 Shivaprakash J. Mutt1,2*
Shivaprakash J. Mutt1,2* Elina Hyppönen3,4,5
Elina Hyppönen3,4,5 Juha Saarnio6Marjo-Riitta Järvelin2,7,8,9
Juha Saarnio6Marjo-Riitta Järvelin2,7,8,9 Karl-Heinz Herzig1,2,10*
Karl-Heinz Herzig1,2,10*- 1Department of Physiology, Institute of Biomedicine, University of Oulu, Oulu, Finland
- 2Biocenter of Oulu, University of Oulu, Oulu, Finland
- 3School of Population Health and Sansom Institute, University of South Australia, Adelaide, SA, Australia
- 4South Australian Health and Medical Research Institute, Adelaide, SA, Australia
- 5Population, Policy and Practice, Institute of Child Health, University College London, London, UK
- 6Department of Surgery, Oulu University Hospital, University of Oulu, Oulu, Finland
- 7Unit of Primary Care, Institute of Health Sciences, University of Oulu, Oulu University Hospital, Oulu, Finland
- 8Department of Children, Young People and Families, National Institute for Health and Welfare, Oulu, Finland
- 9Department of Epidemiology and Biostatistics, and MRC-PHE Center for Environment and Health, School of Public Health, Imperial College London, London, UK
- 10Medical Research Center Oulu and Oulu University Hospital, Oulu, Finland
The pandemic increase in obesity is inversely associated with vitamin D levels. While a higher BMI was causally related to lower 25-hydroxyvitamin D (25(OH)D), no evidence was obtained for a BMI lowering effect by higher 25(OH)D. Some of the physiological functions of 1,25(OH)2D3 (1,25-dihydroxycholecalciferol or calcitriol) via its receptor within the adipose tissue have been investigated such as its effect on energy balance, adipogenesis, adipokine, and cytokine secretion. Adipose tissue inflammation has been recognized as the key component of metabolic disorders, e.g., in the metabolic syndrome. The adipose organ secretes more than 260 different proteins/peptides. However, the molecular basis of the interactions of 1,25(OH)2D3, vitamin D binding proteins (VDBPs) and nuclear vitamin D receptor (VDR) after sequestration in adipose tissue and their regulations are still unclear. 1,25(OH)2D3 and its inactive metabolites are known to inhibit the formation of adipocytes in mouse 3T3-L1 cell line. In humans, 1,25(OH)2D3 promotes preadipocyte differentiation under cell culture conditions. Further evidence of its important functions is given by VDR knock out (VDR−/−) and CYP27B1 knock out (CYP27B1 −/−) mouse models: Both VDR−/− and CYP27B1−/− models are highly resistant to the diet induced weight gain, while the specific overexpression of human VDR in adipose tissue leads to increased adipose tissue mass. The analysis of microarray datasets from human adipocytes treated with macrophage-secreted products up-regulated VDR and CYP27B1 genes indicating the capacity of adipocytes to even produce active 1,25(OH)2D3. Experimental studies demonstrate that 1,25(OH)2D3 has an active role in adipose tissue by modulating inflammation, adipogenesis and adipocyte secretion. Yet, further in vivo studies are needed to address the effects and the effective dosages of vitamin D in human adipose tissue and its relevance in the associated diseases.
Introduction
Adipose tissue is no longer regarded as a simple storage organ since it has been convincingly shown that it secretes more than 260 different proteins/peptides (Lehr et al., 2012). Lean people have about 5 kg of adipose tissue, while in obese and severely obese individuals the adipose tissue/organ could amount to 50 kg or more (Frankenfield et al., 2001). Excess in adipose tissue has been attributed to a variety of diseases including cancer, diabetes, cardiovascular and neurodegenerative diseases and decrease in life expectancy (Adams et al., 2006; Despres and Lemieux, 2006; Kahn et al., 2006; Van Gaal et al., 2006). Adiposity is one of the most serious public health problems, associated with vitamin D insufficiency due to the decreased bioavailability of vitamin D3 (Wortsman et al., 2000). The Institute of Medicine (IOM) recommended 25-hydroxyvitamin D (25(OH)D) levels as reliable biomarker for assessment of Vitamin D status; currently values ≤50 nmol/l or ≤20 ng/ml are considered inadequate or not sufficient and values ≥50 nmol/l or ≥20 ng/ml as adequate or sufficient (Ross et al., 2011) (Figure 1). 25(OH)D levels have been determined by a variety of methods yielding different results. The National Institutes of Health’s Office of Dietary Supplements together with National Institute of Standards and Technology (NIST) therefore developed a standard reference material-972 (SRM-972) for accuracy of laboratory vitamin D measurements (Phinney et al., 2012). A recent study by the D-CarDia consortium employed a Mendelian randomization (MR) approach to establish causality and direction of the association between vitamin D status and obesity measured by body mass index (BMI) using information from 21 adult cohorts (up to 42,024 participants) (Vimaleswaran et al., 2013). The consortium found that a higher BMI was causally related to lower 25(OH)D; no evidence was obtained for a BMI lowering effect of higher 25(OH)D. However, the study did not provide insights into the cellular action of 1,25(OH)2D3 (1,25-dihydroxycholecalciferol or calcitriol). While the knowledge of the effects of 1,25(OH)2D3 as an essential hormone and transcription factor is further emerging, it is increasingly acknowledged that 1,25(OH)2D3 down regulates inflammatory responses in the adipose tissue. The anti-inflammatory effects of 1,25(OH)2D3 might have notable influences on population health and disease prevention, since inflammation is thought to be the underlying cause of a range of metabolic disorders (Hotamisligil, 2006; Huotari and Herzig, 2008; Vlasova et al., 2010).
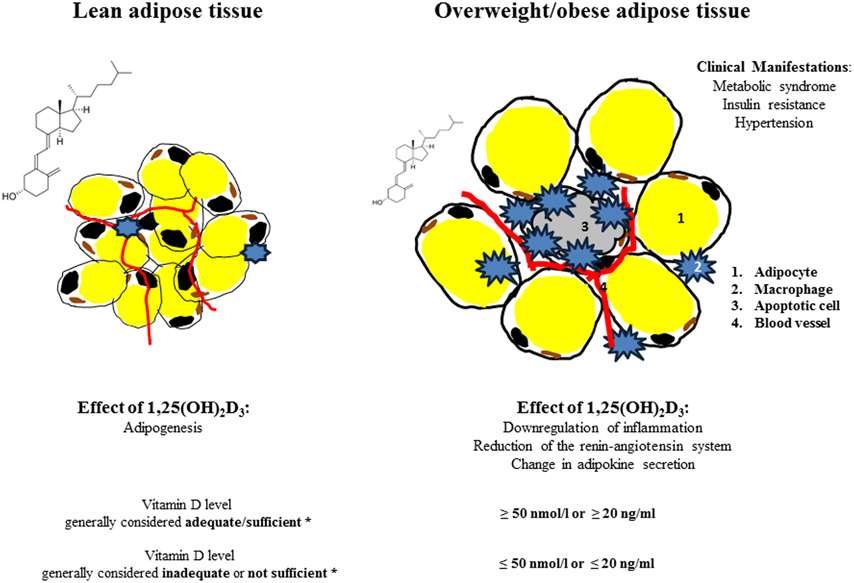
Figure 1. Effect of 1,25(OH)2D3 on adipose tissue/lean and obese state. *Vitamin D levels (25(OH)D) according to the Institute of Medicine, Food and Nutrition Board. Dietary Reference Intakes for Calcium and Vitamin D. Washington, DC: National Academy Press, 2011.
