 Sébastien Lebreton
Sébastien Lebreton Mikael A. Carlsson
Mikael A. Carlsson Peter Witzgall1
Peter Witzgall1- 1Division of Chemical Ecology, Department of Plant Protection Biology, Swedish University of Agricultural Sciences, Alnarp, Sweden
- 2Department of Zoology, Stockholm University, Stockholm, Sweden
Many animals adjust their reproductive behavior according to nutritional state and food availability. Drosophila females for instance decrease their sexual receptivity following starvation. Insulin signaling, which regulates many aspects of insect physiology and behavior, also affects reproduction in females. We show that insulin signaling is involved in the starvation-induced reduction in female receptivity. More specifically, females mutant for the insulin-like peptide 5 (dilp5) were less affected by starvation compared to the other dilp mutants and wild-type flies. Knocking-down the insulin receptor, either in all fruitless-positive neurons or a subset of these neurons dedicated to the perception of a male aphrodisiac pheromone, decreased the effect of starvation on female receptivity. Disrupting insulin signaling in some parts of the brain, including the mushroom bodies even abolished the effect of starvation. In addition, we identified fruitless-positive neurons in the dorso-lateral protocerebrum and in the mushroom bodies co-expressing the insulin receptor. Together, our results suggest that the interaction of insulin peptides determines the tuning of female sexual behavior, either by acting on pheromone perception or directly in the central nervous system.
Introduction
Adjusting reproductive behavior to nutrient availability is a common feature in many animals. Females of many animal species decrease their sexual receptivity to male courtship in response to food deprivation (Kauffman and Rissman, 2004; Pierce et al., 2007; Franssen et al., 2008; Lebreton et al., 2015). In the fruit fly Drosophila melanogaster, feeding, and mating are strongly interconnected. Mating modifies food preference in females (Carvalho et al., 2006; Walker et al., 2015) while feeding regulates their mating behavior (Lebreton et al., 2015, 2016). The molecular mechanisms regulating feeding behavior after mating are fairly well described and involve the transfer of a male component during copulation called Sex Peptide (Carvalho et al., 2006; Ribeiro and Dickson, 2010; Walker et al., 2015). On the other hand, the mechanisms by which feeding and starvation regulate sexual receptivity remain unknown.
One potential system that mechanistically links these two behaviors is insulin signaling as it not only regulates feeding (Broughton et al., 2005; Slaidina et al., 2009; Lebreton et al., 2014) but also various aspects of reproduction such as female attractiveness, egg-laying and remating rate (Yang et al., 2008; Wigby et al., 2011; Kuo et al., 2012). Although, insulin signaling is not necessary for virgin females to be receptive when fed (Wigby et al., 2011; Sakai et al., 2014; Lebreton et al., 2015; Watanabe and Sakai, 2015), so far no study tested the role of this signaling pathway on the sexual behavior of females undergoing a period of starvation. Considering the conserved effect of starvation on sexual receptivity on the one hand, and the fact that insulin signaling both responds to nutrition and regulates reproductive behavior on the other hand, we expected insulin signaling to modulate female sexual receptivity during starvation.
Eight insulin-like peptides (DILP1-8) have been characterized in D. melanogaster while only one insulin receptor is known (InR; Nässel et al., 2013, 2015). DILP2, 3, and 5 are produced in specific cells in the pars intercerebralis called insulin-producing cells (IPCs). In addition, DILP5 is expressed in follicle cells of ovaries and principal cells in renal tubules (Ikeya et al., 2002). DILP6 is, on the other hand, produced by adipose cells (Slaidina et al., 2009). In contrast to other DILPs, DILP7, and DILP8 have been suggested to be more related to relaxin peptides than insulin peptides (Yang et al., 2008; Grönke et al., 2010; Garelli et al., 2015). Whereas, it is unknown whether DILP7 acts through the same InR, it has recently been shown that DILP8 acts via a specific relaxin receptor (Garelli et al., 2015). In addition, two DILPs are expressed almost exclusively during larval stages: DILP1 and DILP4 (Nässel et al., 2015). These different DILPs seem to interact to regulate the fly behavior and metabolism (Grönke et al., 2010; Kannan and Fridell, 2013; Nässel et al., 2013).
DILPs regulate the activity of neuronal circuitries to match behaviors with nutritional status (Wu et al., 2005a,b). Several neuronal networks have been shown to be involved in female sexual receptivity. For instance, neurons expressing the fruitless (fru) gene are necessary for females to be sexually receptive (Demir and Dickson, 2005; Kvitsiani and Dickson, 2006). Similarly, a subset of neurons expressing doublesex (dsx) regulates receptivity, independently of fru (Zhou et al., 2014). Both fru and dsx encode for a transcription factor that are spliced differently in males and females and account for sexually dimorphic traits (Siwicki and Kravitz, 2009). fru is expressed in some pheromone-sensing olfactory sensory neurons (OSNs) expressing the odorant receptors Or67d and Or47b (Stockinger et al., 2005). Or67d detects the male aphrodisiac pheromone cis-vaccenyl acetate (cVA) while Or47b responds to methyl laurate, a compound produced by flies of both sexes (Dweck et al., 2015). Both OSNs expressing Or67d and Or47b have been shown to modulate female receptivity (Kurtovic et al., 2007; Sakurai et al., 2013). In addition to these neurons, an early study found a group of cells in the dorsal anterior brain to be necessary and sufficient to induce receptivity (Tompkins and Hall, 1983), though these cells have never been precisely characterized. Interestingly, these cells seem to be different from those required to perform courtship in males (Tompkins and Hall, 1983), suggesting that they are probably not fru-positive. In young virgin females, acquisition of sexual receptivity is paralleled by the growth of the ovaries and the corpora allata, endocrine glands producing the juvenile hormone (JH, Manning, 1967). Both JH and the corpora allata have been shown to be involved in this switch in female receptivity (Manning, 1967; Ringo et al., 1991). Interestingly, the corpora allata activity is modulated by the insulin pathway (Tu et al., 2005; Belgacem and Martin, 2007). However, whether or not insulin acts on these structures to regulate the female sexual receptivity is unknown.
