 Mariarosaria Santillo
Mariarosaria Santillo Antonio Colantuoni
Antonio Colantuoni Paolo Mondola
Paolo Mondola Bruna Guida
Bruna Guida Simona Damiano
Simona Damiano
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
MINI REVIEW article
Front. Physiol. , 07 July 2015
Sec. Vascular Physiology
Volume 6 - 2015 | https://doi.org/10.3389/fphys.2015.00194
Blood pressure homeostasis is maintained by several mechanisms regulating cardiac output, vascular resistances, and blood volume. At cellular levels, reactive oxygen species (ROS) signaling is involved in multiple molecular mechanisms controlling blood pressure. Among ROS producing systems, NADPH oxidases (NOXs), expressed in different cells of the cardiovascular system, are the most important enzymes clearly linked to the development of hypertension. NOXs exert a central role in cardiac mechanosensing, endothelium-dependent relaxation, and Angiotensin-II (Ang-II) redox signaling regulating vascular tone. The central role of NOXs in redox-dependent cardiovascular cell functions renders these enzymes a promising pharmacological target for the treatment of cardiovascular diseases, including hypertension. The aim of the present review is to focus on the physiological role of the cardiovascular NOX-generating ROS in the molecular and cellular mechanisms affecting blood pressure.
Blood pressure is regulated by a dynamic equilibrium of different complex mechanisms (Opie, 2004; Raven and Chapleau, 2014). The main factor determining the systemic blood pressure is the blood arterial volume that, in turn, depends on the cardiac output and vascular resistances. In addition to the nervous and chemical factors, the cardiac output, is affected by mechanical factors ensuring the adjustment of cardiac output to the venous return and afterload. On the other hand, vascular resistance depends in part on the characteristics of the blood (viscosity) and on the diameter of the vascular lumen. Smooth muscle cell layer of the resistance arteries may contract or relax resulting in a parallel increase or decrease of blood pressure, respectively. Several mechanisms regulate vascular tone. Adrenergic sympathetic fibers exert a vasoconstrictory effect through the activation of alfa1-adrenergic receptors of the vascular smooth muscle cells (VSMCs). In addition, paracrine, hormonal and mechanical mechanisms contribute to the fine regulation of vascular tone modulating blood pressure.
NOX enzymes are membrane NADPH oxidases with the unique role of producing superoxide anions by one electron reduction of oxygen using NAD(P)H as electron donor (Bedard and Krause, 2007).
NADPH oxidase, first discovered in phagocytes (Segal and Jones, 1978) is a multicomponent complex comprising two integral membrane proteins, the catalytic subunit gp91phox (now referred to as NOX2) and p22phox, and the cytosolic components p47phox, p67phox, p40phox, and Rac1 or 2 (Dinauer et al., 1987; Knaus et al., 1991; Babior et al., 2002). Upon stimulation, cytosolic subunits translocate to the membrane activating the enzyme.
Up to now, in mammalian, seven different NOX genes (NOX1 to 5 and DUOX1 and 2) have been identified (Lambeth, 2004) (Figure 1). Like NOX2, also NOX1, NOX3, and NOX4 are associated with p22phox, but the mechanisms of activation are different. NOX1 is activated by membrane translocation of the cytosolic subunits NOXO1, NOXA1, and Rac 1or 2, while NOX3 requires NOXO1 but is still uncertain the role of the other cytosolic subunits. NOX4, NOX5, DUOX 1, and 2 activity is not modulated by cytosolic subunits. DUOX 1 and 2 terminate at N-terminus with an extracellular peroxidase-homology domain (PHD) (Donko et al., 2005) and together with NOX5, are modulated by calcium. NOX4 and DUOX1 and 2 produce hydrogen peroxide instead of superoxide anion (Martin et al., 2006).
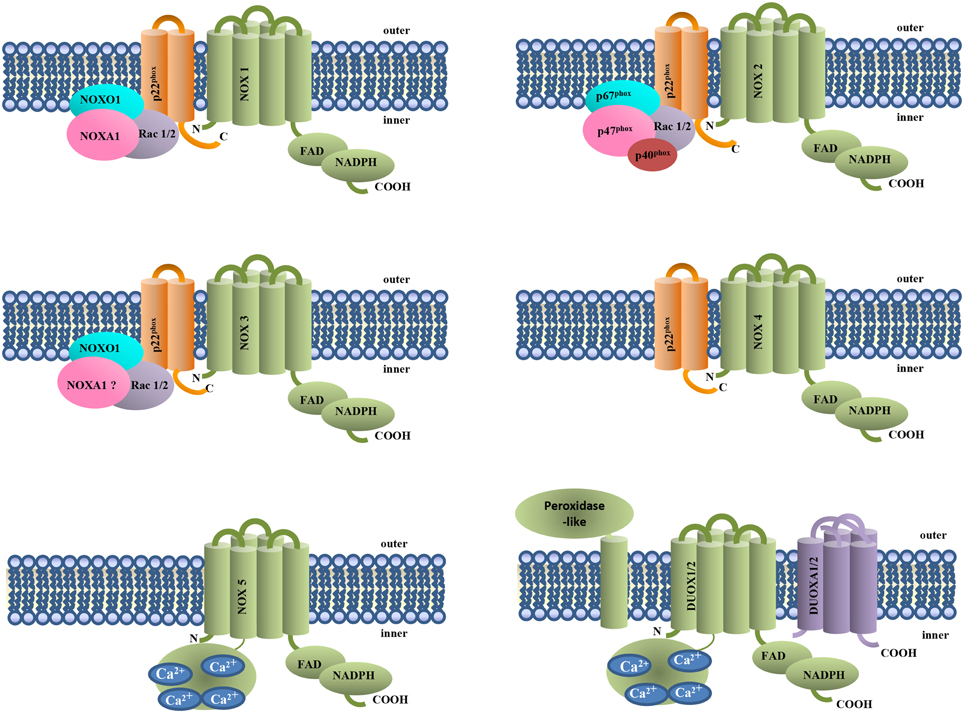
Figure 1. NOX isoforms and regulatory subunits. NOX1-4 are associated to the membrane subunit p22phox. NOX4, NOX5, and DUOX1 and 2 do not require cytosolic subunits for their activity. NOX5 and DUOX1 and 2 activation requires Ca2+ binding to their EF-hand domains.