Vitamin D and Adipogenesis
Adipose tissue expansion is a remarkable process characterized by the enlargement of adipocyte size known as hypertrophy and by the increase in the number of adipocytes known as hyperplasia, which is more strongly associated with severity of obesity (Arner and Spalding, 2010). Both processes emerge through sequential stages of differentiation to form mature adipocytes; this process is called adipogenesis. Mesodermal cells are influenced by various signals like bone morphogenetic proteins (BMPs), fibroblast growth factors (FGFs), transforming-growth factor β (TGFβ) and insulin like growth factor 1 (IGF1) to form preadipocytes (Lowe et al., 2011). Furthermore, preadipocytes undergo differentiation to mature adipocytes by several intracellular signaling molecules (Figure 2) including janus kinase-signal transducer and activator of transcription 3 (JAK-STAT3) (Zhang et al., 2011), glutathione (Vigilanza et al., 2011), SMAD proteins (Jin et al., 2006) and ribosomal protein S6 kinase 1 (S6K1) (Carnevalli et al., 2010) affecting adipogenic transcription factors. In preadipocytes, differentiation factors need to be released from their suppressive signaling molecules such as members of wingless (WNT) family (Ross et al., 2000), protein of the retinoblastoma (Rb) family (Scime et al., 2005), preadipocyte factor 1 (Pref1) (Smas and Sul, 1993) and Necdin, member of the melanoma-associated antigen family of proteins (Fujiwara et al., 2012) to undergo differentiation. The terminal differentiation to mature adipocytes is regulated by a number of transcriptional factors including early key regulator CAAT/enhancer binding proteins (C/EBPβ followed by C/EBPα, C/EBPδ), the master regulator PPARγ and sterol regulatory binding protein 1 (SREBP1) (Payne et al., 2009; White and Stephens, 2010). These transcriptional factors induce expression of various genes related to lipogenesis, lipolysis and insulin sensitivity including fatty acid binding protein (FABP4), lipoprotein lipase (LPL), glucose transporter (GLUT4) and fatty acid synthase (FASN) (Lefterova et al., 2008; Nielsen et al., 2008; Madsen et al., 2014).
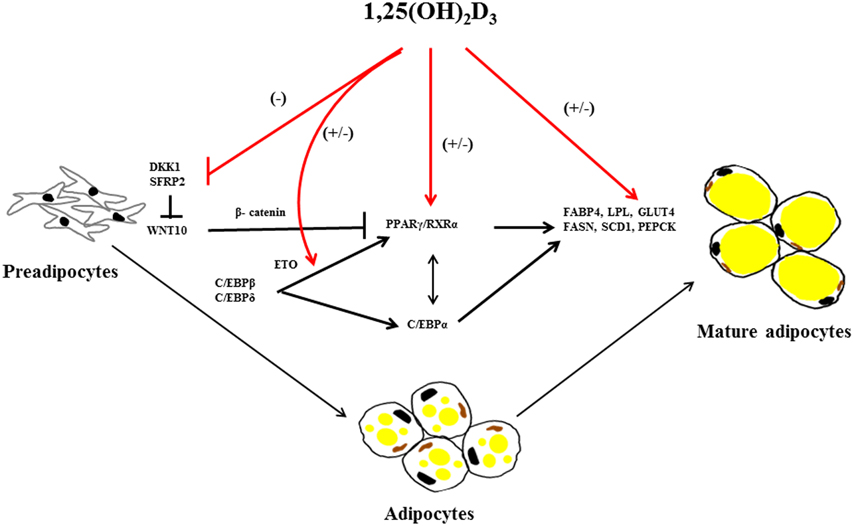
Figure 2. Overview of 1,25(OH)2D3 on regulating factors in adipogenesis. 1,25(OH)2D3 has both stimulating and inhibition effect on adipogenesis. 1,25(OH)2D3 suppress dickkopf1(DKK1) and secreted frizzled-related protein 2 (SFRP2) expression in mouse bone marrow stromal cells (BMSCs) there by suppressing adipogenic peroxisome proliferator-activated receptor γ/retinoid X receptor complex (PPARγ/RXRα) mediated by stabilization of β-catenin through wingless-type MMTV integration site family, member 10 (WNT10) inhibition. Furthermore, 1,25(OH)2D3 regulates several adipogenic mediators during differentiation, proliferation and maturation of mesodermal cells into adipocytes including CCAAT/enhancer-binding proteins α, β, and γ (C/EBPα, β, and γ), C/EBPβ corepressor eight twenty-one (ETO), PPARγ/RXRα, fatty acid binding protein 4 (FABP4), lipoprotein lipase (LPL), fatty acid synthase (FASN), stearoyl-coA desaturase-1 (SCD1), glucose transporter type 4 (GLUT4) and phosphoenolpyruvate carboxykinase (PEPCK). The arrows in the figure indicate activation and blunted line indicate inhibition. The red lines and (+/−) indicate stimulatory or inhibitory effect of 1,25(OH)2D3 on adipogenesis dependent on the species or cellular systems studied.
Investigations of the molecular regulation of 1,25(OH)2D3 on adipogenesis have been conducted in vitro. In mouse 3T3-L1 preadipocytes, 1,25(OH)2D3 inhibits adipogenesis by acting on multiple targets suppressing C/EBPα and PPARγ expression, specifically antagonizing the transacting activity of PPARγ, and sequestering the nuclear receptor retinoic X receptor (RXR), a member nuclear receptor superfamily and down regulating both C/EBP β mRNA expression and C/EBP β nuclear protein levels (Kong and Li, 2006) (Figure 2). 1,25(OH)2D3 stimulates expression of the C/EBP β corepressor, eight twenty-one (ETO), and thus further inhibits the action of any remaining C/EBP β transcriptional effects required for adipogenesis (Blumberg et al., 2006).
Although early studies have established an inhibitory action of 1,25(OH)2D3 in 3T3-L1 preadipocytes differentiation, recently, a more specific effect of 1,25(OH)2D3 on WNT signaling emerged. WNT/β-catenin maintain preadipocytes in their undifferentiated state and thus preventing adipogenesis (Ross et al., 2000). The anti-adipogenic effect of 1,25(OH)2D3 is mediated by maintenance of WNT10B and nuclear β-catenin levels expression levels in 3T3-L1 preadipocytes, thereby suppressing transcription factor PPARγ (Lee et al., 2012). In addition, 1,25(OH)2D3 also inhibited mouse bone marrow stromal cells(BMSCs) differentiation into adipocytes by suppression of dickkopf1 (DKK1) and secreted frizzled-related protein 2 (SFRP2) expression levels via VDR mediated WNT signaling (Cianferotti and Demay, 2007).
