 Tingting Li
Tingting Li Chao Yu
Chao Yu Shougang Zhuang
Shougang Zhuang- 1Department of Nephrology, Shanghai East Hospital, Tongji University School of Medicine, Shanghai, China
- 2Department of Medicine, Alpert Medical School and Rhode Island Hospital, Brown University, Providence, RI, United States
Enhancer of zeste homolog 2 (EZH2) is a histone-lysine N-methyltransferase enzyme that catalyzes the addition of methyl groups to histone H3 at lysine 27, leading to gene silencing. Mutation or over-expression of EZH2 has been linked to many cancers including renal carcinoma. Recent studies have shown that EZH2 expression and activity are also increased in several animal models of kidney injury, such as acute kidney injury (AKI), renal fibrosis, diabetic nephropathy, lupus nephritis (LN), and renal transplantation rejection. The pharmacological and/or genetic inhibition of EZH2 can alleviate AKI, renal fibrosis, and LN, but potentiate podocyte injury in animal models, suggesting that the functional role of EZH2 varies with renal cell type and disease model. In this article, we summarize the role of EZH2 in the pathology of renal injury and relevant mechanisms and highlight EZH2 as a potential therapeutic target for kidney diseases.
Introduction
Epigenetics refers to the heritable change of gene function and phenotype without alteration in a genes DNA sequence (Berger et al., 2009). The core function of epigenetics is to control access to the DNA genetic code both spatially and temporally to ensure orderly gene expression and silencing according to external signals. Epigenetic regulation involves DNA methylation, histone modification, chromatin recombination, and non-coding RNA (Rosner and Hengstschlager, 2012; Roy and Majumdar, 2012). These modifications control the fate of cells, regulate the normal growth, and development of individuals, and are closely related to the occurrence and development of various diseases such as cancers, diabetes, heart disease, and intestinal disease (Filion et al., 2010). The importance and diversity of histone post-translational modification have been widely studied in many epigenetic regulatory mechanisms. Histone modifications include acetylation, methylation, ubiquitination, hydroxylation, phosphorylation, and ADP ribosylation (Graff and Tsai, 2013; Hyun et al., 2017; Bochyńska et al., 2018; Worden et al., 2019). Methylation can occur in histone and non-histone proteins. Histones are proteins that are abundant in lysine and arginine and are found in eukaryotic cell nuclei. Among four core histone proteins (H2A, H2B, H3, and H4), histone H3 is one of the most important epigenetic markers in transcriptional regulation. The methylation of histone H3 mainly occurs on the arginine (R) and lysine (K) residues in its tail (Wei et al., 2009). Histone arginine residues are monomethylated or dimethylated (Blanc and Richard, 2017), while histone lysine can be monomethylated (me1), dimethylated (me2), or trimethylated (me3). Different methylation sites such as H3K4, H3K9, H3K27, H3K36, and H3K79 have been identified (Wang et al., 2009). Among those sites, trimethylation of lysine 27 in histone H3 (H3K27me3) is a key marker of gene silencing, found mainly in gene promoter and enhancer regions (Duan et al., 2020).
H3K27 trimethylation is mainly carried out by Polycomb Repressor Complex (PRC2; Tie et al., 1998; Sanchez-Beato et al., 2006; Yu et al., 2007; Veneti et al., 2017). PRC2 is a histone methyltransferase that mainly trimethylates the histone H3 lysine 27 (H3K27me3; Margueron and Reinberg, 2011; Di Croce and Helin, 2013). The PRC2 complex consists of four components: enhancer of zeste homolog 1 (EZH1) or enhancer of zeste homolog 2 (EZH2), compressor of zeste12 (Suz12), embryonic ectoderm development (EED), and RbAp46/48. EZH1 and EZH2 are the core components of PRC2, while EED can interact with EZH1 or EZH2 to maintain enzyme activity (Kim et al., 2013; Eich et al., 2020). EZH2 is an essential component of PRC2 methylation activity; it can also methylate some non-histone substrates, such as actin, GATA binding protein 4 (GATA4), android receiver (AR), and estrogen receiver (ER). EZH2 can silence tumor suppressor genes and play an important role in cell aging, fate selection, and differentiation (Jacobs and van Lohuizen, 2002; Plath et al., 2003; Martinez and Cavalli, 2006; Aloia et al., 2013). In addition, EZH2-mediated methylation of non-histones such as STAT3 is an important post-translational modification involved in many life processes, such as the cell cycle, DNA repair, cell aging, differentiation, apoptosis, and tumorigenesis (Kim et al., 2013; Eich et al., 2020).
Enhancer of zeste homolog 2 was discovered in 1996 using yeast two-hybrid experiments (Hobert et al., 1996) and its gene is located on the human chromosome 7q35. EZH2 occupies nearly 40 kb in the gene structure, which contains 20 exons, and the open reading frame is distributed on 19 exons (Cardoso et al., 2000). Studies have shown that EZH2 can inhibit the expression of tumor suppressor genes in normal cells, thereby promoting the abnormal proliferation of cells, and stimulating the metastasis of tumor cells. EZH2 exhibits gene amplification and higher levels of expression in many human malignancies, such as gastric cancer, colon cancer, breast cancer, lymphoid hematopoietic tumors, liver cancer, and nephroblastoma (Su et al., 2003; Mimori et al., 2005; Raman et al., 2005; Sudo et al., 2005; Matsukawa et al., 2006; Saramaki et al., 2006; Shi et al., 2007), and is closely related to tumorigenesis and tumor progression.
Increasing evidence shows that EZH2 is associated with a variety of kidney diseases and pathology. In addition to the abnormal expression and activation of EZH2 in renal tumors, its expression levels and activity were also increased in acute kidney injury (AKI; Zhou et al., 2018b), renal fibrosis (Zhou et al., 2016), diabetic nephropathy (DN; Jia et al., 2019b), lupus nephritis (LN; Rohraff et al., 2019), hyperuricemic nephropathy (Shi et al., 2019), and transplanted and aging kidneys (Li et al., 2016; Han and Sun, 2020). Moreover, pharmacological or genetic inhibition of EZH2 can interfere with pathologic fibrosis in these animal models of kidney disease. This article reviews the role of EZH2 in the pathology of renal disease and relevant mechanisms. We also highlight EZH2 as a potential target for ameliorating fibrosis in kidney disease (Table 1).
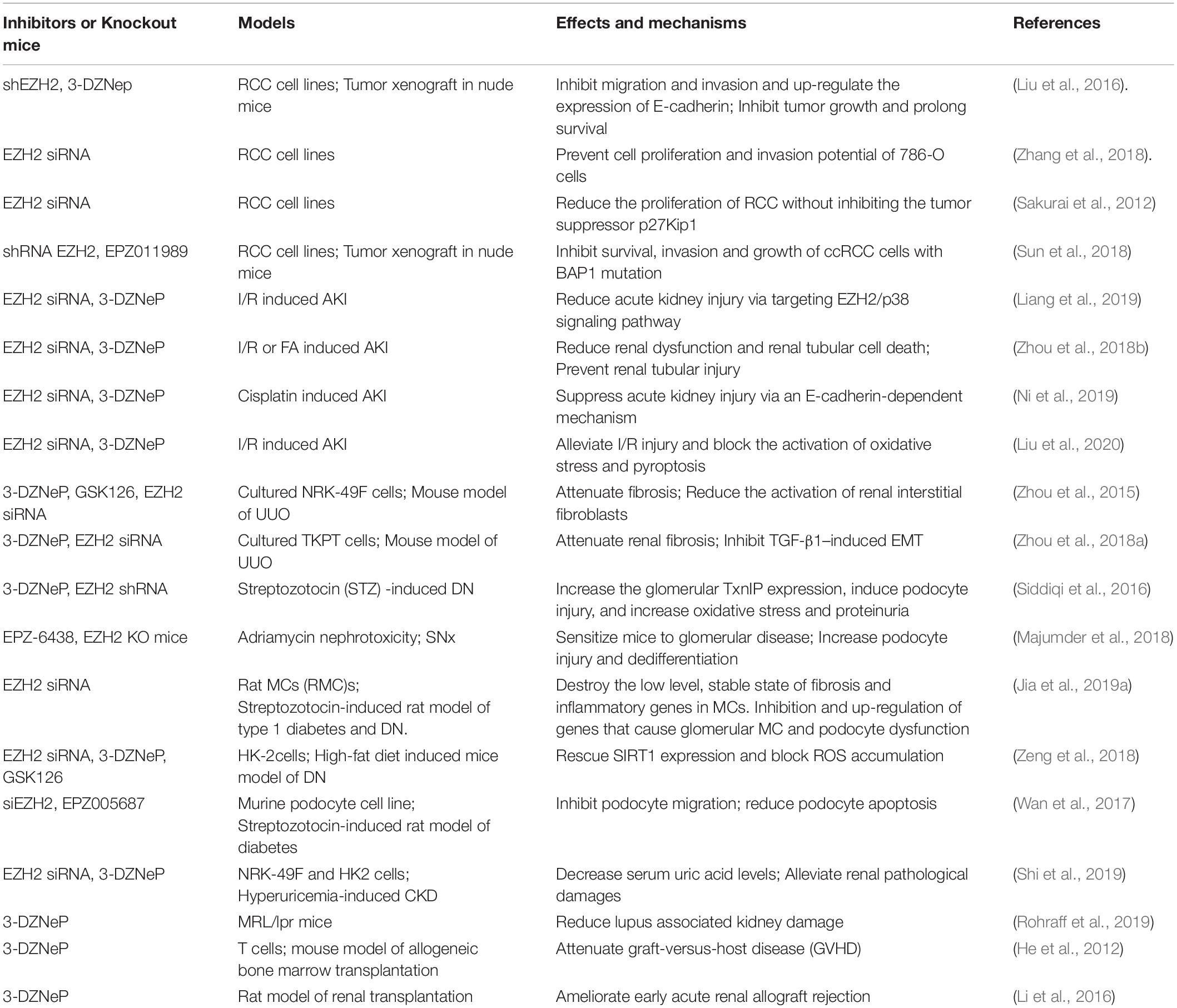
Table 1. EZH2 inhibition on kidney diseases in various in vitro and in vivo models.