In conclusion, despite the fact that insulin is known to both regulate reproductive behavior and respond to food intake/deprivation, its effect in the starvation-induced reduction in female receptivity has not yet been established. We hypothesize that, during starvation, insulin signaling acts on specific neuronal networks to reduce sexual receptivity in order to match nutrition and reproduction. In the present study, we first carefully analyzed the effect of starvation in female mating behavior in Drosophila. We then tested the role of insulin by analyzing the behavior of females deficient for seven different DILPs. Finally, in an attempt to identify some structures on which insulin signaling acts to regulate sexual behavior, we knocked down InR in various brain parts known to be involved in mating behavior or behavioral modulation.
Materials and Methods
Insects
Drosophila flies were reared on a sugar-yeast-cornmeal medium diet under a 12:12 h L:D photoperiod. Virgin flies were collected within 6 h following adult emergence. They were anesthetized with CO2 and separated by sex under a microscope. Flies of the same sex (males and females) were then kept in 30-ml plastic tubes with fresh diet for 3 days before behavioral experiments. For starvation, females were transferred to a humidified piece of cotton for 1, 2, or 3 days before being tested. Flies starved for 3 days were allowed to feed up to 6 h following adult emergence, before they were collected for experiments.
For behavioral experiments, the Dalby strain was used as a wild-type strain (Ruebenbauer et al., 2008). In order to test the effect of the different insulin-like peptides on female receptivity, mutant females for single dilps (dilp1 to 7) or multiple dilps (dilp2-3,5) were used. DILP8, which has been shown to be more related to relaxin peptides than insulin and to act via its own receptor, was not included in this study. In order to identify structures involved in this behavioral modulation, we manipulated insulin signaling in specific parts of the body. For this purpose, InR was knocked-down by crossing a line expressing an InR RNAi (uas-InR RNAi) to lines expressing specific Gal4 drivers (Fru-Gal4 for Fruitless-positive neurons, Aug21-Gal4 for the corpora allata, OK107-Gal4 for the mushroom bodies and Or67d-Gal4 for Or67d-expressing sensory neurons). All mutant and transgenic lines used and their origins are listed in Table 1.
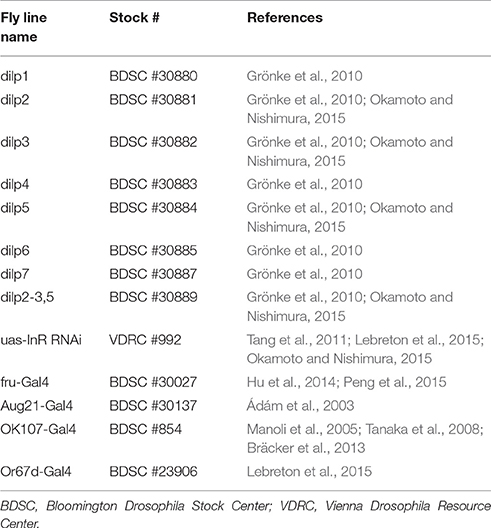
Table 1. Mutant and transgenic fly lines used in this study.
Mating Behavior
One random wild-type (Dalby) 3d-old fed male was introduced together with a virgin female (either wild-type, mutant or transgenic) under a small round inverted plastic cup (45 mm in diameter, 30 mm high) placed on a clean glass plate. Wild-type females were either fed or starved for 1, 2, or 3 days (n = 52, 54, 77, and 47, respectively). Mutant and transgenic females were tested fed or 2-day starved. For mutant and transgenic flies, 33–50 couples were tested for each genotype and feeding condition in order to obtain 30–35 replicates of successful courtship.
Flies were observed for 60 min. Male courtship, courtship latency (time between the beginning of the test and courtship initiation), mating latency (time between the beginning of courtship and copulation initiation) and mating duration were recorded. To estimate female sexual receptivity, only cases where courtship from males was observed were taken into account. The percentage of females accepting to mate with a courting male within the 60-min period was then calculated for each 5-min interval.
Percentages of courting males were analyzed using a χ2-test. Courtship latency, mating latency and mating duration were analyzed using a Generalized Linear Model (GLM) with a Gamma family.
For each wild-type, mutant and transgenic line, an estimated decrease of sexual receptivity was calculated. For this purpose, the area under curve (AUC) was used as a proxy for sexual receptivity (R: auc function in kulife package). The percentage of decrease between AUC of fed and starved females was then calculated.
Regarding the percentage of mating, repeated measurements on the same flies over time were treated as pseudo-replicates and therefore analyzed using a Linear Mixed-effect Model (GLMM, R: glmer function in lme4 package) with a binomial distribution and the “time” factor as a random effect (Crawley, 2007). Similarly, the percentage by which mating was decreased between fed and starved females was compared between mutant and wild-type flies using a GLMM with a Gamma family. When GLM and GLMM showed a significant effect of the treatment or genotype, the test was followed by a multiple comparison test with a FDR correction method (R: glht function in multcomp package).
Statistical analyses were performed in R (R 2.1.1, R Development Core Team, Free Software Foundation Boston, MA, USA).
Immunostaining
Standard immunohistochemical methods were used as earlier described in detail (Carlsson et al., 2010). In brief, dissected brains were fixed for 4 h at 4°C in 4% paraformaldehyde (PFA) and subsequently washed several times in phosphate buffer. The brains were then preincubated overnight in incubation buffer containing 0.01 M phosphate-buffered saline (PBS), 0.25% BSA, 0.25% Triton-X and 3% normal goat serum. Then the brains were incubated with a cocktail of the primary antibodies, mouse monoclonal GFP antibody (1:1,000, Molecular Probes, Invitrogen) and rabbit anti InR (1:1,000, #3021, Cell Signaling Technology) for 72 h at 4°C under gentle agitation. For detection of primary antisera, Alexa goat anti-rabbit 488 and Alexa goat anti-mouse 546 (Invitrogen) were used at a dilution of 1:500 at 4°C overnight, washed in PBS-Tx and PBS and finally mounted in 80% glycerol in PBS.
Imaging
Brains were imaged with a Zeiss LSM 780 confocal microscope (Zeiss Jena, Germany) and stacked images were processed using ZEN 2011 software (Zeiss) and edited for intensity and contrast in Adobe Photoshop CS6.