In contrast, 1,25(OH)2D3 treatment of porcine mesenchymal stem cells (MSCs) stimulated both proliferation and differentiation in a dose dependent manner toward adipocytic phenotype by increasing PPARγ, LPL and adipocyte-binding protein 2 (AP2) mRNA levels (Mahajan and Stahl, 2009). In human tissue, 1,25(OH)2D3 promotes differentiation of already committed subcutaneous preadipocytes through increased expression of adipogenic markers FABP4 and LPL (Nimitphong et al., 2012). Narvaez et al. (2013) demonstrated that mesenchymal cells differentiate in the presence of 1,25(OH)2D3 toward adipocytes with an enhanced lipid accumulation and increased expression of adipogenic marker genes (FASN, FABP4, and PPARγ).
In conclusion, 1,25(OH)2D3 regulates adipogenesis at various levels of the entire differentiation process (Figure 2). However, there are significant differences summarized in Table 1; the reasons for these differences are not clear at the moment—methodological differences as well as physiological roles of the adipose tissue in different species in their environments might affect these processes. Further studies are needed to address the effects of vitamin D in adipose tissue and its relevance in the associated diseases.
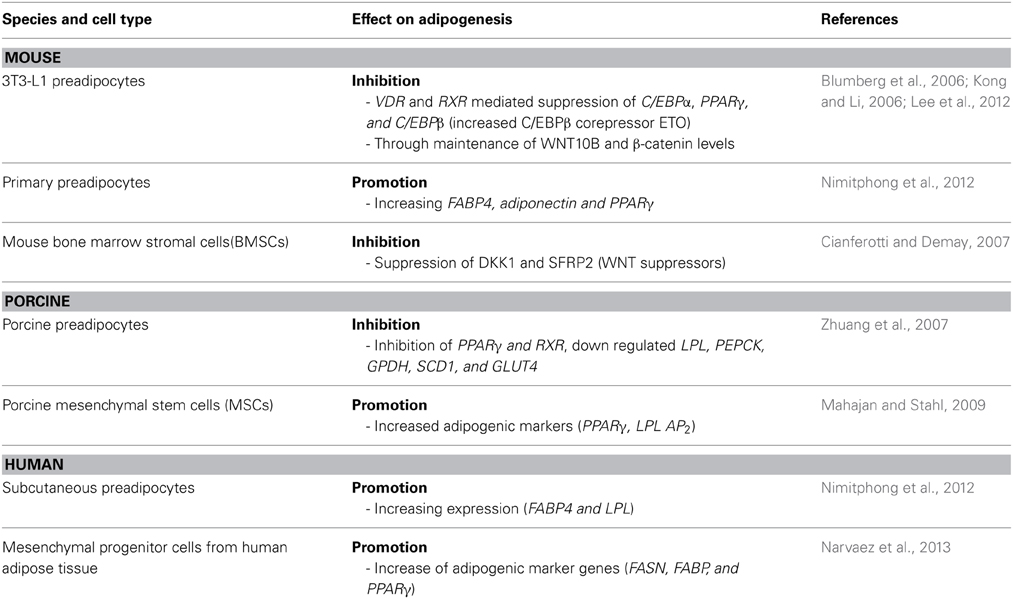
Table 1. Effect of 1,25(OH)2D3 on adipogenesis in different species.
Vitamin D and Adipose Tissue Inflammation
In obesity, adipose tissue undergoes hypertrophic enlargement, which results in an imbalanced blood flow leading to hypoxia, inflammation and macrophage infiltration (Goossens, 2008; Trayhurn, 2013). The hypertrophied adipocytes are characterized by a reduced secretion of adiponectin and increased secretion of several proinflammatory cytokines such as interleukin IL-6, IL-8, TNF-α, resistin and MCP1 (Wellen and Hotamisligil, 2003; Maury and Brichard, 2010; Vlasova et al., 2010).
1,25(OH)2D3 acts at several levels to modulate the function of the immune system (Lemire, 2000). Several in vitro studies in the mouse 3T3-L1 cell line and human adipocytes have demonstrated that 1,25(OH)2D3 inhibits chronic inflammation in adipose tissue (Table 2). However, earlier studies performed in 3T3-L1 and human adipocytes demonstrate contradictory results favoring inflammatory cytokine expression (Sun and Zemel, 2008); the reasons for the contradictory findings are unclear. Recent evidence focuses on the involvement of 1,25(OH)2D3 in the regulation of adipose tissue inflammation by reducing the proinflammatory cytokines secreted from adipose tissue.
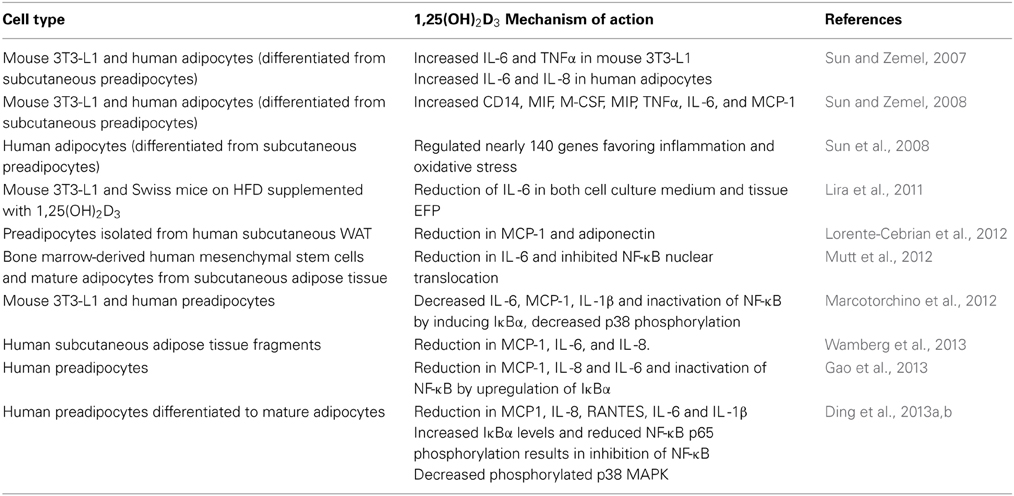
Table 2. 1,25(OH)2D3 and inflammation.
In differentiated adipocytes from human subcutaneous white adipose tissue 1,25(OH)2D3 attenuates TNF-α induced MCP-1 secretion, while it inhibited secretion of adiponectin without affecting its mRNA levels (Lorente-Cebrian et al., 2012). In human subcutaneous adipose tissue fragments 1,25(OH)2D3 reduced IL-1β induced expression of the inflammatory genes MCP-1, IL-6 and IL-8. However, results from the cell culture experiments have not been consistent with the in vivo findings. In a randomized controlled trial including fifty-five obese subjects, oral supplementation of vitamin D 7000 IU per day over 26 weeks did neither affect inflammation markers in the circulation nor in the adipose tissue (Wamberg et al., 2013). In mice on high fat diet, dietary supplementation of 1,25(OH)2D3 (0.05 mg/kg of diet) reduced their IL-6 protein content in epididymal adipose tissue and in the 3T3-L1 cell line stimulated by LPS (Lira et al., 2011).