EZH2 and Renal Cell Carcinoma
Renal cell carcinoma (RCC), also known as renal adenocarcinoma. originates from renal tubular epithelial cells and accounts for over 90% of adult renal malignancies. The worldwide incidence RCC has increased significantly in recent years (Shah et al., 2009). At present, the cause and pathogenesis of RCC are not clear. Epidemiology speculates that the pathogenesis is related to genetics, smoking, obesity, hypertension, and antihypertensive treatment (Grossman et al., 1999). About 70% of renal cancers are associated with gene expression inactivation caused by VHL gene deletion and mutation. VHL gene encodes an E3 ubiquitin ligase complex protein, which can degrade hypoxia inducible factor (HIF). HIF is continuously activated in RCC cells, promoting the transcription of a series of downstream target genes (Mallikarjuna et al., 2018). Recently, it has been reported that epigenetic modification, in particular, EZH2 activation, participates in the occurrence and development of RCC, which provides a new direction for the treatment (Bannister and Kouzarides, 2011).
Studies have demonstrated that EZH2 can promote the development and metastasis of RCC. EZH2 is overexpressed in numerous tumor entities including renal tumor cells (Kim and Roberts, 2016; Sun et al., 2018). EZH2 overexpression leads to increases in H3K27me3, with repression of tumor-suppressor genes such as E-cadherin (Liu et al., 2016). In vitro and in vivo studies confirmed that the abnormal increase of EZH2 can inhibit the expression level of E-cadherin, induce the epithelial stromal transformation of renal cancer cells, and promote the occurrence, development and recurrence of renal cancer. Inhibition of EZH2 with 3-DZNep can reverse these pathological responses (Liu et al., 2016). EZH2 also has growth promoting activity in RCC (Wagener et al., 2010) and can enhance the proliferation and invasion of renal tubular epithelial cells (Zhang et al., 2018). Moreover, EZH2 promotes cell proliferation, migration and angiogenesis by inhibiting expression of tumor suppressor genes such as p27Kip1 and enhancing expression of proto-oncogenes (Sakurai et al., 2012). Thus, inhibition of EZH2 can reduce the survival and invasion of clear cell renal cell carcinoma (ccRCC) cells and the growth of ccRCC in xenografted mice (Sun et al., 2018). Finally, EZH2 in combination with the DNA methyltransferase DNMT can methylate the VHL promoter and inhibit its expression in RCC (Schlesinger et al., 2007).
EZH2 and Acute Kidney Injury
Acute kidney injury is a common pathologic process with high mortality in hospitalized patients (Raimann et al., 2018). It can be caused by ischemia/reperfusion, septicemia, or nephrotoxins (i.e., radiocontrast agents, NSAIDs, etc.) (Zuk and Bonventre, 2016; Moledina et al., 2017; Wu et al., 2017). Acute injury to the kidney usually leads to death of renal tubular epithelial cells, activation of endothelial cells, infiltration of leukocytes, and ultimately renal dysfunction (Sato and Yanagita, 2018; Ronco et al., 2019). In mild injury, adaptive repair mechanisms can restore epithelial integrity, inhibit immune responses and reconstruct a healthy vascular system. On the contrary, severe or persistent damage can lead to inadequate repair (Sato and Yanagita, 2018; Ronco et al., 2019). Tubular cells may experience G2/M cell cycle arrest, senescence, apoptosis or necrosis, leading to release of pro-inflammatory factors (Guzzi et al., 2019). Emerging evidence has shown the role of EZH2-mediated histone modifications in AKI (Ho et al., 2017) (Bomsztyk and Denisenko, 2013).
The abnormal expression or activation of EZH2 is related to the pathogenesis of AKI Zhou et al., 2018b). Initially, it was found that EZH2 is involved in many cellular responses, such as apoptosis and inflammation (Wang Y. et al., 2018; Bamidele et al., 2019). The expression of EZH2 and H3K27me3 is up-regulated in ischemia-reperfusion and folic acid-induced AKI models (Zhou et al., 2018b). Inhibition of EZH2 with 3-DZNep can reduce renal dysfunction and tubular cell death (Zhou et al., 2018b). E-cadherin downregulation mediates disruption of cell-cell adhesion, activation of matrix metalloproteinases, and activation of ERK1/2, and it promotes activation of cell death receptor and regulation of mitochondrial damage. Inhibition of EZH2 can preserve expression of E-cadherin and tight junction protein ZO-1, inhibit expression of matrix metalloproteinase MMP-2 and MMP-9, and suppress phosphorylation of Raf-1 and ERK1/2 in renal tubular cells exposed to oxidative stress (Zhou et al., 2018b). This was confirmed by an in vitro study (Zhou et al., 2018b). In a murine model of cisplatin induced-AKI, inhibition of EZH2 expression by 3-DZNep could also reduce apoptosis of renal tubular cells and ameliorate acute renal injury by restoring expression of E-cadherin (Ni et al., 2019). Both in vitro and in vivo, cisplatin-induced renal tubular cell damage was accompanied by up-regulation of H3K27me3, while 3-DZNep treatment did not affect its expression (Ni et al., 2019). This suggests that 3-DZNeP-elicited renal protection in response to cisplatin exposure may not be through a H3K27me3-mediated mechanism. Recently, it was demonstrated that EZH2 can regulate renal injury by inducing oxidative stress as evidenced by the fact that EZH2 inhibition blocked the production of NOX4 dependent ROS through the ALK5/Smad2/3 signal pathway in an animal model of ischemia/reperfusion-induced AKI (Liu et al., 2020). In the same injury model, EZH2 inhibition also reduced renal dysfunction and tubular injury by regulating p38 signaling, apoptosis and inflammation (Liang et al., 2019). Therefore, EZH2 is an important mediator in the pathogenesis of AKI. Additional studies are needed to describe the mechanism of EZH2 in AKI in more detail.
EZH2 and Renal Fibrosis
Renal fibrosis is a common pathological process in the progression of CKD to end-stage renal disease (ESRD). Renal interstitial fibrosis is considered an example of poor self-healing after injury (Iwano and Neilson, 1991; Inoue et al., 2015; Lovisa et al., 2015). It is mainly manifested by extracellular matrix (ECM) deposition, epithelial to mesenchymal transition (EMT), inflammatory response, and fibroblast activation and proliferation (Iwano and Neilson, 1991; Inoue et al., 2015; Lovisa et al., 2015). Although many studies have been conducted to elucidate its pathogenesis, there is still a lack of effective treatment for renal fibrosis (Boor et al., 2010; Nogueira et al., 2017).
Recent studies have demonstrated that EZH2 plays a critical role in the development of renal fibrosis. EZH2 expression levels are low in normal kidney tissue, but high in mice kidneys following injury and in human kidneys following disease (Zhou et al., 2015). Immunostaining showed that EZH2 was expressed both in tubular epithelial cells and renal interstitial fibroblasts (Zhou et al., 2015). Activation and proliferation of renal fibroblasts produces a large amount of ECM proteins, including fibronectin and collagen I. In vitro, pharmacological inhibition or siRNA mediated silencing of EZH2 reduced the activation of renal interstitial fibroblasts; in vivo, treatment with the EZH2 inhibitor 3-DZNep attenuated UUO-induced fibrosis in an animal model. The anti-fibrotic effect of EZH2 inhibition is related to the inhibition of expression of epidermal growth factor receptor (EGFR) and platelet-derived growth factor receptor (PDGFR) and deactivation of multiple intracellular signaling pathways, including TGFβ/Smad3, AKT and ERK1/2 (Zhou et al., 2015). EZH2 was also identified as a key regulator of epithelial-mesenchymal transition (EMT) in fibrotic kidneys (Zhou et al., 2018a). EZH2 inhibition elicits an anti-EMT effect related to preservation of E-cadherin expression, repression of transcription factors (i.e., Snail, twist), and deactivation of PTEN/Akt and β-catenin signaling pathways (Zhou et al., 2018a). Studies have also shown that EZH2 inhibition can effectively suppress the development of liver fibrosis (Zeybel et al., 2017), skin fibrosis (Tsou et al., 2019), atrial fibrosis (Song et al., 2019), pulmonary fibrosis (Xiao et al., 2016), and peritoneal fibrosis (Shi et al., 2020). These results suggest that EZH2 may serve as a promising therapeutic target for the treatment of fibrosis in many organ systems.
