Mice, Adipose Stem Cell Vascular Niche Impairment Leads to Premature Development of Fat Tissues. Stem Cells 39 (1), 78–91. doi:10.1002/stem.3286
Infanger D. W., Cho Y., Lopez B. S., Mohanan S., Liu S. C., Gursel D., et al. (2013). Glioblastoma Stem Cells Are Regulated by Interleukin-8 Signaling in a Tumoral Perivascular Niche. Cancer Res. 73 (23), 7079–7089. doi:10.1158/0008-5472.CAN-13-1355
Jackson W. F. (2022). Endothelial Ion Channels and Cell-Cell Communication in the Microcirculation. Front. Physiol. 13, 805149. doi:10.3389/fphys.2022.805149
Jing J., Jiang H., Zhang L. (2022). Endothelial Progenitor Cells Promote Neural Stem Cell Proliferation in Hypoxic Conditions through VEGF via the PI3K/AKT Pathway. J. Receptors Signal Transduction 2022, 1–7. doi:10.1080/10799893.2021.2019275
Katsimpardi L., Litterman N. K., Schein P. A., Miller C. M., Loffredo F. S., Wojtkiewicz G. R., et al. (2014). Vascular and Neurogenic Rejuvenation of the Aging Mouse Brain by Young Systemic Factors. Science 344 (6184), 630–634. doi:10.1126/science.1251141
Kaushansky K., Zhan H. (2018). The Regulation of normal and Neoplastic Hematopoiesis Is Dependent on Microenvironmental Cells. Adv. Biol. Regul. 69, 11–15. doi:10.1016/j.jbior.2018.06.003
Kopp H.-G., Avecilla S. T., Hooper A. T., Rafii S. (2005). The Bone Marrow Vascular Niche: Home of HSC Differentiation and Mobilization. Physiology 20, 349–356. doi:10.1152/physiol.00025.2005
Kumar N., Saraber P., Ding Z., Kusumbe A. P. (2021). Diversity of Vascular Niches in Bones and Joints during Homeostasis, Ageing, and Diseases. Front. Immunol. 12, 798211. doi:10.3389/fimmu.2021.798211
Kusumbe A. P., Ramasamy S. K., Itkin T., Mäe M. A., Langen U. H., Betsholtz C., et al. (2016). Age-dependent Modulation of Vascular Niches for Haematopoietic Stem Cells. Nature 532 (7599), 380–384. doi:10.1038/nature17638
Lai C.-Y., Schwartz B. E., Hsu M.-Y. (2012). CD133+ Melanoma Subpopulations Contribute to Perivascular Niche Morphogenesis and Tumorigenicity through Vasculogenic Mimicry. Cancer Res. 72 (19), 5111–5118. doi:10.1158/0008-5472.CAN-12-0624
Li L., Man J., Zhao L. (2021). Hypoxia-CXCL6 axis Affects Arteriolar Niche Remodeling in Acute Myeloid Leukemia. Exp. Biol. Med. (Maywood) 246 (1), 84–96. doi:10.1177/1535370220960675
Li L., Xie T. (2005). Stem Cell Niche: Structure and Function. Annu. Rev. Cel Dev. Biol. 21, 605–631. doi:10.1146/annurev.cellbio.21.012704.131525
Li Q., Ford M. C., Lavik E. B., Madri J. A. (2006). Modeling the Neurovascular Niche: VEGF- and BDNF-Mediated Cross-Talk between Neural Stem Cells and Endothelial Cells: an In Vitro Study. J. Neurosci. Res. 84 (8), 1656–1668. doi:10.1002/jnr.21087
Li Q., Zong Y., Li K., Jie X., Hong J., Zhou X., et al. (2019). Involvement of Endothelial CK2 in the Radiation Induced Perivascular Resistant Niche (PVRN) and the Induction of Radioresistance for Non-small Cell Lung Cancer (NSCLC) Cells. Biol. Res. 52 (1), 22. doi:10.1186/s40659-019-0231-x