Signal transduction of inflammatory pathways in adipose tissue involves activation of NF-κB and translocation of p65 to nucleus mediated by degradation of IκBα (Tourniaire et al., 2013). Mutt et al. (2012) have demonstrated that, 1,25(OH)2D3 suppressed LPS-stimulated IL-6 secretion in human isolated mature and MSC differentiated adipocytes. This was confirmed by Marcotorchino et al. (2012), who demonstrated that 1,25(OH)2D3 inhibits the inflammatory markers in both mouse and human adipocytes via the involvement of p38 MAP kinase and NF-κB classical inflammatory pathway and later by Gao et al. (2013) and Ding et al. (2013a).
In summary, the presence of 1,25(OH)2D3 inhibited chemokine and cytokine secretion in human adipocytes. 1,25(OH)2D3 strongly inhibited the activation of the NF-κB and MAPK signaling pathways, which prevent gene transcription of the proinflammatory factors (Figure 3). 1,25(OH)2D3 has been shown by different groups in different models to significantly reduce inflammation in the adipose tissue. However, further studies are needed to provide more evidence for the physiological relevance and the concentration levels of active 1,25(OH)2D3 in lean and obese subjects required to ameliorate the inflammation and associated complications.
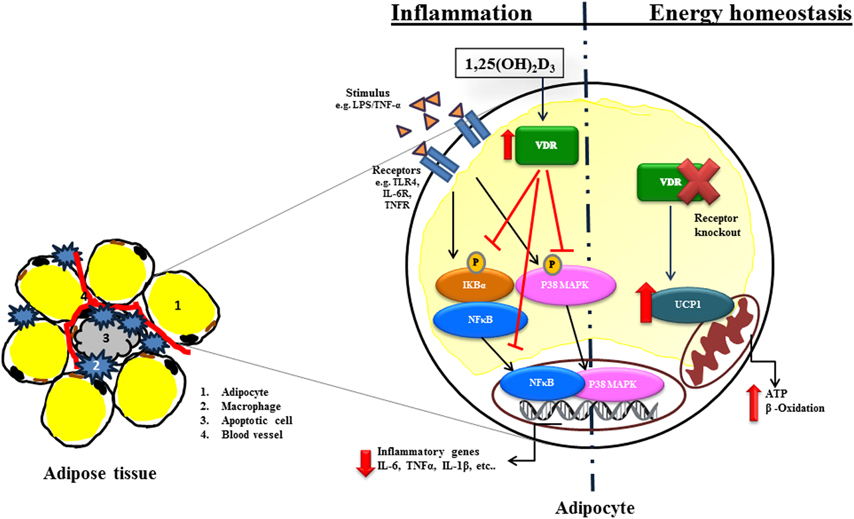
Figure 3. Molecular actions of 1,25(OH)2D3 in inflammation and energy homeostasis in adipocytes. Stimulation via e.g., lipopolysaccharide (LPS), TNF-α via specific receptors e.g., Toll like receptor (TLR), IL-6 receptors (IL-6R) activate Nuclear factor kappa-B (NFκB) or p38 mitogen-activated protein kinase (P38MAPK) signaling dependent transcription of inflammatory genes such as interleukin 6 (IL-6), tumor necrosis factor alpha (TNF-α) and interleukin 1 beta (IL-1β). 1,25(OH)2D3 inhibits inflammation by inhibiting Inhibitor kappa-B (IκBα) phosphorylation and translocation of NFκB as well P38MAPK into the nucleus. Furthermore, 1,25(OH)2D3 affects energy homeostasis. VDR−/− mice increases energy expenditure through uncoupling proteins (UCPs). The arrows in the figure indicate activation and blunted line indicate inhibition. The red lines and arrows indicate the effect of 1,25(OH)2D3 on inflammatory signaling pathway.
Vitamin D and Adipose Tissue Energy Homeostasis
The discovery of VDR expression in adipocytes was the cornerstone for the investigations of the effect of vitamin D on adipose tissue beyond its role in bone metabolism (Stumpf, 1995; Ding et al., 2012). Recent findings in genetically modified mouse models highlighted a new role for vitamin D and its receptor VDR in adipose tissue energy homeostasis. VDR knockout (VDR−/−) mice had reduced body weight and lower serum leptin concentrations, despite of an increased compensatory food intake compared to wild type mice of different genetic background C57BL6 and CD1. VDR−/− mice were highly resistant to high fat diet induced weight gain (Narvaez et al., 2009). In addition, these mice are characterized by a relatively short lifespan, alopecia, osteoporosis, ectopic calcification, progressive loss of hearing and balance (Keisala et al., 2009; Tuohimaa, 2009). Mice lacking CYP27B1 [the (25(OH)D)-1α-hydroxylase enzyme, converts 25(OH)D3 in to 1,25(OH)2D3], displayed features similar to VDR−/− with reduced body weight, hypoleptinemia and hyperphagia. Interestingly, uncoupling protein 1 (UCP-1) expression in white adipose tissue of the VDR−/− mice was increased 25-fold.
In addition to reduced body weight, VDR−/− mice had less body fat and lower levels of plasma triglycerides and cholesterol in comparison to the wild type counterparts even though mice were challenged with a high fat diet (Wong et al., 2009; Weber and Erben, 2013). The depletion of adipose tissue in younger VDR−/− mice progresses with aging and resulted in severe mammary adipose tissue atrophy, along with the increased respiration and energy expenditure (Welsh et al., 2011). The effect on plasma lipid profile and unaltered food intake in these mice was confirmed by an increased β-oxidation rate in isolated adipocytes mediated by the induction of carnitine palmitoyltransferase II (CPTII) (Figure 3). VDR−/− mice had an increased basal metabolism demonstrated by the total energy expenditure, oxygen consumption and CO2 production in comparison with the wild type mice (Wong et al., 2009). In addition, UCP1, UCP2, and UCP3 mRNAs were upregulated in brown adipose tissue of the VDR−/− mice fed high fat diet. In contrast to VDR knock out models with the ablation of the receptor in the whole animal, adipose tissue specific overexpression of human VDR via the adipocyte fatty acid binding protein (aP2) promoter/enhancer element resulted in a decreased energy expenditure and oxygen consumption and thus the mice had an increased body weight and fat mass (Wong et al., 2011).
In conclusion, these transgenic animal models indicate a critical and complex role for 1,25(OH)2D3 and VDR signaling in energy homeostasis. However, notwithstanding the cell and mouse studies, further studies need to explore the role of vitamin D on human adipose tissue metabolism in vivo.
Genetic View on the Actions of VDR in Adipocytes: Integration with Other Tissues
The VDR genomic interactions in different types of cells and tissues have been mapped by in vitro experiments where target cells (primary or secondary) have been treated with 1,25(OH)2D3. Upon stimulation of VDR by its ligand, it forms a heterodimer with RXR and subsequently binds to the vitamin D response elements (VDREs) within the regulatory regions of target genes. The abundance of VDR binding sites and the regulation of changes in gene expressions are analyzed using array technology and the combination of chromatin immunoprecipitation (ChIP) with massive parallel sequencing (ChIP-seq). These advanced techniques have provided novel mechanistic insights of 1,25(OH)2D3 action via VDR in the regulation of cellular metabolism and disease states. However, studies on genome-wide actions of VDR in adipocytes are sparse.