ROS have been for long time considered as toxic byproducts of the chemical utilization of oxygen within the cells and oxidative stress has been linked to the pathogenesis of many disorders (Sies, 1991; Cuda et al., 2002; Sabbatini et al., 2006; Ruocco et al., 2007; Bertoni et al., 2011). It was the discovery of NOX enzymes, that deliberately produce ROS, to highlight the physiological role of ROS (Santillo et al., 2001; Schieber and Chandel, 2014). Many redox-dependent biological processes are controlled through the fine regulation of ROS-producing systems and antioxidant enzymes like glutathione peroxidase, catalase and superoxide dismutases (Mondola et al., 2004; Santillo et al., 2007; Secondo et al., 2008; Cassano et al., 2010; Damiano et al., 2013; Terrazzano et al., 2014).
Multiple physiological functions have been so far attributed to NOX enzymes. DUOX1 and 2 enzymes, first discovered in thyroid with a role in thyroid hormones synthesis (De Deken et al., 2000), are involved also in the innate immunity and cell signaling (Harper et al., 2005; der Vliet, 2008; Bae et al., 2010). NOXs-generated ROS modulate several physiological processes such as cell growth and differentiation or mucin expression and secretion (Ris-Stalpers, 2006; Chan et al., 2009; Damiano et al., 2015).
NOXs are activated by growth factor receptors such as platelet-derived growth factor receptor (Svegliati et al., 2005; Baroni et al., 2006; Gabrielli et al., 2008; Damiano et al., 2012), epidermal growth factor receptor (Damiano et al., 2015) cholinergic receptors (Seru et al., 2004) and many others (Petry et al., 2010). NOX-dependent ROS, in turn, regulate phosphorylation levels of multiple proteins modulated by redox-sensitive kinases and/or protein phosphatase (Brown and Griendling, 2009).
An interesting feature of NOXs biology is that ROS generated by NOXs can act downstream on other ROS generating systems (Lassègue et al., 2012). For example, in endothelial cells, oxidative stress associated with oscillatory shear stress is mediated by NOX-dependent xanthine oxidase activation (McNally et al., 2003). Moreover, there are evidences of a cross talk among NOX homologs. As an example, platelet-derived growth factor induces DUOX1-2 levels in human neuroblastoma cells through NOX2-derived ROS (Damiano et al., 2012). Therefore, the activation of NOXs results in a ROS-induced ROS release that can lead to oxidative stress and associated diseases. In human coronary arteries a correlation between NOXs mRNA expression and severity of atherosclerotic lesions has been shown (Sorescu et al., 2002).
NOXs are expressed in cardiomyocytes and in all cells of the vascular wall including endothelial cells (ECs), vascular smooth muscle cells (VSMCs), and adventitial fibroblasts.
ECs express NOX1, NOX2, NOX4, and NOX5 (Jones et al., 1996; Damico et al., 2012). They regulate cell differentiation, proliferation, migration, angiogenesis and vascular tone. NOX2 has been clearly linked to the reduced bioavailability of endothelium-derived NO (Görlach et al., 2000); NOX1 mediates cell growth, while NOX4 growth suppression, probably depending on their different subcellular localization or agonist stimulation. However, the specific role of the different NOX homologs should be better clarified. NOXs in ECs are in part localized at plasma membrane producing extracellular superoxide with a paracrine function (Barbacanne et al., 2000), and in part preassembled in intracellular compartments also with a perinuclear distribution (Li and Shah, 2002). Their functions are crucial for of ECs activation by different stimuli that rely on redox sensitive signaling molecules. Among the main targets of NOX-derived ROS in ECs there are the transcriptional factors NF-kB, Activated protein-1, hypoxia-inducible factor-1 or p53 that regulate gene expression. Endothelial NOXs also activates signaling cascades acting on protein kinases, (p38 and c-Jun N-terminal kinase, protein kinase B and Src), and/or protein phosphatases, including protein tyrosine phosphatase (Damico et al., 2012).
NOX2 is the main isoform of VSMCs of resistance arteries, while its expression is very low in VSMCs of large vessels (Lassegue and Clempus, 2003). NOX1 and NOX4 are expressed in VSMCs of large arteries with distinct subcellular localization and functions. NOX1, less abundant respect to NOX4, is localized at caveolae and mediates cell proliferation, while NOX4, that induces cell differentiation, is mainly localized at focal adhesions, the sites of tyrosine kinase signaling (Hilenski et al., 2004). Human aortic VSMCs also express NOX5 (Jay et al., 2008). In addition to NOX2, resistance arteries express NOX4, while the expression of NOX1 has not been so far clearly demonstrated.
NOX2 and NOX4 are the most abundantly expressed isoforms in cardiomyocytes. NOX2 is localized in plasma membrane and is modulated by stretch or Ang II (Byrne et al., 2003) (see NOXs in the cardiomyocyte force development). NOX4, involved in cell differentiation (Murray et al., 2013), is constitutively active and is mainly localized in intracellular compartments (Zhang et al., 2010).
Ang II, the major bioactive peptide of the Renin-Angiotensin System (RAS), is involved in many vascular processes including vasoconstriction, fibrosis, hypertrophy, inflammation, and aging (Mehta and Griendling, 2007). These effects are mediated by the interaction of Ang II with AT-1 receptor, while AT-2 receptor activation results in opposite vasodilatatory and antiproliferative effects. Vascular NOXs, including NOX1, NOX2, NOX4, and NOX5, are all regulated by Ang II (Nguyen Dinh Cat et al., 2013) that promotes an increase in blood pressure. The Ang II effects are mainly mediated by NOX-derived ROS signaling (Montezano et al., 2014). In addition to a direct effect on VSMCs, Ang II strengthens the sympathetic vasoconstriction increasing the synthesis and the release of the neurotransmitter at adrenergic varicosities. It has been also demonstrated that NOX-dependent ROS, in turn, activate AT-1 receptor thereby potentiating cell signaling with an auto amplificatory effect (Nishida et al., 2011).
Ang II induces activation of the enzymatic activity and increases expression of NOXs both in cultured VSMCs and intact arteries (Virdis et al., 2004). ROS produced thereby, activate multiple redox sensitive molecule including mitogen-activated protein, non-receptor tyrosine kinases, protein tyrosine phosphatases, calcium channels, and redox-sensitive transcription factors. In addition, Ang II activates tyrosine kinase receptors by transactivation (Cruzado et al., 2005; Li et al., 2010). Activation of these signaling pathways modulates different cellular processes in VSMCs, including contraction that relies on an increase of intracellular calcium levels and on the activation of the RhoA/Rho kinase pathway (Touyz and Schiffrin, 2000; Touyz et al., 2005), leading to an increase of myosin light chain phosphorylation. AT-1 receptor signaling activated by Ang II have an hypertrophic and fibrotic effect on the cardiac cells, mediated in part by endothelin-1 (ET-1) release (Weng et al., 2015). Moreover, Ang II redox signaling in cardiomyocytes is also involved in the Anrep effect (see NOXs in the cardiomyocyte force development). For the role of NOX-dependent Ang II signaling in the endothelium see NOXs in the endothelium-dependent relaxation.
