 Yves Lecarpentier
Yves Lecarpentier Victor Claes2
Victor Claes2- 1Centre de Recherche Clinique, Centre Hospitalier Régional de Meaux, Meaux, France
- 2Department of Pharmaceutical Sciences, University of Antwerp, Wilrijk, Belgium
- 3Institut de Cardiologie, Hôpital de la Pitié-Salpêtière, Paris, France
Circadian clock mechanisms are far-from-equilibrium dissipative structures. Peroxisome proliferator-activated receptors (PPAR alpha, beta/delta, and gamma) play a key role in metabolic regulatory processes, particularly in heart muscle. Links between circadian rhythms (CRs) and PPARs have been established. Mammalian CRs involve at least two critical transcription factors, CLOCK and BMAL1 (Gekakis et al., 1998; Hogenesch et al., 1998). PPAR gamma plays a major role in both glucose and lipid metabolisms and presents circadian properties which coordinate the interplay between metabolism and CRs. PPAR gamma is a major component of the vascular clock. Vascular PPAR gamma is a peripheral regulator of cardiovascular rhythms controlling circadian variations in blood pressure and heart rate through BMAL1. We focused our review on diseases with abnormalities of CRs and with primary or secondary cardiac dysfunction. Moreover, these diseases presented changes in the Wnt/beta-catenin pathway and PPARs, according to two opposed profiles. Profile 1 was defined as follows: inactivation of the Wnt/beta-catenin pathway with increased expression of PPAR gamma. Profile 2 was defined as follows: activation of the Wnt/beta-catenin pathway with decreased expression of PPAR gamma. A typical profile 1 disease is arrhythmogenic right ventricular cardiomyopathy, a genetic cardiac disease which presents mutations of the desmosomal proteins and is mainly characterized by fatty acid accumulation in adult cardiomyocytes mainly in the right ventricle. The link between PPAR gamma dysfunction and desmosomal genetic mutations occurs via inactivation of the Wnt/beta-catenin pathway presenting oscillatory properties. A typical profile 2 disease is type 2 diabetes, with activation of the Wnt/beta-catenin pathway and decreased expression of PPAR gamma. CRs abnormalities are present in numerous pathologies such as cardiovascular diseases, sympathetic/parasympathetic dysfunction, hypertension, diabetes, neurodegenerative diseases, cancer which are often closely inter-related.
Introduction
CRs are biological temporal processes that display endogenous, entrainable free-running periods that last approximately 24 h. They are driven by molecular internal clocks which can be reset by environmental light-dark cycles. Circadian clocks are transcriptionally based molecular mechanisms which comprise feedback loops (Edery, 2000). The molecular basis of CRs have first been clarified in Drosophila and Neurospora, then in cyanobacteria, plants, and mammals (Reppert and Weaver, 2002). All living organisms adjust their physiology and behavior to the 24-h day-night cycle under the governance of circadian clocks. Circadian clocks may provide a selective advantage of anticipation, thus allowing organisms to respond efficiently to various stimuli at the appropriate time. In mammals, sleep-awake and feeding patterns, hormone secretion, heart rate, blood pressure, energy metabolism, and body temperature exhibit CRs. Their disruptions may have deleterious effects. People submitted to shift working, frequent transmeridian air flight, exposure to artificial light exhibit a particularly high incidence of metabolic syndrome and obesity. CR dysfunctions in blood pressure and heart rate, which are both partly regulated by PPAR gamma are involved in arrhythmias which may lead to sudden cardiac death, myocardial infarction or stroke, often occurring at the early morning during the surge in blood pressure. CRs are dissipative structures due to a negative feedback produced by a protein on the expression of its own gene (Goodwin, 1965; Hardin et al., 1990). They operate far-from- equilibrium and generate order spontaneously by exchanging energy with their external environment (Prigogine et al., 1974; Goldbeter, 2002; Lecarpentier et al., 2010).
The Regulatory Sites of Circadian Rhythms
The master regulator site of CRs is the suprachiasmatic nucleus (SCN) inside the hypothalamus in which core clock genes are rhythmically expressed (Weaver, 1998). In addition to this central clock, each organ has its own biological clock system, termed peripheral clock. The SCN and most peripheral tissues such as heart, blood vessels, skeletal muscles, kidneys, liver, and fat, govern numerous functions that are synchronized with the sleep-awake cycle (Zylka et al., 1998). In the cardiovascular system, circadian clocks have been described within numerous mammalian cells, such as cardiomyocytes, vascular smooth muscle cells, endothelial cells, and fibroblasts (McNamara et al., 2001; Nonaka et al., 2001; Durgan et al., 2005, 2007; Takeda et al., 2007). Peripheral clocks have their own regulatory mechanisms, which are specific to each peripheral organ by regulating the expression of clock-controlled genes (Ccg). CRs have been demonstrated in approximately 8–10% of total genes expressed in mouse heart and liver, more than 90% of them depending on self-autonomous local diurnal oscillators (Storch et al., 2002).
Genes and Proteins
Important genes are involved in CRs including Clock (Circadian locomotor output cycles kaput), Bmal1 (brain and muscle aryl-hydrocarbon receptor nuclear translocator-like 1), Cry1 (cryptochrome 1), Cry2 (cryptochrome 2), Per1 (Period 1), Per2 (Period 2), Per3 (Period 3), and Ccg. They organize transcription/translation autoregulatory feedback loops comprising both activating and inhibiting pathways (Reppert and Weaver, 2002; Schibler and Sassone-Corsi, 2002). A complex network is formed by all these genes which interlock feedback and forward subtle loops whose complete time course is approximately 24 h. Clock genes Per1, Per2, Bmal1, and Cry1 display rhythmic expression in human hearts (Leibetseder et al., 2009). At the start of the day, transcription of Clock and Bmal1 begins. The proteins CLOCK and BMAL1 are synthesized and then associate as dimers which bind to regulatory DNA sequences (E-box elements) of the promoters of target genes. CLOCK: BMAL1 dimer activates circadian gene transcription of Period genes (Per1, 2, and 3), Chryptochrome genes (Cry 1 and 2), Rev-erb, Ror (related orphan receptor), and Ccg, and drives their rhythmic expression (Reppert and Weaver, 2001, 2002; Young and Kay, 2001; Canaple et al., 2006; Chen and Yang, 2014). Into the cytoplasm, the PER and CRY proteins dimerize and, after translocation to the nucleus, modulate the transcriptional activity of CLOCK: BMAL1 (Kume et al., 1999). Concentrations of BMAL1 and PER proteins cycle in counterpoint. PER2 is a positive regulator of the Bmal1 loop. Protein CRY is a negative regulator of both Per and Cry loops. ROR alpha enhances Bmal1 transcription (Akashi and Takumi, 2005), while the nuclear receptor Rev-erb inhibits it (Ueda et al., 2002). Rev-erb alpha binds to ROR-responsive element (RORE) in the Bmal1 promoter and represses its transcriptional activity (Preitner et al., 2002). Rev-erb alpha protein is a member of the nuclear receptor family of intracellular transcription factors. The gene Rev-erb alpha is a major regulatory component of the circadian clock (Yin et al., 2006) and among various properties, is involved in the circadian expression of plasminogen activator inhibitor type 1 (Wang et al., 2006).
Circadian Rhythms and Mutations of Genes
Mutations or deletions of clock genes in mice have shown the key role of circadian clocks to ensure the proper timing of metabolic and cardiovascular processes. There is an increased pathological remodeling and vascular injury together with an aberrant CR in Bmal1-knockout and Clock mutant mice (Anea et al., 2009). Aortas from Bmal1-knockout and Clock mutant mice exhibit endothelial dysfunction. Akt (protein kinase B) and subsequent nitric oxide signaling is significantly attenuated in arteries from Bmal1-knockout mice. Bmal1 is a key regulator of myogenesis which may represent a temporal regulatory mechanism to fine-tune myocyte differentiation (Chatterjee et al., 2013). Bmal1 regulates adipogenesis via the Wnt signaling pathway (Guo et al., 2012). Disruption of Bmal1 in mice led to increased adipogenesis, adipocyte hypertrophy, and obesity. Attenuation of Bmal1 function resulted in down-regulation of genes in the canonical Wnt pathway known to suppress adipogenesis. Promoters of these genes, i.e., beta-catenin, Disheveled (Dsh), T cell-enhancing binding (Tcf) display Bmal1 occupancy, indicating direct circadian regulation by Bmal1. Among several abnormalities, deletion of the clock gene Bmal1 in mice adipose tissue induces obesity (Paschos et al., 2012). The cardiomyocyte-specific clock mutant (Ccm) is a mouse model wherein the cardiomyocyte circadian clock is selectively suppressed (Young et al., 2001b,c; Durgan et al., 2006). Ccm presents a temporal suspension of the cardiomyocyte circadian clock at the wake-to-sleep transition (Young, 2009). Numerous mutations of genes will be discussed in the following paragraphs of this review.
Circadian Rhythms and Heart Performance
Loss of synchronization between the internal clock and external stimuli can induce cardiovascular organ damage. Discrepancy in the phases between the central and peripheral clocks also seems to contribute to progression of cardiovascular disorders (Takeda and Maemura, 2011). Peripheral clocks have their own roles specific to each peripheral organ by regulating the expression of Ccg, although the oscillation mechanisms of the peripheral clock are similar to that of the SCN (Takeda et al., 2007). Both the physiological and pathological functions of cardiovascular organs are closely related to CRs. Heart rate, blood pressure and endothelial function show diurnal variations within a day. A profound pattern exists in the time of day at which the death may occur (Takeda and Maemura, 2011). The onset of cardiovascular disorders such as acute coronary syndrome, atrial arrhythmias, and subarachnoid hemorrhage exhibits impairment of diurnal oscillations. Stroke and heart attacks most frequently happen in the morning when blood pressure surges.
Over the course of the day, the normal heart anticipates, responds and adapts to physiological alterations within its environment. Contractile performance, carbohydrate oxidation, fatty acid oxidation (FAO), mitochondrial function, oxygen consumption, and expression of all metabolic genes show diurnal variations. The circadian clock plays an important role in cardiac homeostasis through the anticipation of daily workload. In wild-type mice, the ejection fraction (EF) and the shortening fraction (FS) show circadian variation (Wu et al., 2011). The diurnal variations in EF and FS are altered in mice with disruptions of circadian clock genes and are significantly diminished under an imposed light regimen. The circadian variation in blood pressure and heart rate is disrupted in Bmal1(-/-) and Clock (mut) mice in which core clock genes are deleted or mutated (Curtis et al., 2007). Bmal1 deletion abolishes the 24-h frequency in cardiovascular rhythms. However, a shorter ultradian rhythm remains. Sympathetic adrenal function is disrupted in these mice.
Peroxisome Proliferator-Activated Receptors (PPARs)
PPARs (alpha, beta/delta, and gamma) are nuclear receptors belonging to the nuclear receptor superfamily. They function as transcription factors within the cell nuclei and regulate the expression of several target genes. PPARs play a pivotal role in various physiological and pathological processes, especially in energy metabolism, development, carcinogenesis, extracellular matrix remodeling, and CRs (Lockyer et al., 2009). PPARs heterodimerize with the retinoid X receptor (RXR). PPARs are activated by their respective ligands, either endogenous fatty acids or pharmaceutical drugs which are potential therapeutic agents. Numerous natural and synthetic compounds, i.e., fatty acids, eicosanoids, arachidonic acid, hypolipidemic fibrates activating PPAR alpha, and anti diabetic thiazolidinediones (TZD) activating PPAR gamma, serve as activators of PPARs. PPARs are involved in numerous pathologies such as obesity, dyslipidemia, insulin resistance, type 2 diabetes, hypertension, cardiac hypertrophy (Berger and Moller, 2002; Kelly, 2003). PPAR beta/delta was not studied in this review.
PPARs and Circadian Rhythms
PPARs integrate the mammalian clock and energy metabolism (Chen and Yang, 2014). PPARs have been shown to be rhythmically expressed in mammalian tissues (Yang et al., 2006) and to directly interact with the core clock genes (Inoue et al., 2005). PPAR beta/delta has not been studied in this review.
PPAR alpha
PPAR alpha presents CRs in several organs, i.e. heart, muscles, liver, and kidney (Lemberger et al., 1996; Yang et al., 2006). PPAR alpha expression is stimulated by stress, glucocorticoid hormones, and insulin whose secretion follows CRs (Lemberger et al., 1994). Importantly, PPAR alpha is a direct target of genes (Bmal1 and Clock) through an E-box process (Oishi et al., 2005). The circadian expression of PPAR alpha mRNA is abolished in the liver of homozygous Clock mutant mice and is regulated by the peripheral oscillators in a CLOCK-dependent mechanism. In rodent liver, there is a regulatory feedback loop involving BMAL1 and PPAR alpha in peripheral clocks. This regulation occurs via a direct binding of PPAR alpha on a PPAR alpha response element located in the Bmal1 gene promoter. Moreover, BMAL1 is an upstream regulator of PPAR alpha gene expression (Gervois et al., 1999; Canaple et al., 2006). Several genes such as those encoding for sterol regulatory element binding protein, HMG-CoA reductase, fatty acid synthase, are involved in the lipid metabolism. They display circadian fluctuations, and their activities are diminished or suppressed in PPAR alpha knockout mice (Patel et al., 2001; Gibbons et al., 2002). PPAR alpha directly regulates the transcriptional activity of Bmal1 and Rev-erb alpha through the PPRE located in the promoter site of their respective genes. Per2 interacts with nuclear receptors including PPAR alpha and Rev-Erb alpha and serves as a co-regulator of nuclear receptor-mediated transcription. The PPAR alpha agonist fenofibrate increases transcription and resets circadian expression of Bmal1, Per2, and Rev-erb alpha in mouse liver (Canaple et al., 2006) and cultured hepatocytes (Gervois et al., 1999). Moreover, bezafibrate can phase advance the rhythmic expression of Bmal1, Per2, and Rev-erb alpha in several mouse peripheral tissues (Shirai et al., 2007; Oishi et al., 2008).
PPAR gamma
PPAR gamma exhibits variations in diurnal expression in mouse fat, liver, and blood vessels (Yang et al., 2006; Wang et al., 2008). Deletion of PPAR gamma in mouse suppresses or diminishes diurnal rhythms (Yang et al., 2012). CRs have been analyzed in two strains of whole-body PPAR gamma null mouse models, i.e., Mox2-Cre mice (MoxCre/flox) or induced by tamoxifen (EsrCre/flox/TM). Diurnal variations in blood pressure and heart rate are blunted in MoxCre/flox mice. Impaired rhythmicity of the canonical clock genes is observed in adipose tissue and liver. This shows the important role of PPAR gamma in the coordinated control of circadian clocks, metabolism, and cardiac performance (Yang et al., 2012). Moreover, insulin resistance is correlated with a non-dipper type—i.e., with no blood pressure decrease during the circadian cycle- in essential hypertension. TZD are oral hypoglycemic agents act as insulin sensitizers and possess antihypertensive properties. TZD therapy with pioglitazone transforms the CR of blood pressure from a non-dipper to a dipper type (Anan et al., 2007). PPAR gamma contributes to maintain the diurnal variations of both blood pressure and heart rate.
