 Myra N. Chávez
Myra N. Chávez Geraldine Aedo2
Geraldine Aedo2 Fernando A. Fierro
Fernando A. Fierro Miguel L. Allende
Miguel L. Allende José T. Egaña
José T. Egaña- 1Department of Plastic Surgery and Hand Surgery, University Hospital rechts der Isar, Technische Universität München, Munich, Germany
- 2Department of Biology, FONDAP Center for Genome Regulation, Faculty of Science, Universidad de Chile, Santiago, Chile
- 3Department of Biochemistry and Molecular Biology, FONDAP Advanced Center for Chronic Diseases (ACCDiS) and Center for Molecular Studies of the Cell (CEMC), Faculty of Chemical and Pharmaceutical Sciences, Faculty of Medicine, University of Chile, Santiago, Chile
- 4Department of Cell Biology and Human Anatomy, University of California, Davis, Sacramento, CA, USA
- 5Institute for Medical and Biological Engineering, Schools of Engineering, Biological Sciences and Medicine, Pontifícia Universidad Católica de Chile, Santiago, Chile
Angiogenesis is the process through which new blood vessels are formed from preexisting ones and plays a critical role in several conditions including embryonic development, tissue repair and disease. Moreover, enhanced therapeutic angiogenesis is a major goal in the field of regenerative medicine and efficient vascularization of artificial tissues and organs is one of the main hindrances in the implementation of tissue engineering approaches, while, on the other hand, inhibition of angiogenesis is a key therapeutic target to inhibit for instance tumor growth. During the last decades, the understanding of cellular and molecular mechanisms involved in this process has been matter of intense research. In this regard, several in vitro and in vivo models have been established to visualize and study migration of endothelial progenitor cells, formation of endothelial tubules and the generation of new vascular networks, while assessing the conditions and treatments that either promote or inhibit such processes. In this review, we address and compare the most commonly used experimental models to study angiogenesis in vitro and in vivo. In particular, we focus on the implementation of the zebrafish (Danio rerio) as a model to study angiogenesis and discuss the advantages and not yet explored possibilities of its use as model organism.
Introduction
Angiogenesis is the process through which new blood vessels emanate from preexisting vascular structures. It plays a pivotal role in various physiological and pathological conditions and is orchestrated by the tight interaction between endothelial cells and their niche. While inadequate vessel maintenance or growth leads to tissue ischemia; excessive vascular growth or abnormal remodeling promotes cancer, inflammatory disorders, and retinopathies (Pandya et al., 2006).
Angiogenesis is mainly accomplished through vessel sprouting, which may be divided into four main steps: tip cell formation, tubule morphogenesis and lumen creation, adaptation to tissue needs and, finally, stabilization and maturation of the newly formed vessels (Ribatti and Crivellato, 2012; Neufeld et al., 2014). A non-sprouting mechanism of microvascular growth has been also described, and it involves the increment of vascular surface by insertion of a multitude of transcapillary pillars in a process called “intussusception” (Styp-Rekowska et al., 2011).
Parallel to the study of the angiogenic process, a large number of in vitro and in vivo assays have been developed to study the cellular and molecular mechanisms involved (Cimpean et al., 2011). Each model has its own advantages and disadvantages, and their adequate combination is key to reveal the impact of the element under analysis within the global process.
In vitro assays have been broadly used to answer questions related to specific behaviors of endothelial cells such as proliferation, differentiation, structural organization, cytokine secretion profiling and chemotaxis, as well as the molecular mechanisms associated with angiogenesis (Irvin et al., 2014). Moreover, in vitro systems have helped to identify and validate promising compounds to therapeutically promote or inhibit angiogenesis (Goodwin, 2007), as they are quantitative, easily monitored, reproducible, and provide the confidence necessary for the rapid screening of potential pro- or anti-angiogenic compounds (Weiss et al., 2015). However, important aspects should be considered when assessing the potential of an angiogenic effector using in vitro assays such as the decision over the type or tissue-origin of the endothelial cells being used, and the experimental bias of the protocols being followed (for a more comprehensive discussion see Unger et al., 2002; Staton et al., 2009). Finally, common in vitro experiments do not consider the influence of the vascular niche, which has been shown to be critical in the process of angiogenesis during tissue regeneration (Ribatti and Crivellato, 2012; Kunisaki and Frenette, 2014; Ramasamy et al., 2015).
The complexities of the formation, function and pathology of blood vessels in the context of the living animal mandate the availability of adequate in vivo models in order to confirm the results obtained in vitro. Since the 1970s multiple animal models have been developed in order to understand the physiological mechanisms of blood vessel formation, as well as to validate approaches that either enhance or inhibit the angiogenic process. The mouse model is by far the most common used to study angiogenesis in vivo, with the advantage of being a mammal that in many ways faithfully recapitulates human physiology. However, this animal model can be laborious and expensive to use, especially for screening purposes. Also, the use of mice limits the evaluation of the outcome to a final time point, since de novo or re-vascularization can only be visualized and quantified after euthanizing the animal, hence limiting the understanding of angiogenic dynamics.
As mammalian and most vertebrate tissues are opaque, the introduction of the transparent zebrafish larva as a tool for the examination of the vasculature in the intact animal has gained recent attention. Importantly, several studies have made clear that there is a high degree of molecular conservation in the most important pathways involved in the development and physiology of blood vessels in all vertebrates (reviewed by Baldessari and Mione, 2008; Gore et al., 2012). Furthermore, genetic and pharmacological evidence has shown that there is mutual translatability of findings between zebrafish and human vascular biology (Coultas et al., 2006; Lieschke and Currie, 2007). Thus, the emergence of a simple yet validated discovery and/or screening tool has been welcomed by the community.
In the following sections, we provide a brief overview on the currently available in vitro and in vivo angiogenic assays, describing their most common uses and their potential advantages and limitations. Additionally, we also provide information on the current and potential uses of zebrafish as model to study angiogenesis.
In vitro Models
In vitro angiogenesis models study the behavior of endothelial cells within a controlled environment (Ayata et al., 2015). They are designed to recapitulate the different steps of the angiogenic processes, where endothelial cells are involved, such as cell proliferation, migration, extracellular matrix digestion and invasion, morphogenesis and capillary tube formation (Cimpean et al., 2011). Table 1 summarizes the settings and evaluation parameters of the most commonly used assays focusing on migration, proliferation and tubule formation.
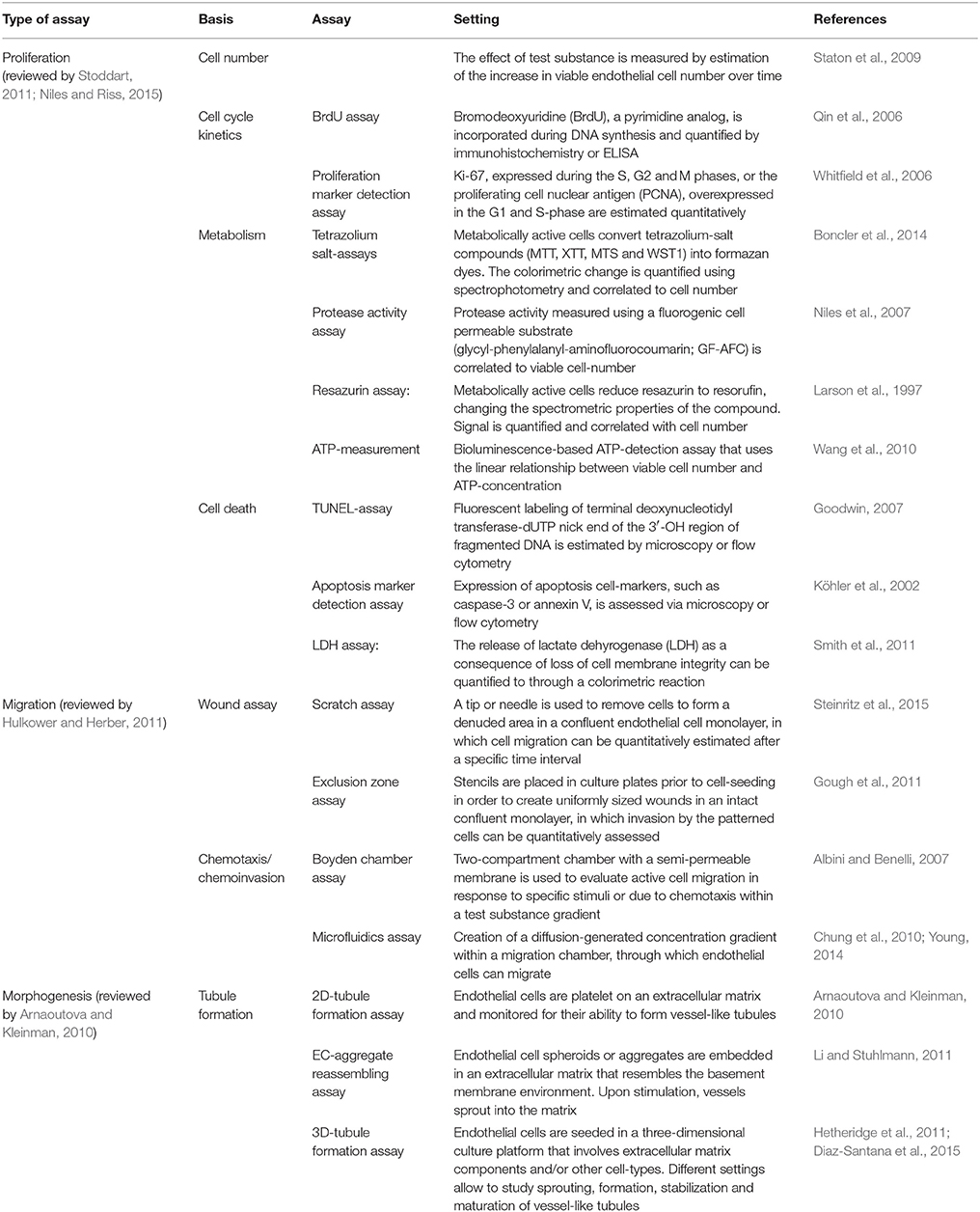
Table 1. In vitro angiogenesis assays.
Proliferation Assays
These assays are conceived to evaluate the effects of a test substance, based on the quantitation of endothelial cell proliferation. They are broadly classified into those that determine net cell number and those that evaluate cell-cycle kinetics (Staton et al., 2004). Cell numbers can be estimated either manually or through automated cell counting. Alternatively, metabolic assays, which have shown a linear correlation with cell density (Niles and Riss, 2015), quantification of DNA synthesis or expression of proliferation markers may be used (reviewed by Whitfield et al., 2006). However, since none of these methods have been explicitly developed for vascular-related cells, it is indispensable to address the target specificity of the test substance, as well as its therapeutic impact based on other angiogenesis-related parameters. Furthermore, proliferation assays should be combined with quantitative methods for estimation of cell death, in order to discard the possibility of cytotoxicity of the test-substance (Kepp et al., 2011).
Migration Assays
Migration assays allow the study of endothelial cell motility and chemotaxis. They evaluate the active migration of cells into a specific area or toward a specific direction as a result of a treatment. The main advantage of the exclusion zone assay (Poujade et al., 2007; Gough et al., 2011), where silicone-based structures, so-called “masks” or “stencils,” are placed on the well bottom to create an cell-empty area, in comparison to a scratch assay (Coomber and Gotlieb, 1990; Yarrow et al., 2004), where a “wound” is created by physically disrupting an endothelial cell monolayer, is the uniformity and hence reproducibility of the denuded area into which confluent endothelial cells will later migrate (reviewed by Hulkower and Herber, 2011).