NOX signaling have a role in endothelium dependent vasorelaxation that is mainly mediated by nitric oxide (NO) generated by endothelial nitric oxide synthase (eNOS). The liposolubile NO diffuses across the membranes reaching VSMCs, where it increases cGMP levels by activating the soluble guanylate cyclase; the subsequent activation of cGMP-dependent kinases leads to a decrease of intracellular calcium levels and relaxation. Superoxide anions produced by NOXs react with NO to produce peroxynitrite (Beckman et al., 1990; Görlach et al., 2000), leading to reduced bioavailability of NO and vasoconstriction. In addition, it has been shown that in aortas of mice with deoxycorticosterone acetate–salt (DOCA-salt) hypertension, ROS produced by NOXs oxidize the eNOS cofactor tetrahydrobiopterin, leading to the uncoupling of eNOS that produces superoxide rather than NO (Landmesser et al., 2003). This mechanism can cooperate with the scavenging effects of superoxide ions, leading to a reduced NO levels and impairment of endothelium-dependent vasorelaxation. This evidence has been also demonstrated in vivo using NOX1 overexpressing mice. In these animals subjected to Ang II induced hypertension, endothelium-dependent relaxation was impaired and bioavailable NO was markedly decreased (Dikalova et al., 2010). In ECs, Ang II activates all NOXs expressed in these cells including NOX5. Ang II-dependent extracellular signaling regulated kinase1/2 activation that mediate growth and inflammation, relies on NOX5 superoxide production. However, unlike the other NOX homologs, NOX5 overexpression increases eNOS activity even if, due to its NO antagonistic action, the overall effect is an impairing of endothelium-dependent relaxation, similarly to the other NOX homologs (Zhang et al., 2008).
NOX-generating ROS contribute to the positive inotropic response to mechanical stretch in cardiomyocytes. Physiological stretch triggers a microtubule-mediated activation of NOX2 localized at t-tubule membranes. This mechanism referred by Prosser et al. (2011) as X-ROS signaling, produces ROS that can diffuse across the membrane to oxidize the RyR2 Ca2+ release channels, located at junctional sarcoplasmic reticulum (J-SR) close to NOX2. Then, ryanodine receptors-2 activation leads to an increase of local cytosolic Ca2+ concentration and force development (Prosser et al., 2011, 2013; Sag et al., 2013). It has been also demonstrated that a cycling cardiomyocyte stretch in vitro results in an increase of ROS levels correlated with the amplitude and the frequency of stretch (Prosser et al., 2013). This mechanism could be of relevant physiological significance in vivo during the normal cyclic stretching and shortening of cardiomyocytes at each heartbeat, where the Ca2+ spark can be dynamically modulated by ROS in dependence of pre-load and heart frequency.
NOX2 is also involved in the slow enhanced increase in intracellular Ca2+ concentration and myocardial contractility due to mechanical stretch, known as Anrep effect. This slow response follows within 1–2 min an increase of the afterload reaching a maximum after 10–15 min. In this case, NOX2-derived ROS mediates Ang II dependent ET-1 release. In cardiomyocytes, Ang II released by mechanical stretch (Sadoshima et al., 1993) induces NOX2 activating auto-AT1 receptors, and induces ET-1 release (Ito et al., 1993). ET-1 signaling activates Na+/H+ exchanger-1 (Akram et al., 2006), that results in an increase of intracellular Na+, inhibition of Na+/Ca2+ exchanger, increase of cytosolic calcium concentration and contraction.
The role of NOXs in cardiomyocytes are not limited to mechanosensing. ROS produced by NOXs and by other sources such as mitochondria, are able to modulate different kinases phosphorylating proteins involved in calcium signaling; sarco/endoplasmic reticulum Ca2+-ATPase, plasma membrane Ca2+ ATPase, L-Type Ca2+ channels and Nav are some examples of molecular target of ROS leading to modulation of intracellular Ca2+ levels linked to myocyte contractility (Sag et al., 2013).
The role of ROS in hypertension has been well documented (Lee and Griendling, 2008; Sirker et al., 2011).
Numerous studies using ROS scavengers or more specific NOX inhibitors, were aimed at evaluating the role of NOXs in the elevation of blood pressure in hypertensive animals (Lassègue et al., 2012). More recently, the involvement of different NOX homologs and of their regulatory subunits in the pathogenesis of hypertension have been investigated using transgenic knockout and overexpressing animal models. These studies evidenced that NOX homologs exert different effects on hypertension.
NOX2 elevation is correlated with hypertension. Indeed, in transgenic mice with endothelial-specific overexpression of NOX2, Ang II causes a greater increase in ROS production and attenuated acetylcholine-induced vasorelaxation compared to wild-type (Murdoch et al., 2011). On the other hand, NOX2 knockout mice show baseline and Ang II-induced blood pressure values significantly lower than that of wild-type animals, even if the increase in blood pressure related to baseline is comparable in the two strains (Wang et al., 2001). Similar results were obtained in p47phox knockout animals in which the lack of Ang II hypertensive response was associated with a strong decrease of Ang II-dependent superoxide production in ECs and VSMCs (Landmesser et al., 2002). Unlike NOX2, cardiomyocyte-targeted NOX4 have protective effects facilitating cardiac adaptation to chronic cardiac pressure overload (Zhang et al., 2010; Schröder et al., 2012). Also endothelial NOX4 exerts beneficial effects. Transgenic mice with endothelium-targeted NOX4 overexpression show enhanced acetylcholine- or histamine-induced vasodilatation than wild-type animals. It is noteworthy to remind that NOX4 produces hydrogen peroxide instead of superoxide so preserving NO bioavailability (Ray et al., 2011). Moreover, when in hypertension or atherosclerosis eNOS was uncoupled to produce superoxide rather than NO (Landmesser et al., 2003), endothelial NOX4-derived H2O2 could mediate compensatory relaxation acting as an endothelium-derived hyperpolarizing factor (Yada et al., 2003).
