 Marco Durante
Marco Durante Kai Schulze
Kai Schulze Sebastien Incerti
Sebastien Incerti Ziad Francis5
Ziad Francis5 Carlos Alberto Guzmán
Carlos Alberto Guzmán- 1Biophysics Department, GSI Helmholtzzentrum für Schwerionenforschung, Darmstadt, Germany
- 2Institute of Condensed Matter Physics, Technische Universität Darmstadt, Darmstadt, Germany
- 3Department of Vaccinology and Applied Microbiology, Helmholtz Zentrum für Infektionsforschung (HZI), Braunschweig, Germany
- 4CNRS/IN2P3, UMR5797, Center d'Études Nucléaires de Bordeaux Gradignan, Université de Bordeaux, Bordeaux, France
- 5Faculty of Sciences, U.R. Mathématiques et Modélisation, Saint Joseph University, Beirut, Lebanon
Virus irradiation has been performed for many decades for basic research studies, sterilization, and vaccine development. The COVID-19 outbreak is currently causing an enormous effort worldwide for finding a vaccine against coronavirus. High doses of γ-rays can be used for the development of vaccines that exploit inactivated virus. This technique has been gradually replaced by more practical methods, in particular the use of chemicals, but irradiation remains a simple and effective method used in some cases. The technique employed for inactivating a virus has an impact on its ability to induce an adaptive immune response able to confer effective protection. We propose here that accelerated heavy ions can be used to inactivate SARS-CoV-2 viruses with small damage to the spike proteins of the envelope and can then provide an intact virion for vaccine development.
Introduction
The coronavirus disease in 2019 (COVID-19), caused by the severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) [1], is an unprecedented health emergency in this century. The World Health Organization declared COVID-19 a pandemic in March 2020. From the start of the pandemic to August 2020, over 22 million cases have been reported worldwide, resulting in over 780,000 casualties. Lacking effective antiviral drugs, the rush to develop an effective vaccine is enormous [2, 3], with over 100 vaccines in pre-clinical evaluation and 10 already in clinical trials [4].
There are several techniques in use to find the most effective vaccine against SARS-CoV-2, including innovative RNA vaccines, viral vector-, or protein-based vaccines [5]. However, the conventional method of using weakened or inactivated viruses is still avidly pursued and has produced some of the most promising vaccines under test [6, 7].
Techniques for virus inactivation are both chemical and physical, the latter including heat, UV, and ionizing radiation (usually γ-rays). The method used for inactivation is important, because the damage to the epitopes will reduce the efficacy of the vaccine. Several studies have measured the impact of different chemical and physical methods on the efficacy of the inactivated virus [8–10]. Chemicals, such as formaldehyde, hydrogen peroxide, binary ethylenimine derivatives, or β-propiolactone, are very practical but can damage the envelope protein and leave toxic residuals. Gamma radiation is therefore still considered a very safe and effective method [11] as shown in many recent reports [12–16].
Virus Radiobiology
Inactivation of virus by radiation has been studied for over a century [17–19]. Virus radiosensitivity is lower when the irradiation is performed in growth medium compared to water and strongly depends on the size of the virion envelope [20, 21]. Viruses are lacking enzymes and are therefore unable to repair any damage in their nucleic acids. These simple targets are therefore the perfect objects to test the target theory of radiation action, introduced by Lea [22]. According to the target theory, the hit probability P for N targets to be hit n times by radiation follows the Poisson distribution:
where v is the target volume and D the radiation dose. In the simple case of N = n = 1, the equation is reduced to the simple single-hit–single-target model:
where F = D·LET is the fluence (in particles/cm2), LET the radiation linear energy transfer (in MeV·cm2/g, often expressed in keV/μm in water), and σ = v/LET is the inactivation cross-section (in cm2). The target theory cannot describe cellular repair effect but is perfectly able to describe the inactivation of the viruses. The survival curves are in fact always linear (in logarithmic scale), and assuming a given energy for the inactivation event, the volume v can be calculated from the slope. Using charged particles, the inactivation cross-section can provide the area of the sensible target, under the assumption that every traversal is lethal [23, 24]. Figure 1 shows typical survival curves of viruses, whereas in Figure 2, we report the inactivation cross-section as a function of LET. The inactivation of the virus is caused by the damage to the nucleic acid, either RNA or DNA. A single-strand break (for single-stranded virus) or a double-strand break is generally sufficient to make the product of the viral nucleic acid not viable. An additional source of inactivation is the damage to the capsid, that can lead to release of the DNA (or RNA) from the viral envelope (Figure 3). Even at high doses, however, this mechanism is less important than direct damage to the DNA [25].
