 Ye Yuan
Ye Yuan Marc-Antonio Padilla2
Marc-Antonio Padilla2 Dunham Clark
Dunham Clark Swathi Yadlapalli
Swathi Yadlapalli
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Physiol. , 09 November 2022
Sec. Chronobiology
Volume 13 - 2022 | https://doi.org/10.3389/fphys.2022.1051544
This article is part of the Research Topic Rising Stars in Chronobiology: 2022 View all 7 articles
Circadian clocks are ∼24-h timekeepers that control rhythms in almost all aspects of our behavior and physiology. While it is well known that subcellular localization of core clock proteins plays a critical role in circadian regulation, very little is known about the spatiotemporal organization of core clock mRNAs and its role in generating ∼24-h circadian rhythms. Here we describe a streamlined single molecule Fluorescence In Situ Hybridization (smFISH) protocol and a fully automated analysis pipeline to precisely quantify the number and subcellular location of mRNAs of Clock, a core circadian transcription factor, in individual clock neurons in whole mount Drosophila adult brains. Specifically, we used ∼48 fluorescent oligonucleotide probes that can bind to an individual Clock mRNA molecule, which can then be detected as a diffraction-limited spot. Further, we developed a machine learning-based approach for 3-D cell segmentation, based on a pretrained encoder-decoder convolutional neural network, to automatically identify the cytoplasm and nuclei of clock neurons. We combined our segmentation model with a spot counting algorithm to detect Clock mRNA spots in individual clock neurons. Our results demonstrate that the number of Clock mRNA molecules cycle in large ventral lateral clock neurons (lLNvs) with peak levels at ZT4 (4 h after lights are turned on) with ∼80 molecules/neuron and trough levels at ZT16 with ∼30 molecules/neuron. Our streamlined smFISH protocol and deep learning-based analysis pipeline can be employed to quantify the number and subcellular location of any mRNA in individual clock neurons in Drosophila brains. Further, this method can open mechanistic and functional studies into how spatiotemporal localization of clock mRNAs affect circadian rhythms.
Almost all organisms have evolved internal clocks to tell time. Circadian clocks are cell-autonomous timekeepers that can be entrained by light (Stanewsky et al., 1998) or temperature (Sehadova et al., 2009; Buhr et al., 2010; Yadlapalli et al., 2018) cycles. These clocks generate ∼24-h oscillations in the expression of many genes and control rhythms in sleep-wake cycles, metabolism, immunity (Hardin et al., 1990; Aronson et al., 1994; Sehgal et al., 1994; Patke et al., 2020). While past studies highlight the critical role of subcellular localization of core clock proteins in controlling circadian rhythms (Curtin et al., 1995; Shafer et al., 2002; Cohen et al., 2014; Zhao et al., 2015; Xiao et al., 2021; Yuan et al., 2022), what remains poorly understood is how clock mRNAs are organized over the circadian cycle and the role of such organization in controlling circadian rhythms. A major drawback of techniques such as qPCR and single-cell RNA-sequencing currently used to study circadian gene expression is lack of subcellular spatial information (Zhang et al., 2014; Ma et al., 2021). Single-molecule RNA Fluorescence In Situ Hybridization (smFISH) has recently emerged as a powerful technique which enables quantitative measurement of the number as well as the subcellular location of mRNA molecules in individual cells (Raj et al., 2008; Trcek et al., 2017). In one recent study, smFISH was adapted to visualize core clock transcripts in Neurospora crassa and it was reported that core clock mRNAs are clustered in the perinuclear cytoplasmic region in a time-of-day dependent manner (Bartholomai et al., 2022).
Drosophila melanogaster is a powerful model system for understanding spatiotemporal regulation of circadian rhythms, as it has a highly conserved clock along with powerful genetic and molecular tools (Dubowy and Sehgal, 2017; Patke et al., 2020). Further, the Drosophila clock network is relatively simple consisting of ∼150 clock neurons (Nitabach and Taghert, 2008; Dubowy and Sehgal, 2017) (Figure 1A). These clock neurons express the key clock proteins–CLOCK (CLK), CYCLE (CYC), PERIOD (PER) and TIMELESS (TIM), that form the core of the molecular clock (Patke et al., 2020) (Figure 1B). CLK and CYC act as the positive transcription factors and PER and TIM act as the core clock repressors. During the activation phase, CLK and CYC drive transcription of hundreds of target genes, including per and tim, by binding to the E-boxes in their promoter regions. PER and TIM, after a time delay, enter the nucleus and inhibit CLK/CYC activity thereby silencing their own expression as well as the expression of other clock-regulated genes. PER and TIM are then degraded leading to the end of the repression phase and the start of the activation phase of a new cycle (Patke et al., 2020). In addition to this core feedback loop, there exists a second interlocked feedback loop that controls rhythmic expression of Clk mRNA (Cyran et al., 2003; Glossop et al., 2003) (Figure 1B). CLK/CYC proteins bind to the E-boxes of the transcription factors, Vrille (VRI) and PAR domain protein 1ε (PDP1ε), creating rhythms in their mRNA with peak levels during early night and trough levels during early day. Interestingly, VRI and PDP1ε protein levels peak at different times during the night, with VRI peaking during the early evening and PDP1ε later in the night. VRI and PDP1ε proteins bind one after the other to the VRI/PDP1ε-binding boxes in the Clk enhancer to repress or activate Clk transcription, respectively, thus resulting in rhythmic Clk mRNA expression.
