
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Pharmacol., 25 April 2024
Sec. Neuropharmacology
Volume 15 - 2024 | https://doi.org/10.3389/fphar.2024.1364630
This article is part of the Research TopicMedicinal Drugs, the Effect on Memories and BeyondView all 7 articles
 Valeska Cid-Jofré1
Valeska Cid-Jofré1 Tamara Bahamondes1Agustina Zúñiga Correa1Ivalú Ahumada Arias1
Tamara Bahamondes1Agustina Zúñiga Correa1Ivalú Ahumada Arias1 Miguel Reyes-Parada1,2*
Miguel Reyes-Parada1,2* Georgina M. Renard1*
Georgina M. Renard1*Mounting evidence from animal models and human studies indicates that psychostimulants can significantly affect social behaviors. This is not surprising considering that the neural circuits underlying the regulation and expression of social behaviors are highly overlapped with those targeted by psychostimulants, which in most cases have strong rewarding and, consequently, addictive properties. In the present work, we provide an overview regarding the effects of illicit and prescription psychostimulants, such as cocaine, amphetamine-type stimulants, methylphenidate or modafinil, upon social behaviors such as social play, maternal behavior, aggression, pair bonding and social cognition and how psychostimulants in both animals and humans alter them. Finally, we discuss why these effects can vary depending on numerous variables such as the type of drug considered, acute versus long-term use, clinical versus recreational consumption, or the presence or absence of concomitant risk factors.
The relationship between our species and drugs is ancient (de Wit and Richards, 2004) and the reasons for drug use are diverse: medical purposes, sociability, relaxation, focus and work, and awakening properties, among others. In the brain there are specific nuclei that organize, activate, and modulate the procurement of natural rewards such as food, social interactions, and sex. Some of these nuclei include the ventral tegmental area (VTA), nucleus accumbens (NAc), prefrontal cortex (PFC), lateral septum (LS) and ventral pallidum (VP), and they are part of the “reward circuitry” or mesocorticolimbic system (Hyman et al., 2006; Koob and Volkow, 2010). Noteworthy, these brain nuclei are not only “activated” by natural but also by other stimuli such as drugs of abuse (Volkow and Morales, 2015). This activation elicits a strong relationship between the effect of the stimulus (i.e., euphoria, pleasure, better attention, or cognitive performance) and the context where it was performed (Berridge and Kringelbach, 2015). Therefore, there exists a learning process leading the individual to repeat the behavior. It has been suggested that one of the main features of drug addiction (drug abuse) versus recreational use is the seeking of the reward without “liking” it (Berridge and Robinson, 2016), regardless of the adverse outcomes (an impulsive and compulsive intake).
Dopamine (DA) projections from VTA to NAc and other limbic structures participate in two crucial features of the rewarding process: motivated behavior and reinforcement of those behaviors. These characteristics are fundamental for drug addiction (Koob and Volkow, 2016; Volkow et al., 2016).
Psychostimulants like cocaine, amphetamine-type stimulants, methylphenidate (MPH), modafinil (MOD) and new psychoactive substances with stimulant effects are globally used. Estimates made by the United Nations in 2019 (Breckenridge et al., 2019), illustrate that there are 18 million cocaine users worldwide and confiscation of psychostimulants only in the Americas is close to 1,215 tons, being cocaine, methamphetamine (MAMPH) and amphetamine (AMPH) the leading drugs, although the percentage varies among continents.
Psychostimulants are drugs that act on the central nervous system (CNS), increasing alertness, and arousal and causing general behavioral excitement (McCreary et al., 2015). Their primary mechanism of action is to enhance the activity of the three main monoamine neurotransmitters: DA, norepinephrine (NE) and/or serotonin (5-HT), which produces an intense activation of several brain pathways underpinning the aforementioned behaviors (Sofuoglu and Sewell, 2009). Cocaine, AMPH, MPH, MOD, and MAMPH have the DA and NE transporter (DAT and NET, respectively) as main targets, and using differential mechanisms lead to an augmentation of the extracellular levels of DA and NE in the synaptic cleft (Goodwin et al., 2009; Underhill et al., 2020). On the other hand, drugs such as 3,4-methylenedioxymethamphetamine (MDMA or ecstasy), which is not necessarily considered a typical psychostimulant, increase mainly 5-HT levels acting on the corresponding transporter (Dunlap et al., 2018; Nichols, 2022). These diverse pharmacodynamics are likely related to some controversial effects that are discussed below. Since psychostimulants can generate various neurobiological effects, they have numerous clinical uses as well (Faraone, 2018). Psychostimulants are used as pharmacotherapy in neuropsychiatric disorders such as narcolepsy and attention-deficit/hyperactivity disorder (ADHD) (Faraone, 2018), but these drugs have been used for other purposes in non-medical settings and off-label fashion especially in young populations (Arria and DuPont, 2010).