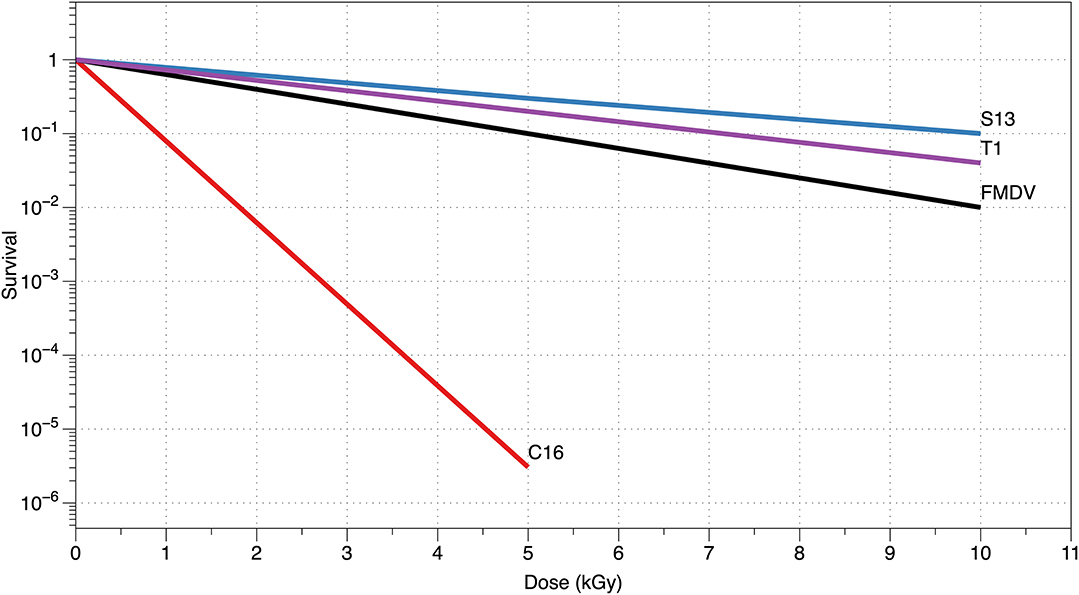
Figure 1. Virus radiosensitivity. Survival of different viruses to X- or γ-rays is plotted vs. the dose (in kGy) for different viruses. C16 bacteriophage [19] is larger (50–70 nm) compared to the S13 [18] and T1 [23] bacteriophages and to the foot-and-mouth-disease picornavirus (FMDV) [21].
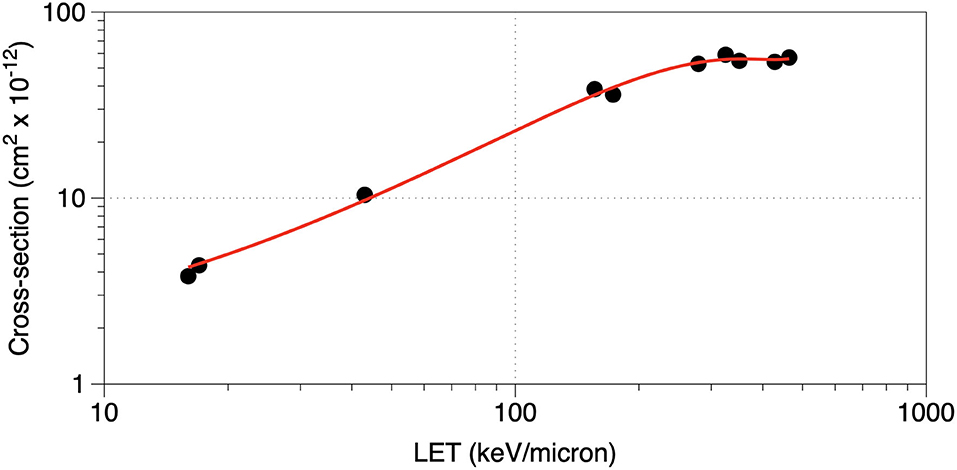
Figure 2. Inactivation efficiency increases with LET. Inactivation cross-section of the bacteriophage T1 plotted vs. the particle LET (in keV/μm in water) following exposure to different heavy ions (He, C, O, F, Ne, and A). Data points combined from references [23, 24]; the line is a guide for the eye.
