 Bin Cai
Bin Cai Ting Cai2
Ting Cai2 Zeyu Feng
Zeyu Feng
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
MINI REVIEW article
Front. Pharmacol. , 13 September 2024
Sec. Ethnopharmacology
Volume 15 - 2024 | https://doi.org/10.3389/fphar.2024.1404172
Cortex Fraxini is a traditional Chinese herb that is widely available, inexpensive, and has low toxicity. Modern pharmacological studies have demonstrated that the active metabolites in Cortex Fraxini, including esculin, esculetin, and fraxetin, exert anti-tumor activities by regulating genes and proteins involved in cancer cell proliferation, apoptosis, invasion, and migration. Additionally, these metabolites play a pivotal role in the regulation of several tumor-associated signaling pathways, including the PI3K/Akt, MAPK/ERK, JAK/STAT3, and Wnt/β-catenin pathways. Due to their pro-apoptotic and anti-proliferative properties in vitro and in vivo, Cortex Fraxini and its active metabolites may be considered as potential candidates for the treatment of tumor. The aim of this review is to highlight the anti-tumor biological activities and underlying mechanisms of action of the active metabolites of Cortex Fraxini, with a view to providing a reference for their further development and application in the treatment of tumors.
Cortex Fraxini (also known as Qinpi in Chinese) is a widely used herbal medicine that belongs to the “heat-clearing” category in traditional Chinese medicine. Cortex Fraxini is derived from the dried bark of Fraxinus rhynchophylla Hance, Fraxinus chinensis Roxb., Fraxinus szaboana Lingelsh., and Fraxinus stylosa Lingelsh. As a well-known traditional Chinese herbal medicine, Cortex Fraxini has been used for more than 2,000 years to treat conjunctivitis, diarrhea, hyperuricemia, bacillary dysentery, and excessive leukorrhea (Li et al., 2015; Wang G. et al., 2017). Modern pharmacological studies of Cortex Fraxini have revealed that it has a range of pharmacological effects, including anti-inflammatory, antioxidant, and antibacterial properties. (Wu et al., 2007). To date, phytochemical investigations have identified a variety of components in Cortex Fraxini, mainly coumarins, lignans, secoiridoids, phenolic acids, flavonoids, phenols, triterpenoids, and steroids (Zheng et al., 2024). Pharmacological studies have shown that coumarins are the primary active metabolites of Cortex Fraxini, including esculin, esculetin, fraxin, and fraxetin (Li C. X. et al., 2022). Moreover, numerous in vivo and in vitro studies have demonstrated that these active metabolites of Cortex Fraxini exert anti-tumor effects through cell cycle regulation, inhibition of tumor cell proliferation, induction of apoptosis, and other mechanisms (Li Z. Y. et al., 2022; Zhang et al., 2022). The anti-tumor effects of these active metabolites have recently attracted the attention of researchers, leading to increased interest in the anti-tumor pharmacological activity of Cortex Fraxini (Figure 1).
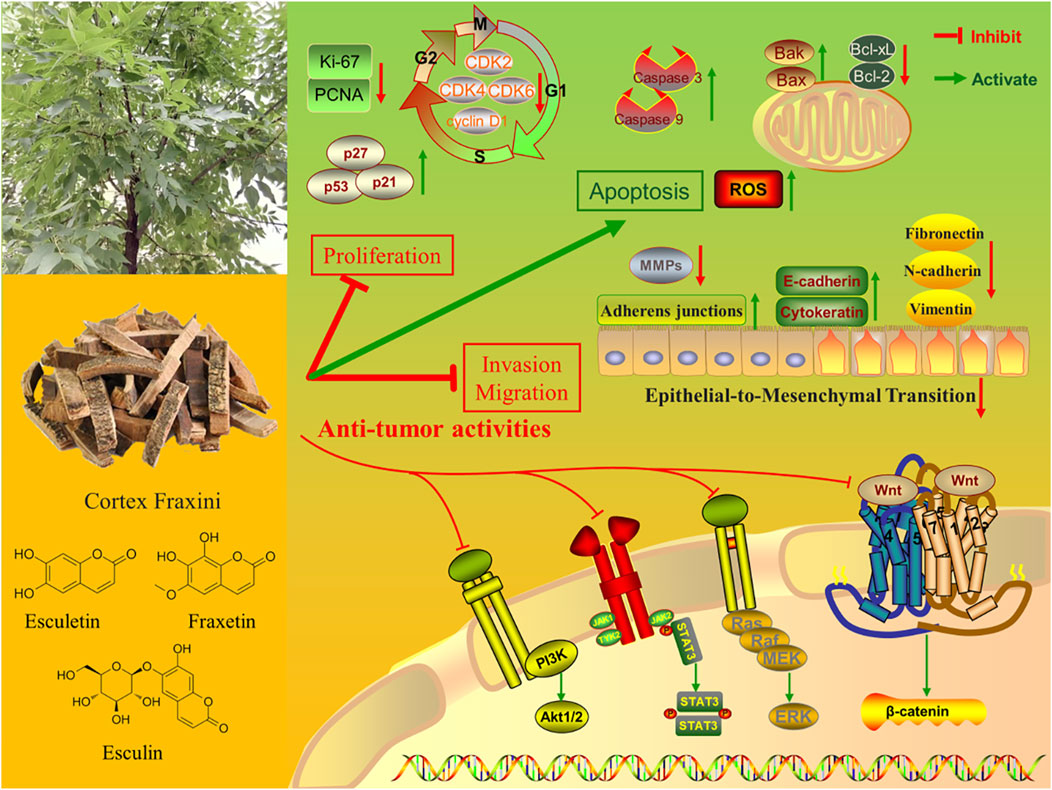
Figure 1. The anti-tumor pharmaceutical activities of Cortex Fraxini and its active metabolites.
Esculin is a glycosidic coumarin derivative (6-beta-glucoside-7-hydroxycoumarin, molecular formula: C15H16O9). The two parts of the molecule (glucose and 7-hydroxycoumarin) are linked by an ester linkage through oxygen. Esculin can be enzymatically hydrolyzed at the 8-glucose linkage to yield two products, esculetin and glucose. Studies have demonstrated that esculin can be distributed throughout the body in a rapid and uniform manner (Li et al., 2013). However, the first-pass effect of esculin was so severe that the calculated oral bioavailability was only 0.62% (Rehman et al., 2015). Esculetin is one of the simplest coumarins with two hydroxyl groups at carbons 6 and 7 (6,7-dihydroxy-2-chromenone, molecular formula: C9H6O4), which is the aglycone metabolite of esculin (Wang et al., 2016). The average oral bioavailability of esculetin was found to be 19%, which was significantly higher than that of esculin (Kwak et al., 2021). Moreover, esculetin demonstrated little cytotoxicity to human normal cells. A limited impact of esculetin on the viability of normal human gastric epithelial cells was observed in an in vitro study (Wang Y. et al., 2017). Fraxetin is a simple hydroxycoumarin (7,8-dihydroxy-6-methoxycoumarin, molecular formula: C10H8O5), and has minimal cytotoxic effects on normal human cells. A report on human melanoma demonstrated that high concentrations of fraxetin only slightly inhibited the viability of the human keratinocyte and melanocyte cell lines (Wróblewska-Łuczka et al., 2022). The content of fraxetin in the Cortex Fraxini was lower than that of esculin and esculetin (Zhou et al., 2008). This paper presents a systematic review of the existing studies on the anti-tumor effects of Cortex Fraxini and its active metabolites. Furthermore, our review is of great significance in exploring the anti-tumor potential of Cortex Fraxini, which may provide a reference for further research on Cortex Fraxini and its active metabolites in anti-tumor pharmaceutical applications.