Another commonly used migration assay follows the principle of the Boyden chamber, first described in 1962, where a semipermeable membrane that only allows active passage of cells is placed in their migration path (Boyden, 1962), sometimes requiring matrix degradation, in which case it is regarded a chemoinvasion assay (Albini and Benelli, 2007), or in response to a test substance (chemotaxis). The use of microfluidic cell culture systems has overcome the difficulties of maintaining a linear gradient of the test compound by introducing microchannel compartments in which a diffusion-generated concentration gradient can be created. Furthermore, they allow live single-cell and cell-population tracking, as well as directionality and velocity estimation (Young, 2014).
Tubule Formation Assays
Tubule formation assays are used to study the assembly of capillary-like structures by adjacent endothelial cells (Arnaoutova et al., 2009). In two-dimensional assays, endothelial cells are usually seeded on extracellular matrices and the spontaneous building of capillary-like networks is analyzed. Quantitation of tubule formation is mainly addressed by immunohistochemistry and analyzed based on four main parameters: average tubule length, number of tubules, tubule area and number of branch points (Staton et al., 2009). They allow to study spontaneous tubule formation due to endothelial cell-to-cell interactions and the assembly of tight-junctions (Vailhé et al., 2001), however they do not resemble the process of sprouting angiogenesis, which is the development of new blood vessels from pre-existing major donor vessels (Ribatti and Crivellato, 2012). Also, the early formed tubules lack lumen and their length and degree of branching differ from real capillaries (Donovan et al., 2001).
On the other hand, three-dimensional culture systems of endothelial cells have been used to study the formation of more complex capillary networks inside extracellular matrix substitutes. They have helped to elucidate the role of support cells, such as fibroblasts (Bishop et al., 1999; Hetheridge et al., 2011), pericytes (Berthod et al., 2012) and adipose stromal cells (Merfeld-Clauss et al., 2010; Verseijden et al., 2010; Sarkanen et al., 2012), as well as the homo- and heterotypic cell-interactions of endothelial cells during vessel-formation, -sprouting and -anastomosis (Ayata et al., 2015; Diaz-Santana et al., 2015). Moreover, three-dimensional tubule formation assays have become an important tool to mimic in vitro microenvironments of tumor vascularization (reviewed by Chwalek et al., 2014; Song et al., 2014). Low standardized settings and the more challenging evaluation of the three-dimensional tubule formation are the main disadvantages of these assays.
Organ Explant Based-Assays
Also known as ex vivo angiogenesis models, these assays aim to analyze the angiogenic sprouting and the growth of vessel capillaries from explanted segments of vasculature. Here, isolated vasculature biopsies are placed generally over three dimensional biological matrices in the presence or absence of a test compound. Explants are then monitored for the outgrowth of vessel tubules extending from the periphery of the explant into the surrounding matrix (Rezzola et al., 2014). Table 2 summarizes the characteristics of the most broadly used ex vivo assays.
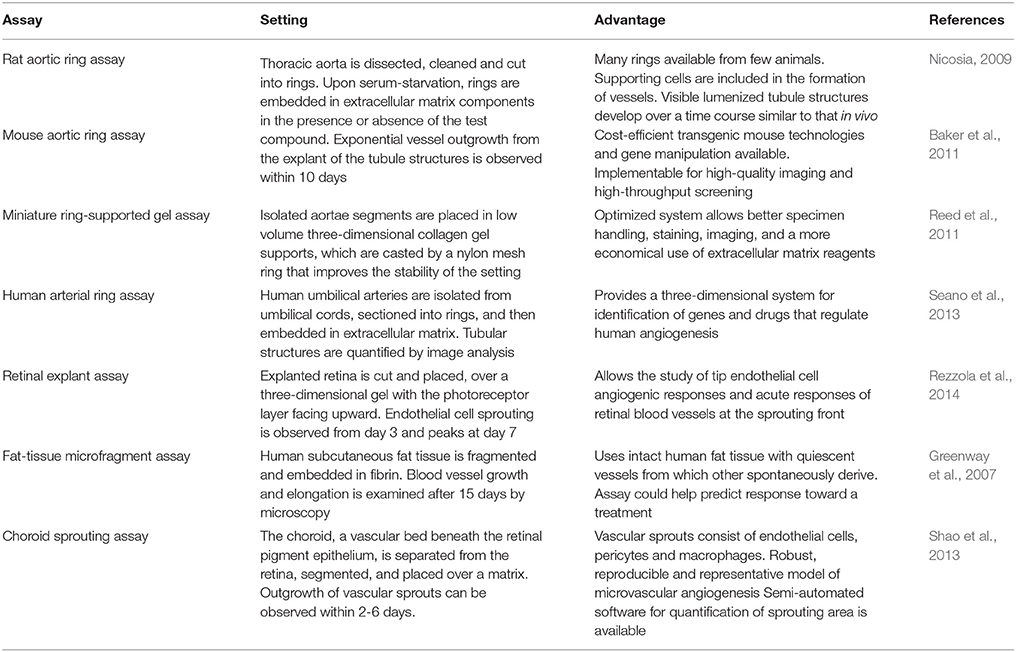
Table 2. Ex vivo angiogenesis assays.
Ex vivo assays have the advantage of working with native quiescent endothelial cells in vivo at the experimental outset (Ucuzian and Greisler, 2007; Staton et al., 2009). Further, because the tissue complexity is preserved, most of the cellular and molecular components involved in angiogenesis are present. As a result, vascular sprouts contain a lumen and a basement membrane, and are composed of a mixed population of endothelial cells, pericytes, fibroblasts, and macrophages (Nicosia et al., 2011). These assays allow the study cell proliferation, migration, tube formation, network branching, perivascular recruitment and vascular remodeling (Baker et al., 2011), in addition to other post-angiogenic mechanisms such as vessel stabilization and regression (Nicosia et al., 2011). Some of the disadvantages compared to in vitro assays are the more demanding technical skills, the limited number of simultaneous samples being processed, and the implicit higher experimental variability (Staton et al., 2009; Rezzola et al., 2014). On the other hand, compared to in vivo assays, ex vivo assays do not consider circulating endothelial progenitors recruited in the angiogenic process and lack the pro-angiogenic stimuli in blood flow (Irvin et al., 2014). Also, the decision over the source of the vascular material should behold that angiogenesis mainly involves the microvasculature rather than the macrovasculature, and that microvessels such as capillaries, small arterioles and venules, are composed of different tissue layers compared to large arteries and veins (Staton et al., 2009).
In vivo Models
Multiple in vivo models have been developed to directly study angiogenesis within an organism, and therefore evaluate the entire process of new blood vessel formation, since they allow to consider all cellular and molecular role players involved, such as supporting cells (e.g., tumor cells, pericytes, smooth muscle cells, and fibroblasts), the extracellular matrix, and the cellular and humoral components in circulating blood (Staton et al., 2004). Most in vivo angiogenesis assays are not designed to understand a specific process, but rather to determine the success of the outcome, with the exception being the zebrafish larva, as we discuss in the next section. Nevertheless, it is also important to point out that one of the main disadvantages of in vivo models is the ethical concerns they raise, and the complications they imply, due to the strict guidelines regulating animal testing in some countries. Again, the zebrafish is exempt, for the most part, from these concerns, especially during larval stages.
Corneal Angiogenesis Assays
As originally developed by Gimbrone et al. (1973), induction of angiogenesis in the cornea is among the most convincing demonstrations of neovascularization, since the cornea is richly innervated, but normally has no blood vessels (Henkind, 1978). In this assay, a stimulus induces the migration of endothelial cells from the edge of the cornea into the space between the corneal epithelium and stromal cells, forming new sprouts directed toward the source of the angiogenic signal. This method has been applied in multiple animal models including rabbit, mouse, rat and guinea pig (Ziche and Morbidelli, 2015). It has been further developed to become quantitative, by incorporation of a contrast-dye such as high molecular weight dextran and imaging analysis. Disadvantages are that it is rather expensive, and that the angiogenic process is rather atypical, since it occurs in a non-vascular environment (Norrby, 2006).
Chorioallantoic Membrane (CAM) Assay
The CAM assay allows the measurement of both inhibition and stimulation of angiogenesis over the vascularized chorioallantoic membrane of a chick embryo, which can develop normally after carefully opening the egg shell to create a window (in ovo), or being placed in a cup outside of the egg shell (ex ovo or in vitro), in order to get access to the CAM. From days 3.5 to 10 after fertilization, highly proliferative and immature endothelial cells rapidly grow a sprouting vascular network, which is then replaced by intussusceptive microvasculature (Ribatti et al., 2001). During early phases, the CAM assay is most suitable to study angiogenic inhibitors. In contrast, the study of pro-angiogenic factors is best accomplished from day 6 to 8, when the rapid embryonic angiogenic development has slowed down. Quantification of angiogenesis is typically based on the directionality of the blood vessels toward the graft/angiogenic stimuli, the number of sprouts, and/or the size/length of the stimulated blood vessels. The CAM assay allows repeated visualizations of the angiogenic process, and it is fast and cost effective, making it suitable for large scale screens. Its major disadvantages are the rather challenging quantification of the outcome, since it is often difficult to distinguish normal angiogenesis from the induced one, and the false positive effects that often occur from inadvertently damaging the CAM (Ribatti et al., 2001; Ribatti, 2008).
Matrigel Plug Assay
Subcutaneous injection of matrigel in mice is a common method to study angiogenesis in vivo in mammals. Matrigel is an extract of the Engelbreth–Holm–Swarm tumor, mostly composed by extracellular matrix proteins and growth factors (Benton et al., 2014). When cold, matrigel is liquid, but becomes solid at body temperature. This property makes simple the injection of matrigel in the midventral abdominal region of mice, where it quickly solidifies forming a “plug” (Akhtar et al., 2002). The injected matrigel can be supplemented with either angiogenic inhibitors or inducers. Then, usually about 2 weeks after injection, infiltration of new blood vessels is determined histologically. A major advantage of this method, is the simplicity to implement it. However, visualization and quantification of differences can be challenging and are mainly based in the histological analysis of explanted plugs at a final experimental point.
Hind Limb Ischemia
A common system to study angiogenesis in vivo from a therapeutic perspective, is the hind limb ischemia (HLI) model (Limbourg et al., 2009). In this case, the femoral artery of mice is ligated causing a strong obstruction of blood flow toward the hind limb. Since originally described (Couffinhal et al., 1998), the HLI protocol has been applied with multiple variations. A common surgical approach is the ligation of the femoral artery at distal and proximal sites, and removal of the intervening arterial fragment (Fierro et al., 2011). Another approach is a single ligation, without arterial excision, where the severity of ischemia depends on the specific site of ligation. Also a gradual arterial occlusion model has been established, by placing ameroid constrictors on the femoral artery (Yang et al., 2008). In all cases, the contralateral hind limb is left intact, as a control. Mice are usually able to recover from this injury naturally, restoring blood flow within approximately 4 weeks, by mechanisms including the formation or enlargement of collateral blood vessels (Sondergaard et al., 2010). Laser scanning Doppler imaging is the best suited method to monitor blood flow restoration upon HLI induction, because it is non-invasive, and can be performed in the same animal at multiple time points. At the end of the experiment, animals can be euthanized for further investigation including histology and gene expression analysis. A negative aspect of scanning Doppler imaging is the sensitivity of the method, since only robust differences can be noticed. Another limitation of this method is that it fails to reveal the exact mechanism underlying the blood flow restoration (e.g., angiogenesis vs. vasculogenesis).
