
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Pharmacol. , 03 March 2025
Sec. Ethnopharmacology
Volume 16 - 2025 | https://doi.org/10.3389/fphar.2025.1534798
This article is part of the Research Topic Food Derived Bioactive Metabolites: Unlocking their Potential Health Benefits and Medical Potential View all 9 articles
 Min Jin1‡Linfang Wei2‡
Min Jin1‡Linfang Wei2‡ Jianhua Wang3
Jianhua Wang3 Yuehong Shen4Lei Gao4Fan Zhao4Qianying Gao4Yifei Ma4Yongyan Sun4Ying Lin4
Yuehong Shen4Lei Gao4Fan Zhao4Qianying Gao4Yifei Ma4Yongyan Sun4Ying Lin4 Guanjie Ji5*†
Guanjie Ji5*† Pingping Cai6*†Rugen Yan4*†
Pingping Cai6*†Rugen Yan4*†Formononetin (FMN) is a common natural metabolite that can be extracted and isolated from some common botanical drugs. In recent years, FMN has garnered increasing attention due to its beneficial biological activities. In this paper, we systematically summarize the sources of FMN and provide a comprehensive review of its pharmacological activities and molecular mechanisms, co-administration, toxicity, derivatives, and drug delivery systems in the last 5 years. The study results found that FMN has a wide range of pharmacological activities in neurological disorders, organ damage and cancer, showing great potential for clinical application and broad prospects. Researchers are exploring various types of delivery systems, including nanoparticle carriers, ligand modifications and polymer microspheres. These advanced delivery systems can enhance the stability of FMN, prolong its release time in vivo, and improve targeting, thereby optimizing its therapeutic efficacy and reducing side effects, and greatly improving its bioavailability. In conclusion, FMN is a natural metabolite with considerable research value, and its diverse biological activities make it a promising candidate for drug development and medical research.
Flavonoids are a class of natural metabolites widely found in plants and foods, belonging to an important branch of polyphenolic compounds, which have a wide range of biological activities, including anti-inflammatory, antidiabetic, and anticancer (Billowria et al., 2024). Because of their different structural characteristics, they are divided into different types, thus displaying various functions. Among them, isoflavones are a unique large class of flavonoids, mainly found in soybeans and legumes, which have estrogenic activity and can be combined with estrogen receptors. Phytoestrogens are not hormones in the traditional sense, but a secondary metabolite product of plants. It is similar to animal estrogen in structure and function, and can have many effects on health, which may help to prevent certain hormone-related diseases, such as breast cancer and osteoporosis (Patra et al., 2023).
FMN is a kind of isoflavone phytoestrogen, which is widely found in plants and common diets. It has important biological activities, and has become the current research hotspot in the field of hormones. Modern research has shown that FMN is one of the active ingredients in some commonly used botanical drugs, such as Astragalus mongholicus, Pueraria montana, and Trifolium pratense. With the deepening of modern pharmacological research, the medicinal value of FMN has been further developed. Currently, the research on the pharmacological effects, combination application and drug delivery system of FMN has been carried out in-depth, indicating that FMN has a variety of pharmacological activities and clinical application potential as a single-component drug, and is expected to become a promising drug. In the past few years, the pharmacological effects of FMN have been widely reported. However, most of the previous reports were scattered and lacked systematic summarization and generalization. Therefore, this article aims to summarize the reports on FMN in the past 5 years, which will provide a basis for the future development and clinical application of FMN.
FMN is a well-known flavonoid with the parent structure of isoflavones, so it is also known as 7-hydroxy-4′-methoxy isoflavone, with the molecular formula C16H12O4, relative molecular weight 268.26 (Křížová et al., 2019), and the chemical structure is shown in Figure 1. Structurally, FMN has only one phenolic hydroxyl group, so it is poorly soluble in water and easily soluble in organic solvents such as methanol, ethyl acetate, and ether. FMN is a secondary metabolite produced by plants, predominantly found in legumes, as shown in Table 1. In addition, it is also present in normal diets, such as milk (Andersen et al., 2009), beer (Lapcík et al., 1998), and coffee (Alves et al., 2010).
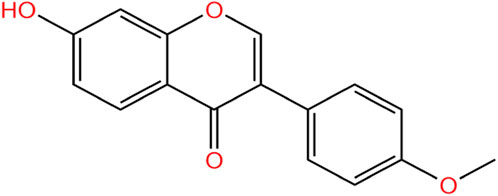
Figure 1. Chemical structures of FMN.
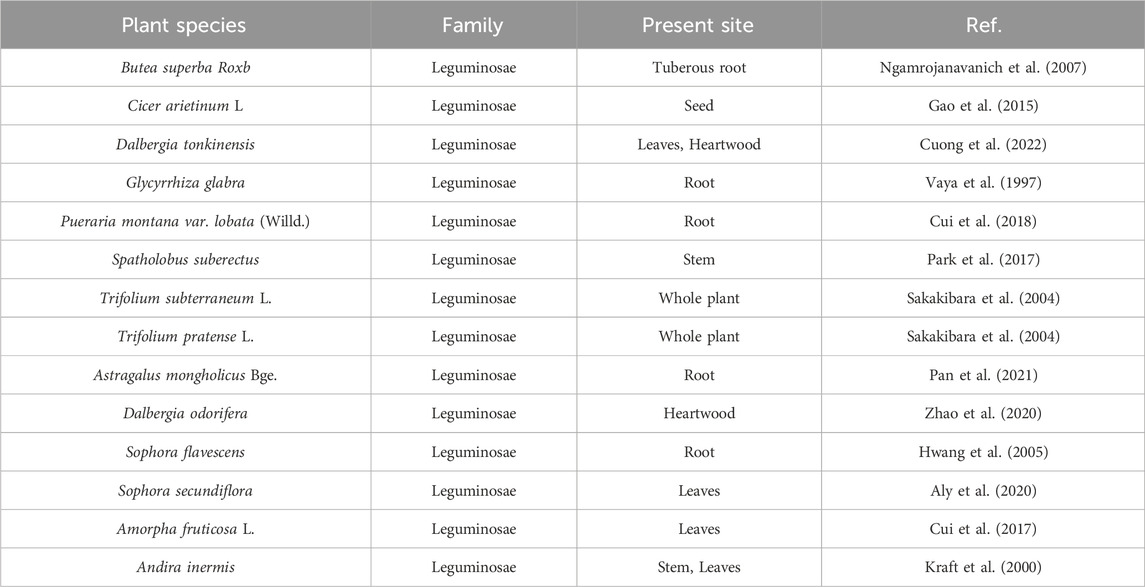
Table 1. Plant origin of FMN.
Alzheimer’s disease (AD) is a progressive neurodegenerative disease and has been listed as a global priority public health problem (Ren and Qu, 2023), and its pathogenesis is related to the hyperphosphorylation of tau protein and Aβ deposition. It had been found that FMN, isolated from Sophora secundiflora, could improve memory problems by restoring the level of oxidative stress in brain tissues and modulating acetylcholinesterase activity. It was a potential neuroprotective agent for further research and development of AD treatment in the pharmaceutical industry (Aly et al., 2020; Sugimoto et al., 2021). Inflammatory responses could lead to vascular damage in the brain, disrupting the blood-brain barrier, promoting Aβ deposition and aggregation, and accelerating the development of AD (Silva et al., 2019). Therefore, inhibition of cerebral vascular inflammation plays a crucial role in the pathogenesis of AD. Fan et al. (2022) treated human brain microvascular endothelial cells (HBMECs) with Aβ25–35 for 12 h and performed quantitative real-time reverse transcription polymerase chain reaction (RT-qPCR). The results of western blot analysis showed that the expression of intracellular adhesion molecule-1 (ICAM-1), vascular cell adhesion molecule-1 (VCAM-1) and E-selectin protein increased, and nuclear factor-κB [NF-κB (p65)] was translocated to the nucleus of HBMECs. But after the administration of FMN, the expression levels of the above three decreased and NF-κB activation was inhibited, which may be related to the release of FMN blocking kelch-like ECH-associated protein-1 (Keap1) and activating the nuclear factor erythroid 2-related factor 2 (Nrf2) signaling pathway. Similarly, FMN exerted anti-neuroinflammatory effects by targeting peroxisome proliferator-activated receptor coactivator-1α (PGC-1α) and bidirectionally regulating NF-κB signaling pathway and Nrf2/Heme oxygenase-1 (HO-1) signaling pathway, thereby inhibiting tau protein hyperphosphorylation. Significantly FMN improve cognitive dysfunction in mice caused by high-fat feeding (Fu et al., 2019).
Neurotransmitters are chemicals in the nervous system that regulate the transmission of signals between neurons. Disruptions in their function can lead to alterations in emotional and cognitive processes, potentially contributing to conditions such as depression and anxiety (Lee, 2013). In mice models of depression established by chronic corticosterone (CORT) injection, FMN significantly increased sucrose preference and shortened the resting time of forced swimming. Using ELISA kits and Western blot analysis, researchers found that FMN decreased serum corticosterone levels and upregulated the protein expression levels of the glucocorticoid receptor (GR) and brain-derived neurotrophic factor (BDNF) in the hippocampus. Additionally, FMN mitigated CORT-induced neuronal damage in the CA1 and CA3 regions of the hippocampus and promoted neurogenesis in this brain area (Zhang et al., 2022). In addition, the inflammatory response could indirectly affect the development of depression by influencing body system diseases. Yang J. et al. (2023) demonstrated for the first time that FMN improved depressive behaviors in mice with myocardial infarction-associated depression. The underlying mechanism involves the reduction of interleukin-6 (IL-6) and IL-17A by inhibiting glycogen synthase kinase-3β (GSK-3β) activity and downregulating downstream signaling molecules, including Notch homolog 1 (Notch1) and CCAAT/enhancer binding protein α (C/EBPα). This, in turn, promoted macrophage/microglia polarization toward the M2 phenotype, reduced neuroinflammation, increased BDNF and 5-hydroxytryptamine (5-HT) levels, and mitigated the progression of depression in mice. To investigate the anxiolytic effect of FMN, Wang X. S. et al. (2019) used a complete Freund’s adjuvant (CFA)-induced anxiety mice model in mice and administered FMN continuously for 14 days. The results showed that FMN significantly increased the open-arm time in the elevated plus maze test and the central area time in the open-field experiment in mice, and had a favorable anxiolytic effect. Molecular mechanism studies showed that FMN may alleviate the inflammation and neuronal hyperexcitability in the basolateral amygdala and alleviate the anxiety-like behavior in mice by inhibiting the NF-κB and N-methyl-D-aspartate (NMDA)/cAMP-response element binding protein (CREB) signaling pathways. Thus, FMN can be used as a new candidate drug for the treatment of depression and anxiety disorders.
Cerebral ischemia-reperfusion (I/R) injury refers to the phenomenon that the injury is further aggravated and deteriorates during reperfusion after reversible ischemic brain injury, which has a high mortality and disability rate. FMN could target Janus kinase 2 (JAK2)/signal transducers and activators of transcription 3 (STAT3) and phosphatidylinositol 3-kinase (PI3K)/protein kinase B (AKT)/extracellular regulated protein kinases (ERK) signaling pathways. It could significantly reduce the level of inflammatory factors, increase the number of dendritic spines in neurons, and increase the expression of βIII-tubulin, growth-associated protein 43 (GAP-43), nerve growth factor (NGF) and BDNF. FMN could protect the neurological dysfunction and pathological changes of brain tissue in rats with arterial occlusion from both anti-inflammatory and neuroprotective aspects (Wu Y. et al., 2020; Yu et al., 2022). Other studies showed that poly(ADPribose) polymerase-1 (PARP-1) and poly (ADP-ribose) glycohydrolase (PARG) played key roles in ischemic neuronal cell death and disease progression. When the activities of both are inhibited, they could effectively reduce the area of cerebral infarction, eliminate the inflammatory response, and restore neurological function in stroke patients (Cuzzocrea and Wang, 2005; Liu et al., 2022). On this basis, Luo et al. (2024) used PARP-1 inhibitor PJ34 and PARG inhibitor ethacridine lactate to investigate whether FMN’s mechanism of action involves PARP-1 and PARG. The result found that FMN significantly reduced PARP1, PARG, apoptosis-inducing factor (AIF), cysteinyl aspartate-specific protease 3 (caspase-3) and p53 protein in rats with cerebral ischemia-reperfusion injury, and increased the expression of endogenous neuroprotective gene Iduna, effectively reducing the area of cerebral infarction and neuronal apoptosis in rats. These findings suggested that FMN may serve as a potential inhibitor of PARP-1 and PARG for the treatment of cerebral I/R injury.
Spinal cord injury (SCI) is a devastating neurological state that leads to impaired sensory and motor functions and has a significant impact on the global healthcare system (Anjum et al., 2020). FMN can improve neurological function and exert neuroprotective effects in spinal cord injury by reducing local apoptosis and inflammatory cell infiltration. Zhou and Zhang (2023) treated PC12 cells with lipopolysaccharide (LPS) and different concentrations of FMN (50 μM, 100 μM, 200 μM). They found that FMN decreased tumor necrosis factor-α (TNF-α), IL-1β, IL-6, p-p65 NF-κB, nucleotide-binding oligomerization domain-like receptor protein 3 (NLRP3) and lactate dehydrogenase (LDH) levels, increased proliferating cell nuclear antigen (PCNA) expression, increased cell viability, inhibited apoptosis, and significantly ameliorated LPS-induced inflammatory injury in neuronal cells. In another report, Fu et al. (2023) demonstrated in vivo and in vitro that FMN attenuated microglial cell inflammatory response to spinal cord injury by inhibiting the epidermal growth factor receptor (EGFR)/mitogen-activated protein kinase (p38 MAPK) signaling pathway, thereby promoting nerve injury repair in rat spinal cord.
Oxaliplatin, a platinum-based chemotherapeutic agent, is commonly used in the treatment of metastatic rectal cancer but is limited in clinical application due to its severe neuropathy. Fang et al. (2020) found that FMN had a favorable ameliorative effect on oxaliplatin-induced peripheral neuropathy and did not affect the chemotherapeutic function of oxaliplatin. Molecular mechanism studies demonstrated that FMN had this effect because it could target the activation of the Nrf2 pathway, increase the activity of the phase II metabolizing enzyme glutathione S-transferase pi 1 (GSTP1), and inhibit oxaliplatin-induced peripheral neuropathy by protecting mitochondrial function.
Diabetic peripheral neuropathy is one of the most common chronic complications of diabetes. The main symptoms include spontaneous refractory pain, which may last for weeks to months. Currently, there are no drugs specifically for the treatment of diabetic peripheral neuropathy on the market. Drugs for the prevention and treatment of diabetic peripheral neuropathy also have strong side effects, which make clinical application difficult (Yang et al., 2022). In both in vivo and in vitro experiments, FMN was shown to be a potential therapeutic agent for diabetic peripheral neuropathy. It promoted the release of NGF by activating silenced information regulatory factor 3 (SIRT3), inhibited oxidative stress, enhanced neuronal survival, restored mitochondrial function, and significantly increased the pain threshold in neuropathic rats, as demonstrated in the hot-plate and tail-dip experiments (Oza and Kulkarni, 2020; Jiang et al., 2023). The neuroprotective mechanism of FMN is shown in Figure 2.
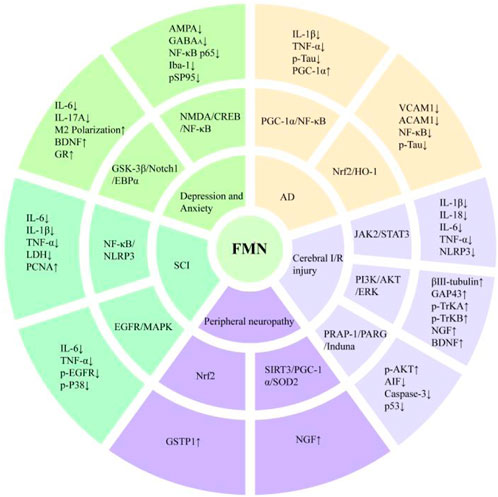
Figure 2. Neuroprotective mechanism of FMN.