The cell cycle is a highly conserved and precisely controlled process that consists of four distinct, ordered phases (G0/G1, S, G2, and M). Uncontrolled cell proliferation is a key factor in tumorigenesis and tumor progression. Cell cycle regulators such as cyclin-dependent kinase 2 (CDK2), CDK4, and cyclin D1 play an important role in this process. CDKs are key regulatory enzymes involved in cell proliferation by regulating cell cycle checkpoints and transcriptional events in response to extracellular and intracellular signals (Ding et al., 2020). Cyclin D1 is a key regulator of the cell cycle and plays a central role in the pathogenesis of cancer by determining uncontrolled cell proliferation (Fuste et al., 2016). Esculetin (10–100 μM) has been demonstrated to exert antiproliferative effects on osteosarcoma LM8 cells by inducing G1 cell cycle arrest. The mechanism of action appears to involve the suppression of CDK4 and cyclin D1 expression (Kimura and Sumiyoshi, 2015). Another study observed that esculetin induced G0/G1 cell cycle arrest in human leukemia cells (HL-60), which was accompanied by the suppression of cyclin D1, cyclin D3, CDK2, and CDK4 (Wang et al., 2019). Additionally, it was shown that esculetin induces cell cycle arrest at the G0/G1 phase in human colon cancer LoVo cells and that the protein expression of cyclin D1 is decreased after treatment with esculetin (Choi et al., 2019). Fraxetin has been demonstrated to induce tumor cell cycle arrest in various cell lines, including Huh7 and Hep3B hepatocellular carcinoma cell lines, where it causes cells to arrest at the G0/G1 phase (Song et al., 2021). Furthermore, fraxetin was shown to effectively induce cell cycle arrest at the G0/G1 phase by downregulating the expression of the G1/S transition regulatory proteins cyclin D1, CDK4, and CDK6 in human non-small cell lung cancer cell lines (HCC827 and H1650) (Zhang G. et al., 2019).
p53 is a primary tumor suppressor gene that functions to inhibit proliferation and eliminate abnormal cells. p53 regulates a variety of genes and activates various responses, including cell cycle arrest and apoptosis (Soussi, 2000). p21 is a potent CDK-binding inhibitor that is transcriptionally controlled by p53 pathways and is a key negative regulator of the cell cycle (Jassim et al., 2021). p27 has the function of binding to and inhibiting cyclin-CDK, leading to cell cycle arrest (Razavipour et al., 2020). Esculin has been reported to reduce the viability and proliferation of MDA-MB-231 triple-negative breast cancer cell lines in a dose- and time-dependent manner. In addition, the levels of p53 and p21 mRNAs and proteins were increased in a concentration-dependent manner after treatment with esculin (Mo et al., 2020). In human colon cancer LoVo cells, esculetin treatment inhibited LoVo cell proliferation and was accompanied by increased expression of p53, p21, and p27 proteins (Choi et al., 2019). Additionally, it was proposed that esculetin inhibits the proliferation of human prostate cancer cell lines (PC3, DU145, and LNCaP) by increasing the expression of p53, p21, and p27 (Turkekul et al., 2018).
The antigen Ki-67, also known as Ki-67 or Marker of Proliferation Ki-67 (MKI67), is a human protein encoded by the MKI67 gene (Davey et al., 2021). Ki-67 protein has been widely used as a proliferation marker for human tumor cells, and its expression is closely related to tumor prognosis (Louis et al., 2023). Proliferating cell nuclear antigen (PCNA) has been found in the nuclei of yeast, plant, and animal cells that are in the process of cell division, suggesting that it plays a role in regulating the cell cycle and/or DNA replication (Strzalka and Ziemienowicz, 2011). PCNA is considered essential for DNA replication in cancer cells and has also been implicated in tumor invasion (Wang et al., 2022). Esculetin was reported to inhibit human colorectal cancer HCT-116 cell proliferation by decreasing the expression of the proliferation-related proteins Ki-67 and PCNA (Yan et al., 2019). In vivo experiments in the HCT-116 subcutaneous tumor-bearing model also demonstrated a significant decrease in the expression of Ki-67 and PCNA following treatment with esculetin. In a xenograft model of colon cancer, treatment with esculetin at 20 mg/kg and 100 mg/kg significantly reduced tumor size by 44% and 64%, respectively. Furthermore, the expression of Ki-67 was suppressed by esculetin treatment in both colon cancer cells and xenograft tumor tissues (Lee et al., 2013). In human pancreatic cancer cell lines (PANC-1 and Patu8988), fraxetin was demonstrated to inhibit cell proliferation and reduce the expression of Ki-67 (Guo et al., 2021). Additionally, fraxetin was demonstrated to inhibit proliferation and reduce the protein expression of Ki-67 in human glioblastoma U251 cells (Qu et al., 2021) (Table 1).
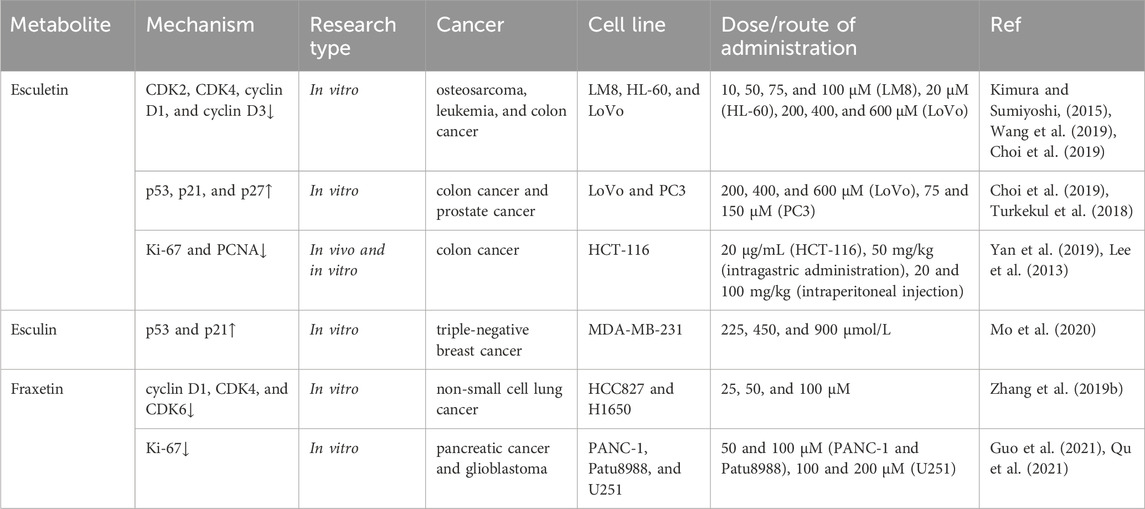
Table 1. Inhibition of cancer cell proliferation.