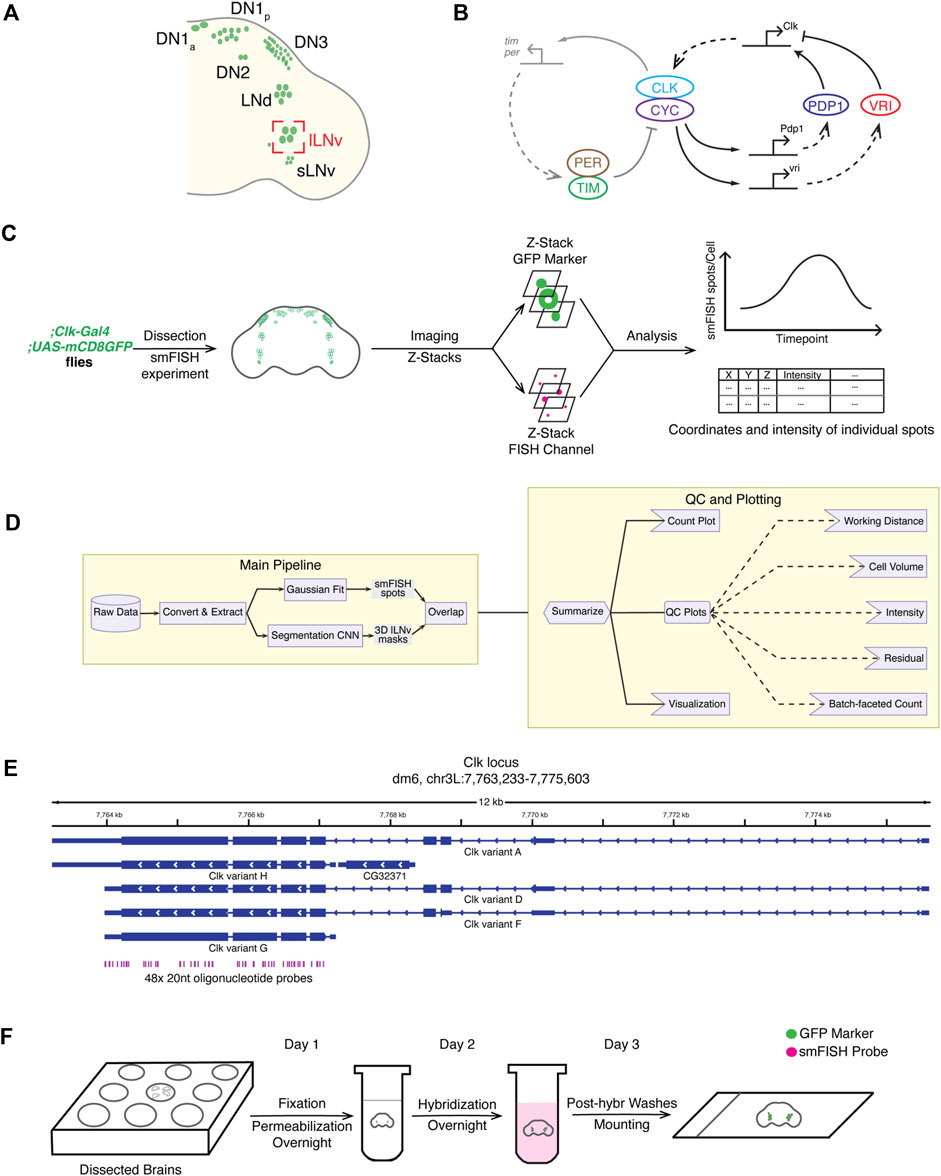
FIGURE 1. Overview of the smFISH method and the data analysis pipeline. (A) Schematic of different clock neuron groups in Drosophila melanogaster brains. Large ventrolateral neurons (lLNvs) are circled in red. (B) Schematic of the molecular clock in Drosophila. The Drosophila circadian clock is composed of two interlocked negative feedback loops. (C) Overview of the full workflow including smFISH experiment and data analysis pipeline. (D) Schematic of the data analysis pipeline. To quantify Clock smFISH spots in lLNvs, two tasks are performed in parallel: Detection of Clock smFISH spots and segmentation of lLNvs. (E) Genomic locations of oligonucleotide probes in the Clock locus. Probes are designed to hybridize with the shared exon sequences to visualize mRNAs of all annotated Clock transcript variants. (F) Schematic of the smFISH procedure.
Here, we describe a streamlined single molecule FISH (smFISH) protocol and a fully automated analysis pipeline to quantify the number and localization of mRNAs of Clock, a key clock transcription factor, in individual clock neurons in whole-mount Drosophila adult brains. We developed a simple, robust smFISH protocol that takes 3 days from brain dissections to slide preparation. Next, we generated an accurate three-dimensional segmentation model of clock neurons using machine learning-based approaches by adopting a pretrained decoder-encoder convolutional neural network model. Using these tools, we visualized and quantified Clock mRNA distribution in Large ventral lateral clock neurons (lLNvs), one of the main Drosophila clock neuron groups. Our results show that Clock mRNA levels cycle in lLNvs, consistent with past biochemical studies and single-cell RNA-seq studies (Ma et al., 2021), with peak levels at ZT4 (ZT-Zeitgeber time, ZT0-time of lights on, ZT12-time of lights off) with ∼80 molecules per neuron and trough levels at ZT16 with ∼30 molecules per neuron. Our study describes a powerful technique to directly visualize and quantify the number and subcellular location of core clock mRNAs in individual neurons, which can open new investigations into the role of spatiotemporal localization of mRNAs in controlling circadian rhythms. The methods described here can be easily adapted to directly visualize gene expression at the single-cell level in Drosophila brains.
To directly visualize the subcellular localization and quantify the number of core clock mRNAs in individual clock neurons, we adapted single molecule RNA-Fluorescence In Situ Hybridization (smFISH) method for use in Drosophila clock neurons and developed a machine-learning based analysis pipeline to analyze the data. To validate our smFISH protocol and data analysis pipeline, we chose to focus on visualizing Clock mRNAs in lLNVs, one of the main groups of clock neurons. We note that the same methods can be applied to visualize mRNAs in any other clock neurons. Below, we describe the smFISH method and analysis pipeline in detail and present our findings.
Our smFISH procedure consists of five parts: smFISH sample preparation, image acquisition, data analysis, quality control check, and final plot generation (Figure 1C). For sample preparation, Drosophila brains are dissected, fixed, and hybridized with smFISH probes with the whole protocol taking 3 days requiring little hands-on time (∼1 h/day). For imaging, we used a glycerol-based mountant for tissue clearing and acquired images using a high-resolution Airyscan confocal microscope, which enabled us to acquire well defined diffraction-limited spots corresponding to individual mRNA molecules. To automate the process of identification of the cytoplasm and nucleus of lLNvs, a group of clock neurons, we developed a custom convolutional neural network (CNN) segmentation model to generate three-dimensional lLNv clock neuron masks. Performance of our CNN segmentation model was validated by manual inspection of CNN-generated masks on a testing dataset. To quantify the intensity and visualize the subcellular location of individual mRNA spots, we used a published spot detection algorithm, AIRLOCALIZE (Lionnet et al., 2011). Finally, we developed an integrated pipeline in which all data analysis workflows are automated by the workflow manager Snakemake (Molder et al., 2021) (Figure 1D). After the data analysis step, we generated quality control plots to verify the consistency of our results, identify noisy image outliers, and optimize analysis parameters. The analysis can be rerun with optimized parameters in a fully automated and reproducible manner. In the next subsections, we describe individual steps of the protocol in detail.
To enable detection of Clock mRNA transcripts, we ordered commercial fluorescent oligonucleotide probes from Biosearch Technologies, Inc. First, we designed ∼48 20 nt short oligonucleotide probes that span across the full-length of all annotated Clock mRNA isoform species covering all exons using the online Stellaris RNA-FISH Probe Designer (Figure 1E). For genes with alternative splicing isoforms, shared exons can be used to design smFISH probes to ensure detection of all isoforms. The designer provides masking of repetitive sequences to improve probe specificity, and we generally start with the most stringent masking level. In our experience, thirty probes are generally adequate, but the minimum number of probes that can yield high signal-to-noise ratio images is likely dependent on microscope setup (e.g., objective numerical aperture, detector sensitivity etc.). If the target mRNA is short or contains repetitive sequences, it might not be possible to design ∼48 smFISH probes targeting the mRNA. In such a case, masking level can be adjusted to increase the number of smFISH probes. Each of the 20 nt probe is conjugated at the 5’ end to a fluorophore and collaborative binding of ∼48 probes to an individual mRNA molecule enables detection of a single mRNA spot by fluorescence microscopy. Upon receipt of the probe pool, we prepare aliquots of 20 mM probe stock in 10 mM Tris-Cl, pH 8.5 and store them in −20°C.
In our smFISH experiments, we typically use transgenic flies in which cell membranes are fluorescently labeled. Specifically, we drive the expression of CD4-tdTomato or CD8-GFP (Han et al., 2011) exclusively in clock neurons by crossing them with pan-clock neuron driver flies, Clk856-GAL4 (Gummadova et al., 2009). On day 1, Drosophila brains are dissected, fixed in 4% paraformaldehyde, and permeabilized in 70% ethanol at 4°C overnight (Figure 1F). Fixation is a crucial step in smFISH experiments, as both under-fixation as well as over-fixation of tissues can result in samples in which cellular morphology is not well preserved. Permeabilization with 70% ethanol is another necessary step to allow the smFISH probes to pass through the cell membrane and enter the cell. On day 2, brains are washed with PBS and then incubated in hybridization solution with probes overnight at 37°C. On day 3, brains are washed with PBS to remove the hybridization solution and mounted on a slide using a glycerol-based mountant for tissue clearing (see Methods section for a detailed protocol). This smFISH protocol is robust, simple to implement, taking 3 days from brain dissections to slide preparation, and produces highly reproducible results across experimental repeats (Figure 1F).