Regarding ADHD, the psychostimulants with the highest number of prescriptions are AMPH and MPH. In addition, MOD is another stimulant that has shown promising results in clinical trials in children with ADHD (Rugino, 2007; Amiri et al., 2008; Goez et al., 2012; Arnold et al., 2014; Faraone, 2018; Caldwell et al., 2020; Zahed et al., 2022). Different studies have reported an over-prescription and illegal use of MOD, MPH and AMPH in young healthy individuals (Lakhan and Kirchgessner, 2012). However, it remains unknown the effects of the illicit use of these psychostimulants after long term (d’Angelo et al., 2017).
Considering our social nature, social interactions are regarded as natural rewards. Interactions with peers are essential for allowing allies to conform, protecting territories or relatives, pair bonding, parental and maternal care of offspring, shared foods, shelter, and more.
Social behaviors can be broadly defined as a modality of communication and/or interaction between two or more individuals of a given species of animals (Chen and Hong, 2018). Humans, as well as other species, depend on social stimuli to make decisions. Thus, for social behavior, individuals use highly complex communication methods, specie-specific sensory cues and dynamic information between conspecifics (Chen and Hong, 2018). These behaviors include a variety of interactions and settings like social group living, social organization, mating, parenting, aggression, social play, and others. It must be considered that different pharmacological interventions may affect social behavior performance and increase or decrease the interactions involved in them.
Studies in rodents using the conditioning place preference (CPP) test have demonstrated the natural tendency to seek social interactions and their reward properties (Douglas et al., 2004; Thiel et al., 2008; Douglas et al., 2018), both in adults and juvenile individuals. Several brain areas have been associated with social behaviors such as PFC, infralimbic cortex (IL), NAc, VTA, hippocampus (Hipp), LS and others (Douglas et al., 2004; Solié et al., 2022; Yan and Rein, 2022). Interestingly, as we mentioned, some of them are also part of the reward circuitry. Therefore, social interactions are regulated by some of the same brain regions activated by drugs of abuse (Figure 1). Given the crucial role of social interaction in normal socio- and neuro-development (O’Connell and Hofmann, 2011; Gunaydin et al., 2014), it is fundamental to unravel how psychostimulants might modify the relationship between reward value and social behaviors. These modifications depend on the length of drug use, dosage, and age of the first use, leading or not to drug addiction (Bardo et al., 2013; Clare et al., 2021).
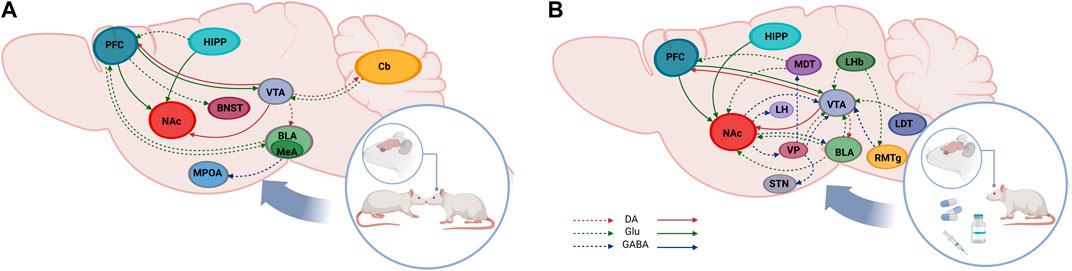
Figure 1. Schematic representation of brain pathways involved in social behavior and reward. Social behaviors are regulated by brain regions and connections that, to some extent, overlap to dose activated by drugs of abuse and natural reinforces (continuous arrows). Sagittal rodent brain section showing the different nuclei of the cortical mesolimbic system and their connections within the social neural circuit (A) and the reward neural circuit (B). Ventral tegmental area (VTA), nucleus accumbens (NAc), prefrontal cortex (PFC), hi ppocampus (HIPP), bed nucleus of the stria terminalis (BNST), medial preoptic area (MPOA), cerebello (Cb), basolateral amygdala (BLA), medial amygdala (MeA), lateral hypothalamus (LH), ventral pallidum (VP), sub thalamic nucleus (STN), rostromedial tegmentum (RMTg) lateral habenula (LHb), laterodorsal tegmentum (LDT), dopamine (DA), glutamate (GLU).
Considering that the neural circuits underlying the regulation and expression of social behaviors are highly overlapped with those targeted by psychostimulants, the aim of the present work is to give a general overview regarding the effects of illicit and prescription psychostimulants upon social behaviors in both animals and humans.
The VTA is a critical brain region involved in social interaction and reward processes. DA neurons from VTA that project to NAc participate in the coding of social and non-social stimuli and this function is modulated by several neurotransmitters and neuropeptides, such as glutamate, GABA, oxytocin (OXT) and vasopressin (AVP), among others.