Esculetin has been shown to induce apoptosis of gastric cancer cell lines (MGC-803, HGC-27, and BGC-823) in a dose-dependent manner with lower toxicity against normal gastric epithelial cells (Wang G. et al., 2017). A mechanistic study revealed that esculetin induced apoptosis in MGC-803 cells by triggering the activation of the mitochondrial apoptotic pathway, increasing cytochrome c release from mitochondria, and activating caspase-3 and caspase-9 activity. In addition, esculetin dose-dependently inhibited the proliferation of human colon cancer LoVo cells and activated caspase-3, caspase-7, and caspase-9 (Choi et al., 2019). In human A549 lung cancer cells, esculin was shown to increase the activity of caspase-3 and caspase-9 (Wang, 2015). It has been demonstrated that esculetin is capable of inducing apoptosis in human leukemia HL-60 cells in a dose-dependent manner. Furthermore, esculetin treatment enhanced the cleavage of caspase-3 and caspase-9 and induced the release of cytochrome c in HL-60 cells (Wang et al., 2019). In another study, esculetin treatment led to Ca2+ influx and induced Ca2+ release from the endoplasmic reticulum in human breast cancer ZR-75-1 cells, thereby activating the Ca2+-associated mitochondrial apoptotic pathway (Chang et al., 2016). Fraxetin was also found to promote apoptosis by the mitochondrial pathway in tumor cells. It was reported that fraxetin inhibited proliferation and induced apoptosis in human endometrial cancer RL95-2 cells. Furthermore, the treatment with fraxetin resulted in a reduction of the mitochondrial membrane potential and an increased expression of caspase-3 and caspase-9 (Xu et al., 2020). Caspase-3 and caspase-9 are both members of the cysteine protease family. Caspase-9 is an initiator caspase; activated caspase-9 cleaves downstream caspases such as caspase-3, caspase-6, and caspase-7 (Kuida, 2000). Caspase-3 is the main executioner of apoptosis; it is cleaved and activated during the early stages of apoptosis to execute apoptosis by cleaving targeted cellular proteins (Eskandari and Eaves, 2022).
Members of the B cell lymphoma-2 (Bcl-2) protein family are key regulators with pro- or anti-apoptotic activities. By affecting mitochondrial membrane permeability changes, the pro- and anti-apoptotic proteins of the Bcl-2 family are involved in the regulation of apoptosis (Qian et al., 2022). Bcl-2 is a well-known anti-apoptotic protein that has been shown to promote carcinogenesis by resisting cell death (Warren et al., 2019). B-cell lymphoma-extra large (Bcl-xL) is also a member of the Bcl-2 family of anti-apoptotic proteins. Bcl-2-associated X protein (Bax) is a pro-apoptotic protein that can commit a cell to programmed death by permeabilizing the outer mitochondrial membrane and subsequently initiating the caspase cascade (Edlich, 2018). Bcl-2 antagonist/killer (Bak) is another pro-apoptotic factor, similar to Bax, that promotes apoptosis and increases susceptibility to apoptosis under appropriate stimulation (Qian et al., 2022). Esculetin was shown to exert a significant inhibitory effect on the viability of hepatocellular carcinoma SMMC-7721 cells in a dose- and time-dependent manner. And esculetin was able to reduce the weight of the tumors in a xenograft model of hepatocellular carcinoma (Wang et al., 2015). Mechanistic studies demonstrated that esculetin induced a mitochondrial-mediated apoptotic pathway in vivo and in vitro, increased Bax protein expression, decreased Bcl-2 protein expression, and caused loss of mitochondrial membrane potential. In oral squamous cell carcinoma cell lines (HN22 and HSC4), esculetin was also shown to induce apoptosis by upregulating Bax and downregulating Bcl-xL (Jeon et al., 2016). Similarly, fraxetin was found to exert regulatory effects on Bcl-2 family members. It was demonstrated that fraxetin suppressed cell viability and induced apoptotic cell death in HT-29 and HCT-116 colon cancer cell lines. Furthermore, fraxetin treatment resulted in an increase in the expression of Bax and Bak and a decrease in the expression of Bcl-2 and Bcl-xL in HT-29 and HCT-116 cells. (Lee et al., 2022). Additionally, fraxetin was shown to upregulate the expression of Bax and downregulate the expression of Bcl-2 in human non-small cell lung cancer cell lines (HCC827 and H1650), thereby inducing apoptosis (Zhang Y. et al., 2019).
Reactive oxygen species (ROS) are a group of highly reactive molecules that play a central role in cell signaling and regulation of apoptosis pathways. It has been demonstrated that ROS are intimately associated with the apoptotic activation of the mitochondrial pathway. Moreover, there is a correlation between ROS and death receptor-induced apoptosis, as well as endoplasmic reticulum stress-induced apoptosis (Redza-Dutordoir and Averill-Bates, 2016). It was reported that esculetin stimulated the generation of intracellular ROS and induced apoptosis in HT-29 colorectal cancer cells (Kim et al., 2015). Pan et al. reported that esculetin inhibited the growth of human gastric cancer cell lines (SGC-7901, MGC-803, and BGC-823) in a dose- and time-dependent manner and reduced the viability of gastric cancer cells by inducing apoptosis. Moreover, it was proposed that elevated intracellular ROS levels were a pivotal factor in esculetin-mediated cell death (Pan et al., 2015). Fraxetin treatment has been shown to promote apoptosis and induce ROS generation in colon cancer cell lines (HT-29 and HCT-116) (Lee et al., 2022), hepatocellular carcinoma cell lines (Huh7 and Hep3B) (Song et al., 2021), and pancreatic cancer cell lines (PANC-1 and Patu8988) (Guo et al., 2021) (Table 2).
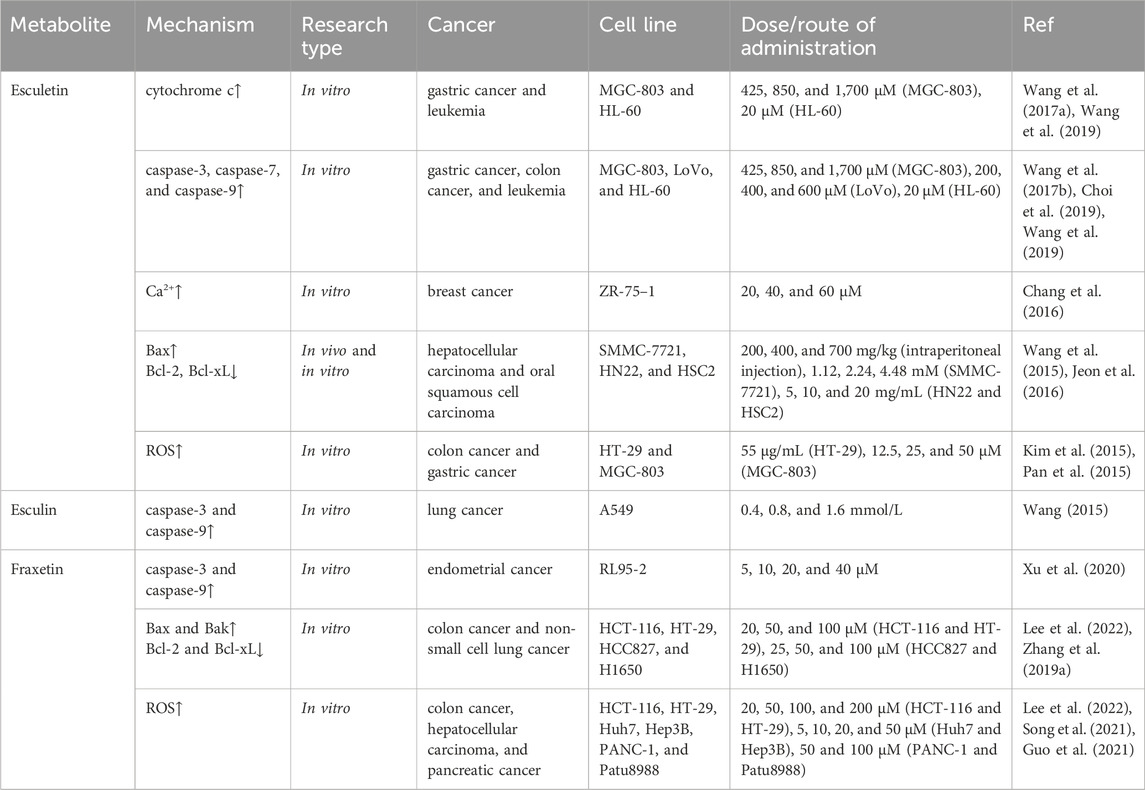
Table 2. Induction of cancer cell apoptosis.