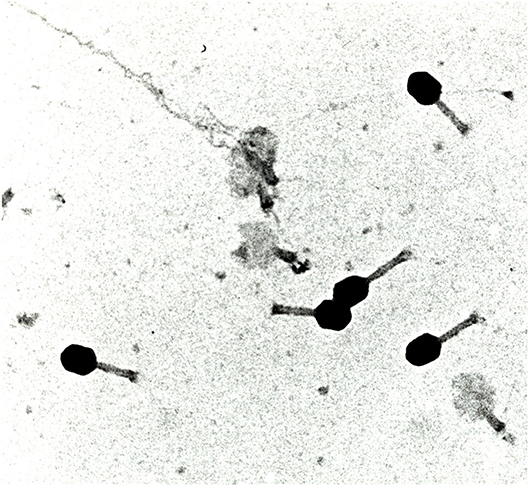
Figure 3. Images of radiation-induced damage in virus. The photo shows the bacteriophage T4 irradiated with protons (details in Ref. [25]). Black heads retain the DNA, and white heads have lost the molecule, which is seen flowing out of the envelope in some viruses. Electron microscope photograph from the Tandem accelerator of the University Federico II, Naples, Italy, courtesy of Prof. Gianfranco Grossi.
For high-energy heavy ions, part of the inactivation can derive from the high-energy electrons emitted along the tracks (δ-rays—see Figure 4). Virus targets were instrumental for the elaboration of the first amorphous track structure models of radiation by Robert Katz [26], where the radial dose is assumed to decrease as r−2 by increasing the distance r from the primary ion track. In the single-hit–single-target model, the inactivation cross-section for heavy ions can be written as a product of the geometrical cross-section times the inactivation probability (1 – S), dependent on the distance r from the track:
where D(r) is the radial dose, D0 is the mean lethal dose (derived from experiments with γ- or X-rays), and R is the maximum track radius (i.e., the range of the δ-rays with maximum energy). The calculation of σ requires several parameters to estimate the radial dose D(r) and the mean inactivation dose D0 from γ-ray experiments. The results of the Katz' model for dry enzymes and viruses are in good agreement with experimental data [27].
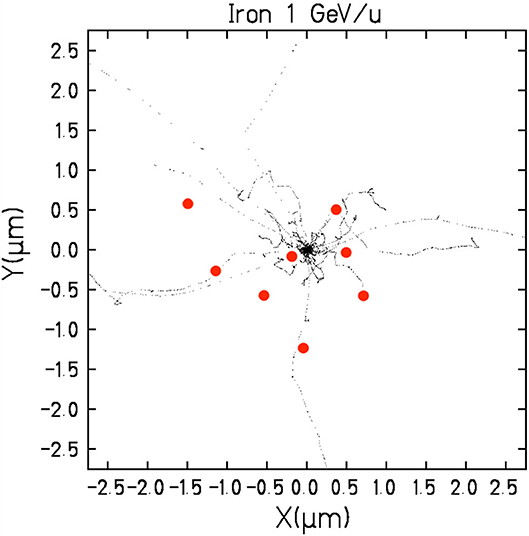
Figure 4. Track structure and virus. Simulation with the TRAX Monte Carlo code of a 1 GeV/n 56Fe-ions in water. Each black dot represents an ionization event. The track is seen exiting from the plane in bird's eye view. Axes are spatial coordinates in μm from the track center. The red spheres give the approximate size of the coronavirus.