Results
Starvation Regulates Female Sexual Receptivity
We found that female sexual receptivity was significantly reduced after a period of starvation. When fed, 96% of virgin females mated within 1 h of being paired with a wild type male. However, this percentage dropped to 70, 37, and 16% after a starvation period of 1, 2, and 3 days, respectively (Figure 1A). Although, starved females also tended to have longer mating latencies, this effect was not significant (Figure 1B).
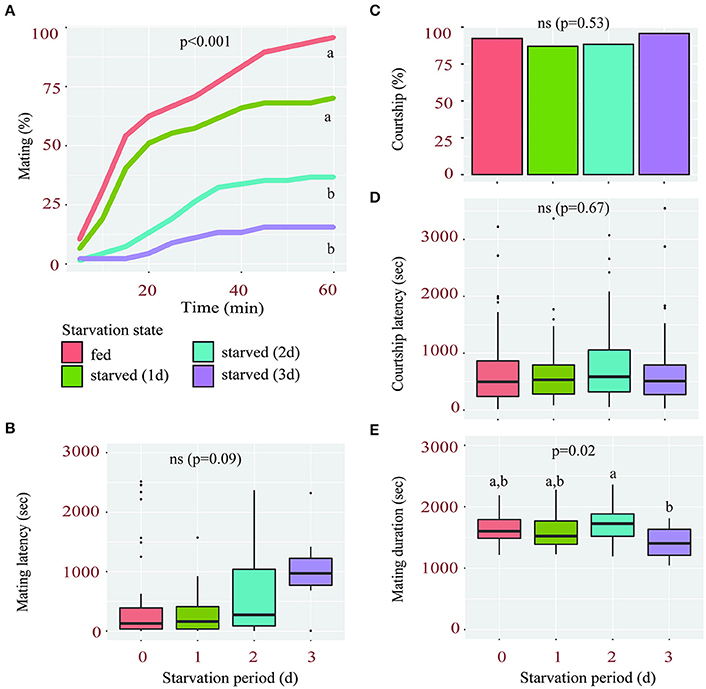
Figure 1. Starvation affects female sexual receptivity. (A) Percentage of wild-type females accepting to mate with a courting male during a 1 h-period according to their starvation state (fed or starved for 1, 2, or 3 days). Linear Mixed-Effect Model (p < 0.001), different letters show statistical significant differences between starvation states. (B) Mating latency (time from courtship initiation to mating) of fed and starved females. (C–E) Percentage of males courting fed and starved females (C), their corresponding courtship latency (D) and mating duration (E).
The drop in sexual receptivity observed in starved females was likely not due to changes in male courtship or mating behavior. First, neither the percentage of courting males (Figure 1C) nor their courtship latency (Figure 1D) was significantly affected by the starvation state of the females they were exposed to, suggesting that all females elicited a similar intensity of courtship. Second, the mean mating duration, a male-regulated aspect of copulation reflecting his investment (Bretman et al., 2009; Wigby et al., 2009), was not significantly shorter in any of the starvation conditions compared to fed females (Figure 1E). Taken together, these findings suggest that the starvation-induced reduction in female receptivity is likely due to altered molecular processes within the female and not reduced male interest.
The Decrease of Sexual Receptivity after Starvation Is Less Pronounced in dilp5 Mutants than in other dilp Mutants and Wild-type Flies
Given the preponderant role of insulin signaling in response to changes in nutritional states, we then tested whether it could be involved in the regulation of sexual receptivity during starvation. We therefore investigated the mating behavior of seven dilp mutant females.
Similar to wild type flies, females carrying a mutation in either one or multiple dilp genes showed significantly reduced sexual receptivity after 2 days of starvation (Figure 2A). Although, the sexual receptivity of fed females varies between lines, the magnitude by which starvation affects sexual receptivity in these lines is overall similar. Indeed, the sexual receptivity of dilp1, dilp2, dilp3, dilp6, and dilp7 mutants is decreased by 64 to 72%. This is similar to what was observed in wild-type flies (Figure 1A, 66%), even though their genetic background differs. Females mutant for dilp4 showed a reduction of 56% of their sexual receptivity. dilp5 mutant females were the least affected with a decrease of only 31%. Indeed, the percentage by which mating was decreased after starvation was significantly lower in dilp5 mutants compared to other single dilp mutants and wild-type flies (Figure 2B). Interestingly, the effect of the lack of DILP5 was not present in a dilp2-3 mutant background (Figure 2, dilp2-3,5 mutants: −72%).
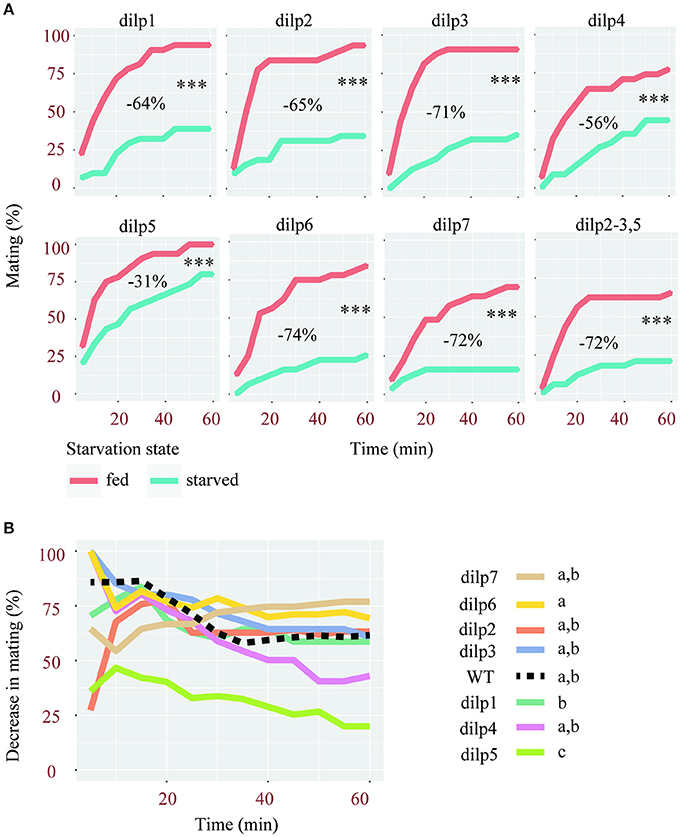
Figure 2. Mutations of single dilps differentially affect female receptivity. (A) Effect of starvation on the receptivity of females, mutant for different dilps. Tested females were either fed or starved for 2 days. ***p < 0.001 (Linear Mixed-Effect Model). For each mutant line is indicated the decrease of sexual receptivity after starvation, estimated based on the area under curves (AUC). (B) Comparison of the percentage of decrease in mating after starvation in single dilp mutant and wild-type females (Linear Mixed-Effect Model, p < 0.001), different letters show statistical significant differences between fly lines.