The epithelial-to-mesenchymal transition (EMT) plays a pivotal role in both developmental processes and the progression of cancer. EMT enables solid tumors to become more malignant, increasing their invasiveness and metastatic activity. Events that occur during EMT include the loss of adherens junctions, the downregulation of epithelial-specific markers (e.g., cytokeratin and E-cadherin), and the upregulation of mesenchymal markers (e.g., fibronectin, N-cadherin, and vimentin) (Zhang and Weinberg, 2018). It was reported that esculetin inhibited the migration and EMT of colorectal cancer HCT-116 cells by downregulating EMT-related proteins N-cadherin, vimentin, and fibronectin (Yan et al., 2019). Xu et al. demonstrated that fraxetin suppressed the expression of EMT-associated markers N-cadherin, snail, and vimentin and increased the expression of E-cadherin in ovarian cancer cell lines (SKOV3 and SW626) (Xu et al., 2023). Furthermore, fraxetin treatment was also found to suppress the invasion and migration of pancreatic cancer PANC-1 and Patu8988 cells by regulating the Slug-E-cadherin axis-dependent EMT process (Guo et al., 2021).
Matrix metalloproteinases (MMPs) are a family of zinc-dependent proteolytic endopeptidases with extracellular matrix remodeling and degradation properties and have long been implicated in cancer initiation, tumor growth, and metastasis (Knapinska et al., 2017). Esculetin was found to suppress the expression of MMP-2 and MMP-7 in HCT-116 cells, thereby inhibiting cell migration. In vivo experiments of the HCT-116 subcutaneous tumor-bearing model also demonstrated that esculetin treatment significantly decreased the expression of MMP-2 and MMP-7 (Yan et al., 2019). Moreover, esculetin was shown to suppress the expression of MMP-2 in osteosarcoma LM8 cells (Kimura and Sumiyoshi, 2015). Qu and colleagues observed that fraxetin inhibited the invasion and migration of human glioblastoma U251 cells in vitro and downregulated the expression levels of MMP-2 and MMP-9 (Qu et al., 2021) (Table 3).

Table 3. Suppression of cancer cell invasion and migration.
The phosphoinositide 3-kinase (PI3K)/protein kinase B (Akt) signaling pathway is one of the most frequently overactivated intracellular pathways in several human cancers (Rascio et al., 2021). The PI3K/Akt pathway plays a multitude of roles in the onset and progression of cancer, including the promotion of cancer cell proliferation and migration as well as the prevention of apoptosis. One study reported that application of esculin to human glioblastoma T98G cells caused apoptosis in approximately 15% of the cells. It was further suggested that this apoptosis-inducing effect may be associated with an increase in the expression of caspase-3 and a decrease in the expression of PI3K (Sumorek-Wiadro et al., 2020). In gastric cancer MGC-803 cells, esculetin was found to promote apoptosis by downregulating the PI3K/Akt signaling pathway. Furthermore, the inhibitory effect of esculetin on the PI3K/Akt pathway was also observed in a nude mouse model subcutaneously inoculated with MGC-803 cells (Wang Y. et al., 2017). Ma et al. reported that fraxetin significantly inhibited the viability, proliferation, migration, and invasion of prostate cancer DU145 cells and induced cell apoptosis in a concentration-dependent manner (Ma et al., 2022). Moreover, the expression of polo-like kinase 4 (PLK4), p-PI3K, and p-Akt was found to be decreased by fraxetin treatment. The researchers postulated that fraxetin may act as a tumor suppressor in prostate cancer by inhibiting PLK4 expression, which in turn inactivates the PI3K/Akt signaling pathway.
Mitogen-activated protein kinase (MAPK) cascades represent a fundamental signaling pathway that regulates a multitude of cellular processes, including proliferation, differentiation, apoptosis, and stress responses (Guo et al., 2020). Extracellular signal-regulated kinase 1/2 (ERK1/2) is a member of the MAPK family, which plays a pivotal role in tumor proliferation, invasion, and metastasis (Ali et al., 2022). The activation of the MAPK/ERK pathway has been demonstrated to promote proliferation and have an anti-apoptotic effect (Bahar et al., 2023). Wang et al. demonstrated that esculetin inhibited the proliferation of human leukemia HL-60 cells in a dose-dependent manner, inducing apoptosis and autophagy in HL-60 cells (Wang et al., 2019). Moreover, esculetin was observed to block the ERK signaling pathway in HL-60 cells in a concentration-dependent manner. Another study demonstrated that esculetin inhibited the cell viability of human leukemia U937 cells by inducing apoptosis and selectively inhibiting the phosphorylation of ERK (Lin et al., 2009). Fraxetin was shown to inhibit the proliferation of MCF-7 breast cancer cells in a dose- and time-dependent manner, as well as induce cell cycle arrest at the G0/G1 phase. Furthermore, it was demonstrated that fraxetin treatment resulted in a reduction in the expression of p-ERK1/2 in MCF-7 cells, indicating that fraxetin may exert an anti-tumor effect through the MAPK/ERK1/2 signaling pathway (Huo et al., 2013).
The signal transducer and activator of transcription 3 (STAT3) pathway is intimately linked to the tumor microenvironment, tumor growth, and metastasis. Excessive STAT3 activation in cancer cells and the tumor microenvironment can be viewed as a neoplastic mimic of an inflammation-driven repair response that collectively drives tumor progression (Huynh et al., 2019). It was demonstrated that esculetin significantly inhibited the proliferation, migration, and invasion of laryngeal cancer Hep-2 cells and significantly inhibited the phosphorylation of Janus kinase 1 (JAK1), JAK2, and STAT3 in Hep-2 cells. Furthermore, esculetin treatment was observed to reduce tumor growth and tumor weight in a dose-dependent manner in laryngeal cancer xenograft mice (Zhang G. et al., 2019). The potential mechanism of action may be attributed to its inhibition of the JAK/STAT3 signaling pathway. Qu et al. found that fraxetin inhibited the proliferation, invasion, and migration of human glioblastoma U251 cells in vitro and significantly reduced tumor volume and weight in vivo. Moreover, fraxetin treatment resulted in a reduction in the expression of p-JAK2 and p-STAT3 in both U251 cells and glioma xenograft mice (Qu et al., 2021). In human pancreatic cancer cell lines (PANC-1 and Patu8988), fraxetin was demonstrated to inhibit cell proliferation and induce mitochondrial-dependent apoptosis. It was demonstrated that fraxetin prevents the formation of STAT3 homodimers through intimate contact with the SH2 domain of STAT3. Thus, fraxetin inhibited STAT3 phosphorylation and blocked the activation of downstream STAT3 signaling pathways (Guo et al., 2021).
The Wnt/β-catenin signaling pathway has been extensively implicated in the pathogenesis of cancers. Aberrant activation of the Wnt/β-catenin signaling pathway is closely associated with an increased prevalence of cancer, the advancement of malignant progression, the development of a poor prognosis, and even an increase in cancer-related mortality (Yu et al., 2021). Esculetin has been demonstrated to suppress the activity of the Wnt/β-catenin signaling pathway in human colorectal cancer cells (HCT116 and SW480), thereby exerting anti-proliferative effects (Kim et al., 2018). Fan et al. reported that esculetin dose- and time-dependently reduced cell viability and decreased the mRNA and protein levels of β-catenin in human hepatocellular carcinoma SMMC-7721 cell lines (Fan et al., 2017). These findings suggest that esculetin may play an anti-tumor role by inhibiting the Wnt/β-catenin signaling pathway (Table 4).