Based on the Katz theory, Liu et al. [28] derived a simple analytical expression for the inactivation cross-sections of viruses:
where is the distance from the track corresponding to the γ-ray mean dose level D0, z is the ion effective charge, β is the ion velocity, R is the maximum track radius, C is a constant depending on the absorbing medium, and A and B are two free parameters. Using A = 3.88 and B = 0.753, the authors fitted very well the published results [28], showing that ion radiosensitivity of viruses can be accurately predicted from the γ-ray radiosensitivity.
A modified version of the amorphous track structure is still used today in treatment planning for heavy ion therapy in cancer patients [29].
Radiation and Vaccines
Beyond the basic radiobiology applications, irradiation of viruses was, since the beginning, used for vaccine development [30]. Despite the fact that the use of chemicals often requires extensive and time-consuming downstream processing in order to detoxify them, it has gradually overcome γ-ray sterilization, as it can easily be applied under good manufacturing practice (GMP) conditions. For instance, influenza viruses of the seasonal flu split or subunit vaccines are inactivated using chemical agents, such as formaldehyde [14]. However, the efficacy of these vaccines usually reaches only 60–70%, and even less in the elderly (~20–30%) [31]. This might be also in part explained by a negative impact of the chemicals on viral surface antigenic structures that are the targets of the human immune system for the elicitation of a protective response. In this regard, γ-irradiated influenza vaccines seem to be more effective not only at stimulating strong antigen-specific antibody production but also at priming cross-reactive cytotoxic T cells, thereby protecting mice against a heterologous influenza virus [32]. Similar results have been observed using gamma radiation for the development of vaccine prototypes against HIV [33], Ebola [12], rotavirus [16], and polio [34].
However, high doses of γ-rays also cause damage to the surface molecules. Radioprotectors can be used to limit this damage [34, 35], but they can also protect the nucleic acids, and therefore the net advantage is dubious. Even if there is not a clear evidence that γ-rays provide a better-quality inactivated virus than chemical methods, there is an increasing demand of these radiation sources to produce inactivated virus with reduced damage to surface antigenic proteins and no requirements to remove chemical compounds after inactivation [36].
Charged Particles for Vaccine Development
A new strategy to reduce the epitope damage while maintaining lesions to the nucleic acids can be the use of a different radiation quality. Electrons produced with linacs are commonly used for sterilization of materials [37]. High-energy electrons were soon used for virus inactivation as replacement of 60Co γ-ray sources [38] and until recently for food sterilization [39]. More recently, low-energy electrons have been explored because they present limited radioprotection problems and can be used in GMP laboratories. A beam of 200-keV electrons maintains the antigenic properties in several inactivated virus [40]. A Monte Carlo simulation of SARS-CoV-2 virus has shown that best results in terms of reduced damage to the spike proteins would be obtained with 2-keV electrons [41]. However, the main drawback of low-energy electrons is their limited range (in water, ~0.45 mm at 200 keV and ~0.2 μm at 2 keV), which makes it impossible to process large volumes of pathogen suspensions as necessary for vaccine manufacturing.
High-energy heavy ions (Figure 4) have instead long penetration distances and reduced attenuation compared to γ-rays and electrons. Compared to sparsely ionizing radiation, they can inactivate the virus with very limited damage to membrane epitopes, because a single high-LET ion can severely damage the nucleic acid but will touch the virus envelope only in the point of entrance and exit (Figure 5). The effectiveness in inactivation per unit dose is lower for particles compared to γ-rays, but the effectiveness per particle traversal increases with LET (Figure 2). For this very reason, we have recently performed a Monte Carlo calculation to evaluate the possible use of heavy ions for the production of SARS-CoV-2 vaccine [42]. The Geant4-DNA extension [43–46] of the Geant4 Monte-Carlo toolkit [47–49] was used to simulate ionizing particle tracks and energy deposition inside the SARS-CoV-2 model. We focused on the ratio of the damage to the spike proteins (SARS-CoV-2 epitope) [50] and strand breaks in the ~30-kbp single-stranded viral RNA. We will call this protein/RNA damage ratio P/R. Figure 6 gives the trend of the P/R ratio as a function of LET. Even if P/R depends not only on LET but also on the track structure [42], the trend in Figure 6 shows the expected advantage of using heavy ions, with a reduction of P/R of about an order of magnitude. Heavy ions such as Fe 1 GeV/n (Figure 4) have ranges of over 25 cm in water-equivalent materials, thus allowing irradiation of plastic boxes containing several cryovials with frozen virus, as often done in γ-irradiation inactivation [51]. High-energy heavy ions require of course large accelerators, but many of them are currently in operation or under construction and have intense programs in applied sciences, especially biomedical research [52].