EZH2 and Diabetic Nephropathy
Diabetic nephropathy is one of the most common chronic complications of diabetes (Alicic et al., 2017; Han et al., 2017; Feldman et al., 2019). The typical characteristics of DN are glomerular hypertrophy, proteinuria, progressive decrease of glomerular filtration rate, and renal fibrosis, which leads to loss of renal function. The pathological changes of diabetes are mainly composed of mesangial hyperplasia, thickening of basement membrane, occlusion of capillary lumen, disorder of podocyte structure, and decrease of podocyte number (Tung et al., 2018). However, the specific molecular mechanism of DN is not completely understood. Recent studies have shown that EZH2 activation is involved in the pathogenesis of DN. EZH2 can regulate oxidative stress in diabetic patients by inhibiting expression of Pax6, a transcription factor, and thus the expression of TxnIP, an endogenous antioxidant inhibitor (Siddiqi et al., 2016). The specific deletion of EZH2 in podocytes induced podocyte damage and dedifferentiation (Siddiqi et al., 2016) and sensitized mice to glomerular disease (Majumder et al., 2018). In human glomerular diseases such as focal segmental glomerulosclerosis and DN, H3K27me3 was decreased in podocytes. H3K27me3 and EZH2 are involved in inhibiting and maintaining the low-level and stable state of fibrosis and inflammation genes in mesangial cells, while H3K27me3 and EZH2 are inhibited by TGF-β, which increases the expression of genes that mediate glomerular mesangial dysfunction and DN, leading to renal dysfunction (Jia et al., 2019a). In contrast, the depletion or inhibition of EZH2 attenuated the increase of reactive oxygen species (ROS) in human renal tubular epithelial cells (HK-2) induced by high glucose (Zeng et al., 2018). Moreover, Wilm’s tumor 1 (WT1) can improve the β-catenin mediated damage of podocytes in DN by antagonizing EZH2, which is manifested in reducing the transformation of the stroma of DN podocytes, maintaining the structural integrity of DN podocytes, reducing apoptosis and oxidative stress of DN podocytes (Wan et al., 2017). These data illustrate that EZH2 activity is necessary for protection against podocyte damage in DN. This is opposite to what has been observed in the murine model of UUO-induced renal fibrosis (Zhou et al., 2015). Currently, it remains unclear about the underlying mechanism by which EZH2 plays distinct roles in different disease models and cells. Since multiple cell types are involved in the pathogenesis of DN, and EZH2 mediated gene regulation is cell-context specific, EZH2 may play the role of a double-edged sword in different cell types in DN (Brasacchio et al., 2009). Further investigation is required to elucidate the gene and signaling pathways regulated by EZH2 during DN development.
EZH2 and Hyperuricemic Nephropathy
Hyperuricemia (HUA) is a purine metabolic disorder, in which the blood uric acid level is higher than the normal level due to the increase of uric acid production and/or the decrease of uric acid excretion. HUA is an independent risk factor for CKD progression (Mok et al., 2012) and has a direct correlation with renal damage (Lin et al., 2014). 10–20% of patients with primary hyperuricemia have evidence of AKI and chronic kidney disease, including chronic uric acid nephropathy, acute uric acid nephropathy and uric acid stones. Long-term HUA can cause renal damage through the urate crystal dependent pathway. Uric acid crystals are deposited in the distal collecting duct and the renal interstitium, causing chronic interstitial nephritis, which may lead to renal interstitial fibrosis (Liu et al., 2015; Yuan et al., 2017; Johnson et al., 2018). In addition, long-term HUA can cause renal damage through an independent pathway of urate crystal formation. In this regard, it has been reported that uric acid can cause renal endothelial dysfunction, renin-angiotensin system (RAS) activation, inflammation, oxidative stress (Sanchez-Lozada et al., 2008; Yu et al., 2010; Filiopoulos et al., 2012), whereas lowering serum uric acid alleviates renal damage or delays its progression (Obermayr et al., 2008; Zhou et al., 2012; Kim et al., 2014).
Although the molecular mechanism of renal damage caused by elevated uric acid levels remains obscure, uric acid has been shown to induce activation of TGF-β receptor and transcription of TGF-β1 target genes (Böttinger, 2007), leading to activation of downstream signaling pathways such as EGFR and ERK1/2 (Joo et al., 2008; Lee et al., 2010). Interestingly, blocking EZH2 by 3-DZNep inhibits TGF-β1-induced activation of renal interstitial fibroblasts in vitro and attenuates ECM protein deposition and α-smooth muscle actin expression in obstructed kidneys (Zhou et al., 2015). Inhibition of EZH2 by 3-DZNep also significantly reduces blood uric acid levels by reducing the activity of serum purine oxidase (XOD) and alleviates HUA-induced renal damage through various mechanisms, including inhibition of the TGF-β1/Smad3 and EGFR/ERK1/2 signaling pathways. Moreover, 3-DZNep was effective in downregulating the levels of various pro-inflammatory chemokines/cytokines and reducing the apoptosis of renal tubular cells (Shi et al., 2019). Therefore, EZH2 may be a potential therapeutic target for reducing renal damage and delaying the development of CKD caused by HUA.
EZH2 and Lupus Nephritis
Systemic lupus erythematosus (SLE) is a chronic autoimmune inflammatory disease characterized by loss of immune tolerance to autoantigens, production of autoantibodies, formation of immune complexes and deposition in different parts of the body, causing inflammation and multiorgan damage (Marshall and Vierstra, 2018). LN is one of the most common and serious complications of SLE, affecting up to 60% of lupus patients by some estimates. LN is thus considered an important cause of chronic kidney disease (Koutsokeras and Healy, 2014; Zhu et al., 2016). The pathogenesis of SLE and LN is complex and exact mechanism(s) are largely unknown. It is generally believed that SLE is caused by genetic, endocrine, environmental factors (infection, ultraviolet radiation, etc.), and abnormal activation of the immune system. Epigenetic changes have also been reported to contribute to the pathogenesis of lupus (Ballestar et al., 2006). In particular, various abnormal patterns of DNA methylation of immune cell types isolated from lupus patients have been found to be related to clinical heterogeneity, interspecific disease variability, lupus onset and remission (Teruel and Sawalha, 2017).
Similar to DNA methylation, histone modifications can lead to abnormal gene expression and contribute to the pathogenesis of SLE (Hu et al., 2008). It has been reported that unregulated EZH2 activation in CD4+ T cells leads to T cell activation and non-Th1 immune responses, prior to transcription activity, and is related to lupus activity. In addition, levels of two microRNAs (miR-101 and miR-26a) targeting and regulating EZH2 in CD4+ T cells of lupus patients were negatively correlated with lupus disease activity (Coit et al., 2016). EZH2 can mark and control functions and survival of effector T cells through microRNAs and the Notch signaling pathway (Zhao et al., 2016). In lupus patients, overexpression of EZH2 leads to the methylation of JAM-A (junctional adhesion molecule A), which may increase the migration of T cells and lead to the invasive exosmosis of T cells. Blocking EZH2 by 3-DZNep and GSK126 can effectively inhibit the adhesion of lupus T cells to human microvascular endothelial cells. In addition, overexpression of EZH2 results in methylation changes of genes involved in gene transcription, ubiquitination and immune response, indicating that EZH2 is involved in various cellular and physiological processes crucial to the survival and function of T cells (Tsou et al., 2018). Inhibition of EZH2 by 3-DZNep significantly reduced the number of pathogenic double negative T cells and production of cytokines and chemokines in lupus prone MRL/lpr mice. Splenomegaly and lymphadenopathy in mice treated with 3-DZNep were significantly reduced. Most importantly, inhibition of EZH2 by 3-DZNep in MRL/lpr mice can reduce renal damage and increase survival rate of MRL/lpr mice. Mice with 3-DZNep treatment have relatively stable albumin:creatinine ratios, and attenuated glomerulonephritis and crescent formation. Glomerular necrosis in the prevention group of mice was significantly relieved as well. Therefore, 3-DZNep elicited inhibition of EZH2 can effectively prevent the progression of renal damage in lupus (Rohraff et al., 2019). Additional studies are necessary to examine whether other EZH2 inhibitors or gene modulators are also effective in ameliorating the pathogenesis of LN.
EZH2 and Aging Kidney
Aging is an irreversible phenomenon characterized by gradual decline of cell function and gradual structural changes of many organ systems. Age-related changes in kidney function include anatomical and physiological changes (Zhou et al., 2008). The histological changes in renal aging mainly include: renal mass reduction, glomerulosclerosis, renal tubular atrophy, renal interstitial fibrosis and arterial intimal fibrosis (Silva, 2005). The partial loss of renal function can be manifested as decreases in renal vascular elasticity, renal blood flow and glomerular filtration rate. At present, mechanisms of renal aging are incompletely studied. Increasing evidence indicates that renal aging is related to epigenetic changes (Painter et al., 2008; Au et al., 2013; Kooman et al., 2014).