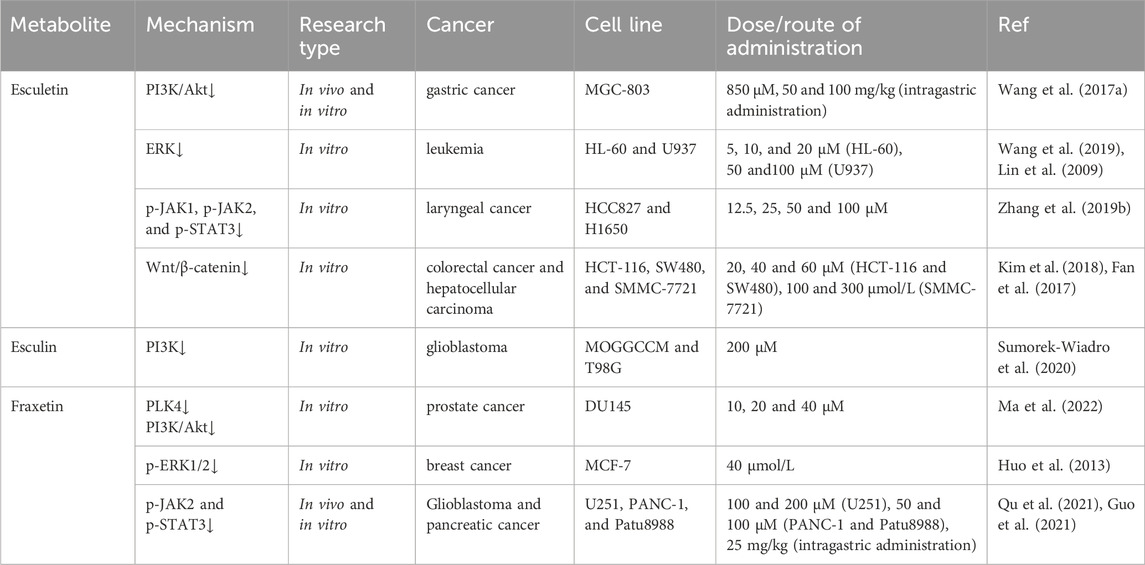
Table 4. Regulation of tumor-associated intracellular signaling pathways.
In the current review, it has been demonstrated that the active metabolites in Cortex Fraxini, such as esculin, esculetin, and fraxetin, have the capacity to inhibit the growth of many different types of cancer, including osteosarcoma, leukemia, colon cancer, prostate cancer, breast cancer, glioblastoma, gastric cancer, lung cancer, hepatocellular carcinoma, and oral squamous cell carcinoma (Figure 2). These active metabolites have been shown to inhibit tumor cell growth through induction of cell cycle arrest, upregulation of p53, p21, and p27, and inhibition of Ki-67 and PCNA expression. Furthermore, it was proposed that they could promote apoptosis in tumor cells by inducing the mitochondrial apoptotic pathway, regulating the Bcl-2 family of proteins, and inducing the production of ROS. Additionally, they were demonstrated to inhibit tumor cell invasion and migration by modulating EMT and inhibiting MMP expression. Furthermore, these active metabolites could regulate several tumor-associated signaling pathways, including the PI3K/Akt, MAPK/ERK, JAK/STAT3, and Wnt/β-catenin pathways. In summary, a large number of studies have been conducted with the aim of elucidating the anti-tumor pharmacological effects and mechanisms of Cortex Fraxini and its active metabolites, including esculin, esculetin, and fraxetin. In view of their pro-apoptotic and anti-proliferative properties in vitro and in vivo, Cortex Fraxini and its active metabolites can be considered potential candidates for the treatment of tumors.
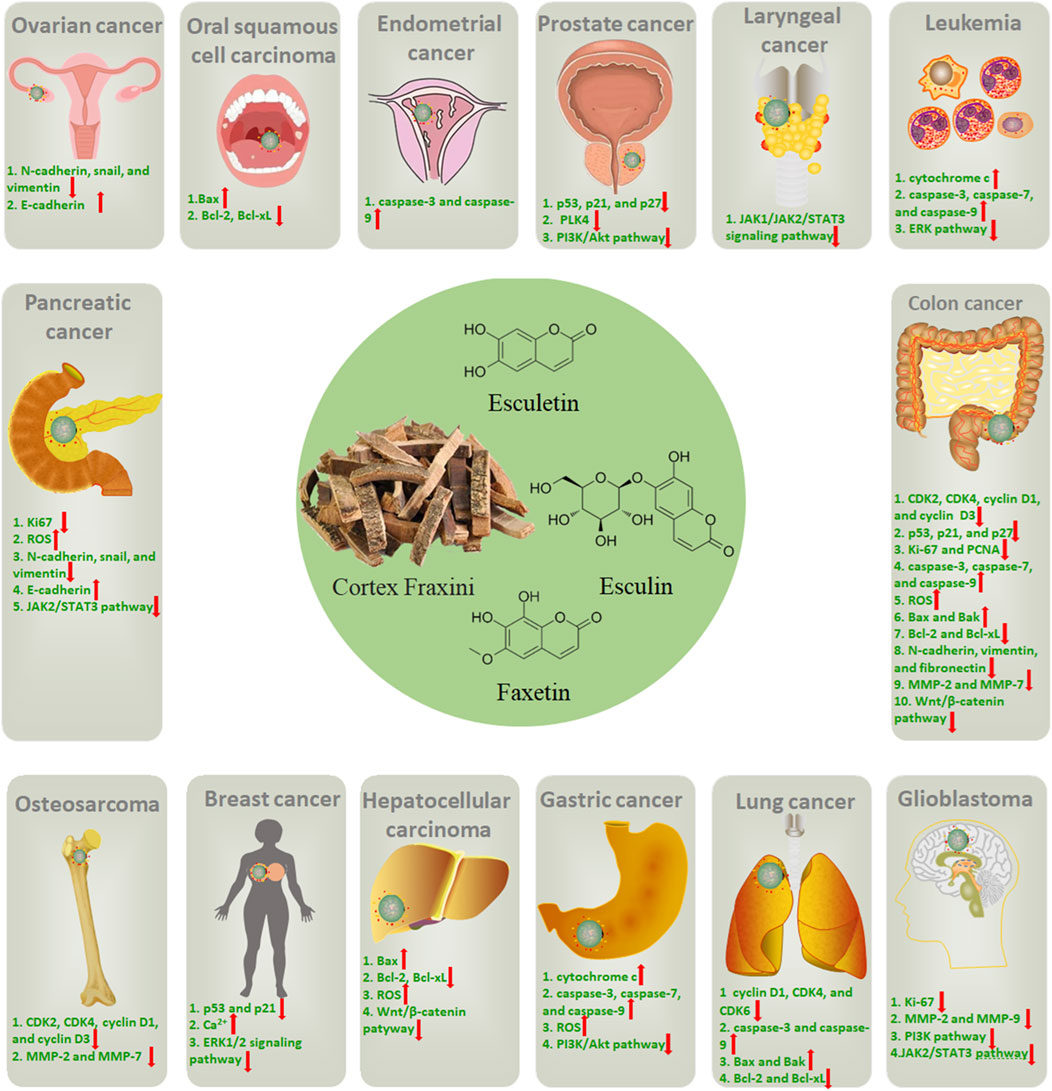
Figure 2. The potential roles of Cortex Fraxini in different types of cancer.