FMN has good therapeutic effects on pulmonary arterial hypertension (PAH), hyperoxia-induced acute lung injury (ALI), and chronic obstructive pulmonary disease (COPD). Cai et al. (2019) used monocrotaline (MCT) to induce PAH in rats and administered FMN. The results indicated that FMN may provide a protective effect against MCT-induced PAH in rats by inhibiting the PI3K/AKT and ERK pathways, decreasing the expression of α-smooth muscle actin (α-SMA), PCNA, and B-cell lymphoma 2 (Bcl-2), thereby preventing the excessive proliferation of pulmonary artery smooth muscle cells (PASMCs) and pulmonary vascular remodeling. Similarly, Wu J. et al. (2020) found that FMN inhibited pulmonary vascular remodeling in MCT-induced PAH rats by down-regulating the expression of transforming growth factor-β1 (TGF-β1), matrix metalloproteinase-2 (MMP-2), MMP-9, collagen types I and III, and fibronectin. It also reduced the dense focal collagen deposition induced by MCT. In addition, FMN alleviated MCT-induced lung inflammation by decreasing the expression of inflammatory cytokines, including TNF-α, IL-1β, and monocyte chemotactic protein-1 (MCP-1). Chen et al. (2021) exposed C57BL/6 mice to hyperoxia for 72 h to establish an ALI model. Intraperitoneal injection of FMN significantly reduced hyperoxia-induced increases in lung water content, pro-inflammatory cytokine levels, and lung neutrophil infiltration in mice. Western blot analysis showed that FMN increased the expression and activity of Nrf2, HO-1, and superoxide dismutase (SOD), thereby promoting the polarization of macrophages to the M2 phenotype. In another report, Li et al. (2024a) established the COPD mice model by exposing mice to cigarette smoke (CS) for 24 weeks and treated bronchial epithelial BEAS-2B cells with CS extract for 24 h to investigate the in vivo and in vitro effects of FMN on COPD. The results showed that FMN inhibited the activation of aryl hydrocarbon receptor (AhR)/CYP1A1 and AKT/mechanistic target of rapamycin (mTOR) signaling pathways, reduced CS-induced inflammatory response, endoplasmic reticulum stress and apoptosis, and effectively improved lung function and airway remodeling in COPD mice, both in vivo and in vitro.
Liao et al. (2021) found that FMN may be a potential active ingredient in the prevention or treatment of liver injury by network pharmacology combined with biochemical assays, which could reduce hepatotoxicity and improve liver function through inflammatory molecular pathways. In the concanavalin A (ConA)-induced autoimmune hepatitis (AIH) mice model, FMN-treated mice showed reduced hepatocyte swelling and inflammatory cell infiltration (Liu G. et al., 2021). Further molecular mechanism studies showed that FMN inhibited the activation of the NF-κB signaling pathway and NLRP3 inflammasome, and reduced the serum levels of alanine aminotransferase (ALT) and aspartate aminotransferase (AST) in mice. At the same time, FMN upregulated the expression of Bcl-2, downregulated the expression of Bcl-2-associated X protein (Bax), cleaved caspase 9, cleaved caspase 3 expression in liver tissue, and inhibited ConA-induced apoptosis in mice hepatocytes.
The process of hepatic ischemia-reperfusion injury involves the regulation of multiple factors, among which mitochondrial damage is one of the causative factors (Cannistrà et al., 2016). Ma et al. (2022) found that FMN could protect against I/R-induced hepatic injury by restoring the function of mitochondrial autophagy. Compared with the model group, the FMN group could effectively improve sinus congestion and cell swelling in rat liver tissue, and reduce serum AST, ALT, TNF-α and IL-1β levels. In addition, through the prohibitin-2 (PHB2)/PTEN-induced putative kinase protein 1 (PINK1)/Parkin signaling pathway, the levels of LC3II, Beclin1, p62, cyclooxygenase-2 (COX2), COX4, MMP and adenosine triphosphate (ATP) were increased, and the liver mitochondrial autophagy function and energy metabolism failure were restored. In addition, during this process, the activity of antioxidant proteins glutathione (GSH), catalase (CAT), GSH-PX in the FMN treatment group recovered, and the levels of reactive oxygen species (ROS) and malondialdehyde (MDA) decreased. These findings suggest that FMN can inhibit oxidative stress in the liver and restore mitochondrial function.
Liver injury is not only a direct lesion at the liver, but other pathologies can also indirectly lead to the occurrence of liver injury, such as cholestasis. Cholestasis is characterized by intracellular bile acid (BA) overload in hepatocytes, based on which FMN can improve hepatic/systemic BA metabolism and protect the liver from liver injury caused by intracellular retention of BA. Yang et al. (2019) induced cholestasis in the mice model using α-naphthylisothiocyanate (ANIT) and used FMN by gavage for 10 days. The results showed that FMN significantly improved gallbladder enlargement and bile color change in the cholestasis model mice. qRT-PCR and Western blot analyses revealed that FMN targeted sirtuin 1 (SIRT1) and peroxisome proliferator-activated receptor alpha (PPARα), reducing the levels of inflammatory markers, promoting bile flow from hepatocytes to the bile ducts, improving liver and portal vein bile acid transport, and inhibiting ANIT-induced liver injury and cholestasis. These findings suggest that FMN may serve as a potential therapeutic strategy for cholestatic liver disease.
Kidney disease has become an significant threat to human death. There are about 850 million patients with kidney disease worldwide, and the global prevalence of chronic kidney disease (CKD) is more than 10% (Feng et al., 2023). Regulation of cell death is a necessary condition for maintaining normal physiological homeostasis. Various forms of cell death, including apoptosis, autophagy, necrosis, and pyroptosis, are involved in the pathogenesis of kidney disease. Iron ptosis is the accumulation of iron-dependent lipid peroxides, which is a new mode of cell death and is associated with acute kidney injury under a variety of stimuli, such as ischemia-reperfusion, sepsis, or toxins, and CKD (Bayır et al., 2023; Li et al., 2024). FMN increased the expression of SLC7A11, glutathione peroxidase 4 (GPX4) and Nrf2, reduced the level of 4-hydroxy-2-nonenal (4-HNE), and negatively regulated iron sagging by inhibiting the translocation of Smad3 from the cytoplasm to nucleus. At the same time, it reduced the expression of fibrosis genes including α-SMA, Col1a1 and fibronectin, and hindered iron apoptosis-related fibrosis, thereby alleviating CKD (Zhu et al., 2023). In addition, FMN also had a good repair effect on drug-induced renal injury. In the rat model of kidney injury induced by gentamycin and methotrexate, FMN prevented kidney tissue damage and renal function reduction caused by gentamycin and methotrexate administration to a large extent (Aladaileh et al., 2019; Althunibat et al., 2022). This was all related to the enhancement of Nrf2, HO-1, GSH and SOD expression, reduction of MDA, TNF-α, IL-1β, COX-2, and inducible nitric oxide synthase (iNOS) levels, and enhancement of renal antioxidant and anti-inflammatory capacity. Hao et al. (2021) demonstrated the protective effect of FMN against renal injury due to cisplatin treatment by in vivo and in vitro experiments. PPARα is a ligand-activated nuclear hormone receptor transcription factor that regulates the expression of genes related to inflammation and cellular lipid metabolism and contributes to the maintenance of renal function (Iwaki et al., 2019). FMN upregulated PPARα expression in a ligand-dependent manner to increase the levels of Nrf2, HO-1, NAD(P)H: quinone oxidoreductase 1 (NQO1), and CAT, decreased the levels of MDA, TNF-α, and IL-1β, inhibited myeloperoxidase (MPO) activity, reduced renal proximal tubular cell apoptosis, oxidative stress, and inflammatory response. Thus, FMN is a promising drug to prevent drug-induced organ damage and can be used as a first-line treatment for acute kidney injury in future studies.
Diabetic nephropathy (DN) is one of the most common and serious complications of diabetes, which is related to the increased morbidity and mortality of diabetic patients (Samsu, 2021). In a high-glucose pathological state, elevated glucose levels stimulate cells to produce excessive ROS, activating various downstream inflammatory signaling pathways. This process induces and accelerates the accumulation of renal inflammatory fibrosis markers, such as fibronectin (FN) and ICAM-1, ultimately leading to renal fibrosis characterized by glomerulosclerosis (Li Y. et al., 2022). Therefore, oxidative stress is considered to be a major pathogenic factor in diabetic nephropathy. In vivo and in vitro experiments demonstrated that FMN could reduce inflammation and fibrosis within the glomerulus by increasing sirtuin-1 (Sirt1) protein levels in renal tissues, activating the Nrf2/ARE signaling pathway, promoting the expression of downstream antioxidant enzymes, inhibiting oxidative stress, and then down-regulating the levels of FN and ICAM-1. Restoring renal function indexes in mice with DN (Zhuang et al., 2020). In addition, FMN could reduce mitochondrial superoxide production and attenuate the loss of mitochondrial membrane power by increasing the expression of PGC-1α, a downstream target of Sirt1, and Mfn2 in the kidney, and decreasing the expression of Drp1 and Fis1, thus improving mitochondrial homeostasis and restoring renal function in rats (Huang et al., 2022). Based on oxidative stress injury, Smad3 directly binds to the promoter region to activate COl-I and COl-III, which promotes extracellular matrix (ECM) synthesis, leading to fibroblast activation and enhancing the pathological process of renal fibrosis (Dan Hu et al., 2023). FMN reduced Smad3, COl1a1, and COl3a1 mRNA and protein expression, attenuated α-SMA protein overactivation, reduced collagen production and deposition, and improved renal fiber structure (Lv et al., 2020). The above studies have shown that FMN can be used as a potential drug for the treatment of diabetic nephropathy.
Gastric ulcer is a disease characterized by chronic inflammation and barrier damage of the gastric mucosa, affecting 5%–10% of the population. Its etiology is closely related to excessive drinking, use of non-steroidal anti-inflammatory drugs and infection with helicobacter pylori (Mousa et al., 2019). Due to the side effects and resistance to conventional medications such as cimetidine, amoxicillin, and omeprazole, there is a need to find new therapeutic approaches. Rubab et al. (2022) used ethanol-induced gastric ulcer rats model to explore whether Caragana ambigua had anti-gastric ulcer effect and used liquid chromatography-mass spectrometry for component analysis. The results showed that FMN in C. ambigua significantly reduced the gastric ulcer index in the gastric tissue of model rats, and increased the pH value of gastric juice, gastric parietal protein and gastric mucus content. The results of histopathological observation showed that FMN could also significantly reduce gastric mucosal injury, submucosal edema and leukocyte infiltration in rats. In the rats model of gastric ulcers stimulated by non-steroidal anti-inflammatory drugs, FMN also played the same role (Mendonça et al., 2020). Yi et al. (2022) found that FMN could significantly improve the phenomenon of exfoliation necrosis and a large number of inflammatory cell infiltration in some areas of gastric mucosa induced by acetic acid in rats. At the same time, FMN upregulated the levels of CD34 protein, tight junction protein, vascular endothelial growth factor (VEGF) and nitric oxide (NO) in a dose-dependent manner, downregulated the level of human endothelin-1 (ET-1), and promoted gastric mucosal angiogenesis. In addition, compared with the model group, FMN could effectively reduce the levels of TNF-α, IL-1β, IL-6, p-P65/P65 and MPO activity, and increase the level of p-IκBα/IκBα, suggesting that FMN could play an anti-gastric ulcer role by inhibiting the activation of NF-κB signaling pathway. The above studies have shown that FMN can treat gastric ulcers through damage repair and anti-inflammatory effects. The organ protective mechanism of FMN is shown in Figure 3.
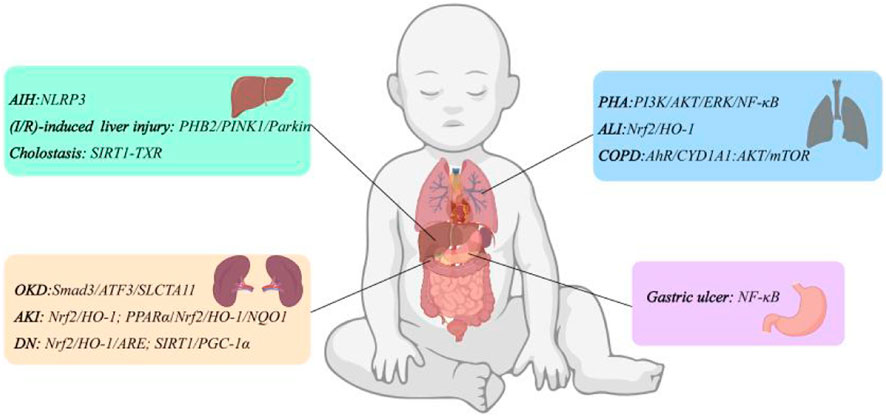
Figure 3. Organ protective mechanism of FMN.
Atherosclerosis (AS) is a chronic inflammatory disease characterized by lipid deposition and persistent inflammation within the arterial wall (Zhu et al., 2018). In the early stage of AS, the primary trigger of AS is vascular endothelial dysfunction, which is caused by vascular endothelial cell damage and hemodynamics (Wang et al., 2023). Zhang et al. (2021) discovered through network pharmacology and in vitro experiments that FMN alleviated inflammation, oxidative stress, and apoptosis responses in HUVECs induced by oxidized low-density lipoprotein by activating peroxisome proliferator-activated receptor-γ (PPAR-γ) signaling. He et al. (2024) further demonstrated that FMN upregulated the expression of α7nAChR, CD206, IL-10, SHIP1, and Arg-1, while down-regulating the expression of CD68, iNOS, COX-2, miR155-5p, IL-6, and IL-1β. FMN also promoted macrophage polarization to the M2 phenotype, thereby reducing inflammation and atherosclerosis in ApoE^−/− mice by inhibiting the JAK/STAT signaling pathway. In addition, foam cells play a key role in the development of AS, composed of macrophages, vascular smooth muscle cells, and other types of cells that adhere to endothelial cells, ingesting accumulated lipids and lipid products as foci for growing lesions (Galindo et al., 2023). Ma et al. (2020) found that FMN reduced vascular smooth muscle cell and macrophage-derived foam cell formation and their accumulation in the arterial wall, which may be associated with decreased SRA expression and reduced monocyte adhesion. Therefore, foam cells may serve as a potential breakthrough for FMN in the treatment of AS.
Zhou et al. (2022) found that FMN can reduce vascular endothelial injury caused by deep vein thrombosis in rats by increasing the expression of endothelial nitric oxide synthase (eNOS) and NO production. On this basis, Wu et al. (2020) further determined through in vivo and in vitro experiments that the mechanism by which FMN exerted its effects was due to the activation of the ERK1/2 and AKT signaling pathways, which in turn significantly enhanced endothelial function. In addition, FMN dose-dependently increased the levels of insulin-like growth factor 1 (IGF-1), ICAM-1, and VEGF, and promoted the proliferation, migration, and angiogenesis of HUVECs (Liang et al., 2019).
Diabetes induces vascular endothelial dysfunction, a critical and initiating factor in the development of diabetic vascular complications (Shi and Vanhoutte, 2017). The JAK/STAT signaling pathway is involved in the development of diabetic vascular complications (Banes-Berceli et al., 2007). FMN acted as an inhibitor of JAK, significantly reducing the phosphorylation and mRNA expression of JAK2 and STAT, as well as down-regulating the protein and mRNA levels of IL-1β and ICAM-1. Additionally, FMN restored NO synthesis and phenylephrine-induced contraction and acetylcholine-induced relaxation in the aortic tissues of rats on a high-glucose diet in a dose-dependent manner (Zhou et al., 2019). These findings suggested that FMN may be a new candidate natural metabolite for preventing and treating diabetic vascular complications.
FMN was protective against both cardiac I/R injury and cardiac fibrosis. For cardiac I/R injury, FMN significantly attenuated cardiac dysfunction, infarct size, and release of cardiac markers, and inhibited the elevation of inflammatory factors TNF-α, IL-1β, and IL-6 in rats with cardiac I/R injury, primarily by inhibiting the activation of the ROS-TXNIP-NLRP3 pathway (Wang et al., 2020). Regarding cardiac fibrosis, FMN improved mitochondrial function by downregulating MOAB mRNA expression and upregulating the mRNA expression of aldehyde dehydrogenase 2 (ALDH2) and hydroxyacyl-CoA dehydrogenase (HADH). Additionally, it significantly inhibited isoproterenol-induced cardiac fibrosis and the expression of fibrosis-related proteins and genes in mice (Qian et al., 2024).
Osteoporosis is a common clinical pathologic bone disease, which is the result of the imbalance between the functions of osteoblasts and osteoclasts and is most common in the elderly and postmenopausal women. Estrogen levels decline in postmenopausal women, leading to disruptions in the differentiation and activation of osteoblasts, reduced synthesis and deposition of new bone, and ultimately osteoporosis. FMN belonged to isoflavones and had estrogen-like effects. It can promote osteoblast differentiation by activating the p38 MAPK/Smad/bone morphogenetic protein (BMP) signaling pathway (Soundharrajan et al., 2019). In addition, enhanced osteoclast resorption is another important mechanism in the development of osteoporosis. Under the stimulation of inflammatory factors, mouse bone marrow-derived macrophages (BMMs) can differentiate into osteoclasts, increasing osteoclast bone phagocytosis capacity and leading to increased bone loss (de Villiers, 2023). The joint inflammatory response caused by CoCrMo particles produced by the implanted prosthetic biomaterials will also induce the formation, differentiation, and maturation of osteoclasts, and ultimately transform the homeostasis of bone metabolism into osteolysis (Yin et al., 2023). In response to this process, FMN could prevent this process by inhibiting the activation of the NF-κB/MAPK signaling pathway and inhibit osteolysis triggered by osteoclast over-resorption (Yu et al., 2023), making it a potential therapeutic agent for the prevention and treatment of periprosthetic osteolysis. It can be seen that FMN can improve osteoporosis disease by reducing the inflammatory response, inhibiting BMMs differentiation to osteoclasts, and reducing osteoclast resorption, which is instructive for the treatment of osteoporosis.