The goal is to obtain clock neuron images where smFISH spots and the neuron cell membrane marker are clearly visible and can be distinguished from the background. In general, image acquisition parameters should be optimized taking into account both the clock neuron marker and smFISH probe channels, and the same settings need to be reused across all experimental conditions and repeats to ensure data consistency. Filter settings must be compatible with the fluorophore used and appropriate laser power and detector gain should be determined. While larger laser power and detector gain will yield higher pixel intensities, high laser power will lead to significant bleaching during acquisition and high detector gain will lead to increased random noise. Finally, to further suppress random noise, pixel averaging can be performed to obtain distinct diffraction-limited spots in the smFISH probe channels and clear cell boundaries in the neuron marker channel across all Z-slices.
The goal of the segmentation model is to assign a classification label to each pixel of the Z-stack image, i.e., to determine whether a pixel can be assigned to a ‘clock neuron’ or to the background. The clock neuron marker used in our studies, CD8-GFP, not only labels the plasma membrane but also the components of the secretory pathway intracellularly. Therefore, CD8-GFP cell marker intensity varies significantly throughout the cytoplasm, making conventional segmentation methods such as simple thresholding [e.g., Otsu method (Otsu, 1979)] ineffective. To solve this issue, we adopted a publicly available pretrained encoder-decoder convolutional neural network (CNN) model based on the U-Net architecture (Ronneberger et al., 2015) for semantic segmentation of clock neurons. In short, the CNN model computes complex features based on the original clock neuron marker signal (i.e., encoding) and then assigns classification labels based on the computed features (i.e., decoding). Publicly available pretrained CNN models are typically developed with two-dimensional image datasets (Ronneberger et al., 2015). To provide an accurate segmentation model of 3-dimensional clock neurons, for each Z-slice we input the following images to the CNN model: 1) current Z-slice, 2) a maximum intensity projection (MIP) of neighboring Z-slices within 0.4 μm in one direction, 3) a MIP of neighboring 0.4 μm slices in another Z-direction. (Figure 2A).
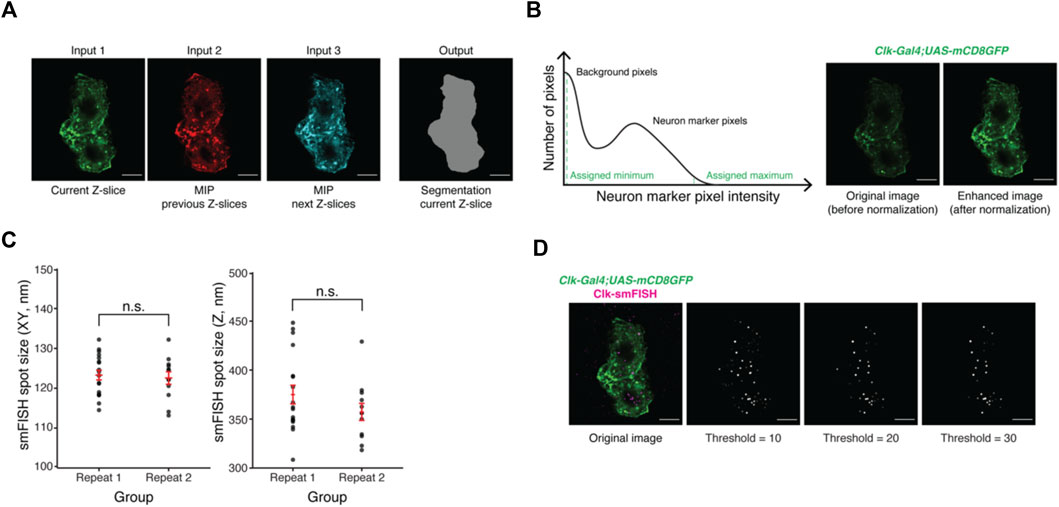
FIGURE 2. Development of the machine-learning based data analysis pipeline. (A) Representative images of current and projections of neighboring Z-slices of lLNvs. Maximum intensity projections (MIP) of neighboring Z-slices are used to incorporate three-dimensional information of lLNvs into the segmentation model. (B) Panel on the left is an illustration of a histogram stretching algorithm. Panel on the right shows representative images of lLNvs before and after normalization. The goal of normalization is to obtain comparable image contrast regardless of original experimental conditions. (C) Experimentally measured smFISH spot size in XY and Z directions. The values are found to be highly consistent across different biological replicates. (D) Representative images of detected Clock smFISH spots in lLNvs with different threshold settings. Proper threshold selection is important for correct identification of the smFISH spots. Scale bars, 5 μm. Statistical test used is unpaired, two-tailed Student’s t-tests assuming unequal variances. Individual data points, mean, and s.e.m. (standard error of mean) are shown.
Further, during model training, we found that CD8-GFP intensity level may vary significantly across different experimental conditions, which hindered the performance of the model. To address this issue, we adopted a MATLAB-based histogram stretching algorithm (MATLAB and Image Processing Toolbox, 2021) to normalize the levels of CD8-GFP intensities among different images. Briefly, the distribution of cell marker pixel intensities follows a bimodal distribution; most pixels will show near-zero values (background pixels) while some show significantly higher values (true CD8GFP signal) (Figure 2B, left panel). We then take the 2% dimmest and brightest intensities and assign them to be the minimum and maximum values. Figure 2B panel on the right shows the original image, in which CD8-GFP signal is dim, and the enhanced image after normalization.
One of the caveats to adopting complex CNN models to generate segmentation masks is the limited training set size which may lead to suboptimal models. To efficiently train our CNN image segmentation model, we augmented (i.e., increased the size of) our training dataset by computationally generating images with varying brightness levels from the original training data set (Dong et al., 2017). We note that lLNv clock neurons are typically contiguous to each other, which makes it difficult to identify individual lLNvs using our segmentation model. In fact, our segmentation model provides an outline of all the lLNvs in each hemi-brain (Supplementary Video S1). We verified the performance of the segmentation model by visual inspection of testing datasets, and we were able to generate segmentation masks of the cytoplasm and the nucleus of lLNvs with ∼10 manually annotated Z-stacks (Supplementary Video S1).
We adopted the publicly available spot counting package, AIRLOCALIZE (Lionnet et al., 2011), for mRNA spot detection and quantification. In short, diffraction-limited spots are modeled as a three-dimensional Gaussian function with lateral (i.e., XY) symmetry. Parameters of the Gaussian function, essentially sizes of the spot in both XY and Z directions, can be measured using a graphical interface provided by the AIRLOCALIZE package. Using our Airyscan confocal microscope, we found that the spot sizes in both XY (∼120 nm) and Z (∼400 nm) directions are highly reproducible (Figure 2C).