Although numerous studies have been conducted to investigate the anti-tumor pharmacological effects of the active metabolites of Cortex Fraxini, there are still some deficiencies that require further research and improvement in the future. Firstly, given that many of the anti-tumor effects of Cortex Fraxini are only phenotypic variations, in-depth research to explore the complex mechanistic aspects should be a priority. Secondly, the majority of studies to date have employed in vitro experiments as the primary methodology, with a paucity of further proof from in vivo studies. Therefore, the utilization of tumor animal models should be enhanced in future studies to verify the aforementioned anti-tumor mechanisms. Thirdly, although coumarins are the primary active metabolites of Cortex Fraxini, the roles of its other metabolites, including lignans, secoiridoids, phenylethanol glycosides, flavonoids, and triterpenoids, in its anti-tumor activities require further elucidation. Fourthly, how to improve the bioavailability of the metabolites is also a future research direction. To address this challenge, some researchers have constructed a nanostructured lipid carrier loaded with esculetin, which improves its oral bioavailability by 1.7 times, thus enhancing its pharmacological role (Shi et al., 2023). Another study developed an esculetin-loaded mixed micellar delivery system that specifically increased the biological availability of esculetin by 3.06 times (Li et al., 2023). Fifthly, according to existing reports, the active metabolites of Cortex Fraxini have inhibitory effects on many types of tumors, but whether their anti-tumor activity differs among different tumors deserves further investigation.
BC: Conceptualization, Funding acquisition, Writing–original draft, Writing–review and editing. TC: Funding acquisition, Writing–original draft. ZF: Formal Analysis, Supervision, Writing–review and editing. HZ: Conceptualization, Writing–review and editing.
The author(s) declare that financial support was received for the research, authorship, and/or publication of this article. This work was supported by National Natural Science Foundation of China (No. 82205110), Natural Science Foundation of Jiangsu Province (No. BK20210067), Top Talent Support Program for Young and Middle-aged People of Wuxi Health Committee (No. BJ2023066).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Akt, protein kinase B; Bak, Bcl-2 antagonist/killer; Bax, Bcl-2-associated X protein; Bcl-2, B cell lymphoma-2; Bcl-xL, B-cell lymphoma-extra large; CDK, cyclin-dependent kinase; EMT, epithelial-to-mesenchymal transition; ERK, extracellular signal-regulated kinase; JAK, Janus kinase; MAPK, mitogen-activated protein kinase; MEK, mitogen-activated protein kinase; MMP, matrix metalloproteinase; PCNA, proliferating cell nuclear antigen; PI3K, phosphoinositide 3-kinase; PLK4, polo-like kinase 4; Ras, rat sarcoma; ROS, reactive oxygen species; STAT3, signal transducer and activator of transcription 3; TYK2, tyrosine kinase 2.
Ali, E. S., Akter, S., Ramproshad, S., Mondal, B., Riaz, T. A., Islam, M. T., et al. (2022). Targeting Ras-ERK cascade by bioactive natural products for potential treatment of cancer: an updated overview. Cancer. Cell. Int. 22, 246. doi:10.1186/s12935-022-02666-z
Bahar, M. E., Kim, H. J., and Kim, D. R. (2023). Targeting the RAS/RAF/MAPK pathway for cancer therapy: from mechanism to clinical studies. Signal. Transduct. Target. Ther. 8, 455. doi:10.1038/s41392-023-01705-z
Chang, H. T., Chou, C. T., Lin, Y. S., Shieh, P., Kuo, D. H., Jan, C. R., et al. (2016). Esculetin, a natural coumarin compound, evokes Ca(2+) movement and activation of Ca(2+)-associated mitochondrial apoptotic pathways that involved cell cycle arrest in ZR-75-1 human breast cancer cells. Tumour. Biol. 37, 4665–4678. doi:10.1007/s13277-015-4286-1
Choi, Y. J., Lee, C. M., Park, S. H., and Nam, M. J. (2019). Esculetin induces cell cycle arrest and apoptosis in human colon cancer LoVo cells. Environ. Toxicol. 34, 1129–1136. doi:10.1002/tox.22815
Davey, M. G., Hynes, S. O., Kerin, M. J., Miller, N., and Lowery, A. J. (2021). Ki-67 as a prognostic biomarker in invasive breast cancer. Cancers. (Basel) 13, 4455. doi:10.3390/cancers13174455
Ding, L., Cao, J., Lin, W., Chen, H., Xiong, X., Ao, H., et al. (2020). The roles of cyclin-dependent kinases in cell-cycle progression and therapeutic strategies in human breast cancer. Int. J. Mol. Sci. 21, 1960. doi:10.3390/ijms21061960
Edlich, F. (2018). BCL-2 proteins and apoptosis: recent insights and unknowns. Biochem. Biophys. Res. Commun. 500, 26–34. doi:10.1016/j.bbrc.2017.06.190
Eskandari, E., and Eaves, C. J. (2022). Paradoxical roles of caspase-3 in regulating cell survival, proliferation, and tumorigenesis. J. Cell. Biol. 221, e202201159. doi:10.1083/jcb.202201159
Fan, X., Du, H., Sun, Y., Jiang, J., Wang, Z., Yin, W., et al. (2017). Suppression of the Wnt signaling pathway may contribute to the inhibition of proliferation of human hepatocellular carcinoma SMMC-7721 cells by esculetin. Oncol. Lett. 14, 1731–1736. doi:10.3892/ol.2017.6364
Fuste, N. P., Ferrezuelo, F., and Gari, E. (2016). Cyclin D1 promotes tumor cell invasion and metastasis by cytoplasmic mechanisms. Mol. Cell. Oncol. 3, e1203471. doi:10.1080/23723556.2016.1203471
Guo, Y., Xiao, Y., Guo, H., Zhu, H., Chen, D., Wang, J., et al. (2021). The anti-dysenteric drug fraxetin enhances anti-tumor efficacy of gemcitabine and suppresses pancreatic cancer development by antagonizing STAT3 activation. Aging. (Albany NY) 13, 18545–18563. doi:10.18632/aging.203301
Guo, Y. J., Pan, W. W., Liu, S. B., Shen, Z. F., Xu, Y., and Hu, L. L. (2020). ERK/MAPK signalling pathway and tumorigenesis. Exp. Ther. Med. 19, 1997–2007. doi:10.3892/etm.2020.8454
Huo, H. N., Xie, K. P., Wang, L. M., Jiang, Y., Zou, W., and Xie, M. J. (2013). Relationship between the inhibitory effect of fraxetin on breast cancer and estrogen signaling pathway. Sheng. Li. Xue. Bao. 65, 323–328.