Human studies aimed at linking NOX dysfunction with hypertension, have shown that some polymorphisms in the gene encoding p22 phox and affecting enzymatic activity, are associated with hypertension (Zalba et al., 2005). Moreover, it has been demonstrated that in human arteries Ang II increases superoxide levels and that this effect, mediated by NOX, is inhibited by AT1 receptor antagonists (Berry et al., 2000). However, results with AT1 antagonists are more convincing in patients with coronary artery disease (Hornig et al., 2001) rather than with hypertensive subjects (Ghiadoni et al., 2000).
Finally, another interesting aspect of NOX involvement in blood pressure homeostasis impairment is related to cigarette smoke, a risk factor of hypertension. Cigarette smoke condensate exposures have been correlated with ROS production, downregulation of enzymatic antioxidant cellular systems and cell toxicity (Russo et al., 2011). NOX2 activation by cigarette smoke have been demonstrated in isolated blood vessels and cultured ECs and VSMCs, suggesting a role of NOX-derived ROS in endothelial dysfunction associated with hypertension (Kim et al., 2014).
NOXs are now being considered as target of pharmacological intervention in patients with hypertension (Cai et al., 2003). Homolog-specific NOX inhibitors have been recently developed. This class of drugs opens the possibility to affect ROS production without altering ROS beneficial effects. The peptide-based inhibitors like NOX2ds-TAT (Csányi et al., 2011) and the NOX1 targeting inhibitor, NOXA1ds (Ranayhossaini et al., 2013) are the most reliable in in vitro experiments. However, due to the low bio-availability, peptide inhibitors are not promising therapeutic tools.
A number of small molecule NOXs selective inhibitors have been developed. Among them there are NOX1 and NOX 4 selective inhibitors like GKT137831, GKT136901, and GKT901 (Takac et al., 2012). The first one is in a phase 2 trial in patients with diabetic nephropathy and is also subjected to a series of preclinical studies for its application in different disease including cardiovascular disease.
Multiple molecular mechanisms regulating blood pressure involve NOX signaling. Of great importance is the central role of NOXs in angiotensin signaling, in the availability of NO and in the cardiac mechanosensing. A general scheme of the main overall effects of NOX-mediated signaling in the cells of the cardiovascular system leading to blood pressure modulation are shown in Figure 2.
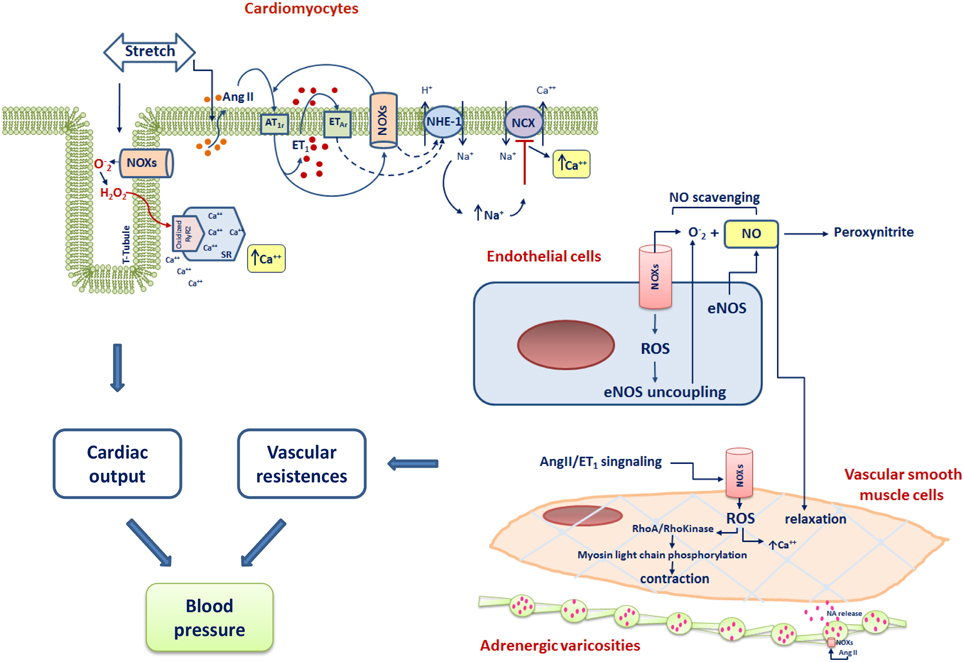
Figure 2. Scheme of the main NOXs-dependent cardiovascular mechanisms involved in the control of blood pressure. RyR2, Ryanodine Receptor type 2; ET-1, endothelin-1; NA, noradrenaline; AT1r, Angiotensin I type 1 receptor; ET1, endothelin 1; ETAR, Endothelin type A receptor; NHE-1, Na+/H+ exchanger-1; NCX, Na+/Ca2+ exchanger; e-NOS, endothelial nitric oxide synthase.