Epigenetic histone modification plays a role in aging (Sen et al., 2016), especially trimethylation of 27 lysine (H3K27me3) of histone H3, which is directly related to life span and aging in different models (Jin et al., 2011; Ma et al., 2018). Previous studies have shown that EZH2 expression is related to abnormal expression of genes in aging animal models (Shumaker et al., 2006; Bracken et al., 2007; Chen et al., 2009). For example, overexpression of EZH2 can prevent stem cell failure and aging (De Haan and Gerrits, 2007) while pharmacological inhibition of EZH2 can dysregulate tissue regeneration in aged mice (Nishiguchi et al., 2018). Methylation of CpG islands related to aging may overlap with the regulatory regions of cancer genes such as c1ql3. EZH2 interacts with these regulatory regions in mice, and the occupancy of EZH2 may decrease with age at c1ql3. EZH2 is part of the protein mechanism of forming the aging epigenome (Mozhui and Pandey, 2017). A recent study found that H3K27me3 regulated the expression of Klotho in the kidney of aging mice. A decrease of Klotho levels is an important mechanism of aging. The epigenetic down-regulation of Klotho gene expression is at least partly due to the histone 3 modification of the Klotho promoter. Aging plays a role by up-regulating H3K27me3 and down-regulating hyperphosphorylation of Klotho and mTOR in renal tubules. Inhibition of EZH2 with GSK343 or EED226 was able to reduce H3K27me3 recruitment to the Klotho promoter (Han and Sun, 2020). The expression level of EZH2 decreased in older mice (Han and Sun, 2020). At present, no study has confirmed the obvious correlation between the expression of EZH2 and H3K27me3 in the kidney of aging mice or adult tissues (Margueron et al., 2008). Studies have pointed out that renal aging can up-regulate the ECM laminin genes by down-regulating 5mC and H3K27me3 in the promoter region of the ECM laminin gene. Reduction of H3K27me3 levels by 3-DZNep can inhibit expression of the laminin gene in the ECM, but administration of a more specific EZH2 inhibitor, GSK-126, did not inhibit expression of the laminin gene in ECM (Denisenko et al., 2018). Therefore, the exact mechanism(s) by which EZH2 contributes to renal aging needs further investigation.
EZH2 and Kidney Transplantation Rejection
Currently, renal transplantation is the most effective treatment for ESRD. However, acute rejection (AR) is a common adverse reaction after kidney transplantation, usually occurring weeks to months after transplantation. Rejection after transplantation is caused by the recipient’s immune system. Recognition of the graft as a foreign body stimulates secretion of various inflammatory factors to attack the graft. Eliminating the foreign body reaction and maintaining the stability of the internal environment would improve the long-term survival of the transplanted kidney. Transplantation rejection includes T-cell-mediated rejection and antibody mediated rejection (Haas et al., 2018). Acute T-cell mediated rejection is an inflammatory reaction, involving extensive T lymphocytes infiltration of the allograft and activation (Yang C. et al., 2015). T cell mediated AR of renal transplantation includes mononuclear interstitial infiltration and tubulitis with intima-intimal arteritis. Epigenetic modification is involved in T cell-mediated AR of renal transplantation (Cuddapah et al., 2010).
Enhancer of zeste homolog 2 plays an important role in maintaining T cell numbers and functions. It has been documented that EZH2 is essential for the expansion of T-effector cells (Yang X. P. et al., 2015) as well as differentiation and characteristic maintenance of regulatory T cells necessary for maintaining immune homeostasis (DuPage et al., 2015). EZH2 can also stimulate the expression of T cell multifunctional cytokines by activating the Notch pathway, and promoting T cell survival by Bcl-2 expression (Zhao et al., 2016). In addition, EZH2 can protect key T cell development regulators from DNA methylation so that they can be activated in a subsequent differentiation stage (Wang C. et al., 2018). Treatment with 3-DZNep, a EZH2 inhibitor can induce selective apoptosis of alloantigen-activated T cells and arrest persistent graft-versus-host disease (GVHD) in mice after allogeneic bone marrow transplantation (He et al., 2012). A recent study was the first to demonstrate the relationship between EZH2 and allograft rejection. EZH2 in T cells was increased after kidney transplantation in a rat model of kidney transplantation; inhibition of EZH2 by DZNep reduced AR, reduced injury and inflammatory infiltration of the transplanted kidney. The cellular mechanisms are related to the inhibition of activation and proliferation of alloreactive T cells, impairment of production of inflammatory factors, and increased apoptosis of alloreactive T cells in the transplanted kidney and peripheral blood) (Li et al., 2016). However, the specific mechanism(s) of EZH2 in AR of renal transplantation remains to be further studied.
Conclusion and Perspectives
Enhancer of zeste homolog 2 is highly expressed in renal tumors and many kidney diseases. Abnormal expression or activation of EZH2 can lead to development and progression of renal tumors and several kidney diseases as indicated in this review. The molecular mechanisms of EZH2-mediated renal pathology are associated with renal tubular cell injury, podocyte dedifferentiation, renal interstitial fibroblast proliferation, production of multiple cytokines/chemokines and infiltration of inflammatory cells. Because EZH2 regulates expression of diverse genes and activation of multiple signaling pathways associated with pathogenesis of disease, its functional role may vary with cell types, tissues and disease models. In this context, EZH2 activation has been shown to contribute to renal tubular cell death, but protects against podocyte injury. Therefore, further studies are necessary to elucidate the detailed mechanistic actions of EZH2 in the pathogenesis and progression of kidney diseases.
Given that preclinical studies have demonstrated that EZH2 inhibitors attenuate some renal diseases in animal models, EZH2 inhibitors alone, or in combination with other drugs may provide beneficial effects to ameliorate or prevent kidney diseases. This is encouraged by recent approval of tazemetostat (EPZ-6438), one of EZH2 inhibitors, for treatment of adult patients with relapsed or refractory (R/R) follicular lymphoma by the FDA (Gulati et al., 2018). Among the 203 patients who were evaluated for efficacy, responses were seen in 24% patients who had received tazemetostat administered as a single agent in both tumor types and in EZH2 mutant and WT tumor (Gulati et al., 2018). In addition to tazemetostat, other EZH2 inhibitors such as GSK126 and CPT-1205 are being tested in treating lymphoma and several other solid tumor types (Eich et al., 2020). Currently, there are still no clinical trials of EZH2 inhibitors for the treatment of kidney disease of any underlying cause. Based on the evidence showing the efficacy of EZH2 inhibitors in animal models of kidney disease, clinical trials assessing the effect of EZH2 inhibition may hold out the promise of treatment for some forms of progressive kidney disease in humans. However, since genetic and pharmacological inhibition of EZH2 potentiates podocyte injury in murine models of glomerular disease (Majumder et al., 2018), EZH2 inhibitors may not be suitable for treatment of renal diseases associated with podocyte injury.
Author Contributions
TL and CY drafted the article. SZ edited the manuscript. All the authors reviewed the manuscript and approved is for publication.
Funding
This work was supported by the National Natural Science Foundation of China (81670623 and 81830021 to SZ), the Branch Grant of National Key Grants of the Ministry of Science and Technology (2018YFA0108802 to SZ), and the US National Institutes of Health (1R01DK113256-01A1 to SZ).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We appreciate Dr. George Bayliss for editing this manuscript.