Huynh, J., Chand, A., Gough, D., and Ernst, M. (2019). Therapeutically exploiting STAT3 activity in cancer - using tissue repair as a road map. Nat. Rev. Cancer. 19, 82–96. doi:10.1038/s41568-018-0090-8
Jassim, M. M. A., Rasool, K. H., and Mahmood, M. M. (2021). p53, p21, and cyclin d1 protein expression patterns in patients with breast cancer. Vet. World. 14, 2833–2838. doi:10.14202/vetworld.2021.2833-2838
Jeon, Y. J., Cho, J. H., Lee, S. Y., Choi, Y. H., Park, H., Jung, S., et al. (2016). Esculetin induces apoptosis through EGFR/PI3K/akt signaling pathway and nucleophosmin relocalization. J. Cell. Biochem. 117, 1210–1221. doi:10.1002/jcb.25404
Kim, A. D., Han, X., Piao, M. J., Hewage, S. R., Hyun, C. L., Cho, S. J., et al. (2015). Esculetin induces death of human colon cancer cells via the reactive oxygen species-mediated mitochondrial apoptosis pathway. Environ. Toxicol. Pharmacol. 39, 982–989. doi:10.1016/j.etap.2015.03.003
Kim, W. K., Byun, W. S., Chung, H. J., Oh, J., Park, H. J., Choi, J. S., et al. (2018). Esculetin suppresses tumor growth and metastasis by targeting Axin2/E-cadherin axis in colorectal cancer. Biochem. Pharmacol. 152, 71–83. doi:10.1016/j.bcp.2018.03.009
Kimura, Y., and Sumiyoshi, M. (2015). Antitumor and antimetastatic actions of dihydroxycoumarins (esculetin or fraxetin) through the inhibition of M2 macrophage differentiation in tumor-associated macrophages and/or G1 arrest in tumor cells. Eur. J. Pharmacol. 746, 115–125. doi:10.1016/j.ejphar.2014.10.048
Knapinska, A. M., Estrada, C. A., and Fields, G. B. (2017). The roles of matrix metalloproteinases in pancreatic cancer. Prog. Mol. Biol. Transl. Sci. 148, 339–354. doi:10.1016/bs.pmbts.2017.03.004
Kuida, K. (2000). Caspase-9. Int. J. Biochem. Cell. Biol. 32, 121–124. doi:10.1016/s1357-2725(99)00024-2
Kwak, J. H., Kim, Y., Staatz, C. E., and Baek, I. H. (2021). Oral bioavailability and pharmacokinetics of esculetin following intravenous and oral administration in rats. Xenobiotica 51, 811–817. doi:10.1080/00498254.2021.1925774
Lee, M., Yang, C., Park, S., Song, G., and Lim, W. (2022). Fraxetin induces cell death in colon cancer cells via mitochondria dysfunction and enhances therapeutic effects in 5-fluorouracil resistant cells. J. Cell. Biochem. 123, 469–480. doi:10.1002/jcb.30187
Lee, S. Y., Lim, T. G., Chen, H., Jung, S. K., Lee, H. J., Lee, M. H., et al. (2013). Esculetin suppresses proliferation of human colon cancer cells by directly targeting β-catenin. Cancer. Prev. Res. (Phila). 6, 1356–1364. doi:10.1158/1940-6207.CAPR-13-0241
Li, C. X., Li, J. C., Lai, J., and Liu, Y. (2022a). The pharmacological and pharmacokinetic properties of esculin: a comprehensive review. Phytother. Res. 36, 2434–2448. doi:10.1002/ptr.7470
Li, J. J., Zhou, S. Y., Zhang, H., Lam, K. H., Lee, S. M., Yu, P. H., et al. (2015). Cortex fraxini (qingpi) protects rat pheochromocytoma cells against 6-hydroxydopamine-induced apoptosis. Park. Dis. 2015, 532849. doi:10.1155/2015/532849
Li, X., Xia, X., Zhang, J., Adu-Frimpong, M., Shen, X., Yin, W., et al. (2023). Preparation, physical characterization, pharmacokinetics and anti-hyperglycemic activity of esculetin-loaded mixed micelles. J. Pharm. Sci. 112, 148–157. doi:10.1016/j.xphs.2022.06.022
Li, Y., Guo, H., Wu, Y., Geng, Q., Dong, D., Wu, H., et al. (2013). A sensitive and selective method for determination of aesculin in cortex fraxini by liquid chromatography quadrupole time-of-flight tandem mass spectrometry and application in pharmacokinetic study. J. Anal. Methods. Chem. 2013, 432465. doi:10.1155/2013/432465
Li, Z. Y., Li, X. K., Lin, Y., Feng, N., Zhang, X. Z., Li, Q. L., et al. (2022b). A comparative study of three chemometrics methods combined with excitation-emission matrix fluorescence for quantification of the bioactive compounds aesculin and aesculetin in Cortex Fraxini. Front. Chem. 10, 984010. doi:10.3389/fchem.2022.984010
Lin, T. H., Lu, F. J., Yin, Y. F., and Tseng, T. H. (2009). Enhancement of esculetin on arsenic trioxide-provoked apoptosis in human leukemia U937 cells. Chem. Biol. Interact. 180, 61–68. doi:10.1016/j.cbi.2009.01.011
Louis, D. M., Nair, L. M., Vallonthaiel, A. G., Narmadha, M. P., and Vijaykumar, D. K. (2023). Ki 67: a promising prognostic marker in early breast cancer-a review article. Indian. J. Surg. Oncol. 14, 122–127. doi:10.1007/s13193-022-01631-6
Ma, Z., Sun, Y., and Peng, W. (2022). Fraxetin down-regulates polo-like kinase 4 (PLK4) to inhibit proliferation, migration and invasion of prostate cancer cells through the phosphatidylinositol 3-kinase (PI3K)/protein kinase B (Akt) pathway. Bioengineered 13, 9345–9356. doi:10.1080/21655979.2022.2054195
Mo, M., Chen, M. J., Huang, Y., Jiang, W., Qin, Q. H., Liang, Z. J., et al. (2020). Esculin inhibits proliferation of triple negative breast cancer cells by down-regulating FBI-1. Zhonghua. Zhong. Liu. Za. Zhi 42, 629–634. doi:10.3760/cma.j.cn112152-20191001-00642
Pan, H., Wang, B. H., Lv, W., Jiang, Y., and He, L. (2015). Esculetin induces apoptosis in human gastric cancer cells through a cyclophilin D-mediated mitochondrial permeability transition pore associated with ROS. Chem. Biol. Interact. 242, 51–60. doi:10.1016/j.cbi.2015.09.015
Qian, S., Wei, Z., Yang, W., Huang, J., Yang, Y., and Wang, J. (2022). The role of BCL-2 family proteins in regulating apoptosis and cancer therapy. Front. Oncol. 12, 985363. doi:10.3389/fonc.2022.985363
Qu, L., Lin, P., Lin, M., Ye, S., Papa Akuetteh, P. D., and Youyou, Z. (2021). Fraxetin inhibits the proliferation and metastasis of glioma cells by inactivating JAK2/STAT3 signaling. Evid. Based. Complement. Altern. Med. 2021, 1–10. doi:10.1155/2021/5540139
Rascio, F., Spadaccino, F., Rocchetti, M. T., Castellano, G., Stallone, G., Netti, G. S., et al. (2021). The pathogenic role of PI3K/AKT pathway in cancer onset and drug resistance: an updated review. Cancers. (Basel). 13, 3949. doi:10.3390/cancers13163949
Razavipour, S. F., Harikumar, K. B., and Slingerland, J. M. (2020). p27 as a transcriptional regulator: new roles in development and cancer. Cancer. Res. 80, 3451–3458. doi:10.1158/0008-5472.CAN-19-3663
Redza-Dutordoir, M., and Averill-Bates, D. A. (2016). Activation of apoptosis signalling pathways by reactive oxygen species. Biochim. Biophys. Acta. 1863, 2977–2992. doi:10.1016/j.bbamcr.2016.09.012
Rehman, S. U., Kim, I. S., Kang, K. S., and Yoo, H. H. (2015). HPLC determination of esculin and esculetin in rat plasma for pharmacokinetic studies. J. Chromatogr. Sci. 53, 1322–1327. doi:10.1093/chromsci/bmv014
Shi, F., Yin, W., Adu-Frimpong, M., Li, X., Xia, X., Sun, W., et al. (2023). In-vitro and in-vivo evaluation and anti-colitis activity of esculetin-loaded nanostructured lipid carrier decorated with DSPE-MPEG2000. J. Microencapsul. 40, 442–455. doi:10.1080/02652048.2023.2215345
Song, J., Ham, J., Hong, T., Song, G., and Lim, W. (2021). Fraxetin suppresses cell proliferation and induces apoptosis through mitochondria dysfunction in human hepatocellular carcinoma cell lines Huh7 and Hep3B. Pharmaceutics 13, 112. doi:10.3390/pharmaceutics13010112
Soussi, T. (2000). The p53 tumor suppressor gene: from molecular biology to clinical investigation. Ann. N. Y. Acad. Sci. 910, 121–137. doi:10.1111/j.1749-6632.2000.tb06705.x
Strzalka, W., and Ziemienowicz, A. (2011). Proliferating cell nuclear antigen (PCNA): a key factor in DNA replication and cell cycle regulation. Ann. Bot. 107, 1127–1140. doi:10.1093/aob/mcq243
Sumorek-Wiadro, J., Zajac, A., Langner, E., Skalicka-Wozniak, K., Maciejczyk, A., Rzeski, W., et al. (2020). Antiglioma potential of coumarins combined with sorafenib. Molecules 25, 5192. doi:10.3390/molecules25215192
Turkekul, K., Colpan, R. D., Baykul, T., Ozdemir, M. D., and Erdogan, S. (2018). Esculetin inhibits the survival of human prostate cancer cells by inducing apoptosis and arresting the cell cycle. J. Cancer. Prev. 23, 10–17. doi:10.15430/JCP.2018.23.1.10
Wang, G., Lu, M., Yao, Y., Wang, J., and Li, J. (2017a). Esculetin exerts antitumor effect on human gastric cancer cells through IGF-1/PI3K/Akt signaling pathway. Eur. J. Pharmacol. 814, 207–215. doi:10.1016/j.ejphar.2017.08.025
Wang, J. (2015). [Esculin induces A549 lung cancer cell apoptosis by inhibiting Ras/ERK pathway]. Chinese. Traditional. Pat. Med. 37, 40–43.