In conclusion, among all flies deficient for one or several DILPs, only the lack of DILP5 significantly reduced the effect of starvation.
Disrupting the Insulin Signaling in Specific Neuronal Circuitries Inhibits the Effect of Starvation on Sexual Receptivity
Although, the effect was less pronounced in dilp5 mutants, the sexual receptivity of all dilp mutants was significantly reduced after 2 days of starvation, suggesting that insulin signaling is not involved. However, compensatory mechanisms exist among DILPs (Broughton et al., 2008; Grönke et al., 2010), which may have led to a masking effect when the expression of a single dilp was abolished. Since all DILPs act via a single insulin receptor (InR), disrupting InR and therefore all DILP signaling could reveal hidden effects. We therefore knocked-down InR in specific parts of the body, using specific Gal4 drivers. These drivers were chosen because they target specific sites known to regulate different aspects of Drosophila behavior. Fruitless-positive neurons (Fru-Gal4) are involved in sexually dimorphic behavior (Siwicki and Kravitz, 2009), Or67d-expressing OSNs (a sub-population of Fruitless-positive neurons, Or67d-Gal4) detect the male aphrodisiac pheromone cVA (Datta et al., 2008), cells of the corpora allata (Aug21-Gal4) are necessary for young virgin females to become sexually receptive (Manning, 1966, 1967) and the mushroom bodies (OK107-Gal4) are involved various behavioral processes such as courtship and decision making (Zars, 2000). Of note, OK107-Gal4 also drives expression to some extent in the pars intercerebralis, optic lobes, the subesophageal ganglion, the tritocerebrum, and the antennal lobes (Aso et al., 2009).
In all control lines (uas-InR RNAi and Gal4 lines) sexual receptivity was negatively affected by starvation (Figure 3A). However, the genetic background appears to have a substantial effect on the amplitude by which starvation affects female sexual receptivity (Figure 3A). Of note, the uas-InR RNAi line itself was only little affected (reduction of 20% of the sexual receptivity after starvation, Figure 3A). Nevertheless, knocking-down InR in Fruitless-positive neurons, or only in a subset of these neurons (Or67d-OSNs) reduced the effect of starvation on receptivity (with a reduction of only 8 to 10%). In fact, the difference in sexual receptivity between starved and fed females was not statistically significant in these flies. Similarly, when knocking-down InR in the mushroom bodies using the OK107-Gal4 driver, the effect of starvation was abolished. Interestingly, in this case, starved females were even slightly more receptive than fed females, although this effect was not statistically significant (increased receptivity of 8% in starved flies). In contrast, we did not observe any effect by expressing an InR RNAi in the corpora allata, with an effect intermediate to those of the two control lines (Figure 3A).
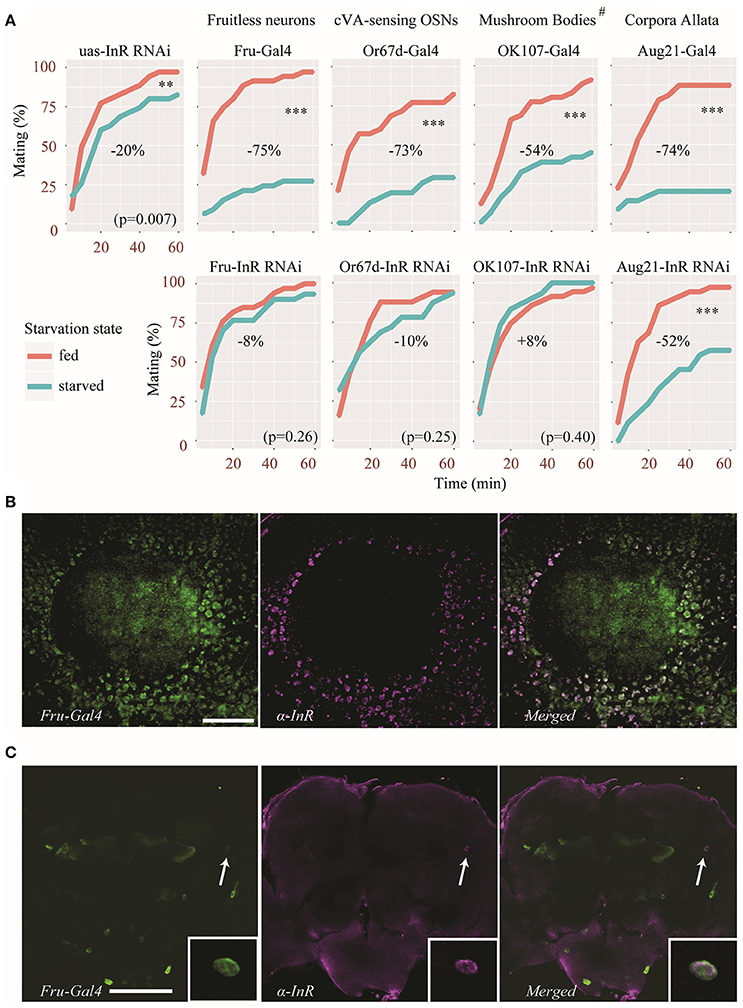
Figure 3. Insulin signaling acts on specific neuronal circuitry to regulate sexual receptivity during starvation. (A) An InR RNAi was expressed in Fruitless-positive neurons, the corpora allata, the mushroom bodies and cVA-sensing neurons, using specific Gal4 drivers. Top panels show the behavior of control parental lines. Bottom panels show the behavior of flies in which the InR RNAi was expressed. Estimated difference in sexual receptivity (based on the calculation of the area under curve) after starvation is indicated for each line. **p < 0.01, ***p < 0.001 (Linear Mixed-Effect Model). #OK107-Gal4 also drives expression to some extent to other parts of the brain (see text for details). (B) InR immunoreactivity (magenta) in Kenyon cells in the calyx (CA) of a mushroom body also showing Fru-Gal4 driven GFP expression (green). Scale bar = 20 μm. (C) Colocalization of InR immunoreactivity (magenta) in a pair of neurons showing Fru-Gal4 driven GFP expression (green) in the dorso-lateral protocerebrum (arrow). The inset shows a detail of the co-expression (from a different focal plane). Scale bar = 100 μm.