Recent microarray studies of human adipocytes and preadipocytes incubated with macrophage-conditioned medium derived from U937 monocytes, confirmed the induction of genes associated with the metabolism and action of 1,25(OH)2D3, including CYP27B1 and VDR (Trayhurn et al., 2011). An earlier single microarray study in human subcutaneous adipose tissue derived preadipocytes differentiated to adipocytes demonstrated 237 1,25(OH)2D3 responsive genes (cell proliferation, angiogenesis, cell cycle, inflammation and response to oxidative stress) (Sun et al., 2008).
Most recent studies in the other cell types such as monocytes, primary CD4+ T-lymphocytes, adenocarcinoma, hepatic stellate and lymphoblastoid cell lines (LCLs) (Ramagopalan et al., 2010; Heikkinen et al., 2011; Meyer et al., 2012; Ding et al., 2013b; Handel et al., 2013; Tuoresmäki et al., 2014) contribute to a systemic understanding of 1,25(OH)2D3 induced gene regulation. Depending on the cell type, concentration and length of 1,25(OH)2D3 incubation approximately 2000 VDR genomic binding sites have been found in these studies. Yet, alterations in DNA accessibility in cell lines after short-term stimulation with 1,25(OH)2D3 may not reflect the physiological 1,25(OH)2D3 levels in vivo due to the different tissue environment and sympathetic influence. In primary CD4+ lymphocyte cells, isolated from nine healthy individuals with measured serum 25(OH)D levels, VDR binding sites ranged from 200 to 7118 across the genome and the corresponding 25(OH)D levels directly correlated with the number of VDR binding sites, suggesting far greater number of VDR binding sites in 1,25(OH)2D3 sufficient than the insufficient subjects (Handel et al., 2013).
Genome-wide VDR cistromes are not available in adipocytes, but recent VDR binding sites in other cell types has been mapped with ChIP-seq from both upstream and downstream of the transcription start site. Further genome wide view actions of VDR in adipocytes as well as integration of other tissue specific cell types are warranted.
Conclusion and Future Directions
Adipose tissue acts in addition to nutrient storage as an active endocrine organ. In the obese state, sub-clinical inflammation increases the risk of a variety of chronic diseases. Vitamin D deficiency is common in overweight and obese individuals, and it is possible that lower circulating concentrations may contribute to increases in metabolic risk. A genome-wide association study of 25(OH)D concentrations in 33996 individuals of European descent from 15 cohorts found variants near genes involved in cholesterol synthesis, hydroxylation, and vitamin D transport affect vitamin D status (Wang et al., 2010). Genetic variation at these loci identifies individuals who have a substantially increased risk of vitamin D insufficiency.
On the cellular level, 1,25(OH)2D3 has a significant role in adipogenesis and inflammation which might be species dependent. Holick et al. (1989) demonstrated that the peak circulating concentrations of 25(OH)D in the elderly are about 30% of that of the young. These findings suggest that there will be significant challenges in the translation of the finding from models and non-human primates to the targeted human populations (healthy, diseased, black, white, age, BMI, geographical latitude, race). More evidence accumulates that one dose does not fit all (Powe et al., 2014). Powe and colleagues evaluated vitamin D binding proteins (VDBP) and 25(OH)D levels in black and white Americans. Black adult Americans had low 25(OH)D levels and with the threshold of 20 or 30 ng/ml, 77–96% of them would be classified as vitamin D deficient. Surprisingly, the black study participants had higher bone mineral density, higher calcium levels and only slightly higher parathyroid levels than the white study participants due to VDBP gene polymorphisms (rs7041 and rs4588). The authors speculated that the low levels of VDBP might protect against the adverse effects of vitamin D deficiency. Sufficient levels of this essential hormone and the development of potent novel vitamin D receptor analogs (Peräkylä et al., 2005; Leyssens et al., 2014), which could be easily and cheaply substituted, are beneficial in the maintenance of health and prevention of a number of diseases associated with vitamin D deficiency. Recent systemic review and meta-analysis summary of observational studies and randomized interventions investigated the association between the circulating 25(OH)D concentrations and cause specific mortality in 900,000 subjects in 26 countries (Chowdhury et al., 2014). There was an inverse association of mortality risk and vitamin D levels, yet the observed association could be direct [suboptimal 25(OH)D concentrations] or indirect through higher BMI or disadvantageous social circumstances. Thus, prospective intervention studies are needed to establish potential causal associations between vitamin D levels and disease outcomes.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgment
This work was support in part by the grants from the Finnish Academy project 139900, Finnish Cultural Foundation, North Ostrobothnia Regional fund.
Abbreviations
25(OH)D, 25-hydroxyvitamin D; 1,25(OH)2D3, 1,25-dihydroxycholecalciferol; VDBPs, Vitamin D binding proteins; VDR, Vitamin D receptor; DKK1, dickkopf 1; SFRP2, Frizzled-related protein 2; BMSCs, bone marrow stromal cells; PPARγ, peroxisome proliferator-activated receptor gamma; RXRα, retinoid X receptor alpha; WNT10, wingless-type MMTV integration site family member 10; C/EBP (α, β, and γ), CCAAT/enhancer-binding proteins (α, β, and γ); ETO, C/EBPβ corepressor eight twenty-one; FABP4, fatty acid binding protein 4; LPL, lipoprotein lipase; FASN, fatty acid synthase; SCD1, stearoyl-coA desaturase 1; GLUT4, glucose transporter type 4; PEPCK, phosphoenolpyruvate carboxykinase; LPS, lipopolysaccharide; TLR, toll like receptor; IL-6R, IL-6 receptors; NFκB, nuclear factor kappa-B; P38MAPK, p38 mitogen-activated protein kinase; IL-6, interleukin 6; TNF-α, tumor necrosis factor alpha; IL-1β , interleukin 1 beta; IκBα, inhibitor kappa-B; UCPs, uncoupling proteins; VDR −/−, VDR knock out; CYP27B1−/−, CYP27B1 knock out; BMI, Body mass index; BMPs, bone morphogenetic proteins; FGFs, fibroblast growth factors; TGFβ , transforming-growth factor β ; IGF1, insulin like growth factor 1; JAK-STAT3, janus kinase-signal transducer and activator of transcription 3; S6K1, ribosomal protein S6 kinase 1; WNT, wingless family; Rb, protein of retinoblastoma family; Pref1, preadipocyte factor 1; Necdin, melanoma-associated antigen family of proteins member; SREBP1, sterol regulatory binding protein 1; MSCs, mesenchymal stem cells; AP2, adipocyte-binding protein 2; MCP1, monocyte chemoattractant protein 1; CYP27B1,(25(OH)D)-1α-hydroxylase; CPTII, carnitine palmitoyltransferase II; WAT, White adipose tissue; VDREs, vitamin D response elements; ChIP-seq, chromatin immunoprecipitation—sequencing; LCLs, lymphoblastoid cell lines.