Li W., Terada Y., Tyurina Y. Y., Tyurin V. A., Bery A. I., Gauthier J. M., et al. (2022). Necroptosis Triggers Spatially Restricted Neutrophil-Mediated Vascular Damage during Lung Ischemia Reperfusion Injury. Proc. Natl. Acad. Sci. U.S.A. 119 (10), e2111537119. doi:10.1073/pnas.2111537119
Li Y., Zhang P., Liu Y., Liu W., Yin N. (2016). Helium Preconditioning Protects the Brain against Hypoxia/ischemia Injury via Improving the Neurovascular Niche in a Neonatal Rat Model. Behav. Brain Res. 314, 165–172. doi:10.1016/j.bbr.2016.08.015
Lin C. H. S., Zhang Y., Kaushansky K., Zhan H. (2018). JAK2V617F-bearing Vascular Niche Enhances Malignant Hematopoietic Regeneration Following Radiation Injury. Haematologica 103 (7), 1160–1168. doi:10.3324/haematol.2017.185736
Lo A. Y., Lai A. W., Ng T., Hung V. L., Tam K., Cheung C. S., et al. (2022). Exacerbated VEGF Up-Regulation Accompanies Diabetes-Aggravated Hemorrhage in Mice after Experimental Cerebral Ischemia and Delayed Reperfusion. Neural Regen. Res. 17 (7), 1566–1575. doi:10.4103/1673-5374.330612
Lui K. (2014). VEGF-A: the Inductive Angiogenic Factor for Development, Regeneration and Function of Pancreatic Beta Cells. Cscr 9 (5), 396–400. doi:10.2174/1574888x09666140710100603
Luo L.-l., Han J.-x., Wu S.-r., Kasim V. (2022). Intramuscular Injection of Sotagliflozin Promotes Neovascularization in Diabetic Mice through Enhancing Skeletal Muscle Cells Paracrine Function. Acta Pharmacol. Sin. Online ahead of print. doi:10.1038/s41401-022-00889-4
Lv Y., Fu L. (2018). The Potential Mechanism for Hydroxysafflor Yellow A Attenuating Blood-Brain Barrier Dysfunction via Tight junction Signaling Pathways Excavated by an Integrated Serial Affinity Chromatography and Shotgun Proteomics Analysis Approach. Neurochem. Int. 112, 38–48. doi:10.1016/j.neuint.2017.10.012
Madri J. A. (2009). Modeling the Neurovascular Niche: Implications for Recovery from CNS Injury. J. Physiol. Pharmacol. 60 (Suppl. 4), 95–104. doi:10.1013/2009/51441418
Mammoto A., Mammoto T. (2019). Vascular Niche in Lung Alveolar Development, Homeostasis, and Regeneration. Front. Bioeng. Biotechnol. 7, 318. doi:10.3389/fbioe.2019.00318
Marchio P., Guerra-Ojeda S., Vila J. M., Aldasoro M., Victor V. M., Mauricio M. D. (2019). Targeting Early Atherosclerosis: a Focus on Oxidative Stress and Inflammation. Oxid Med. Cel Longev 2019, 8563845. doi:10.1155/2019/8563845
Miwa Y., Hayashi T., Suzuki S., Abe S., Onishi I., Kirimura S., et al. (2013). Up-regulated Expression of CXCL12 in Human Spleens with Extramedullary Haematopoiesis. Pathology 45 (4), 408–416. doi:10.1097/PAT.0b013e3283613dbf
Morita K., Tokushige C., Maeda S., Kiyose H., Noura M., Iwai A., et al. (2018). RUNX Transcription Factors Potentially Control E-Selectin Expression in the Bone Marrow Vascular Niche in Mice. Blood Adv. 2 (5), 509–515. doi:10.1182/bloodadvances.2017009324
Nambirajan A., Sharma M. C., Gupta R. K., Suri V., Singh M., Sarkar C. (2014). Study of Stem Cell Marker Nestin and its Correlation with Vascular Endothelial Growth Factor and Microvascular Density in Ependymomas. Neuropathol. Appl. Neurobiol. 40 (6), 714–725. doi:10.1111/nan.12097