For instance, when VTA glutamatergic projections to NAc are activated, they stimulate DA release in NAc, and place preference behavior is reinforced. However, these glutamatergic projections can reinforce this behavior independently of DA release (Zell et al., 2020). OXT alters the readiness of the brain to orient to social stimuli via the activation of DA neurons in VTA (Groppe et al., 2013). Moreover, when activation occurs at this level, social interaction generates rewarding experiences by promoting prosocial behaviors (Hung et al., 2017). For example, it has been shown that in male Syrian hamsters there is an increase in VTA neuronal activation during social stimuli. Specifically, the number of c-Fos immunoreactive cells in the VTA of Syrian hamsters after OXT release was higher in males who experienced direct social interaction compared with socially naïve males. This indicates that social experience enhances the response of this area to social stimuli (Hung et al., 2017). Another work done in Syrian hamster demonstrated that activation of the OXT receptor (OXTR), but not the vasopressin type V1a receptor (V1aR) in the VTA is crucial for the encoding of social interactions as reward (Song et al., 2016).
The VP is a GABAergic nucleus with a small population of glutamatergic projections (Soares-Cunha and Heinsbroek, 2023). This nucleus is part of a larger network of limbic brain areas that mediates the salience of several rewards (food, social affiliation, sex, and others) (Soares-Cunha and Heinsbroek, 2023). It also acts as a central point for inputs from the medial amygdala (MeA), VTA, and lateral hypothalamus (LH), among other structures, and the VP projects back to its input sources, including the NAc and VTA for reciprocal information exchange (Ahrens et al., 2018; Zhou et al., 2022).
Considering the multiple connections with areas related to core processes related to reward (motivation, arousal, motor control), VP is seen as a control center that regulates limbic signals and translates them into motor outcomes (Ottenheimer et al., 2020; Soares-Cunha and Heinsbroek, 2023). Regarding motivated behaviors and natural rewards, in a study using cell specific fiber photometry it was shown that inhibition of glutamatergic neurons in VP reduces the animals’ ability to detect salient stimuli, and those neurons got activated in response to several reward related stimuli (of social and non-social nature) (Wang et al., 2020).
Fiber photometry recordings revealed that VTA GABA projection neurons respond to food (reward). This response is relatively invariable (the outcome does not change as animals learn that a cue predicts reward availability) and the extent of this response is completely correlated with the size and deliciousness of the reward. Moreover, VP neurons respond to both reward consumption and seeking–associated motor activity. Additionally, when the VTA-to-VP GABA pathway is chemogenetically stimulated, an increased activity in a portion of VP neurons responsive to reward is observed. Interestingly, this effect was shown only in those neurons related to motivated motor activity but not in those related to reward consumption (Zhou et al., 2022). Additionally, optogenetic stimulation of these neurons enhances the performance of freely moving mice in a cued reward task and maintains a high incentive for reward during a progressive ratio schedule of reinforcement (Zhou et al., 2022).
NAc is a significant structure of the ventral striatum involved in the responses related to reward, motivated and goal-directed behaviors, and drugs of abuse. (Ambroggi et al., 2011; West and Carelli, 2016; Piantadosi et al., 2018). For example, in young animals there is an increase in neuronal activity (c-Fos immunoreactivity) after social play (a motivated young social behavior) (van Kerkhof et al., 2014). Furthermore, GABA antagonists can modify the length of the play (van Kerkhof et al., 2013). In adults, sexual consummatory behavior increases DA release in NAc in males (Sun et al., 2018; Sun et al., 2020). Moreover, a study showed DA release in both naïve and pair-bonded males in response to pups in prairie voles (Lei et al., 2017).
Altogether, these data identify the importance of DA, glutamate, and GABAergic neurons within the VP-VTA-NAc circuit in the salience, learning, and seeking of natural rewards. Furthermore, the neuronal response is in accordance with the salience magnitude of the reward, and this does not change during a cue learning protocol, reflecting the importance of VP-VTA-NAc communication on reward and motivation for both natural and artificial rewards.
The PFC has a complex mix of inhibitory interneurons and receives inputs from neuromodulators such as acetylcholine (ACh), DA, and OXT, being suggested as a top-down control system for decision-making, goal-directed behaviors and complex cognitive processing (Miller and Cohen, 2001; Hoover and Vertes, 2007; Anastasiades and Carter, 2021). Indeed, the PFC projects to several brain areas (NAc, Hipp and amygdala) that are known to influence sociability (Franklin et al., 2017). Lesions of regions within the PFC in rodents have demonstrated its importance in social functioning. For example, the lesion of the rat orbitofrontal cortex increases aggressive behavior (Rudebeck et al., 2007). Furthermore, it has been shown that, in rats, PFC was activated (c-Fos activity) during social interaction (Wall et al., 2012), also showing the involvement of the PFC in social behavior.
It has been proposed that the medial PFC (mPFC) is also pivotal for normal social behavior. A portion of mPFC neurons increases activity when social stimuli (unfamiliar mouse), but not when non-social stimuli (an object) are presented (Levy et al., 2019). In humans, different reports show impairments in mPFC activity and altered social behavior in individuals with autism spectrum disorder (von dem Hagen et al., 2013; Karen Pierce et al., 2004).
In summary, the PFC is important for generating appropriate social responses and controlling execution function, by evaluating and interpreting social information of the moment within the context of previous experiences (Franklin et al., 2017; Levy et al., 2019).