InR is Expressed in Fruitless-Positive Neurons
Our behavioral analysis revealed that knocking down InR in Fruitless-positive neurons diminished the effect of starvation on female receptivity. We therefore performed immunostaining on fly brains in order to visualize which Fruitless-positive neurons express the insulin receptor. A large number of Fruitless-positive Kenyon cells were also immunoreactive to the insulin receptor antibody as can be observed in Figure 3B. In addition, we found InR immunolabeling in a pair of Fruitless-positive anterior dorso-lateral neurons (Figure 3C). Thus, at least in Kenyon cells of the mushroom bodies and in a few protocerebral cells, InR and Fru are co-expressed.
Discussion
Drosophila females need nutrients to produce eggs and a nutrient rich substrate to lay their eggs (Drummond-Barbosa and Spradling, 2001; Becher et al., 2012). When food is scarce it would therefore be beneficial for flies to decrease their sexual behavior and to focus on food searching instead. On the other hand, female flies can store sperm and use it several days later when conditions are suitable (Qazi et al., 2003). It could therefore be optimal for females to remain receptive for short periods of food deprivation. Several insulin peptides produced in specific spatiotemporal patterns acting through one single receptor enables a fine-scale regulation of behaviors in response to changes in physiology. The expression of the different dilps is differentially affected by food quality or food deprivation (Bai et al., 2012; Whitaker et al., 2014; Post and Tatar, 2016). For instance, both starvation and dietary restriction reduce the expression of dilp5 but increase the expression of dilp6, while the expression of dilp2 is not affected by either condition (Bai et al., 2012; Whitaker et al., 2014). Our results suggest that DILP5 might be involved in the decrease of receptivity during non-feeding stages. Indeed, dilp5 mutant females were less affected by starvation than other dilp mutants. The effect of the lack of DILP5 was no longer observed in the simultaneous absence of DILP2 and DILP3. Although, we cannot completely rule out background mutation effects, this suggests that DILP5 might interact with other DILPs to finely tune female sexual receptivity.
Insulin is known to act on the olfactory system to modulate odor sensitivity after feeding (Root et al., 2011). Moreover, normal InR expression in Or67d-expressing (Fruitless-positive) OSNs is necessary for fed females to be attracted to a blend of food odors and cVA (Lebreton et al., 2015), a pheromone promoting sexual receptivity (Kurtovic et al., 2007). Our results suggest that insulin signaling in Fruitless-positive neurons, and more specifically in Or67d OSNs may decrease sexual receptivity during starvation.
Fruitless-positive cells other than pheromone-sensing neurons can also be involved. We found different Fruitless-positive cells in the protocerebrum that strongly express InR. First of all, a large number of Kenyon cells in the calyx of the mushroom bodies express both Fruitless and the insulin receptor. Additionally, we found one pair of neurons with somata located in the anterior dorso-lateral protocerebrum. We could not trace any processes from these somata and do thus not know what neuropils they innervate. However, the fact that InR immunostaining was observed in Fruitless neurons, most of which were Kenyon cells, corroborate our behavioral results. Indeed, the sexual receptivity of females in which insulin signaling was knocked down in the mushroom bodies was not affected by starvation. Interestingly, the mushroom bodies are not required for virgin females to be receptive (Neckameyer, 1998), suggesting that these structures may regulate the activity of neuronal networks inducing sexual receptivity. However, this result must be take with caution, given the fact that the Gal4 line we used to target the mushroom bodies also drives expression to some extent in other brain tissues (Aso et al., 2009). Further experiments will be necessary to confirm that the mushroom bodies are indeed responsible for this effect.
Insulin signaling not only modulates neuronal activity in adults but also shapes neuronal networks during development (Song et al., 2003). The effects we observed in our study may therefore be the consequence of a developmental defect of specific neuronal circuitry rather than a direct effect of insulin on these neurons during starvation. However, Fruitless-positive neurons being required for females to be receptive (Kvitsiani and Dickson, 2006), we would expect fed females to be unreceptive if the disruption of insulin signaling had altered the connectivity of these neurons during development, which was not the case. This suggests that insulin acts on these neurons during adult stage to modulate sexual receptivity. This is different for the mushroom bodies, which are not necessary for females to be receptive (Neckameyer, 1998). Knocking down InR specifically during development or specifically in adults will be necessary to disentangle these two possible modes of action of insulin.
In contrast with Fruitless neurons and the mushroom bodies, we did not observe any effect of the corpora allata in the insulin-dependent control of sexual receptivity, whereas these structures have been linked to the development of receptivity in virgin females (Manning, 1966, 1967). This result should however be taken with caution, considering the behavioral variability displayed by the different transgenic lines, which would have prevented us from observing subtle changes. Nonetheless, our results suggest that the structures that generate behaviors (such as the corpora allata) and those modulating these behaviors (for example the mushroom bodies) can be different and the underlying mechanisms uncoupled.
Taken together, Drosophila flies adjust their sexual behavior to match their nutritional state. Together with other hormonal pathways (Lebreton et al., 2016), insulin regulates some aspects of sexual activity (Wigby et al., 2011; Kuo et al., 2012; Sakai et al., 2014; Watanabe and Sakai, 2015), both after food intake (Lebreton et al., 2015) and after a period of starvation. Our results suggest that specific insulin peptides regulate female receptivity, possibly by acting on pheromone perception at the periphery or directly in the central nervous system. Indeed, the mushroom bodies probably play a major role in the insulin-dependent effect of starvation on female sexual receptivity. The next step will be to untangle the specific neuronal circuitry involved.