Rev-Erb alpha, an orphan nuclear receptor and a core clock component, is expressed after PPAR gamma activation with rosiglitazone in rat. Activated PPAR gamma induces Rev-Erb alpha promoter activity by binding to the response element Rev-DR2. Mutations of the 5′ or 3′ half-sites of the response element suppress PPAR gamma binding and transcriptional activation (Fontaine et al., 2003). PGC-1 alpha, a transcriptional co-activator that regulates energy metabolism, is rhythmically expressed in the liver and skeletal muscle of mice. PGC-1 alpha stimulates the expression of clock genes, notably Bmal1 and Rev-erb alpha, through co-activation of the ROR family of orphan nuclear receptors. Mice lacking PGC-1 alpha show abnormal CRs of activity, body temperature, and metabolic rate (Liu et al., 2007a). Nocturnin, a circadian-regulated gene, promotes adipogenesis by stimulating PPAR gamma nuclear translocation (Kawai et al., 2010). Nocturnin binds to PPAR gamma and stimulates its transcriptional activity whereas its deletion suppresses PPAR gamma oscillations (Green et al., 2007). The hormone-dependent interaction of the nuclear receptor RXR alpha with CLOCK negatively regulates CLOCK: BMAL1-mediated transcriptional activation of clock gene expression in vascular cells. RXR alpha can phase shift Per2 mRNA rhythmicity, providing a molecular mechanism for hormonal control of clock gene expression (McNamara et al., 2001).
Canonical Wnt/Beta-Catenin Pathway
Beta-catenin plays a key role during epithelial-mesenchymal transition (EMT), that characterizes normal embryonic development, tissue regeneration and cancer proliferation (Heuberger and Birchmeier, 2010). Beta-catenin is a normal constituent of the zonula adherens, a major cell-to-cell adhesion complex in pavement-like tissues. During EMT, the loss of cadherins disrupts the zonula adherens, thus liberating beta-catenin into the cytoplasm. This molecule then migrates to the cell nucleus where it activates the Wnt/beta-catenin target genes. A hallmark of the canonical Wnt pathway activation is the elevation of cytoplasmic beta-catenin protein levels, the subsequent nuclear translocation and further activation of beta-catenin specific gene transcription (Ben-Ze'ev and Geiger, 1998; Klymkowsky et al., 1999; Zhurinsky et al., 2000; Moon et al., 2002; Maeda et al., 2004; Sen-Chowdhry et al., 2005; Garcia-Gras et al., 2006), (Figure 1). In the absence of Wnt ligands, beta-catenin is recruited into a destruction complex that contains adenomatous polyposis coli (APC) and Axin, which facilitate the phosphorylation of beta-catenin by glycogen synthase kinase 3- beta (GSK3-beta). GSK3-beta phosphorylates the N-terminal domain of beta-catenin, thereby targeting it for ubiquitination and proteasomal degradation. In the presence of a Wnt ligand, the binding of Wnt to Frizzled (Fzd) leads to activation of the phosphoprotein Disheveled (Dsh). Dsh recruits Axin and the destruction complex to the plasma membrane, where Axin directly binds to the cytoplasmic tail of LRP5/6. Axin is degraded, which decreases beta-catenin degradation. The activation of Dsh also leads to the inhibition of GSK3-beta by phosphorylation, which further reduces the phosphorylation and degradation of beta-catenin. The beta-catenin degradation complex is inactivated with recruitment of axin to the plasma membrane, thus stabilizing the non-phosphorylated beta-catenin which translocates to the nucleus. Beta-catenin binds to T cell/lymphoid-enhancing binding (Tcf/Lef) transcription factors. The resulting complex becomes active by displacing Grouchos, leading to activation of numerous target genes.
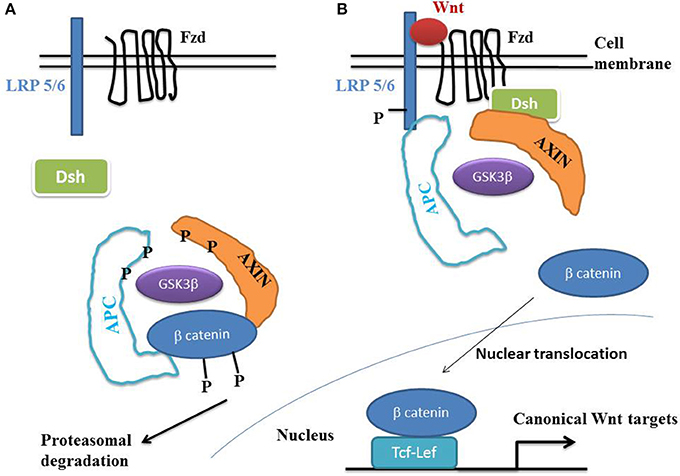
Figure 1. The Wnt/beta-catenin pathway. (A) In the absence of Wnt, cytosolic beta-catenin is phosphorylated by GSK3 beta. APS and AXIN complex with GSK3 beta and beta-catenin to enhance the destruction process into the proteasome. Phosphorylated beta-catenin is recognized by the ubiquitin ligase beta -TrCP, ubiquinated and degraded. The Wnt pathway is in an “off state.” (B) In the presence of Wnt, Wnt binds both Frizzled and LRP5/6 receptors to initiate GRK5/6-mediated LRP phosphorylation and disheveled-mediated Frizzled internalization. Disheveled membrane translocation leads to dissociation of the AXIN/APC/GSK3 beta complex. Beta-catenin phosphorylation is inhibited and accumulates into the cytosol. Beta-catenin then translocates to the nucleus to bind Lef-Tcf co-transcription factors, which induces the Wnt-response gene transcription. Abbreviations: APC, adenomatous polyposis coli; Dsh, Disheveled; GSK3 beta, glycogen synthase kinase 3 beta; LRP5/6, low density lipoprotein receptor-related protein 5/6; Fzd, Frizzled.
Canonical Wnt/Beta-Catenin Pathway and PPAR Gamma
Numerous studies have shown the direct interaction between beta-catenin and PPAR gamma (Moldes et al., 2003; Jansson et al., 2005; Garcia-Gras et al., 2006). PPAR gamma activation inhibits the beta-catenin activation of Tcf/Lef transcription factors (Lu and Carson, 2010). The TZD PPAR gamma agonists troglitazone, rosiglitazone, and pioglitazone, and the non-TZD PPAR gamma activator GW1929 inhibit the beta-catenin-induced transcription in a PPAR gamma dependent manner. Activation of the Wnt-beta catenin pathway leads to osteogenesis, not adipogenesis and its inhibition leads to an increase in transcription of PPAR gamma. Osteogenic pathway is linked to the stimulation of Wnt signal leading to the final transcriptional activation of early osteogenic markers such as RUNX-2 and ALP, mediated by beta-catenin. Conversely, the adipogenic pathway involves inhibition of Wnt pathway leading to ubiquitination/degradation of beta-catenin which results in the transcription of PPAR gamma, a pivotal initiator of adipogenesis. The canonical Wnt/beta-catenin-PPAR gamma system determines the molecular switching of osteablastogenesis vs. adipogenesis (Takada et al., 2009). PPAR gamma is a prime inducer of adipogenesis that inhibits osteoblastogenesis. Two different pathways switch the cell fate decision from adipocytes to osteoblasts by suppressing the transactivation function of PPAR gamma. TNF-alpha- and IL-1-induced TAK1/TAB1/NIK signaling cascade attenuate PPAR gamma-mediated adipogenesis by inhibiting the binding of PPAR gamma to the DNA response element. PPAR gamma suppresses Wnt/beta-catenin signaling during adipogenesis (Moldes et al., 2003). Wnt/beta-catenin pathway operates to maintain the undifferentiated state of preadipocytes by inhibiting adipogenic gene expression. Importantly, there is a reciprocal relationship between beta-catenin expression and PPAR gamma activity.
Diseases Associated with Deactivation of the Wnt/Beta-Catenin Pathway and Increased Expression of PPAR Gamma
Numerous diseases present a common denominator: activation of the Wnt/beta-catenin pathway decreased and the expression of PPAR gamma increased. In most cases, expression of PPAR alpha decreases.
Arrhythmogenic right ventricular cardiomyopathy (ARVC)
ARVC is a rare human disease characterized by the development of a fibro-fatty tissue in both ventricles, prominently involving the right ventricular (RV) myocardium (Marcus et al., 1982; Fontaine et al., 1999) (Figure 2). Cardiac dysfunction progressively develops, initially located at the RV and becoming biventricular in about 20% of cases (Richardson et al., 1996; Hebert et al., 2004). ARVC is most often an autosomic family-related disease. Genetic mutations have been identified in about 50% of cases, occurring among the five desmosomal proteins so far identified in the ventricular cardiomyocyte, i.e., desmoglein 2 (DSG2), desmocollin 2 (DSC2), plakophilin 2 (PKP2), plakoglobin (PG), and desmoplakin (DSP) (Basso et al., 2009; Fressard et al., 2010). PPAR abnormalities have been reported in ARVC with an increase in PPAR gamma and a decrease in PPAR alpha in RV (Djouadi et al., 2009).
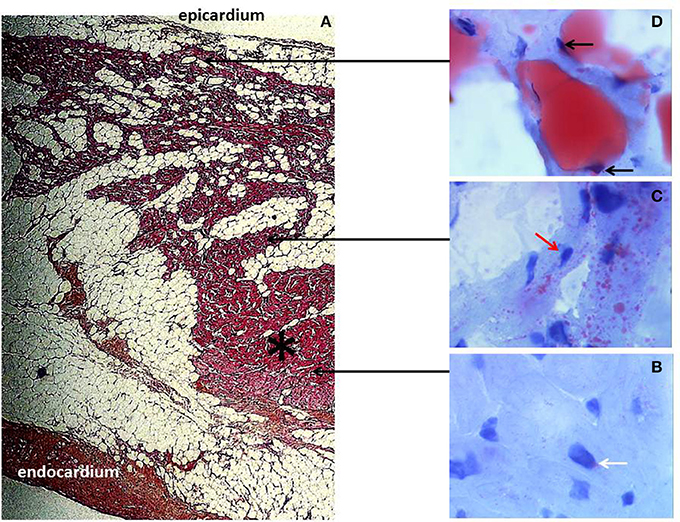
Figure 2. Arrhythmogenic right ventricular cardiomyopathy (ARVC) histology. (A) Typical morphology of right ventricular transmural free wall section in a terminal ARVC heart transplant specimen, showing extensive fibro-fatty replacement. A mid-mural residual muscular core (black asterisk) is well-identified. Fibrosis is prominently located at the subendocardium. Note the layer of normal subepicardial fat (Hematoxylin Eosin Saffron staining, original magnification × 10). (B–D) are fresh tissue snap frozen fragments representative of regions referred to as muscular (B,C) and fatty myocardium, (D) respectively, stained with oil red O (original magnification × 50). The red staining indicates neutral lipid accumulation. (B) Note the normal discrete perinuclear staining of the cardiomyocytes (white arrow) within the well-preserved myocardial core. (C) In contrast, there is an abnormal major accumulation of fatty droplets (not visible under standard staining) dispersed within the cells of the mid mural muscular zone located above the residual muscular core and surrounded by fatty tissue. Note the remaining normal central position of the nucleus within the myocardial cells (red arrow). (D) Finally, there is a direct transdifferentiation of myocardial cells into adipocytes within the upper muscular zone bordering the normal subepicardial fat. Take notice of the major confluence of the fatty droplets as well as of the final aspect of total fatty transformation with migration of the nucleus beneath the cell membrane (black arrows).
Molecular mechanisms underlying ARVC are now better understood. The link between PPAR gamma dysfunction and desmosomal genetic mutations implicates the Wnt/beta-catenin pathway. Thus, the suppression of canonical Wnt/beta-catenin signaling by nuclear PG recapitulates the phenotype of ARVD by exhibiting fat accumulation in cardiomyocytes, enhanced myocyte apoptosis, ventricular dysfunction, and ventricular arrhythmias in transgenic mice (Garcia-Gras et al., 2006). The desmosomal PG also known as gamma-catenin, has structural and functional similarities to beta-catenin, which is the effector for canonical Wnt signaling (Moon et al., 2002). PG interacts and competes with beta-catenin at multiple cellular levels with a net negative effect on the canonical Wnt/beta-catenin signaling pathway through Tcf/Lef transcription factors (Ben-Ze'ev and Geiger, 1998; Klymkowsky et al., 1999; Zhurinsky et al., 2000; Maeda et al., 2004). Mutating the desmosomal protein DSP by impairing desmosome assembly set free gamma-catenin from the desmosomes. As a consequence, gamma catenin translocates to the nucleus and after competition with beta-catenin suppresses signaling through the canonical Wnt/beta-catenin-Tcf/Lef pathway. Suppression of DSP expression responsible for human ARVC, leads to nuclear localization of PG and to suppression of canonical Wnt/beta-catenin-Tcf/Lef1 signaling in cultured atrial myocytes and in mouse hearts (Sen-Chowdhry et al., 2005). Tcf/Lef1 suppression induces a transcriptional switch from myogenesis to adipogenesis (Ross et al., 2000). This leads to enhanced adipogenesis, fibrogenesis, and myocyte apoptosis, thus summarizing the phenotype of human ARVC (Corrado et al., 1997).
Cardiac hypoxia
Hypoxia up-regulates expression of PPAR gamma angiopoietin-related gene in cardiomyocytes (Belanger et al., 2002). Hypoxia-inducible factor 1 alpha (HIF1 alpha) inhibits PPAR alpha expression during hypoxia (Narravula and Colgan, 2001). Hypoxia leads to activation of HIF1 alpha (Krishnan et al., 2009) and several genes involved in the regulation of glucose transporters, glycolytic enzyme, and pyruvate deshydrogenase kinase (PDK1). HIF1 alpha overexpression in vitro leads to triacylglycerol accumulation, and reduced FAO due to inhibition of PPAR alpha. Cardiac hypoxia represents a pathological state where expression of PPAR alpha is reduced whereas that of PPAR gamma is increased. Hypoxia triggers a cascade of cellular metabolic responses including a decrease in mitochondrial oxidative flux (Huss et al., 2001). Under hypoxic conditions, myocytes exhibit significant accumulation of intracellular neutral lipid consistent with reduced carnitine palmitoyltransferase-1 (CPT-1) activity and diminished FAO capacity. Hypoxia reduces PPAR alpha/RXR binding activity and had no effect on the nuclear level of PPAR alpha protein. Hypoxia reduced the nuclear and cellular RXR levels and deactivates PPAR alpha by reducing the availability of its obligate partner RXR. In rat models of systemic hypoxia (Razeghi et al., 2001), cardiac hypoxia induces a decrease in heart muscle transcript levels of PPAR alpha and PPAR alpha-regulated genes (PDK4), muscle CPT-1, and malonyl-CoA decarboxylase. This explains the increased reliance of the heart for glucose during hypoxia.