Osteoarthritis (OA) is a degenerative joint disease characterized by cartilage degeneration and inflammatory responses (Abramoff and Caldera, 2020). Studies had shown that FMN reduces the expression of cartilage-degrading enzymes, such as MMP-1, MMP-3, and MMP-13, in primary rat chondrocytes, effectively counteracting IL-1β-induced catabolism and enhancing chondrocyte viability (Cho et al., 2019). Furthermore, FMN inhibited the inflammatory response by blocking the phosphorylation of related components in the NF-κB/MAPK signaling pathway. It also reduced the levels of cartilage catabolic markers, including MMP-3, MMP-13, and thrombospondin motifs 5 (ADAMTS5), in a concentration-dependent manner, while increasing the level of the cartilage-specific marker COL2A1 and improving bone and joint damage in OA mice (Ni et al., 2023). In addition to targeting chondrocyte degradation, FMN also reduced NO production and PGE2 expression, upregulated the levels of type II collagen and Aggrecan, and thus improved chondrocyte ECM synthesis and inhibited cartilage surface destruction and bone mineral formation in OA rats by modulating the PTEN/AKT/NF-κB signaling pathway, and this change was dose-dependent (Jia et al., 2022).
Atopic dermatitis is a recurrent, chronic, non-infectious inflammatory skin disease characterized by persistent itching. It is the largest component of non-fatal diseases worldwide and mainly occurs in infants and children (Sroka-Tomaszewska and Trzeciak, 2021). Previous studies showed that FMN inhibited the expression of intracellular murine double minute 2 (MDM2), hypoxia-inducible factor 1 alpha (HIF-1α), and NF-κB, while reducing the levels of thymic stromal lymphopoietin (TSLP) and VEGF (Han et al., 2023). This results in decreased ear swelling and thickness in mice with acute ear edema, suggesting that FMN had potential as a treatment for inflammatory skin diseases. On this basis, Yuan et al. (2021) demonstrated that FMN could effectively alleviate AD symptoms in vivo and in vitro experiments. The underlying mechanism appeared to involve the activation of the G protein-coupled estrogen receptor (GPER), which upregulated A20 protein and mRNA expression. This, in turn, affected the ubiquitination of signal transducer and activator of transcription 1 (STAT1) and STAT3, thereby inhibiting TSLP production. Thus, TSLP is identified as a key target of FMN in the treatment of AD.
Psoriasis is a common chronic inflammatory skin disease, which is mainly characterized by distinctive erythematous plaques with white scales (Kamiya et al., 2019). Currently, mild psoriasis is mainly treated by topical therapy with steroids, vitamin D analogs, calcium-modulated phosphatase inhibitors, keratolytic agents, and phototherapy, while moderate and severe psoriasis is mainly treated by systemic therapy (Calabrese et al., 2023). Although these therapies are effective in the short term, they are associated with potential side effects and contraindications, limiting their suitability for long-term use. Therefore, there is an urgent need for affordable, safe, and effective treatments. In vitro experiments found that FMN could significantly inhibit the growth of HaCaT cells, and 20 μM of FMN could minimize the levels of TNF-α and IL-6, which had good anti-inflammatory activity (Xu et al., 2024). This result was confirmed in vivo experiments, and it was found that the interferon (IFN) signaling pathway was inhibited, which could effectively reduce the expression of related inflammatory chemokines, and significantly improve the erythema, scales and thickness of skin lesions in the psoriasis mice model.
Diabetes is a metabolic disease characterized by hyperglycemia and is mainly classified as Type I and Type II, of which Type 1 diabetes mellitus (T1DM) is caused by the destruction of pancreatic β-cells, resulting in an almost complete loss of, or a severe deficiency in, insulin secretion (American Diabetes Association, 2011). Pancreatic cell damage caused by oxidative stress stimulation is one of the key factors promoting the development of T1DM. Based on this, Chen et al. (2024) demonstrated through in vivo and in vitro experiments that FMN binds to Keap1, activating the Keap1/Nrf2 signaling pathway. This promoted the translocation of Nrf2 from the cytoplasm to the nucleus, enhanced the expression of antioxidant proteins HO-1 and NQO1, and prevented excessive ROS production in pancreatic cells. As a result, FMN effectively mitigated alloxan-induced pancreatic β-cell and DNA damage, lowered blood glucose levels, and increased insulin content.
Non-alcoholic fatty liver disease (NAFLD) is one of the most common liver diseases worldwide, characterized by steatosis caused by the accumulation of triglycerides in hepatocytes (Sheka et al., 2020). Non-alcoholic steatohepatitis (NASH) is a progressive form of NAFLD, which can further develop into cirrhosis and hepatocellular carcinoma with the development of time. Liao et al. (2024) established the mice model of NASH using a methionine-choline-deficient diet. Following intervention with FMN, liver function and hepatocyte steatosis were significantly improved. This effect was attributed to FMN’s ability to increase the expression and activity of SIRT1, promote the deacetylation of PGC-1α, and enhance the transcriptional activity of PPARα. These effects led to the upregulation of fatty acid oxidation (FAO) capacity, as well as increased levels of carnitine, acyl-CoA dehydrogenase medium-chain (ACADM), and carnitine palmitoyl transferase 1A (CPT1A), thus promoting liver fatty acid β-oxidation and regulating lipid metabolism. Finally, the steatosis of hepatocytes in NASH mice was improved. Furthermore, if it developed into NASH, FMN promoted the nuclear translocation of transcription factor EB (TFEB) by activating adenosine monophosphate-activated protein kinase (AMPK). This activation enhanced lysosome biogenesis and the fusion of autophagosomes with lysosomes, alleviating autophagic flux blockage and inducing lipid droplet degradation to prevent cellular lipid accumulation (Wang et al., 2019). In addition, FMN could act as a prebiotic to regulate intestinal microbial flora, thereby improving host metabolism and preventing obesity (Naudhani et al., 2021).
Oxidative stress is a disorder of cellular function and environmental homeostasis caused by the accumulation of overproduction of ROS, which can trigger multiple cellular pathways and induce organismal damage (Tang et al., 2022). Nrf is known as the“main regulator”of the antioxidant response and is a key redox-sensitive transcription factor. When Nrf is activated, it will promote the expression of phase II detoxification enzymes and antioxidant enzymes, improve oxidative stress, promote cell survival, and maintain cellular redox homeostasis (Hybertson et al., 2011). Flap necrosis is a common postoperative complication, and hypoxia leads to secondary damage to the flap tissue. By increasing the expression of Nrf2, FMN could activate the expression of downstream target antioxidant/phase II detoxification enzymes to play a transcriptional function, thereby increasing the activity of SOD and GSH-Px. At the same time, the expression of VEGF was increased and angiogenesis was enhanced (Li et al., 2022). It could be seen that FMN exerted a strong antioxidant effect through the Nrf2-driven antioxidant defense system to restore flap tissue damage. In addition, FMN could also inhibit the sustained production of ROS through the Nrf2/HO-1 pathway, alleviate acute kidney injury due to oxidative stress and improve kidney function in rats (Aladaileh et al., 2019). The main pharmacological activity mechanism of FMN is shown in Table 2.

Table 2. The main pharmacological activity mechanism of FMN.
FMN has good therapeutic effects against inflammatory diseases, such as rhinitis, mastitis, and keratitis. Huang et al. (2021) reported that FMN could inhibit IL-13-induced inflammation and mucus formation in JME/CF15 cells by activating the SIRT1/Nrf2 signaling pathway, suggesting that FMN may be a promising drug for the treatment of allergic rhinitis. Xiang et al. (2022) found that FMN could protect the breast by inhibiting the inflammatory response and enhancing the integrity of the blood-milk barrier. Specifically, FMN significantly reduced the production of inflammatory factors TNF-α and IL-1β by inhibiting the NF-kB signaling pathway and concentration-dependently increased the expression of AhR and inhibited the phosphorylation of Src, thus exerting an anti-mastitis effect. Additionally, FMN increased the expression of mammary epithelial tight junction proteins (claudin-3, occludin, and ZO-1), which enhanced the integrity of the blood-milk barrier and provided protection to the mammary gland. In the model of fungal keratitis induced by Aspergillus fumigatus in mice, researchers observed that TSLP and TSLPR were overexpressed in epithelial cells and infiltrating immune cells. After treatment with FMN, the expression of TSLP, TSLPR, and inflammatory factors was reduced, ROS production decreased, and the migration of macrophages and neutrophils was inhibited, alleviating the cornea’s inflammatory response (Feng et al., 2024). In addition, FMN enhanced the migration of HCECs from human corneal cells and promoted corneal epithelial repair. Thus, it could be seen that FMN had the potential to be an inhibitor of TSLP to treat fungal keratitis.
COX-2 is expressed in many types of tumor cells, which regulates cell growth and participates in tumorigenesis and carcinogenesis through mitosis (Cheng et al., 2012). There is increasing evidence suggesting that COX-2 could serve as a potential target to reduce cancer risk (Hashemi Goradel et al., 2019). The continuous program of mitosis is known as the cell cycle, which consists of the G0, G1, S, G2, and M phases. The G1 phase is an important period that affects cell division and development. It is mediated by cyclin-dependent kinases (CDKs) and regulatory cyclin subunits, which determine whether cells further self-renew and develop (Massagué, 2004). Therefore, the G1 phase of the cell cycle can be used as a key target to control the proliferation of tumor cells. Chen et al. (2023) demonstrated that FMN effectively inhibited the proliferation of KYSE170 and KYSE150 cells by significantly reducing the mRNA and protein expression levels of COX-2 and cyclin D1, while inducing G1 phase arrest. The incidence of esophageal cancer in mice treated with FMN was significantly reduced at 18 weeks (0/15 vs. 2/15) and 24 weeks (6/15 vs. 13/15). In addition, FMN also exhibited a strong anti-proliferative effect on MCF-7 and MDA-MB-468 cells, where it significantly increased the proportion of G0/G1 phase cells after treatment (Liu et al., 2024).
EGFR is a tyrosine kinase receptor that is often overexpressed or mutated in patients with non-small cell lung cancer (NSCLC), leading to uncontrolled cell proliferation and tumor formation (Riely et al., 2024). Tyrosine kinase inhibitors, such as gefitinib, erlotinib and osimertinib, have become the first-line treatment for patients with EGFR activation mutations. However, its clinical application is limited due to drug resistance. FMN was found to have the potential to become a new EGFR inhibitor and show a significant inhibitory effect on both osimertinib-sensitive and resistant non-small cell lung cancer cells, which could greatly reduce the generation of drug resistance (Yu et al., 2020). Further in vitro and in vivo studies demonstrated that FMN binds to both WT and mutant EGFR, reducing EGFR kinase activity and inhibiting downstream signaling. This, in turn, activated GSK-3β and decreased the expression of myeloid leukemia sequence 1 (Mcl-1), without causing significant toxicity to the vital organs of mice. These findings highlight the importance of developing targeted therapies for the inhibition of tumor proliferation.
HIF-1 is a heterodimeric protein consisting of HIF-1α protein and HIF-1β protein, which can activate the expression of target genes involved in angiogenesis, such as VEGF (Semenza, 2003). During cancer cell proliferation, tumor cell growth is promoted by increasing VEGF expression, enhancing vascular permeability, and stimulating tumor angiogenesis (Ahmad and Nawaz, 2022). Therefore, VEGF can be used as a target to inhibit tumor proliferation. Zhang et al. (2019) established Balb/c nude mice model of cervical cancer by inoculating HeLa cells and gavaged FMN every 7 days for a month. The results showed that during the drug intervention, the mice in the FMN group had no obvious adverse reactions and were in good condition, while the mice in the cisplatin group had poor appetite, listlessness, and reduced activity. The tumor inhibition rate in the FMN group was comparable to that in the cisplatin group, and the tumor inhibition rate was 50.17%. Further analysis using RT-qPCR and western blotting showed that the expression levels of HIF-1α, VEGF mRNA, and protein in the cervical cancer tissues of mice in the FMN group were significantly lower than those in the positive control group. It could be seen that FMN inhibited the proliferation and growth of cervical cancer cells by inhibiting the expression of HIF-1-α and VEGF. In addition, FMN also had a significant inhibitory effect on the proliferation of bladder cancer cells. Zhou H. et al. (2023) used two bladder cancer cell lines, TM4 and 5,637, to explore the mechanism of action of FMN in the treatment of bladder cancer through transcriptomics. After 24, 48, and 72 h of FMN treatment, the growth of bladder cancer cell lines T24 and 5,637 was inhibited in a dose-dependent manner, without affecting the viability of non-tumor cells. Further analysis revealed that the above phenomenon was attributed to the fact that FMN mediated VEGF expression through the regulation of fibroblast growth factor receptor (FGFR) and its receptor, which in turn impaired tumor angiogenesis and inhibited cancer cell proliferation.
The ERK pathway is a classical and important pathway in the MAPK signaling pathway, which transmits extracellular signals to the nucleus and regulates the expression of related genes, and is highly expressed in aggressive cancer cells (Krueger et al., 2001; Guo et al., 2022), such as nasopharyngeal carcinoma. In addition to the compounds clinically used for the treatment of nasopharyngeal carcinoma, FMN is also considered to be a therapeutically promising compound, which can achieve antiproliferative and invasive effects through effective inhibition of the oncogenic ERK1/2 pathway and the Lamin A/C signaling pathway, thereby modulating the nasopharyngeal carcinoma tumor microenvironment and inhibiting the migration and invasion of cancer cells (Ying et al., 2019).
Apoptosis is a form of programmed cell death that plays a crucial role in normal embryonic development, maintenance of homeostasis in adult tissues, and suppression of carcinogenesis (Willis et al., 2003). Mitochondrial function is closely related to cancer development, and mitochondrial dysfunction, induced by aberrant oncogenic and tumor suppressor signals, alters cellular metabolic pathways, disrupts redox homeostasis, and contributes to resistance to apoptosis (Luo et al., 2020). The Bcl-2 family of proteins, mainly located in the outer membrane of the mitochondrion, is a key factor in regulating cellular death and an upstream regulator of the caspases cascade, which can be disrupted by disrupting the inner membrane of the mitochondrion. It can effectively promote the release of pro-regulatory factors from mitochondria by disrupting the inner mitochondrial membrane and drive apoptosis (Bruckheimer et al., 1998). FMN, as a candidate anticancer drug, could release cytochrome C (cyto C) directly through the mitochondrial pathway and activate the cascade reaction of caspase-9, caspase-3 and PARP, which ultimately lead to FaDu cell death (Oh et al., 2020). In addition, both in vivo and in vitro experiments demonstrated that FMN upregulated the levels of LDH, Bax, Apaf-1, and caspase-3 in a dose-dependent manner, and downregulated the levels of ERα, p-PI3KCATyr317, p-AKTSer473 (Hu et al., 2019). Based on these findings, we concluded that FMN induced osteosarcoma cell death via a mitochondria-dependent signaling pathway.
Mitochondrial autophagy is a critical cellular process and a key regulatory mechanism for the removal of damaged mitochondria, essential for maintaining cellular homeostasis (Qiu et al., 2021). In addition, more and more data suggest that mitochondrial autophagy plays an important role in the occurrence, development, and treatment of many tumors. Li et al. (2023) collected clinical samples from triple negative breast cancer (TNBC) patients and explored the effect of FMN on TNBC. The results demonstrated that FMN inhibited mitosis by inactivating the BACH1/p53 signaling pathway, promoted the release of cyto C and cysteine asparaginase, and mediated mitochondrial autophagy, resulting in mitochondrial dysfunction. In this process, FMN delivered ROS to mitochondria to release cyto C and activated caspase-3 and caspase-9 cascade reactions to induce apoptosis in MCF7 cells. These findings suggest that FMN may exert its anti-tumor function by hindering mitochondrial function in breast cancer cells.