Ethics Statement
All the experiments described in the manuscript were performed with laboratory-reared insects. No recommendations from an ethical committee were required.
Author Contributions
SL and PW conceived the project. SL designed, performed and analyzed the behavioral experiments. MC performed the immunostaining and the acquisition of confocal microscopy images. SL wrote the first draft of the manuscript. All authors contributed to the final version of the manuscript.
Funding
SL was supported by Carl Tryggers Stiftelse för Vetenskaplig Forskning (grant number: CTS 13:512).
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The reviewer ML and handling Editor declared their shared affiliation.
References
Ádám, G., Perrimon, N., and Noselli, S. (2003). The retinoic-like juvenile hormone controls the looping of left-right asymmetric organs in Drosophila. Development 130, 2397–2406. doi: 10.1242/dev.00460
Aso, Y., Grübel, K., Busch, S., Friedrich, A. B., and Tanimoto, H. (2009). The mushroom body of adult Drosophila characterized by GAL4 drivers. J. Neurogenet. 23, 156–172. doi: 10.1080/01677060802471718
Bai, H., Kang, P., and Tatar, M. (2012). Drosophila insulin-like peptide-6 (dilp6) expression from fat body extends lifespan and represses secretion of Drosophila insulin-like peptide-2 from the brain. Aging Cell 11, 978–985. doi: 10.1111/acel.12000
Becher, P. G., Flick, G., Rozpędowska, E., Schmidt, A., Hagman, A., Lebreton, S., et al. (2012). Yeast, not fruit volatiles mediate Drosophila melanogaster attraction, oviposition and development. Funct. Ecol. 26, 822–828. doi: 10.1111/j.1365-2435.2012.02006.x
Belgacem, Y. H., and Martin, J. R. (2007). Hmgcr in the corpus allatum controls sexual dimorphism of locomotor activity and body size via the insulin pathway in Drosophila. PLoS ONE 2:e187. doi: 10.1371/journal.pone.0000187
Bräcker, L. B., Siju, K. P., Varela, N., Aso, Y., Zhang, M., Hein, I., et al. (2013). Report essential role of the mushroom body in context-dependent CO2 avoidance in Drosophila. Curr. Biol. 23, 1228–1234. doi: 10.1016/j.cub.2013.05.029
Bretman, A., Fricke, C., and Chapman, T. (2009). Plastic responses of male Drosophila melanogaster to the level of sperm competition increase male reproductive fitness. Proc. R. Soc. B Biol. Sci. 276, 1705–1711. doi: 10.1098/rspb.2008.1878
Broughton, S. J., Piper, M. D. W., Ikeya, T., Bass, T. M., Jacobson, J., Driege, Y., et al. (2005). Longer lifespan, altered metabolism, and stress resistance in Drosophila from ablation of cells making insulin-like ligands. Proc. Natl. Acad. Sci. U.S.A. 102, 3105–3110. doi: 10.1073/pnas.0405775102
Broughton, S., Alic, N., Slack, C., Bass, T., Ikeya, T., Vinti, G., et al. (2008). Reduction of DILP2 in Drosophila Triages a metabolic phenotype from lifespan revealing redundancy and compensation among DILPs. PLoS ONE 3:e3721. doi: 10.1371/journal.pone.0003721
Carlsson, M. A., Diesner, M., Schachtner, J., and Nässel, D. R. (2010). Multiple neuropeptides in the Drosophila antennal lobe suggest complex modulatory circuits. J. Comp. Neurol. 518, 3359–3380. doi: 10.1002/cne.22405
Carvalho, G. B., Kapahi, P., Anderson, D. J., and Benzer, S. (2006). Allocrine modulation of feeding behavior by the Sex Peptide of Drosophila. Curr. Biol. 16, 692–696. doi: 10.1016/j.cub.2006.02.064
Datta, S. R., Vasconcelos, M. L., Ruta, V., Luo, S., Wong, A., Demir, E., et al. (2008). The Drosophila pheromone cVA activates a sexually dimorphic neural circuit. Nature 452, 473–477. doi: 10.1038/nature06808
Demir, E., and Dickson, B. J. (2005). Fruitless splicing specifies male courtship behavior in Drosophila. Cell 121, 785–794. doi: 10.1016/j.cell.2005.04.027
Drummond-Barbosa, D., and Spradling, A. C. (2001). Stem cells and their progeny respond to nutritional changes during Drosophila Oogenesis. Dev. Biol. 231, 265–278. doi: 10.1006/dbio.2000.0135
Dweck, H. K. M., Ebrahim, S. A. M., Thoma, M., Mohamed, A. A. M., Keesey, I. W., Trona, F., et al. (2015). Pheromones mediating copulation and attraction in Drosophila. Proc. Natl. Acad. Sci. U.S.A. 112, E2829–E2835. doi: 10.1073/pnas.1504527112
Franssen, C. M., Tobler, M., Riesch, R., García de León, F. J., Tiedemann, R., Schlupp, I., et al. (2008). Sperm production in an extremophile fish, the cave molly (Poecilia mexicana, Poeciliidae, Teleostei). Aquat. Ecol. 17, 685–692. doi: 10.1007/s10452-007-9128-9
Garelli, A., Heredia, F., Casimiro, A. P., Macedo, A., Nunes, C., Garcez, M., et al. (2015). Dilp8 requires the neuronal relaxin receptor Lgr3 to couple growth to developmental timing. Nat. Commun. 6:8732. doi: 10.1038/ncomms9732
Grönke, S., Clarke, D.-F., Broughton, S., Andrews, T. D., and Partridge, L. (2010). Molecular evolution and functional characterization of Drosophila insulin-like peptides. PLoS Genet. 6:e1000857. doi: 10.1371/journal.pgen.1000857
Hu, Y., Han, Y., Wang, X., and Xue, L. (2014). Aging-related neurodegeneration eliminates male courtship choice in Drosophila. Neurobiol. Aging 35, 2174–2178. doi: 10.1016/j.neurobiolaging.2014.02.026