PPAR gamma co-activator 1 alpha (PGC-1 alpha) is a major regulator of mitochondrial biogenesis and activity in the cardiac muscle. Hypoxia stimulates the expression of PGC-1 alpha in cardiac myocytes (Zhu et al., 2010). PGC-1 alpha stimulates the expression of clock genes, particularly Bmal1 and Rev-erb alpha (Liu et al., 2007a). Mice lacking PGC-1alpha present abnormal diurnal rhythms of activity, body temperature, and metabolic rate. Overexpression of PGC-1 alpha inhibits clock gene expression in both heart and skeletal muscles and decreases the expression of PPAR alpha. PGC-1 alpha overexpression abolishes the diurnal variation of EF (Wu et al., 2011) and plays an important role on cardiac function by regulating CRs of metabolic genes.
Ephrins belong to the family of receptor tyrosine kinases. Interestingly, Ephrin-Eph cell signaling is linked to the Wnt/beta catenin pathway (Clevers and Batlle, 2006) and favorably influences cardiomyocyte viability which ultimately preserves cardiac function after myocardial infarction. Ephrin-Eph signaling could potentially be a new therapeutic target in the treatment of myocardial infarction (O'Neal et al., 2013). In non re-perfused hearts of mice with a functional deletion of the CR gene mPer2, myocardial infarct size is reduced. A decrease in infarct size in mPer2-M mouse hearts following ischemia-reperfusion injury and ischemic preconditioning is observed and improves preservation of myocardial viability. In the mPer2-mutant mouse myocardium cardio-protection occurs via the mechanisms connecting cardiac events, mitochondrial function, and mPer2 (Virag et al., 2013).
Cardiac hypertrophy and cardiac overload
Development of cardiac hypertrophy and progression to heart failure induce a change in myocardial metabolism, characterized by a switch from fatty acid utilization to glycolysis, and lipid accumulation. PPAR gamma and HIF-1 alpha are key mediators of lipid anabolism and glycolysis, respectively. They are jointly up-regulated in hypertrophic cardiomyopathy and cooperate to mediate key changes in cardiac metabolism (Krishnan et al., 2009). In response to pathological stress, HIF-1 alpha activates glycolytic genes, and PPAR gamma. This results in increased glycolytic flux, glucose-to-lipid conversion via the glycerol-3-phosphate pathway, and contractile dysfunction. Ventricular deletion of HIF1 alpha in mice prevents hypertrophy-induced PPAR gamma activation, the consequent metabolic re-programming, and contractile dysfunction. HIF-1 alpha and PPAR gamma protein expression is up-regulated in human and mouse cardiac hypertrophy. HIF 1 alpha directly activates PPAR gamma transcription. PPAR gamma is a key downstream effector of HIF-1 alpha-driven triacylglycerol accumulation in cardiomyocytes (Krishnan et al., 2009).
In pathological hypertrophied heart, PPAR alpha expression and activity are diminished, leading to a reduction in the capacity for FAO and increased rate of glucose utilization (Barger et al., 2000). Alpha 1-adrenergic agonist-induced hypertrophy of cardiomyocytes in culture results in a switch in energy substrate preference from fatty acids to glucose and in a significant decrease in palmitate oxidation rates together with a reduction in the expression of the gene encoding muscle carnitine palmitoyltransferase 1 (M-CPT1). Cardiac myocyte transfection has shown that M-CPT1 promoter activity is repressed during cardiomyocyte hypertrophic growth, an effect involving a PPAR alpha response element. Hypertrophied myocytes exhibited reduced capacity for cellular lipid homeostasis, as evidenced by intracellular fat accumulation. Thus, during cardiomyocyte hypertrophic growth, PPAR alpha is deactivated at several levels, leading to diminished capacity for myocardial lipid metabolism. The functional consequences of this metabolic switch from lipid to glucose may serve to preserve ventricular function in the context of chronic pressure overload (Young et al., 2001a). During cardiac pressure overload-induced cardiac hypertrophy, the diurnal variation of metabolic gene expression is completely suppressed and the cardiac performance is impaired (Young et al., 2001b). The induction of clock output genes is attenuated in the pressure-overloaded hypertrophied heart, providing evidence for a diminished ability of the hypertrophied heart to anticipate and subsequently to adapt to physiological alterations during the day (Young et al., 2001c).
Osteoporosis
The Wnt pathway induces differentiation of bone-forming cells (osteoblasts) and suppresses the development of bone-resorbing cells (osteoclasts). It is controlled by antagonists that interact either with Wnt proteins (Wnts) or with Wnt co-receptors. Wnts function as key regulators in osteogenic differentiation of mesenchymal stem cells and bone formation. Aberrant Wnt pathways are associated with many osteogenic diseases (Rawadi and Roman-Roman, 2005; Canalis, 2013). Both human genetics and animal studies have pointed out the role of the Wnt/LRP5 pathway as a major regulator of bone mass. In mice, down-regulation or neutralization of Wnt antagonists enhances bone formation. Mutations in LRP5 cause primary osteoporosis by reducing Wnt signaling activity and result in decreased bone formation (Korvala et al., 2012). Heterozygous PPAR gamma-deficient mice exhibit high bone mass by stimulating osteoblastogenesis from bone marrow progenitors. Inhibition of PPAR gamma increases osteoblastogenesis and bone mass in male C57BL/6 Mice (Duque et al., 2013). PPAR gamma inhibits osteoblast differentiation (Wan et al., 2007).
Cardiovascular disease and osteoporosis are common age-related conditions associated with significant morbidity and mortality. An increasing body of biological and epidemiological evidences provides support for a link between cardiovascular disease and osteoporosis that cannot be explained by age alone (Farhat and Cauley, 2008). Several hypotheses have been proposed to explain the link between osteoporosis and cardiovascular disease including shared risk factors, common pathophysiological mechanisms and common genetic factors.
Alzheimer disease (AD)
AD is a progressive neurodegenerative disorder, neuropathologically characterized by amyloid-beta (Abeta) plaques, and hyperphosphorylated tau accumulation with hereditary missense mutations in the amyloid precursor protein or presenilin-1 and -2 (PSEN1 and PSEN2) genes. Presenilins are involved in modulating beta-catenin stability; therefore familial AD-linked PSEN-mediated effects can reduce the Wnt pathway (Boonen et al., 2009). Tau phosphorylation is mediated by GSK-3 beta, a key antagonist of the Wnt pathway. Sustained loss of function of Wnt/beta-catenin signaling underlies the onset and progression of AD (Inestrosa and Toledo, 2008; De Ferrari et al., 2014). Downregulation of Wnt signaling induced by Abeta is associated with AD progression. Persistent activation of Wnt signaling through Wnt ligands, or inhibition of negative regulators of Wnt signaling, such as Dickkopf-1 and GSK-3 beta are able to protect against Abeta toxicity and ameliorate cognitive performance in AD (Wan et al., 2014). A relationship between amyloid-beta-peptide -induced neurotoxicity and a decrease in the cytoplasmatic levels of beta-catenin has been observed. Although PPAR gamma is elevated in the brain of AD individuals (Jiang et al., 2008), activation of the Wnt signaling pathway may be proposed as a therapeutic target for the treatment of AD.
In old mice engineered to lack Bmal1, there is evidence of brain cell damage that looked similar to that seen in AD. BMAL1 in a complex with CLOCK regulates cerebral redox homeostasis and connects impaired clock gene function to neurodegeneration (Musiek et al., 2013). Altered CR synchronization has been reported in the brain of AD patients (Cermakian et al., 2011). CR disturbances affect as many as a quarter of AD patients. Alterations in the SCN and melatonin secretion are the major factors linked with CR abnormalities. Daytime agitation, night-time insomnia, and restlessness are among the common behavioral alterations observed in AD. Normally, in the interstitial fluid, Abeta has a diurnal fluctuation with low levels during sleep and peak levels during wake. Prolonged wake and/or orexin administration increase levels of the Abeta in the interstitial fluid of the brain in mice. Orexin antagonist reduces amyloid deposits in brain areas. There is a strong causal association between AD and cardiovascular disease. Several cardiovascular risk factors including hypertension and diabetes are also risk factors for dementia (Stampfer, 2006).
Bipolar disorder and schizophrenia
The Wnt pathway and its key enzyme, GSK 3 beta, which antagonizes the canonical Wnt pathway, play an important role in regulating synaptic plasticity, cell survival, and CRs in the mature central nervous system. This pathway is implicated in the pathophysiology and treatment of bipolar disorder (Gould and Manji, 2002; Valvezan and Klein, 2012). GSK3- beta-inhibitor lithium chloride enhances activation of Wnt canonical signaling (Hedgepeth et al., 1997; Sinha et al., 2005; Galli et al., 2013). Lithium activates downstream components of the Wnt signaling pathway in vivo, leading to an increase of the beta-catenin protein. GSK3-beta phosphorylates and stabilizes the orphan nuclear receptor Rev-erb alpha, a negative component of the circadian clock. Lithium treatment of cells leads to rapid proteasomal degradation of Rev-erb alpha and activation of clock gene Bmal1 (Yin et al., 2006). The origin of cyclicity in bipolar disorders has been shown by means of a computational approach, and this disease enters the class of dissipative structures (Goldbeter, 2013). Valproate, an effective medication for the prevention and treatment of mood symptoms in bipolar disorder causes a decrease of PPAR gamma signaling (Lan et al., 2008). Many cardiovascular complications are seen in bipolar disorder (Swartz and Fagiolini, 2012).
An emerging role for Wnt and GSK-3 beta signaling pathways has been found in schizophrenia (Singh, 2013). Sleep and circadian rhythm disruption are seen in schizophrenia (Wulff et al., 2012). Schizophrenia increases risks of cardiovascular disease, particularly coronary heart disease, dyslipidemia, diabetes and hypertension (Hennekens et al., 2005; Andreassen et al., 2013).
Diseases Associated with Activation of the Wnt/Beta-Catenin Pathway and Decreased Expression of PPAR Gamma
Numerous diseases present a common denominator: the Wnt/beta-catenin pathway is overexpressed and the PPAR gamma expression is decreased. This explains why type 2 diabetes is commonly associated with hypertension, sympathetic- parasympathetic abnormalities, and cancers and why CR disruptions are often observed among these pathologies. PPAR alpha expression is often increased in these diseases.
Impaired sympathetic-parasympathetic system
PPAR gamma and sympathetic nerve activity (SNA) antagonistically regulate energy metabolism and cardiovascular function with the former promoting anabolism and vasorelaxation and the later favoring catabolism and vasoconstriction (Yang et al., 2013). Systemic inactivation of PPAR gamma can be generated constitutively by using Mox2-Cre mice (MoxCre/flox) or inducibly by using the tamoxifen system (EsrCre/flox/TM). There is an increase in heart rate in both strains of null mice. PPAR gamma deletion causes the activation of SNA. Rosuvastatin increases vascular endothelial PPAR gamma expression and corrects blood pressure variability in obese dyslipemic mice (Desjardins et al., 2008). Sympathetic adrenal function is disrupted in both Bmal1(-/-) and Clock (mut) mice (Curtis et al., 2007). Although a shorter ultradian rhythm remains, Bmal1 deletion abolishes the 24-h frequency in cardiovascular rhythms. In humans, heart rate variability has been shown to be driven by an intrinsic mechanism (Hu et al., 2004; Ivanov et al., 2007). CRs and sleep modulate the sympathetic-parasympathetic balance. Sleep deprivation induces a decrease in the global variability, and an imbalance of the autonomous nervous system (ANS) with an increase in sympathetic activity and a loss of parasympathetic predominance. Human individuals homozygous for the longer allele PER3(5/5) compared with PER3(4/4) subjects present an elevated sympathetic predominance and a reduction of parasympathetic activity (Viola et al., 2008). In mice, selective deletion of the Bmal1 activator PPAR gamma in the vasculature induces a diminution in heart rate circadian variations (Wang et al., 2008). The CCM mouse model exhibits a decrease in heart rate. Conversely, this model does not present differences in systolic, diastolic, and mean blood pressures as compared with controls (Bray et al., 2008).
Type 2 diabetes
PPAR alpha activity and its downstream targets are abnormally activated in the diabetic heart, leading to a marked increase in both fatty acid uptake and oxidation (Finck et al., 2005). Chronic activation of the cardiac PPAR alpha pathway which occurs in the diabetic heart, contributes to myocardial lipid accumulation and diabetic cardiomyopathy (Finck et al., 2002). Diabetes alters the circadian clock in the heart. The clock in the heart loses normal synchronization with its environment during diabetes. Diabetes and fasting activate the expression of cardiac FAO. Excessive fatty acid import and oxidation may be a cause of pathological cardiac remodeling in the diabetic heart (Finck and Kelly, 2002). In type 2 diabetes, PPAR alpha is overexpressed and expression of PPAR gamma is deceased. Some TZD PPAR agonists are used to treat type 2 diabetes. The Wnt/beta-catenin signaling pathway is involved in diabetes mellitus (Ip et al., 2012). Expression of PGC-1 alpha is down-regulated in muscles of type 2 diabetic subjects (Liang and Ward, 2006). PGC-1 alpha activates the expression of insulin-sensitive GLUT4 in skeletal muscle and plays a role in preventing insulin resistance and type 2 diabetes mellitus.
The mammalian clock (Bmal1, Clock, Cry1, Cry2, Per1, Per2, and Per3) expresses CRs and the phases of these CRs are altered in the hearts from streptozotocin-induced diabetic rats (Young et al., 2002). Two BMAL1 haplotypes are associated with type 2 diabetes and hypertension. This provides evidence of a causative role of Bmal1 variants in pathological components of the metabolic syndrome (Woon et al., 2007). Rhythmic control of insulin release is deregulated in humans with diabetes. Disruption of the clock components Clock and Bmal1 leads to hypoinsulinemia and type 2 diabetes. Pancreatic islets express self-sustained circadian gene and protein oscillations of the transcription factors CLOCK and BMAL1. The phase of oscillation of the islet genes is delayed in circadian mutant mice, and both Clock and Bmal1 mutants show impaired glucose tolerance, reduced insulin secretion and defects in size and proliferation of pancreatic islets (Marcheva et al., 2010). In rodent models of type II diabetes, mean blood pressure is mildly elevated. The elevation in blood pressure is accompanied by changes in the circadian variation of blood pressure as demonstrated in type 2 diabetes (db/db) mice. The daytime fall in blood pressure in mice is significantly blunted in type 2 diabetes db/db mice (Rudic and Fulton, 2009).