Social play is the first non-mother directed social behavior in young animals. It is characterized by exaggerated and distinct behavioral patterns related to social, sexual, and aggressive behavior (Vanderschuren et al., 2016). It is part of normal social behavior and is considered an indicator of emotional wellbeing, general health, and welfare of animals (Ahloy-Dallaire et al., 2018). This behavior is essential for social, cognitive, emotional, and motor abilities development, having a role in the environment and social context adaptation (Vanderschuren et al., 1997; Vanderschuren et al., 2016). Social play behavior is a highly pleasurable rewarding activity, especially in young individuals (Trezza et al., 2010) across species. This behavior starts around weaning and continues until early adolescence (Vanderschuren et al., 2016), where the type of response in the play depends on age and sex (Pellis and Pellis, 1987; Pellis and Pellis, 1990).
In rodents, social play is also known as ‘rough and tumble play or play fighting’ and it is characterized by highly physical and vigorous movement patterns between peers. In the context of juvenile rats, social play behavior can be divided into two basic components such as pouncing (interaction in which an animal nose rubs the partner’s nape “asking for play”) and pinning (interaction in which an animal stands over the ventral surface of another animal “wanting to play”), along with grooming, chasing, and boxing (Homberg et al., 2007; Argue and McCarthy, 2015a; Argue and McCarthy, 2015b). Furthermore, in rodents this type of behavior is accompanied by physical, facial, and vocal (ultrasonic vocalization) signals with ludic intention (Vanderschuren et al., 2016).
Thus, social play generates emotional excitement and bonding having a neurobiological function in development. Some studies have shown how animals with play deprivation generate higher levels of anxiety (Lukkes et al., 2009; Vanderschuren et al., 2016) and hence higher sensitivity to drugs (Whitaker et al., 2013; Lesscher et al., 2015; Heilig et al., 2016). Play is sensitive to several factors including motivational state, isolation periods and housing conditions. For example, in young Sprague Dawley rats, different housing conditions produce high or low levels of playing, and isolation usually diminishes general social interactions in young and adult rats regardless of sex (Varlinskaya and Spear, 2008).
Psychostimulants can profoundly alter social play behavior due to their effects on brain mechanisms involved in reward (Trezza et al., 2014; Knell, 2022). For instance, AMPH, MPH, and cocaine suppress social play. It has been shown that AMPH and MPH have a suppressive effect on social play through the stimulation of α2 adrenoceptors but not through DA receptors (Achterberg et al., 2014). The inhibitory effects of MPH on social play are mediated by a distributed network of prefrontal and limbic subcortical regions involved in cognitive control and emotional processes. Thus, microinjections of MPH in the infralimbic cortex, anterior cingulate cortex, basolateral amygdala and habenula produce a decrease in social play behavior (Achterberg et al., 2015). The motivation for social play is mediated by DA receptors stimulation and the suppression in the expression of play is mediated by stimulation of adrenergic α2 receptors, suggesting that the decrease in social play behavior promoted by MPH is regulated by a network that could work in parallel (Achterberg et al., 2016). Besides, it has been shown that chronic treatment with MOD in pre-adolescent rats, changes dopaminergic neurotransmission in the NAc decreasing DA release induced by a depolarizing stimulus, and suggesting a decrease in the ability of rats to perceive the rewarding effects of social play (Cid-Jofre et al., 2021).
In addition, cocaine, MPH and AMPH have demonstrated to impair social play behavior when are administrated acutely (Vanderschuren et al., 2008; Manduca et al., 2016), but not after chronic MPH (Bolanos et al., 2003). These data suggest that the extent of psychostimulant exposure is crucial for determining behavioral and molecular outputs in the brain reward circuit such as receptor expression, second messenger signaling, transporter function, and trafficking, among others (Anderson and Pierce, 2005).
Aggressive behavior is a form of social communication, used to exert control over the social environment. It is characterized by a series of specific sequenced patterns that vary across the species in terms of frequency and duration. These are usually aimed at intruders considered rivals and are meant to suppress their reproductive success (Bartholow, 2018).
Aggressive behavior can be offensive or defensive. The offensive behavior is typically displayed by the resident and is characterized by introductory and threatening demonstrations towards the intruder intended to damage him, sometimes including an attack or behaviors such as aggressive groom, charge, and chase, among others (Koolhaas et al., 2013). The intruder usually displays in response a defensive behavior, characterized by the lack of initiative, evade, flee, jump, and others, which can result in non-intentional damage (Olivier and Young, 2002; Koolhaas et al., 2013).
It is important to note that aggression and violence are not the same. Aggressive behaviors are social responses considered normal and adaptive in animals and humans. On the other hand, violence is a harmful form of offensive aggression, given that it is a form of behavior regardless of the context that may include attacks towards vulnerable body spots such as paws, belly, and throat. In contrast, the aggression is usually directed to the back and the neck, both considered non-vulnerable targets for the attack (Koolhaas et al., 2013). It has been shown in mice a phenomenon known as “addiction to aggression.” Aggression is highly sought due to its gratifying effects, despite negative consequences in the short and long term, registering high rates of relapse, which reminds some features observed in drug addiction (Golden et al., 2019).