References
Alicic, R. Z., Rooney, M. T., and Tuttle, K. R. (2017). Diabetic kidney disease: challenges, progress, and possibilities. Clin. J. Am. Soc. Nephrol. 12, 2032–2045. doi: 10.2215/cjn.11491116
Aloia, L., Di Stefano, B., and Di Croce, L. (2013). Polycomb complexes in stem cells and embryonic development. Development 140, 2525–2534. doi: 10.1242/dev.091553
Au, C. P., Raynes-Greenow, C. H., Turner, R. M., Carberry, A. E., and Jeffery, H. (2013). Fetal and maternal factors associated with neonatal adiposity as measured by air displacement plethysmography: a large cross-sectional study. Early Hum. Dev. 89, 839–843. doi: 10.1016/j.earlhumdev.2013.07.028
Ballestar, E., Esteller, M., and Richardson, B. C. (2006). The epigenetic face of systemic lupus erythematosus. J. Immunol. 176, 7143–7147. doi: 10.4049/jimmunol.176.12.7143
Bamidele, A. O., Svingen, P. A., Sagstetter, M. R., Sarmento, O. F., Gonzalez, M., Braga Neto, M. B., et al. (2019). Disruption of FOXP3-EZH2 interaction represents a pathobiological mechanism in intestinal inflammation. Cell Mol. Gastroenterol. Hepatol. 7, 55–71. doi: 10.1016/j.jcmgh.2018.08.009
Bannister, A. J., and Kouzarides, T. (2011). Regulation of chromatin by histone modifications. Cell Res. 21, 381–395. doi: 10.1038/cr.2011.22
Berger, S. L., Kouzarides, T., Shiekhattar, R., and Shilatifard, A. (2009). An operational definition of epigenetics. Genes Dev. 23, 781–783. doi: 10.1101/gad.1787609
Blanc, R. S., and Richard, S. (2017). Arginine methylation: the coming of age. Mol. Cell 65, 8–24. doi: 10.1016/j.molcel.2016.11.003
Bochyńska, A., Lüscher-Firzlaff, J., and Lüscher, B. (2018). Modes of interaction of KMT2 histone H3 lysine 4 methyltransferase/COMPASS complexes with chromatin. Cells 7:17. doi: 10.3390/cells7030017
Bomsztyk, K., and Denisenko, O. (2013). Epigenetic alterations in acute kidney injury. Semin. Nephrol. 33, 327–340. doi: 10.1016/j.semnephrol.2013.05.005
Boor, P., Ostendorf, T., and Floege, J. (2010). Renal fibrosis: novel insights into mechanisms and therapeutic targets. Nat. Rev. Nephrol. 6, 643–656. doi: 10.1038/nrneph.2010.120
Böttinger, E. P. (2007). TGF-beta in renal injury and disease. Semin.Nephrol. 27, 309–320. doi: 10.1016/j.semnephrol.2007.02.009
Bracken, A. P., Kleine-Kohlbrecher, D., Dietrich, N., Pasini, D., Gargiulo, G., Beekman, C., et al. (2007). The Polycomb group proteins bind throughout the INK4A-ARF locus and are disassociated in senescent cells. Genes Dev. 21, 525–530. doi: 10.1101/gad.415507
Brasacchio, D., Okabe, J., Tikellis, C., Balcerczyk, A., George, P., Baker, E. K., et al. (2009). Hyperglycemia induces a dynamic cooperativity of histone methylase and demethylase enzymes associated with gene-activating epigenetic marks that coexist on the lysine tail. Diabetes 58, 1229–1236. doi: 10.2337/db08-1666
Cardoso, C., Mignon, C., Hetet, G., Grandchamps, B., Fontes, M., and Colleaux, L. (2000). The human EZH2 gene: genomic organisation and revised mapping in 7q35 within the critical region for malignant myeloid disorders. Eur. J. Hum. Genet. 8, 174–180. doi: 10.1038/sj.ejhg.5200439
Chen, H., Gu, X., Su, I. H., Bottino, R., Contreras, J. L., Tarakhovsky, A., et al. (2009). Polycomb protein Ezh2 regulates pancreatic beta-cell Ink4a/Arf expression and regeneration in diabetes mellitus. Genes Dev. 23, 975–985. doi: 10.1101/gad.1742509
Coit, P., Dozmorov, M. G., Merrill, J. T., McCune, W. J., Maksimowicz-McKinnon, K., Wren, J. D., et al. (2016). Epigenetic reprogramming in naive CD4+ T cells favoring T cell activation and Non-Th1 effector T cell immune response as an early event in lupus flares. Arthr. Rheumatol. 68, 2200–2209. doi: 10.1002/art.39720
Cuddapah, S., Barski, A., and Zhao, K. (2010). Epigenomics of T cell activation, differentiation, and memory. Curr. Opin. Immunol. 22, 341–347. doi: 10.1016/j.coi.2010.02.007
De Haan, G., and Gerrits, A. (2007). Epigenetic control of hematopoietic stem cell aging the case of Ezh2. Ann. N. Y. Acad. Sci. 1106, 233–239. doi: 10.1196/annals.1392.008
Denisenko, O., Mar, D., Trawczynski, M., and Bomsztyk, K. (2018). Chromatin changes trigger laminin genes dysregulation in aging kidneys. Aging (Albany N. Y.) 10, 1133–1145. doi: 10.18632/aging.101453
Di Croce, L., and Helin, K. (2013). Transcriptional regulation by Polycomb group proteins. Nat.Struct. Mol. Bio.l. 20, 1147–1155. doi: 10.1038/nsmb.2669
Duan, R., Du, W., and Guo, W. (2020). EZH2: a novel target for cancer treatment. J. Hematol. Oncol. 13:104. doi: 10.1186/s13045-020-00937-8
DuPage, M., Chopra, G., Quiros, J., Rosenthal, W. L., Morar, M. M., Holohan, D., et al. (2015). The chromatin-modifying enzyme Ezh2 is critical for the maintenance of regulatory T cell identity after activation. Immunity 42, 227–238. doi: 10.1016/j.immuni.2015.01.007
Eich, M. L., Athar, M., Ferguson, J. E. III, and Varambally, S. (2020). EZH2-targeted therapies in cancer: hype or a reality. Cancer Res. 80, 5449–5458. doi: 10.1158/0008-5472.CAN-20-2147
Feldman, E. L., Callaghan, B. C., Pop-Busui, R., Zochodne, D. W., Wright, D. E., Bennett, D. L., et al. (2019). Diabetic neuropathy. Nat. Rev. Dis. Primers 5:41. doi: 10.1038/s41572-019-0092-1
Filion, G. J., van Bemmel, J. G., Braunschweig, U., Talhout, W., Kind, J., Ward, L. D., et al. (2010). Systematic protein location mapping reveals five principal chromatin types in Drosophila cells. Cell 143, 212–224. doi: 10.1016/j.cell.2010.09.009
Filiopoulos, V., Hadjiyannakos, D., and Vlassopoulos, D. (2012). New insights into uric acid effects on the progression and prognosis of chronic kidney disease. Ren. Fail 34, 510–520. doi: 10.3109/0886022X.2011.653753
Graff, J., and Tsai, L. H. (2013). Histone acetylation: molecular mnemonics on the chromatin. Nat. Rev. Neurosci. 14, 97–111. doi: 10.1038/nrn3427
Grossman, E., Messerli, F. H., and Goldbourt, U. (1999). Does diuretic therapy increase the risk of renal cell carcinoma? Am. J. Cardiol. 83, 1090–1093. doi: 10.1016/s0002-9149(99)00021-1
Gulati, N., Beguelin, W., and Giulino-Roth, L. (2018). Enhancer of zeste homolog 2 (EZH2) inhibitors. Leuk. Lymphoma 59, 1574–1585.
Guzzi, F., Cirillo, L., Roperto, R. M., Romagnani, P., and Lazzeri, E. (2019). Molecular Mechanisms of the acute kidney injury to chronic kidney disease transition: an updated view. Int. J. Mol. Sci. 20:941. doi: 10.3390/ijms20194941
Haas, M., Loupy, A., Lefaucheur, C., Roufosse, C., Glotz, D., Seron, D., et al. (2018). The Banff 2017 Kidney Meeting Report: Revised diagnostic criteria for chronic active T cell-mediated rejection, antibody-mediated rejection, and prospects for integrative endpoints for next-generation clinical trials. Am. J. Transplant 18, 293–307. doi: 10.1111/ajt.14625
Han, Q., Zhu, H., Chen, X., and Liu, Z. (2017). Non-genetic mechanisms of diabetic nephropathy. Front. Med. 11:319. doi: 10.1007/s11684-017-0569-9
Han, X., and Sun, Z. (2020). Epigenetic Regulation of KL (Klotho) via H3K27me3 (Histone 3 Lysine [K] 27 Trimethylation) in Renal Tubule Cells. Hypertension 75, 1233–1241. doi: 10.1161/HYPERTENSIONAHA.120.14642
He, S., Wang, J., Kato, K., Xie, F., Varambally, S., Mineishi, S., et al. (2012). Inhibition of histone methylation arrests ongoing graft-versus-host disease in mice by selectively inducing apoptosis of alloreactive effector T cells. Blood 119, 1274–1282. doi: 10.1182/blood-2011-06-364422
Ho, T. H., Kapur, P., Eckel-Passow, J. E., Christie, A., Joseph, R. W., Serie, D. J., et al. (2017). Multicenter Validation of Enhancer of Zeste Homolog 2 Expression as an Independent Prognostic Marker in Localized Clear Cell Renal Cell Carcinoma. J. Clin. Oncol. 35, 3706–3713. doi: 10.1200/JCO.2017.73.3238
Hobert, O., Jallal, B., and Ullrich, A. (1996). Interaction of Vav with ENX-1, a putative transcriptional regulator of homeobox gene expression. Mol. Cell Biol. 16, 3066–3073. doi: 10.1128/mcb.16.6.3066
Hu, N., Qiu, X., Luo, Y., Yuan, J., Li, Y., Lei, W., et al. (2008). Abnormal histone modification patterns in lupus CD4+ T cells. J. Rheumatol. 35, 804–810.
Hyun, K., Jeon, J., Park, K., and Kim, J. (2017). Writing, erasing and reading histone lysine methylations. Exp. Mol. Med. 49:e324. doi: 10.1038/emm.2017.11
Inoue, T., Umezawa, A., Takenaka, T., Suzuki, H., and Okada, H. (2015). The contribution of epithelial-mesenchymal transition to renal fibrosis differs among kidney disease models. Kidney Int. 87, 233–238. doi: 10.1038/ki.2014.235
Iwano, M., and Neilson, E. G. (1991). Mechanisms of tubulointerstitial fibrosis. Kidney Int. 39, 550–556.
Jacobs, J. J., and van Lohuizen, M. (2002). Polycomb repression: from cellular memory to cellular proliferation and cancer. Biochim. Biophys. Acta 1602, 151–161. doi: 10.1016/s0304-419x(02)00052-5
Jia, Y., Reddy, M. A., and Das, S. (2019a). Dysregulation of histone H3 lysine 27 trimethylation in transforming growth factor-β1-induced gene expression in mesangial cells and diabetic kidney. J. Biol. Chem. 294, 12695–12707.