Hypertension
PPAR gamma in vascular muscle plays a role in the regulation of vascular tone and blood pressure. Thus, mutations in PPAR gamma induce severe hypertension and type 2 diabetes. Transgenic mice with mutations in PPAR gamma in smooth muscle present vascular dysfunction and severe systolic hypertension (Halabi et al., 2008). PPAR gamma ligands lower blood pressure in both animals and humans. PPAR gamma agonist rosiglitazone improves vascular function and lowers blood pressure in hypertensive transgenic mice (Ryan et al., 2004). In mice, after vascular PPAR gamma deletion, circadian variations of blood pressure and heart rate are dampened through a dysregulation of Bmal1 (Wang et al., 2008). In a null mouse model with specific disruption of PPAR gamma in endothelial cells, PPAR gamma appears to be an important regulator of blood pressure and heart rate mimicking type 2 diabetes, and mediates the antihypertensive effects of rosiglitazone (Nicol et al., 2005). PPAR gamma regulates the renin-angiotensin system activity in the hypothalamic paraventricular nucleus and ameliorates peripheral manifestations of heart failure (Yu et al., 2012). Activation of PPAR gamma down-regulates the renin-angiotensin system. PPAR gamma is expressed in key brain areas involved in cardiovascular and autonomic regulation. Activation of central PPAR gamma reduces sympathetic excitation and improves peripheral manifestations of heart failure by inhibiting brain renin-angiotensin system activity. PPAR gamma ligands lower blood pressure in both animals and humans, possibly via the PPAR gamma-mediated inhibition of the angiotensin II type 1 receptor expression which results in the suppression of the renin-angiotensin system (Sugawara et al., 2010). Genetic variation in BMAL1 is associated with the development of hypertension in man. BMAL1 dysfunction is associated with susceptibility to hypertension and type 2 diabetes. In conditions of constant darkness, Cry1/Cry2 deficient mice are hypertensive in the daytime (Rudic and Fulton, 2009). Targeted deletion of Bmal1 in mice (Bmal1-KO) abolishes the CR in blood pressure. Mice with targeted deletion of PPAR gamma in the endothelium (EC-PPAR gamma-KO) exhibit a striking phenotypic resemblance to endothelial cell (EC)-specific deletion of Bmal1 (EC-Bmal1-KO). The loss of PPAR gamma in the aorta of both EC-PPAR gamma-KO mice leads to reduced expression of Bmal1, Cry1, Cry2, and Per2. The ability of PPAR gamma to modulate blood pressure arises in part from its ability to transactivate Bmal1.
Atherosclerosis
Wnt/beta-catenin signaling plays a key role in atherosclerosis (Wang et al., 2002). Besides Wnt/beta-catenin, GSK3-beta acts as a beta-catenin independent signal, and plays a crucial role in the regulation of cell proliferation and vascular homeostasis. The progression of atherosclerosis is prevented by PPAR gamma ligands in both animals and humans (Sugawara et al., 2010). Monocyte adhesion to vascular endothelium is one of the early processes in the development of atherosclerosis (Lee et al., 2006). Activation of the canonical Wnt/beta-catenin pathway enhances monocyte adhesion to endothelial cells.
Cardiac-restricted overexpression of PPAR alpha (MHC-PPAR)
In mice with cardiac-restricted overexpression of PPAR alpha (MHC-PPAR), the expression of PPAR alpha target genes is increased whereas that of genes involved in glucose transport and utilization is repressed (Finck et al., 2002). The metabolic phenotype of MHC-PPAR mice mimics that of the diabetic heart. MHC-PPAR hearts exhibits profiles of diabetic cardiomyopathy including ventricular hypertrophy, activation of gene markers of pathological hypertrophic growth, and systolic ventricular dysfunction. Transgenic mice overexpressing PPAR alpha in muscle (MCK-PPAR alpha mice) developed glucose intolerance. Skeletal muscle of MCK-PPAR alpha mice exhibits increased FAO rates and reduced insulin-stimulated glucose uptake. The effects on muscle glucose uptake imply transcriptional repression of the GLUT4 gene.
Aging
Aging is associated with various heart diseases, and this may be attributable, in part, to the prolonged exposure of the heart to cardiovascular risk factors. However, aging is also associated with heart disorders such as diastolic dysfunction that are not necessarily linked to the risk factors for cardiovascular diseases. A mechanistic link between Wnt signaling and premature aging or aging-related phenotypes has been demonstrated (Naito et al., 2010). Tissues and organs from klotho-deficient animals showevidence of increased Wnt signaling. Both in vitro and in vivo, continuous Wnt exposure triggers accelerated cellular senescence. Thus, klotho appears to be a Wnt antagonist (Brack et al., 2007; Liu et al., 2007b). Specific mutations in the human gene encoding lamin A cause premature aging. In mice and humans, these mutations affect adult stem cells by interfering with the Wnt signaling pathway (Meshorer and Gruenbaum, 2008). Overexpression of Per in the fruit fly Drosophila melanogaster enhances long-term memory, while in Per null flies memory is impaired. This supports a link for circadian genes in the processes of learning and memory (Sakai et al., 2004). In aged animals, the normal photonic stimulation of Per1 expression is reduced. The free-running period of Per1–luc rhythmicity is shortened in aged animals and the amplitude of Clock and Bmal1 expression is decreased (Kolker et al., 2003).
Neurodegenerative diseases
The common denominator overexpression of the Wnt/beta-catenin pathway and the consequent decrease in PPAR gamma expression play a central role in numerous neurodegenerative diseases (Clevers, 2006a; MacDonald et al., 2009; Yang, 2012). PPAR gamma agonists could potentially inhibit neuro-inflammation and subsequently neurodegeneration. This may partially occur through the ability of PPAR: RXR heterodimers to antagonize NFκB mediated gene transcription of several inflammatory mediators such as COX-2, iNOS, and various proinflammatory cytokines. It is not surprising that abnormalities of the cardiovascular system and CRs dysfunction are often associated with neurodegenerative pathologies. Sleep disturbances may predict manifestation of neurodegenerative diseases (Postuma and Montplaisir, 2009).
Huntington disease (HD)
HD is a dominantly inherited cytosine-adenine-guanine (CAG) repeat disorder with expanded polyglutamine (polyQ) tracts in huntingtin, causing striatal and cortical degeneration (Walker, 2007). Huntingtin interacts with beta-catenin, beta -TrCP, and axin. Normal huntingtin acts as a scaffold protein, promoting the beta-catenin degradation by facilitating the recognition of beta-catenin by beta -TrCP within the destruction complex (Godin et al., 2010). The binding of beta-catenin to the destruction complex is altered in HD. The presence of an abnormal polyQ expansion in mutant huntingtin leads to a decreased binding to beta-catenin therefore impairing the binding of beta-catenin to the destruction complex and subsequently resulting in beta-catenin accumulation into the cytosol. Thus, beta-catenin levels are up-regulated in HD. Mutant huntingtin alters the stability and levels of beta-catenin. Reducing the canonical Wnt signaling pathway confers protection against mutant huntingtin toxicity in Drosophila (Dupont et al., 2012). Knockdown of Wnt ligands improves the survival of HD flies. Overexpression of armadillo/beta-catenin destruction complex component (AXIN, APC2, or GSK3-beta) increases the lifespan of HD flies.
Early-onset of cardiovascular disease is the second leading cause of death in HD patients. Due to the ubiquitous expression of huntingtin, all cell types with high energetic levels can be impaired. Expression of mutant huntingtin induces cardiac dysfunction in the transgenic model of HD (line R6/2). R6/2 mice develop cardiac dysfunction with cardiac remodeling (e.g. hypertrophy, fibrosis, apoptosis, beta1 adrenergic receptor down-regulation) (Mihm et al., 2007). R6/1 transgenic mice exhibit profound autonomic nervous system-cardiac dysfunction involving both sympathetic and parasympathetic systems, leading to cardiac arrhythmias, and sudden death (Kiriazis et al., 2012). A baroreceptor reflex dysfunction has been described in the BACHD mouse model of HD (Schroeder et al., 2011). Several studies report dysfunction of the autonomic nervous system in HD patients. This may contribute to the increased incidence of cardiovascular events in this patient population that often leads to death. There is a blunted response of the baroreceptor reflex as well as a significantly higher daytime blood pressure in BACHD mice compared to WT controls, which are both indications of autonomic dysfunction. In humans, autonomic dysfunction is present even in the middle stages of HD and affects both the sympathetic and parasympathetic systems (Andrich et al., 2002). Sleep and wake regions of the brain including the brainstem, thalamus, hypothalamus, and cortex are also affected in HD (Kremer et al., 1991). The SCN pacemaker is functional in HD mouse models, so a dysfunction of the circadian circuitry has been proposed to contribute to circadian abnormalities (Pallier and Morton, 2009). Central and peripheral clock gene expression is altered (Maywood et al., 2010). The sleep/wake cycle is disrupted in HD patients characterized by sleep fragmentation at night and delayed sleep phase (Aziz et al., 2010).
Amyotrophic lateral sclerosis (ALS)
ALS is a neurodegenerative disease resulting in the progressive loss of upper and lower limb motoneurons and leading to gradual muscle weakening ultimately causing paralysis and death. The Wnt/beta-catenin pathway plays a role in the neurodegeneration of motor neurons in an in vitro model of ALS (Pinto et al., 2013). In ALS, a potentially therapeutic pathway may be the activation by PPAR gamma agonists due to their ability to block the neuropathological damage caused by inflammation (Kiaei, 2008). The neuroprotective effect of pioglitazone has been demonstrated in G93A SOD1 transgenic mouse model of ALS and shows a significant increase in their survival. In ALS, PPAR gamma controls natural protective mechanisms against lipid peroxidation (Benedusi et al., 2012).
Ataxin-2 gene (ATX2) is linked to a number of neurodegenerative disorders in humans including ALS and Parkinson disease (PD). ATX2 protein inhibits the production of certain proteins and plays a crucial role in the control of the circadian sleep/wake cycle. ATX2 regulates the expression of the circadian protein Per in Drosophila. By reducing expression of ATX2 in Drosophila, the flies are active two and half hours longer. Patients suffering from a form of the neurodegenerative disease spinocerebellar ataxia caused by ATX2 mutations also experience rapid eye movement sleep disruptions. ATX2 is necessary for PER accumulation in circadian pacemaker neurons and thus determines period length of circadian behavior. ATX2 is required for the function of TWENTY-FOUR, an activator of PER translation. In humans with ALS, CR of cortisol is impaired (Patacchioli et al., 2003). Both sympathetic and parasympathetic dysfunctions are observed in ALS (Druschky et al., 1999). There are sleep-wake disturbances in patients with ALS (Lo Coco et al., 2011). In human ALS, heart failure is a frequent common cause of death (Gdynia et al., 2006).
Parkinson disease (PD)
In a mouse model of PD, a cross talk between inflammatory and Wnt/beta-catenin signaling pathways is involved (L'Episcopo et al., 2012). The Wnt1 regulated Frizzled-1/beta-catenin signaling pathway controls the mesencephalic dopaminergic neuron-astrocyte crosstalk (L'Episcopo et al., 2011). The PPAR gamma agonist pioglitazone modulates inflammation and induces neuroprotection in PD monkeys (Swanson et al., 2011) and mice (Schintu et al., 2009). Expanded glutamine repeats of the ATX2 protein have been identified in fronto-temporal lobar degeneration in PD (Ross et al., 2011). Moreover, a peripheral molecular clock, as reflected in the dampened expression of the clock gene Bmal1 in leukocytes is altered in PD patients (Cai et al., 2010). There is a disappearance of CRs in a PD dog model (Hineno et al., 1992). Sleep disturbances in PD may be related to CR dysfunction (Hack et al., 2014). Sleep complaints are present in almost half of PD patients. PD patients exhibit increased sleep latency and reduced sleep efficiency. In PD, there is a sustained elevation of serum cortisol levels, reduced circulating melatonin levels, and altered Bmal1 expression (Breen et al., 2014). PD causes dysfunction of the diurnal autonomic cardiovascular regulation. This dysfunction is profound in patients with severe PD (Haapaniemi et al., 2001).
Multiple sclerosis (MS)
Wnt signaling is involved in the MS pathogenesis (Yuan et al., 2012). Mice with experimental autoimmune encephalomyelitis (EAE) have been widely used as a MS model with central nervous system demyelination, neuro-inflammation, and motor impairments. Wnt3a, a Wnt ligand for the canonical pathway, is significantly increased in the spinal cord dorsal horn (SCDH) of the EAE mice. Beta-catenin is also significantly up-regulated. Wnt signaling pathways are up-regulated in the SCDH of the EAE mice and aberrant activation of Wnt signaling contributes to the development of EAE-related chronic pain. PPAR gamma agonists modulate the development of experimental EAE (Drew et al., 2008). Moreover, the risk of myocardial infarction, stroke, heart failure, and atrial fibrillation or flutter is increased in MS patients (Jadidi et al., 2013).
Friedreich ataxia (FRDA)
FRDA is a debilitating, life-shortening, degenerative neuromuscular disorder, due to frataxin (FXN) deficiency. FRDA is characterized by neuronal degeneration and heart failure, which are due to loss of transcription of the FXN gene caused by a trinucleotide repeat expansion. FXN is a mitochondrial protein involved in iron–sulfur-cluster biogenesis, serving to bind and transfer iron to key electron transport complexes and cytochrome C. Diabetes mellitus and serious heart dysfunction (hypertrophic cardiomyopathy) are associated in most cases. The PPAR gamma agonist Azelaoyl PAF increases FXN protein and mRNA expression in human neuroblastoma cells SKNBE and in primary fibroblasts from skin biopsies from FRDA patients. This offers new implications for the FRDA therapy (Marmolino et al., 2009). It has been shown a coordinate dysregulation of the PPAR gamma co-activator PGC-1 alpha and transcription factor Srebp1 in cellular and animal models of FXN deficiency, and in cells from FRDA patients. A genetic modulation of the PPAR gamma pathway affects FXN levels in vitro, supporting PPAR gamma as a new therapeutic target in FRDA (Coppola et al., 2009).
Colon cancer
Activation of beta-catenin-Tcf signaling has been observed in colon cancer (Morin et al., 1997). Activation of the Wnt signaling pathway via mutation of the APC gene is a critical event in the development of colon cancer (Najdi et al., 2011). Inherited mutations in APC lead to the development of non-invasive colonic adenomas (polyps). Wnt pathway activation is a driving force in the development of adenomas. Activation of the Wnt/beta-catenin signaling pathway decreases PPAR gamma activity in colon cancer cells (Jansson et al., 2005) and a loss-of-function mutations in PPAR gamma is associated with human colon cancer (Sarraf et al., 1999).
Colorectal cancer is linked to CR dysregulation (Savvidis and Koutsilieris, 2012). Down-regulation of Per2 increases beta-catenin protein levels and its target cyclin D, leading to cell proliferation in colon cancer cell lines and colonic polyp formation. Per2 gene activation suppresses tumorigenesis in colon by down-regulation of beta-catenin. Increased beta-catenin affects the circadian clock and enhances PER2 protein degradation in colon cancer. Suppression of human beta-catenin expression inhibits cellular proliferation in intestinal adenomas. Disruption of the peripheral intestinal CRs may contribute to intestinal epithelial neoplastic transformation of human colorectal cancer. The circadian expression of dihydropyrimidine dehydrogenase, an enzyme that is implicated in the metabolism of the anticancer drug 5-fluorouracil, may be regulated by Per1 in high-grade colon tumors. The ephrin-Eph cell pathway is linked to the Wnt/beta catenin pathway and is involved in colon cancer (Clevers, 2006a,b).
Functional bowel disorders are associated with autonomic disturbance (Tougas, 2000). People with type 2 diabetes have an increased risk of developing colorectal cancer. Diabetes is associated with a higher risk of colon cancer (Yuhara et al., 2011). Heart disease increases at twice the risk of bowel cancer. Colon cancer and coronary artery disease are known to share similar risk factors (smoking, high-fat diet, obesity, diabetes, high blood pressure, and sedentary lifestyle) which increase the risk of colon cancer. This suggests that the two diseases may be connected (Chan et al., 2007). People with coronary artery disease are more likely to develop colon cancer than those without.