Vascularization during Dermal Wound Repair
Our group has developed a full skin defect model that presents several advantages compared to the in vivo models presented above, which are intrinsic to the nature of skin. Among others: transparency, large surface, easy manipulation, external location and tissue homogeneity (Egaña et al., 2008). In this model, full skin defects are surgically created bilaterally on the back of mice, and the skin excision is replaced by biodegradable scaffolds, which can be modified to contain a specific angiogenic stimuli. Typically, after two weeks animals are euthanized, and tissue vascularization is quantified as follows: the skin, including the implanted scaffold, is removed and quickly placed over a light source. During trans-illumination, a digital picture is taken, and is later analyzed by digital segmentation (Schenck et al., 2014). This method does not affect cell integrity post mortem, allowing further analysis such as histology or protein/RNA extraction.
The Skinfold Chamber and Ear Assays
Four major types of in vivo models have been developed to observe the angiogenic process in two dimensions: the rat mesentery window assay (Norrby, 2011), the hamster cheek pouch assay (Monti-Hughes et al., 2015), the dorsal skinfold chamber adapted to mice, hamsters and rats (Lehr et al., 1993; Harder et al., 2014; Irvin et al., 2014), and the rabbit ear chamber assay (Clark et al., 1931; Ichioka et al., 1997). These techniques, developed as early as in the 1940s, rely on semi-transparent tissue or the implantation of a transparent chamber that allows an easy and direct visualization and quantification of the angiogenic process, including blood vessel density and blood flow velocity. In particular, the implementation of intravital microscopy along with epifluorescence, confocal and multiphoton techniques, offers the possibility of repetitive, direct, and quantitative measurements of several microcirculatory parameters, as well as microvasculature imaging at an unparalleled subcellular-resolution (Taqueti and Jaffer, 2013). However, these methods are invasive, and may cause great discomfort to animals. In addition, some methods such as the implantation of a dorsal window chamber in mice, are cumbersome (Palmer et al., 2011) and therefore difficult to implement in a number of animals sufficient for adequate technical replicates.
Zebrafish as a Model for Angiogenesis Research
While the models described above have provided essential information and platforms for discovery of therapeutic targets and drugs, many questions about the biology of vascular cells and how they build the circulatory system remain unresolved. Above all, the relevance of the models is often hindered by the inaccessibility of the tissue in live animals, and much of what we know has been derived from fixed material or indirect assays. Zebrafish provides a series of advantages as a model of study due to its rapid development, optical transparency, high number of offspring and straightforward strategies for forward and reverse genetic manipulation. Furthermore, the early development of a cardiovascular system in the transparent zebrafish embryo and larva translates into a unique opportunity for direct observation of blood flow and the development of the system's related organs in both wild type and transgenic fish, without the need for complex instrumentation. Lastly, genetic studies have revealed conservation of the molecular pathways between fish and mammals making research in vascular biology in teleosts directly translatable into potentially relevant information for human health.
As the restrictions on the experimental use of mammalian models for research increase, the zebrafish emerges as a convenient alternative. Larvae can be used in massive numbers in genetic or pharmacological screens, at stages in which they lack the legal status of a “vertebrate animal” yet have all of the physiological functions of the adult, including a hematovascular system. Circulation begins 24 h after fertilization, with a simple, yet functional blood circuit. The embryos and larvae, can be kept for the first five days of life in small wells in microtiter plates, in only a few hundred microliters of water. This is the pharmacologists dream since as many replicates of the experiment as one desires can be done and dilutions of each drug can be tested ad libitum.
Two decades ago, the generation of the first stable transgenic zebrafish line was reported. Since then, hundreds of transgenic lines have been developed both for expression of reporter proteins or for expression of diverse proteins for functional studies (Udvadia and Linney, 2003). At the same time, efficient mutagenesis protocols have allowed forward-genetic screening in the context of angiogenesis, generating valuable collections of mutants (Jin et al., 2007). Traditionally, gene function in zebrafish has been assessed using chemically or insertionally induced mutants that required large scale unbiased screens to identify phenotypes related with the process or organ of interest (Gaiano et al., 1996; Haffter et al., 1996). While common antisense technologies were not generally applicable to the zebrafish, the advent of oligonucleotide substitutes named morpholinos, enabled the knockdown of endogenous genes by either blocking translation of the mRNA or splicing of the pre-mRNA (Nasevicius et al., 2000). The ease of this technology spurred its widespread use, even though it presented some limitations such as the induction of undesired off-target effects or the progressive loss of the effect at late developmental stages because of diminishing activity over time. The zebrafish toolkit has been recently enriched with the introduction of gene editing technologies such as TALENs (Transcription activator-like effector nucleases, Bedell et al., 2012), and CRISPR (Clustered regularly-interspaced short palindromic repeats)-Cas based strategies (Hwang et al., 2013). As long as genomic sequence is available for the targeted locus, any gene can be mutated efficiently and permanently in the germ line; the efficiency is often high enough such that recessive phenotypes can be seen already in the injected animals. Further, the CRISPR-Cas9 system has been adapted for high throughput mutagenesis in zebrafish so that dozens of genes can be mutated in a single experiment (Varshney et al., 2015). Recently, phenotypic inconsistencies between genomic mutations induced by CRSPR-Cas9 and knockdown via morpholinos have emerged (Kok et al., 2015). It is likely that these two gene loss-of-function strategies differ in their penetrance given that genetic lesions might induce compensatory reactions in the genome obscuring the gene's function. Many authors believe that a combination of strategies is desirable when analyzing a particular gene and that it is unwise (as has been agreed by communities using other model organisms) to rely only on a gene knockdown phenotype to assign gene function (Lawson, 2016).
Despite its success and popularity, those working with the zebrafish model must consider complementing their studies with mammalian systems, if they wish to validate the knowledge gained for potential clinical applications. Gene and protein functional conservation is high, but not absolute, and obviously there are important physiological differences to be dealt with. Aquatic and terrestrial life pose unique challenges that impact on many organs, most notably the respiratory system and, thus, cardiovascular architecture. In fish, only the embryo and larva are transparent, making studies in adults just as difficult as in mammals. The small size of embryos makes some observations challenging (i.e., requiring sophisticated microscopy and imaging) and they are also developing systems, which means they are constantly in a state of change and growth. Thus, the zebrafish, with all of its attributes, should be considered a starting point for discovery and a model that can offer new hypotheses to be tested further in other models.
Vascular Development in Zebrafish
Transgenic technology has enhanced the inherent in vivo imaging capabilities that zebrafish larvae may offer to the investigator. Though vessels and blood flow can easily be visualized with a simple dissecting scope, it was with the introduction of tissue specific expression of fluorescent proteins that vascular and blood development could be examined in great detail. Confocal microscopy and time lapse imaging can both be carried out with live specimens which allows detailed morphogenetic movements and cell shape changes to be followed directly. Thus, vascular development has been described in great detail, both from the anatomical and cellular point of view and with a comprehensive examination of the molecular players involved (reviewed by Gore et al., 2012; Schuermann et al., 2014).
Most of the strategies which have been followed to create stable transgenic lines with vascular-specific phenotypes are based on gene-specific promoters. Both autologous and heterologous promoters have been shown to work. Table 3 lists some of the transgenic lines, which have been designed and developed for the visualization and analysis of the vascular system. Before a complete and reliable zebrafish genome sequence was available, the promoter of a related gene from another species, most commonly a mammalian one (Baldessari and Mione, 2008), was used. However, the reporter protein expression in zebrafish did not always exactly recapitulate that of the orthologous one, because of the differences in promoter elements among species. For example, the zebrafish Tg(tie2:GFP)s849 line encoding the promoter for the murine tie2-gene (a vascular-specific tyrosine kinase receptor activated by angiopoietin ligands), successfully drove GFP expression in endothelial cells, but also showed substantial nonvascular expression in the hindbrain and the posterior neural tube, and the overall level of expression was proportionally lower compared to that in mice (Motoike et al., 2000). On the other hand, the fli1a and scl zebrafish genes, have been used as early markers of vascular and hematopoietic lateral mesoderm. While the expression of fli1a is restricted to endothelial cells, a subset of early circulating myeloid cells, and cranial neural crest derivatives (Brown et al., 2000), the expression of scl is specific for the hematopoietic lineage at later stages (Gering et al., 1998).
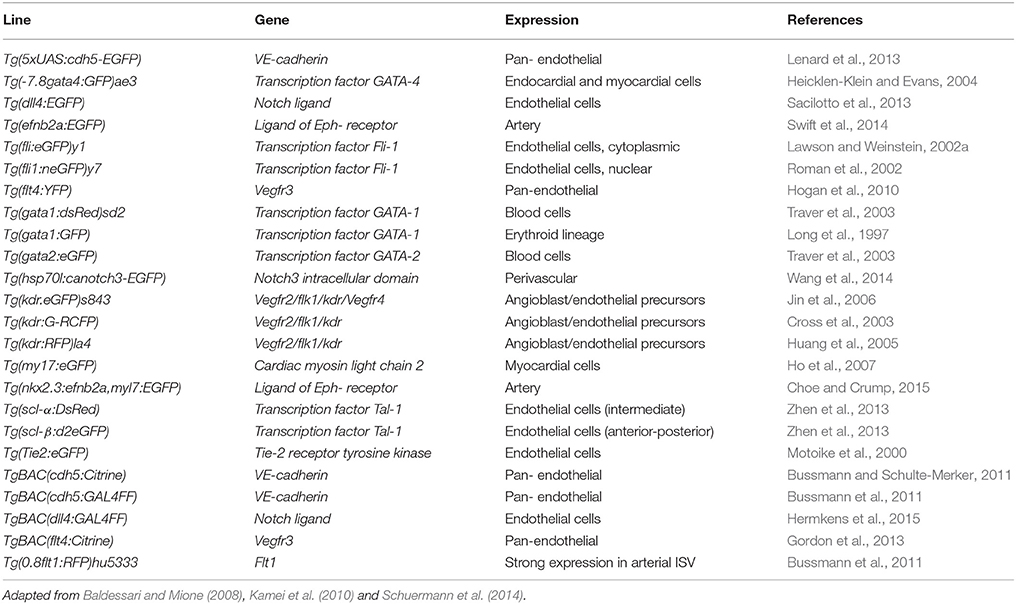
Table 3. Transgenic zebrafish lines generated for the study and visualization of the vascular system.
The development of the vascular anatomy of the zebrafish has been extensively described and has been proven to share high similarity with other vertebrates (Isogai et al., 2001; Ellertsdóttir et al., 2010; Gore et al., 2012). Many of the studies on vascular development have been achieved by using molecular tracers during the early embryonic stages of zebrafish. One of such strategies is the injection of fluorescent microspheres, and their detection after lumenization and anastomosis of the vascular network is complete (Küchler et al., 2006). This strategy has also been used to compare the development of blood and lymphatic vasculature in zebrafish (Coffindaffer-Wilson et al., 2011). Transgenic zebrafish lines have been also employed to track individual cell growth during vascular development. Using fluorescent endothelial cell markers, it is possible to observe the proliferative and migratory behaviors of single cells, and different kinds of cell types during the embryo-to-larva transition. Combining transgenic lines expressing different fluorescent proteins, it was possible to observe two cell types simultaneously. For instance, it was possible to track both endothelial progenitors and erythrocytes while following the vascular network development and the initiation of blood circulation (Lawson and Weinstein, 2002a,b; Herwig et al., 2011; Kimura et al., 2013). Moreover, combining nuclear and cell membrane specific fluorescent tags has allowed the examination of single cell morphological dynamics in living larvae during vessel formation (Yu et al., 2015). Finally, the development of stable transgenic zebrafish lines has been a valuable resource for tissue specific gene expression as well as inducible gene expression (Udvadia and Linney, 2003). The implementation of these strategies enabled the study of the sequence of events involved in the establishment of the first circulatory loop in zebrafish embryos, which consists in the connection between the heart with the dorsal aorta and the cardinal posterior vein back to the heart. Other blood vessels, which are characteristic and highly accessible in the zebrafish embryos and larvae are the intersegmental vessels, which emerge from the dorsal aorta into the embryonic trunk and tail, and later grow into the anastomosing dorsal longitudinal vessels (Strilić et al., 2009).