References
Adams, K. F., Schatzkin, A., Harris, T. B., Kipnis, V., Mouw, T., Ballard-Barbash, R., et al. (2006). Overweight, obesity, and mortality in a large prospective cohort of persons 50 to 71 years old. N. Engl. J. Med. 355, 763–778. doi: 10.1056/NEJMoa055643
Arner, P., and Spalding, K. L. (2010). Fat cell turnover in humans. Biochem. Biophys. Res. Commun. 396, 101–104. doi: 10.1016/j.bbrc.2010.02.165
Blumberg, J. M., Tzameli, I., Astapova, I., Lam, F. S., Flier, J. S., and Hollenberg, A. N. (2006). Complex role of the vitamin D receptor and its ligand in adipogenesis in 3T3-L1 cells. J. Biol. Chem. 281, 11205–11213. doi: 10.1074/jbc.M510343200
Carnevalli, L. S., Masuda, K., Frigerio, F., Le Bacquer, O., Um, S. H., Gandin, V., et al. (2010). S6K1 plays a critical role in early adipocyte differentiation. Dev. Cell. 18, 763–774. doi: 10.1016/j.devcel.2010.02.018
Chowdhury, R., Kunutsor, S., Vitezova, A., Oliver-Williams, C., Chowdhury, S., Kiefte-de-Jong, J. C., et al. (2014). Vitamin D and risk of cause specific death: systematic review and meta-analysis of observational cohort and randomised intervention studies. BMJ 348:g1903. doi: 10.1136/bmj.g1903.
Cianferotti, L., and Demay, M. B. (2007). VDR-mediated inhibition of DKK1 and SFRP2 suppresses adipogenic differentiation of murine bone marrow stromal cells. J. Cell. Biochem. 101, 80–88. doi: 10.1002/jcb.21151
Despres, J. P., and Lemieux, I. (2006). Abdominal obesity and metabolic syndrome. Nature 444, 881–887. doi: 10.1038/nature05488
Ding, C., Gao, D., Wilding, J., Trayhurn, P., and Bing, C. (2012). Vitamin D signalling in adipose tissue. Br. J. Nutr. 108, 1915–1923. doi: 10.1017/S0007114512003285
Ding, C., Wilding, J. P., and Bing, C. (2013a). 1,25-dihydroxyvitamin D3 protects against macrophage-induced activation of NFkappaB and MAPK signalling and chemokine release in human adipocytes. PLoS ONE 8:e61707. doi: 10.1371/journal.pone.0061707
Ding, N., Yu, R. T., Subramaniam, N., Sherman, M. H., Wilson, C., Rao, R., et al. (2013b). A vitamin D receptor/SMAD genomic circuit gates hepatic fibrotic response. Cell 153, 601–613. doi: 10.1016/j.cell.2013.03.028
Frankenfield, D. C., Rowe, W. A., Cooney, R. N., Smith, J. S., and Becker, D. (2001). Limits of body mass index to detect obesity and predict body composition. Nutrition 17, 26–30. doi: 10.1016/S0899-9007(00)00471-8
Fujiwara, K., Hasegawa, K., Ohkumo, T., Miyoshi, H., Tseng, Y. H., and Yoshikawa, K. (2012). Necdin controls proliferation of white adipocyte progenitor cells. PLoS ONE 7:e30948. doi: 10.1371/journal.pone.0030948
Gao, D., Trayhurn, P., and Bing, C. (2013). 1,25-Dihydroxyvitamin D3 inhibits the cytokine-induced secretion of MCP-1 and reduces monocyte recruitment by human preadipocytes. Int. J. Obes. (Lond) 37, 357–365. doi: 10.1038/ijo.2012.53
Goossens, G. H. (2008). The role of adipose tissue dysfunction in the pathogenesis of obesity-related insulin resistance. Physiol. Behav. 94, 206–218 doi: 10.1016/j.physbeh.2007.10.010
Handel, A. E., Sandve, G. K., Disanto, G., Berlanga-Taylor, A. J., Gallone, G., Hanwell, H., et al. (2013). Vitamin D receptor ChIP-seq in primary CD4+ cells: relationship to serum 25-hydroxyvitamin D levels and autoimmune disease. BMC Med. 11:163. doi: 10.1186/1741-7015-11-163
Heikkinen, S., Vaisanen, S., Pehkonen, P., Seuter, S., Benes, V., and Carlberg, C. (2011). Nuclear hormone 1alpha,25-dihydroxyvitamin D3 elicits a genome-wide shift in the locations of VDR chromatin occupancy. Nucleic Acids Res. 39, 9181–9193. doi: 10.1093/nar/gkr654
Holick, M. F., Matsuoka, L. Y., and Wortsman, J. (1989). Age, vitamin D, and solar ultraviolet. Lancet 2, 1104–1105. doi: 10.1016/S0140-6736(89)91124-0
Hotamisligil, G. S. (2006). Inflammation and metabolic disorders. Nature 444, 860–867. doi: 10.1038/nature05485
Huotari, A., and Herzig, K. H. (2008). Vitamin D and living in northern latitudes–an endemic risk area for vitamin D deficiency. Int. J. Circumpolar Health 67, 164–178. doi: 10.3402/ijch.v67i2-3.18258
Jin, W., Takagi, T., Kanesashi, S. N., Kurahashi, T., Nomura, T., Harada, J., et al. (2006). Schnurri-2 controls BMP-dependent adipogenesis via interaction with Smad proteins. Dev. Cell. 10, 461–471. doi: 10.1016/j.devcel.2006.02.016
Kahn, S. E., Hull, R. L., and Utzschneider, K. M. (2006). Mechanisms linking obesity to insulin resistance and type 2 diabetes. Nature 444, 840–846. doi: 10.1038/nature05482
Keisala, T., Minasyan, A., Lou, Y. R., Zou, J., Kalueff, A. V., Pyykkö, I., et al. (2009). Premature aging in vitamin D receptor mutant mice. J. Steroid Biochem. Mol. Biol. 115, 91–97. doi: 10.1016/j.jsbmb.2009.03.007
Kong, J., and Li, Y. C. (2006). Molecular mechanism of 1,25-dihydroxyvitamin D3 inhibition of adipogenesis in 3T3-L1 cells. Am. J. Physiol. Endocrinol. Metab. 290, E916–E924. doi: 10.1152/ajpendo.00410.2005
Lee, H., Bae, S., and Yoon, Y. (2012). Anti-adipogenic effects of 1,25-dihydroxyvitamin D3 are mediated by the maintenance of the wingless-type MMTV integration site/beta-catenin pathway. Int. J. Mol. Med. 30, 1219–1224. doi: 10.3892/ijmm.2012.1101
Lefterova, M. I., Zhang, Y., Steger, D. J., Schupp, M., Schug, J., Cristancho, A., et al. (2008). PPARgamma and C/EBP factors orchestrate adipocyte biology via adjacent binding on a genome-wide scale. Genes Dev. 22, 2941–2952. doi: 10.1101/gad.1709008