Synthesis
Circadian rhythms (CRs) are particularly fascinating phenomena. They go very far back in evolution. The existence of a CR is the signature of instability. Beyond a point of bifurcation, an unstable thermodynamic system can evolve spontaneously into a periodic state. These periodic oscillations correspond to a phenomenon of self-organization in time and have been called “dissipative structures” (Prigogine et al., 1974). Dissipative structures are far-from-equilibrium systems, such as cyclones, hurricanes, lasers, Bénard cells, Belousov–Zhabotinsky reactions, Turing structures, circadian rhythms, and more generally most of the living organisms. CRs are based on the existence of negative feedback loops. Oscillatory behavior gradually has been integrated into the living world to become one of its major characteristics. In the cardiovascular system, circadian genes show properties of anticipation and this makes it possible to coordinate lipid and carbohydrate metabolism with the cardiovascular function, especially for blood pressure and heart rate. Dysfunction of CRs can be associated with serious clinical problems and may induce a negative impact on quality of life, sometimes with a poor prognosis. Abnormalities of circadian gene function may result in the occurrence of metabolic syndrome, obesity, and even more seriously, stroke, or myocardial infarction.
Two major systems interfere with circadian genes, namely the canonical Wnt pathway, and the PPAR system. In some cases of diseases presenting profiles 1 or 2, there is not always evidence of the exact influence of CRs on the Wnt-beta catenin-PPAR gamma pathway and cardiac function. PPAR gamma controls the circadian Clock-Bmal1 genes in the vascular system. Importantly, there is an opposite between activation of the Wnt pathway and PPAR gamma. This is attested by their respective profiles in numerous diseases, either cardiovascular diseases or pathologies with cardiovascular complications. There is a subtle thermodynamic regulation of CRs that run far-from-equilibrium; moreover, there is the need to maintain the balance between the two systems canonical Wnt and PPAR gamma. Indeed, activation of the Wnt system with inactivation of PPAR gamma favors diabetes, hypertension, several cancers, and neurodegenerative diseases. The reverse is observed in ARVD, osteoporosis, Alzheimer disease, bipolar disorder, schizophrenia, and myocardial ischemia. The extreme complexity of the Wnt-PPAR systems and their numerous inter-related pathways partly explain their involvement in numerous diseases. We remain surprised by both the number and the importance of these diseases, causing considerable morbidity, and mortality and heavy social and economic costs.
The discovery and use of new agonist or antagonist pharmacological agents acting on PPARs, and more generally, directly or indirectly implied in the canonical Wnt system, are particularly important. This leads to numerous novel therapeutic approaches. PPAR gamma is a key regulator of lipid metabolism and its activation by some TZD is used for the treatment of type 2 diabetes and protects against atherosclerosis. However, some TZD have been reported to cause a higher rate of fractures in human patients. Pharmacological inhibition of PPAR gamma represents a potential therapeutic approach for age-related bone loss. Induction of the Wnt pathway or inhibition of Wnt antagonists may offer therapeutic opportunities in treating bone disorders, including osteoporosis. Antibodies targeting the Wnt inhibitor sclerostin lead to increased bone mineral density in post-menopausal women. Lithium, often used to treat bipolar disorder, blocks a Wnt antagonist, decreasing the patient's risk of fractures. Lithium exerts effects on components of the Wnt signaling pathway. The Wnt signaling pathway plays an important role in the treatment of bipolar disorder. The future development of selective GSK3- beta inhibitors may have considerable utility not only for the treatment of bipolar disorder but also for a variety of neurodegenerative disorders. Therapies targeting the Wnt pathway are not without risk, and may lead to over-activation of Wnt/catenin and its association with many tumors. However, it is conceivable that targeting Wnt inhibitors may predispose the individuals to tumorigenic phenotypes.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We would like to thank Mr. Vincent Gobert and Dr. Michel Grivaux, Director of the Clinical Research Center of the Meaux Hospital, France.
Abbreviations
CR, circadian rhythm; PPAR, Peroxisome proliferator-activated receptor; SCN, suprachiasmatic nucleus; APC, adenomatous polyposis coli; Dsh, Disheveled GSK3-beta, glycogen synthase kinase 3-beta; LRP5/6, low density lipoprotein receptor-related protein 5/6; Fzd, Frizzled; Tcf/Lef, T cell/lymphoid-enhancing binding; Clock, Circadian locomotor output cycles kaput; Ccm, cardiomyocyte-specific clock mutant; FAO, fatty acid oxidation; RXR, retinoid X receptor; PPRE, peroxisome proliferator response element; TZD, thiazolidinedione; Bmal1, Brain and muscle aryl-hydrocarbon receptor nuclear translocator-like; Cry, cryptochrome; Per, Period; Ccg, clock-controlled genes; Ror, related orphan receptor; TZD, thiazolidinediones; ARVC, arrhythmogenic right ventricular cardiomyopathy; DSG, desmoglein; DSC, desmocollin; PKP, plakophilin; PG, plakoglobin; DSP, desmoplakin; PDK, pyruvate deshydrogenase kinase; HIF1 alpha, hypoxia-inducible factor 1 alpha; PGC-1 alpha, PPAR gamma co-activator 1 alpha; M-CPT1, muscle carnitine palmitoyltransferase 1; ANS, autonomous nervous system; AD, Alzheimer Disease; HD, Huntington disease; ALS, Amyotrophic lateral sclerosis; PD, Parkinson disease; MS, Multiple sclerosis; FRDA, Friedreich ataxia; Abeta, amyloid-beta; FXN, Frataxin.
References
Akashi, M., and Takumi, T. (2005). The orphan nuclear receptor RORalpha regulates circadian transcription of the mammalian core-clock Bmal1. Nat. Struct. Mol. Biol. 12, 441–448. doi: 10.1038/nsmb925
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Anan, F., Masaki, T., Fukunaga, N., Teshima, Y., Iwao, T., Kaneda, K., et al. (2007). Pioglitazone shift circadian rhythm of blood pressure from non-dipper to dipper type in type 2 diabetes mellitus. Eur. J. Clin. Invest. 37, 709–714. doi: 10.1111/j.1365-2362.2007.01854.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Andreassen, O. A., Djurovic, S., Thompson, W. K., Schork, A. J., Kendler, K. S., O'Donovan, M. C., et al. (2013). Improved detection of common variants associated with schizophrenia by leveraging pleiotropy with cardiovascular-disease risk factors. Am. J. Hum. Genet. 92, 197–209. doi: 10.1016/j.ajhg.2013.01.001
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Andrich, J., Schmitz, T., Saft, C., Postert, T., Kraus, P., Epplen, J. T., et al. (2002). Autonomic nervous system function in Huntington's disease. J. Neurol. Neurosurg. Psychiatry 72, 726–731. doi: 10.1136/jnnp.72.6.726
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Anea, C. B., Zhang, M., Stepp, D. W., Simkins, G. B., Reed, G., Fulton, D. J., et al. (2009). Vascular disease in mice with a dysfunctional circadian clock. Circulation 119, 1510–1517. doi: 10.1161/CIRCULATIONAHA.108.827477
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Aziz, N. A., Anguelova, G. V., Marinus, J., Lammers, G. J., and Roos, R. A. (2010). Sleep and circadian rhythm alterations correlate with depression and cognitive impairment in Huntington's disease. Parkinsonism Relat. Disord. 16, 345–350. doi: 10.1016/j.parkreldis.2010.02.009
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Barger, P. M., Brandt, J. M., Leone, T. C., Weinheimer, C. J., and Kelly, D. P. (2000). Deactivation of peroxisome proliferator-activated receptor-alpha during cardiac hypertrophic growth. J. Clin. Invest. 105, 1723–1730. doi: 10.1172/JCI9056
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Basso, C., Corrado, D., Marcus, F. I., Nava, A., and Thiene, G. (2009). Arrhythmogenic right ventricular cardiomyopathy. Lancet 373, 1289–1300. doi: 10.1016/S0140-6736(09)60256-7
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Belanger, A. J., Lu, H., Date, T., Liu, L. X., Vincent, K. A., Akita, G. Y., et al. (2002). Hypoxia up-regulates expression of peroxisome proliferator-activated receptor gamma angiopoietin-related gene (PGAR) in cardiomyocytes: role of hypoxia inducible factor 1alpha. J. Mol. Cell. Cardiol. 34, 765–774. doi: 10.1006/jmcc.2002.2021
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Benedusi, V., Martorana, F., Brambilla, L., Maggi, A., and Rossi, D. (2012). The peroxisome proliferator-activated receptor gamma (PPARgamma) controls natural protective mechanisms against lipid peroxidation in amyotrophic lateral sclerosis. J. Biol. Chem. 287, 35899–35911. doi: 10.1074/jbc.M112.366419
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Ben-Ze'ev, A., and Geiger, B. (1998). Differential molecular interactions of beta-catenin and plakoglobin in adhesion, signaling and cancer. Curr. Opin. Cell Biol. 10, 629–639. doi: 10.1016/S0955-0674(98)80039-2
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Berger, J., and Moller, D. E. (2002). The mechanisms of action of PPARs. Annu. Rev. Med. 53, 409–435. doi: 10.1146/annurev.med.53.082901.104018
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Boonen, R. A., van Tijn, P., and Zivkovic, D. (2009). Wnt signaling in Alzheimer's disease: up or down, that is the question. Ageing Res. Rev. 8, 71–82. doi: 10.1016/j.arr.2008.11.003
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Brack, A. S., Conboy, M. J., Roy, S., Lee, M., Kuo, C. J., Keller, C., et al. (2007). Increased Wnt signaling during aging alters muscle stem cell fate and increases fibrosis. Science 317, 807–810. doi: 10.1126/science.1144090
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Bray, M. S., Shaw, C. A., Moore, M. W., Garcia, R. A., Zanquetta, M. M., Durgan, D. J., et al. (2008). Disruption of the circadian clock within the cardiomyocyte influences myocardial contractile function, metabolism, and gene expression. Am. J. Physiol. Heart. Circ. Physiol. 294, H1036–H1047. doi: 10.1152/ajpheart.01291.2007
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Breen, D. P., Vuono, R., Nawarathna, U., Fisher, K., Shneerson, J. M., Reddy, A. B., et al. (2014). Sleep and circadian rhythm regulation in early Parkinson disease. JAMA Neurol. 71, 589–595. doi: 10.1001/jamaneurol.2014.65
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Cai, Y., Liu, S., Sothern, R. B., Xu, S., and Chan, P. (2010). Expression of clock genes Per1 and Bmal1 in total leukocytes in health and Parkinson's disease. Eur. J. Neurol. 17, 550–554. doi: 10.1111/j.1468-1331.2009.02848.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Canalis, E. (2013). Wnt signalling in osteoporosis: mechanisms and novel therapeutic approaches. Nat. Rev. Endocrinol. 9, 575–583. doi: 10.1038/nrendo.2013.154
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Canaple, L., Rambaud, J., Dkhissi-Benyahya, O., Rayet, B., Tan, N. S., Michalik, L., et al. (2006). Reciprocal regulation of brain and muscle Arnt-like protein 1 and peroxisome proliferator-activated receptor alpha defines a novel positive feedback loop in the rodent liver circadian clock. Mol. Endocrinol. 20, 1715–1727. doi: 10.1210/me.2006-0052
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Cermakian, N., Lamont, E. W., Boudreau, P., and Boivin, D. B. (2011). Circadian clock gene expression in brain regions of Alzheimer's disease patients and control subjects. J. Biol. Rhythms 26, 160–170. doi: 10.1177/0748730410395732
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Chan, A. O., Jim, M. H., Lam, K. F., Morris, J. S., Siu, D. C., Tong, T., et al. (2007). Prevalence of colorectal neoplasm among patients with newly diagnosed coronary artery disease. JAMA 298, 1412–1419. doi: 10.1001/jama.298.12.1412
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Chatterjee, S., Nam, D., Guo, B., Kim, J. M., Winnier, G. E., Lee, J., et al. (2013). Brain and muscle Arnt-like 1 is a key regulator of myogenesis. J. Cell Sci. 126, 2213–2224. doi: 10.1242/jcs.120519
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Chen, L., and Yang, G. (2014). PPARs integrate the mammalian clock and energy metabolism. PPAR Res. 2014:653017. doi: 10.1155/2014/653017
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Clevers, H. (2006a). Colon cancer–understanding how NSAIDs work. N. Engl. J. Med. 354, 761–763. doi: 10.1056/NEJMcibr055457
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Clevers, H. (2006b). Wnt/beta-catenin signaling in development and disease. Cell 127, 469–480. doi: 10.1016/j.cell.2006.10.018
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Clevers, H., and Batlle, E. (2006). EphB/EphrinB receptors and Wnt signaling in colorectal cancer. Cancer Res. 66, 2–5. doi: 10.1158/0008-5472.CAN-05-3849
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Coppola, G., Marmolino, D., Lu, D., Wang, Q., Cnop, M., Rai, M., et al. (2009). Functional genomic analysis of frataxin deficiency reveals tissue-specific alterations and identifies the PPARgamma pathway as a therapeutic target in Friedreich's ataxia. Hum. Mol. Genet. 18, 2452–2461. doi: 10.1093/hmg/ddp183
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Corrado, D., Basso, C., Thiene, G., McKenna, W. J., Davies, M. J., Fontaliran, F., et al. (1997). Spectrum of clinicopathologic manifestations of arrhythmogenic right ventricular cardiomyopathy/dysplasia: a multicenter study. J. Am. Coll. Cardiol. 30, 1512–1520. doi: 10.1016/S0735-1097(97)00332-X
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Curtis, A. M., Cheng, Y., Kapoor, S., Reilly, D., Price, T. S., and Fitzgerald, G. A. (2007). Circadian variation of blood pressure and the vascular response to asynchronous stress. Proc. Natl. Acad. Sci. U.S.A. 104, 3450–3455. doi: 10.1073/pnas.0611680104
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
De Ferrari, G., Avila, M., Medina, M., Perez-Palma, E., Bustos, B., and Alarcon, M. (2014). Wnt/beta-Catenin Signaling in Alzheimer's Disease. CNS Neurol. Disord. Drug Targets 13, 745–754.
Desjardins, F., Sekkali, B., Verreth, W., Pelat, M., De Keyzer, D., Mertens, A., et al. (2008). Rosuvastatin increases vascular endothelial PPARgamma expression and corrects blood pressure variability in obese dyslipidaemic mice. Eur. Heart J. 29, 128–137. doi: 10.1093/eurheartj/ehm540
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Djouadi, F., Lecarpentier, Y., Hebert, J. L., Charron, P., Bastin, J., and Coirault, C. (2009). A potential link between PPAR signaling and the pathogenesis of arrhythmogenic right ventricular cardiomyopathy (ARVC). Cardiovasc. Res. 84, 83–90. doi: 10.1093/cvr/cvp183
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Drew, P. D., Xu, J., and Racke, M. K. (2008). PPAR-gamma: therapeutic potential for multiple sclerosis. PPAR Res. 2008:627463. doi: 10.1155/2008/627463.