MicroRNAs are small non-coding regulatory RNAs ranging from 17 to 25 nucleotides. In the past few years, research on microRNA-based therapies for cancer has begun to emerge. The expression of microRNAs can reflect either carcinogenic or tumor-suppressive effects, making them useful for tumor diagnosis and prognosis (Lee and Dutta, 2009). TNBC is the most refractory breast cancer type and the second leading cause of cancer-related deaths worldwide (Li S. et al., 2022). Currently, therapeutic options for TNBC are limited to surgery and chemotherapy. However, because it is a highly heterogeneous cancer with specific mutational signaling, targeted therapy is a promising option for the treatment of TNBC. FMN demonstrated targeted potential in the treatment of TNBC. By up-regulating the expression of miR-195 and miR-545, it could inhibit the expression of CDK4 and Raf-1 in the development of TNBC, thereby inhibiting the proliferation, migration and invasion of MDA-MB-231 and BT-549 cells (Wu et al., 2021). However, the experiment has not been validated by in vivo experiments, so it lacks the comprehensiveness and reliability of the experimental results, and the follow-up needs to be supplemented with in vivo experiments to verify the actual effect and safety in organisms. Li et al. (2020) found that FMN could inhibit the viability of osteosarcoma MG-63 cells and induce apoptosis. It was found by dual luciferase assay, western blot and qRT-PCR that FMN targetedly increased the expression of PTEN by reducing the expression of miR-214-3p. In addition, FMN also showed anti-gastric cancer activity (Wang and Zhao, 2021). In vitro experiments, FMN inhibited the proliferation of gastric cancer cells SGC-7901 and MGC-803 in a dose-dependent manner and significantly reduced cell colony formation, which was related to the decreased expression of miR-542-5p. The anti-gastric cancer potential of FMN in vivo was confirmed using an SGC-7901 xenograft model. Compared with the control group, the xenograft tumor volume of the FMN group was smaller, the tumor weight was lighter, and the expression of miR-542-5p was decreased. The results were consistent with in vitro experiments. In summary, natural metabolites combined with molecular targets have great prospects in treating cancer.
Programmed death 1 (PD-1) is a protein expressed on T-cells. When it binds to the PD-L1 receptor on immune cells, it disrupts the effector function of lymphocytes and inhibits T-cell proliferation, thereby facilitating tumor escape from immune system attack (Safi et al., 2021). Liu et al. (2024), Wang et al. (2022) found that FMN could inhibit the occurrence of breast cancer and cervical cancer by interfering with the activation of PD-L1, but their mechanisms were different. For breast cancer, FMN mainly interfered with PD-L1 activation by inhibiting the STING-NF-κB signaling pathway. For cervical cancer, FMN mainly inhibited MYC protein expression and STAT3 activation through RAS/ERK and JAK1/STAT3 signaling pathways to inhibit PD-L1 synthesis. At the same time, it also promoted lysosomal biogenesis and induced PD-L1 degradation. Consequently, FMN could block the binding of PD-1 to PD-L1, and restore T cell activity, thereby enhancing the ability of the immune system to attack tumors. These findings suggest that FMN has the potential to serve as a PD-1/PD-L1 inhibitor for clinical use. The anti-tumor mechanism of FMN is shown in Figure 4.

Figure 4. Anti-cancer mechanism of FMN.
FMN has been reported to possess other pharmacological activities, such as antibacterial, antiviral, and antiallergic. Feng et al. (2024) found that FMN inhibited the growth of aspergillus fumigatus at concentrations ranging from 8–512 μg/mL, and impeded the formation of biofilm at a concentration range of 256 μg/mL. In addition, FMN also showed good inhibitory effects against streptococcus suis (Wang et al., 2020). Cui et al. (2021) reported that FMN had anti-cat calicivirus effect. When combined with icariin, it could play a synergistic effect. Zhou et al. (2023) suggested that FMN may be a novel anti-allergic drug with great potential to alleviate pseudoallergic reactions. In models of C48/80-stimulated BMMCs and RBL-2H3 cells, FMN inhibited immunoglobulin E-independent degranulation of dependent mast cells by reducing the release of β-hexose and histamine and the expression of inflammatory factors. Moreover, in the 2,4-dinitrobenzene-induced AD mice model, FMN reduced the local allergic reaction of AD model mice by increasing the expression of filaggrin and loricrine. Liu et al. (2021) used CKD rats and TNF-α-induced C2C12 myotubes to establish in vivo and in vitro muscle atrophy models, and found that FMN improved muscle atrophy caused by chronic renal dysfunction. This was related to FMN increasing the phosphorylation of PI3K/Akt/FoxO3 a pathway and the level of myogenic differentiation factor D (MyoD) and myogenin, thereby improving the proliferation and differentiation dysfunction of satellite cells. In the mice model of caerulein-induced pancreatitis, the researchers used FMN for continuous intragastric administration for 7 days. The results showed that FMN significantly improved pancreatic edema in mice by activating the Keap1/Nrf2 signaling pathway and reducing the levels of ROS, NLRP3 and inflammatory factors (Yang et al., 2023). In addition, FMN could also upregulate the expression of colonic tight junction proteins, restore intestinal mucosal barrier function, improve the intestinal flora environment, and provide a new solution for the treatment of pancreatitis. Park et al. (2023) believed that FMN could be used as a non-hormone treatment option for endometriosis. Through in vivo and in vitro experiments, FMN was found to achieve therapeutic effects on endometriosis by decreasing the expression of p27, p-STAT3, and p-ERK within the endometrium.
For many cancers, the combined treatment of drugs is superior to the effect of single treatment. This is due to the uniqueness of each drug treatment, when combined, synergistic effects can be exerted (Plana et al., 2022). The mechanisms of combination therapy for FMN are shown in Table 3.
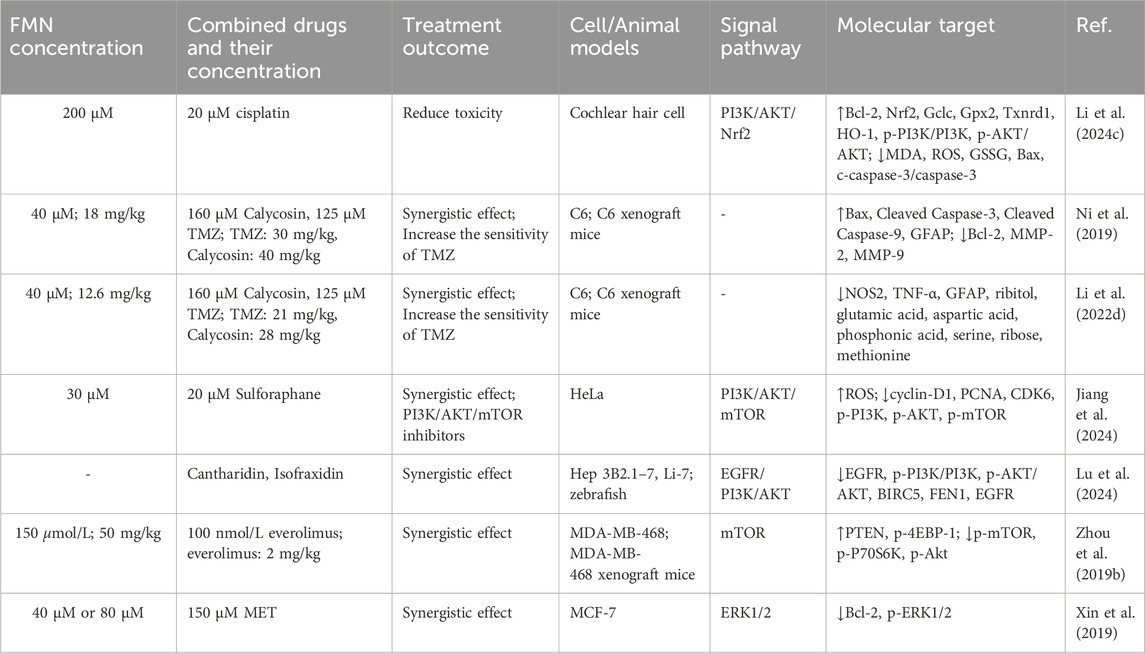
Table 3. The mechanism of combination therapy.
Cisplatin is a common chemotherapeutic drug in the clinic, and ototoxicity is an unavoidable side effect during its use, which can damage the inner ear and impair both hearing and balance functions in patients (Wang et al., 2023). Currently, no effective treatment exists to repair the hair cell damage induced by cisplatin. Based on this, Li et al. (2024) used cisplatin to culture cochlear hair cells in vitro and treated them with FMN, and found that FMN effectively reduced the accumulation of ROS and mitochondrial damage in hair cells by activating the PI3K/AKT-Nrf2 signaling pathway, restored the balance of GSH/GSSG. Therefore, FMN was a potential therapeutic agent for cisplatin-induced ototoxicity.
Glioma is a common malignant tumor of the brain, and temozolomide (TMZ) is currently the first-line drug used in clinical practice for postoperative or non-surgical chemotherapy of gliomas, which can easily penetrate the lesion site through the blood-brain barrier, but the acquired and intrinsic resistance of TMZ limits its application (Xu et al., 2020). In addition to using FMN alone, the combination of FMN and mullein isoflavones may enhance the antitumor effects of TMZ. In vivo and in vitro experiments showed that the IC50 of TMZ was significantly reduced from 1000 μM to 125 μM when the low effective concentration of calycosin (160 μM) and FMN (40 μM) were combined with TMZ. The combination of the three greatly improved the sensitivity of TMZ, significantly inhibited the growth and migration of C6 cells, and the toxicity of the drugs to the heart, liver, spleen, lung and kidney of mice was small (Ni et al., 2019). In addition, based on metabolomics and molecular biology studies, nitric oxide synthase 2 (NOS2) was a key target for FMN and calycosin to regulate TMZ in the treatment of glioma. FMN and calycosin reduced the secretion of TNF-α in the tumor area by inhibiting the expression of NOS2, and regulated amino acid metabolism, especially glutamic acid, ornithine, aspartic acid, proline and arginine, thereby increasing the cytotoxicity of TMZ on malignant glioma, inhibiting tumor growth and infiltration in C6 glioma rats, and reducing cell aggregation and GFAP expression in C6 glioma area (Li et al., 2022). The above studies showed that cancer cells acquire drug resistance because multi-gene abnormalities in tumor cells allow them to evade the effects of these drugs when patients receive treatment for a single pathway. Therefore, combination therapy had better control effects on multiple targets and a lower risk of drug resistance, which had great application prospects for treating cancer.
The PI3K/Akt/mTOR signaling pathway plays a critical role in the apoptosis and proliferation of various cancer cells. The activation of Akt/mTOR can initiate a signaling cascade that regulates apoptosis, survival, growth and proliferation, thus promoting tumorigenesis (Li et al., 2017). The combination of FMN (30 μM) and sulforaphane (20 μM) exhibited a significant synergistic effect (CI = 0.57), and inhibited the expression of cyclin-D1, PCNA and CDK6 in HeLa cells by down-regulating the PI3K/Akt/mTOR signaling pathway, reducing the proportion of cells in G0/G1 phase, hindering the entry of cells into S phase, and leading to the death of HeLa cells (Jiang et al., 2024). In addition, the combination of the two could promote the generation of ROS in HeLa cells, leading to oxidative damage, which was also one of the causes of HeLa cell death. It could be seen that the combination of FMN and sulforaphane could be used as an inhibitor of PI3K/AKT/mTOR, which had important clinical significance in the treatment of cervical cancer. Aidi injection is a widely used Chinese patent medicine for the treatment of hepatocellular carcinoma. Lu et al. (2024) used Hep3B2.1-7, Li-7 cells and zebrafish to explore the anti-hepatocarcinoma mechanism of Aidi injection. The results showed that the strong anti-hepatocarcinoma effect of Aidi injection was the result of the combined treatment of cantharidin, FMN and isofraxidin. Meta-analysis showed that BIRC5, FEN1 and EGFR could be the key targets of Aidi injection against hepatocellular carcinoma, and the three chemical components exerted synergistic inhibitory effects through their targets, respectively, which revealed the mechanism of anti-hepatocellular carcinoma of Aidi injection, and also provided a typical example for the pharmacological evaluation of other proprietary Chinese medicines.
In another report, FMN could enhance the anti-tumor effect of everolimus on MDA-MB-468 cells, increasing the apoptosis rate by 27.9%. There was no significant loss of appetite in mice with the MDA-MB-468 cell xenograft model, and the tumor volume was reduced to half (Zhou et al., 2019). It could be seen that FMN combined with everolimus can safely inhibit tumor growth. RT-qPCR and western blot analysis revealed that both FMN and everolimus inhibited the expression of p-mTOR and p-P70s6k and increased the expression of PTEN and p-4EBP-1. However, FMN alone inhibited p-Akt expression, whereas everolimus had no inhibitory effect on this. When Akt siRNA was used to silence Akt, everolimus did not affect the expression of mTOR, and FMN still reversed the expression of p-4EBP-1 and p-P70s6k. It could be seen that the synergistic effect of FMN on everolimus plays a role by inhibiting Akt activity and inhibiting the mTOR signaling pathway. In addition, Xin et al. (2019) also found that FMN-assisted metformin (MET) in the treatment of breast cancer, which had great application prospects. The combination of 40 μM FMN and 150 μM MET or the combination of 80 μM FMN and 150 μM MET could reduce the proliferation rate of MCF-7 cells. Compared with the monotherapy, the levels of Bcl-2 and p-ERK1/2 were greatly reduced, and the apoptosis rate was increased.
FMN had been reported to be cytotoxic to human-immortalized epidermal cells HaCaT with an IC50 of 40.64 μM (Xu et al., 2024). Yu et al. (2023) treated BMMs with FMN at concentrations up to 20 μM without any significant cytotoxicity. However, Ni et al. (2023) further found that FMN had a greater effect on the viability of BMMs when the concentration reached 40 μM or higher, with an IC50 of 29.61 μM. Cho et al. (2019) evaluated the effect of FMN on the viability of rat primary chondrocytes using the MTT assay, and the results showed that, even at a concentration of 100 μM, FMN did not affect rat primary chondrocyte viability. In addition, FMN had no toxic effect on the growth of normal ovarian cell lines (Zhang et al., 2018). In another report, Pingale and Gupta (2023) carried out acute toxicity tests and subacute toxicity tests of FMN by intraperitoneal injection in mice. The results showed that the acute dose of FMN was 300 mg/kg, the LD50 was 103.6 mg/kg, and the NOAEL was 50 mg/kg. During the subacute toxicity experiment, the body weight, food intake, water intake and behavior of the animals did not change, and the organs did not have any toxic effects and pathological damage. Therefore, it was proved that FMN was safe and non-toxic and could be used for pharmacological and therapeutic purposes. In summary, FMN may be a relatively safe natural compound component. However, there is still a lack of subchronic toxicity, genotoxicity and reproductive toxicity experiments, and the follow-up should be expanded to carry out clinical trials of FMN.
FMN has low water solubility, which limits its application in the pharmaceutical industry. To solve the water solubility problem of FMN, Zhao et al. (2021) succinylated FMN using Bacillus amyloliquefaciens FJ18 to form the compound FMN-7-O-β-D (6″-O-succinyl)-D-glucoside (FMP), which compared to FMN, the water solubility was increased more than 106-fold. Moreover, in the mice model of isoprenaline-induced acute ischemia, the compound significantly attenuated cell membrane damage during myocardial ischemia by scavenging free radicals in serum, increasing CAT and SOD activities, and decreasing lactate dehydrogenase (LDH) activity. In another study, Yan et al. (2024) prepared seven derivatives of FMN from FMN. Among them, 9-butyl-3-(4-methoxyphenyl)-9,10-dihydro-4H,8H-chromeno[8,7-e][1,3]oxazin-4-one(2) and 9-(furan-3-ylmethyl)-3-(4-methoxyphenyl)-9,10-dihydro-4H,8H-chromeno-[8,7-e][1,3]oxazin-4-one possessed the ability to promote bone formation and inhibit osteoclastogenesis, which was superior to the effect of ipliflavone, suggesting that this compound could be a patentable candidate for anti-osteoporosis. In addition, Zuo et al. (2022) used structural modification to replace the methoxy group at position 40 of FMN with the phenoxy group to obtain a new compound that greatly improved the antihypertensive effect of FMN. Kim et al. (2019) utilized microbial transformation technology to synthesize a novel compound, FMN 7-O-phosphate. Compared with FMN, FMP showed lower cytotoxicity in RAW264.7 cells and significantly improved cell viability. At the same time, it inhibited the production of pro-inflammatory factors in a dose-dependent manner and decreased the mRNA expression of inducible iNOS and COX-2, which could be used as a promising candidate compound for new anti-inflammatory drugs.