Wang, J., Lu, M. L., Dai, H. L., Zhang, S. P., Wang, H. X., and Wei, N. (2015). Esculetin, a coumarin derivative, exerts in vitro and in vivo antiproliferative activity against hepatocellular carcinoma by initiating a mitochondrial-dependent apoptosis pathway. Braz. J. Med. Biol. Res. 48, 245–253. doi:10.1590/1414-431X20144074
Wang, X., Yang, C., Zhang, Q., Wang, C., Zhou, X., Zhang, X., et al. (2019). In vitro anticancer effects of esculetin against human leukemia cell lines involves apoptotic cell death, autophagy, G0/G1 cell cycle arrest and modulation of Raf/MEK/ERK signalling pathway. J. BUON. 24, 1686–1691.
Wang, Y., Zhao, M., Ou, Y., Zeng, B., Lou, X., Wang, M., et al. (2016). Metabolic profile of esculin in rats by ultra high performance liquid chromatography combined with Fourier transform ion cyclotron resonance mass spectrometry. J. Chromatogr. B. Anal. Technol. Biomed. Life. Sci. 1020, 120–128. doi:10.1016/j.jchromb.2016.03.027
Wang, Y., Zhao, M., Ye, H., Shao, Y., Yu, Y., Wang, M., et al. (2017b). Comparative pharmacokinetic study of the main components of cortex fraxini after oral administration in normal and hyperuricemic rats. Biomed. Chromatogr. 31. doi:10.1002/bmc.3934
Wang, Y. L., Wu, W. R., Lin, P. L., Shen, Y. C., Lin, Y. Z., Li, H. W., et al. (2022). The functions of PCNA in tumor stemness and invasion. Int. J. Mol. Sci. 23, 5679. doi:10.3390/ijms23105679
Warren, C. F. A., Wong-Brown, M. W., and Bowden, N. A. (2019). BCL-2 family isoforms in apoptosis and cancer. Cell. death. Dis. 10, 177. doi:10.1038/s41419-019-1407-6
Wróblewska-Łuczka, P., Grabarska, A., Góralczyk, A., Marzęda, P., and Łuszczki, J. J. (2022). Fraxetin interacts additively with cisplatin and mitoxantrone, antagonistically with docetaxel in various human melanoma cell lines-an isobolographic analysis. Int. J. Mol. Sci. 24, 212. doi:10.3390/ijms24010212
Wu, C., Huang, M., Lin, Y., Ju, H., and Ching, H. (2007). Antioxidant properties of Cortex Fraxini and its simple coumarins. Food. Chem. 104, 1464–1471. doi:10.1016/j.foodchem.2007.02.023
Xu, H., Zhang, J., Wang, Q., Li, Y., and Zhang, B. (2020). Fraxetin inhibits the proliferation of RL95-2 cells through regulation of metabolism. Int. J. Clin. Exp. Pathol. 13, 1500–1505.
Xu, R., Ruan, Y., Zhang, L., Gu, Y., and Liu, M. (2023). Fraxetin suppresses the proliferation, migration, and invasion of ovarian cancer cells by inhibiting the TLR4/STAT3 signaling pathway. Immunopharmacol. Immunotoxicol. 45, 287–294. doi:10.1080/08923973.2022.2141643
Yan, L., Yu, H. H., Liu, Y. S., Wang, Y. S., and Zhao, W. H. (2019). Esculetin enhances the inhibitory effect of 5-Fluorouracil on the proliferation, migration and epithelial-mesenchymal transition of colorectal cancer. Cancer. Biomark. 24, 231–240. doi:10.3233/CBM-181764
Yu, F., Yu, C., Li, F., Zuo, Y., Wang, Y., Yao, L., et al. (2021). Wnt/β-catenin signaling in cancers and targeted therapies. Signal. Transduct. Target. Ther. 6, 307. doi:10.1038/s41392-021-00701-5
Zhang, G., Xu, Y., and Zhou, H. F. (2019a). Esculetin inhibits proliferation, invasion, and migration of laryngeal cancer in vitro and in vivo by inhibiting Janus kinas (JAK)-Signal transducer and activator of transcription-3 (STAT3) activation. Med. Sci. Monit. 25, 7853–7863. doi:10.12659/MSM.916246
Zhang, L., Xie, Q., and Li, X. (2022). Esculetin: a review of its pharmacology and pharmacokinetics. Phytother. Res. 36, 279–298. doi:10.1002/ptr.7311
Zhang, Y., Wang, L., Deng, Y., Zhao, P., Deng, W., Zhang, J., et al. (2019b). Fraxetin suppresses proliferation of non-small-cell lung cancer cells via preventing activation of signal transducer and activator of transcription 3. Tohoku. J. Exp. Med. 248, 3–12. doi:10.1620/tjem.248.3
Zhang, Y., and Weinberg, R. A. (2018). Epithelial-to-mesenchymal transition in cancer: complexity and opportunities. Front. Med. 12, 361–373. doi:10.1007/s11684-018-0656-6
Zheng, Z., Sun, C., Zhong, Y., Shi, Y., Zhuang, L., Liu, B., et al. (2024). Fraxini cortex: progresses in phytochemistry, pharmacology and ethnomedicinal uses. J. Ethnopharmacol. 325, 117849. doi:10.1016/j.jep.2024.117849
Zhou, L., Kang, J., Fan, L., Ma, X. C., Zhao, H. Y., Han, J., et al. (2008). Simultaneous analysis of coumarins and secoiridoids in Cortex Fraxini by high-performance liquid chromatography-diode array detection-electrospray ionization tandem mass spectrometry. J. Pharm. Biomed. Anal. 47, 39–46. doi:10.1016/j.jpba.2007.12.009
Keywords: Cortex Fraxini, anti-tumor, esculin, esculetin, fraxetin
Citation: Cai B, Cai T, Feng Z and Zhu H (2024) The possible anti-tumor actions and mechanisms of active metabolites from Cortex Fraxini. Front. Pharmacol. 15:1404172. doi: 10.3389/fphar.2024.1404172
Received: 20 March 2024; Accepted: 02 September 2024;
Published: 13 September 2024.
Edited by:
Mohammad Reza Khazdair, Birjand University of Medical Sciences, IranReviewed by:
Manel Ben Hammouda, Duke University, United StatesCopyright © 2024 Cai, Cai, Feng and Zhu. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Huanhuan Zhu, ZnN5eTAxMTU5QG5qdWNtLmVkdS5jbg==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.