Jia, Y., Reddy, M. A., Das, S., Oh, H. J., Abdollahi, M., Yuan, H., et al. (2019b). Dysregulation of histone H3 lysine 27 trimethylation in transforming growth factor-beta1-induced gene expression in mesangial cells and diabetic kidney. J. Biol. Chem. 294, 12695–12707. doi: 10.1074/jbc.RA119.007575
Jin, C., Li, J., Green, C. D., Yu, X., Tang, X., Han, D., et al. (2011). Histone demethylase UTX-1 regulates C. elegans life span by targeting the insulin/IGF-1 signaling pathway. Cell Metab. 14, 161–172. doi: 10.1016/j.cmet.2011.07.001
Johnson, R. J., Bakris, G. L., Borghi, C., Chonchol, M. B., Feldman, D., Lanaspa, M. A., et al. (2018). Hyperuricemia, acute and chronic kidney disease, hypertension, and cardiovascular disease: report of a scientific workshop organized by the national kidney foundation. Am. J. Kidney Dis. 71, 851–865. doi: 10.1053/j.ajkd.2017.12.009
Joo, C. K., Kim, H. S., Park, J. Y., Seomun, Y., Son, M. J., and Kim, J. T. (2008). Ligand release-independent transactivation of epidermal growth factor receptor by transforming growth factor-beta involves multiple signaling pathways. Oncogene 27, 614–628. doi: 10.1038/sj.onc.1210649
Kim, E., Kim, M., Woo, D. H., Shin, Y., Shin, J., Chang, N., et al. (2013). Phosphorylation of EZH2 activates STAT3 signaling via STAT3 methylation and promotes tumorigenicity of glioblastoma stem-like cells. Cancer Cell 23, 839–852. doi: 10.1016/j.ccr.2013.04.008
Kim, I. Y., Lee, D. W., Lee, S. B., and Kwak, I. S. (2014). The role of uric acid in kidney fibrosis: experimental evidences for the causal relationship. Biomed. Res. Int. 2014:638732. doi: 10.1155/2014/638732
Kim, K. H., and Roberts, C. W. (2016). Targeting EZH2 in cancer. Nat. Med. 22, 128–134. doi: 10.1038/nm.4036
Kooman, J. P., Kotanko, P., Schols, A. M., Shiels, P. G., and Stenvinkel, P. (2014). Chronic kidney disease and premature ageing. Nat. Rev. Nephrol. 10, 732–742. doi: 10.1038/nrneph.2014.185
Koutsokeras, T., and Healy, T. (2014). Systemic lupus erythematosus and lupus nephritis. Nat. Rev. Drug Discov. 13, 173–174. doi: 10.1038/nrd4227
Lee, E., Yi, J. Y., Chung, E., and Son, Y. (2010). Transforming growth factorbeta(1) transactivates EGFR via an H(2)O(2)-dependent mechanism in squamous carcinoma cell line. Cancer Lett. 290, 43–48. doi: 10.1016/j.canlet.2009.08.022
Li, L., Zhang, Y., Xu, M., Rong, R., Wang, J., and Zhu, T. (2016). Inhibition of histone methyltransferase EZH2 ameliorates early acute renal allograft rejection in rats. BMC Immunol. 17:41. doi: 10.1186/s12865-016-0179-3
Liang, H., Huang, Q., Liao, M. J., Xu, F., Zhang, T., He, J., et al. (2019). EZH2 plays a crucial role in ischemia/reperfusion-induced acute kidney injury by regulating p38 signaling. Inflamm. Res. 68, 325–336. doi: 10.1007/s00011-019-01221-3
Lin, B., Shao, L., Luo, Q., Ou-yang, L., Zhou, F., Du, B., et al. (2014). Prevalence of chronic kidney disease and its association with metabolic diseases: a cross-sectional survey in Zhejiang province, Eastern China. BMC Nephrol. 15:36. doi: 10.1186/1471-2369-15-36
Liu, H., Chen, Z., Weng, X., Chen, H., Du, Y., Diao, C., et al. (2020). Enhancer of zeste homolog 2 modulates oxidative stress-mediated pyroptosis in vitro and in a mouse kidney ischemia-reperfusion injury model. Faseb J. 34, 835–852. doi: 10.1096/fj.201901816R
Liu, L., Xu, Z., Zhong, L., Wang, H., Jiang, S., Long, Q., et al. (2016). Enhancer of zeste homolog 2 (EZH2) promotes tumour cell migration and invasion via epigenetic repression of E-cadherin in renal cell carcinoma. BJU Int. 117, 351–362. doi: 10.1111/bju.12702
Liu, N., Wang, L., Yang, T., Xiong, C., Xu, L., Shi, Y., et al. (2015). EGF receptor inhibition alleviates hyperuricemic nephropathy. J. Am. Soc. Nephrol. 26, 2716–2729. doi: 10.1681/ASN.2014080793
Lovisa, S., LeBleu, V. S., Tampe, B., Sugimoto, H., Vadnagara, K., Carstens, J. L., et al. (2015). Epithelial-to-mesenchymal transition induces cell cycle arrest and parenchymal damage in renal fibrosis. Nat. Med. 21, 998–1009. doi: 10.1038/nm.3902
Ma, Z., Wang, H., Cai, Y., Wang, H., Niu, K., Wu, X., et al. (2018). Epigenetic drift of H3K27me3 in aging links glycolysis to healthy longevity in Drosophila. Elife 7:35368. doi: 10.7554/eLife.35368
Majumder, S., Thieme, K., Batchu, S. N., Alghamdi, T. A., Bowskill, B. B., Kabir, M. G., et al. (2018). Shifts in podocyte histone H3K27me3 regulate mouse and human glomerular disease. J. Clin. Invest. 128, 483–499. doi: 10.1172/jci95946
Mallikarjuna, P., Sitaram, R. T., Landstrom, M., and Ljungberg, B. (2018). VHL status regulates transforming growth factor-beta signaling pathways in renal cell carcinoma. Oncotarget 9, 16297–16310. doi: 10.18632/oncotarget.24631
Margueron, R., Li, G., Sarma, K., Blais, A., Zavadil, J., Woodcock, C. L., et al. (2008). Ezh1 and Ezh2 maintain repressive chromatin through different mechanisms. Mol. Cell 32, 503–518. doi: 10.1016/j.molcel.2008.11.004
Margueron, R., and Reinberg, D. (2011). The Polycomb complex PRC2 and its mark in life. Nature 469, 343–349. doi: 10.1038/nature09784
Marshall, R. S., and Vierstra, R. D. (2018). To save or degrade: balancing proteasome homeostasis to maximize cell survival. Autophagy 14, 2029–2031. doi: 10.1080/15548627.2018.1515531
Martinez, A. M., and Cavalli, G. (2006). The role of polycomb group proteins in cell cycle regulation during development. Cell Cycle 5, 1189–1197. doi: 10.4161/cc.5.11.2781
Matsukawa, Y., Semba, S., Kato, H., Ito, A., Yanagihara, K., and Yokozaki, H. (2006). Expression of the enhancer of zeste homolog 2 is correlated with poor prognosis in human gastric cancer. Cancer Sci. 97, 484–491. doi: 10.1111/j.1349-7006.2006.00203.x
Mimori, K., Ogawa, K., Okamoto, M., Sudo, T., Inoue, H., and Mori, M. (2005). Clinical significance of enhancer of zeste homolog 2 expression in colorectal cancer cases. Eur. J. Surg. Oncol. 31, 376–380. doi: 10.1016/j.ejso.2004.11.001
Mok, Y., Lee, S. J., Kim, M. S., Cui, W., Moon, Y. M., and Jee, S. H. (2012). Serum uric acid and chronic kidney disease: the Severance cohort study. Nephrol. Dial. Transplant 27, 1831–1835. doi: 10.1093/ndt/gfr530
Moledina, D. G., Hall, I. E., Thiessen-Philbrook, H., Reese, P. P., Weng, F. L., Schroppel, B., et al. (2017). Performance of serum creatinine and kidney injury biomarkers for diagnosing histologic acute tubular injury. Am. J. Kidney Dis. 70, 807–816. doi: 10.1053/j.ajkd.2017.06.031
Mozhui, K., and Pandey, A. K. (2017). Conserved effect of aging on DNA methylation and association with EZH2 polycomb protein in mice and humans. Mech. Ageing Dev. 162, 27–37. doi: 10.1016/j.mad.2017.02.006
Ni, J., Hou, X., Wang, X., Shi, Y., Xu, L., Zheng, X., et al. (2019). 3-deazaneplanocin A protects against cisplatin-induced renal tubular cell apoptosis and acute kidney injury by restoration of E-cadherin expression. Cell Death Dis. 10:355. doi: 10.1038/s41419-019-1589-y