To detect smFISH spots in an image, the AIRLOCALIZE algorithm first applies a global threshold to reject noise in the image. Then, the algorithm finds candidates of smFISH spots by identifying locally brightest pixels. Finally, Gaussian fitting is performed using the identified candidate spot locations and a list of “true” smFISH spots is obtained. In Figure 2D, we show the effect of applying different threshold settings on smFISH spot detection. In this example image, the number of detected spots in lLNvs changes with threshold setting. At the minimum threshold tested, we were able to detect ∼70 smFISH spots. At the maximum threshold tested, ∼30 spots were detected (Figure 2D). The global threshold therefore affects smFISH spot detection results and should be set empirically. Typically, we find that the same threshold can be used for all images acquired in one experimental batch. Moreover, our analysis pipeline (discussed in the next section) provides spot visualization capabilities, which can be used to optimize the threshold parameter (Supplementary Video S1).
To ensure reproducibility and aid optimization of analysis parameters, we developed an automated analysis pipeline using the Snakemake workflow management system (Molder et al., 2021) (Figure 1F). First, the raw data is converted to a lossless, open-source OME-TIFF format by the Bio-Formats package (Linkert et al., 2010). Next, smFISH and cell marker images are used for spot detection and cell segmentation, respectively. To obtain the list of smFISH spots within clock neurons, we only consider smFISH spots whose centroids overlap with the lLNv segmentation masks. Finally, quality control plots and images of all intermediate outputs (segmentation masks and detected smFISH spots within clock neurons) are generated together with the final output plots. In the final output plots, we report the number of detected smFISH spots per clock neuron and the locations and intensities of smFISH spots (Figure 1F). Quality control plots are discussed in more detail in the following section. The data analysis pipeline with detailed documentation is publicly available online at Github repository (https://github.com/yeyuan98/rna_fish_analysis).
Using our smFISH method and data analysis pipeline, we performed smFISH experiments to detect Clock mRNA transcripts in large ventral lateral neurons (lLNvs) in whole-mount Drosophila brains across the circadian cycle. We entrained the flies to 12 h Light: 12 h Dark cycles (‘ZT0’ refers to time of lights on and “ZT12” refers to time of lights off) and performed smFISH experiments at various timepoints across the circadian cycle (Figure 3A). We observed ∼80 Clock mRNAs per lLNv at ZT4 (peak) and ∼30 mRNAs per lLNv at ZT16 (trough) (Figures 3B,C; Supplementary Figure S1), consistent with past qPCR and single-cell RNA-seq studies (Ma et al., 2021).
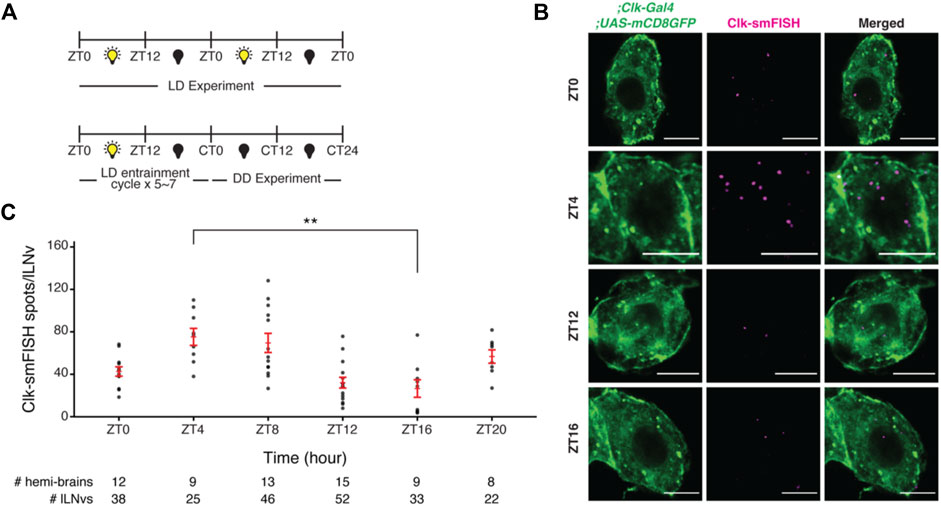
FIGURE 3. Visualization and quantification of Clock mRNA molecules in lLNvs over the circadian cycle. (A) Entrainment schedule for light-dark (LD) or constant darkness (DD) experiments. We performed Clock smFISH experiments using Clk-GAL4>UAS-CD8GFP flies under 12 h Light: 12 h Dark conditions. (B) Representative images of Clock smFISH spots in lLNVs at various timepoints over the circadian cycle. “ZT” refers to Zeitgeber Time; ZT0 denotes the time of lights on and ZT12 denotes the time of lights off. Clock mRNA spots are shown in magenta. (C) Quantification of number of Clock smFISH spots per individual lLNvs over the circadian cycle in wild-type flies. Scale bars, 5 μm. Statistical test used is unpaired, two-tailed Student’s t-test assuming unequal variance. *p < 0.05, **p < 0.005, n.s.-not significant. Individual data points, mean, and s.e.m. (standard error of mean) are shown.
Quality control is an essential component of our fully automated data analysis pipeline, enabling us to verify the validity of our experimental results. As a large number of Z-stack images are generated in each experiment, unbiased consistency verification and identification of noisy images are vital for our workflow. Towards this end, we developed a series of quality control plots. First, we generated intensity histogram plots of all detected smFISH spots in individual images. As individual mRNAs are expected to be bound by a similar number of fluorescent oligonucleotide probes, the histogram plots of smFISH spot intensities are expected to show predominantly a single peak with consistent distribution among different samples. The individual and merged intensity histogram plots generated from our experimental data show that Clock smFISH signal intensities in lLNvs from wild-type flies are highly consistent (Figure 4A). Next, as we usually scan ∼10 μm in the Z-axis for imaging lLNvs in a hemi-brain, we examined how smFISH spot intensity varies across Z position in the acquired images (Figures 4B,D). Again, as individual mRNAs are bound by a similar number of fluorescent oligonucleotide probes, if working distance is sufficient smFISH spot intensity should not depend on Z-position. Indeed, >80% of our acquired images show no significant correlation between smFISH spot intensity values and Z position of the spots. Finally, equivalent diameters of segmentation masks of lLNvs demonstrate consistent volumes expected for lLNv neurons (Figures 4C,D). With these quality controls, we were able to confirm the consistency and validity of our experimental data. Intermediate results of the pipeline including segmentation masks and identified smFISH spots are readily available for visual inspection (Supplementary Videos S1,S2).