Nguyen T. S., Lapidot T., Ruf W. (2018). Extravascular Coagulation in Hematopoietic Stem and Progenitor Cell Regulation. Blood 132 (2), 123–131. doi:10.1182/blood-2017-12-768986
Nikolova G., Strilic B., Lammert E. (2007). The Vascular Niche and its Basement Membrane. Trends Cel Biol. 17 (1), 19–25. doi:10.1016/j.tcb.2006.11.005
Oh M., Zhang Z., Mantesso A., Oklejas A. E., Nör J. E. (2020). Endothelial-Initiated Crosstalk Regulates Dental Pulp Stem Cell Self-Renewal. J. Dent Res. 99 (9), 1102–1111. doi:10.1177/0022034520925417
Ohab J. J., Fleming S., Blesch A., Carmichael S. T. (2006). A Neurovascular Niche for Neurogenesis after Stroke. J. Neurosci. 26 (50), 13007–13016. doi:10.1523/JNEUROSCI.4323-06.2006
Olerud J., Johansson Å., Carlsson P.-O. (2009). Vascular Niche of Pancreatic Islets. Expert Rev. Endocrinol. Metab. 4 (5), 481–491. doi:10.1586/eem.09.25
Owen-Woods C., Kusumbe A. (2022). Fundamentals of Bone Vasculature: Specialization, Interactions and Functions. Semin. Cel Develop. Biol. 123, 36–47. doi:10.1016/j.semcdb.2021.06.025
Palikuqi B., Nguyen D.-H. T., Li G., Schreiner R., Pellegata A. F., Liu Y., et al. (2020). Adaptable Haemodynamic Endothelial Cells for Organogenesis and Tumorigenesis. Nature 585 (7825), 426–432. doi:10.1038/s41586-020-2712-z
Pasquier J., Ghiabi P., Chouchane L., Razzouk K., Rafii S., Rafii A. (2020). Angiocrine Endothelium: from Physiology to Cancer. J. Transl Med. 18 (1), 52. doi:10.1186/s12967-020-02244-9
Pitt L. A., Tikhonova A. N., Hu H., Trimarchi T., King B., Gong Y., et al. (2015). CXCL12-Producing Vascular Endothelial Niches Control Acute T Cell Leukemia Maintenance. Cancer Cell 27 (6), 755–768. doi:10.1016/j.ccell.2015.05.0020
Poller W. C., Nahrendorf M., Swirski F. K. (2020). Hematopoiesis and Cardiovascular Disease. Circ. Res. 126 (8), 1061–1085. doi:10.1161/CIRCRESAHA.120.315895
Poulos M. G., Gars E. J., Gutkin M. C., Kloss C. C., Ginsberg M., Scandura J. M., et al. (2014). Activation of the Vascular Niche Supports Leukemic Progression and Resistance to Chemotherapy. Exp. Hematol. 42 (11), 976–986. doi:10.1016/j.exphem.2014.08.003
Psaltis P. J., Harbuzariu A., Delacroix S., Holroyd E. W., Simari R. D. (2011). Resident Vascular Progenitor Cells-Diverse Origins, Phenotype, and Function. J. Cardiovasc. Trans. Res. 4 (2), 161–176. doi:10.1007/s12265-010-9248-9
Qian C., Schoemaker R. G., van Gilst W. H., Yu B., Roks A. J. M. (2008). Regenerative Cell Therapy and Pharmacotherapeutic Intervention in Heart Failure. Nhjl 16 (9), 305–309. doi:10.1007/BF03086169
Rafii S., Butler J. M., Ding B.-S. (2016). Angiocrine Functions of Organ-specific Endothelial Cells. Nature 529 (7586), 316–325. doi:10.1038/nature17040
Ribatti D., Basile A., Ruggieri S., Vacca A. (2014). Bone Marrow Vascular Niche and the Control of Angiogenesis in Multiple Myeloma. Front. Biosci. 19, 304–311. doi:10.2741/4209