The current challenge in the NOX biology research field is represented by the better understanding of the mechanisms by which NOX isoforms exert their differential biological effects leading to the development of substances able to modulate specific redox-dependent cell functions.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Akram, S., Teong, H. F., Fliegel, L., Pervaiz, S., and Clement, M. V. (2006). Reactive oxygen species-mediated regulation of the Na+-H+ exchanger 1 gene expression connects intracellular redox status with cells sensitivity to death triggers. Cell Death Differ. 13, 628–641. doi: 10.1038/sj.cdd.4401775
Babior, B. M., Lambeth, J. D., and Nauseef, W. (2002). The neutrophil NADPH oxidase. Arch. Biochem. Biophys. 397, 342–344. doi: 10.1006/abbi.2001.2642
Bae, Y. S., Choi, M. K., and Lee, W. J. (2010). Dual oxidase in mucosal immunity and host-microbe homeostasis. Trends Immunol. 31, 278–287. doi: 10.1016/j.it.2010.05.003
Barbacanne, M. A., Souchard, J. P., Darblade, B., Liou, J. P., Nepveu, F., Ppy, B., et al. (2000). Detection of superoxide anion realeased extracellularly by endothelial cells using cytochrome c reduction, ESR, fluorescence and lucigenin-enhanced chemiluminescence technique. Free Radic. Biol. Med. 29, 388–396. doi: 10.1016/S0891-5849(00)00336-1
Baroni, S. S., Santillo, M., Bevilacqua, F., Luchetti, M., Spadoni, T., Mancini, M., et al. (2006). Stimulatory autoantibodies to the PDGF receptor in systemic sclerosis. N. Eng. J. Med. 354, 2667–2676. doi: 10.1056/NEJMoa052955
Beckman, J. S., Beckman, T. W., Chen, J., Marshall, P. A., and Freeman, B. A. (1990). Apparent hydroxyl radical production by peroxynitrite: implications for endothelial injury from nitric oxide and superoxide. Proc. Natl. Acad. Sci. U.S.A. 87, 1620–1624. doi: 10.1073/pnas.87.4.1620
Bedard, K., and Krause, K. H. (2007). The NOX family of ROS-generating NADPH oxidases: physiology and pathophysiology. Physiol. Rev. 87, 245–313. doi: 10.1152/physrev.00044.2005
Berry, C., Hamilton, C. A., Brosnan, M. J., Magill, F. G., Berg, G. A., McMurray, J. J. V., et al. (2000). Investigation into the sources of superoxide in human blood vessels: angiotensin II increases superoxide production in human internal mammary arteries. Circulation 101, 2206–2212. doi: 10.1161/01.CIR.101.18.2206
Bertoni, A., Giuliano, P., Galgani, M., Rotoli, D., Ulianich, L., Adornetto, A., et al. (2011). Early and late events induced by polyQ-expanded proteins: identification of a common pathogenic property of polYQ-expanded proteins. J. Biol. Chem. 286, 4727–4741. doi: 10.1074/jbc.M110.156521
Brown, D. I., and Griendling, K. K. (2009). Nox proteins in signal transduction. Free Radic. Biol. Med. 47, 1239–1253. doi: 10.1016/j.freeradbiomed.2009.07.023
Byrne, J. A., Grieve, D. J., Bendall, J. K., Li, J. M., Gove, C., Lambeth, J. D., et al. (2003). Contrasting roles of NADPH oxidase isoforms in pressure-overload versus angiotensin II-induced cardiac hypertrophy. Circ. Res. 93, 802–805. doi: 10.1161/01.RES.0000099504.30207.F5
Cai, H., Griendling, K. K., and Harrison, D. G. (2003). The vascular NAD(P)H oxidases as therapeutic targets in cardiovascular diseases. Trends Pharmacol. Sci. 24, 471–478. doi: 10.1016/S0165-6147(03)00233-5
Cassano, S., Agnese, S., D'amato, V., Papale, M., Garbi, C., Castagnola, P., et al. (2010). ROS, Ki-Ras and mitochondrial SOD co-operate in NGF-induced differentiation of PC12 cells. J. Biol. Chem. 285, 24141–24153. doi: 10.1074/jbc.M109.098525
Chan, E. C., Jiang, F., Peshavariya, H. M., and Dusting, G. J. (2009). Regulation of cell proliferation by NADPH oxidase-mediated signaling: potential roles in tissue repair, regenerative medicine and tissue engineering. Pharmacol. Ther. 122, 97–108. doi: 10.1016/j.pharmthera.2009.02.005
Cruzado, M. C., Risler, N. R., Miatello, R. M., Yao, G., Schiffrin, E. L., and Touyz, R. M. (2005). Vascular smooth muscle cell NAD(P)H oxidase activity during the development of hypertension: effect of angiotensin II and role of insulinlike growth factor-1 receptor transactivation. Am. J. Hypertens. 18, 81–87. doi: 10.1016/j.amjhyper.2004.09.001
Csányi, G., Cifuentes-Pagano, E., Al Ghouleh, I., Ranayhossaini, D. J., Egaña, L., Lopes, L. R., et al. (2011). Nox2 B-loop peptide, Nox2ds, specifically inhibits the NADPH oxidase Nox2. Free Radic. Biol. Med. 51, 1116–1125. doi: 10.1016/j.freeradbiomed.2011.04.025
Cuda, G., Paternò, R., Ceravolo, R., Candigliota, M., Pernotti, N., Perticone, F., et al. (2002). Protection of human endothelial cells from oxidative stress: role of Ras-ERK1/2 signaling. Circulation 105, 968–974. doi: 10.1161/hc0802.104324
Damiano, S., Fusco, R., Morano, A., De Mizio, M., Paternò, R., De Rosa, A., et al. (2012). Reactive oxygen species regulate the levels of dual oxidase (duox1-2) in human neuroblastoma cells. PLoS ONE 7:e34405. doi: 10.1371/journal.pone.0034405
Damiano, S., Morano, A., Ucci, V., Accetta, R., Mondola, P., Paternò, R., et al. (2015). Dual oxidase 2 generated reactive oxygen species selectively mediate the induction of mucins by epidermal growth factor in enterocytes. Int. J. Biochem. Cell Biol. 60C, 8–18. doi: 10.1016/j.biocel.2014.12.014
Damiano, S., Petrozziello, T., Ucci, V., Amente, S., Santillo, M., and Mondola, P. (2013). Cu-Zn superoxide dismutase activates muscarinic acetylcholine M1 receptor pathway in neuroblastoma cells. Mol. Cell. Neurosci. 52, 31–37. doi: 10.1016/j.mcn.2012.11.001
Damico, R., Zulueta, J. J., and Hassoun, P. M. (2012). Pulmonary endothelial cell NOX. Am. J. Respir. Cell Mol. Biol. 47, 129–139. doi: 10.1165/rcmb.2010-0331RT