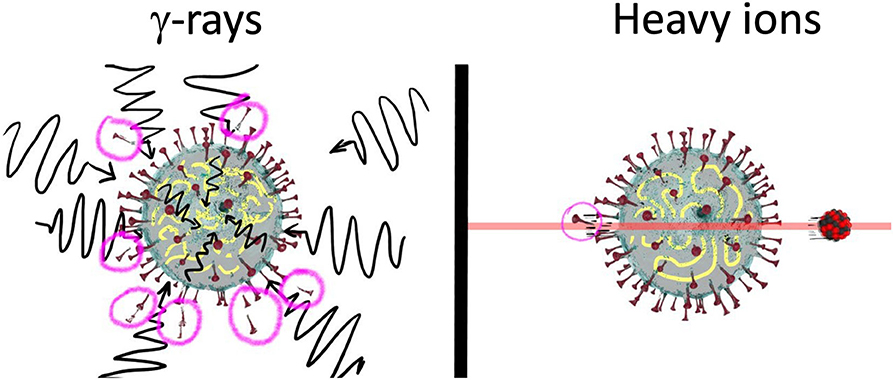
Figure 5. High- and low-LET radiation for virus inactivation. Schematic representation of the action of sparsely and densely ionizing radiation on SARS-CoV-2. High doses of γ-rays can inactivate the virus, but will damage many membrane proteins, whereas single (or few) heavy ion traversals will produce limited membrane damage while maintaining a high inactivation probability. Sparing of membrane epitopes is essential to elicit the immune response toward vaccine generation.
Figure 6. Simulated ratio of damages to membrane and nucleic acids. Geant4-DNA simulation of the ratio of the damages to spike proteins and RNA (P/R ratio) for radiation of different LET (in keV/μm in water). The curve provides only the trend as a function of the LET alone, the actual points showing the complex dependence from the velocity and charge can be found in reference [42].
Conclusions
Ionizing radiation has been used for decades to inactivate viruses. Early studies have contributed to our understanding of radiation action in living organisms. Inactivated viruses are still an important tool for vaccine development, and ionizing radiation has been used for years to this goal. One of the main problems of inactivated viruses is the damage to epitopes, which might reduce their ability to elicit an effective protective immune response post-vaccination. We have shown that densely ionizing heavy ions are potentially ideal to inactivate the virus with minimal damage to the envelope proteins and may therefore represent a new powerful tool for the development of vaccines against SARS-CoV-2 and other viruses.
Author Contributions
MD proposed the idea. KS and CG contributed to the section on vaccine production and virus. SI, ZF, and SZ contributed to the section on physics and simulation and in particular produced the results in Figure 6. All authors contributed to the article and approved the submitted version.
Funding
This work was partly supported by EU Horizon 2020 grant 730983 (INSPIRE).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank Daria Boscolo for the TRAX simulation image in Figure 4 and Serena Durante for the cartoon in Figure 5.
References
1. Wu F, Zhao S, Yu B, Chen YM, Wang W, Song ZG, et al. A new coronavirus associated with human respiratory disease in China. Nature. (2020) 579:265–9. doi: 10.1038/s41586-020-2008-3
2. Amanat F, Krammer F. SARS-CoV-2 vaccines: status report. Immunity. (2020) 52:583–9. doi: 10.1016/j.immuni.2020.03.007
3. Kaur SP, Gupta V. COVID-19 vaccine: a comprehensive status report. Virus Res. (2020) 288:198114. doi: 10.1016/j.virusres.2020.198114
4. WHO. Draft Landscape of COVID 19 Candidate Vaccines. Available online at: https://www.who.int/who-documents-detail/draft-landscape-of-covid-19-candidate-vaccines (accessed May 12, 2020).