Ribatti D., Tamma R., Annese T. (2021). The Role of Vascular Niche and Endothelial Cells in Organogenesis and Regeneration. Exp. Cel Res. 398 (1), 112398. doi:10.1016/j.yexcr.2020.112398
Rodrigues G., Hoshino A., Kenific C. M., Matei I. R., Steiner L., Freitas D., et al. (2019). Tumour Exosomal CEMIP Protein Promotes Cancer Cell Colonization in Brain Metastasis. Nat. Cel Biol 21 (11), 1403–1412. doi:10.1038/s41556-019-0404-4
Rubalcaba J. G., Verberk W. C. E. P., Hendriks A. J., Saris B., Woods H. A. (2020). Oxygen Limitation May Affect the Temperature and Size Dependence of Metabolism in Aquatic Ectotherms. Proc. Natl. Acad. Sci. U.S.A. 117 (50), 31963–31968. doi:10.1073/pnas.2003292117
Saçma M., Pospiech J., Bogeska R., de Back W., Mallm J.-P., Sakk V., et al. (2019). Haematopoietic Stem Cells in Perisinusoidal Niches Are Protected from Ageing. Nat. Cel Biol 21 (11), 1309–1320. doi:10.1038/s41556-019-0418-y
Sasine J. P., Yeo K. T., Chute J. P. (2017). Concise Review: Paracrine Functions of Vascular Niche Cells in Regulating Hematopoietic Stem Cell Fate. Stem Cell Transl Med 6 (2), 482–489. doi:10.5966/sctm.2016-0254
Sato Y., Uchida Y., Hu J., Young-Pearse T. L., Niikura T., Mukouyama Y.-s. (2017). Soluble APP Functions as a Vascular Niche Signal that Controls Adult Neural Stem Cell Number. Development 144 (15), 2730–2736. doi:10.1242/dev.143370
Schiffer D., Annovazzi L., Casalone C., Corona C., Mellai M. (2018). Glioblastoma: Microenvironment and Niche Concept. Cancers 11 (1), 5. doi:10.3390/cancers11010005
Sharma A., Shiras A. (2016). Cancer Stem Cell-Vascular Endothelial Cell Interactions in Glioblastoma. Biochem. Biophysical Res. Commun. 473 (3), 688–692. doi:10.1016/j.bbrc.2015.12.022
Shen Q., Goderie S. K., Jin L., Karanth N., Sun Y., Abramova N., et al. (2004). Endothelial Cells Stimulate Self-Renewal and Expand Neurogenesis of Neural Stem Cells. Science 304 (5675), 1338–1340. doi:10.1126/science.1095505
Shido K., Chavez D., Cao Z., Ko J. L., Rafii S., Ding B.-S. (2017). Platelets Prime Hematopoietic-Vascular Niche to Drive Angiocrine-Mediated Liver Regeneration. Sig Transduct Target. Ther. 2, 16044. doi:10.1038/sigtrans.2016.44
Si S., Nakajima-Takagi Y., Iga T., Tsuji M., Hou L., Oshima M., et al. (2018). Hematopoietic Insults Damage Bone Marrow Niche by Activating P53 in Vascular Endothelial Cells. Exp. Hematol. 63, 41–51. doi:10.1016/j.exphem.2018.04.006
Singh A., Veeriah V., Xi P., Labella R., Chen J., Romeo S. G., et al. (2019). Angiocrine Signals Regulate Quiescence and Therapy Resistance in Bone Metastasis. JCI Insight 4 (13), e125679. doi:10.1172/jci.insight.125679
Singhal M., Augustin H. G. (2020). Beyond Angiogenesis: Exploiting Angiocrine Factors to Restrict Tumor Progression and Metastasis. Cancer Res. 80 (4), 659–662. doi:10.1158/0008-5472.CAN-19-3351
Singhal M., Gengenbacher N., Abdul Pari A. A., Kamiyama M., Hai L., Kuhn B. J., et al. (2021b). Temporal Multi-Omics Identifies LRG1 as a Vascular Niche Instructor of Metastasis. Sci. Transl Med. 13 (609), eabe6805. doi:10.1126/scitranslmed.abe6805