De Deken, X., Wang, D., Many, M. C., Costagliola, S., Libert, F., Vassart, G., et al. (2000). Cloning of two human thyroid cDNAs encoding new members of the NADPH oxidase family. J. Biol. Chem. 275, 23227–23233. doi: 10.1074/jbc.M000916200
der Vliet, A. (2008). NADPH oxidases in lung biology and pathology: host defence enzymes, and more. Free Radical. Biol. Med. 44, 938–955. doi: 10.1016/j.freeradbiomed.2007.11.016
Dikalova, A. E., Go'ngora, M. C., Harrison, D. G., Lambeth, J. D., Dikalov, S., and Griendling, K. K. (2010). Upregulation of Nox1 in vascular smooth muscle leads to impaired endothelium dependent relaxation via eNOS uncoupling. Am. J. Physiol. Heart Circ. Physiol. 299, H673–H679. doi: 10.1152/ajpheart.00242.2010
Dinauer, M. C., Orkin, S. H., Brown, R., Jesaitis, A. J., and Parkos, C. A. (1987). The glycoprotein encoded by the X-linked chronic granulomatous disease locus is a component of the neutrophil cytochrome b complex. Nature 327, 717–720. doi: 10.1038/327717a0
Donko, A., Péterfi, Z., Sum, A., Leto, T., and Geiszt, M. (2005). Dual oxidases. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 360, 2301–2308. doi: 10.1098/rstb.2005.1767
Gabrielli, A., Svegliati, S., Moroncini, G., Pomponio, G., Santillo, M., and Avvedimento, E. V. (2008). Oxidative stress and the pathogenesis of scleroderma: the murrell's hypothesis revisited. Semin. Immunopathol. 30, 329–337. doi: 10.1007/s00281-008-0125-4
Ghiadoni, L., Virdis, A., Magagna, A., Taddei, S., and Salvetti, A. (2000). Effect of the angiotensin II type 1 receptor blocker candesartan on endothelial function in patients with essential hypertension. Hypertension 35, 501–506. doi: 10.1161/01.HYP.35.1.501
Görlach, A., Brandes, R. P., Nguyen, K., Amidi, M., Dehghani, F., and Busse, R. (2000). A gp91phox containing NADPH oxidase selectively expressed in endothelial cells is a major source of oxygen radical generation in the arterial wall. Circ. Res. 87, 26–32. doi: 10.1161/01.RES.87.1.26
Harper, R. W., Xu, C., Eiserich, J. P., Chen, Y., Kao, C. Y., Thai, P., et al. (2005). Differential regulation of dual NADPH oxidases/peroxidases, Duox1 and Duox2, by Th1 and Th2 cytokines in respiratory tract epithelium. FEBS Lett. 579, 4911–4917. doi: 10.1016/j.febslet.2005.08.002
Hilenski, L. L., Clempus, R. E., Quinn, M. T., Lambeth, J. D., and Griendling, K. K. (2004). Distinct subcellular localizations of Nox1 and Nox4 in vascular smooth muscle cells. Arterioscler. Thromb. Vasc. Biol. 24, 677–683. doi: 10.1161/01.ATV.0000112024.13727.2c
Hornig, B., Landmesser, U., Kohler, C., Ahlersmann, D., Spiekermann, S., Christoph, A., et al. (2001). Comparative effect of ACE inhibition and angiotensin II type 1 receptor antagonism on bioavailability of nitric oxide in patients with coronary artery disease: role of superoxide dismutase. Circulation 103, 799–805. doi: 10.1161/01.CIR.103.6.799
Ito, H., Hirata, Y., Adachi, S., Tanaka, M., Tsujino, M., Koike, A., et al. (1993). Endothelin-1 is an autocrine/paracrine factor in the mechanism of angiotensin II-induced hypertrophy in cultured rat cardiomyocytes. J. Clin. Invest. 92, 398–403. doi: 10.1172/JCI116579
Jay, D. B., Papaharalambus, C. A., Seidel-Rogol, B., Dikalova, A. E., Lassegue, B., and Griendling, K. K. (2008). Nox5 mediates PDGF-induced proliferation in human aortic smooth muscle cells. Free Radic. Biol. Med. 45, 329–335. doi: 10.1016/j.freeradbiomed.2008.04.024
Jones, S. A., O'Donnell, V. B., Wood, J. D., Broughton, J. P., Hughes, E. J., and Jones, O. T. (1996). Expression of phagocyte NADPH oxidase components in human endothelial cells. Am. J. Physiol. 271, 1626–1634.
Kim, M., Han, C. H., and Lee, M. Y. (2014). NADPH oxidase and the cardiovascular toxicity associated with smoking. Toxicol. Res. 30, 149–157. doi: 10.5487/TR.2014.30.3.149
Knaus, U. G., Heyworth, P. G., Evans, T., Curnutte, J. T., and Bokoch, G. M. (1991). Regulation of phagocyte oxygen radical production by the GTP-binding protein Rac 2. Science 254, 1512–1515. doi: 10.1126/science.1660188
Lambeth, J. D. (2004). NOX enzymes and the biology of reactive oxygen. Nat. Rev. Immunol. 4, 181–189. doi: 10.1038/nri1312
Landmesser, U., Cai, H., Dikalov, S., McCann, L., Hwang, J., Jo, H., et al. (2002). Role of p47(phox) in vascular oxidative stress and hypertension caused by angiotensin II. Hypertension 40, 511–515. doi: 10.1161/01.HYP.0000032100.23772.98
Landmesser, U., Dikalov, S., Price, S. R., McCann, L., Fukai, T., Holland, S. M., et al. (2003). Oxidation of tetrahydrobiopterin leads to uncoupling of endothelial cell nitric oxide synthase in hypertension. J. Clin. Invest. 111, 1201–1209. doi: 10.1172/JCI200314172
Lassegue, B., and Clempus, R. E. (2003). Vascular NAD(P)H oxidases: specific features, expression, and regulation. Am. J. Physiol. Regul. Integr. Comp. Physiol. 285, R277–R297. doi: 10.1152/ajpregu.00758.2002
Lassègue, B., San Martín, A., and Griendling, K. K. (2012). Biochemistry, physiology, and pathophysiology of NADPH oxidases in the cardiovascular system. Circ. Res. 110, 1364–1390. doi: 10.1161/CIRCRESAHA.111.243972
Lee, M. Y., and Griendling, K. K. (2008). Redox signaling, vascular function, and hypertension. Antioxid. Redox Signal. 10, 1045–1059. doi: 10.1089/ars.2007.1986
Li, J. M., and Shah, A. M. (2002). Intracellular localization and preassembly of the NADPH oxidase complex in cultured endothelial cells. J. Biol. Chem. 277, 19952–19960. doi: 10.1074/jbc.M110073200
Li, Y., Le'vesque, L. O., and Anand-Srivastava, M. B. (2010). Epidermal growth factor receptor transactivation by endogenous vasoactive peptides contributes to hyperproliferation of vascular smooth muscle cells of SHR. Am. J. Physiol. Heart Circ. Physiol. 299, H1959–H1967. doi: 10.1152/ajpheart.00526.2010