Several studies have focused on the impact of psychostimulants on aggressive behavior. Currently, mounting evidence shows that the use of drugs like AMPH, MAMPH, and ecstasy influences aggressive behavior, whereas the effect of MOD on this type of behavior has been less studied (Dawe et al., 2009; Machalova et al., 2012). Machalova and colleagues (Machalova et al., 2012) showed in an animal model of social conflict interaction (agonistic behavior model), that MOD inhibits attacks in aggressive mice and increases aggression in shy mice. On the other hand, acutely administrated MAMPH in aggressive mice produces a dose-dependent inhibition of aggressive behavior and an increase in timid behavior at high doses (Machalova et al., 2012). Finally, when they administered MDMA, both types of mice (aggressive and shy) showed an increase in the frequency of timid displays and a decrease in aggressive behavior was observed only in aggressive mice (Machalova et al., 2012). Similar results have been shown in BALB/cJ mice, an animal model of ADHD and conduct disorder, where the administration of 3 mg/kg intraperitoneal MPH produces a decrease in aggressive behavior while 10 mg/kg increases aggression, showing a dose-dependent effect (Jager et al., 2019). The first studies done with AMPH showed similar results (Miczek, 1974; Miczek and Tidey, 1989). Currently, the National Institute on Drug Abuse (NIDA, USA) directly relates the chronic use of MAMPH with violent behavior (NIDA, 2023). However, an increase in aggression levels due to the therapeutic use of AMPH has not been reported (O’Malley et al., 2022).
Regarding cocaine effects on aggression, controversial results have been reported. For instance, exposure to a single high dose of cocaine in adult rats produces a decrease in defensive behaviors in the resident-intruder paradigm (Alves et al., 2014), whereas when a social interaction test was used, the results were opposite (Rademacher et al., 2002). In isolated adult mice that receive cocaine in a single-binge administration, an increase in their defense behavior was observed (Estelles et al., 2004).
As indicated by the evidence, psychostimulants effects on aggressive behavior are diverse. This variability could be related to different paradigms, drug dose, age, type of administration (acute or chronic), and the specific neurobiological effects of each drug.
Maternal care is a complex behavior that directly influences the survival chances of the offspring (Fischer and O’Connell, 2018). This behavior includes building a nest and crouching over pups to keep them warm, nursing pups to provide nutrition, grooming them, protecting them from intruders, and retrieving pups back to the nest when they become displaced. The neurobiology of maternal care has been frequently studied in rodents, since females display very well characterized maternal behaviors towards their offspring that are easily distinguished and measured (Stolzenberg and Mayer, 2019).
Maternal care is sensitive to several environmental factors such as drugs of abuse, experience, housing conditions, isolation, and pharmacological manipulations. For instance, Perry and colleagues studied the effects of 3 days of MAMPH administration at three different doses on parental behavior in prairie voles. They found that the lowest dose (0.2 mg/kg) was enough to reduce the contact with the pups (Perry et al., 2019). When MAMPH injections (5 mg/kg) were used prenatally in pregnant rats, during the first or second half of the gestation period, several physiological and sensorimotor effects were observed in the offspring, although there were no differences in maternal behavior (Malinová-Ševčíková et al., 2014). On the other hand, repeated MPH (5 mg/kg) exposure during the lactation period impairs both maternal and adult offspring behavior. Regarding maternal behavior, the latency of retrieving pups was increased accompanied by a lower number of dams retrieving their pups (Ponchio et al., 2015).
In an interesting study with a fostering approach, the maternal behavior in juvenile offspring that were reared by dams exposed to intermittent cocaine was poor (Johns et al., 2007). Besides, chronic cocaine administration twice daily to pregnant rats for the entire gestational period elicited those dams to spend significantly less time next to the pups on postpartum day 1, and at postpartum day five, pups engaged in less ultrasonic vocalizations (Lippard et al., 2015). In contrast, another study showed that 10 days of cocaine administration to females prior to mating was enough to increase subsequent maternal behavior (Nephew and Febo, 2010). In particular, these mothers retrieved pups more quickly, spent more time taking care of the pups, and were more aggressive towards a male intruder on day 2 of lactation compared to control animals. However, no effects were observed on days 9 and 16 of lactation (Nephew and Febo, 2010).
In summary, maternal behavior is a set of complex behaviors towards pups with their survival as a major goal. As with other social behaviors, caring for pups is a rewarding experience that can be positively or negatively affected by psychostimulants.
Pair bonding is the social behavior associated with the ability to form intense and selective social attachments (pair bonds) usually, but not exclusively, related to sexual attraction and the partner preference for mating. The brain areas associated with this behavior are PFC, NAc and VP among others.
The rodent prairie vole (Microtus ochrogaster) is an animal widely used to study pair bonding and the effects of drugs of abuse on the neurobiological mechanisms behind this behavior. This species is highly social and monogamous (although now it is known that there are two species, one monogamous and the other promiscuous) and forms long-term and exclusive pair bonds with a partner after mating (McGraw and Young, 2010). In this case, preference for the partner over a stranger conspecific is considered a pair bond index and is usually measured as time or percentage of total time of close physical contact between the animals.