Singhal M., Gengenbacher N., Abdul Pari A. A., Kamiyama M., Hai L., Kuhn B. J., et al. (2021a). Temporal Multi-Omics Identifies LRG1 as a Vascular Niche Instructor of Metastasis. Sci. Transl. Med. 13 (609), eabe6805. doi:10.1126/scitranslmed.abe6805
Stolz D. B., Sims-Lucas S. (2015). Unwrapping the Origins and Roles of the Renal Endothelium. Pediatr. Nephrol. 30 (6), 865–872. doi:10.1007/s00467-014-2798-3
Tanaka M., Koyama T., Sakurai T., Kamiyoshi A., Ichikawa-Shindo Y., Kawate H., et al. (2016). The Endothelial Adrenomedullin-RAMP2 System Regulates Vascular Integrity and Suppresses Tumour Metastasis. Cardiovasc. Res. 111 (4), 398–409. doi:10.1093/cvr/cvw166
Tracy E. P., Stielberg V., Rowe G., Benson D., Nunes S. S., Hoying J. B., et al. (2022). State of the Field: Cellular and Exosomal Therapeutic Approaches in Vascular Regeneration. Am. J. Physiol. Heart Circ. Physiol. 322 (4), H647–H680. doi:10.1152/ajpheart.00674.2021
Tsai T. L., Wang B., Squire M. W., Guo L. W., Li W. J. (2015). Endothelial Cells Direct Human Mesenchymal Stem Cells for Osteo- and Chondro-Lineage Differentiation through Endothelin-1 and AKT Signaling. Stem Cel Res Ther 6 (1), 88. doi:10.1186/s13287-015-0065-6
Tung H. H., Lee S. L. (2017). Physical Binding of Endothelial MCAM and Neural Transmembrane Protease Matriptase-Novel Cell Adhesion in Neural Stem Cell Vascular Niche. Sci. Rep. 7 (1), 4946. doi:10.1038/s41598-017-05131-4
Turpin A., Sharif A., Stoven L., Blond S., Maurage C. A., Le Rhun É. (2015). La niche des cellules souches tumorales dans le glioblastome : des aspects fondamentaux au ciblage thérapeutique [The stem cell niche in glioblastoma: from fundamental aspects to targeted therapies]. Bull. Cancer 102 (1), 24–33. doi:10.1016/j.bulcan.2014.07.001
Willert K., Brown J. D., Danenberg E., Duncan A. W., Weissman I. L., Reya T., et al. (2003). Wnt Proteins Are Lipid-Modified and Can Act as Stem Cell Growth Factors. Nature 423 (6938), 448–452. doi:10.1038/nature01611
Winkler I. G., Barbier V., Nowlan B., Jacobsen R. N., Forristal C. E., Patton J. T., et al. (2012). Vascular Niche E-Selectin Regulates Hematopoietic Stem Cell Dormancy, Self Renewal and Chemoresistance. Nat. Med. 18 (11), 1651–1657. doi:10.1038/nm.2969
Wu L., Mo W., Zhang Y., Zhou M., Li Y., Zhou R., et al. (2017). Vascular and Perivascular Niches, but Not the Osteoblastic Niche, Are Numerically Restored Following Allogeneic Hematopoietic Stem Cell Transplantation in Patients with Aplastic Anemia. Int. J. Hematol. 106 (1), 71–81. doi:10.1007/s12185-017-2217-1
Wurmser A. E., Nakashima K., Summers R. G., Toni N., D'Amour K. A., Lie D. C., et al. (2004). Cell Fusion-independent Differentiation of Neural Stem Cells to the Endothelial Lineage. Nature 430 (6997), 350–356. doi:10.1038/nature02604
Yamagata K. (2019). Soy Isoflavones Inhibit Endothelial Cell Dysfunction and Prevent Cardiovascular Disease. J. Cardiovasc. Pharmacol. 74 (3), 201–209. doi:10.1097/FJC.0000000000000708