5. Callaway E. The race for coronavirus vaccines: a graphical guide. Nature. (2020) 580:576–7. doi: 10.1038/d41586-020-01221-y
6. Gao Q, Bao L, Mao H, Wang L, Xu K, Yang M, et al. Development of an inactivated vaccine candidate for SARS-CoV-2. Science. (2020) 369:77–81. doi: 10.1126/science.abc1932
7. Wang H, Zhang Y, Huang B, Deng W, Quan Y, Wang W, et al. Development of an inactivated vaccine candidate, BBIBP-CorV, with potent protection against SARS-CoV-2. Cell. (2020) 182:713–21.e9. doi: 10.1016/j.cell.2020.06.008
8. Astill J, Alkie T, Yitbarek A, Taha-Abdelaziz K, Bavananthasivam J, Nagy É, et al. Examination of the effects of virus inactivation methods on the induction of antibody- and cell-mediated immune responses against whole inactivated H9N2 avian influenza virus vaccines in chickens. Vaccine. (2018) 36:3908–16. doi: 10.1016/j.vaccine.2018.05.093
9. Furuya Y, Regner M, Lobigs M, Koskinen A, Mullbacher A, Alsharifi M. Effect of inactivation method on the cross-protective immunity induced by whole “killed” influenza A viruses and commercial vaccine preparations. J Gen Virol. (2010) 91:1450–60. doi: 10.1099/vir.0.018168-0
10. Sabbaghi A, Miri SM, Keshavarz M, Zargar M, Ghaemi A. Inactivation methods for whole influenza vaccine production. Rev Med Virol. (2019) 29:e2074. doi: 10.1002/rmv.2074
11. Furuya Y. Return of inactivated whole-virus vaccine for superior efficacy. Immunol Cell Biol. (2012) 90:571–8. doi: 10.1038/icb.2011.70
12. Marzi A, Halfmann P, Hill-Batorski L, Feldmann F, Shupert WL, Neumann G, et al. An Ebola whole-virus vaccine is protective in nonhuman primates. Science. (2015) 348:439–42. doi: 10.1126/science.aaa4919
13. Arama C, Troye-Blomberg M. The path of malaria vaccine development: challenges and perspectives. J Intern Med. (2014) 275:456–66. doi: 10.1111/joim.12223
14. Pica N, Palese P. Toward a universal influenza virus vaccine: prospects and challenges. Annu Rev Med. (2013) 64:189–202. doi: 10.1146/annurev-med-120611-145115
15. Motamedi-Sedeh F, Soleimanjahi H, Jalilian AR, Mahravani H, Shafaee K, Sotoodeh M, et al. Development of protective immunity against inactivated Iranian isolate of foot-and-mouth disease virus type O/IRN/2007 using gamma ray-irradiated vaccine on BALB/c mice and Guinea pigs. Intervirology. (2015) 58:190–6. doi: 10.1159/000433538
16. Shahrudin S, Chen C, David SC, Singleton EV, Davies J, Kirkwood CD, et al. Gamma-irradiated rotavirus: a possible whole virus inactivated vaccine. PLoS ONE. (2018) 13:e0198182. doi: 10.1371/journal.pone.0198182
17. Bachofer CS. Direct Effect of X-rays on bacterial viruses, modified by physical state, in relation to the target theory. Science. (1953) 117:280–2. doi: 10.1126/science.117.3037.280
18. Pollard E. The action of ionizing radiation on viruses. Adv Virus Res. (1954) 2:109–51. doi: 10.1016/S0065-3527(08)60531-X
19. Wollman E, Holweck F, Luria S. Effect of radiations on bacteriophage C16. Nature. (1940) 145:935–6. doi: 10.1038/145935b0
20. Sullivan R, Fassolitis AC, Larkin EP, Read RB, Peeler JT. Inactivation of thirty viruses by gamma radiation. Appl Microbiol. (1971) 22:61–65. doi: 10.1128/AEM.22.1.61-65.1971