A remarkable feature of zebrafish compared to other vertebrates, is that they rely on passive oxygen diffusion during the early embryonic stages rather than oxygen perfusion, as the completion of the vascular development takes place after hatching. Moreover, the generation and characterization of zebrafish mutants has shown that embryos are able to sustain normal development even in absence of a functional vascular system or in the absence of blood (Stainier et al., 1995; Isogai et al., 2003). This attribute has made the analysis of late phenotypes related to circulatory system malformations possible, whereas they are lethal and hence impossible to study in living mammals (reviewed by Isogai et al., 2001; Wilkinson and van Eeden, 2014). A prime example of the power of the genetic approach was the study of the zebrafish gridlock mutant (Peterson et al., 2004). The gridlock mutation causes a syndrome similar to human aortic coarctation disrupting blood flow in the aorta. Further, mutant animals were used to design a small molecule screen that would detect reversal of the phenotype upon treatment and several compounds were found to have such an effect. Table 4 summarizes some of the most remarkable vascular zebrafish mutant lines described thus far.
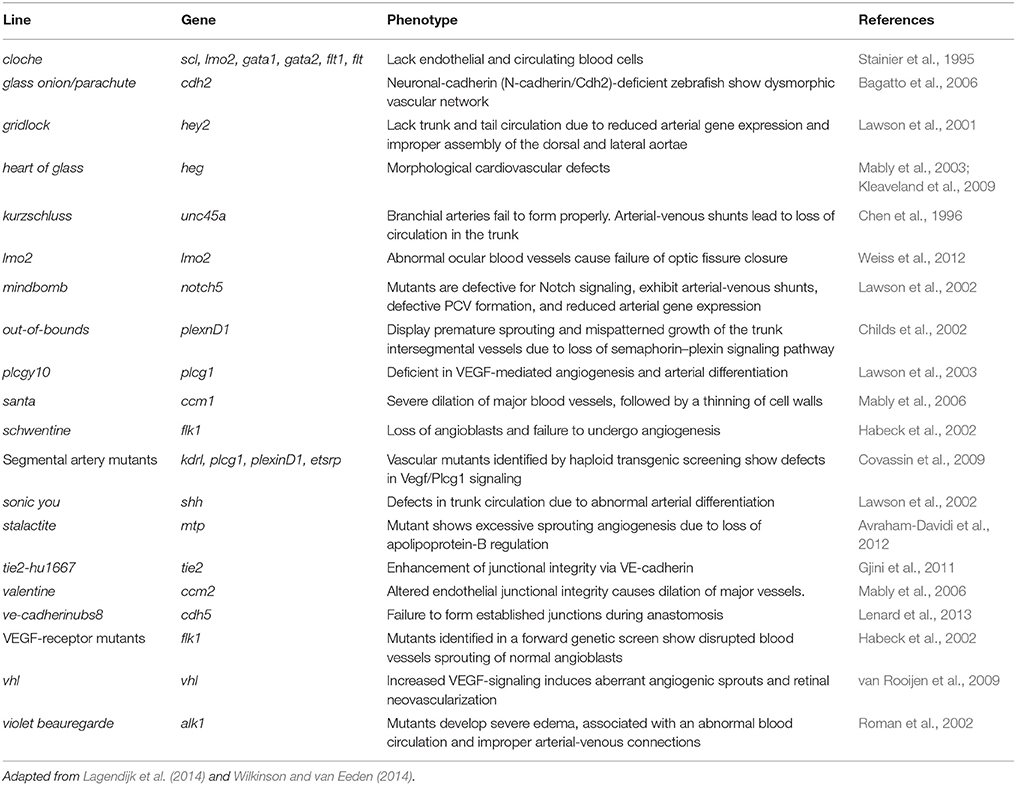
Table 4. Zebrafish vascular mutants.
Finally, experimental analysis of blood vessels during zebrafish development has also relied on common techniques for visualizing gene and protein expression. In order to observe the expression of endogenous genes in zebrafish embryos and larvae, two methods are available: in situ hybridization and immunohistochemistry. While neither of these methods was specifically developed for the zebrafish vasculature studies, an increasing number of tools and protocols are becoming available that facilitate these strategies (Kamei et al., 2010; Thisse and Thisse, 2014).
Vascular Regeneration
The zebrafish is a broadly known model for studies on tissue regeneration. In this regard, its capacity to regenerate its organs and limbs is remarkable even in adult stages. The caudal fin, in particular, provides an ideal tissue for studies related to vascular regeneration in adult zebrafish due to its simple thin architecture and relative transparency (Poss et al., 2003). While caudal fin regeneration in zebrafish larvae takes a few days, it has been demonstrated that the adult caudal fin is capable of full regeneration after successive amputations within a couple of weeks (Azevedo et al., 2011). The caudal fin amputation model has been extensively used to study the orchestration of the mechanisms involved in regeneration, such as cell differentiation, migration and patterning, which lead to the restoration of the fin's original morphology and functionality (Pfefferli and Jaźwińska, 2015). In a landmark study, Xu et al. (2014) showed that regenerating vessels in the regenerating tail fin originate from vein-derived cells that acquire angiogenic potential. These cells migrate singly or collectively and organize into vessel in response to chemokine signaling (reviewed by Hasan and Siekmann, 2015). However, the applicability of this model to the study of vascular regeneration could be much more widely exploited. For instance, the ablation of single vessels or vessel interruption has not been addressed in the zebrafish. A new technique called electroablation (Moya-Díaz et al., 2014) has been shown to be useful for inducing small tissue lesions including blood vessel ablation in the adult tail fin.
The zebrafish larval vascular network has been subject of numerous screens over the past decade. Key to this effort was the development of the Tg(fli1:EGFP)y1 transgenic line (Lawson and Weinstein, 2002a), that fluorescently labels endothelial cells throughout life (Figure 1) and enables the visualization of the microvasculature in this tissue. However, most screens to date have used the larval vasculature to find molecules that disrupt (positively or negatively) the normal pattern of blood vessels. Only a few screens have examined the role of the vasculature on tissue regeneration, even though these transgenic fish could be a remarkable tool to allow the study of the effects of test substances and genetic interference on vessel growth and restoration. As an example, Bayliss et al. examined the requirement for blood vessels in caudal fin regeneration using adult fish (Bayliss et al., 2006). In this work, the authors conclude that up to ~1 mm avascular caudal fin tissue can be regenerated, though, for regeneration of the full limb, angiogenesis is required. Further, they showed that the model can be implemented for antiangiogenic drug screening, as it is possible to selectively inhibit highly active, abnormal vessels while leaving quiescent vessels intact.
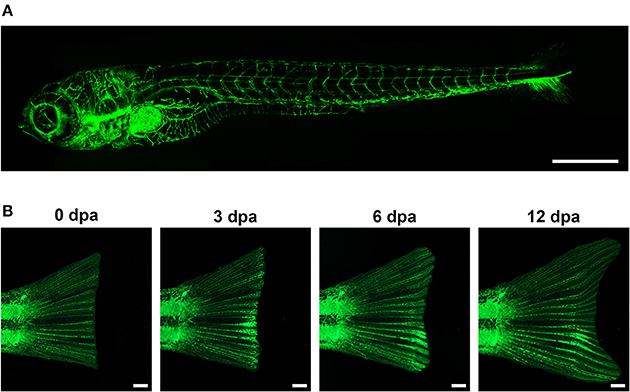
Figure 1. In the transgenic zebrafish line Tg(fli1:EGFP)y1, the promoter for the endothelial marker fli1 drives the specific expression of EGFP in blood vessels. This allows the visualization, and hence the analysis of the vasculature during zebrafish embryonic development (A, Lawson and Weinstein, 2002a), and during adult vessel regeneration upon tail fin amputation (B, Huang et al., 2003). Scale bar represents 500 μm in (A), and 1 mm in (B).
Since angiogenesis is one of the main focuses of vascular regeneration research, models for this type of vascular development and growth have been developed. In the embryo, the intersegmental vessels form by angiogenic sprouting from the dorsal aorta and have been the target of studies using drugs or genetic perturbations (Schuermann et al., 2014). Further, since it has been shown that mammalian malignant cells can be xenotransplanted into zebrafish embryos and that they can form tumors (Haldi et al., 2006), models for tumor angiogenesis have been developed (Tobia et al., 2011). We have also shown recently (Chávez et al., 2016), that angiogenic sprouting can also be induced by xenotransplantation of cells expressing the recombinant vascular endothelial growth factor (VEGF), in this case plant cells. Thus, the factors governing angiogenic growth and inhibition are amenable to be examined in vivo in these contexts.
High Throughput Screens
As previously mentioned, zebrafish larvae are optically transparent until 5 days after fertilization allowing direct observation of internal tissues. This feature, coupled with the use of transgenic zebrafish lines with fluorescently labeled organs and cells, has allowed for straightforward assays to be developed to assess either positive or negative effects of chemicals or genetic perturbations on vascular integrity (Raghunath et al., 2009; Taylor et al., 2010). For instance, by using transgenic lines in a genetic screen, numerous vascular-specific mutations were identified (Covassin et al., 2009), while a chemical screen has revealed compounds that restored a normal phenotype in mutant fish (Hill et al., 2005; Asnani and Peterson, 2014).
How relevant are drug screens carried out in fish to human biology? As most human genes have a fish ortholog and sequence conservation is high, most teleost proteins targeted by drugs will predict an effect on its human counterpart (Tran et al., 2007). The relevance of this type of approach is highlighted by the fact that several small molecules identified in zebrafish are currently in clinical trial phase (MacRae and Peterson, 2015). Furthermore, these assays can be scaled into high throughput screens due to the fact that the zebrafish larvae, 2–3 mm at 3 days post-fertilization, can be arrayed into microwell plates and examined manually or automatedly by the thousands. Large chemical libraries can be screened for direct effects on the tissue of interest as compounds readily permeate the animal, and minimal amounts of each compound are required (drugs are supplied diluted in only a few ml in aqueous solution). The readout can be exceedingly simple: usually a perturbation of the normal or expected anatomical structure or cellular behavior is sought. While it is possible to visually screen hundreds of fish for a phenotype as it has been classically done (i.e., double blind scoring), there are automated and semi-automated systems for image acquisition and analysis as well as software that can quantitatively detect subtle effects (Pardo-Martin et al., 2010; Tamplin and Zon, 2010).
Conclusions and Perspectives
Since the 1990s experimentation on animals has increasingly emphasized the “three Rs”: reduction (minimize the number of animals), refinement (maximize the amount of data obtained) and replacement, (substitute with in vitro studies, when possible; Mayer et al., 1994). Here, we have enumerated a series of alternative models for the study of vascular development and regeneration. In vitro studies are accessible and offer controlled conditions for manipulation, but they lack the complexity found in living tissues. As mammalian models present the closest substitutes for humans, they should be preferred as the final validation step when proposing a therapy. However, these organisms can only be used in small numbers due to the cost, cumbersomeness of the experimental designs and ethical concerns. We describe the zebrafish model as an attractive alternative because it combines the relevance of in vivo assays with the simplicity and versatility of in vitro assays. In larvae, access to the developing vasculature is straightforward thanks to fluorophore-tagged strains and the small size of the animals makes the use of high-throughput strategies possible. In adults, the tailfin is equally convenient as a model tissue as regenerating vessels are directly observable at all stages and the animals are suitable for experimental manipulation with compounds, for instance. The advent of new genome modification techniques opens up even more tools for the vascular biologist as new therapeutic targets can be identified through mutational analysis.