Ikeya, T., Galic, M., Belawat, P., Nairz, K., and Hafen, E. (2002). Nutrient-dependent expression of insulin-like peptides from neuroendocrine cells in the CNS contributes to growth regulation in Drosophila. Curr. Biol. 12, 1293–1300. doi: 10.1016/S0960-9822(02)01043-6
Kannan, K., and Fridell, Y.-W. C. (2013). Functional implications of Drosophila insulin-like peptides in metabolism, aging, and dietary restriction. Front. Physiol. 4:288. doi: 10.3389/fphys.2013.00288
Kauffman, A. S., and Rissman, E. F. (2004). A critical role for the evolutionarily conserved gonadotropin-releasing hormone II: mediation of energy status and female sexual behavior. Endocrinology 145, 3639–3646. doi: 10.1210/en.2004-0148
Kuo, T.-H., Fedina, T. Y., Hansen, I., Dreisewerd, K., Dierick, H. A., Yew, J. Y., et al. (2012). Insulin signaling mediates sexual attractiveness in Drosophila. PLoS Genet. 8:e1002684. doi: 10.1371/journal.pgen.1002684
Kurtovic, A., Widmer, A., and Dickson, B. J. (2007). A single class of olfactory neurons mediates behavioural responses to a Drosophila sex pheromone. Nature 446, 542–546. doi: 10.1038/nature05672
Kvitsiani, D., and Dickson, B. J. (2006). Shared neural circuitry for female and male sexual behaviours in Drosophila positive selection on gene expression in the human brain. Curr. Biol. 16, R355–R356. doi: 10.1016/j.cub.2006.04.025
Lebreton, S., Mansourian, S., Bigarreau, J., and Dekker, T. (2016). The Adipokinetic hormone receptor modulates sexual behavior, Pheromone perception and pheromone production in a sex-specific and starvation-dependent manner in Drosophila melanogaster. Front. Ecol. Evol. 3:151. doi: 10.3389/fevo.2015.00151
Lebreton, S., Trona, F., Borrero-Echeverry, F., Bilz, F., Grabe, V., Becher, P. G., et al. (2015). Feeding regulates sex pheromone attraction and courtship in Drosophila females. Sci. Rep. 5:13132. doi: 10.1038/srep13132
Lebreton, S., Witzgall, P., Olsson, M., and Becher, P. G. (2014). Dietary glucose regulates yeast consumption in adult Drosophila males. Front. Physiol. 5:504. doi: 10.3389/fphys.2014.00504
Manning, A. (1966). Corpus allatum and sexual receptivity in female Drosophila melanogaster. Nature 211, 1321–1322. doi: 10.1038/2111321b0
Manning, A. (1967). The control of sexual receptivity in female Drosophila. Anim. Behav. 15, 239–260. doi: 10.1016/0003-3472(67)90006-1
Manoli, D. S., Foss, M., Villella, A., Taylor, B. J., Hall, J. C., and Baker, B. S. (2005). Male-specific fruitless specifies the neural substrates of Drosophila courtship behaviour. Nature 436, 395–400. doi: 10.1038/nature03859
Nässel, D. R., Kubrak, O. I., Liu, Y., Luo, J., and Lushchak, O. V. (2013). Factors that regulate insulin producing cells and their output in Drosophila. Front. Physiol. 4:252. doi: 10.3389/fphys.2013.00252
Nässel, D. R., Liu, Y., and Luo, J. (2015). Insulin/IGF signaling and its regulation in Drosophila. Gen. Comp. Endocrinol. 221, 255–266. doi: 10.1016/j.ygcen.2014.11.021
Neckameyer, W. S. (1998). Dopamine and mushroom bodies in Drosophila : experience-dependent and -independent aspects of sexual behavior. Learn. Mem. 5, 157–166.
Okamoto, N., and Nishimura, T. (2015). Signaling from glia and cholinergic neurons controls nutrient-dependent production of an insulin-like peptide for Drosophila body growth. Dev. Cell 35, 295–310. doi: 10.1016/j.devcel.2015.10.003
Peng, F., Zhao, Y., Huang, X., Chen, C., Sun, L., Zhuang, L., et al. (2015). Loss of Polo ameliorates APP- induced Alzheimer's disease-like symptoms in Drosophila. Sci. Rep. 5:16816. doi: 10.1038/srep16816
Pierce, A. A., Iwueke, I., and Ferkin, M. H. (2007). Food deprivation and the role of estradiol in mediating sexual behaviors in meadow voles. Physiol. Behav. 90, 353–361. doi: 10.1016/j.physbeh.2006.09.034
Post, S., and Tatar, M. (2016). Nutritional geometric profiles of insulin / IGF expression in Drosophila melanogaster. PLoS ONE 11:e0155628. doi: 10.1371/journal.pone.0155628
Qazi, M. C. B., Heifetz, Y., and Wolfner, M. F. (2003). The developments between gametogenesis and fertilization: ovulation and female sperm storage in Drosophila melanogaster. Dev. Biol. 256, 195–211. doi: 10.1016/S0012-1606(02)00125-2
Ribeiro, C., and Dickson, B. J. (2010). Sex peptide receptor and neuronal TOR/S6K signaling modulate nutrient balancing in Drosophila. Curr. Biol. 20, 1000–1005. doi: 10.1016/j.cub.2010.03.061
Ringo, J., Werczberger, R., Altaratz, M., and Segal, D. (1991). Female sexual receptivity is defective in juvenile hormone-deficient mutants of the apterous gene of Drosophila melanogaster. Behav. Genet. 21, 453–469. doi: 10.1007/BF01066724
Root, C. M., Ko, K. I., Jafari, A., and Wang, J. W. (2011). Presynaptic facilitation by neuropeptide signaling mediates odor-driven food search. Cell 145, 133–144. doi: 10.1016/j.cell.2011.02.008