FMN can act as a Bax agonist to exert anti-cancer effects. To improve its bioavailability, Jia et al. (2023) modified its structure and obtained a new compound 7-((1-(4-fluorobenzyl)-1H-1,2,3-triazol-4-yl)methoxy)-3-(4-methoxyphenyl)-4H-chromen-4-one. The compound could significantly inhibit the growth of the A549 cell line, downregulate the expression of Bcl-2 in cells, upregulate the expression of Bax, and promote apoptosis. Its efficacy was 40 times and 6.94 times greater than that of FMN and adriamycin, respectively, indicating that the derivative had strong potential as an anti-cancer lead compound. Based on the natural FMN, Yao et al. (2019) designed and synthesized a FMN-coumarin hybrid product by molecular hybridization strategy. Compared with FMN, the hybrid product could significantly inhibit the growth and migration of gastric cancer SGC7901 cells through the wnt/β-Catenin and AKT/mTOR pathways, and could be used as an inhibitor of SIRT1 to inhibit its expression, thereby inducing cancer cell death. In vivo, the hybrid effectively inhibited the growth of SGC7901 xenograft tumors in nude mice without significantly affecting their body weight. Similarly, Yang et al. (2019) inserted the FMN fragment into the C4 hydroxyl position of podophyllotoxin to form a new hybrid product, which had a stronger ability to resist A549 tumor growth than podophyllotoxin. This was because FMN could be inserted into the αβ-interface to form hydrogen bonds, destroy the cytoskeleton of tubulin, and enhance anti-tumor ability. The chemical structure of FMN derivatives is shown in Figure 5.
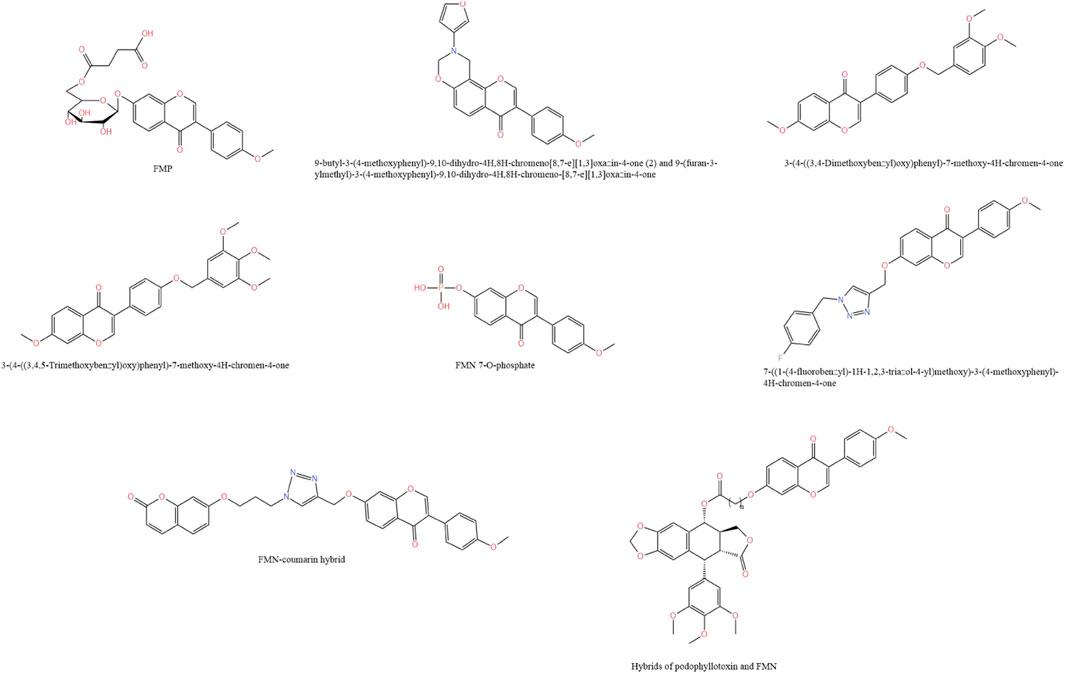
Figure 5. Chemical structure of FMN derivatives.
Drug delivery system is the use of carriers or technologies to effectively and safely deliver drugs to specific targets in the body, which has the characteristics of optimizing the therapeutic effect of drugs, high solubility and long retention time in biological systems (Prakash, 2023). Since FMN is characterized by low water solubility and low bioavailability, limiting its application in various areas. Therefore, research based on nano-delivery systems has emerged in recent years. The following text should be added: The FMN delivery systems is shown in Figure 6.
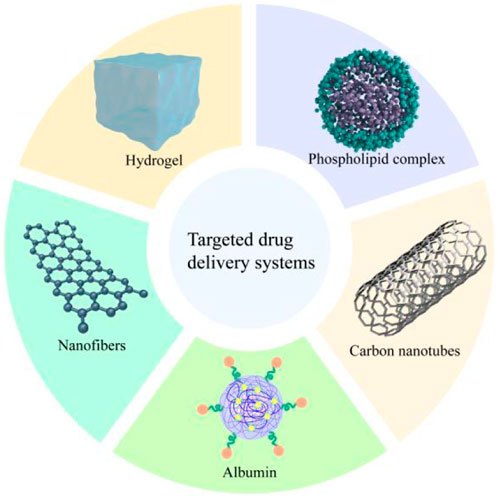
Figure 6. FMN’s delivery system.
Carbon nanotubes (CNs) are small, flexible, robust, inert, and electrically conductive materials that have been investigated for treating spinal cord injuries due to their electrical properties and nano-size. However, CNs have low hydrophilicity and poor dispersibility, which tend to lead to aggregation, so it is usually necessary to functionalize the CNs to avoid its aggregation (Imani et al., 2016). de Vasconcelos et al. (2020) used FMN to functionalize CNs, which not only improved the stability of CNs, but also further stimulated neuronal growth and cell membrane fusion through FMN, and even reduced inflammatory response. On this basis, in order to enhance the bio-interface with nerve cells, gelatin methacryloyl hydrogel was used as a scaffold, which could retain the nanocomposite of multi-walled carbon nanotubes and FMN at the inflammation site, and was directly used for the repair of neural tissues in the diseased site.
Ouyang et al. (2023) used bovine serum albumin (BSA) as raw material, and nanoprecipitation with 20:1 as the optimal ratio of albumin to FMN to prepare FMN-containing nanoparticles FMN@BSA. The nanoparticles improved the bioavailability of FMN by promoting the preferential accumulation of albumin’s affinity for SPARC protein overexpressed in lung inflammation and pulmonary fibrosis, and alleviated bleomycin-induced lung inflammation and interstitial deposition in mice by blocking NLRP3 inflammasome to slow down the pyroptosis process of macrophages. It could be seen that the nanoparticles could be used as a macrophage pyroptosis inhibitor. Compared with the commonly used first-line therapeutic drug pirfenidone, FMN@BSA was a natural product as the active ingredient, which had lower toxicity and stronger clinical transferability.
It has been previously shown that β-cyclodextrin can improve the solubility of FMN (Bhardwaj and Purohit, 2023). On this basis, Wang et al. (2021), in order to improve the oral bioavailability of FMN, prepared instantly soluble nanofibers containing FMN/methyl-β-cyclodextrin (FMN/Me-β-CD) inclusion complex, which increased the solubility of FMN by about 50-fold in 20 mM Me-β-Cd aqueous solution. After that, polyvinyl alcohol nanofiber webs containing FMN/Me-β-CD inclusion complex was prepared by electrostatic spinning method, which could be dissolved in artificial saliva in about 2 s. The rapid dissolution of FMN in artificial saliva was greatly improved, which would be beneficial to the application of FMN in oral diseases. In addition, Agarwal et al. (2024) had also made efforts to improve the oral bioavailability of FMN. The researchers used phospholipid (PL) as a carrier to prepare FMN phospholipid complex (FNT-PC) using the solvent evaporation method and characterized it. The results showed that FNT-PC exhibited amorphous geometry, and the water solubility was 4 times higher than FMN. In addition, in vivo pharmacokinetic studies in rats by gavage showed that FNT-pc also reduced first-pass metabolism, increased gastric mucosal permeability, and significantly improved oral bioavailability.
As mentioned earlier, FMN has excellent antitumor activity, and to further enhance its clinical application value and therapeutic effect, the use of the nano-delivery system to effectively improve the bioavailability and targeting of FMN is a promising strategy. It had been reported that some researchers used epidermal growth factor receptor-targeting peptide (GE11)-modified nanoparticles (GE-NPs) loaded with paclitaxel and hyaluronic acid (HA)-modified nanoparticles (HA-NPs) encapsulated with FMN, and self-assembled both of them to construct binary nanoparticles modified by GE11 and HA dual ligands (HAGE-DTX/FMN-NPs). The binary nanoparticles showed a stronger inhibitory effect on PC3 cells with a cellular uptake rate of 59.6% compared with the single ligand-modified nanoparticles and free drug. PC3 cells were injected into the right side of Balb/c nude mice to prepare cancer xenografts by subcutaneous injection and treated by tail vein injection, and it was found that HAGE-DTX/FMN-NPs were highly aggregated at the tumor site, whereas the single ligand-modified nanoparticles and the free drug had more distribution of the drug in the liver and kidneys, which showed that the binary nanoparticles modified with dual ligands have better targeting and bioavailability and can avoid unwanted side effects (Dong et al., 2022).
Cao et al. (2023) used microfluidic technology to prepare loaded FMN microspheres, which could encapsulate and release the drug in a controlled manner. In vitro experiments showed that the microspheres inhibited the proliferation, migration, and apoptosis of A549 cells better than FMN, and had good anti-tumor selectivity and low toxicity to normal cells. In addition, the microspheres could induce intracellular ROS production, leading to oxidative damage and better antitumor effects.
In recent years, more and more attention has been paid to developing natural metabolite, mainly because they are derived from plants with low toxicity and high efficiency. FMN is one of the important discoveries of novel natural metabolite with considerable medicinal value. It is widely found in plants and daily diets, and is also detected in some chinese patent medicines, such as Qiliqiangxin capsule (Zhang et al., 2018), Zhenqi Fuzheng capsules (Liu et al., 2015), Guanjiekang (Wu et al., 2016) and Naoxintong capsule. This paper reviews the source, chemical properties, pharmacological activities and molecular mechanisms, co-administration, toxicity, derivatives and drug delivery system of FMN, which can provide a basis for in-depth research and clinical application of FMN.
FMN is involved in various disease processes, such as neurological disorders, organ damage and cancer, by regulating various factors, enzyme activities and signaling pathways, and it is evident that FMN has multiple pharmacodynamic effects. However, the current study still has certain flaws and limitations. First, studies on FMN have mainly focused on cellular and animal models, and the multiple pharmacological mechanisms have not been fully clarified. Although the treatment of the nervous system, organ injury and cancer is outstanding, it is still in the preliminary stage, without in-depth study, and lacks large-scale and long-term clinical trials to verify its safety and effectiveness. And the lack of FMN metabolic process, can not fully understand the drug circulation in the body. Therefore, in the future, in addition to the use of traditional in vivo and in vitro experimental analysis, other research methods should be strengthened, such as metabolomics, proteomics and genomics analysis, in order to better study the absorption, distribution and metabolic process of FMN, and how to act on the target to play a therapeutic effect. Through these studies, we can better understand the regulatory role and overall effect of FMN at the cellular molecular level. Second, to realize the maximum potential of its clinical application, further exploration of its quantitative relationship, side effects, and toxicity is needed, and this information is critical for assessing its safety and feasibility in clinical application. Finally, Although toxicity studies have been conducted on FMN, most of the tests have been at the cellular level, and more comprehensive toxicity tests such as subchronic toxicity, genotoxicity and reproductive toxicity are lacking. It is recommended to use different animal models to validate the toxicity results and extend the experimental period to obtain more data. Finally, systematic clinical trials, including randomized controlled trials, long-term efficacy observation and adverse reaction evaluation, are needed to fully understand the actual effect and safety of FMN in different diseases.
As previously mentioned, FMN has the characteristics of poor water solubility, low bioavailability and insufficient targeting. To solve those problems, structural modification and nano-delivery systems can be used as a promising solution. Numerous scholars have utilized nano-delivery systems to modulate the particle size, surface properties and carrier materials of drugs, which can significantly improve the bioavailability and targeting of FMN, and show good application prospects. At present, although studies have confirmed that the nano-delivery system has a significant improvement on the shortcomings of FMN, there is a lack of long-term safety and toxicological evaluation, and further research is needed in the future to ensure its reliability and safety in clinical applications. Meanwhile, the design of the delivery system should be further optimized to achieve higher drug loading capacity and more precise targeted release. In addition to the use of drug delivery systems alone, FMN can be combined with other treatments, such as immunotherapy, to enhance the therapeutic effect and improve the prognosis of patients; gene editing technology can be used to solve the disease directly from the root cause by precisely modifying the disease-causing genes. In the future, with the gradual enrichment of knowledge, the pharmacological activity of FMN and its molecular mechanism will be further elucidated, and the study of its structural modification and delivery system will be further optimized and innovated, which will have a new impact on the future pharmaceutical industry.
MJ: Writing–original draft. LW: Writing–review and editing. JW: Software, Writing–review and editing. YuS: Software, Writing–review and editing. LG: Data curation, Writing–review and editing. FZ: Data curation, Writing–review and editing. QG: Formal Analysis, Writing–review and editing. YM: Formal Analysis, Writing–review and editing. YoS: Supervision, Writing–review and editing. YL: Supervision, Writing–review and editing. GJ: Writing–review and editing. PC: Writing–review and editing. RY: Funding acquisition, Writing–review and editing.