Author Contributions
MC, GA, FF, MA, and TE all contributed to the conception of this manuscript, as well as to the acquisition and critical analysis of the intellectual content reviewed in this work. MC and GA were responsible for generating the images shown in Figure 1. The authors declare no potential conflict of interest, and approve this manuscript as the final version to be published.
Funding
This work was partially financed by ICGEB (CRP/CHI11-01), FONDECYT Postdoctorado 2016 (Nr. 3160086) to MC, and the FONDAP Center for Genome Regulation (Nr. 15090007) to both MA and JE. The publication of this work was supported by the German Research Foundation (DFG) and the Technische Universität München within the funding program Open Access Publishing.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
Akhtar, N., Dickerson, E. B., and Auerbach, R. (2002). The sponge/matrigel angiogenesis assay. Angiogenesis 5, 75–80. doi: 10.1023/A:1021507031486
Albini, A., and Benelli, R. (2007). The chemoinvasion assay: a method to assess tumor and endothelial cell invasion and its modulation. Nat. Protoc. 2, 504–511. doi: 10.1038/nprot.2006.466
Arnaoutova, I., George, J., Kleinman, H. K., and Benton, G. (2009). The endothelial cell tube formation assay on basement membrane turns 20: state of the science and the art. Angiogenesis 12, 267–274. doi: 10.1007/s10456-009-9146-4
Arnaoutova, I., and Kleinman, H. K. (2010). In vitro angiogenesis: endothelial cell tube formation on gelled basement membrane extract. Nat. Protoc. 5, 628–635. doi: 10.1038/nprot.2010.6
Asnani, A., and Peterson, R. T. (2014). The zebrafish as a tool to identify novel therapies for human cardiovascular disease. Dis. Model. Mech. 7, 763–767. doi: 10.1242/dmm.016170
Avraham-Davidi, I., Ely, Y., Pham, V. N., Castranova, D., Grunspan, M., Malkinson, G., et al. (2012). ApoB-containing lipoproteins regulate angiogenesis by modulating expression of VEGF receptor 1. Nat. Med. 18, 967–973. doi: 10.1038/nm.2759
Ayata, R., Chabaud, S., Auger, M., and Pouliot, R. (2015). Behaviour of endothelial cells in a tridimensional in vitro environment. Biomed Res. Int. 2015:630461. doi: 10.1155/2015/630461
Azevedo, A., Grotek, B., Jacinto, A., Weidinger, G., Saúde, L., and Karl, M. (2011). The regenerative capacity of the zebrafish caudal fin is not affected by repeated amputations. PLoS ONE 6:e22820. doi: 10.1371/journal.pone.0022820
Bagatto, B., Francl, J., Liu, B., and Liu, Q. (2006). Cadherin2 (N-cadherin) plays an essential role in zebrafish cardiovascular development. BMC Dev. Biol. 6:23. doi: 10.1186/1471-213X-6-23
Baker, M., Robinson, S. D., Lechertier, T., Barber, P. R., Tavora, B., D'Amico, G., et al. (2011). Use of the mouse aortic ring assay to study angiogenesis. Nat. Protoc. 7, 89–104. doi: 10.1038/nprot.2011.435
Baldessari, D., and Mione, M. (2008). How to create the vascular tree? (Latest) help from the zebrafish. Pharmacol. Ther. 118, 206–230. doi: 10.1016/j.pharmthera.2008.02.010
Bayliss, P. E., Bellavance, K. L., Whitehead, G. G., Abrams, J. M., Aegerter, S., Robbins, H. S., et al. (2006). Chemical modulation of receptor signaling inhibits regenerative angiogenesis in adult zebrafish. Nat. Chem. Biol. 2, 265–273. doi: 10.1038/nchembio778
Bedell, V. M., Wang, Y., Campbell, J. M., Poshusta, T. L., Starker, C. G., Krug, R. G., et al. (2012). In vivo genome editing using a high-efficiency TALEN system. Nature 491, 114–118. doi: 10.1038/nature11537
Benton, G., Arnaoutova, I., George, J., Kleinman, H. K., and Koblinski, J. (2014). Matrigel: from discovery and ECM mimicry to assays and models for cancer research. Adv. Drug Deliv. Rev. 79–80, 3–18. doi: 10.1016/j.addr.2014.06.005
Berthod, F., Symes, J., Tremblay, N., Medin, J., and Auger, F. (2012). Spontaneous fibroblast−derived pericyte recruitment in a human tissue−engineered angiogenesis model in vitro. J. Cell. Physiol. 227, 2130–2137. doi: 10.1002/jcp.22943
Bishop, E. T., Bell, G. T., Bloor, S., Broom, I. J., Hendry, N. F., and Wheatley, D. N. (1999). An in vitro model of angiogenesis: basic features. Angiogenesis 3, 335–344. doi: 10.1023/A:1026546219962
Boncler, M., Różalski, M., Krajewska, U., Podsȩdek, A., and Watala, C. (2014). Comparison of PrestoBlue and MTT assays of cellular viability in the assessment of anti-proliferative effects of plant extracts on human endothelial cells. J. Pharmacol. Toxicol. Methods 69, 9–16. doi: 10.1016/j.vascn.2013.09.003
Boyden, S. (1962). The chemotactic effect of mixtures of antibody and antigen on polymorphonuclear leucocytes. J. Exp. Med. 115, 453–466. doi: 10.1084/jem.115.3.453
Brown, L. A., Rodaway, A. R., Schilling, T. F., Jowett, T., Ingham, P. W., Patient, R. K., et al. (2000). Insights into early vasculogenesis revealed by expression of the ETS-domain transcription factor Fli-1 in wild-type and mutant zebrafish embryos. Mech. Dev. 90, 237–252. doi: 10.1016/S0925-4773(99)00256-7
Bussmann, J., and Schulte-Merker, S. (2011). Rapid BAC selection for tol2-mediated transgenesis in zebrafish. Development 138, 4327–4332. doi: 10.1242/dev.068080
Bussmann, J., Wolfe, S. A., and Siekmann, A. F. (2011). Arterial-venous network formation during brain vascularization involves hemodynamic regulation of chemokine signaling. Development 138, 1717–1726. doi: 10.1242/dev.059881
Chávez, M. N., Schenck, T. L., Hopfner, U., Centeno-Cerdas, C., Somlai-Schweiger, I., Schwarz, C., et al. (2016). Towards autotrophic tissue engineering: photosynthetic gene therapy for regeneration. Biomaterials 75, 25–36. doi: 10.1016/j.biomaterials.2015.10.014
Chen, J. N., Haffter, P., Odenthal, J., Vogelsang, E., Brand, M., Eeden, F. J., et al. (1996). Mutations affecting the cardiovascular system and other internal organs in zebrafish. Development 123, 293–302.
Childs, S., Chen, J.-N. N., Garrity, D. M., and Fishman, M. C. (2002). Patterning of angiogenesis in the zebrafish embryo. Development 129, 973–982.
Choe, C. P., and Crump, J. G. (2015). Eph-Pak2a signaling regulates branching of the pharyngeal endoderm by inhibiting late-stage epithelial dynamics. Development 142, 1089–1094. doi: 10.1242/dev.115774
Chung, S., Sudo, R., Vickerman, V., Zervantonakis, I. K., and Kamm, R. D. (2010). Microfluidic platforms for studies of angiogenesis, cell migration, and cell-cell interactions. Sixth International Bio-Fluid Mechanics Symposium and Workshop March 28-30, 2008 Pasadena, California. Ann. Biomed. Eng. 38, 1164–1177. doi: 10.1007/s10439-010-9899-3
Chwalek, K., Bray, L., and Werner, C. (2014). Tissue-engineered 3D tumor angiogenesis models: potential technologies for anti-cancer drug discovery. Adv. Drug Deliv. Rev. 79–80, 3039. doi: 10.1016/j.addr.2014.05.006
Cimpean, A.-M., Ribatti, D., and Raica, M. (2011). A brief history of angiogenesis assays. Int. J. Dev. Biol. 55, 377382. doi: 10.1387/ijdb.103215ac
Clark, E. R., Hitschler, W. J., Kirby-Smith, H. T., Rex, R. O., and Smith, J. H. (1931). General observations on the ingrowth of new blood vessels into standardized chambers in the rabbit's ear, and the subsequent changes in the newly grown vessels over a period of months. Anat. Rec. 50, 29–167. doi: 10.1002/ar.1090500203
Coffindaffer-Wilson, M., Craig, M. P., and Hove, J. R. (2011). Determination of lymphatic vascular identity and developmental timecourse in zebrafish (Danio rerio). Lymphology 44, 1–12.
Coomber, B. L., and Gotlieb, A. I. (1990). In vitro endothelial wound repair. Interaction of cell migration and proliferation. Arteriosclerosis 10, 215–222. doi: 10.1161/01.ATV.10.2.215
Couffinhal, T., Silver, M., Zheng, L. P., Kearney, M., Witzenbichler, B., and Isner, J. M. (1998). Mouse model of angiogenesis. Am. J. Pathol. 152, 1667–1679.
Coultas, L., Chawengsaksophak, K., and Rossant, J. (2006). Endothelial cells and VEGF in vascular development. Nature 438, 937–945. doi: 10.1038/nature04479
Covassin, L. D., Siekmann, A. F., Kacergis, M. C., Laver, E., Moore, J. C., Villefranc, J. A., et al. (2009). A genetic screen for vascular mutants in zebrafish reveals dynamic roles for Vegf/Plcg1 signaling during artery development. Dev. Biol. 329, 212–226. doi: 10.1016/j.ydbio.2009.02.031
Cross, L. M., Cook, M. A., Lin, S., Chen, J.-N. N., and Rubinstein, A. L. (2003). Rapid analysis of angiogenesis drugs in a live fluorescent zebrafish assay. Arterioscler. Thromb. Vasc. Biol. 23, 911–912. doi: 10.1161/01.ATV.0000068685.72914.7E
Diaz-Santana, A., Shan, M., and Stroock, A. D. (2015). Endothelial cell dynamics during anastomosis in vitro. Integr. Biol. 7, 454–466. doi: 10.1039/C5IB00052A
Donovan, D., Brown, N. J., Bishop, E. T., and Lewis, C. E. (2001). Comparison of three in vitro human “angiogenesis” assays with capillaries formed in vivo. Angiogenesis 4, 113–121. doi: 10.1023/A:1012218401036
Egaña, J., Condurache, A., Lohmeyer, J., Kremer, M., Stöckelhuber, B., Lavandero, S., et al. (2008). Ex vivo method to visualize and quantify vascular networks in native and tissue engineered skin. Langenbeck's Arch. Surg. 394, 349–356. doi: 10.1007/s00423-008-0333-3
Ellertsdóttir, E., Lenard, A., Blum, Y., Krudewig, A., Herwig, L., Affolter, M., et al. (2010). Vascular morphogenesis in the zebrafish embryo. Dev. Biol. 341, 56–65. doi: 10.1016/j.ydbio.2009.10.035
Fierro, F. A., Kalomoiris, S., Sondergaard, C. S., and Nolta, J. A. (2011). Effects on proliferation and differentiation of multipotent bone marrow stromal cells engineered to express growth factors for combined cell and gene therapy. Stem Cells 29, 1727–1737. doi: 10.1002/stem.720
Gaiano, N., Amsterdam, A., Kawakami, K., Allende, M., Becker, T., and Hopkins, N. (1996). Insertional mutagenesis and rapid cloning of essential genes in zebrafish. Nature 383, 829–832. doi: 10.1038/383829a0
Gering, M., Rodaway, A. R., Göttgens, B., Patient, R. K., and Green, A. R. (1998). The SCL gene specifies haemangioblast development from early mesoderm. EMBO J. 17, 4029–4045. doi: 10.1093/emboj/17.14.4029
Gimbrone, M. A., Leapman, S. B., Cotran, R. S., and Folkman, J. (1973). Tumor angiogenesis: iris neovascularization at a distance from experimental intraocular tumors. J. Natl. Cancer Inst. 50, 219–228.