21. Smolko EE, Lombardo JH. Virus inactivation studies using ion beams, electron and gamma irradiation. Nucl Instrum Methods Phys Res B. (2005) 236:249–53. doi: 10.1016/j.nimb.2005.04.055
23. Schambra PE, Hutchinson F. The action of fast heavy ions on biological material: II. Effects on T1 and ϕX-174 bacteriophage and double-strand and single-strand DNA. Radiat Res. (1964) 23:514–26. doi: 10.2307/3571746
24. Fluke DJ, Brustad T, Birge AC. Inactivation of Dry T-1 bacteriophage by helium ions, carbon ions, and oxygen ions: comparison of effect for tracks of various ion density. Radiat Res. (1960) 13:788–808. doi: 10.2307/3570854
25. Gialanella G, Grossi GF, Macchiato MF, Napolitano M, Speranza PR. Contributions of various types of damage to inactivation of T4 bacteriophage by protons. Radiat Res. (1983) 96:462–75. doi: 10.2307/3576113
26. Butts JJ, Katz R. Theory of RBE for heavy ion bombardment of dry enzymes and viruses. Radiat Res. (1967) 30:855–71. doi: 10.2307/3572151
27. Chunxiang Z, Dunn DE, Katz R. Radial distribution of dose and cross-sections for the inactivation of dry enzymes and viruses. Radiat Prot Dosimetry. (1985) 13:215–8. doi: 10.1093/rpd/13.1-4.215
28. Liu X, Mianfeng Li, Yu KN. Inactivation cross section of ions for dry enzymes and viruses. Phys Med Biol. (1994) 39:1757–61. doi: 10.1088/0031-9155/39/10/016
29. Friedrich T, Scholz U, Elsässer T, Durante M, Scholz M. Calculation of the biological effects of ion beams based on the microscopic spatial damage distribution pattern. Int J Radiat Biol. (2012) 88:103–7. doi: 10.3109/09553002.2011.611213
30. Pollard E. Inactivation of viruses for vaccine purposes using ionizing radiation and heat. Yale J Biol Med. (1957) 29:436–43.
31. Simpson CR, Lone NI, Kavanagh K, Ritchie LD, Robertson C, Sheikh A, McMenamin J. Trivalent inactivated seasonal influenza vaccine effectiveness for the prevention of laboratory-confirmed influenza in a Scottish population 2000 to 2009. Eurosurveillance. (2015) 20:21043. doi: 10.2807/1560-7917.ES2015.20.8.21043
32. Alsharifi M, Müllbacher A. The γ-irradiated influenza vaccine and the prospect of producing safe vaccines in general. Immunol Cell Biol. (2010) 88:103–4. doi: 10.1038/icb.2009.81
33. Kang CY, Gao Y. Killed whole-HIV vaccine; employing a well established strategy for antiviral vaccines. AIDS Res Ther. (2017) 14:47. doi: 10.1186/s12981-017-0176-5
34. Tobin GJ, Tobin JK, Gaidamakova EK, Wiggins TJ, Bushnell RV, Lee WM, et al. A novel gamma radiation-inactivated sabin-based polio vaccine. PLoS ONE. (2020) 15:e0228006. doi: 10.1371/journal.pone.0228006
35. Gaidamakova EK, Myles IA, McDaniel DP, Fowler CJ, Valdez PA, Naik S, et al. Preserving immunogenicity of lethally irradiated viral and bacterial vaccine epitopes using a radio- protective Mn2+-peptide complex from deinococcus. Cell Host Microbe. (2012) 12:117–24. doi: 10.1016/j.chom.2012.05.011
36. Seo HS. Application of radiation technology in vaccines development. Clin Exp Vaccine Res. (2015) 4:145–58. doi: 10.7774/cevr.2015.4.2.145
37. Lancker M, Bastiaansen L. Electron-beam sterilization. Trends and developments. Med Device Technol. (2000) 11:18–21.
38. Traub FB, Friedemann U, Brasch A, Huber W, Kaplan H. High intensity electrons as a tool for preparation of vaccines. J Immunol. (1951) 67:379–84.