Lehr, S., Hartwig, S., Lamers, D., Famulla, S., Muller, S., Hanisch, F. G., et al. (2012). Identification and validation of novel adipokines released from primary human adipocytes. Mol. Cell. Proteomics 11:M111.010504. doi: 10.1074/mcp.M111.010504
Lemire, J. (2000). 1,25-Dihydroxyvitamin D3–a hormone with immunomodulatory properties. Z. Rheumatol. 59, 24–27. doi: 10.1007/s003930070034
Leyssens, C., Verlinden, L., and Verstuyf, A. (2014). The future of vitamin D analogs. Front. Physiol. 5:122. doi: 10.3389/fphys.2014.00122
Lira, F. S., Rosa, J. C., Cunha, C. A., Ribeiro, E. B., do Nascimento, C. O., Oyama, L. M., et al. (2011). Supplementing alpha-tocopherol (vitamin E) and vitamin D3 in high fat diet decrease IL-6 production in murine epididymal adipose tissue and 3T3-L1 adipocytes following LPS stimulation. Lipids Health Dis. 10:37. doi: 10.1186/1476-511X-10-37
Lorente-Cebrian, S., Eriksson, A., Dunlop, T., Mejhert, N., Dahlman, I., Astrom, G., et al. (2012). Differential effects of 1alpha,25-dihydroxycholecalciferol on MCP-1 and adiponectin production in human white adipocytes. Eur. J. Nutr. 51, 335–342. doi: 10.1007/s00394-011-0218-z
Lowe, C. E., O’Rahilly, S., and Rochford, J. J. (2011). Adipogenesis at a glance. J. Cell. Sci. 124, 2681–2686. doi: 10.1242/jcs.079699
Madsen, M. S., Siersbaek, R., Boergesen, M., Nielsen, R., and Mandrup, S. (2014). Peroxisome proliferator-activated receptor gamma and c/ebpalpha synergistically activate key metabolic adipocyte genes by assisted loading. Mol. Cell. Biol. 34, 939–954. doi: 10.1128/MCB.01344-13
Mahajan, A., and Stahl, C. H. (2009). Dihydroxy-cholecalciferol stimulates adipocytic differentiation of porcine mesenchymal stem cells. J. Nutr. Biochem. 20, 512–520. doi: 10.1016/j.jnutbio.2008.05.010
Marcotorchino, J., Gouranton, E., Romier, B., Tourniaire, F., Astier, J., Malezet, C., et al. (2012). Vitamin D reduces the inflammatory response and restores glucose uptake in adipocytes. Mol. Nutr. Food Res. 56, 1771–1782. doi: 10.1002/mnfr.201200383
Maury, E., and Brichard, S. M. (2010). Adipokine dysregulation, adipose tissue inflammation and metabolic syndrome. Mol. Cell. Endocrinol. 314, 1–16. doi: 10.1016/j.mce.2009.07.031
Meyer, M. B., Goetsch, P. D., and Pike, J. W. (2012). VDR/RXR and TCF4/beta-catenin cistromes in colonic cells of colorectal tumor origin: impact on c-FOS and c-MYC gene expression. Mol. Endocrinol. 26, 37–51. doi: 10.1210/me.2011-1109
Mutt, S. J., Karhu, T., Lehtonen, S., Lehenkari, P., Carlberg, C., Saarnio, J., et al. (2012). Inhibition of cytokine secretion from adipocytes by 1,25-dihydroxyvitamin D(3) via the NF-kappaB pathway. FASEB J. 26, 4400–4407. doi: 10.1096/fj.12-210808
Narvaez, C. J., Matthews, D., Broun, E., Chan, M., and Welsh, J. (2009). Lean phenotype and resistance to diet-induced obesity in vitamin D receptor knockout mice correlates with induction of uncoupling protein-1 in white adipose tissue. Endocrinology 150, 651–661. doi: 10.1210/en.2008-1118
Narvaez, C. J., Simmons, K. M., Brunton, J., Salinero, A., Chittur, S. V., and Welsh, J. E. (2013). Induction of STEAP4 correlates with 1,25-dihydroxyvitamin D3 stimulation of adipogenesis in mesenchymal progenitor cells derived from human adipose tissue. J. Cell. Physiol. 228, 2024–2036. doi: 10.1002/jcp.24371
Nielsen, R., Pedersen, T. A., Hagenbeek, D., Moulos, P., Siersbaek, R., Megens, E., et al. (2008). Genome-wide profiling of PPARgamma: RXR and RNA polymerase II occupancy reveals temporal activation of distinct metabolic pathways and changes in RXR dimer composition during adipogenesis. Genes Dev. 22, 2953–2967. doi: 10.1101/gad.501108
Nimitphong, H., Holick, M. F., Fried, S. K., and Lee, M. J. (2012). 25-hydroxyvitamin D(3) and 1,25-dihydroxyvitamin D(3) promote the differentiation of human subcutaneous preadipocytes. PLoS ONE 7:e52171. doi: 10.1371/journal.pone.0052171
Payne, V. A., Au, W. S., Lowe, C. E., Rahman, S. M., Friedman, J. E., O’Rahilly, S., et al. (2009). C/EBP transcription factors regulate SREBP1c gene expression during adipogenesis. Biochem. J. 425, 215–223. doi: 10.1042/BJ20091112
Peräkylä, M., Malinen, M., Herzig, K. H., and Carlberg, C. (2005). Gene regulatory potential of nonsteroidal vitamin D receptor ligands. Mol. Endocrinol. 19, 2060–2073. doi: 10.1210/me.2004-0417
Phinney, K. W., Bedner, M., Tai, S. S., Vamathevan, V. V., Sander, L. C., Sharpless, K. E., et al. (2012). Development and certification of a standard reference material for vitamin D metabolites in human serum. Anal. Chem. 84, 956–962. doi: 10.1021/ac202047n
Powe, C. E., Karumanchi, S. A., and Thadhani, R. (2014). Vitamin D-binding protein and vitamin D in blacks and whites. N. Engl. J. Med. 370, 880–881. doi: 10.1056/NEJMoa1306357
Ramagopalan, S. V., Heger, A., Berlanga, A. J., Maugeri, N. J., Lincoln, M. R., Burrell, A., et al. (2010). A ChIP-seq defined genome-wide map of vitamin D receptor binding: associations with disease and evolution. Genome Res. 20, 1352–1360. doi: 10.1101/gr.107920.110
Ross, A., Taylor, C., Yaktine, A., and Del Valle, H. (2011). Dietary Reference Intakes for Calcium and Vitamin D. Institute of Medicine Report. Washington, DC: The National Academies Press.