Yan X., Dai X., He L., Ling X., Shao M., Zhang C., et al. (2017). A Novel CXCR4 Antagonist Enhances Angiogenesis via Modifying the Ischaemic Tissue Environment. J. Cel Mol Med 21 (10), 2298–2307. doi:10.1111/jcmm.13150
Yu D. C., Chen J., Ding Y. T. (2010). Hypoxic and Highly Angiogenic Non-tumor Tissues Surrounding Hepatocellular Carcinoma: the 'niche' of Endothelial Progenitor Cells. Int. J. Mol. Sci. 11 (8), 2901–2909. doi:10.3390/ijms11082901
Zeng Z., Li Y., Pan Y., Lan X., Song F., Sun J., et al. (2018a). Cancer-derived Exosomal miR-25-3p Promotes Pre-metastatic Niche Formation by Inducing Vascular Permeability and Angiogenesis. Nat. Commun. 9 (1), 5395. doi:10.1038/s41467-018-07810-w
Zhan H., Lin C., Segal Y., Kaushansky K. (2018b). The JAK2V617F-Bearing Vascular Niche Promotes Clonal Expansion in Myeloproliferative Neoplasms. Leukemia 32 (2), 462–469. doi:10.1038/leu.2017.233
Zhang C. D., Liu M., Wang X. L., Chen S., Fu X. J., Li G., et al. (2022a). Mechanism of CircANKRD36 Regulating Cell Heterogeneity and Endothelial Mesenchymal Transition in Aortic Valve Stromal Cells by Regulating miR-599 and TGF-β Signaling Pathway. Int. J. Cardiol. 352, 104–114. doi:10.1016/j.ijcard.2022.01.043
Zhang S., Zhao L., Shen L., Xu D., Huang B., Wang Q., et al. (2012). Comparison of Various Niches for Endothelial Progenitor Cell Therapy on Ischemic Myocardial Repair: Coexistence of Host Collateralization and Akt-Mediated Angiogenesis Produces a superior Microenvironment. Arterioscler Thromb. Vasc. Biol. 32 (4), 910–923. doi:10.1161/ATVBAHA.111.244970
Zhang T., Lei M. L., Zhou H., Chen Z. Z., Peng Shi P. (2022b). Phylogenetic Relationships of the Zokor Genus Eospalax (Mammalia, Rodentia, Spalacidae) Inferred from Whole-Genome Analyses, with Description of a New Species Endemic to Hengduan Mountains. Zool Res. 43 (3), 331–342. doi:10.24272/j.issn.2095-8137.2022.045
Zhang Z., Dong Z., Lauxen I. S., Filho M. S., Nör J. E. (2014). Endothelial Cell-Secreted EGF Induces Epithelial to Mesenchymal Transition and Endows Head and Neck Cancer Cells with Stem-like Phenotype. Cancer Res. 74 (10), 2869–2881. doi:10.1158/0008-5472.CAN-13-2032
Keywords: vascular endothelial niche, tumor metastasis, blood, chemokine, adhesion molecules, tight junction, inflammatory factor
Citation: Lei Z, Hu X, Wu Y, Fu L, Lai S, Lin J, Li X and Lv Y (2022) The Role and Mechanism of the Vascular Endothelial Niche in Diseases: A Review. Front. Physiol. 13:863265. doi: 10.3389/fphys.2022.863265
Received: 27 January 2022; Accepted: 13 April 2022;
Published: 27 April 2022.
Edited by:
Francesco Moccia, University of Pavia, ItalyReviewed by:
Anjali P. Kusumbe, University of Oxford, United KingdomCopyright © 2022 Lei, Hu, Wu, Fu, Lai, Lin, Li and Lv. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Yanni Lv, eWFubmlsdjIyNUBuY3UuZWR1LmNu
†These authors have contributed equally to this work
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.