In adult prairie voles, it has been reported inhibition of partner preference after AMPH exposure in both sexes (Liu et al., 2010; Young et al., 2014). Notably, the effects of AMPH were observed exclusively on partner preference since other related behaviors such as mating frequency or locomotor activity were not affected. Studies performed in monogamous species have suggested that neuropeptides such as OXT and AVP facilitate pair bond (Rigney et al., 2022). For instance, it has been reported that OXT administration into the mPFC restores both partner preference and NAc DA levels, impaired by AMPH administration (Young et al., 2014). On the other hand, in agreement with the protective effects of social bonding on drug abuse, Liu and colleagues (Liu et al., 2011) have demonstrated that pair bonding prevents AMPH-induced CPP, through a D1R-mediated mechanism. Moreover, as shown in female prairie voles, AMPH exposure increases DA levels in the NAc but not in mPFC or VTA, increases D1R mRNA in the NAc and decreases D2R mRNA in VTA. These data indicate that the effects of AMPH upon pair bonding in female prairie voles exhibit nuclei and receptor specificity (Young et al., 2011).
In the same way, a study using a continuous 2 bottle choice procedure (one bottle with tap water and other bottle with MAMPH), showed that MAMPH had an inhibitory effect in partner preference, an effect that is accompanied by a decrease of OXT levels in the paraventricular nucleus of the hypothalamus (Hostetler et al., 2016).
Globally, these works highlight the profound effects of psychostimulants in pair bonding. Since, OXT modulates the DA system and the reward process, drugs acting on OXT receptors appear promising as treatments for conditions in which these behaviors are altered.
Table 1 summarizes the results of the studies evaluating the effects of different psychostimulants upon diverse social behaviors in animal models.
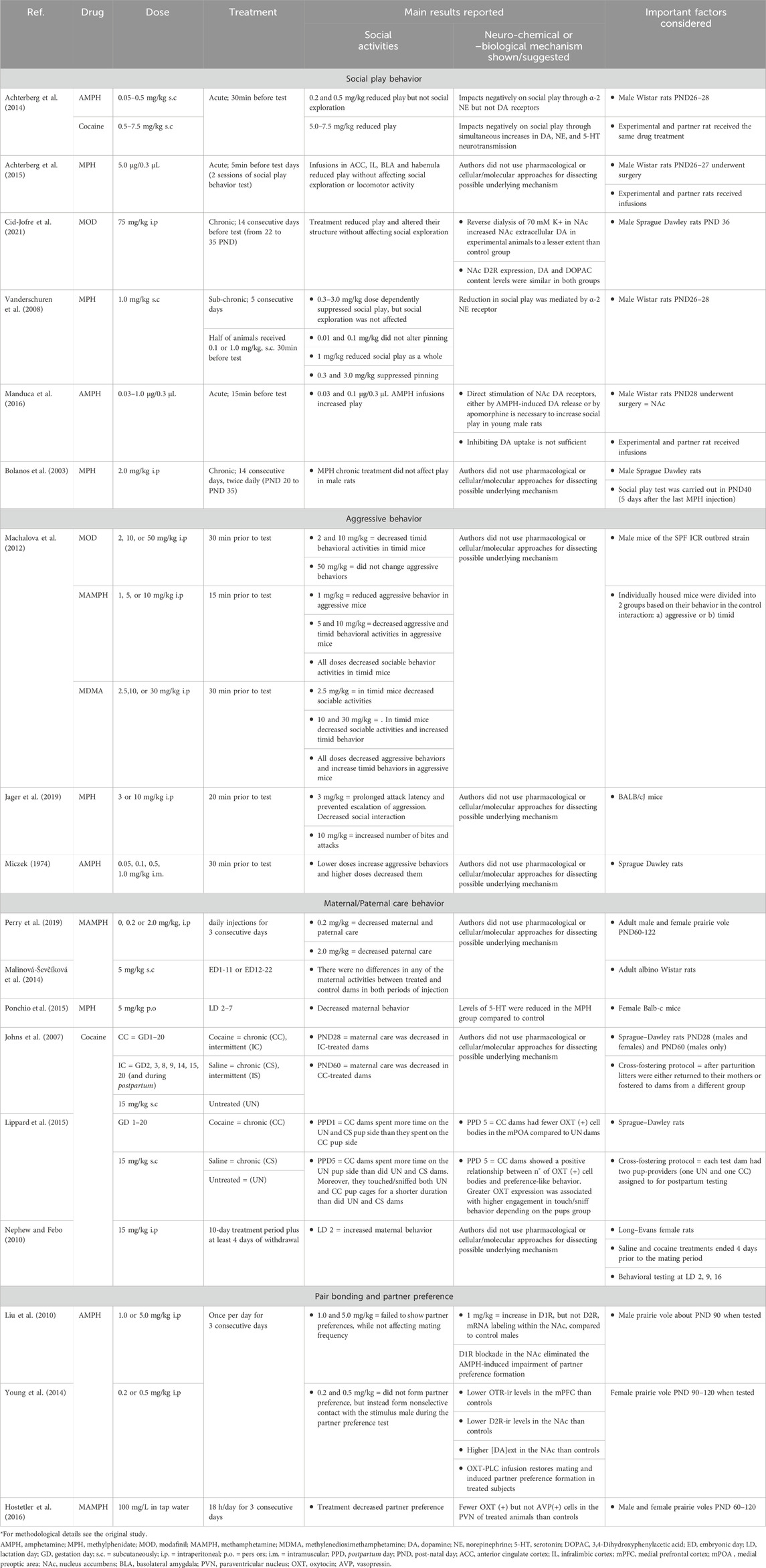
Table 1. Animal research summary in social play, aggression, maternal care, pair bonding and partner preference studies*.