39. Mark Plavsic RWN. Efficacy of electron beam for viral inactivation. J Microb Biochem Technol. (2015) 7:173–76. doi: 10.4172/1948-5948.1000200
40. Fertey J, Bayer L, Grunwald T, Pohl A, Beckmann J, Gotzmann G, et al. Pathogens inactivated by low-energy-electron irradiation maintain antigenic properties and induce protective immune responses. Viruses. (2016) 8:319. doi: 10.3390/v8110319
41. Feng G, Liu L, Cui W, Wang F. Electron beam irradiation on novel coronavirus (COVID-19): A Monte–Carlo simulation. Chin Phys B. (2020) 29:048703. doi: 10.1088/1674-1056/ab7dac
42. Francis Z, Incerti S, Zein SA, Lampe N, Guzman CA, Durante M. Monte Carlo Simulation of SARS-CoV-2 Radiation-Induced Inactivation for Vaccine Development. (2020). Available online at: http://arxiv.org/abs/2005.06201 (accessed August 10, 2020).
43. Incerti S, Ivanchenko A, Karamitros M, Mantero A, Moretto P, Tran HN, et al. Comparison of GEANT4 very low energy cross section models with experimental data in water. Med Phys. (2010) 37:4692–708. doi: 10.1118/1.3476457
44. Incerti S, Kyriakou I, Bernal MA, Bordage MC, Francis Z, Guatelli S, et al. Geant4-DNA example applications for track structure simulations in liquid water: a report from the Geant4-DNA Project. Med Phys. (2018) 45:e722–39. doi: 10.1002/mp.13048
45. Incerti S, Baldacchino G, Bernal M, Capra R, Champion C, Francis Z, et al. The GEANT4-DNA project. Int J Model Simul Sci Comput. (2010) 01:157–78. doi: 10.1142/S1793962310000122
46. Bernal MA, Bordage MC, Brown JMC, Davídková M, Delage E, El Bitar Z, et al. Track structure modeling in liquid water: a review of the Geant4-DNA very low energy extension of the Geant4 Monte Carlo simulation toolkit. Phys Medica Eur J Med Phys. (2015) 31:861–74. doi: 10.1016/j.ejmp.2015.10.087
47. Agostinelli S, Allison J, Amako K, Apostolakis J, Araujo H, Arce P, et al. Geant4—a simulation toolkit. Nucl Instrum Methods Phys Res A. (2003) 506:250–303. doi: 10.1016/S0168-9002(03)01368-8
48. Allison J, Amako K, Apostolakis J, Araujo H, Dubois PA, Asai M, et al. Geant4 developments and applications. IEEE Trans Nucl Sci. (2006) 53:270–8. doi: 10.1109/TNS.2006.869826
49. Allison J, Amako K, Apostolakis J, Arce P, Asai M, Aso T, et al. Recent developments in Geant4. Nucl Instrum Methods Phys Res A. (2016) 835:186–225. doi: 10.1016/j.nima.2016.06.125
50. Walls AC, Park YJ, Tortorici MA, Wall A, McGuire AT, Veesler D. Structure, function, and antigenicity of the SARS-CoV-2 spike glycoprotein. Cell. (2020) 181:281–92.e6. doi: 10.1016/j.cell.2020.02.058
51. David SC, Lau J, Singleton EV, Babb R, Davies J, Hirst TR, et al. The effect of gamma-irradiation conditions on the immunogenicity of whole-inactivated Influenza A virus vaccine. Vaccine. (2017) 35:1071–9. doi: 10.1016/j.vaccine.2016.12.044
Keywords: COVID-19, SARS-CoV-2, virus, gamma rays, heavy ions
Citation: Durante M, Schulze K, Incerti S, Francis Z, Zein S and Guzmán CA (2020) Virus Irradiation and COVID-19 Disease. Front. Phys. 8:565861. doi: 10.3389/fphy.2020.565861
Received: 26 May 2020; Accepted: 24 August 2020;
Published: 20 October 2020.
Edited by:
Zhen Cheng, Stanford University, United StatesReviewed by:
David B. Stout, Independent Researcher, Culver City, United StatesHui Shi, Stanford Bio-X, Stanford University, United States
Copyright © 2020 Durante, Schulze, Incerti, Francis, Zein and Guzmán. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Marco Durante, bS5kdXJhbnRlQGdzaS5kZQ==