Gjini, E., Hekking, L. H., Küchler, A., Saharinen, P., Wienholds, E., Post, J.-A. A., et al. (2011). Zebrafish Tie-2 shares a redundant role with Tie-1 in heart development and regulates vessel integrity. Dis. Models Mech. 4, 57–66. doi: 10.1242/dmm.005033
Goodwin, A. M. (2007). In vitro assays of angiogenesis for assessment of angiogenic and anti-angiogenic agents. Microvasc. Res. 74, 172–183. doi: 10.1016/j.mvr.2007.05.006
Gordon, K., Schulte, D., Brice, G., Simpson, M. A., Roukens, M. G., Impel, A., et al. (2013). Mutation in vascular endothelial growth factor-C, a ligand for vascular endothelial growth factor receptor-3, is associated with autosomal dominant milroy-like primary lymphedema. Circ. Res. 112, 956–960. doi: 10.1161/CIRCRESAHA.113.300350
Gore, A. V., Monzo, K., Cha, Y. R., Pan, W., and Weinstein, B. M. (2012). Vascular development in the zebrafish. Cold Spring Harb. Perspect. Med. 2:a006684. doi: 10.1101/cshperspect.a006684
Gough, W., Hulkower, K. I., Lynch, R., McGlynn, P., Uhlik, M., Yan, L., et al. (2011). A quantitative, facile, and high-throughput image-based cell migration method is a robust alternative to the scratch assay. J. Biomol. Screen. 16, 155–163. doi: 10.1177/1087057110393340
Greenway, F. L., Liu, Z., Yu, Y., Caruso, M. K., Roberts, A. T., Lyons, J., et al. (2007). An assay to measure angiogenesis in human fat tissue. Obes. Surg. 17, 510–515. doi: 10.1007/s11695-007-9089-z
Habeck, H., Odenthal, J., Walderich, B., Maischein, H., and Schulte-Merker, S. (2002). Analysis of a zebrafish VEGF receptor mutant reveals specific disruption of angiogenesis. Curr. Biol. 12, 1405–1412. doi: 10.1016/S0960-9822(02)01044-8
Haffter, P., Granato, M., Brand, M., Mullins, M. C., Hammerschmidt, M., Kane, D. A., et al. (1996). The identification of genes with unique and essential functions in the development of the zebrafish, Danio rerio. Development 123, 1–36.
Haldi, M., Ton, C., Seng, W. L., and McGrath, P. (2006). Human melanoma cells transplanted into zebrafish proliferate, migrate, produce melanin, form masses and stimulate angiogenesis in zebrafish. Angiogenesis 9, 139–151. doi: 10.1007/s10456-006-9040-2
Harder, Y., Schmauss, D., Wettstein, R., Egaña, J. T. T., Weiss, F., Weinzierl, A., et al. (2014). Ischemic tissue injury in the dorsal skinfold chamber of the mouse: a skin flap model to investigate acute persistent ischemia. J. Vis. Exp. 93:e51900. doi: 10.3791/51900
Hasan, S. S., and Siekmann, A. F. (2015). The same but different: signaling pathways in control of endothelial cell migration. Curr. Opin. Cell Biol. 36, 86–92. doi: 10.1016/j.ceb.2015.07.009
Heicklen-Klein, A., and Evans, T. (2004). T-box binding sites are required for activity of a cardiac GATA-4 enhancer. Dev. Biol. 267, 490–504. doi: 10.1016/j.ydbio.2003.09.042
Henkind, P. (1978). Ocular neovascularization. The Krill memorial lecture. Am. J. Ophthalmol. 85, 287–301. doi: 10.1016/S0002-9394(14)77719-0
Hermkens, D. M., Impel, A., van, Urasaki, A., Bussmann, J., Duckers, H. J., and Schulte-Merker, S. (2015). Sox7 controls arterial specification in conjunction with hey2 and efnb2 function. Development 142, 1695–1704. doi: 10.1242/dev.117275
Herwig, L., Blum, Y., Krudewig, A., Ellertsdottir, E., Lenard, A., Belting, H.-G. G., et al. (2011). Distinct cellular mechanisms of blood vessel fusion in the zebrafish embryo. Curr. Biol. 21, 1942–1948. doi: 10.1016/j.cub.2011.10.016
Hetheridge, C., Mavria, G., and Mellor, H. (2011). Uses of the in vitro endothelial-fibroblast organotypic co-culture assay in angiogenesis research. Biochem. Soc. Trans. 39, 1597–1600. doi: 10.1042/BST20110738
Hill, A. J., Teraoka, H., Heideman, W., and Peterson, R. E. (2005). Zebrafish as a model vertebrate for investigating chemical toxicity. Toxicol. Sci. 86, 6–19. doi: 10.1093/toxsci/kfi110
Ho, Y.-L. L., Lin, Y.-H. H., Tsai, I.-J. J., Hsieh, F.-J. J., and Tsai, H.-J. J. (2007). In vivo assessment of cardiac morphology and function in heart-specific green fluorescent zebrafish. J. Formos. Med. Assoc. 106, 181–186. doi: 10.1016/S0929-6646(09)60238-2
Hogan, B. M., Herpers, R., Witte, M., Heloterä, H., Alitalo, K., Duckers, H. J., et al. (2010). Vegfc/Flt4 signalling is suppressed by Dll4 in developing zebrafish intersegmental arteries. Development 136, 4001–4009. doi: 10.1242/dev.039990
Huang, C. C., Lawson, N. D., Weinstein, B. M., and Johnson, S. L. (2003). reg6 is required for branching morphogenesis during blood vessel regeneration in zebrafish caudal fins. Dev. Biol. 264, 263–274. doi: 10.1016/j.ydbio.2003.08.016
Huang, H., Zhang, B., Hartenstein, P. A., Chen, J. N., and Lin, S. (2005). NXT2 is required for embryonic heart development in zebrafish. BMC Dev. Biol. 5:7. doi: 10.1186/1471-213X-5-7
Hulkower, K. I., and Herber, R. L. (2011). Cell migration and invasion assays as tools for drug discovery. Pharmaceutics 3, 107–124. doi: 10.3390/pharmaceutics3010107
Hwang, W. Y., Fu, Y., Reyon, D., Maeder, M. L., Tsai, S. Q., Sander, J. D., et al. (2013). Efficient genome editing in zebrafish using a CRISPR-Cas system. Nat. Biotechnol. 31, 227–229. doi: 10.1038/nbt.2501
Ichioka, S., Shibata, M., Kosaki, K., Sato, Y., Harii, K., and Kamiya, A. (1997). Effects of shear stress on wound-healing angiogenesis in the rabbit ear chamber. J. Surg. Res. 72, 29–35. doi: 10.1006/jsre.1997.5170
Irvin, M. W., Zijlstra, A., Wikswo, J. P., and Pozzi, A. (2014). Techniques and assays for the study of angiogenesis. Exp. Biol. Med. 239, 14761488. doi: 10.1177/1535370214529386
Isogai, S., Horiguchi, M., and Weinstein, B. M. (2001). The vascular anatomy of the developing zebrafish: an atlas of embryonic and early larval development. Dev. Biol. 230, 278–301. doi: 10.1006/dbio.2000.9995
Isogai, S., Lawson, N. D., Torrealday, S., Horiguchi, M., and Weinstein, B. M. (2003). Angiogenic network formation in the developing vertebrate trunk. Development 130, 5281–5290. doi: 10.1242/dev.00733
Jin, S.-W. W., Beis, D., Mitchell, T., Chen, J.-N. N., and Stainier, D. Y. (2006). Cellular and molecular analyses of vascular tube and lumen formation in zebrafish. Development 132, 5199–5209. doi: 10.1242/dev.02087
Jin, S.-W. W., Herzog, W., Santoro, M. M., Mitchell, T. S., Frantsve, J., Jungblut, B., et al. (2007). A transgene-assisted genetic screen identifies essential regulators of vascular development in vertebrate embryos. Dev. Biol. 307, 29–42. doi: 10.1016/j.ydbio.2007.03.526
Kamei, M., Isogai, S., Pan, W., and Weinstein, B. M. (2010). Imaging blood vessels in the zebrafish. Methods Cell Biol. 100, 27–54. doi: 10.1016/B978-0-12-384892-5.00002-5
Kepp, O., Galluzzi, L., Lipinski, M., Yuan, J., and Kroemer, G. (2011). Cell death assays for drug discovery. Nat. Rev. Drug Discov. 10, 221–237. doi: 10.1038/nrd3373
Kimura, E., Deguchi, T., Kamei, Y., Shoji, W., Yuba, S., and Hitomi, J. (2013). Application of infrared laser to the zebrafish vascular system: gene induction, tracing, and ablation of single endothelial cells. Arterioscler. Thromb. Vasc. Biol. 33, 1264–1270. doi: 10.1161/ATVBAHA.112.300602
Kleaveland, B., Zheng, X., Liu, J. J., Blum, Y., Tung, J. J., Zou, Z., et al. (2009). Regulation of cardiovascular development and integrity by the heart of glass-cerebral cavernous malformation protein pathway. Nat. Med. 15, 169–176. doi: 10.1038/nm.1918
Köhler, C., Orrenius, S., and Zhivotovsky, B. (2002). Evaluation of caspase activity in apoptotic cells. J. Immunol. Methods 265, 97–110 doi: 10.1016/s0022-1759(02)00073-x
Kok, F. O., Shin, M., Ni, C.-W. W., Gupta, A., Grosse, A. S., Impel, A., et al. (2015). Reverse genetic screening reveals poor correlation between morpholino-induced and mutant phenotypes in zebrafish. Dev. Cell 32, 97–108. doi: 10.1016/j.devcel.2014.11.018
Küchler, A. M., Gjini, E., Peterson-Maduro, J., Cancilla, B., Wolburg, H., and Schulte-Merker, S. (2006). Development of the zebrafish lymphatic system requires VEGFC signaling. Curr. Biol. 16, 1244–1248. doi: 10.1016/j.cub.2006.05.026
Kunisaki, Y., and Frenette, P. S. (2014). Influences of vascular niches on hematopoietic stem cell fate. Int. J. Hematol. 99, 699–705. doi: 10.1007/s12185-014-1580-4
Lagendijk, A. K., Yap, A. S., and Hogan, B. M. (2014). Endothelial cell-cell adhesion during zebrafish vascular development. Cell Adh. Migr. 8, 136–145. doi: 10.4161/cam.28229
Larson, E. M., Doughman, D. J., Gregerson, D. S., and Obritsch, W. F. (1997). A new, simple, nonradioactive, nontoxic in vitro assay to monitor corneal endothelial cell viability. Invest. Ophthalmol. Vis. Sci. 38, 1929–1933.