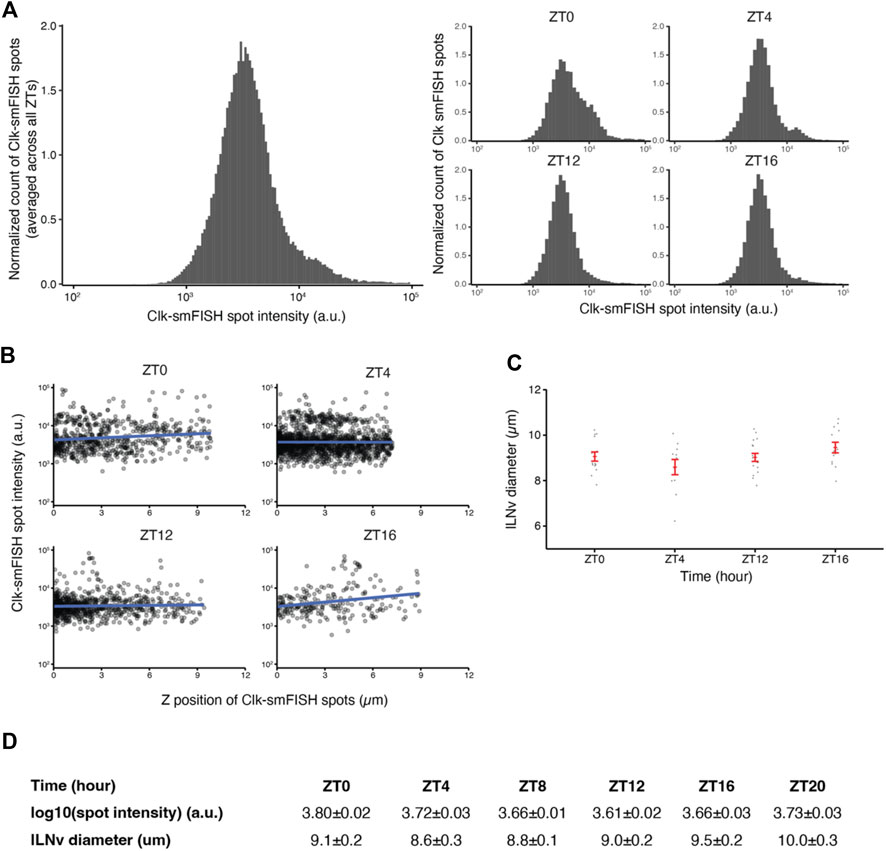
FIGURE 4. Quality control plots for Clock smFISH spots in lLNvs. (A) Histogram plots of Clock smFISH spot intensities in lLNvs. Panel on the left shows averaged data across all ZT’s over the circadian cycle, and panels on the right show data from individual ZT timepoints. (B) Clock smFISH spot intensity distribution across all Z-slices of representative lLNvs at different ZT’s over the circadian cycle. (C) Equivalent diameters of lLNvs segmentation masks at different ZT’s. (D) Summary table of the quality control metrics corresponding to Clock smFISH spot intensities and lLNv diameters at different ZT’s over the circadian cycle. Individual data points, mean, and s.e.m. (standard error of mean) are shown.
To further validate our findings, we examined how the number of Clock mRNAs are affected over the circadian cycle in various mutants. We found that Clock mRNA transcript numbers do not cycle in per null mutant flies (Konopka and Benzer, 1971) and remain low throughout the circadian cycle (Figures 5A,C; Supplementary Figure S2). These results are consistent with past studies which reported that CLK protein fluorescence levels are lower in per mutant flies compared to control flies (Andreazza et al., 2015). Further, we tested how Clock mRNA levels are affected upon clock neuron-specific RNAi-knockdown of PDP1 and VRILLE, transcriptional regulators of Clock. Our smFISH results demonstrate that knockdown of PDP1 abolished Clock mRNA rhythms with lower number of Clock mRNA molecules at different timepoints over the circadian cycle, consistent with PDP1’s function as a Clock activator (Figures 5B,D; Supplementary Figure S3). As expected, knockdown of VRILLE, which is a transcriptional repressor of Clock, led to a higher number of Clock mRNA molecules over the circadian cycle (Figures 5B,E; Supplementary Figure S3).
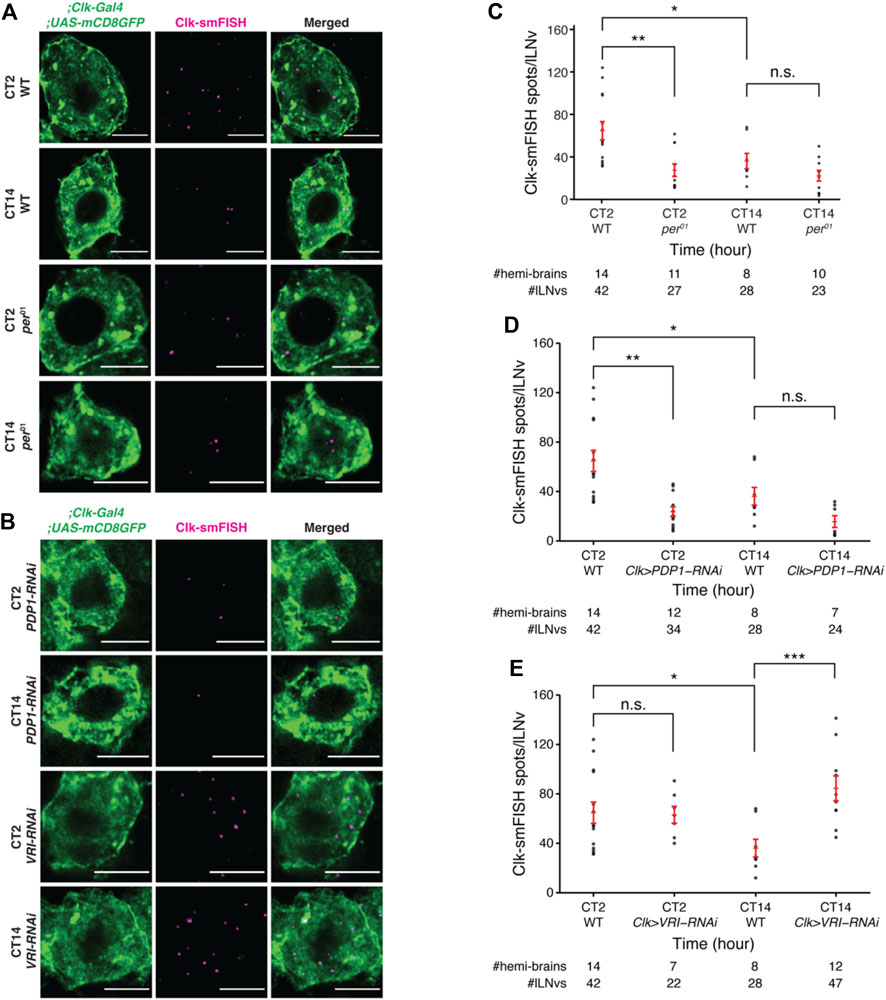
FIGURE 5. Clock mRNA rhythms are disrupted in per01 null mutant and clock neuron specific-knockdown of PDP1 and VRI. (A) Representative images of Clock smFISH spots in lLNvs from control and per01 mutant flies in constant darkness conditions. “CT” refers to Circadian Time. Clock smFISH spot count is lower at both the timepoints over the circadian cycle in per01 mutants. (B) Representative images of Clock smFISH spots in lLNvs from Clk-Gal4; UAS-PDP1-RNAi and Clk-Gal4; UAS-VRI-RNAi flies at different CT’s over the circadian cycle. (C) Quantification of number of Clock smFISH spots per individual lLNvs over the circadian cycle in per01 mutant flies. (D,E) Quantification of the number of Clock smFISH spots per lLNv in PDP1-RNAi (D), and VRI-RNAi (E) flies at different CT’s. Scale bars, 5 μm. Statistical test used is unpaired, two-tailed Student’s t-test assuming unequal variance. *p < 0.05, **p < 0.005, ***p < 0.0005, n.s.-not significant. Individual data points, mean, and s.e.m. (standard error of mean) are shown.
Here, we present a streamlined single molecule RNA-fluorescent in situ hybridization (smFISH) protocol and a machine-learning based data analysis pipeline that enables direct visualization of the subcellular location of individual mRNAs and precise quantification of the total number of mRNA molecules in clock neurons. Using this method, we show that Clock mRNAs cycle in lLNvs with peak levels (∼80 molecules/lLNv) at ZT4 and trough levels (∼30 molecules/lLNv) at ZT16. We validated our smFISH experimental protocol and data analysis pipeline by analyzing Clock mRNA levels in wildtype and mutant conditions.