Martin, K. D., Frederick, L. M., von Loehneysen, K., Dinauer, M. C., and Knaus, U. G. (2006). Functional analysis of nox4 reveals unique characteristics compared to other NADPH oxidases. Cell. Signal. 18, 69–82. doi: 10.1016/j.cellsig.2005.03.023
McNally, J. S., Davis, M. E., Giddens, D. P., Saha, A., Hwang, J., Dikalov, S., et al. (2003). Role of xanthine oxidoreductase and NAD(P)H oxidase in endothelial superoxide production in response to oscillatory shear stress. Am. J. Physiol. Heart Circ. Physiol. 285, H2290–H22977. doi: 10.1152/ajpheart.00515.2003
Mehta, P. K., and Griendling, K. K. (2007). Angiotensin II cell signaling: physiological and pathological effects in the cardiovascular system. Am. J. Physiol. Cell Physiol. 292, C82–C97. doi: 10.1152/ajpcell.00287.2006
Mondola, P., Santillo, M., Serù, R., Damiano, S., Alvino, C., Ruggiero, G., et al. (2004). Cu, Zn superoxide dismutase increases intracellular calcium levels via a phospholipase C-protein kinase C pathway in SK-N-BE neuroblastoma cells. Biochem. Biophys. Res. Commun. 324, 887–892. doi: 10.1016/j.bbrc.2004.09.131
Montezano, A. C., Nguyen Dinh Cat, A., Rios, F. J., and Touyz, R. M. (2014). Angiotensin II and vascular injury. Curr. Hypertens. Rep. 16, 431. doi: 10.1007/s11906-014-0431-2
Murdoch, C. E., Alom-Ruiz, S. P., Wang, M., Zhang, M., Walker, S., Yu, B., et al. (2011). Role of endothelial NOX2-NADPHoxidase in angiotensin II-induced hypertension and vasomotor dysfunction. Basic Res. Cardiol. 106, 527–538. doi: 10.1007/s00395-011-0179-7
Murray, T. V., Smyrnias, I., Shah, A. M., and Brewer, A. C. (2013). NADPH oxidase 4 regulates cardiomyocyte differentiation via redox activation of c-Jun protein and the cis-regulation of GATA-4 gene transcription. J. Biol. Chem. 288, 15745–15759. doi: 10.1074/jbc.M112.439844
Nguyen Dinh Cat, A., Montezano, A. C., Burger, D., and Touyz, R. M. (2013). Angiotensin II, NADPH oxidase, and redox signaling in the vasculature. Antioxid. Redox Signal. 19, 1110–1120. doi: 10.1089/ars.2012.4641
Nishida, M., Kitajima, N., Saiki, S., Nakaya, M., and Kurose, H. (2011). Regulation of angiotensin II receptor signaling by cysteine modification of NF-jB. Nitric Oxide 25, 112–117. doi: 10.1016/j.niox.2010.10.003
Opie, L. H. (2004). Heart Physiology. From Cell to Circulation. 4th Edn. Philadelphia: Lippincott Williams and Wilkins.
Petry, A., Weitnauer, M., and Görlach, A. (2010). Receptor activation of NADPH oxidases. Antioxid. Redox Signal. 13, 467–487. doi: 10.1089/ars.2009.3026
Prosser, B. L., Ward, C. W., and Lederer, W. J. (2011). X-ROS signaling: rapid mechano-chemo transduction in heart. Science 333, 1440–1445. doi: 10.1126/science.1202768
Prosser, B. L., Ward, C. W., and Lederer, W. J. (2013). X-ROS signalling is enhanced and graded by cyclic cardiomyocyte stretch. Cardiovasc. Res. 98, 307–314. doi: 10.1093/cvr/cvt066
Ranayhossaini, D. J., Rodriguez, A. I., Sahoo, S., Chen, B. B., Mallampalli, R. K., Kelley, E. E., et al. (2013). Selective recapitulation of conserved and non conserved regions of putative NOXA1 protein activation domain confers isoform-specific inhibition of Nox1 oxidase and attenuation of endothelial cell migration. J. Biol. Chem. 288, 36437–36450. doi: 10.1074/jbc.M113.521344
Raven, P. B., and Chapleau, M. W. (2014). Blood pressure regulation XI: overview and future research directions Eur. J. Appl. Physiol. 114, 579–586. doi: 10.1007/s00421-014-2823-z
Ray, R., Murdoch, C. E., Wang, M., Santos, C. X., Zhang, M., Alom-Ruiz, S., et al. (2011). Endothelial Nox4 NADPH oxidase enhances vasodilatation and reduces blood pressure in vivo. Arterioscler. Thromb. Vasc. Biol. 31, 1368–1376. doi: 10.1161/ATVBAHA.110.219238
Ris-Stalpers, C. (2006). Physiology and pathophysiology of the DUOXes. Antioxid. Redox Signal. 8, 1563–1572. doi: 10.1089/ars.2006.8.1563
Ruocco, A., Santillo, M., Cicale, M., Serù, R., Cuda, G., Anrather, J., et al. (2007). Farnesyl transferase inhibitors induce neuroprotection by inhibiting Ha-Ras signalling pathway. Eur. J. Neurosci. 26, 3261–3266. doi: 10.1111/j.1460-9568.2007.05935.x
Russo, M., Cocco, S., Secondo, A., Adornetto, A., Bassi, A., Nunziata, A., et al. (2011). Cigarette smoke condensate causes a decrease of the gene expression of Cu-Zn superoxide dismutase, Mn superoxide dismutase, glutathione peroxidase, catalase, and free radical-induced cell injury in SH-SY5Y human neuroblastoma cells. Neurotox. Res. 19, 49–54. doi: 10.1007/s12640-009-9138-6
Sabbatini, M., Santillo, M., Pisani, A., Paternò, R., Uccello, F., Serù, R., et al. (2006). Inhibition of Ras/ERK1/2 signaling protects against postischemic renal injury. Am. J. Physiol. Renal Physiol. 290, F1408–F1415. doi: 10.1152/ajprenal.00304.2005
Sadoshima, J., Xu, Y., Slayter, H. S., and Izumo, S. (1993). Autocrine release of angiotensin II mediates stretch-induced hypertrophy of cardiac myocytes in vitro. Cell 75, 977–984. doi: 10.1016/0092-8674(93)90541-W
Sag, C. M., Wagner, S., and Maier, L. S. (2013). Role of oxidants on calcium and sodium movement in healthy and diseased cardiacmyocytes. Free Radical. Biol. Med. 63, 338–349. doi: 10.1016/j.freeradbiomed.2013.05.035
Santillo, M., Mondola, P., Serù, R., Annella, T., Cassano, S., Ciullo, I., et al. (2001). Opposing functions of Ki- and Ha-Ras genes in the regulation of redox signals. Curr. Biol. 11, 614–619. doi: 10.1016/S0960-9822(01)00159-2
Santillo, M., Secondo, A., Serù, R., Damiano, S., Garbi, C., Taverna, E., et al. (2007). Evidence of calcium- and SNARE-dependent release of CuZn superoxide dismutase from rat pituitary GH3 cells and synaptosomes in response to depolarization. J. Neurochem. 102, 679–685. doi: 10.1111/j.1471-4159.2007.04538.x