The author(s) declare that financial support was received for the research, authorship, and/or publication of this article. The authors gratefully acknowledge the financial support from the Fifth Batch of National Clinical Talents Training Program in Traditional Chinese Medicine by the State Administration of Traditional Chinese Medicine (SATCM) (SATCM Human Resources Education Letter (2022) No. 1), the Qilu Biancang Traditional Chinese Medicine talent training project (Lu health Letter No. (2024) 78), and the Nanjing University Of Chinese Medicine Integrated Traditional Chinese and Western Medicine Discipline Excellent Innovative Talent Cultivation Project (035062005004-28).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declare that no Generative AI was used in the creation of this manuscript.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Abramoff, B., and Caldera, F. E. (2020). Osteoarthritis: pathology, diagnosis, and treatment options. Med. Clin. N. Am. 104 (2), 293–311. doi:10.1016/j.mcna.2019.10.007
Agarwal, A., Wahajuddin, M., Chaturvedi, S., Singh, S. K., Rashid, M., Garg, R., et al. (2024). Formulation and characterization of phytosomes as drug delivery system of formononetin: an effective anti-osteoporotic agent. Curr. Drug Deliv. 21 (2), 261–270. doi:10.2174/1567201820666230124114906
Ahmad, A., and Nawaz, M. I. (2022). Molecular mechanism of VEGF and its role in pathological angiogenesis. J. Cell. Biochem. 123 (12), 1938–1965. doi:10.1002/jcb.30344
Aladaileh, S. H., Hussein, O. E., Abukhalil, M. H., Saghir, S. A. M., Bin-Jumah, M., Alfwuaires, M. A., et al. (2019). Formononetin upregulates Nrf2/HO-1 signaling and prevents oxidative stress, inflammation, and kidney injury in methotrexate-induced rats. Antioxidants (Basel) 8 (10), 430. doi:10.3390/antiox8100430
Althunibat, O. Y., Abukhalil, M. H., Aladaileh, S. H., Qaralleh, H., Al-Amarat, W., Alfwuaires, M. A., et al. (2022). Formononetin ameliorates renal dysfunction, oxidative stress, inflammation, and apoptosis and upregulates Nrf2/HO-1 signaling in a rat model of gentamicin-induced nephrotoxicity. Front. Pharmacol. 13, 916732. doi:10.3389/fphar.2022.916732
Alves, R. C., Almeida, I. M., Casal, S., and Oliveira, M. B. (2010). Isoflavones in coffee: influence of species, roast degree, and brewing method. J. Agric. Food Chem. 58 (5), 3002–3007. doi:10.1021/jf9039205
Aly, S. H., Elissawy, A. M., Fayez, A. M., Eldahshan, O. A., Elshanawany, M. A., and Singab, A. N. B. (2020). Neuroprotective effects of Sophora secundiflora, Sophora tomentosa leaves and formononetin on scopolamine-induced dementia. Nat. Prod. Res. 35 (24), 5848–5852. doi:10.1080/14786419.2020.1795853
American Diabetes Association (2011). Diagnosis and classification of diabetes mellitus. Diabetes Care 34 (Suppl. 1), S62–S69. doi:10.2337/dc11-S062
Andersen, C., Nielsen, T. S., Purup, S., Kristensen, T., Eriksen, J., Søegaard, K., et al. (2009). Phyto-oestrogens in herbage and milk from cows grazing white clover, red clover, lucerne or chicory-rich pastures. Animal 3 (8), 1189–1195. doi:10.1017/s1751731109004613
Anjum, A., Yazid, M. D., Fauzi Daud, M., Idris, J., Ng, A. M. H., Selvi Naicker, A., et al. (2020). Spinal cord injury: pathophysiology, multimolecular interactions, and underlying recovery mechanisms. Int. J. Mol. Sci. 21 (20), 7533. doi:10.3390/ijms21207533
Banes-Berceli, A. K., Ketsawatsomkron, P., Ogbi, S., Patel, B., Pollock, D. M., and Marrero, M. B. (2007). Angiotensin II and endothelin-1 augment the vascular complications of diabetes via JAK2 activation. Am. J. Physiol. Heart Circ. Physiol. 293 (2), H1291–H1299. doi:10.1152/ajpheart.00181.2007
Bayır, H., Dixon, S. J., Tyurina, Y. Y., Kellum, J. A., and Kagan, V. E. (2023). Ferroptotic mechanisms and therapeutic targeting of iron metabolism and lipid peroxidation in the kidney. Nat. Rev. Nephrol. 19 (5), 315–336. doi:10.1038/s41581-023-00689-x
Bhardwaj, V. K., and Purohit, R. (2023). A comparative study on inclusion complex formation between formononetin and β-cyclodextrin derivatives through multiscale classical and umbrella sampling simulations. Carbohydr. Polym. 310, 120729. doi:10.1016/j.carbpol.2023.120729
Billowria, K., Ali, R., Rangra, N. K., Kumar, R., and Chawla, P. A. (2024). Bioactive flavonoids: a comprehensive review on pharmacokinetics and analytical aspects. Crit. Rev. Anal. Chem. 54 (5), 1002–1016. doi:10.1080/10408347.2022.2105641
Bruckheimer, E. M., Cho, S. H., Sarkiss, M., Herrmann, J., and McDonnell, T. J. (1998). The Bcl-2 gene family and apoptosis. Adv. Biochem. Eng. Biotechnol. 62, 75–105. doi:10.1007/BFb0102306
Cai, C., Xiang, Y., Wu, Y., Zhu, N., Zhao, H., Xu, J., et al. (2019). Formononetin attenuates monocrotaline-induced pulmonary arterial hypertension via inhibiting pulmonary vascular remodeling in rats. Mol. Med. Rep. 20 (6), 4984–4992. doi:10.3892/mmr.2019.10781
Calabrese, L., Malvaso, D., Antonelli, F., Mannino, M., Peris, K., and Chiricozzi, A. (2023). Investigational systemic drugs for moderate to severe plaque psoriasis: what's new? Expert Opin. Investig. Drugs 32 (3), 229–243. doi:10.1080/13543784.2023.2184684
Cannistrà, M., Ruggiero, M., Zullo, A., Gallelli, G., Serafini, S., Maria, M., et al. (2016). Hepatic ischemia reperfusion injury: a systematic review of literature and the role of current drugs and biomarkers. Int. J. Surg. 33 (Suppl. 1), S57–S70. doi:10.1016/j.ijsu.2016.05.050
Cao, X., Li, Q., Li, X., Liu, Q., Liu, K., Deng, T., et al. (2023). Enhancing anticancer efficacy of formononetin microspheres via microfluidic fabrication. AAPS PharmSciTech 24 (8), 241. doi:10.1208/s12249-023-02691-9
Chen, H., Lou, Y., Lin, S., Tan, X., Zheng, Y., Yu, H., et al. (2024). Formononetin, a bioactive isoflavonoid constituent from Astragalus membranaceus (Fisch.) Bunge, ameliorates type 1 diabetes mellitus via activation of Keap1/Nrf2 signaling pathway: an integrated study supported by network pharmacology and experimental validation. J. Ethnopharmacol. 322, 117576. doi:10.1016/j.jep.2023.117576
Chen, L., Xing, D., Guo, L. R., Jin, J., and Li, S. (2023). Formononetin, an active component of Astragalus membranaceus, inhibits the pathogenesis and progression of esophageal cancer through the COX-2/cyclin D1 Axis. Clin. Lab. 69 (3). doi:10.7754/Clin.Lab.2022.220403
Chen, Y., Wei, D., Zhao, J., Xu, X., and Chen, J. (2021). Reduction of hyperoxic acute lung injury in mice by Formononetin. PLoS One 16 (1), e0245050. doi:10.1371/journal.pone.0245050
Cheng, C. W., Liu, Y. F., Yu, J. C., Wang, H. W., Ding, S. L., Hsiung, C. N., et al. (2012). Prognostic significance of cyclin D1, β-catenin, and MTA1 in patients with invasive ductal carcinoma of the breast. Ann. Surg. Oncol. 19 (13), 4129–4139. doi:10.1245/s10434-012-2541-x
Cho, I. A., Kim, T. H., Lim, H., Park, J. H., Kang, K. R., Lee, S. Y., et al. (2019). Formononetin antagonizes the interleukin-1β-induced catabolic effects through suppressing inflammation in primary rat chondrocytes. Inflammation 42 (4), 1426–1440. doi:10.1007/s10753-019-01005-1
Cui, T., Tang, S., Liu, C., Li, Z., Zhu, Q., You, J., et al. (2018). Three new isoflavones from the Pueraria Montana var. lobata (Willd.) and their bioactivities. Nat. Prod. Res. 32 (23), 2817–2824. doi:10.1080/14786419.2017.1385008
Cui, X., Guo, J., Lai, C. S., Pan, M. H., Ma, Z., Guo, S., et al. (2017). Analysis of bioactive constituents from the leaves of Amorpha fruticosa L. J. Food Drug Anal. 25 (4), 992–999. doi:10.1016/j.jfda.2016.10.006
Cui, Z., Wang, Q., Li, D., Zhao, S., Zhang, Q., Tan, Y., et al. (2021). Icariin, formononetin and caffeic acid phenethyl ester inhibit feline calicivirus replication in vitro. Arch. Virol. 166 (9), 2443–2450. doi:10.1007/s00705-021-05107-w
Cuong, N. M., Son, N. T., Nhan, N. T., Fukuyama, Y., Ahmed, A., Saponara, S., et al. (2022). Vietnamese dalbergia tonkinensis: a promising source of mono- and bifunctional vasodilators. Molecules 27 (14), 4505. doi:10.3390/molecules27144505
Cuzzocrea, S., and Wang, Z. Q. (2005). Role of poly(ADP-ribose) glycohydrolase (PARG) in shock, ischemia and reperfusion. Pharmacol. Res. 52 (1), 100–108. doi:10.1016/j.phrs.2005.02.009
Dan Hu, Q., Wang, H. L., Liu, J., He, T., Tan, R. Z., Zhang, Q., et al. (2023). Btg2 promotes focal segmental glomerulosclerosis via smad3-dependent podocyte-mesenchymal transition. Adv. Sci. (Weinh) 10 (32), e2304360. doi:10.1002/advs.202304360
de Vasconcelos, A. C. P., Morais, R. P., Novais, G. B., da, S. B. S., Menezes, L. R. O., Dos Santos, S., et al. (2020). In situ photocrosslinkable formulation of nanocomposites based on multi-walled carbon nanotubes and formononetin for potential application in spinal cord injury treatment. Nanomedicine 29, 102272. doi:10.1016/j.nano.2020.102272
de Villiers, T. J. (2023). Bone health and menopause: osteoporosis prevention and treatment. Best. Pract. Res. Clin. Endocrinol. Metab. 38, 101782. doi:10.1016/j.beem.2023.101782
Dong, Z., Wang, Y., Guo, J., Tian, C., Pan, W., Wang, H., et al. (2022). Prostate cancer therapy using docetaxel and formononetin combination: hyaluronic acid and epidermal growth factor receptor targeted peptide dual ligands modified binary nanoparticles to facilitate the in vivo anti-tumor activity. Drug Des. Devel Ther. 16, 2683–2693. doi:10.2147/dddt.S366622
Fan, M., Li, Z., Hu, M., Zhao, H., Wang, T., Jia, Y., et al. (2022). Formononetin attenuates Aβ(25-35)-induced adhesion molecules in HBMECs via Nrf2 activation. Brain Res. Bull. 183, 162–171. doi:10.1016/j.brainresbull.2022.03.009
Fang, Y., Ye, J., Zhao, B., Sun, J., Gu, N., Chen, X., et al. (2020). Formononetin ameliorates oxaliplatin-induced peripheral neuropathy via the KEAP1-NRF2-GSTP1 axis. Redox Biol. 36, 101677. doi:10.1016/j.redox.2020.101677
Feng, Q., Yang, Y., Ren, K., Qiao, Y., Sun, Z., Pan, S., et al. (2023). Broadening horizons: the multifaceted functions of ferroptosis in kidney diseases. Int. J. Biol. Sci. 19 (12), 3726–3743. doi:10.7150/ijbs.85674
Feng, Z., Gu, L., Lin, J., Wang, Q., Yu, B., Yao, X., et al. (2024). Formononetin protects against Aspergillus fumigatus Keratitis: targeting inflammation and fungal load. Int. Immunopharmacol. 132, 112046. doi:10.1016/j.intimp.2024.112046
Fu, H., Li, M., Huan, Y., Wang, X., Tao, M., Jiang, T., et al. (2023). Formononetin inhibits microglial inflammatory response and contributes to spinal cord injury repair by targeting the EGFR/MAPK pathway. Immunol. Invest 52 (4), 399–414. doi:10.1080/08820139.2023.2183135
Fu, X., Qin, T., Yu, J., Jiao, J., Ma, Z., Fu, Q., et al. (2019). Formononetin ameliorates cognitive disorder via PGC-1α pathway in neuroinflammation conditions in high-fat diet-induced mice. CNS Neurol. Disord. Drug Targets 18 (7), 566–577. doi:10.2174/1871527318666190807160137
Galindo, C. L., Khan, S., Zhang, X., Yeh, Y. S., Liu, Z., and Razani, B. (2023). Lipid-laden foam cells in the pathology of atherosclerosis: shedding light on new therapeutic targets. Expert Opin. Ther. Targets 27 (12), 1231–1245. doi:10.1080/14728222.2023.2288272
Gao, Y., Yao, Y., Zhu, Y., and Ren, G. (2015). Isoflavone content and composition in chickpea (Cicer arietinum L.) sprouts germinated under different conditions. J. Agric. Food Chem. 63 (10), 2701–2707. doi:10.1021/jf5057524
Guo, P., Liu, L., Yang, X., Li, M., Zhao, Q., and Wu, H. (2022). Irisin improves BBB dysfunction in SAP rats by inhibiting MMP-9 via the ERK/NF-κB signaling pathway. Cell Signal 93, 110300. doi:10.1016/j.cellsig.2022.110300
Han, N.-R., Park, H.-J., Ko, S.-G., and Moon, P.-D. (2023). The protective effect of a functional food consisting of Astragalus membranaceus, Trichosanthes kirilowii, and Angelica gigas or its active component formononetin against inflammatory skin disorders through suppression of TSLP via MDM2/HIF1α signaling pathways. Foods 12 (2), 276. doi:10.3390/foods12020276
Hao, Y., Miao, J., Liu, W., Peng, L., Chen, Y., and Zhong, Q. (2021). Formononetin protects against cisplatin-induced acute kidney injury through activation of the PPARα/Nrf2/HO-1/NQO1 pathway. Int. J. Mol. Med. 47 (2), 511–522. doi:10.3892/ijmm.2020.4805
Hashemi Goradel, N., Najafi, M., Salehi, E., Farhood, B., and Mortezaee, K. (2019). Cyclooxygenase-2 in cancer: a review. J. Cell Physiol. 234 (5), 5683–5699. doi:10.1002/jcp.27411
He, Y., Cai, Y., Wei, D., Cao, L., He, Q., and Zhang, Y. (2024). Elucidating the mechanisms of formononetin in modulating atherosclerotic plaque formation in ApoE-/- mice. BMC Cardiovasc Disord. 24 (1), 121. doi:10.1186/s12872-024-03774-6
Hu, W., Wu, X., Tang, J., Xiao, N., Zhao, G., Zhang, L., et al. (2019). In vitro and in vivo studies of antiosteosarcoma activities of formononetin. J. Cell Physiol. 234 (10), 17305–17313. doi:10.1002/jcp.28349
Huang, J., Chen, X., and Xie, A. (2021). Formononetin ameliorates IL-13-induced inflammation and mucus formation in human nasal epithelial cells by activating the SIRT1/Nrf2 signaling pathway. Mol. Med. Rep. 24 (6), 832. doi:10.3892/mmr.2021.12472
Huang, Q., Chen, H., Yin, K., Shen, Y., Lin, K., Guo, X., et al. (2022). Formononetin attenuates renal tubular injury and mitochondrial damage in diabetic nephropathy partly via regulating sirt1/PGC-1α pathway. Front. Pharmacol. 13, 901234. doi:10.3389/fphar.2022.901234
Hwang, J. S., Lee, S. A., Hong, S. S., Lee, K. S., Lee, M. K., Hwang, B. Y., et al. (2005). Monoamine oxidase inhibitory components from the roots of Sophora flavescens. Arch. Pharm. Res. 28 (2), 190–194. doi:10.1007/bf02977714
Hybertson, B. M., Gao, B., Bose, S. K., and McCord, J. M. (2011). Oxidative stress in health and disease: the therapeutic potential of Nrf2 activation. Mol. Aspects Med. 32 (4-6), 234–246. doi:10.1016/j.mam.2011.10.006
Imani, S., Zagari, Z., Rezaei Zarchi, S., Jorjani, M., and Nasri, S. (2016). Functional recovery of carbon nanotube/nafion nanocomposite in rat model of spinal cord injury. Artif. Cells Nanomed Biotechnol. 44 (1), 144–149. doi:10.3109/21691401.2015.1024844
Iwaki, T., Bennion, B. G., Stenson, E. K., Lynn, J. C., Otinga, C., Djukovic, D., et al. (2019). PPARα contributes to protection against metabolic and inflammatory derangements associated with acute kidney injury in experimental sepsis. Physiol. Rep. 7 (10), e14078. doi:10.14814/phy2.14078
Jia, C., Hu, F., Lu, D., Jin, H., Lu, H., Xue, E., et al. (2022). Formononetin inhibits IL-1β-induced inflammation in human chondrocytes and slows the progression of osteoarthritis in rat model via the regulation of PTEN/AKT/NF-κB pathway. Int. Immunopharmacol. 113 (Pt A), 109309. doi:10.1016/j.intimp.2022.109309