In addition to transcriptional regulation, post-transcriptional regulation plays a prominent role in regulating gene expression. Localization of mRNAs to specific subcellular structures (e.g., P-bodies, stress granules, nuclear speckles) has been shown to be vital for post-transcriptional regulation in many different biological processes (Decker and Parker, 2012; Fei et al., 2017; Hubstenberger et al., 2017). To visualize mRNA localization, RNA fluorescence in situ hybridization (RNA-FISH) methods have been widely adapted in the past in many model systems (Lawrence and Singer, 1986; Femino et al., 1998; Fowlkes et al., 2008). In general, RNA-FISH technique uses a number of oligonucleotide probes that can specifically bind to specific sequences of a mRNA, and direct detection (as shown in our Clock smFISH experiments) or amplification-based methods (e.g., hybridization chain reaction, HCR (Schwarzkopf et al., 2021)) can be used to visualize the mRNAs. While amplification-based methods offer more flexibility with regard to target length and yield better signal-to-noise ratios, direct detection methods generate individual diffraction-limited spots for each single mRNA molecule with well-defined intensity and shape profiles.
smFISH protocol requires hybridization of tens of ∼20 nt oligonucleotides to the same RNA molecule of interest. Therefore, a relatively long transcript (∼1.5 kb) is required for adequate signal-to-noise ratio (SNR). If short transcripts or specific parts of transcripts (e.g., individual exons or introns) need to be visualized, amplification-based methods such as hybridization chain reaction (HCR) may be adopted (Schwarzkopf et al., 2021). However, while spatial information is typically retained in amplification-based methods, precise quantification of the number of mRNA molecules might require optimization as signal amplification kinetics must be determined empirically. While smFISH and HCR protocols involve fixing the sample and hybridizing it with fluorescent oligonucleotide probes, MS2-MCP system enables live imaging of individual mRNA molecules with high temporal and spatial resolution (Lionnet et al., 2011; Tutucci et al., 2018). Briefly, RNA loops derived from the bacteriophage MS2 can be inserted in the 3′UTR of an mRNA of interest and co-expressed with MCP, which is a MS2 coat protein that can tightly bind to MS2 binding sites, fused to fluorescent proteins. While the MS2-MCP system enables live imaging of individual mRNA molecules, however, addition of stem-loops to the mRNA molecule of interest may affect its function and/or transcription/degradation kinetics. In conclusion, our smFISH method and the data analysis pipeline presented here is a powerful tool that can be employed to study subcellular organization of any clock mRNA molecule in Drosophila clock neurons.
No statistical calculations were used to pre-determine sample sizes. Samples were not randomized for the smFISH experiments and blinding was not used during the data analysis step. We reported the sample sizes for each of our experiments in the figures.
Flies were raised on cornmeal-yeast-sucrose food under a 12:12 Light:Dark cycle at 25°C and 60–70% humidity. The following flies used in the study were previously described or obtained from the Bloomington Stock Center: Clk-GAL4 (Gummadova et al., 2009) (expressed in all clock neuron classes), per01 (Konopka and Benzer, 1971), UAS-CD8GFP (Han et al., 2011), UAS-Pdp1-RNAi (BL-40863), UAS-vri-RNAi (BL-40862). Specific genotypes of flies used in the experiments are presented in the figures.
Prepare all solutions with RNase-free tubes and reagents and filtered pipette tips. Aliquot reagent stocks to reduce chance of contamination.
Fixation solution: 4% formaldehyde in 1X phosphate-buffered saline (PBS) and is made from 16% EM-grade formaldehyde (Electron Microscopy Sciences 15710) and 10X PBS (Invitrogen AM9625) in RNase-free water.
Permeabilization solution: 70% ethanol diluted from 200 proof ethanol (Thermo Scientific T038181000) in RNase-free water.
Hybridization solution: 2X saline-sodium citrate (SSC, Invitrogen AM9763), 10% dextran sulfate (Thermo Scientific AC433240050), 1 mg/mL E.coli tRNA (Roche 10109541001), 2 mM ribonucleoside vanadyl complex (RVC, New England Biolabs S1402S), 0.5% bovine serum albumin (BSA, Roche 10711454001), 10% formamide (Sigma, F9037), and 5 μM oligonucleotide probes conjugated to fluorophores (Stellaris, LGC Biosearch Technologies) in RNase-free water.
Wash solution: 2X SSC and 10% formamide in RNase-free water.
We prepare 20% (weight/volume) dextran sulfate stock from powder and store at 4°C. Fixation and permeabilization solutions are prepared fresh on day of experiment. RVC and formamide aliquots are used one-time once thawed.
1. Prepare fixative solution. For each sample, put 500 uL of fixative to a well of a clean glass dish and leave on ice.
2. Dissect at least 10 brains in 1X PBS for each sample. Put dissected brains directly into the fixative on ice in the glass dish. Dissections should be completed within 30 min.
3. Place the glass dish on a nutator and incubate for 20 min at room temperature.
4. Remove the fixative and perform three quick rinses with 1X PBS.
5. Remove solution and wash twice with 1X PBS, 5 min per wash at room temperature with nutation.
6. Remove 1X PBS completely and add 500 uL of permeabilization solution to the glass dish. With a P1000 tip, carefully transfer the brains into a protein low-bind tube. Avoid damaging the brains during transfer.
7. Fill the tubes with extra permeabilization solution to a final volume of 1 ml. Incubate the sample overnight at 4°C with nutation, protected from light.
8. Prepare hybridization solution, 100 μL per sample and leave on ice protected from light. Prepare wash solution, 1 ml per sample and leave at room temperature. Hybridization solution is viscous and should be well-mixed by pipetting (>10 times).
9. Aspirate permeabilization solution and add 1 ml of wash solution. Incubate 5 min at room temperature with nutation.
10. Aspirate the wash solution completely and add 100 μL of hybridization solution containing probe(s). Gently pipette to make sure all brains are collected in the solution.
11. Incubate overnight in a 37°C incubator, protected from light.
12. Add 1 ml of wash solution without removing the hybridization solution. Gently pipette once to mix without touching the brains. Incubate at 37°C for 30 min.
13. Aspirate off the solution and wash one more time with 1 ml of wash solution at 37°C for 30 min. Supplement the wash solution with 1 μg/ml DAPI for nucleus staining.
14. Remove the wash solution and mount the samples in Prolong glass mountant. Make sure the brains are mounted in the appropriate orientation to visualize the clock neurons. Seal the samples immediately after mounting and leave at room temperature overnight protected from light.
15. The samples are ready for imaging the next day.
Flies were entrained to Light-Dark (LD) cycles with lights on for 12 h and off for 12 h for 5–7 days, and then released into complete darkness (DD) for 6–7 more days. Zeitgeber Time (ZT) 0 marks the time of lights on, and ZT12 marks the time of lights off. Circadian time (CT) refers to times in complete darkness (DD) and CT0 is the start of the subjective light phase and CT12 is the start of the subjective dark phase, the times when the light transitions would have occurred had the LD cycle continued.