Psychostimulants could also affect human social skills and behavior (Henry et al., 2009; Fox et al., 2011; Morgan and Marshall, 2013; Preller et al., 2014; Shreffler et al., 2022). However, as seen with animal models, the effects of psychostimulants upon social behavior can be controversial and even opposite depending on the type of drug analyzed, whether they are used in clinical or recreational/abuse contexts, the time of use, the age of the user, etc. Thus, for example, it has been shown that in general terms, chronic psychostimulant (e.g., cocaine or MAMPH) abusers tend to have some impairment in key social skills (Henry et al., 2009; Fox et al., 2011; Morgan and Marshall, 2013; Preller et al., 2014; Shreffler et al., 2022) and reduced social networks, and also show a series of alterations in brain networks underlying social behaviors (see, for example, (Aron and Paulus, 2007; Monterosso et al., 2007; Kim et al., 2010; Mackey and Paulus, 2013)).In contrast, patients using psychostimulants in clinical settings (e.g., MPH for the treatment of ADHD) exhibit improvements in some of the same aforementioned social skills (Fantozzi et al., 2021a; Fantozzi et al., 2021b; Levi-Shachar et al., 2021). Beyond these global considerations, most of the reports highlight that there is a lack of research in humans regarding psychostimulants and social behaviors since the majority of the works are related to their physiological effects, addictive properties, and potential treatments for their abuse. Here, we give an overview of some recent studies that show a relationship between psychostimulant consumption and modifications in sociality, empathy and aggression in humans.
One of the most studied drugs in relation with social behavior is MDMA. In 2012, MDMA was estimated to be the 3rd recreational drug among adults (between 18 and 25 years old) and in the same year, nearly 869,000 consumed it for the first time in the USA (Substance Abuse and Mental Health Services Administration, 2014). In addition, MDMA is under examination as a potential counterpart for psychotherapy (Feduccia et al., 2023; Kangaslampi and Zijlmans, 2023). In various reports it has been described that MDMA has a prosocial effect. For instance, Wardle and de Wit (Wardle and de Wit, 2014) analyzed the effects of MDMA using a three-session within-subjects design over 36 healthy volunteers of both sexes. In females only, MDMA (1.5 mg/kg) both increased smiling (zygomatic muscle activity) and reduced frown (corrugator muscle activity) in response to happy facial expressions, indicating more positive reactions to positive facial expressions. Therefore, MDMA altered emotional perception processing and response. It also impacts behavior and perception in an actual social scenario with a partner (Wardle and de Wit, 2014).
In a non-social and controlled laboratory setup, the administration of MDMA (from 0.5 to 2.0 mg/kg) increased (relative to placebo) self-report ratings of a broad range of socially relevant mood states. In this line, there is some evidence suggesting that MDMA prosocial effect depends on the social setting used in research. When MDMA (1.0 mg/kg single administration) was analyzed in volunteers, the increased feelings of confidence only appeared in subjects that were accompanied by other participants (exposed to MDMA too) (Kirkpatrick and de Wit, 2015). Interestingly, MDMA (0.75 mg/kg) increased the rating of loneliness (Bedi et al., 2010) in individuals who received the drug alone in isolated laboratory setups. In a study of free speech, MDMA increased the use of social and sexual words, as well as words referring to death, (Baggott et al., 2015), suggesting that consequences on speech reflect underlying alterations in social mood states. Thus, these findings suggest that MDMA generates prosocial feelings and different mental states in humans in controlled laboratory conditions that depend on social settings.
Studying several domains of social cognition in cocaine, MAMPH, and non-medical MPH users, interesting results have been described. For instance, a negative impact in the total performance for emotion cognition was observed only in MAMPH users. For emotional empathy, cocaine and MAMPH score lower than controls. Regarding cocaine users they show decreased social and moral decision-making, and lower social reward compared to controls [reviewed in (Quednow, 2017). As we mentioned previously, social network size is influenced by psychostimulant abuse. In both cases, cocaine and MPH users exhibit smaller social networks (Quednow, 2017).
More recently, social decision-making was analyzed in males with MAMPH use disorder. Applying a modified dictator monetary game where two scenarios were present (disadvantageous and advantageous), the pro-sociality behavior was measured. If the subject chose no money or less money for themselves against the hypothetical strangers, the choice was selected as pro-social. In the disadvantageous context, males with MAMPH use disorder made fewer pro-social choices compared to controls, however, in the advantageous context authors reported no differences in pro-social choices between groups (Li et al., 2022). Although the pro-social decisions could be different if the interaction were face to face, this study suggests that in disadvantageous scenarios MAMPH male users have less consideration of other’s benefits as compared to healthy controls.