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Druschky, A., Spitzer, A., Platsch, G., Claus, D., Feistel, H., Druschky, K., et al. (1999). Cardiac sympathetic denervation in early stages of amyotrophic lateral sclerosis demonstrated by 123I-MIBG-SPECT. Acta Neurol. Scand. 99, 308–314. doi: 10.1111/j.1600-0404.1999.tb00681.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Dupont, P., Besson, M. T., Devaux, J., and Lievens, J. C. (2012). Reducing canonical Wingless/Wnt signaling pathway confers protection against mutant Huntingtin toxicity in Drosophila. Neurobiol. Dis. 47, 237–247. doi: 10.1016/j.nbd.2012.04.007
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Duque, G., Li, W., Vidal, C., Bermeo, S., Rivas, D., and Henderson, J. (2013). Pharmacological inhibition of PPARgamma increases osteoblastogenesis and bone mass in male C57BL/6 mice. J. Bone Miner. Res. 28, 639–648. doi: 10.1002/jbmr.1782
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Durgan, D. J., Hotze, M. A., Tomlin, T. M., Egbejimi, O., Graveleau, C., Abel, E. D., et al. (2005). The intrinsic circadian clock within the cardiomyocyte. Am. J. Physiol. Heart Circ. Physiol. 289, H1530–H1541. doi: 10.1152/ajpheart.00406.2005
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Durgan, D. J., Moore, M. W., Ha, N. P., Egbejimi, O., Fields, A., Mbawuike, U., et al. (2007). Circadian rhythms in myocardial metabolism and contractile function: influence of workload and oleate. Am. J. Physiol. Heart Circ. Physiol. 293, H2385–H2393. doi: 10.1152/ajpheart.01361.2006
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Durgan, D. J., Trexler, N. A., Egbejimi, O., McElfresh, T. A., Suk, H. Y., Petterson, L. E., et al. (2006). The circadian clock within the cardiomyocyte is essential for responsiveness of the heart to fatty acids. J. Biol. Chem. 281, 24254–24269. doi: 10.1074/jbc.M601704200
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Farhat, G. N., and Cauley, J. A. (2008). The link between osteoporosis and cardiovascular disease. Clin. Cases Miner. Bone Metab. 5, 19–34.
Finck, B. N., Bernal-Mizrachi, C., Han, D. H., Coleman, T., Sambandam, N., LaRiviere, L. L., et al. (2005). A potential link between muscle peroxisome proliferator- activated receptor-alpha signaling and obesity-related diabetes. Cell Metab. 1, 133–144. doi: 10.1016/j.cmet.2005.01.006
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Finck, B. N., and Kelly, D. P. (2002). Peroxisome proliferator-activated receptor alpha (PPARalpha) signaling in the gene regulatory control of energy metabolism in the normal and diseased heart. J. Mol. Cell. Cardiol. 34, 1249–1257. doi: 10.1006/jmcc.2002.2061
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Finck, B. N., Lehman, J. J., Leone, T. C., Welch, M. J., Bennett, M. J., Kovacs, A., et al. (2002). The cardiac phenotype induced by PPARalpha overexpression mimics that caused by diabetes mellitus. J. Clin. Invest. 109, 121–130. doi: 10.1172/JCI0214080
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Fontaine, C., Dubois, G., Duguay, Y., Helledie, T., Vu-Dac, N., Gervois, P., et al. (2003). The orphan nuclear receptor Rev-Erbalpha is a peroxisome proliferator-activated receptor (PPAR) gamma target gene and promotes PPARgamma-induced adipocyte differentiation. J. Biol. Chem. 278, 37672–37680. doi: 10.1074/jbc.M304664200
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Fontaine, G., Fontaliran, F., Hebert, J. L., Chemla, D., Zenati, O., Lecarpentier, Y., et al. (1999). Arrhythmogenic right ventricular dysplasia. Annu. Rev. Med. 50, 17–35. doi: 10.1146/annurev.med.50.1.17
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Fressard, V., Duthoit, G., Donal, E., Probst, V., Deharo, J.-C., Chevalier, P., et al. (2010). Desmosomal genes analysis in arrhythmogenic right ventricular dysplasia/cardiomyopathy: spectrum of mutations and clinical impact in practice. Europace 12, 861–868. doi: 10.1093/europace/euq104
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Galli, C., Piemontese, M., Lumetti, S., Manfredi, E., Macaluso, G. M., and Passeri, G. (2013). GSK3b-inhibitor lithium chloride enhances activation of Wnt canonical signaling and osteoblast differentiation on hydrophilic titanium surfaces. Clin. Oral Implants Res. 24, 921–927. doi: 10.1111/j.1600-0501.2012.02488.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Garcia-Gras, E., Lombardi, R., Giocondo, M. J., Willerson, J. T., Schneider, M. D., Khoury, D. S., et al. (2006). Suppression of canonical Wnt/beta-catenin signaling by nuclear plakoglobin recapitulates phenotype of arrhythmogenic right ventricular cardiomyopathy. J. Clin. Invest. 116, 2012–2021. doi: 10.1172/JCI27751
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Gdynia, H. J., Kurt, A., Endruhn, S., Ludolph, A. C., and Sperfeld, A. D. (2006). Cardiomyopathy in motor neuron diseases. J. Neurol. Neurosurg. Psychiatry 77, 671–673. doi: 10.1136/jnnp.2005.078600
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Gekakis, N., Staknis, D., Nguyen, H. B., Davis, F. C., Wilsbacher, L. D., King, D. P., et al. (1998). Role of the CLOCK protein in the mammalian circadian mechanism. Science 280, 1564–1569. doi: 10.1126/science.280.5369.1564
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Gervois, P., Chopin-Delannoy, S., Fadel, A., Dubois, G., Kosykh, V., Fruchart, J. C., et al. (1999). Fibrates increase human REV-ERBalpha expression in liver via a novel peroxisome proliferator-activated receptor response element. Mol. Endocrinol. 13, 400–409.
Gibbons, G. F., Patel, D., Wiggins, D., and Knight, B. L. (2002). The functional efficiency of lipogenic and cholesterogenic gene expression in normal mice and in mice lacking the peroxisomal proliferator-activated receptor-alpha (PPAR-alpha). Adv. Enzyme Regul. 42, 227–247. doi: 10.1016/S0065-2571(01)00033-4
Godin, J. D., Poizat, G., Hickey, M. A., Maschat, F., and Humbert, S. (2010). Mutant huntingtin-impaired degradation of beta-catenin causes neurotoxicity in Huntington's disease. EMBO J. 29, 2433–2445. doi: 10.1038/emboj.2010.117
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Goldbeter, A. (2002). Computational approaches to cellular rhythms. Nature 420, 238–245. doi: 10.1038/nature01259
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Goldbeter, A. (2013). Origin of cyclicity in bipolar disorders: a computational approach. Pharmacopsychiatry 46(Suppl. 1), S44–S52. doi: 10.1055/s-0033-1341502
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Goodwin, B. C. (1965). Oscillatory behavior in enzymatic control processes. Adv. Enzyme Regul. 3, 425–438. doi: 10.1016/0065-2571(65)90067-1
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Gould, T. D., and Manji, H. K. (2002). The Wnt signaling pathway in bipolar disorder. Neuroscientist 8, 497–511. doi: 10.1177/107385802237176
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Green, C. B., Douris, N., Kojima, S., Strayer, C. A., Fogerty, J., Lourim, D., et al. (2007). Loss of Nocturnin, a circadian deadenylase, confers resistance to hepatic steatosis and diet-induced obesity. Proc. Natl. Acad. Sci. U.S.A. 104, 9888–9893. doi: 10.1073/pnas.0702448104
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Guo, B., Chatterjee, S., Li, L., Kim, J. M., Lee, J., Yechoor, V. K., et al. (2012). The clock gene, brain and muscle Arnt-like 1, regulates adipogenesis via Wnt signaling pathway. FASEB J. 26, 3453–3463. doi: 10.1096/fj.12-205781
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Haapaniemi, T. H., Pursiainen, V., Korpelainen, J. T., Huikuri, H. V., Sotaniemi, K. A., and Myllyla, V. V. (2001). Ambulatory ECG and analysis of heart rate variability in Parkinson's disease. J. Neurol. Neurosurg. Psychiatry 70, 305–310. doi: 10.1136/jnnp.70.3.305
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Hack, N., Fayad, S. M., Monari, E. H., Akbar, U., Hardwick, A., Rodriguez, R. L., et al. (2014). An eight-year clinic experience with clozapine use in a Parkinson's disease clinic setting. PLoS ONE 9:e91545. doi: 10.1371/journal.pone.0091545
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Halabi, C. M., Beyer, A. M., de Lange, W. J., Keen, H. L., Baumbach, G. L., Faraci, F. M., et al. (2008). Interference with PPAR gamma function in smooth muscle causes vascular dysfunction and hypertension. Cell Metab. 7, 215–226. doi: 10.1016/j.cmet.2007.12.008
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Hardin, P. E., Hall, J. C., and Rosbash, M. (1990). Feedback of the Drosophila period gene product on circadian cycling of its messenger RNA levels. Nature 343, 536–540. doi: 10.1038/343536a0
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Hebert, J. L., Chemla, D., Gerard, O., Zamani, K., Quillard, J., Azarine, A., et al. (2004). Angiographic right and left ventricular function in arrhythmogenic right ventricular dysplasia. Am. J. Cardiol. 93, 728–733. doi: 10.1016/j.amjcard.2003.11.055
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Hedgepeth, C. M., Conrad, L. J., Zhang, J., Huang, H. C., Lee, V. M., and Klein, P. S. (1997). Activation of the Wnt signaling pathway: a molecular mechanism for lithium action. Dev. Biol. 185, 82–91. doi: 10.1006/dbio.1997.8552
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Hennekens, C. H., Hennekens, A. R., Hollar, D., and Casey, D. E. (2005). Schizophrenia and increased risks of cardiovascular disease. Am. Heart J. 150, 1115–1121. doi: 10.1016/j.ahj.2005.02.007
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Heuberger, J., and Birchmeier, W. (2010). Interplay of cadherin-mediated cell adhesion and canonical wnt signaling. Cold Spring Harb. Perspect. Biol. 2:a002915. doi: 10.1101/cshperspect.a002915
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Hineno, T., Mizobuchi, M., Hiratani, K., Inami, Y., and Kakimoto, Y. (1992). Disappearance of circadian rhythms in Parkinson's disease model induced by 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine in dogs. Brain Res. 580, 92–99. doi: 10.1016/0006-8993(92)90930-8
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Hogenesch, J. B., Gu, Y. Z., Jain, S., and Bradfield, C. A. (1998). The basic-helix-loop-helix-PAS orphan MOP3 forms transcriptionally active complexes with circadian and hypoxia factors. Proc. Natl. Acad. Sci. U.S.A. 95, 5474–5479. doi: 10.1073/pnas.95.10.5474
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Hu, K., Ivanov, P., Hilton, M. F., Chen, Z., Ayers, R. T., Stanley, H. E., et al. (2004). Endogenous circadian rhythm in an index of cardiac vulnerability independent of changes in behavior. Proc. Natl. Acad. Sci. U.S.A. 101, 18223–18227. doi: 10.1073/pnas.0408243101
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Huss, J. M., Levy, F. H., and Kelly, D. P. (2001). Hypoxia inhibits the peroxisome proliferator-activated receptor alpha/retinoid X receptor gene regulatory pathway in cardiac myocytes: a mechanism for O2-dependent modulation of mitochondrial fatty acid oxidation. J. Biol. Chem. 276, 27605–27612. doi: 10.1074/jbc.M100277200
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Inestrosa, N. C., and Toledo, E. M. (2008). The role of Wnt signaling in neuronal dysfunction in Alzheimer's Disease. Mol. Neurodegener. 3:9. doi: 10.1186/1750-1326-3-9
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Inoue, I., Shinoda, Y., Ikeda, M., Hayashi, K., Kanazawa, K., Nomura, M., et al. (2005). CLOCK/BMAL1 is involved in lipid metabolism via transactivation of the peroxisome proliferator-activated receptor (PPAR) response element. J. Atheroscler. Thromb. 12, 169–174. doi: 10.5551/jat.12.169
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Ip, W., Chiang, Y. T., and Jin, T. (2012). The involvement of the wnt signaling pathway and TCF7L2 in diabetes mellitus: the current understanding, dispute, and perspective. Cell Biosci. 2:28. doi: 10.1186/2045-3701-2-28
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Ivanov, P., Hu, K., Hilton, M. F., Shea, S. A., and Stanley, H. E. (2007). Endogenous circadian rhythm in human motor activity uncoupled from circadian influences on cardiac dynamics. Proc. Natl. Acad. Sci. U.S.A. 104, 20702–20707. doi: 10.1073/pnas.0709957104
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Jadidi, E., Mohammadi, M., and Moradi, T. (2013). High risk of cardiovascular diseases after diagnosis of multiple sclerosis. Mult. Scler. 19, 1336–1340. doi: 10.1177/1352458513475833
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Jansson, E. A., Are, A., Greicius, G., Kuo, I. C., Kelly, D., Arulampalam, V., et al. (2005). The Wnt/beta-catenin signaling pathway targets PPARgamma activity in colon cancer cells. Proc. Natl. Acad. Sci. U.S.A. 102, 1460–1465. doi: 10.1073/pnas.0405928102
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Jiang, Q., Heneka, M., and Landreth, G. E. (2008). The role of peroxisome proliferator-activated receptor-gamma (PPARgamma) in Alzheimer's disease: therapeutic implications. CNS Drugs 22, 1–14. doi: 10.2165/00023210-200822010-00001
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Kawai, M., Green, C. B., Lecka-Czernik, B., Douris, N., Gilbert, M. R., Kojima, S., et al. (2010). A circadian-regulated gene, Nocturnin, promotes adipogenesis by stimulating PPAR-gamma nuclear translocation. Proc. Natl. Acad. Sci. U.S.A. 107, 10508–10513. doi: 10.1073/pnas.1000788107
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Kelly, D. P. (2003). PPARs of the heart: three is a crowd. Circ. Res. 92, 482–484. doi: 10.1161/01.RES.0000064382.46274.95
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Kiaei, M. (2008). Peroxisome proliferator-activated receptor-gamma in amyotrophic lateral sclerosis and Huntington's disease. PPAR Res. 2008:418765. doi: 10.1155/2008/418765.