Nishiguchi, M. A., Spencer, C. A., Leung, D. H., and Leung, T. H. (2018). Aging suppresses skin-derived circulating SDF1 to promote full-thickness tissue regeneration. Cell Rep. 24, 3383–3392.e3385. doi: 10.1016/j.celrep.2018.08.054
Nogueira, A., Pires, M. J., and Oliveira, P. A. (2017). Pathophysiological Mechanisms of renal fibrosis: a review of animal models and therapeutic strategies. In Vivo 31, 1–22. doi: 10.21873/invivo.11019
Obermayr, R. P., Temml, C., Gutjahr, G., Knechtelsdorfer, M., Oberbauer, R., and Klauser-Braun, R. (2008). Elevated uric acid increases the risk for kidney disease. J. Am. Soc. Nephrol. 19, 2407–2413. doi: 10.1681/ASN.2008010080
Painter, R. C., Osmond, C., Gluckman, P., Hanson, M., Phillips, D. I., and Roseboom, T. J. (2008). Transgenerational effects of prenatal exposure to the Dutch famine on neonatal adiposity and health in later life. Bjog 115, 1243–1249. doi: 10.1111/j.1471-0528.2008.01822.x
Plath, K., Fang, J., Mlynarczyk-Evans, S. K., Cao, R., Worringer, K. A., Wang, H., et al. (2003). Role of histone H3 lysine 27 methylation in X inactivation. Science 300, 131–135. doi: 10.1126/science.1084274
Raimann, J. G., Riella, M. C., and Levin, N. W. (2018). International Society of Nephrology’s 0by25 initiative (zero preventable deaths from acute kidney injury by 2025): focus on diagnosis of acute kidney injury in low-income countries. Clin. Kidney J. 11, 12–19. doi: 10.1093/ckj/sfw134
Raman, J. D., Mongan, N. P., Tickoo, S. K., Boorjian, S. A., Scherr, D. S., and Gudas, L. J. (2005). Increased expression of the polycomb group gene, EZH2, in transitional cell carcinoma of the bladder. Clin Cancer Res. 11(24 Pt 1), 8570–8576. doi: 10.1158/1078-0432.CCR-05-1047
Rohraff, D. M., He, Y., Farkash, E. A., Schonfeld, M., Tsou, P. S., and Sawalha, A. H. (2019). Inhibition of EZH2 ameliorates lupus-like disease in MRL/lpr mice. Arthritis Rheumatol. 71, 1681–1690. doi: 10.1002/art.40931
Ronco, C., Bellomo, R., and Kellum, J. A. (2019). Acute kidney injury. Lancet 394, 1949–1964. doi: 10.1016/S0140-6736(19)32563-2
Rosner, M., and Hengstschlager, M. (2012). Targeting epigenetic readers in cancer. N. Engl. J. Med. 367, 1764–1765. doi: 10.1056/NEJMc1211175
Roy, S., and Majumdar, A. P. (2012). Cancer stem cells in colorectal cancer: genetic and epigenetic changes. J. Stem Cell Res. Ther. 7:6. doi: 10.4172/2157-7633.S7-006
Sakurai, T., Bilim, V. N., Ugolkov, A. V., Yuuki, K., Tsukigi, M., Motoyama, T., et al. (2012). The enhancer of zeste homolog 2 (EZH2), a potential therapeutic target, is regulated by miR-101 in renal cancer cells. Biochem. Biophys. Res. Commun. 422, 607–614. doi: 10.1016/j.bbrc.2012.05.035
Sanchez-Beato, M., Sanchez, E., Gonzalez-Carrero, J., Morente, M., Diez, A., Sanchez-Verde, L., et al. (2006). Variability in the expression of polycomb proteins in different normal and tumoral tissues. A pilot study using tissue microarrays. Mod. Pathol. 19, 684–694. doi: 10.1038/modpathol.3800577
Sanchez-Lozada, L. G., Soto, V., Tapia, E., Avila-Casado, C., Sautin, Y. Y., Nakagawa, T., et al. (2008). Role of oxidative stress in the renal abnormalities induced by experimental hyperuricemia. Am. J. Physiol. Renal Physiol. 295, F1134–F1141. doi: 10.1152/ajprenal.00104.2008
Saramaki, O. R., Tammela, T. L., Martikainen, P. M., Vessella, R. L., and Visakorpi, T. (2006). The gene for polycomb group protein enhancer of zeste homolog 2 (EZH2) is amplified in late-stage prostate cancer. Genes Chromosomes Cancer 45, 639–645. doi: 10.1002/gcc.20327
Sato, Y., and Yanagita, M. (2018). Immune cells and inflammation in AKI to CKD progression. Am. J. Physiol. Renal. Physiol. 315, F1501–F1512. doi: 10.1152/ajprenal.00195.2018
Schlesinger, Y., Straussman, R., Keshet, I., Farkash, S., Hecht, M., Zimmerman, J., et al. (2007). Polycomb-mediated methylation on Lys27 of histone H3 pre-marks genes for de novo methylation in cancer. Nat. Genet. 39, 232–236. doi: 10.1038/ng1950
Sen, P., Shah, P. P., Nativio, R., and Berger, S. L. (2016). Epigenetic mechanisms of longevity and aging. Cell 166, 822–839. doi: 10.1016/j.cell.2016.07.050
Shah, A. R., Lazar, E. L., and Atlas, A. B. (2009). Tracheal diverticula after tracheoesophageal fistula repair: case series and review of the literature. J. Pediatr. Surg. 44, 2107–2111. doi: 10.1016/j.jpedsurg.2009.04.036
Shi, B., Liang, J., Yang, X., Wang, Y., Zhao, Y., Wu, H., et al. (2007). Integration of estrogen and Wnt signaling circuits by the polycomb group protein EZH2 in breast cancer cells. Mol. Cell Biol. 27, 5105–5119. doi: 10.1128/MCB.00162-07
Shi, Y., Tao, M., Wang, Y., Zang, X., Ma, X., Qiu, A., et al. (2020). Genetic or pharmacologic blockade of enhancer of zeste homolog 2 inhibits the progression of peritoneal fibrosis. J. Pathol. 250, 79–94. doi: 10.1002/path.5352
Shi, Y., Xu, L., Tao, M., Fang, L., Lu, J., Gu, H., et al. (2019). Blockade of enhancer of zeste homolog 2 alleviates renal injury associated with hyperuricemia. Am. J. Physiol. Renal. Physiol. 316, F488–F505. doi: 10.1152/ajprenal.00234.2018
Shumaker, D. K., Dechat, T., Kohlmaier, A., Adam, S. A., Bozovsky, M. R., Erdos, M. R., et al. (2006). Mutant nuclear lamin A leads to progressive alterations of epigenetic control in premature aging. Proc. Natl. Acad. Sci. U.S.A. 103, 8703–8708. doi: 10.1073/pnas.0602569103
Siddiqi, F. S., Majumder, S., Thai, K., Abdalla, M., Hu, P., Advani, S. L., et al. (2016). The histone methyltransferase enzyme enhancer of zeste homolog 2 protects against podocyte oxidative stress and renal injury in diabetes. J. Am. Soc. Nephrol. 27, 2021–2034. doi: 10.1681/asn.2014090898
Silva, F. G. (2005). The aging kidney: a review – part I. Int. Urol. Nephrol. 37, 185–205. doi: 10.1007/s11255-004-0873-6
Song, S., Zhang, R., Mo, B., Chen, L., Liu, L., Yu, Y., et al. (2019). EZH2 as a novel therapeutic target for atrial fibrosis and atrial fibrillation. J. Mol. Cell Cardiol. 135, 119–133. doi: 10.1016/j.yjmcc.2019.08.003
Su, I. H., Basavaraj, A., Krutchinsky, A. N., Hobert, O., Ullrich, A., Chait, B. T., et al. (2003). Ezh2 controls B cell development through histone H3 methylation and Igh rearrangement. Nat. Immunol. 4, 124–131. doi: 10.1038/ni876
Sudo, T., Utsunomiya, T., Mimori, K., Nagahara, H., Ogawa, K., Inoue, H., et al. (2005). Clinicopathological significance of EZH2 mRNA expression in patients with hepatocellular carcinoma. Br. J. Cancer 92, 1754–1758. doi: 10.1038/sj.bjc.6602531
Sun, C., Zhao, C., Li, S., Wang, J., Zhou, Q., Sun, J., et al. (2018). EZH2 Expression is increased in BAP1-mutant renal clear cell carcinoma and is related to poor prognosis. J. Cancer 9, 3787–3796. doi: 10.7150/jca.26275
Teruel, M., and Sawalha, A. H. (2017). Epigenetic variability in systemic lupus erythematosus: what we learned from genome-wide DNA methylation studies. Curr. Rheumatol. Rep. 19:32. doi: 10.1007/s11926-017-0657-5
Tie, F., Furuyama, T., and Harte, P. J. (1998). The Drosophila Polycomb Group proteins ESC and E(Z) bind directly to each other and co-localize at multiple chromosomal sites. Development 125, 3483–3496.