Lawson, N. D. (2016). Reverse genetics in zebrafish: mutants, morphants, and moving forward. Trends Cell Biol. 2, 77–79. doi: 10.1016/j.tcb.2015.11.005
Lawson, N. D., Mugford, J. W., Diamond, B. A., and Weinstein, B. M. (2003). Phospholipase C gamma-1 is required downstream of vascular endothelial growth factor during arterial development. Genes Develop. 17, 1346–1351. doi: 10.1101/gad.1072203
Lawson, N. D., Scheer, N., Pham, V. N., Kim, C. H., Chitnis, A. B., Campos-Ortega, J. A., et al. (2001). Notch signaling is required for arterial-venous differentiation during embryonic vascular development. Development 128, 3675–3683.
Lawson, N. D., Vogel, A. M., and Weinstein, B. M. (2002). Sonic hedgehog and vascular endothelial growth factor act upstream of the Notch pathway during arterial endothelial differentiation. Dev. Cell 3, 127–136. doi: 10.1016/S1534-5807(02)00198-3
Lawson, N. D., and Weinstein, B. M. (2002a). In vivo imaging of embryonic vascular development using transgenic zebrafish. Dev. Biol. 248, 307–318. doi: 10.1006/dbio.2002.0711
Lawson, N. D., and Weinstein, B. M. (2002b). Arteries and veins: making a difference with zebrafish. Nat. Rev. Genet. 3, 674–682. doi: 10.1038/nrg888
Lehr, H. A., Leunig, M., Menger, M. D., Nolte, D., and Messmer, K. (1993). Dorsal skinfold chamber technique for intravital microscopy in nude mice. Am. J. Pathol. 143, 1055–1062.
Lenard, A., Ellertsdottir, E., Herwig, L., Krudewig, A., Sauteur, L., Belting, H.-G. G., et al. (2013). In vivo analysis reveals a highly stereotypic morphogenetic pathway of vascular anastomosis. Dev. Cell 25, 492–506. doi: 10.1016/j.devcel.2013.05.010
Li, J., and Stuhlmann, H. (2011). In vitro imaging of angiogenesis using embryonic stem cell-derived endothelial cells. Stem Cells Dev. 21, 331–342. doi: 10.1089/scd.2010.0587
Lieschke, G. J., and Currie, P. D. (2007). Animal models of human disease: zebrafish swim into view. Nat. Rev. Genet. 8, 353–367. doi: 10.1038/nrg2091
Limbourg, A., Korff, T., Napp, L. C., Schaper, W., Drexler, H., and Limbourg, F. P. (2009). Evaluation of postnatal arteriogenesis and angiogenesis in a mouse model of hind-limb ischemia. Nat. Protoc. 4, 1737–1746. doi: 10.1038/nprot.2009.185
Long, Q., Meng, A., Wang, H., Jessen, J. R., Farrell, M. J., and Lin, S. (1997). GATA-1 expression pattern can be recapitulated in living transgenic zebrafish using GFP reporter gene. Development 124, 4105–4111.
Mably, J. D., Chuang, L. P., Serluca, F. C., Mohideen, M. A., Chen, J. N., and Fishman, M. C. (2006). santa and valentine pattern concentric growth of cardiac myocardium in the zebrafish. Development 133, 3139–3146. doi: 10.1242/dev.02469
Mably, J. D., Mohideen, M. A., Burns, C. G., Chen, J.-N. N., and Fishman, M. C. (2003). heart of glass regulates the concentric growth of the heart in zebrafish. Curr. Biol. 13, 2138–2147. doi: 10.1016/j.cub.2003.11.055
MacRae, C. A., and Peterson, R. T. (2015). Zebrafish as tools for drug discovery. Nat. Rev. Drug Discov. 10, 721–731. doi: 10.1038/nrd4627
Mayer, F. L., Whalen, E. A., and Rheins, L. A. (1994). A regulatory overview of alternatives to animal testing: United States, Europe, and Japan. J. Toxicol. Cutaneous Ocul. Toxicol. 13, 3–22. doi: 10.3109/15569529409037506
Merfeld-Clauss, S., Gollahalli, N., March, K. L., and Traktuev, D. O. (2010). Adipose tissue progenitor cells directly interact with endothelial cells to induce vascular network formation. Tissue Eng. A 16, 2953–2966. doi: 10.1089/ten.tea.2009.0635
Monti-Hughes, A., Aromando, R. F., Pérez, M. A., Schwint, A. E., and Itoiz, M. E. (2015). The hamster cheek pouch model for field cancerization studies. Periodontol. 2000 67, 292–311. doi: 10.1111/prd.12066
Motoike, T., Loughna, S., Perens, E., Roman, B. L., Liao, W., Chau, T. C., et al. (2000). Universal GFP reporter for the study of vascular development. Genesis 28, 75–81. doi: 10.1002/1526-968X(200010)28:2<75::AID-GENE50>3.0.CO;2-S
Moya-Díaz, J., Peña, O. A., Sánchez, M., Ureta, D. A., Reynaert, N. G., Anguita-Salinas, C., et al. (2014). Electroablation: a method for neurectomy and localized tissue injury. BMC Dev. Biol. 14:7. doi: 10.1186/1471-213X-14-7
Nasevicius, A., Larson, J., and Ekker, S. C. (2000). Distinct requirements for zebrafish angiogenesis revealed by a VEGF-A morphant. Yeast 17, 294–301. doi: 10.1002/1097-0061(200012)17:4<294::AID-YEA54>3.0.CO;2-5
Neufeld, S., Planas-Paz, L., and Lammert, E. (2014). Blood and lymphatic vascular tube formation in mouse. Seminars Cell Dev. Biol. 31, 115123. doi: 10.1016/j.semcdb.2014.02.013
Nicosia, R. F. (2009). The aortic ring model of angiogenesis: a quarter century of search and discovery. J. Cell. Mol. Med. 13, 4113–4136. doi: 10.1111/j.1582-4934.2009.00891.x
Nicosia, R., Zorzi, P., Ligresti, G., Morishita, A., and Aplin, A. (2011). Paracrine regulation of angiogenesis by different cell types in the aorta ring model. Int. J. Dev. Biol. 55, 447–453. doi: 10.1387/ijdb.103222rn
Niles, A. L., Moravec, R. A., Eric Hesselberth, P., Scurria, M. A., Daily, W. J., and Riss, T. L. (2007). A homogeneous assay to measure live and dead cells in the same sample by detecting different protease markers. Anal. Biochem. 366, 197–206. doi: 10.1016/j.ab.2007.04.007
Niles, A. L., and Riss, T. L. (2015). Multiplexed viability, cytotoxicity, and caspase activity assays. Methods Mol. Biol. 1219, 21–33. doi: 10.1007/978-1-4939-1661-0_3
Norrby, K. (2006). In vivo models of angiogenesis. J. Cell. Mol. Med. 10, 588–612. doi: 10.1111/j.1582-4934.2006.tb00423.x
Palmer, G. M., Fontanella, A. N., Shan, S., Hanna, G., Zhang, G., Fraser, C. L., et al. (2011). In vivo optical molecular imaging and analysis in mice using dorsal window chamber models applied to hypoxia, vasculature and fluorescent reporters. Nat. Protoc. 6, 1355–1366. doi: 10.1038/nprot.2011.349
Pandya, N., Dhalla, N., and Santani, D. (2006). Angiogenesis—a new target for future therapy. Vascul. Pharmacol. 44, 265274. doi: 10.1016/j.vph.2006.01.005
Pardo-Martin, C., Chang, T.-Y., Koo, B., Gilleland, C., Wasserman, S., and Yanik, M. (2010). High-throughput in vivo vertebrate screening. Nat. Methods 7, 634–636. doi: 10.1038/nmeth.1481
Peterson, R. T., Shaw, S. Y., Peterson, T. A., Milan, D. J., Zhong, T. P., Schreiber, S. L., et al. (2004). Chemical suppression of a genetic mutation in a zebrafish model of aortic coarctation. Nat. Biotechnol. 22, 595–599. doi: 10.1038/nbt963
Pfefferli, C., and Jaźwińska, A. (2015). The art of fin regeneration in zebrafish. Regeneration 2, 72–83. doi: 10.1002/reg2.33
Poss, K. D., Keating, M. T., and Nechiporuk, A. (2003). Tales of regeneration in zebrafish. Dev. Dyn. 226, 202–210. doi: 10.1002/dvdy.10220
Poujade, M., Grasland-Mongrain, E., Hertzog, A., Jouanneau, J., Chavrier, P., Ladoux, B., et al. (2007). Collective migration of an epithelial monolayer in response to a model wound. Proc. Natl. Acad. Sci. U.S.A. 104, 15988–15993. doi: 10.1073/pnas.0705062104
Qin, L., Zeng, H., and Zhao, D. (2006). Requirement of protein kinase D tyrosine phosphorylation for VEGF-A165-induced angiogenesis through its interaction and regulation of phospholipase Cgamma phosphorylation. J. Biol. Chem. 281, 32550–32558. doi: 10.1074/jbc.M604853200
Raghunath, M., Wong, Y., Farooq, M., and Ge, R. (2009). Pharmacologically induced angiogenesis in transgenic zebrafish. Biochem. Biophys. Res. Commun. 378, 766–771. doi: 10.1016/j.bbrc.2008.11.127
Ramasamy, S. K., Kusumbe, A. P., and Adams, R. H. (2015). Regulation of tissue morphogenesis by endothelial cell-derived signals. Trends Cell Biol. 25, 148–157. doi: 10.1016/j.tcb.2014.11.007
Reed, M., Damodarasamy, M., and Vernon, R. (2011). Angiogenesis in vitro utilizing murine vascular explants in miniaturized 3-dimensional collagen gels. Open Circ. Vasc. J. 4, 12–17. doi: 10.2174/1877382601104010012
Rezzola, S., Belleri, M., Gariano, G., Ribatti, D., Costagliola, C., Semeraro, F., et al. (2014). In vitro and ex vivo retina angiogenesis assays. Angiogenesis 17, 429–442. doi: 10.1007/s10456-013-9398-x
Ribatti, D. (2008). Chick embryo chorioallantoic membrane as a useful tool to study angiogenesis. Int. Rev. Cell Mol. Biol. 270, 181–224. doi: 10.1016/S1937-6448(08)01405-6
Ribatti, D., and Crivellato, E. (2012). “Sprouting angiogenesis,” a reappraisal. Dev. Biol. 372, 157–165. doi: 10.1016/j.ydbio.2012.09.018
Ribatti, D., Nico, B., Vacca, A., Roncali, L., Burri, P. H., and Djonov, V. (2001). Chorioallantoic membrane capillary bed: a useful target for studying angiogenesis and anti-angiogenesis in vivo. Anat. Rec. 264, 317–324. doi: 10.1002/ar.10021
Roman, B. L., Pham, V. N., Lawson, N. D., Kulik, M., Childs, S., Lekven, A. C., et al. (2002). Disruption of acvrl1 increases endothelial cell number in zebrafish cranial vessels. Development 129, 3009–3019.