Ross, S. E., Hemati, N., Longo, K. A., Bennett, C. N., Lucas, P. C., Erickson, R. L., et al. (2000). Inhibition of adipogenesis by Wnt signaling. Science 289, 950–953. doi: 10.1126/science.289.5481.950
Scime, A., Grenier, G., Huh, M. S., Gillespie, M. A., Bevilacqua, L., Harper, M. E., et al. (2005). Rb and p107 regulate preadipocyte differentiation into white versus brown fat through repression of PGC-1alpha. Cell. Metab. 2, 283–295. doi: 10.1016/j.cmet.2005.10.002
Smas, C. M., and Sul, H. S. (1993). Pref-1, a protein containing EGF-like repeats, inhibits adipocyte differentiation. Cell 73, 725–734. doi: 10.1016/0092-8674(93)90252-L
Stumpf, W. E. (1995). Vitamin D sites and mechanisms of action: a histochemical perspective. reflections on the utility of autoradiography and cytopharmacology for drug targeting. Histochem. Cell Biol. 104, 417–427. doi: 10.1007/BF01464331
Sun, X., Morris, K. L., and Zemel, M. B. (2008). Role of calcitriol and cortisol on human adipocyte proliferation and oxidative and inflammatory stress: a microarray study. J. Nutrigenet. Nutrigenomics 1, 30–48. doi: 10.1159/000109873
Sun, X., and Zemel, M. B. (2007). Calcium and 1,25-dihydroxyvitamin D3 regulation of adipokine expression. Obesity (Silver Spring) 15, 340–348. doi: 10.1038/oby.2007.540
Sun, X., and Zemel, M. B. (2008). Calcitriol and calcium regulate cytokine production and adipocyte-macrophage cross-talk. J. Nutr. Biochem. 19, 392–399. doi: 10.1016/j.jnutbio.2007.05.013
Tourniaire, F., Romier-Crouzet, B., Lee, J. H., Marcotorchino, J., Gouranton, E., Salles, J., et al. (2013). Chemokine expression in inflamed adipose tissue is mainly mediated by NF-kappaB. PLoS ONE 8:e66515. doi: 10.1371/journal.pone.0066515
Trayhurn, P., O’Hara, A., and Bing, C. (2011). Interrogation of microarray datasets indicates that macrophage-secreted factors stimulate the expression of genes associated with vitamin D metabolism (VDR and CYP27B1) in human adipocytes. Adipobiology 3, 29–34.
Trayhurn, P. (2013). Hypoxia and adipose tissue function and dysfunction in obesity. Physiol. Rev. 93, 1–21. doi: 10.1152/physrev.00017.2012
Tuohimaa, P. (2009). Vitamin D and aging. J. Steroid Biochem. Mol. Biol. 114, 78–84. doi: 10.1016/j.jsbmb.2008.12.020
Tuoresmäki, P., Väisänen, S., Neme, A., Heikkinen, S., and Carlberg, C. (2014). Patterns of genome-wide VDR locations. PLoS ONE 9:e96105. doi: 10.1371/journal.pone.0096105
Van Gaal, L. F., Mertens, I. L., and De Block, C. E. (2006). Mechanisms linking obesity with cardiovascular disease. Nature 444, 875–880. doi: 10.1038/nature05487
Vigilanza, P., Aquilano, K., Baldelli, S., Rotilio, G., and Ciriolo, M. R. (2011). Modulation of intracellular glutathione affects adipogenesis in 3T3-L1 cells. J. Cell. Physiol. 226, 2016–2024. doi: 10.1002/jcp.22542
Vimaleswaran, K. S., Berry, D. J., Lu, C., Tikkanen, E., Pilz, S., Hiraki, L. T., et al. (2013). Causal relationship between obesity and vitamin D status: bi-directional mendelian randomization analysis of multiple cohorts. PLoS Med. 10:e1001383. doi: 10.1371/journal.pmed.1001383
Vlasova, M., Purhonen, A. K., Jarvelin, M. R., Rodilla, E., Pascual, J., and Herzig, K. H. (2010). Role of adipokines in obesity-associated hypertension. Acta Physiol. (Oxf). 200, 107–127. doi: 10.1111/j.1748-1716.2010.02171.x
Wamberg, L., Cullberg, K. B., Rejnmark, L., Richelsen, B., and Pedersen, S. B. (2013). Investigations of the anti-inflammatory effects of vitamin D in adipose tissue: results from an in vitro study and a randomized controlled trial. Horm. Metab. Res. 45, 456–462. doi: 10.1055/s-0032-1331746
Wang, T. J., Zhang, F., Richards, J. B., Kestenbaum, B., van Meurs, J. B., Berry, D., et al. (2010). Common genetic determinants of vitamin D insufficiency: a genome-wide association study. Lancet 376, 180–188. doi: 10.1016/S0140-6736(10)60588-0
Weber, K., and Erben, R. G. (2013). Differences in triglyceride and cholesterol metabolism and resistance to obesity in male and female vitamin D receptor knockout mice. J. Anim. Physiol. Anim. Nutr. (Berl.) 97, 675–683. doi: 10.1111/j.1439-0396.2012.01308.x
Wellen, K. E., and Hotamisligil, G. S. (2003). Obesity-induced inflammatory changes in adipose tissue. J. Clin. Invest. 112, 1785–1788. doi: 10.1172/JCI20514
Welsh, J., Zinser, L. N., Mianecki-Morton, L., Martin, J., Waltz, S. E., James, H., et al. (2011). Age-related changes in the epithelial and stromal compartments of the mammary gland in normocalcemic mice lacking the vitamin D3 receptor. PLoS ONE 6:e16479. doi: 10.1371/journal.pone.0016479
White, U. A., and Stephens, J. M. (2010). Transcriptional factors that promote formation of white adipose tissue. Mol. Cell. Endocrinol. 318, 10–14. doi: 10.1016/j.mce.2009.08.023
Wong, K. E., Kong, J., Zhang, W., Szeto, F. L., Ye, H., Deb, D. K., et al. (2011). Targeted expression of human vitamin D receptor in adipocytes decreases energy expenditure and induces obesity in mice. J. Biol. Chem. 286, 33804–33810. doi: 10.1074/jbc.M111.257568
Wong, K. E., Szeto, F. L., Zhang, W., Ye, H., Kong, J., Zhang, Z., et al. (2009). Involvement of the vitamin D receptor in energy metabolism: regulation of uncoupling proteins. Am. J. Physiol. Endocrinol. Metab. 296, E820–E828. doi: 10.1152/ajpendo.90763.2008
Wortsman, J., Matsuoka, L. Y., Chen, T. C., Lu, Z., and Holick, M. F. (2000). Decreased bioavailability of vitamin D in obesity. Am. J. Clin. Nutr. 72, 690–693.
Zhang, K., Guo, W., Yang, Y., and Wu, J. (2011). JAK2/STAT3 pathway is involved in the early stage of adipogenesis through regulating C/EBPbeta transcription. J. Cell. Biochem. 112, 488–497. doi: 10.1002/jcb.22936
Keywords: 1,25-dihydroxycholecalciferol or calcitriol, vitamin D binding protein, gene regulation, adipose tissue, adipogenesis, secretion, adipokines
Citation: Mutt SJ, Hyppönen E, Saarnio J, Järvelin M-R and Herzig K-H (2014) Vitamin D and adipose tissue—more than storage. Front. Physiol. 5:228. doi: 10.3389/fphys.2014.00228
Received: 16 March 2014; Accepted: 02 June 2014;
Published online: 24 June 2014.
Edited by:
Carsten Carlberg, University of Eastern Finland, FinlandReviewed by:
Nina Eikelis, Baker IDI Heart and Diabetes Institute, AustraliaJoEllen Welsh, State University of New York at Albany Cancer Research Center, USA
Copyright © 2014 Mutt, Hyppönen, Saarnio, Järvelin and Herzig. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Shivaprakash J. Mutt and Karl-Heinz Herzig, Institute of Biomedicine and Biocenter of Oulu, University of Oulu, Aapistie 5, PO Box 5000, FIN-90014 Oulu, Finland e-mail:c2hpdmFwcmFrYXNoLmphZ2FsdXJAb3VsdS5maQ==;a2FybC1oZWluei5oZXJ6aWdAb3VsdS5maQ==