Schieber, M., and Chandel, N. S. (2014). ROS function in redox signaling and oxidative stress. Curr. Biol. 24, R453–R462. doi: 10.1016/j.cub.2014.03.034
Schröder, K., Zhang, M., Benkhoff, S., Mieth, A., Pliquett, R., Kosowski, J., et al. (2012). NOX4 is a protective reactive oxygen species generating vascular NADPH oxidase. Circ. Res. 110, 1217–1225. doi: 10.1161/CIRCRESAHA.112.267054
Secondo, A., De Mizio, M., Zirpoli, L., Santillo, M., and Mondola, P. (2008). The Cu-Zn superoxide dismutase (SOD1) inhibits ERK phosphorylation by muscarinic receptor modulation in rat pituitary GH3 cells. Biochem. Biophys. Res. Commun. 376, 143–147. doi: 10.1016/j.bbrc.2008.08.110
Segal, A. W., and Jones, O. T. (1978). Novel cytochrome b system in phagocytic vacuoles of human granulocytes. Nature 276, 515–517. doi: 10.1038/276515a0
Seru, R., Mondola, P., Damiano, S., Svegliati, S., Agnese, S., Avvedimento, E. V., et al. (2004). HaRas activates the NADPH oxidase complex in human neuroblastoma cells via extracellular signal-regulated kinase 1/2 pathway. J. Neurochem. 91, 613–622. doi: 10.1111/j.1471-4159.2004.02754.x
Sies, H. (1991). Oxidative stress: from basic research to clinical application. Am. J. Med. 91, 31S–38S. doi: 10.1016/0002-9343(91)90281-2
Sirker, A., Zhang, M., and Shah, A. M. (2011). NADPH oxidases in cardiovascular disease: insights from in vivo models and clinical studies. Basic Res. Cardiol. 106, 735–747. doi: 10.1007/s00395-011-0190-z
Sorescu, D., Weiss, D., Lassègue, B., Clempus, R. E., Szöcs, K., Sorescu, G. P., et al. (2002). Superoxide production and expression of nox family proteins in human atherosclerosis. Circulation 105, 1429–1435. doi: 10.1161/01.CIR.0000012917.74432.66
Svegliati, S., Cancello, R., Sambo, P., Luchetti, M., Paroncini, P., Orlandini, G., et al. (2005). PDGF and reactive oxygen species (ROS) regulate ras protein levels in primary human fibroblasts via ERK1/2: amplification of ROS and ras in systemic sclerosis fibroblasts. J. Biol. Chem. 280, 36474–36482. doi: 10.1074/jbc.M502851200
Takac, I., Schröder, K., and Brandes, R. P. (2012). The nox family of NADPH oxidases: friend or foe of the vascular system? Curr. Hypertens. Rep. 14, 70–78. doi: 10.1007/s11906-011-0238-3
Terrazzano, G., Rubino, V., Damiano, S., Sasso, A., Petrozziello, T., Ucci, V., et al. (2014). T cell activation induces CuZn superoxide dismutase (SOD)-1 intracellular re-localization, production and secretion. Biochim. Biophys. Acta. 1843, 265–274. doi: 10.1016/j.bbamcr.2013.10.020
Touyz, R. M., and Schiffrin, E. L. (2000). Signal transduction mechanisms mediating the physiological and pathophysiological actions of angiotensin II in vascular smooth muscle cells. Pharmacol. Rev. 52, 639–672.
Touyz, R. M., Yao, G., Quinn, M. T., Pagano, P. J., and Schiffrin, E. L. (2005). p47phox associates with the cytoskeleton through cortactin in human vascular smooth muscle cells: role in NAD(P)H oxidase regulation by angiotensin II. Arterioscler. Thromb. Vasc. Biol. 25, 512–518. doi: 10.1161/01.ATV.0000154141.66879.98
Virdis, A., Neves, M. F., Amiri, F., Touyz, R. M., and Schiffrin, E. L. (2004). Role of NAD(P)H oxidase on vascular alterations in angiotensin II-infused mice. J. Hypertens. 22, 535–542. doi: 10.1097/00004872-200403000-00016
Wang, H. D., Xu, S., Johns, D. G., Du, Y., Quinn, M. T., Cayatte, A. J., et al. (2001). Role of NADPH oxidase in the vascular hypertrophic and oxidative stress response to angiotensin II in mice. Circ. Res. 88, 947–953. doi: 10.1161/hh0901.089987
Weng, X., Yu, L., Liang, P., Li, L., Dai, X., Zhou, B., et al. (2015). A crosstalk between chromatin remodeling and histone H3K4 methyltransferase complexes in endothelial cells regulates angiotensin II induced cardiac hypertrophy. J. Mol. Cell. Cardiol. 82, 48–58. doi: 10.1016/j.yjmcc.2015.02.010
Yada, T., Shimokawa, H., Hiramatsu, O., Kajita, T., Shigeto, F., Goto, M., et al. (2003). Hydrogen peroxide, an endogenous endothelium-derived hyperpolarizing factor, plays an important role in coronary autoregulation in vivo. Circulation 107, 1040–1045. doi: 10.1161/01.CIR.0000050145.25589.65
Zalba, G., San José, G., Moreno, U. M., Fortuno, A., and Diez, J. (2005). NADPH oxidase-mediated oxidative stress: genetic studies of the p22phox gene in hypertension Antioxid. Redox Signal. 7, 1327–1336. doi: 10.1089/ars.2005.7.1327
Zhang, M., Brewer, A. C., Schröder, K., Santos, C. X., Grieve, D. J., Wang, M., et al. (2010). NADPH oxidase-4 mediates protection against chronic load-induced stress in mouse hearts by enhancing angiogenesis. Proc. Natl. Acad. Sci. U.S.A. 107, 18121–18126. doi: 10.1073/pnas.1009700107
Keywords: NOX, blood pressure, reactive oxygen species, redox signaling, cardiomyocytes, endothelial cells, vascular smooth muscle cells, Ang II signaling
Citation: Santillo M, Colantuoni A, Mondola P, Guida B and Damiano S (2015) NOX signaling in molecular cardiovascular mechanisms involved in the blood pressure homeostasis. Front. Physiol. 6:194. doi: 10.3389/fphys.2015.00194
Received: 02 April 2015; Accepted: 22 June 2015;
Published: 07 July 2015.
Edited by:
Lacolley Patrick, Institut National de la Santé et de la Recherche Médicale, FranceReviewed by:
Zsolt Bagi, Georgia Regents University, USACopyright © 2015 Santillo, Colantuoni, Mondola, Guida and Damiano. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Mariarosaria Santillo, Dipartimento di Medicina Clinica e Chirurgia, Università di Napoli “Federico II,” Via S. Pansini, 5, 80131 Naples, Italy,bWFyc2FudGlAdW5pbmEuaXQ=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.