Sacilotto, N., Monteiro, R., Fritzsche, M., Becker, P. W., Sanchez-Del-Campo, L., Liu, K., et al. (2013). Analysis of Dll4 regulation reveals a combinatorial role for Sox and Notch in arterial development. Proc. Natl. Acad. Sci. U.S.A. 110, 11893–11898. doi: 10.1073/pnas.1300805110
Sarkanen, J.-R. R., Vuorenpää, H., Huttala, O., Mannerström, B., Kuokkanen, H., Miettinen, S., et al. (2012). Adipose stromal cell tubule network model provides a versatile tool for vascular research and tissue engineering. Cells Tissues Organs 196, 385–397. doi: 10.1159/000336679
Schenck, T. L., Chávez, M. N., Condurache, A. P., Hopfner, U., Rezaeian, F., Machens, H.-G. G., et al. (2014). A full skin defect model to evaluate vascularization of biomaterials in vivo. J. Vis. Exp. 90:e51428. doi: 10.3791/51428
Schuermann, A., Helker, C. S., and Herzog, W. (2014). Angiogenesis in zebrafish. Semin. Cell Dev. Biol. 31, 106–114. doi: 10.1016/j.semcdb.2014.04.037
Seano, G., Chiaverina, G., Gagliardi, P. A., di Blasio, L., Sessa, R., Bussolino, F., et al. (2013). Modeling human tumor angiogenesis in a three-dimensional culture system. Blood 121, e129–e137. doi: 10.1182/blood-2012-08-452292
Shao, Z., Friedlander, M., Hurst, C., Cui, Z., Pei, D., Evans, L., et al. (2013). Choroid sprouting assay: an ex vivo model of microvascular angiogenesis. PLoS ONE 8:e69552. doi: 10.1371/journal.pone.0069552
Smith, S. M., Wunder, M. B., Norris, D. A., and Shellman, Y. G. (2011). A simple protocol for using a LDH-based cytotoxicity assay to assess the effects of death and growth inhibition at the same time. PLoS ONE 6:e26908. doi: 10.1371/journal.pone.0026908
Sondergaard, C. S., Hess, D. A., Maxwell, D. J., Weinheimer, C., Rosová, I., Creer, M. H., et al. (2010). Human cord blood progenitors with high aldehyde dehydrogenase activity improve vascular density in a model of acute myocardial infarction. J. Transl. Med. 8:24. doi: 10.1186/1479-5876-8-24
Song, H.-H., Park, K., and Gerecht, S. (2014). Hydrogels to model 3D in vitro microenvironment of tumor vascularization. Adv. Drug Deliv. Rev. 79–80, 19–29. doi: 10.1016/j.addr.2014.06.002
Stainier, D. Y., Weinstein, B. M., Detrich, H. W., Zon, L. I., and Fishman, M. C. (1995). Cloche, an early acting zebrafish gene, is required by both the endothelial and hematopoietic lineages. Development 121, 3141–3150.
Staton, C. A., Reed, M. W., and Brown, N. J. (2009). A critical analysis of current in vitro and in vivo angiogenesis assays. Int. J. Exp. Pathol. 90, 195–221. doi: 10.1111/j.1365-2613.2008.00633.x
Staton, C. A., Stribbling, S. M., Tazzyman, S., Hughes, R., Brown, N. J., and Lewis, C. E. (2004). Current methods for assaying angiogenesis in vitro and in vivo. Int. J. Exp. Pathol. 85, 233–248. doi: 10.1111/j.0959-9673.2004.00396.x
Steinritz, D., Schmidt, A., Balszuweit, F., Thiermann, H., Ibrahim, M., Bölck, B., et al. (2015). Assessment of endothelial cell migration after exposure to toxic chemicals. J. Vis. Exp. 101:e52768. doi: 10.3791/52768
Stoddart, M. J. (2011). Cell viability assays: introduction. Methods Mol. Biol. 740, 1–6. doi: 10.1007/978-1-61779-108-6_1
Strilić, B., Kucera, T., Eglinger, J., Hughes, M. R., McNagny, K. M., Tsukita, S., et al. (2009). The molecular basis of vascular lumen formation in the developing mouse aorta. Dev. Cell 17, 505–515. doi: 10.1016/j.devcel.2009.08.011
Styp-Rekowska, B., Hlushchuk, R., Pries, A. R., and Djonov, V. (2011). Intussusceptive angiogenesis: pillars against the blood flow. Acta Physiol. 202, 213–223. doi: 10.1111/j.1748-1716.2011.02321.x
Swift, M. R., Pham, V. N., Castranova, D., Bell, K., Poole, R. J., and Weinstein, B. M. (2014). SoxF factors and Notch regulate nr2f2 gene expression during venous differentiation in zebrafish. Dev. Biol. 390, 116–125. doi: 10.1016/j.ydbio.2014.03.018
Tamplin, O., and Zon, L. (2010). Fishing at the cellular level. Nat. Methods 7, 600–601. doi: 10.1038/nmeth0810-600
Taqueti, V. R., and Jaffer, F. A. (2013). High-resolution molecular imaging via intravital microscopy: illuminating vascular biology in vivo. Integr. Biol. 5, 278–290. doi: 10.1039/C2IB20194A
Taylor, K., Grant, N., Temperley, N., and Patton, E. (2010). Small molecule screening in zebrafish: an in vivo approach to identifying new chemical tools and drug leads. Cell Commun. Signal. 8:11. doi: 10.1186/1478-811X-8-11
Thisse, B., and Thisse, C. (2014). In situ hybridization on whole-mount zebrafish embryos and young larvae. Methods Mol. Biol. 1211, 53–67. doi: 10.1007/978-1-4939-1459-3_5
Tobia, C., De Sena, G., and Presta, M. (2011). Zebrafish embryo, a tool to study tumor angiogenesis. Int. J. Dev. Biol. 55, 505–509. doi: 10.1387/ijdb.103238ct
Tran, T. C., Sneed, B., Haider, J., Blavo, D., White, A., Aiyejorun, T., et al. (2007). Automated, quantitative screening assay for antiangiogenic compounds using transgenic zebrafish. Cancer Res. 67, 11386–11392. doi: 10.1158/0008-5472.CAN-07-3126
Traver, D., Paw, B. H., Poss, K. D., Penberthy, W. T., Lin, S., and Zon, L. I. (2003). Transplantation and in vivo imaging of multilineage engraftment in zebrafish bloodless mutants. Nat. Immunol. 4, 1238–1246. doi: 10.1038/ni1007
Ucuzian, A., and Greisler, H. (2007). In vitro models of angiogenesis. World J. Surg. 31, 654–663. doi: 10.1007/s00268-006-0763-4
Udvadia, A. J., and Linney, E. (2003). Windows into development: historic, current, and future perspectives on transgenic zebrafish. Dev. Biol. 256, 1–17. doi: 10.1016/S0012-1606(02)00083-0
Unger, R. E., Krump-Konvalinkova, V., Peters, K., and Kirkpatrick, C. J. (2002). In vitro expression of the endothelial phenotype: comparative study of primary isolated cells and cell lines, including the novel cell line HPMEC-ST1.6R. Microvasc. Res. 64, 384–397. doi: 10.1006/mvre.2002.2434
Vailhé, B., Vittet, D., and Feige, J. J. (2001). In vitro models of vasculogenesis and angiogenesis. Lab. Invest. 81, 439–452. doi: 10.1038/labinvest.3780252
van Rooijen, E., Voest, E. E., Logister, I., Korving, J., Schwerte, T., Schulte-Merker, S., et al. (2009). Zebrafish mutants in the von Hippel-Lindau tumor suppressor display a hypoxic response and recapitulate key aspects of Chuvash polycythemia. Blood 113, 6449–6460. doi: 10.1182/blood-2008-07-167890
Varshney, G. K., Pei, W., LaFave, M. C., Idol, J., Xu, L., Gallardo, V., et al. (2015). High-throughput gene targeting and phenotyping in zebrafish using CRISPR/Cas9. Genome Res. 25, 1030–1042. doi: 10.1101/gr.186379.114
Verseijden, F., Posthumus-van Sluijs, S. J., Pavljasevic, P., Hofer, S. O., Osch, G. J., and van Farrell, E. (2010). Adult human bone marrow- and adipose tissue-derived stromal cells support the formation of prevascular-like structures from endothelial cells in vitro. Tissue Eng. A 16, 101–114. doi: 10.1089/ten.tea.2009.0106
Wang, P., Henning, S. M., and Heber, D. (2010). Limitations of MTT and MTS-based assays for measurement of antiproliferative activity of green tea polyphenols. PLoS ONE 5:e10202. doi: 10.1371/journal.pone.0010202
Wang, Y., Pan, L., Moens, C. B., and Appel, B. (2014). Notch3 establishes brain vascular integrity by regulating pericyte number. Development 141, 307–317. doi: 10.1242/dev.096107
Weiss, A., Ding, X., Beijnum, J., Wong, I., Wong, T., Berndsen, R., et al. (2015). Rapid optimization of drug combinations for the optimal angiostatic treatment of cancer. Angiogenesis. 18, 233–244. doi: 10.1007/s10456-015-9462-9
Weiss, O., Kaufman, R., Michaeli, N., and Inbal, A. (2012). Abnormal vasculature interferes with optic fissure closure in lmo2 mutant zebrafish embryos. Dev. Biol. 369, 191–198. doi: 10.1016/j.ydbio.2012.06.029
Whitfield, M. L., George, L. K., Grant, G. D., and Perou, C. M. (2006). Common markers of proliferation. Nat. Rev. Cancer 6, 99–106. doi: 10.1038/nrc1802
Wilkinson, R. N., and van Eeden, F. J. (2014). The zebrafish as a model of vascular development and disease. Prog. Mol. Biol. Transl. Sci. 124, 93–122. doi: 10.1016/B978-0-12-386930-2.00005-7
Xu, C., Hasan, S. S., Schmidt, I., Rocha, S. F., Pitulescu, M. E., Bussmann, J., et al. (2014). Arteries are formed by vein-derived endothelial tip cells. Nat. Commun. 5, 5758. doi: 10.1038/ncomms6758
Yang, Y., Tang, G., Yan, J., Park, B., Hoffman, A., Tie, G., et al. (2008). Cellular and molecular mechanism regulating blood flow recovery in acute versus gradual femoral artery occlusion are distinct in the mouse. J. Vasc. Surg. 48, 1546–1558. doi: 10.1016/j.jvs.2008.07.063
Yarrow, J. C., Perlman, Z. E., Westwood, N. J., and Mitchison, T. J. (2004). A high-throughput cell migration assay using scratch wound healing, a comparison of image-based readout methods. BMC Biotechnol. 4:21. doi: 10.1186/1472-6750-4-21
Young, E. W. (2014). Advances in microfluidic cell culture systems for studying angiogenesis. J. Lab. Autom. 18, 427–436. doi: 10.1177/2211068213495206
Yu, J. A., Castranova, D., Pham, V. N., and Weinstein, B. M. (2015). Single-cell analysis of endothelial morphogenesis in vivo. Development 142, 2951–2961. doi: 10.1242/dev.123174
Zhen, F., Lan, Y., Yan, B., Zhang, W., and Wen, Z. (2013). Hemogenic endothelium specification and hematopoietic stem cell maintenance employ distinct Scl isoforms. Development 140, 3977–3985. doi: 10.1242/dev.097071
Keywords: Danio rerio, vascular development, vessel regeneration, angiogenesis assay, high-throughput screening assays, endothelial markers
Citation: Chávez MN, Aedo G, Fierro FA, Allende ML and Egaña JT (2016) Zebrafish as an Emerging Model Organism to Study Angiogenesis in Development and Regeneration. Front. Physiol. 7:56. doi: 10.3389/fphys.2016.00056
Received: 25 September 2015; Accepted: 05 February 2016;
Published: 08 March 2016.
Edited by:
Francisco J. Rivera, Paracelsus Private Medical University of Salzburg, AustriaReviewed by:
Grzegorz Sowa, University of Missouri School of Medicine, USAEirini Trompouki, Max Planck Institute for Immunobiology and Epigenetics, Germany
Copyright © 2016 Chávez, Aedo, Fierro, Allende and Egaña. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Miguel L. Allende, bWFsbGVuZGVAdS51Y2hpbGUuY2w=;
José T. Egaña, anRlQHVjLmNs