Jia, W. D., Bai, X., Ma, Q. Q., Bian, M., Bai, C. M., Li, D., et al. (2023). Synthesis, molecular docking studies of formononetin derivatives as potent Bax agonists for anticancer activity. Nat. Prod. Res. 39, 423–437. doi:10.1080/14786419.2023.2269592
Jiang, P., Jiang, W., Li, X., and Zhu, Q. (2024). Combination of formononetin and sulforaphane natural drug repress the proliferation of cervical cancer cells via impeding PI3K/AKT/mTOR pathway. Appl. Biochem. Biotechnol. 196, 6726–6744. doi:10.1007/s12010-024-04873-y
Jiang, W., Hu, T., Ye, C., Hu, M., Yu, Q., Sun, L., et al. (2023). Formononetin attenuates high glucose-induced neurotoxicity by negatively regulating oxidative stress and mitochondrial dysfunction in Schwann cells via activation of SIRT3. Food Chem. Toxicol. 182, 114156. doi:10.1016/j.fct.2023.114156
Kamiya, K., Kishimoto, M., Sugai, J., Komine, M., and Ohtsuki, M. (2019). Risk factors for the development of psoriasis. Int. J. Mol. Sci. 20 (18), 4347. doi:10.3390/ijms20184347
Kim, M. S., Park, J. S., Chung, Y. C., Jang, S., Hyun, C. G., and Kim, S. Y. (2019). Anti-inflammatory effects of formononetin 7-O-phosphate, a novel biorenovation product, on LPS-stimulated RAW 264.7 macrophage cells. Molecules 24 (21), 3910. doi:10.3390/molecules24213910
Kraft, C., Jenett-Siems, K., Siems, K., Gupta, M. P., Bienzle, U., and Eich, E. (2000). Antiplasmodial activity of isoflavones from Andira inermis. J. Ethnopharmacol. 73 (1-2), 131–135. doi:10.1016/s0378-8741(00)00285-3
Křížová, L., Dadáková, K., Kašparovská, J., and Kašparovský, T. (2019). Isoflavones. Molecules 24 (6), 1076. doi:10.3390/molecules24061076
Krueger, J. S., Keshamouni, V. G., Atanaskova, N., and Reddy, K. B. (2001). Temporal and quantitative regulation of mitogen-activated protein kinase (MAPK) modulates cell motility and invasion. Oncogene 20 (31), 4209–4218. doi:10.1038/sj.onc.1204541
Lapcík, O., Hill, M., Hampl, R., Wähälä, K., and Adlercreutz, H. (1998). Identification of isoflavonoids in beer. Steroids 63 (1), 14–20. doi:10.1016/s0039-128x(97)00104-9
Lee, M. (2013). Neurotransmitters and microglial-mediated neuroinflammation. Curr. Protein Pept. Sci. 14 (1), 21–32. doi:10.2174/1389203711314010005
Lee, Y. S., and Dutta, A. (2009). MicroRNAs in cancer. Annu. Rev. Pathol. 4, 199–227. doi:10.1146/annurev.pathol.4.110807.092222
Li, H., Jiang, R., Lou, L., Jia, C., Zou, L., and Chen, M. (2022b). Formononetin improves the survival of random skin flaps through PI3K/Akt-Mediated Nrf2 antioxidant defense system. Front. Pharmacol. 13, 901498. doi:10.3389/fphar.2022.901498
Li, K., Shen, H., Lu, M., Chen, J., Yin, Q., and Li, P. (2020). Formononetin inhibits osteosarcoma cell proliferation and promotes apoptosis by regulating the miR-214-3p/phosphatase and tensin homolog pathway. Transl. Cancer Res. 9 (8), 4914–4921. doi:10.21037/tcr-20-2296
Li, L., Sun, J. X., Wang, X. Q., Liu, X. K., Chen, X. X., Zhang, B., et al. (2017). Notoginsenoside R7 suppresses cervical cancer via PI3K/PTEN/Akt/mTOR signaling. Oncotarget 8 (65), 109487–109496. doi:10.18632/oncotarget.22721
Li, S., Li, J., Fan, Y., Huang, T., Zhou, Y., Fan, H., et al. (2022d). The mechanism of formononetin/calycosin compound optimizing the effects of temozolomide on C6 malignant glioma based on metabolomics and network pharmacology. Biomed. Pharmacother. 153, 113418. doi:10.1016/j.biopha.2022.113418
Li, S., Zhu, L., He, Y., and Sun, T. (2023). Formononetin enhances the chemosensitivity of triple negative breast cancer via BTB domain and CNC homolog 1-mediated mitophagy pathways. Acta Biochim. Pol. 70 (3), 533–539. doi:10.18388/abp.2020_6466
Li, X., Jiang, X., Zeng, R., Lai, X., Wang, J., Liu, H., et al. (2024a). Formononetin attenuates cigarette smoke-induced COPD in mice by suppressing inflammation, endoplasmic reticulum stress, and apoptosis in bronchial epithelial cells via AhR/CYP1A1 and AKT/mTOR signaling pathways. Phytother. Res. 38 (3), 1278–1293. doi:10.1002/ptr.8104
Li, X., Lu, L., Hou, W., Huang, T., Chen, X., Qi, J., et al. (2022a). Epigenetics in the pathogenesis of diabetic nephropathy. Acta Biochim. Biophys. Sin. (Shanghai) 54 (2), 163–172. doi:10.3724/abbs.2021016
Li, X., Ma, T. K., Wang, P., Shi, H., Hai, S., Qin, Y., et al. (2024b). HOXD10 attenuates renal fibrosis by inhibiting NOX4-induced ferroptosis. Cell Death Dis. 15 (6), 398. doi:10.1038/s41419-024-06780-w
Li, Y., Wu, J., Yu, H., Lu, X., and Ni, Y. (2024c). Formononetin ameliorates cisplatin-induced hair cell death via activation of the PI3K/AKT-Nrf2 signaling pathway. Heliyon 10 (1), e23750. doi:10.1016/j.heliyon.2023.e23750
Li, Y., Zhang, H., Merkher, Y., Chen, L., Liu, N., Leonov, S., et al. (2022c). Recent advances in therapeutic strategies for triple-negative breast cancer. J. Hematol. Oncol. 15 (1), 121. doi:10.1186/s13045-022-01341-0
Liang, C., Zhou, A., Sui, C., and Huang, Z. (2019). The effect of formononetin on the proliferation and migration of human umbilical vein endothelial cells and its mechanism. Biomed. Pharmacother. 111, 86–90. doi:10.1016/j.biopha.2018.12.049
Liao, J., Xie, X., Wang, N., Wang, Y., Zhao, J., Chen, F., et al. (2024). Formononetin promotes fatty acid β-oxidation to treat non-alcoholic steatohepatitis through SIRT1/PGC-1α/PPARα pathway. Phytomedicine 124, 155285. doi:10.1016/j.phymed.2023.155285
Liao, L., Huang, L., Wei, X., Yin, L., Wei, X., and Li, T. (2021). Bioinformatic and biochemical studies of formononetin against liver injure. Life Sci. 272, 119229. doi:10.1016/j.lfs.2021.119229
Liu, G., Zhao, W., Bai, J., Cui, J., Liang, H., and Lu, B. (2021a). Formononetin protects against concanavalin-A-induced autoimmune hepatitis in mice through its anti-apoptotic and anti-inflammatory properties. Biochem. Cell Biol. 99 (2), 231–240. doi:10.1139/bcb-2020-0197
Liu, H., Wang, Z., and Liu, Z. (2024). Formononetin restrains tumorigenesis of breast tumor by restraining STING-NF-κb and interfering with the activation of PD-L1. Discov. Med. 36 (182), 613–620. doi:10.24976/Discov.Med.202436182.58
Liu, L., Hu, R., You, H., Li, J., Liu, Y., Li, Q., et al. (2021b). Formononetin ameliorates muscle atrophy by regulating myostatin-mediated PI3K/Akt/FoxO3a pathway and satellite cell function in chronic kidney disease. J. Cell Mol. Med. 25 (3), 1493–1506. doi:10.1111/jcmm.16238
Liu, S., Luo, W., and Wang, Y. (2022). Emerging role of PARP-1 and PARthanatos in ischemic stroke. J. Neurochem. 160 (1), 74–87. doi:10.1111/jnc.15464
Liu, X. H., Zhu, R. J., Hu, F., Guo, L., Yang, Y. L., and Feng, S. L. (2015). Tissue distribution of six major bio-active components after oral administration of Zhenqi Fuzheng capsules to rats using ultra-pressure liquid chromatography-tandem mass spectrometry. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 986-987, 44–53. doi:10.1016/j.jchromb.2015.01.033
Lu, S., Huang, J., Zhang, J., Wu, C., Huang, Z., Tao, X., et al. (2024). The anti-hepatocellular carcinoma effect of Aidi injection was related to the synergistic action of cantharidin, formononetin, and isofraxidin through BIRC5, FEN1, and EGFR. J. Ethnopharmacol. 319 (Pt 2), 117209. doi:10.1016/j.jep.2023.117209
Luo, J., Cai, Y., Wei, D., Cao, L., He, Q., and Wu, Y. (2024). Formononetin alleviates cerebral ischemia-reperfusion injury in rats by targeting the PARP-1/PARG/Iduna signaling pathway. Brain Res. 1829, 148845. doi:10.1016/j.brainres.2024.148845
Luo, Y., Ma, J., and Lu, W. (2020). The significance of mitochondrial dysfunction in cancer. Int. J. Mol. Sci. 21 (16), 5598. doi:10.3390/ijms21165598
Lv, J., Zhuang, K., Jiang, X., Huang, H., and Quan, S. (2020). Renoprotective effect of formononetin by suppressing Smad3 expression in Db/Db mice. Diabetes Metab. Syndr. Obes. 13, 3313–3324. doi:10.2147/dmso.S272147
Ma, C., Xia, R., Yang, S., Liu, L., Zhang, J., Feng, K., et al. (2020). Formononetin attenuates atherosclerosis via regulating interaction between KLF4 and SRA in apoE(-/-) mice. Theranostics 10 (3), 1090–1106. doi:10.7150/thno.38115
Ma, Z., Zhang, D., Sun, J., Zhang, Q., Qiao, Y., Zhu, Y., et al. (2022). Formononetin inhibits hepatic I/R-Induced injury through regulating PHB2/PINK1/parkin pathway. Oxid. Med. Cell Longev. 2022, 6481192. doi:10.1155/2022/6481192
Massagué, J. (2004). G1 cell-cycle control and cancer. Nature 432 (7015), 298–306. doi:10.1038/nature03094
Mendonça, M. A. A. d., Ribeiro, A. R. S., Lima, A. K. d., Bezerra, G. B., Pinheiro, M. S., Albuquerque-Júnior, R. L. C. d., et al. (2020). Red propolis and its dyslipidemic regulator formononetin: evaluation of antioxidant activity and gastroprotective effects in rat model of gastric ulcer. Nutrients 12 (10), 2951. doi:10.3390/nu12102951
Mousa, A. M., El-Sammad, N. M., Hassan, S. K., Madboli, A. E. N. A., Hashim, A. N., Moustafa, E. S., et al. (2019). Antiulcerogenic effect of Cuphea ignea extract against ethanol-induced gastric ulcer in rats. BMC Complementary Altern. Med. 19 (1), 345. doi:10.1186/s12906-019-2760-9
Naudhani, M., Thakur, K., Ni, Z. J., Zhang, J. G., and Wei, Z. J. (2021). Formononetin reshapes the gut microbiota, prevents progression of obesity and improves host metabolism. Food Funct. 12 (24), 12303–12324. doi:10.1039/d1fo02942h
Ngamrojanavanich, N., Loontaisong, A., Pengpreecha, S., Cherdshewasart, W., Pornpakakul, S., Pudhom, K., et al. (2007). Cytotoxic constituents from butea superba roxb. J. Ethnopharmacol. 109 (2), 354–358. doi:10.1016/j.jep.2006.07.034
Ni, K. N., Ye, L., Zhang, Y. J., Fang, J. W., Yang, T., Pan, W. Z., et al. (2023). Formononetin improves the inflammatory response and bone destruction in knee joint lesions by regulating the NF-kB and MAPK signaling pathways. Phytother. Res. 37 (8), 3363–3379. doi:10.1002/ptr.7810
Ni, Q., Fan, Y., Zhang, X., Fan, H., and Li, Y. (2019). In vitro and in vivo study on glioma treatment enhancement by combining temozolomide with calycosin and formononetin. J. Ethnopharmacol. 242, 111699. doi:10.1016/j.jep.2019.01.023
Oh, J. S., Kim, T. H., Park, J. H., Lim, H., Cho, I. A., You, J. S., et al. (2020). Formononetin induces apoptotic cell death through the suppression of mitogen-activated protein kinase and nuclear factor-κB phosphorylation in FaDu human head and neck squamous cell carcinoma cells. Oncol. Rep. 43 (2), 700–710. doi:10.3892/or.2019.7432
Ouyang, B., Deng, L., Yang, F., Shi, H., Wang, N., Tang, W., et al. (2023). Albumin-based formononetin nanomedicines for lung injury and fibrosis therapy via blocking macrophage pyroptosis. Mater Today Bio 20, 100643. doi:10.1016/j.mtbio.2023.100643
Oza, M. J., and Kulkarni, Y. A. (2020). Formononetin ameliorates diabetic neuropathy by increasing expression of SIRT1 and NGF. Chem. Biodivers. 17 (6), e2000162. doi:10.1002/cbdv.202000162
Pan, C., Wang, H., Shan, H., and Lü, H. (2021). Preparative isolation and purification of calycosin and formononetin from astragali radix using hydrolytic extraction combined with high speed countercurrent chromatography. J. Chromatogr. Sci. 59 (5), 412–418. doi:10.1093/chromsci/bmab021
Park, W., Ahn, C. H., Cho, H., Kim, C. K., Shin, J., and Oh, K. B. (2017). Inhibitory effects of flavonoids from spatholobus suberectus on sortase A and sortase A-mediated aggregation of Streptococcus mutans. J. Microbiol. Biotechnol. 27 (8), 1457–1460. doi:10.4014/jmb.1704.04001
Park, Y., Choo, S. P., Jung, G. S., Kim, S., Lee, M. J., Im, W., et al. (2023). Formononetin inhibits progression of endometriosis via regulation of p27, pSTAT3, and progesterone receptor: in vitro and in vivo studies. Nutrients 15 (13), 3001. doi:10.3390/nu15133001
Patra, S., Gorai, S., Pal, S., Ghosh, K., Pradhan, S., and Chakrabarti, S. (2023). A review on phytoestrogens: current status and future direction. Phytother. Res. 37 (7), 3097–3120. doi:10.1002/ptr.7861
Pingale, T. D., and Gupta, G. L. (2023). Acute and sub-acute toxicity study reveals no dentrimental effect of formononetin in mice upon repeated i.p. dosing. Toxicol. Mech. Methods 33 (8), 688–697. doi:10.1080/15376516.2023.2234026
Plana, D., Palmer, A. C., and Sorger, P. K. (2022). Independent drug action in combination therapy: implications for precision oncology. Cancer Discov. 12 (3), 606–624. doi:10.1158/2159-8290.Cd-21-0212
Prakash, S. (2023). Nano-based drug delivery system for therapeutics: a comprehensive review. Biomed. Phys. Eng. Express 9 (5), 052002. doi:10.1088/2057-1976/acedb2
Qian, L., Xu, H., Yuan, R., Yun, W., and Ma, Y. (2024). Formononetin ameliorates isoproterenol induced cardiac fibrosis through improving mitochondrial dysfunction. Biomed. Pharmacother. 170, 116000. doi:10.1016/j.biopha.2023.116000
Qiu, Y. H., Zhang, T. S., Wang, X. W., Wang, M. Y., Zhao, W. X., Zhou, H. M., et al. (2021). Mitochondria autophagy: a potential target for cancer therapy. J. Drug Target 29 (6), 576–591. doi:10.1080/1061186x.2020.1867992
Ren, Y., and Qu, S. (2023). Constituent isoflavones of Puerariae radix as a potential neuroprotector in cognitive impairment: evidence from preclinical studies. Ageing Res. Rev. 90, 102040. doi:10.1016/j.arr.2023.102040
Riely, G. J., Wood, D. E., Ettinger, D. S., Aisner, D. L., Akerley, W., Bauman, J. R., et al. (2024). Non-small cell lung cancer, version 4.2024, NCCN clinical practice guidelines in oncology. J. Natl. Compr. Canc Netw. 22 (4), 249–274. doi:10.6004/jnccn.2204.0023
Rubab, F., Ijaz, H., Hussain, S., Munir, A., Stuppner, S., Jakschitz, T., et al. (2022). Gastroprotective effects of Caragana ambigua stocks on ethanol-induced gastric ulcer in rats supported by LC–MS/MS characterization of formononetin and biochanin A. J. Sci. Food Agric. 102 (15), 7030–7038. doi:10.1002/jsfa.12064
Safi, M., Ahmed, H., Al-Azab, M., Xia, Y.-l., Shan, X., Al-radhi, M., et al. (2021). PD-1/PDL-1 inhibitors and cardiotoxicity; molecular, etiological and management outlines. J. Adv. Res. 29, 45–54. doi:10.1016/j.jare.2020.09.006
Sakakibara, H., Viala, D., Ollier, A., Combeau, A., and Besle, J. M. (2004). Isoflavones in several clover species and in milk from goats fed clovers. Biofactors 22 (1-4), 237–239. doi:10.1002/biof.5520220147
Samsu, N. (2021). Diabetic nephropathy: challenges in pathogenesis, diagnosis, and treatment. Biomed. Res. Int. 2021, 1497449. doi:10.1155/2021/1497449
Semenza, G. L. (2003). Targeting HIF-1 for cancer therapy. Nat. Rev. Cancer 3 (10), 721–732. doi:10.1038/nrc1187
Sheka, A. C., Adeyi, O., Thompson, J., Hameed, B., Crawford, P. A., and Ikramuddin, S. (2020). Nonalcoholic steatohepatitis: a review. Jama 323 (12), 1175–1183. doi:10.1001/jama.2020.2298