Tsou, P. S., Campbell, P., Amin, M. A., Coit, P., Miller, S., Fox, D. A., et al. (2019). Inhibition of EZH2 prevents fibrosis and restores normal angiogenesis in scleroderma. Proc. Natl. Acad. Sci. U.S.A. 116, 3695–3702. doi: 10.1073/pnas.1813006116
Tsou, P. S., Coit, P., Kilian, N. C., and Sawalha, A. H. (2018). EZH2 Modulates the DNA methylome and controls T Cell adhesion through junctional adhesion molecule a in lupus patients. Arthr. Rheumatol. 70, 98–108. doi: 10.1002/art.40338
Tung, C. W., Hsu, Y. C., Shih, Y. H., Chang, P. J., and Lin, C. L. (2018). Glomerular mesangial cell and podocyte injuries in diabetic nephropathy. Nephrology (Carlton) 23(Suppl. 4) 32–37. doi: 10.1111/nep.13451
Veneti, Z., Gkouskou, K. K., and Eliopoulos, A. G. (2017). Polycomb repressor complex 2 in genomic instability and cancer. Int. J. Mol. Sci. 18:1657. doi: 10.3390/ijms18081657
Wagener, N., Macher-Goeppinger, S., Pritsch, M., Husing, J., Hoppe-Seyler, K., Schirmacher, P., et al. (2010). Enhancer of zeste homolog 2 (EZH2) expression is an independent prognostic factor in renal cell carcinoma. BMC Cancer 10:524. doi: 10.1186/1471-2407-10-524
Wan, J., Hou, X., Zhou, Z., Geng, J., Tian, J., Bai, X., et al. (2017). WT1 ameliorates podocyte injury via repression of EZH2/beta-catenin pathway in diabetic nephropathy. Free Radic. Biol. Med. 108, 280–299. doi: 10.1016/j.freeradbiomed.2017.03.012
Wang, C., Oshima, M., Sato, D., Matsui, H., Kubota, S., Aoyama, K., et al. (2018). Ezh2 loss propagates hypermethylation at T cell differentiation-regulating genes to promote leukemic transformation. J. Clin. Invest. 128, 3872–3886. doi: 10.1172/JCI94645
Wang, Y., Fan, L., Cui, C., Wang, Y., and Liang, T. (2018). EZH2 inhibition promotes methyl jasmonate-induced apoptosis of human colorectal cancer through the Wnt/beta-catenin pathway. Oncol. Lett. 16, 1231–1236. doi: 10.3892/ol.2018.8779
Wang, Z., Schones, D. E., and Zhao, K. (2009). Characterization of human epigenomes. Curr. Opin. Genet. Dev. 19, 127–134. doi: 10.1016/j.gde.2009.02.001
Wei, G., Wei, L., Zhu, J., Zang, C., Hu-Li, J., Yao, Z., et al. (2009). Global mapping of H3K4me3 and H3K27me3 reveals specificity and plasticity in lineage fate determination of differentiating CD4+ T cells. Immunity 30, 155–167. doi: 10.1016/j.immuni.2008.12.009
Worden, E. J., Hoffmann, N. A., Hicks, C. W., and Wolberger, C. (2019). Mechanism of Cross-talk between H2B Ubiquitination and H3 Methylation by Dot1L. Cell 176, 1490–1501.e1412. doi: 10.1016/j.cell.2019.02.002
Wu, C. L., Su, T. C., Chang, C. C., Kor, C. T., Chang, C. H., Yang, T. H., et al. (2017). Tubular peroxiredoxin 3 as a predictor of renal recovery from acute tubular necrosis in patients with chronic kidney disease. Sci. Rep. 7:43589. doi: 10.1038/srep43589
Xiao, X., Senavirathna, L. K., Gou, X., Huang, C., Liang, Y., and Liu, L. (2016). EZH2 enhances the differentiation of fibroblasts into myofibroblasts in idiopathic pulmonary fibrosis. Physiol. Rep. 4:12915. doi: 10.14814/phy2.12915
Yang, C., Zhang, Y., Wang, J., Li, L., Wang, L., Hu, M., et al. (2015). A novel cyclic helix B peptide inhibits dendritic cell maturation during amelioration of acute kidney graft rejection through Jak-2/STAT3/SOCS1. Cell Death Dis. 6:e1993. doi: 10.1038/cddis.2015.338
Yang, X. P., Jiang, K., Hirahara, K., Vahedi, G., Afzali, B., Sciume, G., et al. (2015). EZH2 is crucial for both differentiation of regulatory T cells and T effector cell expansion. Sci. Rep. 5:10643. doi: 10.1038/srep10643
Yu, J., Yu, J., Rhodes, D. R., Tomlins, S. A., Cao, X., Chen, G., et al. (2007). A polycomb repression signature in metastatic prostate cancer predicts cancer outcome. Cancer Res. 67, 10657–10663. doi: 10.1158/0008-5472.CAN-07-2498
Yu, M. A., Sanchez-Lozada, L. G., Johnson, R. J., and Kang, D. H. (2010). Oxidative stress with an activation of the renin-angiotensin system in human vascular endothelial cells as a novel mechanism of uric acid-induced endothelial dysfunction. J. Hypertens 28, 1234–1242.
Yuan, J., Shen, Y., Yang, X., Xie, Y., Lin, X., Zeng, W., et al. (2017). Thymosin beta4 alleviates renal fibrosis and tubular cell apoptosis through TGF-beta pathway inhibition in UUO rat models. BMC Nephrol. 18:314. doi: 10.1186/s12882-017-0708-1
Zeng, S., Wu, X., Chen, X., Xu, H., Zhang, T., and Xu, Y. (2018). Hypermethylated in cancer 1 (HIC1) mediates high glucose induced ROS accumulation in renal tubular epithelial cells by epigenetically repressing SIRT1 transcription. Biochim. Biophys. Acta Gene Regul. Mech. 1861, 917–927. doi: 10.1016/j.bbagrm.2018.08.002
Zeybel, M., Luli, S., Sabater, L., Hardy, T., Oakley, F., Leslie, J., et al. (2017). A Proof-of-concept for epigenetic therapy of tissue fibrosis: inhibition of liver fibrosis progression by 3-deazaneplanocin A. Mol. Ther. 25, 218–231. doi: 10.1016/j.ymthe.2016.10.004
Zhang, D., Yang, X. J., Luo, Q. D., Fu, D. L., Li, H. L., Li, H. C., et al. (2018). EZH2 enhances the invasive capability of renal cell carcinoma cells via activation of STAT3. Mol. Med. Rep. 17, 3621–3626. doi: 10.3892/mmr.2017.8363
Zhao, E., Maj, T., Kryczek, I., Li, W., Wu, K., Zhao, L., et al. (2016). Cancer mediates effector T cell dysfunction by targeting microRNAs and EZH2 via glycolysis restriction. Nat. Immunol. 17, 95–103. doi: 10.1038/ni.3313
Zhou, X., Xiong, C., Tolbert, E., Zhao, T. C., Bayliss, G., and Zhuang, S. (2018a). Targeting histone methyltransferase enhancer of zeste homolog-2 inhibits renal epithelial-mesenchymal transition and attenuates renal fibrosis. Faseb J. 32:fj201800237R. doi: 10.1096/fj.201800237R
Zhou, X., Zang, X., Guan, Y., Tolbert, T., Zhao, T. C., Bayliss, G., et al. (2018b). Targeting enhancer of zeste homolog 2 protects against acute kidney injury. Cell Death Dis. 9:1067. doi: 10.1038/s41419-018-1012-0
Zhou, X., Zang, X., Ponnusamy, M., Masucci, M. V., Tolbert, E., Gong, R., et al. (2016). Enhancer of zeste homolog 2 inhibition attenuates renal fibrosis by maintaining Smad7 and phosphatase and tensin homolog expression. J. Am. Soc. Nephrol. 27, 2092–2108. doi: 10.1681/ASN.2015040457
Zhou, X., Zang, X., Ponnusamy, M., Masucci, M. V., and Zhuang, S. (2015). Enhancer of zeste homolog 2 inhibition attenuates renal fibrosis by maintaining smad7 and phosphatase and tensin homolog expression. J. Am. Soc. Nephrol. 27:2092.
Zhou, X. J., Saxena, R., Liu, Z., Vaziri, N. D., and Silva, F. G. (2008). Renal senescence in 2008: progress and challenges. Int. Urol. Nephrol. 40, 823–839. doi: 10.1007/s11255-008-9405-0
Zhou, Y., Fang, L., Jiang, L., Wen, P., Cao, H., He, W., et al. (2012). Uric acid induces renal inflammation via activating tubular NF-kappaB signaling pathway. PLoS One 7:e39738. doi: 10.1371/journal.pone.0039738
Zhu, H., Mi, W., Luo, H., Chen, T., Liu, S., Raman, I., et al. (2016). Whole-genome transcription and DNA methylation analysis of peripheral blood mononuclear cells identified aberrant gene regulation pathways in systemic lupus erythematosus. Arthritis Res. Ther. 18:162. doi: 10.1186/s13075-016-1050-x
Keywords: acute kidney injury, chronic kidney disease, diabetic nephropathy, renal cell carcinoma, epigenetic regulation, histone methyltransferase (HMT), enhancer of zeste homolog 2 (EZH2), renal fibrosis
Citation: Li T, Yu C and Zhuang S (2021) Histone Methyltransferase EZH2: A Potential Therapeutic Target for Kidney Diseases. Front. Physiol. 12:640700. doi: 10.3389/fphys.2021.640700
Received: 11 December 2020; Accepted: 11 January 2021;
Published: 18 February 2021.
Edited by:
Xiao-ming Meng, Anhui Medical University, ChinaReviewed by:
Baoxue Yang, Peking University, ChinaHaiyong Chen, The University of Hong Kong, Hong Kong
Copyright © 2021 Li, Yu and Zhuang. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Shougang Zhuang, szhuang@lifespan.org; gangzhuang@hotmail.com