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Kiriazis, H., Jennings, N. L., Davern, P., Lambert, G., Su, Y., Pang, T., et al. (2012). Neurocardiac dysregulation and neurogenic arrhythmias in a transgenic mouse model of Huntington's disease. J. Physiol. 590, 5845–5860. doi: 10.1113/jphysiol.2012.238113
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Klymkowsky, M. W., Williams, B. O., Barish, G. D., Varmus, H. E., and Vourgourakis, Y. E. (1999). Membrane-anchored plakoglobins have multiple mechanisms of action in Wnt signaling. Mol. Biol. Cell 10, 3151–3169. doi: 10.1091/mbc.10.10.3151
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Kolker, D. E., Fukuyama, H., Huang, D. S., Takahashi, J. S., Horton, T. H., and Turek, F. W. (2003). Aging alters circadian and light-induced expression of clock genes in golden hamsters. J. Biol. Rhythms 18, 159–169. doi: 10.1177/0748730403251802
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Korvala, J., Juppner, H., Makitie, O., Sochett, E., Schnabel, D., Mora, S., et al. (2012). Mutations in LRP5 cause primary osteoporosis without features of OI by reducing Wnt signaling activity. BMC Med. Genet. 13:26. doi: 10.1186/1471-2350-13-26
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Kremer, H. P., Roos, R. A., Dingjan, G. M., Bots, G. T., Bruyn, G. W., and Hofman, M. A. (1991). The hypothalamic lateral tuberal nucleus and the characteristics of neuronal loss in Huntington's disease. Neurosci. Lett. 132, 101–104. doi: 10.1016/0304-3940(91)90443-W
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Krishnan, J., Suter, M., Windak, R., Krebs, T., Felley, A., Montessuit, C., et al. (2009). Activation of a HIF1alpha-PPARgamma axis underlies the integration of glycolytic and lipid anabolic pathways in pathologic cardiac hypertrophy. Cell Metab. 9, 512–524. doi: 10.1016/j.cmet.2009.05.005
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Kume, K., Zylka, M. J., Sriram, S., Shearman, L. P., Weaver, D. R., Jin, X., et al. (1999). mCRY1 and mCRY2 are essential components of the negative limb of the circadian clock feedback loop. Cell 98, 193–205. doi: 10.1016/S0092-8674(00)81014-4
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Lan, M. J., Yuan, P., Chen, G., and Manji, H. K. (2008). Neuronal peroxisome proliferator-activated receptor gamma signaling: regulation by mood-stabilizer valproate. J. Mol. Neurosci. 35, 225–234. doi: 10.1007/s12031-008-9056-8
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Lecarpentier, Y., Claes, V., and Hebert, J. L. (2010). PPARs, cardiovascular metabolism, and function: near- or far-from-equilibrium pathways. PPAR Res. 2010:783273. doi: 10.1155/2010/783273
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Lee, D. K., Nathan Grantham, R., Trachte, A. L., Mannion, J. D., and Wilson, C. L. (2006). Activation of the canonical Wnt/beta-catenin pathway enhances monocyte adhesion to endothelial cells. Biochem. Biophys. Res. Commun. 347, 109–116. doi: 10.1016/j.bbrc.2006.06.082
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Leibetseder, V., Humpeler, S., Svoboda, M., Schmid, D., Thalhammer, T., Zuckermann, A., et al. (2009). Clock genes display rhythmic expression in human hearts. Chronobiol. Int. 26, 621–636. doi: 10.1080/07420520902924939
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Lemberger, T., Saladin, R., Vazquez, M., Assimacopoulos, F., Staels, B., Desvergne, B., et al. (1996). Expression of the peroxisome proliferator-activated receptor alpha gene is stimulated by stress and follows a diurnal rhythm. J. Biol. Chem. 271, 1764–1769. doi: 10.1074/jbc.271.3.1764
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Lemberger, T., Staels, B., Saladin, R., Desvergne, B., Auwerx, J., and Wahli, W. (1994). Regulation of the peroxisome proliferator-activated receptor alpha gene by glucocorticoids. J. Biol. Chem. 269, 24527–24530.
L'Episcopo, F., Serapide, M. F., Tirolo, C., Testa, N., Caniglia, S., Morale, M. C., et al. (2011). A Wnt1 regulated Frizzled-1/beta-Catenin signaling pathway as a candidate regulatory circuit controlling mesencephalic dopaminergic neuron-astrocyte crosstalk: therapeutical relevance for neuron survival and neuroprotection. Mol. Neurodegener. 6:49. doi: 10.1186/1750-1326-6-49
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
L'Episcopo, F., Tirolo, C., Testa, N., Caniglia, S., Morale, M. C., Deleidi, M., et al. (2012). Plasticity of subventricular zone neuroprogenitors in MPTP (1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine) mouse model of Parkinson's disease involves cross talk between inflammatory and Wnt/beta-catenin signaling pathways: functional consequences for neuroprotection and repair. J. Neurosci. 32, 2062–2085. doi: 10.1523/JNEUROSCI.5259-11.2012
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Liang, H., and Ward, W. F. (2006). PGC-1alpha: a key regulator of energy metabolism. Adv. Physiol. Educ. 30, 145–151. doi: 10.1152/advan.00052.2006
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Liu, C., Li, S., Liu, T., Borjigin, J., and Lin, J. D. (2007a). Transcriptional coactivator PGC-1alpha integrates the mammalian clock and energy metabolism. Nature 447, 477–481. doi: 10.1038/nature05767
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Liu, H., Fergusson, M. M., Castilho, R. M., Liu, J., Cao, L., Chen, J., et al. (2007b). Augmented Wnt signaling in a mammalian model of accelerated aging. Science 317, 803–806. doi: 10.1126/science.1143578
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Lockyer, P., Schisler, J. C., Patterson, C., and Willis, M. S. (2009). Minireview: won't get fooled again: the nonmetabolic roles of peroxisome proliferator-activated receptors (PPARs) in the heart. Mol. Endocrinol. 24, 1111–1119. doi: 10.1210/me.2009-0374
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Lo Coco, D., Mattaliano, P., Spataro, R., Mattaliano, A., and La Bella, V. (2011). Sleep-wake disturbances in patients with amyotrophic lateral sclerosis. J. Neurol. Neurosurg. Psychiatry 82, 839–842. doi: 10.1136/jnnp.2010.228007
Lu, D., and Carson, D. A. (2010). Repression of beta-catenin signaling by PPAR gamma ligands. Eur. J. Pharmacol. 636, 198–202. doi: 10.1016/j.ejphar.2010.03.010
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
MacDonald, B. T., Tamai, K., and He, X. (2009). Wnt/beta-catenin signaling: components, mechanisms, and diseases. Dev. Cell 17, 9–26. doi: 10.1016/j.devcel.2009.06.016
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Maeda, O., Usami, N., Kondo, M., Takahashi, M., Goto, H., Shimokata, K., et al. (2004). Plakoglobin (gamma-catenin) has TCF/LEF family-dependent transcriptional activity in beta-catenin-deficient cell line. Oncogene 23, 964–972. doi: 10.1038/sj.onc.1207254
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Marcheva, B., Ramsey, K. M., Buhr, E. D., Kobayashi, Y., Su, H., Ko, C. H., et al. (2010). Disruption of the clock components CLOCK and BMAL1 leads to hypoinsulinaemia and diabetes. Nature 466, 627–631. doi: 10.1038/nature09253
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Marcus, F. I., Fontaine, G. H., Guiraudon, G., Frank, R., Laurenceau, J. L., Malergue, C., et al. (1982). Right ventricular dysplasia: a report of 24 adult cases. Circulation 65, 384–398. doi: 10.1161/01.CIR.65.2.384
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Marmolino, D., Acquaviva, F., Pinelli, M., Monticelli, A., Castaldo, I., Filla, A., et al. (2009). PPAR-gamma agonist Azelaoyl PAF increases frataxin protein and mRNA expression: new implications for the Friedreich's ataxia therapy. Cerebellum 8, 98–103. doi: 10.1007/s12311-008-0087-z
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Maywood, E. S., Fraenkel, E., McAllister, C. J., Wood, N., Reddy, A. B., Hastings, M. H., et al. (2010). Disruption of peripheral circadian timekeeping in a mouse model of Huntington's disease and its restoration by temporally scheduled feeding. J. Neurosci. 30, 10199–10204. doi: 10.1523/JNEUROSCI.1694-10.2010
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
McNamara, P., Seo, S. B., Rudic, R. D., Sehgal, A., Chakravarti, D., and FitzGerald, G. A. (2001). Regulation of CLOCK and MOP4 by nuclear hormone receptors in the vasculature: a humoral mechanism to reset a peripheral clock. Cell 105, 877–889. doi: 10.1016/S0092-8674(01)00401-9
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Meshorer, E., and Gruenbaum, Y. (2008). Gone with the Wnt/Notch: stem cells in laminopathies, progeria, and aging. J. Cell Biol. 181, 9–13. doi: 10.1083/jcb.200802155
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Mihm, M. J., Amann, D. M., Schanbacher, B. L., Altschuld, R. A., Bauer, J. A., and Hoyt, K. R. (2007). Cardiac dysfunction in the R6/2 mouse model of Huntington's disease. Neurobiol. Dis. 25, 297–308. doi: 10.1016/j.nbd.2006.09.016
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Moldes, M., Zuo, Y., Morrison, R. F., Silva, D., Park, B. H., Liu, J., et al. (2003). Peroxisome-proliferator-activated receptor gamma suppresses Wnt/beta-catenin signalling during adipogenesis. Biochem. J. 376, 607–613. doi: 10.1042/BJ20030426
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Moon, R. T., Bowerman, B., Boutros, M., and Perrimon, N. (2002). The promise and perils of Wnt signaling through beta-catenin. Science 296, 1644–1646. doi: 10.1126/science.1071549
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Morin, P. J., Sparks, A. B., Korinek, V., Barker, N., Clevers, H., Vogelstein, B., et al. (1997). Activation of beta-catenin-Tcf signaling in colon cancer by mutations in beta-catenin or APC. Science 275, 1787–1790. doi: 10.1126/science.275.5307.1787
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Musiek, E. S., Lim, M. M., Yang, G., Bauer, A. Q., Qi, L., Lee, Y., et al. (2013). Circadian clock proteins regulate neuronal redox homeostasis and neurodegeneration. J. Clin. Invest. 123, 5389–5400. doi: 10.1172/JCI70317
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Naito, A. T., Shiojima, I., and Komuro, I. (2010). Wnt signaling and aging-related heart disorders. Circ. Res. 107, 1295–1303. doi: 10.1161/CIRCRESAHA.110.223776
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Najdi, R., Holcombe, R. F., and Waterman, M. L. (2011). Wnt signaling and colon carcinogenesis: beyond APC. J. Carcinog. 10:5. doi: 10.4103/1477-3163.78111
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Narravula, S., and Colgan, S. P. (2001). Hypoxia-inducible factor 1-mediated inhibition of peroxisome proliferator-activated receptor alpha expression during hypoxia. J. Immunol. 166, 7543–7548. doi: 10.4049/jimmunol.166.12.7543
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Nicol, C. J., Adachi, M., Akiyama, T. E., and Gonzalez, F. J. (2005). PPARgamma in endothelial cells influences high fat diet-induced hypertension. Am. J. Hypertens. 18, 549–556. doi: 10.1016/j.amjhyper.2004.10.032
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Nonaka, H., Emoto, N., Ikeda, K., Fukuya, H., Rohman, M. S., Raharjo, S. B., et al. (2001). Angiotensin II induces circadian gene expression of clock genes in cultured vascular smooth muscle cells. Circulation 104, 1746–1748. doi: 10.1161/hc4001.098048
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Oishi, K., Shirai, H., and Ishida, N. (2005). CLOCK is involved in the circadian transactivation of peroxisome-proliferator-activated receptor alpha (PPARalpha) in mice. Biochem. J. 386, 575–581. doi: 10.1042/BJ20041150
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Oishi, K., Shirai, H., and Ishida, N. (2008). PPARalpha is involved in photoentrainment of the circadian clock. Neuroreport 19, 487–489. doi: 10.1097/WNR.0b013e3282f7968f
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
O'Neal, W. T., Griffin, W. F., Dries-Devlin, J. L., Kent, S. D., Chen, J., Willis, M. S., et al. (2013). Ephrin-Eph signaling as a potential therapeutic target for the treatment of myocardial infarction. Med. Hypotheses 80, 738–744. doi: 10.1016/j.mehy.2013.02.024
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Pallier, P. N., and Morton, A. J. (2009). Management of sleep/wake cycles improves cognitive function in a transgenic mouse model of Huntington's disease. Brain Res. 1279, 90–98. doi: 10.1016/j.brainres.2009.03.072
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Paschos, G. K., Ibrahim, S., Song, W. L., Kunieda, T., Grant, G., Reyes, T. M., et al. (2012). Obesity in mice with adipocyte-specific deletion of clock component Arntl. Nat. Med. 18, 1768–1777. doi: 10.1038/nm.2979
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Patacchioli, F. R., Monnazzi, P., Scontrini, A., Tremante, E., Caridi, I., Brunetti, E., et al. (2003). Adrenal dysregulation in amyotrophic lateral sclerosis. J. Endocrinol. Invest. 26, RC23–RC25. doi: 10.1007/BF03349149
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Patel, D. D., Knight, B. L., Wiggins, D., Humphreys, S. M., and Gibbons, G. F. (2001). Disturbances in the normal regulation of SREBP-sensitive genes in PPAR alpha-deficient mice. J. Lipid Res. 42, 328–337.