All flies used for smFISH experiments were placed in density-controlled food vials (4 females and 4 males) and entrained for 5–7 days in incubators. We performed all our smFISH experiments on 5–7 day old male or female flies, and did not notice any differences in our experimental results. The fly genotype and ZT was as described in the figure legends. We used the GAL4/UAS system to express transgenes in clock neurons in the brain. We imaged our samples using a Zeiss LSM800 laser scanning confocal microscope with AiryScan super-resolution module (125 nm lateral and 350 nm axial resolution). We have acquired our images using a 63 × Plan-Apochromat Oil (N.A. 1.4) objective and 405, 488, and 561, and 647 nm laser lines.
All experiments were repeated at least twice independently. All statistical significance tests presented are unpaired, two-tailed Student’s t-tests assuming unequal variances.
The raw data supporting the conclusion of this article will be made available by the authors, without undue reservation.
YY, Conceptualization, Data curation, Formal analysis, Validation, Investigation, Visualization, Methodology, Writing—original draft, Writing—review and editing; M-AP and DC, Formal analysis, Visualization; SY, Conceptualization, Investigation, Supervision, Funding acquisition, Methodology, Writing—original draft, Writing—review and editing.
This work was supported by funds from the NIH (Grant No. R35GM133737 to SY), Alfred P. Sloan Fellowship (to SY) and McKnight Scholar Award (to SY).
The authors acknowledge help from Yukiko Yamashita and Jaclyn Fingerhut in adapting smFISH protocol for use in adult Drosophila brains. The authors also thank Yangbo Xiao for aiding in the preparation of the figures.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fphys.2022.1051544/full#supplementary-material
Supplementary Figure S1 | Zoomed-out representative image of Clock smFISH spots in all lLNvs. Here we show a representative Z-slice of all four lLNvs observed in a hemi-brain at ZT4. Scale bars, 5 μm.
Supplementary Figure S2 | Quality control plots for Clock smFISH spots in lLNvs from per01 null mutant flies. (A) Histogram plots of Clock smFISH spot intensities in lLNvs from per01 (left) and wildtype (right) flies at different CT’s over the circadian cycle. (B) Equivalent diameters of lLNvs segmentation masks from per01 (left) and wildtype (right) flies at different CT’s. (C) Clock smFISH spot intensity distribution across all Z-slices of representative lLNvs from per01 (left) and wildtype (right) flies at different CT’s. Statistical test used is unpaired, two-tailed Student’s t-test assuming unequal variance. *p < 0.05, n.s.-not significant. Individual data points, mean, and s.e.m. (standard error of mean) are shown.
Supplementary Figure S3 | Quality control plots for Clock smFISH spots in lLNvs from Clk>Pdp1-RNAi and Clk>vri-RNAi mutants. (A,B) Histogram plots of Clock smFISH spot intensities in lLNvs at different CT’s over the circadian cycle in PDP-RNAi (A) and VRI-RNAi (B) conditions. (C,D) Equivalent diameters of lLNvs segmentation masks at different CT’s in PDP-RNAi (C) and VRI-RNAi (D) flies.
Supplementary Video S1 | Representative segmentation mask of lLNv neurons. A Z-stack video showing a group of 4 lLNv clock neurons marked with CD8-GFP (left panel) and representative segmentation masks from our CNN model (right panel). Scale bar: 4 μm.
Supplementary Video S2 | Visualization of detected Clock smFISH spots in lLNvs. A Z-stack video showing Clock mRNA probe signals in magenta (Quasar 670 fluorophore) and clock neurons in green (CD8-GFP) in the left panel. Right panel shows visualization of detected Clock smFISH spots in lLNvs using AIRLOCALIZE algorithm. Scale bar: 4 μm.
Andreazza S., Bouleau S., Martin B., Lamouroux A., Ponien P., Papin C., et al. (2015). Daytime CLOCK dephosphorylation is controlled by STRIPAK complexes in Drosophila. Cell Rep. 11, 1266–1279. doi:10.1016/j.celrep.2015.04.033
Aronson B. D., Johnson K. A., Loros J. J., Dunlap J. C. (1994). Negative feedback defining a circadian clock - autoregulation of the clock gene-frequency. Science 263, 1578–1584. doi:10.1126/science.8128244
Bartholomai B. M., Gladfelter A. S., Loros J. J., Dunlap J. C. (2022). PRD-2 mediates clock-regulated perinuclear localization of clock gene RNAs within the circadian cycle of Neurospora. Proc. Natl. Acad. Sci. U. S. A. 119, e2203078119. doi:10.1073/pnas.2203078119
Buhr E. D., Yoo S. H., Takahashi J. S. (2010). Temperature as a universal resetting cue for mammalian circadian oscillators. Science 330, 379–385. doi:10.1126/science.1195262
Cohen S. E., Erb M. L., Selimkhanov J., Dong G., Hasty J., Pogliano J., et al. (2014). Dynamic localization of the cyanobacterial circadian clock proteins. Curr. Biol. 24, 1836–1844. doi:10.1016/j.cub.2014.07.036
Curtin K. D., Huang Z. J., Rosbash M. (1995). Temporally regulated nuclear entry of the Drosophila period protein contributes to the circadian clock. Neuron 14, 365–372. doi:10.1016/0896-6273(95)90292-9
Cyran S. A., Buchsbaum A. M., Reddy K. L., Lin M. C., Glossop N. R. J., Hardin P. E., et al. vrille, Pdp1, and dClock form a second feedback loop in the Drosophila circadian clock. Cell 112, 329–341. doi:10.1016/S0092-8674(03)00074-62003).