Ruebenbauer, A., Schlyter, F., Hansson, B. S., Löfstedt, C., and Larsson, M. C. (2008). Genetic variability and robustness of host odor preference in Drosophila melanogaster. Curr. Biol. 18, 1438–1443. doi: 10.1016/j.cub.2008.08.062
Sakai, T., Watanabe, K., Ohashi, H., Sato, S., Inami, S., and Shimada, N. (2014). Insulin-producing cells regulate the sexual receptivity through the painless TRP channel in Drosophila virgin females. PLoS ONE 9:e88175. doi: 10.1371/journal.pone.0088175
Sakurai, A., Koganezawa, M., Yasunaga, K., Emoto, K., and Yamamoto, D. (2013). Select interneuron clusters determine female sexual receptivity in Drosophila. Nat. Commun. 4:1825. doi: 10.1038/ncomms2837
Siwicki, K. K., and Kravitz, E. A. (2009). Fruitless, doublesex and the genetics of social behavior in Drosophila melanogaster. Curr. Opin. Neurobiol. 19, 200–206. doi: 10.1016/j.conb.2009.04.001
Slaidina, M., Delanoue, R., Gronke, S., Partridge, L., and Léopold, P. (2009). A Drosophila Insulin-like peptide promotes growth during Nonfeeding States. Dev. Cell 17, 874–884. doi: 10.1016/j.devcel.2009.10.009
Song, J., Wu, L., Chen, Z., Kohanski, R. A., and Pick, L. (2003). Axons guided by insulin receptor in Drosophila visual system. Science 300, 502–506. doi: 10.1126/science.1081203
Stockinger, P., Kvitsiani, D., Rotkopf, S., Tirian, L., and Dickson, B. J. (2005). Neural circuitry that governs Drosophila male courtship behavior. Cell 121, 795–807. doi: 10.1016/j.cell.2005.04.026
Tanaka, N. K., Tanimoto, H., and Ito, K. E. I. (2008). Neuronal assemblies of the Drosophila mushroom body. J. Comp. Neurol. 508, 711–755. doi: 10.1002/cne.21692
Tang, H. Y., Smith-Caldas, M. S. B., Driscoll, M. V., Salhadar, S., and Shingleton, A. W. (2011). FOXO regulates organ-specific phenotypic plasticity in Drosophila. PLoS Genet. 7:e1002373. doi: 10.1371/journal.pgen.1002373
Tompkins, L., and Hall, J. C. (1983). Identification of brain sites controlling female receptivity in mosaics of Drosophila melanogaster. Genetics 103, 179–195.
Tu, M.-P., Yin, C.-M., and Tatar, M. (2005). Mutations in insulin signaling pathway alter juvenile hormone synthesis in Drosophila melanogaster. Gen. Comp. Endocrinol. 142, 347–356. doi: 10.1016/j.ygcen.2005.02.009
Walker, S. J., Corrales-Carvajal, V. M., and Ribeiro, C. (2015). Postmating circuitry modulates salt taste processing to increase reproductive output in Drosophila. Curr. Biol. 25, 2621–2630. doi: 10.1016/j.cub.2015.08.043
Watanabe, K., and Sakai, T. (2015). Knockout mutations of insulin-like peptide genes enhance sexual receptivity in Drosophila virgin females. Genes Genet. Syst. 90, 237–241. doi: 10.1266/ggs.15-00025
Whitaker, R., Gil, M. P., Ding, F., Tatar, M., Helfand, S. L., and Neretti, N. (2014). Dietary switch reveals fast coordinated gene expression changes in Drosophila melanogaster. Aging (Albany. NY). 6, 355–368. doi: 10.18632/aging.100662
Wigby, S., Sirot, L. K., Linklater, J. R., Buehner, N., Calboli, F. C. F., Bretman, A., et al. (2009). Seminal fluid protein allocation and male reproductive success. Curr. Biol. 19, 751–757. doi: 10.1016/j.cub.2009.03.036
Wigby, S., Slack, C., Grönke, S., Martinez, P., Calboli, F. C. F., Chapman, T., et al. (2011). Insulin signalling regulates remating in female Drosophila. Proc. R. Soc. B Biol. Sci. U.S.A. 278, 424–431. doi: 10.1098/rspb.2010.1390
Wu, Q., Zhang, Y., Xu, J., and Shen, P. (2005a). Regulation of hunger-driven behaviors by neural ribosomal S6 kinase in Drosophila. Proc. Natl. Acad. Sci. U.S.A. 102, 13289–13294. doi: 10.1073/pnas.0501914102
Wu, Q., Zhao, Z., and Shen, P. (2005b). Regulation of aversion to noxious food by Drosophila neuropeptide Y- and insulin-like systems. Nat. Neurosci. 8, 1350–1355. doi: 10.1038/nn1540
Yang, C., Belawat, P., Hafen, E., Jan, L. Y., and Jan, Y. (2008). Drosophila egg-laying site selection as a system to study simple decision-making processes. Science 319, 1679–1683. doi: 10.1126/science.1151842
Zars, T. (2000). Behavioral functions of the insect mushroom bodies. Curr. Opin. Neurobiol. 10, 790–795. doi: 10.1016/S0959-4388(00)00147-1
Keywords: mating behavior, feeding state, insulin, fruitless, mushroom bodies
Citation: Lebreton S, Carlsson MA and Witzgall P (2017) Insulin Signaling in the Peripheral and Central Nervous System Regulates Female Sexual Receptivity during Starvation in Drosophila. Front. Physiol. 8:685. doi: 10.3389/fphys.2017.00685
Received: 11 May 2017; Accepted: 25 August 2017;
Published: 08 September 2017.
Edited by:
Carolina E. Reisenman, University of California, Berkeley, United StatesReviewed by:
Nilay Yapici, Cornell University, United StatesToshiro Aigaki, Tokyo Metropolitan University, Japan
Meghan Laturney, University of California, Berkeley, United States
Copyright © 2017 Lebreton, Carlsson and Witzgall. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Sébastien Lebreton, c2ViYXN0aWVuLmxlYnJldG9uQHVuaXYtYW11LmZy
†Present Address: Sébastien Lebreton, Aix-Marseille Université, Centre National de la Recherche Scientifique, UMR7288, Institut de Biologie du Développement de Marseille, Marseille, France