Pinto, C., Cardenas, P., Osses, N., and Henriquez, J. P. (2013). Characterization of Wnt/beta-catenin and BMP/Smad signaling pathways in an in vitro model of amyotrophic lateral sclerosis. Front. Cell. Neurosci. 7:239. doi: 10.3389/fncel.2013.00239
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Postuma, R. B., and Montplaisir, J. (2009). Predicting Parkinson's disease - why, when, and how? Parkinsonism Relat. Disord. 15(Suppl. 3), S105–S109. doi: 10.1016/S1353-8020(09)70793-X
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Preitner, N., Damiola, F., Lopez-Molina, L., Zakany, J., Duboule, D., Albrecht, U., et al. (2002). The orphan nuclear receptor REV-ERBalpha controls circadian transcription within the positive limb of the mammalian circadian oscillator. Cell 110, 251–260. doi: 10.1016/S0092-8674(02)00825-5
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Prigogine, I., Nicolis, G., and Babloyantz, A. (1974). Nonequilibrium problems in biological phenomena. Ann. N.Y. Acad. Sci. 231, 99–105. doi: 10.1111/j.1749-6632.1974.tb20557.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Rawadi, G., and Roman-Roman, S. (2005). Wnt signalling pathway: a new target for the treatment of osteoporosis. Expert Opin. Ther. Targets 9, 1063–1077. doi: 10.1517/14728222.9.5.1063
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Razeghi, P., Young, M. E., Abbasi, S., and Taegtmeyer, H. (2001). Hypoxia in vivo decreases peroxisome proliferator-activated receptor alpha-regulated gene expression in rat heart. Biochem. Biophys. Res. Commun. 287, 5–10. doi: 10.1006/bbrc.2001.5541
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Reppert, S. M., and Weaver, D. R. (2001). Molecular analysis of mammalian circadian rhythms. Annu. Rev. Physiol. 63, 647–676. doi: 10.1146/annurev.physiol.63.1.647
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Reppert, S. M., and Weaver, D. R. (2002). Coordination of circadian timing in mammals. Nature 418, 935–941. doi: 10.1038/nature00965
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Richardson, P., McKenna, W., Bristow, M., Maisch, B., Mautner, B., O'Connell, J., et al. (1996). Report of the 1995 World Health Organization/International Society and Federation of Cardiology Task Force on the Definition and Classification of cardiomyopathies. Circulation 93, 841–842. doi: 10.1161/01.CIR.93.5.841
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Ross, O. A., Rutherford, N. J., Baker, M., Soto-Ortolaza, A. I., Carrasquillo, M. M., DeJesus-Hernandez, M., et al. (2011). Ataxin-2 repeat-length variation and neurodegeneration. Hum. Mol. Genet. 20, 3207–3212. doi: 10.1093/hmg/ddr227
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Ross, S. E., Hemati, N., Longo, K. A., Bennett, C. N., Lucas, P. C., Erickson, R. L., et al. (2000). Inhibition of adipogenesis by Wnt signaling. Science 289, 950–953. doi: 10.1126/science.289.5481.950
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Rudic, R. D., and Fulton, D. J. (2009). Pressed for time: the circadian clock and hypertension. J. Appl. Physiol. 107, 1328–1338. doi: 10.1152/japplphysiol.00661.2009
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Ryan, M. J., Didion, S. P., Mathur, S., Faraci, F. M., and Sigmund, C. D. (2004). PPAR(gamma) agonist rosiglitazone improves vascular function and lowers blood pressure in hypertensive transgenic mice. Hypertension 43, 661–666. doi: 10.1161/01.HYP.0000116303.71408.c2
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Sakai, T., Tamura, T., Kitamoto, T., and Kidokoro, Y. (2004). A clock gene, period, plays a key role in long-term memory formation in Drosophila. Proc. Natl. Acad. Sci. U.S.A. 101, 16058–16063. doi: 10.1073/pnas.0401472101
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Sarraf, P., Mueller, E., Smith, W. M., Wright, H. M., Kum, J. B., Aaltonen, L. A., et al. (1999). Loss-of-function mutations in PPAR gamma associated with human colon cancer. Mol. Cell 3, 799–804. doi: 10.1016/S1097-2765(01)80012-5
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Savvidis, C., and Koutsilieris, M. (2012). Circadian rhythm disruption in cancer biology. Mol. Med. 18, 1249–1260. doi: 10.2119/molmed.2012.00077
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Schibler, U., and Sassone-Corsi, P. (2002). A web of circadian pacemakers. Cell 111, 919–922. doi: 10.1016/S0092-8674(02)01225-4
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Schintu, N., Frau, L., Ibba, M., Caboni, P., Garau, A., Carboni, E., et al. (2009). PPAR-gamma-mediated neuroprotection in a chronic mouse model of Parkinson's disease. Eur. J. Neurosci. 29, 954–963. doi: 10.1111/j.1460-9568.2009.06657.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Schroeder, A. M., Loh, D. H., Jordan, M. C., Roos, K. P., and Colwell, C. S. (2011). Baroreceptor reflex dysfunction in the BACHD mouse model of Huntington's disease. PLoS Curr 3:RRN1266. doi: 10.1371/currents.RRN1266
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Sen-Chowdhry, S., Syrris, P., and McKenna, W. J. (2005). Genetics of right ventricular cardiomyopathy. J. Cardiovasc. Electrophysiol. 16, 927–935. doi: 10.1111/j.1540-8167.2005.40842.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Shirai, H., Oishi, K., Kudo, T., Shibata, S., and Ishida, N. (2007). PPARalpha is a potential therapeutic target of drugs to treat circadian rhythm sleep disorders. Biochem. Biophys. Res. Commun. 357, 679–682. doi: 10.1016/j.bbrc.2007.04.002
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Singh, K. K. (2013). An emerging role for Wnt and GSK3 signaling pathways in schizophrenia. Clin. Genet. 83, 511–517. doi: 10.1111/cge.12111
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Sinha, D., Wang, Z., Ruchalski, K. L., Levine, J. S., Krishnan, S., Lieberthal, W., et al. (2005). Lithium activates the Wnt and phosphatidylinositol 3-kinase Akt signaling pathways to promote cell survival in the absence of soluble survival factors. Am. J. Physiol. Renal Physiol. 288, F703–F713. doi: 10.1152/ajprenal.00189.2004
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Stampfer, M. J. (2006). Cardiovascular disease and Alzheimer's disease: common links. J. Intern. Med. 260, 211–223. doi: 10.1111/j.1365-2796.2006.01687.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Storch, K. F., Lipan, O., Leykin, I., Viswanathan, N., Davis, F. C., Wong, W. H., et al. (2002). Extensive and divergent circadian gene expression in liver and heart. Nature 417, 78–83. doi: 10.1038/nature744
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Sugawara, A., Uruno, A., Kudo, M., Matsuda, K., Yang, C. W., and Ito, S. (2010). Effects of PPARgamma on hypertension, atherosclerosis, and chronic kidney disease. Endocr. J. 57, 847–852. doi: 10.1507/endocrj.K10E-281
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Swanson, C. R., Joers, V., Bondarenko, V., Brunner, K., Simmons, H. A., Ziegler, T. E., et al. (2011). The PPAR-gamma agonist pioglitazone modulates inflammation and induces neuroprotection in parkinsonian monkeys. J. Neuroinflammation 8:91. doi: 10.1186/1742-2094-8-91
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Swartz, H. A., and Fagiolini, A. (2012). Cardiovascular disease and bipolar disorder: risk and clinical implications. J. Clin. Psychiatry 73, 1563–1565. doi: 10.4088/JCP.12ac08227
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Takada, I., Kouzmenko, A. P., and Kato, S. (2009). Wnt and PPARgamma signaling in osteoblastogenesis and adipogenesis. Nat. Rev. Rheumatol. 5, 442–447. doi: 10.1038/nrrheum.2009.137
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Takeda, N., and Maemura, K. (2011). Circadian clock and cardiovascular disease. J. Cardiol. 57, 249–256. doi: 10.1016/j.jjcc.2011.02.006
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Takeda, N., Maemura, K., Horie, S., Oishi, K., Imai, Y., Harada, T., et al. (2007). Thrombomodulin is a clock-controlled gene in vascular endothelial cells. J. Biol. Chem. 282, 32561–32567. doi: 10.1074/jbc.M705692200
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Tougas, G. (2000). The autonomic nervous system in functional bowel disorders. Gut 47(Suppl. 4), iv78–iv80. discussion: iv87.
Ueda, H. R., Chen, W., Adachi, A., Wakamatsu, H., Hayashi, S., Takasugi, T., et al. (2002). A transcription factor response element for gene expression during circadian night. Nature 418, 534–539. doi: 10.1038/nature00906
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Valvezan, A. J., and Klein, P. S. (2012). GSK-3 and Wnt signaling in neurogenesis and bipolar disorder. Front. Mol. Neurosci. 5:1. doi: 10.3389/fnmol.2012.00001
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Viola, A. U., James, L. M., Archer, S. N., and Dijk, D. J. (2008). PER3 polymorphism and cardiac autonomic control: effects of sleep debt and circadian phase. Am. J. Physiol. Heart Circ. Physiol. 295, H2156–H2163. doi: 10.1152/ajpheart.00662.2008
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Virag, J. A., Anderson, E. J., Kent, S. D., Blanton, H. D., Johnson, T. L., Moukdar, F., et al. (2013). Cardioprotection via preserved mitochondrial structure and function in the mPer2-mutant mouse myocardium. Am. J. Physiol. Heart Circ. Physiol. 305, H477–H483. doi: 10.1152/ajpheart.00914.2012
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Walker, F. O. (2007). Huntington's disease. Lancet 369, 218–228. doi: 10.1016/S0140-6736(07)60111-1
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Wan, W., Xia, S., Kalionis, B., Liu, L., and Li, Y. (2014). The role of Wnt signaling in the development of Alzheimer's disease: a potential therapeutic target? Biomed. Res. Int. 2014:301575. doi: 10.1155/2014/301575
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Wan, Y., Chong, L. W., and Evans, R. M. (2007). PPAR-gamma regulates osteoclastogenesis in mice. Nat. Med. 13, 1496–1503. doi: 10.1038/nm1672
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Wang, J., Yin, L., and Lazar, M. A. (2006). The orphan nuclear receptor Rev-erb alpha regulates circadian expression of plasminogen activator inhibitor type 1. J. Biol. Chem. 281, 33842–33848. doi: 10.1074/jbc.M607873200
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Wang, N., Yang, G., Jia, Z., Zhang, H., Aoyagi, T., Soodvilai, S., et al. (2008). Vascular PPARgamma controls circadian variation in blood pressure and heart rate through Bmal1. Cell Metab. 8, 482–491. doi: 10.1016/j.cmet.2008.10.009
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Wang, X., Xiao, Y., Mou, Y., Zhao, Y., Blankesteijn, W. M., and Hall, J. L. (2002). A role for the beta-catenin/T-cell factor signaling cascade in vascular remodeling. Circ. Res. 90, 340–347. doi: 10.1161/hh0302.104466
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Weaver, D. R. (1998). The suprachiasmatic nucleus: a 25-year retrospective. J. Biol. Rhythms 13, 100–112. doi: 10.1177/074873098128999952
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Woon, P. Y., Kaisaki, P. J., Braganca, J., Bihoreau, M. T., Levy, J. C., Farrall, M., et al. (2007). Aryl hydrocarbon receptor nuclear translocator-like (BMAL1) is associated with susceptibility to hypertension and type 2 diabetes. Proc. Natl. Acad. Sci. U.S.A. 104, 14412–14417. doi: 10.1073/pnas.0703247104
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Wu, X., Liu, Z., Shi, G., Xing, L., Wang, X., Gu, X., et al. (2011). The circadian clock influences heart performance. J. Biol. Rhythms 26, 402–411. doi: 10.1177/0748730411414168
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Wulff, K., Dijk, D. J., Middleton, B., Foster, R. G., and Joyce, E. M. (2012). Sleep and circadian rhythm disruption in schizophrenia. Br. J. Psychiatry 200, 308–316. doi: 10.1192/bjp.bp.111.096321
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Yang, G., Jia, Z., Aoyagi, T., McClain, D., Mortensen, R. M., and Yang, T. (2012). Systemic PPARgamma deletion impairs circadian rhythms of behavior and metabolism. PLoS ONE 7:e38117. doi: 10.1371/journal.pone.0038117
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Yang, T., Aoyagi, T., Dowtown, M., Liu, M., Whitehead, K., and Jia, Z. (2013). Whole-body deletion of PPAR in mice increases sympathetic nerve activity. Circulation 62:A399.
Yang, X., Downes, M., Yu, R. T., Bookout, A. L., He, W., Straume, M., et al. (2006). Nuclear receptor expression links the circadian clock to metabolism. Cell 126, 801–810. doi: 10.1016/j.cell.2006.06.050
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Yang, Y. (2012). Wnt signaling in development and disease. Cell Biosci. 2:14. doi: 10.1186/2045-3701-2-14
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Yin, L., Wang, J., Klein, P. S., and Lazar, M. A. (2006). Nuclear receptor Rev-erbalpha is a critical lithium-sensitive component of the circadian clock. Science 311, 1002–1005. doi: 10.1126/science.1121613
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Young, M. E. (2009). Anticipating anticipation: pursuing identification of cardiomyocyte circadian clock function. J. Appl. Physiol. 107, 1339–1347. doi: 10.1152/japplphysiol.00473.2009
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Young, M. E., Laws, F. A., Goodwin, G. W., and Taegtmeyer, H. (2001a). Reactivation of peroxisome proliferator-activated receptor alpha is associated with contractile dysfunction in hypertrophied rat heart. J. Biol. Chem. 276, 44390–44395. doi: 10.1074/jbc.M103826200
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Young, M. E., Razeghi, P., Cedars, A. M., Guthrie, P. H., and Taegtmeyer, H. (2001b). Intrinsic diurnal variations in cardiac metabolism and contractile function. Circ. Res. 89, 1199–1208. doi: 10.1161/hh2401.100741
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Young, M. E., Razeghi, P., and Taegtmeyer, H. (2001c). Clock genes in the heart: characterization and attenuation with hypertrophy. Circ. Res. 88, 1142–1150. doi: 10.1161/hh1101.091190
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Young, M. E., Wilson, C. R., Razeghi, P., Guthrie, P. H., and Taegtmeyer, H. (2002). Alterations of the circadian clock in the heart by streptozotocin-induced diabetes. J. Mol. Cell. Cardiol. 34, 223–231. doi: 10.1006/jmcc.2001.1504
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Young, M. W., and Kay, S. A. (2001). Time zones: a comparative genetics of circadian clocks. Nat. Rev. Genet. 2, 702–715. doi: 10.1038/35088576
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Yu, Y., Zhang, Z. H., Wei, S. G., Weiss, R. M., and Felder, R. B. (2012). Peroxisome proliferator-activated receptor-gamma regulates inflammation and renin-angiotensin system activity in the hypothalamic paraventricular nucleus and ameliorates peripheral manifestations of heart failure. Hypertension 59, 477–484. doi: 10.1161/HYPERTENSIONAHA.111.182345
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Yuan, S., Shi, Y., and Tang, S. J. (2012). Wnt signaling in the pathogenesis of multiple sclerosis-associated chronic pain. J. Neuroimmune Pharmacol. 7, 904–913. doi: 10.1007/s11481-012-9370-3
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Yuhara, H., Steinmaus, C., Cohen, S. E., Corley, D. A., Tei, Y., and Buffler, P. A. (2011). Is diabetes mellitus an independent risk factor for colon cancer and rectal cancer? Am. J. Gastroenterol. 106, 1911–1192. doi: 10.1038/ajg.2011.301
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Zhu, L., Wang, Q., Zhang, L., Fang, Z., Zhao, F., Lv, Z., et al. (2010). Hypoxia induces PGC-1alpha expression and mitochondrial biogenesis in the myocardium of TOF patients. Cell Res. 20, 676–687. doi: 10.1038/cr.2010.46
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Zhurinsky, J., Shtutman, M., and Ben-Ze'ev, A. (2000). Differential mechanisms of LEF/TCF family-dependent transcriptional activation by beta-catenin and plakoglobin. Mol. Cell. Biol. 20, 4238–4252. doi: 10.1128/MCB.20.12.4238-4252.2000
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Zylka, M. J., Shearman, L. P., Weaver, D. R., and Reppert, S. M. (1998). Three period homologs in mammals: differential light responses in the suprachiasmatic circadian clock and oscillating transcripts outside of brain. Neuron 20, 1103–1110. doi: 10.1016/S0896-6273(00)80492-4
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Keywords: circadian rhythms, Wnt/beta-catenin, PPAR, diabetes, hypertension, myocardial ischemia, neurodegenerative diseases, colon cancer
Citation: Lecarpentier Y, Claes V, Duthoit G and Hébert J-L (2014) Circadian rhythms, Wnt/beta-catenin pathway and PPAR alpha/gamma profiles in diseases with primary or secondary cardiac dysfunction. Front. Physiol. 5:429. doi: 10.3389/fphys.2014.00429
Received: 16 July 2014; Accepted: 15 October 2014;
Published online: 04 November 2014.
Edited by:
Jitka A. I. Virag, Brody School of Medicine, USAReviewed by:
James Todd Pearson, Monash University, AustraliaMing Gong, University of Kentucky, USA
Copyright © 2014 Lecarpentier, Claes, Duthoit and Hébert. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Yves Lecarpentier, Centre de Recherche Clinique, Hôpital de Meaux, 6-8 rue Saint Fiacre, 77100 Meaux, France e-mail: yves.c.lecarpentier@gmail.com