Decker C. J., Parker R. (2012). P-Bodies and stress granules: Possible roles in the control of translation and mRNA degradation. Cold Spring Harb. Perspect. Biol. 4, a012286. doi:10.1101/cshperspect.a012286
Dong H., Yang G., Liu F. D., Mo Y. H., Guo Y. K. (2017). Automatic brain tumor detection and segmentation using U-net based fully convolutional networks. Comm. Com. Inf. S. C. 723, 506–517. doi:10.1007/978-3-319-60964-5_44
Dubowy C., Sehgal A. (2017). Circadian rhythms and sleep in Drosophila melanogaster. Genetics 205, 1373–1397. doi:10.1534/genetics.115.185157
Fei J., Jadaliha M., Harmon T. S., Li I. T. S., Hua B., Hao Q., et al. (2017). Quantitative analysis of multilayer organization of proteins and RNA in nuclear speckles at super resolution. J. Cell Sci. 130, 4180–4192. doi:10.1242/jcs.206854
Femino A. M., Fay F. S., Fogarty K., Singer R. H. (1998). Visualization of single RNA transcripts in situ. Science 280, 585–590. doi:10.1126/science.280.5363.585
Fowlkes C. C., Hendriks C. L. L., Keranen S. V. E., Weber G. H., Rubel O., Huang M. Y., et al. (2008). A quantitative spatiotemporal atlas of gene expression in the Drosophila blastoderm. Cell 133, 364–374. doi:10.1016/j.cell.2008.01.053
Glossop N. R. J., Houl J. H., Zheng H., Ng F. S., Dudek S. M., Hardin P. E. (2003). VRILLE feeds back to control circadian transcription of Clock in the Drosophila circadian oscillator. Neuron 37, 249–261. doi:10.1016/S0896-6273(03)00002-3
Gummadova J. O., Coutts G. A., Glossop N. R. (2009). Analysis of the Drosophila Clock promoter reveals heterogeneity in expression between subgroups of central oscillator cells and identifies a novel enhancer region. J. Biol. Rhythms 24, 353–367. doi:10.1177/0748730409343890
Han C., Jan L. Y., Jan Y. N. (2011). Enhancer-driven membrane markers for analysis of nonautonomous mechanisms reveal neuron-glia interactions in Drosophila. Proc. Natl. Acad. Sci. U. S. A. 108, 9673–9678. doi:10.1073/pnas.1106386108
Hardin P. E., Hall J. C., Rosbash M. (1990). Feedback of the Drosophila period gene product on circadian cycling of its messenger RNA levels. Nature 343, 536–540. doi:10.1038/343536a0
Hubstenberger A., Courel M., Benard M., Souquere S., Ernoult-Lange M., Chouaib R., et al. (2017). P-body purification reveals the condensation of repressed mRNA regulons. Mol. Cell 68, 144–157. doi:10.1016/j.molcel.2017.09.003
Konopka R. J., Benzer S. (1971). Clock mutants of Drosophila melanogaster. Proc. Natl. Acad. Sci. U. S. A. 68, 2112–2116. doi:10.1073/pnas.68.9.2112
Lawrence J. B., Singer R. H. (1986). Intracellular localization of messenger RNAs for cytoskeletal proteins. Cell 45, 407–415. doi:10.1016/0092-8674(86)90326-0
Linkert M., Rueden C. T., Allan C., Burel J. M., Moore W., Patterson A., et al. (2010). Metadata matters: Access to image data in the real world. J. Cell Biol. 189, 777–782. doi:10.1083/jcb.201004104
Lionnet T., Czaplinski K., Darzacq X., Shav-Tal Y., Wells A. L., Chao J. A., et al. (2011). A transgenic mouse for in vivo detection of endogenous labeled mRNA. Nat. Methods 8, 165–170. doi:10.1038/nmeth.1551
Ma D., Przybylski D., Abruzzi K. C., Schlichting M., Li Q., Long X., et al. (2021). A transcriptomic taxonomy of Drosophila circadian neurons around the clock. Elife 10, e63056. doi:10.7554/eLife.63056
MATLAB and image processing Toolbox release, Natick, Massachusetts, United States: The MathWorks, Inc.(2021).
Molder F., Jablonski K. P., Letcher B., Hall M. B., Tomkins-Tinch C. H., Sochat V., et al. (2021). Sustainable data analysis with Snakemake. F1000Res. 10, 33. doi:10.12688/f1000research.29032.2
Nitabach M. N., Taghert P. H. (2008). Organization of the Drosophila circadian control circuit. Curr. Biol. 18, R84–R93. doi:10.1016/j.cub.2007.11.061
Otsu N. A threshold selection method from gray-level histograms. IEEE Trans. Syst. Man. Cybern. 9, 62–66. doi:10.1109/Tsmc.1979.43100761979).
Patke A., Young M. W., Axelrod S. (2020). Molecular mechanisms and physiological importance of circadian rhythms. Nat. Rev. Mol. Cell Biol. 21, 67–84. doi:10.1038/s41580-019-0179-2
Raj A., van den Bogaard P., Rifkin S. A., van Oudenaarden A., Tyagi S. (2008). Imaging individual mRNA molecules using multiple singly labeled probes. Nat. Methods 5, 877–879. doi:10.1038/nmeth.1253
Ronneberger O., Fischer P., Brox T. (2015). U-Net: Convolutional networks for biomedical image segmentation. Lect. Notes Comput. S. C. 9351, 234–241. doi:10.1007/978-3-319-24574-4_28
Schwarzkopf M., Liu M. C., Schulte S. J., Ives R., Husain N., Choi H. M. T., et al. (2021). Hybridization chain reaction enables a unified approach to multiplexed, quantitative, high-resolution immunohistochemistry and in situ hybridization. Development 148, dev199847. doi:10.1242/dev.199847
Sehadova H., Glaser F. T., Gentile C., Simoni A., Giesecke A., Albert J. T., et al. (2009). Temperature entrainment of Drosophila's circadian clock involves the gene nocte and signaling from peripheral sensory tissues to the brain. Neuron 64, 251–266. doi:10.1016/j.neuron.2009.08.026
Sehgal A., Price J. L., Man B., Young M. W. (1994). Loss of circadian behavioral rhythms and per RNA oscillations in the Drosophila mutant timeless. Science 263, 1603–1606. doi:10.1126/science.8128246
Shafer O. T., Rosbash M., Truman J. W. (2002). Sequential nuclear accumulation of the clock proteins period and timeless in the pacemaker neurons of Drosophila melanogaster. J. Neurosci. 22, 5946–5954. doi:10.1523/JNEUROSCI.22-14-05946.2002
Stanewsky R., KanekoM. , Emery P., Beretta B., Wager-Smith K., Kay S. A., et al. (1998). The cryb mutation identifies cryptochrome as a circadian photoreceptor in Drosophila. Cell 95, 681–692. doi:10.1016/s0092-8674(00)81638-4
Trcek T., Lionnet T., Shroff H., Lehmann R. (2017). mRNA quantification using single-molecule FISH in Drosophila embryos. Nat. Protoc. 12, 1326–1348. doi:10.1038/nprot.2017.030
Tutucci E., Vera M., Biswas J., Garcia J., Parker R., Singer R. H. (2018). An improved MS2 system for accurate reporting of the mRNA life cycle. Nat. Methods 15, 81–89. doi:10.1038/nmeth.4502
Xiao Y., Yuan Y., Jimenez M., Soni N., Yadlapalli S. (2021). Clock proteins regulate spatiotemporal organization of clock genes to control circadian rhythms. Proc. Natl. Acad. Sci. U. S. A. 118, e2019756118. doi:10.1073/pnas.2019756118
Yadlapalli S., Jiang C., Bahle A., Reddy P., Meyhofer E., Shafer O. T. (2018). Circadian clock neurons constantly monitor environmental temperature to set sleep timing. Nature 555, 98–102. doi:10.1038/nature25740
Yuan Y., Xiao Y., Yadlapalli S. (2022). The role of spatiotemporal organization and dynamics of clock complexes in circadian regulation. Curr. Opin. Cell Biol. 78, 102129. doi:10.1016/j.ceb.2022.102129
Zhang R., Lahens N. F., Ballance H. I., Hughes M. E., Hogenesch J. B. (2014). A circadian gene expression atlas in mammals: Implications for biology and medicine. Proc. Natl. Acad. Sci. U. S. A. 111, 16219–16224. doi:10.1073/pnas.1408886111
Keywords: circadian rhythms, single molecule RNA-fluorescence in situ hybridization (smFISH), machine learning-based analysis pipeline, spatiotemporal organization of clock mRNAs, Drosophila clock neurons
Citation: Yuan Y, Padilla M-A, Clark D and Yadlapalli S (2022) Streamlined single-molecule RNA-FISH of core clock mRNAs in clock neurons in whole mount Drosophila brains. Front. Physiol. 13:1051544. doi: 10.3389/fphys.2022.1051544
Received: 22 September 2022; Accepted: 17 October 2022;
Published: 09 November 2022.
Edited by:
Joanna C. Chiu, University of California, Davis, United StatesReviewed by:
Luoying Zhang, Huazhong University of Science and Technology, ChinaCopyright © 2022 Yuan, Padilla, Clark and Yadlapalli. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Ye Yuan, eWV5dUB1bWljaC5lZHU=; Swathi Yadlapalli, c3dhdGhpQHVtaWNoLmVkdQ==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.