Shi, Y., and Vanhoutte, P. M. (2017). Macro- and microvascular endothelial dysfunction in diabetes. J. Diabetes 9 (5), 434–449. doi:10.1111/1753-0407.12521
Silva, M. V. F., Loures, C. d.M. G., Alves, L. C. V., de Souza, L. C., Borges, K. B. G., and Carvalho, M. d.G. (2019). Alzheimer’s disease: risk factors and potentially protective measures. J. Biomed. Sci. 26 (1), 33. doi:10.1186/s12929-019-0524-y
Soundharrajan, I., Kim, D. H., Kuppusamy, P., and Choi, K. C. (2019). Modulation of osteogenic and myogenic differentiation by a phytoestrogen formononetin via p38MAPK-dependent JAK-STAT and Smad-1/5/8 signaling pathways in mouse myogenic progenitor cells. Sci. Rep. 9 (1), 9307. doi:10.1038/s41598-019-45793-w
Sroka-Tomaszewska, J., and Trzeciak, M. (2021). Molecular mechanisms of atopic dermatitis pathogenesis. Int. J. Mol. Sci. 22 (8), 4130. doi:10.3390/ijms22084130
Sugimoto, M., Ko, R., Goshima, H., Koike, A., Shibano, M., and Fujimori, K. (2021). Formononetin attenuates H(2)O(2)-induced cell death through decreasing ROS level by PI3K/Akt-Nrf2-activated antioxidant gene expression and suppressing MAPK-regulated apoptosis in neuronal SH-SY5Y cells. Neurotoxicology 85, 186–200. doi:10.1016/j.neuro.2021.05.014
Tang, Y., Zhou, X., Cao, T., Chen, E., Li, Y., Lei, W., et al. (2022). Endoplasmic reticulum stress and oxidative stress in inflammatory diseases. DNA Cell Biol. 41 (11), 924–934. doi:10.1089/dna.2022.0353
Vaya, J., Belinky, P. A., and Aviram, M. (1997). Antioxidant constituents from licorice roots: isolation, structure elucidation and antioxidative capacity toward LDL oxidation. Free Radic. Biol. Med. 23 (2), 302–313. doi:10.1016/s0891-5849(97)00089-0
Wang, D. S., Yan, L. Y., Yang, D. Z., Lyu, Y., Fang, L. H., Wang, S. B., et al. (2020a). Formononetin ameliorates myocardial ischemia/reperfusion injury in rats by suppressing the ROS-TXNIP-NLRP3 pathway. Biochem. Biophys. Res. Commun. 525 (3), 759–766. doi:10.1016/j.bbrc.2020.02.147
Wang, G., Liu, H., Liu, Y., Li, H., Li, Z., Shao, G., et al. (2020b). Formononetin alleviates Streptococcus suis infection by targeting suilysin. Microb. Pathog. 147, 104388. doi:10.1016/j.micpath.2020.104388
Wang, J. Y., Jiang, M. W., Li, M. Y., Zhang, Z. H., Xing, Y., Ri, M., et al. (2022). Formononetin represses cervical tumorigenesis by interfering with the activation of PD-L1 through MYC and STAT3 downregulation. J. Nutr. Biochem. 100, 108899. doi:10.1016/j.jnutbio.2021.108899
Wang, W. S., and Zhao, C. S. (2021). Formononetin exhibits anticancer activity in gastric carcinoma cell and regulating miR-542-5p. Kaohsiung J. Med. Sci. 37 (3), 215–225. doi:10.1002/kjm2.12322
Wang, X., Shen, Y., Shang, M., Liu, X., and Munn, L. L. (2023a). Endothelial mechanobiology in atherosclerosis. Cardiovasc Res. 119 (8), 1656–1675. doi:10.1093/cvr/cvad076
Wang, X., Zhou, Y., Wang, D., Wang, Y., Zhou, Z., Ma, X., et al. (2023b). Cisplatin-induced ototoxicity: from signaling network to therapeutic targets. Biomed. Pharmacother. 157, 114045. doi:10.1016/j.biopha.2022.114045
Wang, X. S., Guan, S. Y., Liu, A., Yue, J., Hu, L. N., Zhang, K., et al. (2019a). Anxiolytic effects of Formononetin in an inflammatory pain mouse model. Mol. Brain 12 (1), 36. doi:10.1186/s13041-019-0453-4
Wang, Y., Deng, Z., Wang, X., Shi, Y., Lu, Y., Fang, S., et al. (2021). Formononetin/methyl-β-cyclodextrin inclusion complex incorporated into electrospun polyvinyl-alcohol nanofibers: enhanced water solubility and oral fast-dissolving property. Int. J. Pharm. 603, 120696. doi:10.1016/j.ijpharm.2021.120696
Wang, Y., Zhao, H., Li, X., Wang, Q., Yan, M., Zhang, H., et al. (2019b). Formononetin alleviates hepatic steatosis by facilitating TFEB-mediated lysosome biogenesis and lipophagy. J. Nutr. Biochem. 73, 108214. doi:10.1016/j.jnutbio.2019.07.005
Willis, S., Day, C. L., Hinds, M. G., and Huang, D. C. (2003). The Bcl-2-regulated apoptotic pathway. J. Cell Sci. 116 (Pt 20), 4053–4056. doi:10.1242/jcs.00754
Wu, J., Kong, M., Lou, Y., Li, L., Yang, C., Xu, H., et al. (2020c). Simultaneous activation of erk1/2 and Akt signaling is critical for formononetin-induced promotion of endothelial function. Front. Pharmacol. 11, 608518. doi:10.3389/fphar.2020.608518
Wu, J., Xie, Y., Xiang, Z., Wang, C., Zhou, H., and Liu, L. (2016). Simultaneous determination of multiple components in Guanjiekang in rat plasma via the UPLC-MS/MS method and its application in pharmacokinetic study. Molecules 21 (12), 1732. doi:10.3390/molecules21121732
Wu, J., Xu, W., Ma, L., Sheng, J., Ye, M., Chen, H., et al. (2021). Formononetin relieves the facilitating effect of lncRNA AFAP1-AS1-miR-195/miR-545 axis on progression and chemo-resistance of triple-negative breast cancer. Aging (Albany NY) 13 (14), 18191–18222. doi:10.18632/aging.203156
Wu, Q. L., Cheng, Y. Q., Liu, A. J., and Zhang, W. D. (2020a). Formononetin recovered injured nerve functions by enhancing synaptic plasticity in ischemic stroke rats. Biochem. Biophys. Res. Commun. 525, 67–72. doi:10.1016/j.bbrc.2020.02.015
Wu, Y., Cai, C., Yang, L., Xiang, Y., Zhao, H., and Zeng, C. (2020b). Inhibitory effects of formononetin on the monocrotaline-induced pulmonary arterial hypertension in rats. Mol. Med. Rep. 21 (3), 1192–1200. doi:10.3892/mmr.2020.10911
Xiang, K., Shen, P., Gao, Z., Liu, Z., Hu, X., Liu, B., et al. (2022). Formononetin protects LPS-induced mastitis through suppressing inflammation and enhancing blood-milk barrier integrity via AhR-induced Src inactivation. Front. Immunol. 13, 814319. doi:10.3389/fimmu.2022.814319
Xin, M., Wang, Y., Ren, Q., and Guo, Y. (2019). Formononetin and metformin act synergistically to inhibit growth of MCF-7 breast cancer cells in vitro. Biomed. Pharmacother. 109, 2084–2089. doi:10.1016/j.biopha.2018.09.033
Xu, H. T., Zheng, Q., Tai, Z. G., Jiang, W. C., Xie, S. Q., Luo, Y., et al. (2024). Formononetin attenuates psoriasiform inflammation by regulating interferon signaling pathway. Phytomedicine 128, 155412. doi:10.1016/j.phymed.2024.155412
Xu, S., Tang, L., Li, X., Fan, F., and Liu, Z. (2020). Immunotherapy for glioma: current management and future application. Cancer Lett. 476, 1–12. doi:10.1016/j.canlet.2020.02.002
Yan, X. J., Wang, Z. J., Wang, H., Wei, M. Z., Chen, Y. C., Zhao, Y. L., et al. (2024). Formononetin derivative for osteoporosis by simultaneous regulating osteoblast and osteoclast. J. Nat. Prod. 87 (8), 2004–2013. doi:10.1021/acs.jnatprod.4c00437
Yang, C., Xie, Q., Zeng, X., Tao, N., Xu, Y., Chen, Y., et al. (2019b). Novel hybrids of podophyllotoxin and formononetin inhibit the growth, migration and invasion of lung cancer cells. Bioorg Chem. 85, 445–454. doi:10.1016/j.bioorg.2019.02.019
Yang, J., Sha, X., Wu, D., Wu, B., Pan, X., Pan, L. L., et al. (2023b). Formononetin alleviates acute pancreatitis by reducing oxidative stress and modulating intestinal barrier. Chin. Med. 18 (1), 78. doi:10.1186/s13020-023-00773-1
Yang, K., Wang, Y., Li, Y. W., Chen, Y. G., Xing, N., Lin, H. B., et al. (2022). Progress in the treatment of diabetic peripheral neuropathy. Biomed. Pharmacother. 148, 112717. doi:10.1016/j.biopha.2022.112717
Yang, S., Wei, L., Xia, R., Liu, L., Chen, Y., Zhang, W., et al. (2019a). Formononetin ameliorates cholestasis by regulating hepatic SIRT1 and PPARα. Biochem. Biophys. Res. Commun. 512 (4), 770–778. doi:10.1016/j.bbrc.2019.03.131
Yang, Y., Huang, T., Zhang, H., Li, X., Shi, S., Tian, X., et al. (2023a). Formononetin improves cardiac function and depressive behaviours in myocardial infarction with depression by targeting GSK-3β to regulate macrophage/microglial polarization. Phytomedicine 109, 154602. doi:10.1016/j.phymed.2022.154602
Yao, J. N., Zhang, X. X., Zhang, Y. Z., Li, J. H., Zhao, D. Y., Gao, B., et al. (2019). Discovery and anticancer evaluation of a formononetin derivative against gastric cancer SGC7901 cells. Invest New Drugs 37 (6), 1300–1308. doi:10.1007/s10637-019-00767-7
Yi, L., Lu, Y., Yu, S., Cheng, Q., and Yi, L. (2022). Formononetin inhibits inflammation and promotes gastric mucosal angiogenesis in gastric ulcer rats through regulating NF-κB signaling pathway. J. Recept Signal Transduct. Res. 42 (1), 16–22. doi:10.1080/10799893.2020.1837873
Yin, Z., Gong, G., Liu, X., and Yin, J. (2023). Mechanism of regulating macrophages/osteoclasts in attenuating wear particle-induced aseptic osteolysis. Front. Immunol. 14, 1274679. doi:10.3389/fimmu.2023.1274679
Ying, K., Liu, Y., Zhang, C., and Shangguan, M. (2019). Medical findings of nasopharyngeal carcinoma patients and anti-tumor benefits of formononetin. Eur. J. Pharmacol. 861, 172619. doi:10.1016/j.ejphar.2019.172619
Yu, L., Zhang, Y., Chen, Q., He, Y., Zhou, H., Wan, H., et al. (2022). Formononetin protects against inflammation associated with cerebral ischemia-reperfusion injury in rats by targeting the JAK2/STAT3 signaling pathway. Biomed. Pharmacother. 149, 112836. doi:10.1016/j.biopha.2022.112836
Yu, X., Gao, F., Li, W., Zhou, L., Liu, W., and Li, M. (2020). Formononetin inhibits tumor growth by suppression of EGFR-Akt-Mcl-1 axis in non-small cell lung cancer. J. Exp. Clin. Cancer Res. 39 (1), 62. doi:10.1186/s13046-020-01566-2
Yu, X., Yang, B., Chen, B., Wu, Q., Ren, Z., Wang, D., et al. (2023). Inhibitory effects of Formononetin on CoCrMo particle-induced osteoclast activation and bone loss through downregulating NF-κB and MAPK signaling. Cell Signal 106, 110651. doi:10.1016/j.cellsig.2023.110651
Yuan, W., Chen, Y., Zhou, Y., Bao, K., Yu, X., Xu, Y., et al. (2021). Formononetin attenuates atopic dermatitis by upregulating A20 expression via activation of G protein-coupled estrogen receptor. J. Ethnopharmacol. 266, 113397. doi:10.1016/j.jep.2020.113397
Zhang, B., Hao, Z., Zhou, W., Zhang, S., Sun, M., Li, H., et al. (2021). Formononetin protects against ox-LDL-induced endothelial dysfunction by activating PPAR-γ signaling based on network pharmacology and experimental validation. Bioengineered 12 (1), 4887–4898. doi:10.1080/21655979.2021.1959493
Zhang, C., Zhu, L., Lu, S., Li, M., Bai, M., Li, Y., et al. (2022). The antidepressant-like effect of formononetin on chronic corticosterone-treated mice. Brain Res. 1783, 147844. doi:10.1016/j.brainres.2022.147844
Zhang, J., Liu, L., Wang, J., Ren, B., Zhang, L., and Li, W. (2018). Formononetin, an isoflavone from Astragalus membranaceus inhibits proliferation and metastasis of ovarian cancer cells. J. Ethnopharmacol. 221, 91–99. doi:10.1016/j.jep.2018.04.014
Zhang, Y., Chen, C., and Zhang, J. (2019). Effects and significance of formononetin on expression levels of HIF-1α and VEGF in mouse cervical cancer tissue. Oncol. Lett. 18 (3), 2248–2253. doi:10.3892/ol.2019.10567
Zhao, L., Han, J., Liu, J., Fan, K., Yuan, T., Han, J., et al. (2021). A novel formononetin derivative promotes anti-ischemic effects on acute ischemic injury in mice. Front. Microbiol. 12, 786464. doi:10.3389/fmicb.2021.786464
Zhao, X., Zhang, S., Liu, D., Yang, M., and Wei, J. (2020). Analysis of flavonoids in dalbergia odorifera by ultra-performance liquid chromatography with tandem mass spectrometry. Molecules 25 (2), 389. doi:10.3390/molecules25020389
Zhou, H., Wang, P., Qin, X., Zhang, X., Lai, K. P., and Chen, J. (2023a). Comparative transcriptomic analysis and mechanistic characterization revealed the use of formononetin for bladder cancer treatment. Food Funct. 14 (12), 5787–5804. doi:10.1039/d2fo03962a
Zhou, Q., Zhang, W., Li, T., Tang, R., Li, C., Yuan, S., et al. (2019b). Formononetin enhances the tumoricidal effect of everolimus in breast cancer MDA-MB-468 cells by suppressing the mTOR pathway. Evid. Based Complement. Altern. Med. 2019, 9610629. doi:10.1155/2019/9610629
Zhou, Z., and Zhang, P. (2023). Formononetin ameliorates the LPS-induced inflammatory response and apoptosis of neuronal cells via NF-κB/NLRP3 signaling pathway. Funct. Integr. Genomics 23 (4), 321. doi:10.1007/s10142-023-01247-1
Zhou, Z., Zhou, H., Zou, X., Wang, X., and Yan, M. (2022). Formononetin regulates endothelial nitric oxide synthase to protect vascular endothelium in deep vein thrombosis rats. Int. J. Immunopathol. Pharmacol. 36, 3946320221111117. doi:10.1177/03946320221111117
Zhou, Z., Zhou, X., Dong, Y., Li, M., and Xu, Y. (2019a). Formononetin ameliorates high glucose-induced endothelial dysfunction by inhibiting the JAK/STAT signaling pathway. Mol. Med. Rep. 20 (3), 2893–2901. doi:10.3892/mmr.2019.10512
Zhou, Z. W., Zhu, X. Y., Li, S. Y., Lin, S. E., Zhu, Y. H., Ji, K., et al. (2023b). Formononetin inhibits mast cell degranulation to ameliorate compound 48/80-induced pseudoallergic reactions. Molecules 28 (13), 5271. doi:10.3390/molecules28135271
Zhu, B., Ni, Y., Gong, Y., Kang, X., Guo, H., Liu, X., et al. (2023). Formononetin ameliorates ferroptosis-associated fibrosis in renal tubular epithelial cells and in mice with chronic kidney disease by suppressing the Smad3/ATF3/SLC7A11 signaling. Life Sci. 315, 121331. doi:10.1016/j.lfs.2022.121331
Zhu, Y., Xian, X., Wang, Z., Bi, Y., Chen, Q., Han, X., et al. (2018). Research progress on the relationship between atherosclerosis and inflammation. Biomolecules 8 (3), 80. doi:10.3390/biom8030080
Zhuang, K., Jiang, X., Liu, R., Ye, C., Wang, Y., Wang, Y., et al. (2020). Formononetin activates the Nrf2/ARE signaling pathway via Sirt1 to improve diabetic renal fibrosis. Front. Pharmacol. 11, 616378. doi:10.3389/fphar.2020.616378
Keywords: formononetin, pharmacology, drug combination, toxicity, derivatives, drug delivery systems
Citation: Jin M, Wei L, Wang J, Shen Y, Gao L, Zhao F, Gao Q, Ma Y, Sun Y, Lin Y, Ji G, Cai P and Yan R (2025) Formononetin: a review of its source, pharmacology, drug combination, toxicity, derivatives, and drug delivery systems. Front. Pharmacol. 16:1534798. doi: 10.3389/fphar.2025.1534798
Received: 26 November 2024; Accepted: 31 January 2025;
Published: 03 March 2025.
Edited by:
Ruyu Yao, Chinese Academy of Sciences (CAS), ChinaReviewed by:
Zaixiang Lou, Jiangnan University, ChinaCopyright © 2025 Jin, Wei, Wang, Shen, Gao, Zhao, Gao, Ma, Sun, Lin, Ji, Cai and Yan. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Guanjie Ji, amlndWFuamllMzZAMTYzLmNvbQ==; Pingping Cai, cGluZ3BpbmdjYWlAMTI2LmNvbQ==; Rugen Yan, eXlhb2xlQG5qdWNtLmVkdS5jbg==
†These authors have contributed equally to this work
‡These authors have contributed equally to this work and share first authorship
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.