On the other hand, there is evidence that ADHD patients have some problems with emotion recognition and social skills. MPH is the first-line medication for this disorder and several works have studied the effect of the drug on different aspects of social performance. Alkalay and Dan (Alkalay and Dan, 2022), after reviewing 15 studies on the impact of MPH on social performance in children between 6 and 14 years old with ADHD, concluded that MPH seems to improve the social deficit after weeks of treatment, increasing the capacity to recognize emotions and decreasing conflictive behaviors. However, as mentioned, these authors recognize that still controversial results exist and further studies are required to confirm conclusions.
On the other hand, the administration of lisdexamphetamine (LDX) to parents diagnosed with ADHD improved parenting behaviors. The pharmacological intervention reduced ADHD symptoms -negative talk during free play and quiet time components. Also, parents treated with LDX made over four times more praising statements, reduced their number of verbalizations during the homework task, and reduced the percentage of demands during tasks (Waxmonsky et al., 2014).
High aggression is a problematic social behavior that can be explained, among other causes, by a deficit of social cognition skills that could be impaired by the use of drugs. Impulsivity, a characteristic related to drug abuse, has also been associated with aggressive behaviors. In addition, although not well studied, the use of psychostimulants seems to be a cause of aggression. A study performed in medical students assessed the relationship between the use of psychostimulants for non-medical reasons and aggressive-hostility behavior. The study was carried out using a 73-question anonymous survey that included the Zuckerman–Kuhlman Personality Questionnaire in which aggressive behavior is described as a “tendency to express verbal aggression and show rudeness” among other qualities such as “thoughtlessness, vengefulness, spitefulness, quick temper and impatient behavior.” Results showed that the use of psychostimulants is related to aggressive behavior at least in this student sample (Bucher et al., 2013).
Among psychostimulants, several studies have shown that the use of amphetamine-related drugs, in particular MAMPH, increases aggressive behaviors or trait aggression (Sekine et al., 2006; Plüddemann et al., 2010; Payer et al., 2012; Lederer et al., 2016). A study assessed the magnitude of deficit in social cognition skills and its relation with aggression in MAMPH dependent individuals, compared with MAMPH users with psychosis and a control group. The criteria used were the facial morphing “emotion recognition task” with emotions such as anger, fear, happiness, and sadness, the “reading the mind in the eyes task” and the aggression questionnaire. The MAMPH dependent groups showed impairment in social cognition skills and higher levels of aggression, although there were no associations between them (Uhlmann et al., 2018).
Although it is well known that the use of MAMPH increases aggression, little is known about the underlying biological mechanisms. It has been shown that MAMPH withdrawal increases aggressive levels and decreases the expression of serotonin transporters in several brain regions like the midbrain, striatum and cortex (Sekine et al., 2006). Other studies have shown that MAMPH dependent individuals exhibiting higher aggression levels also showed higher activity in PFC and occipital cortex (Payer et al., 2012) and a decrease of frontal white matter (Lederer et al., 2016). Altogether these results suggest that MAMPH abuse increases aggression due to, at least in part, the alteration in frontal cortex functions.
The use of alcohol is related to aggression, and on many occasions people combine the use of alcohol with other drugs. It has been shown that in amphetamine-type stimulant users, the combination with alcohol increases aggression and hostility only in MAMPH users but not in those who combine alcohol with MDMA (EllenLeslie et al., 2017). Another risk factor that could increase aggressive behavior in MAMPH users is post-traumatic stress disorder (Wahlstrom et al., 2015). These results highlight the importance of different risk factors that could increase the levels of aggression in psychostimulant users.
Regarding cocaine, less is known about the relationship between this drug and aggression. In a 2002 study, Moeller and colleagues (Moeller et al., 2002) investigated the effects of cocaine on impulsivity. In brief, impulsivity was measured in both cocaine dependent subjects and controls using an impulsiveness scale and a monetary delayed behavioral task using a computer. The results showed a higher impulsivity in cocaine dependent volunteers compared to control individuals and this outcome was not related to a history of aggressive behavior (Moeller et al., 2002). Importantly, the impulsiveness scale applied did not distinguish between social and non-social related impulsive behavior. In another study, detoxified cocaine-dependent patients had higher levels of physical aggression and hostility, and also, higher levels of dysfunctional impulsivity than healthy control subjects (Roozen et al., 2011). On the other hand, in the context of ADHD treatment, it has been shown that the treatment with MPH was effective on aggressive behavior when ADHD is comorbid with oppositional defiant disorder and aggression (Masi et al., 2017). However, this study has several limitations (non-randomized, non-blinded and few patients), and therefore more studies are needed to elucidate the effect of psychostimulants on aggressive behaviors, when used as a therapeutic treatment.
Table 2 summarizes the results of the studies evaluating the effects of different psychostimulants upon diverse social behaviors in humans.
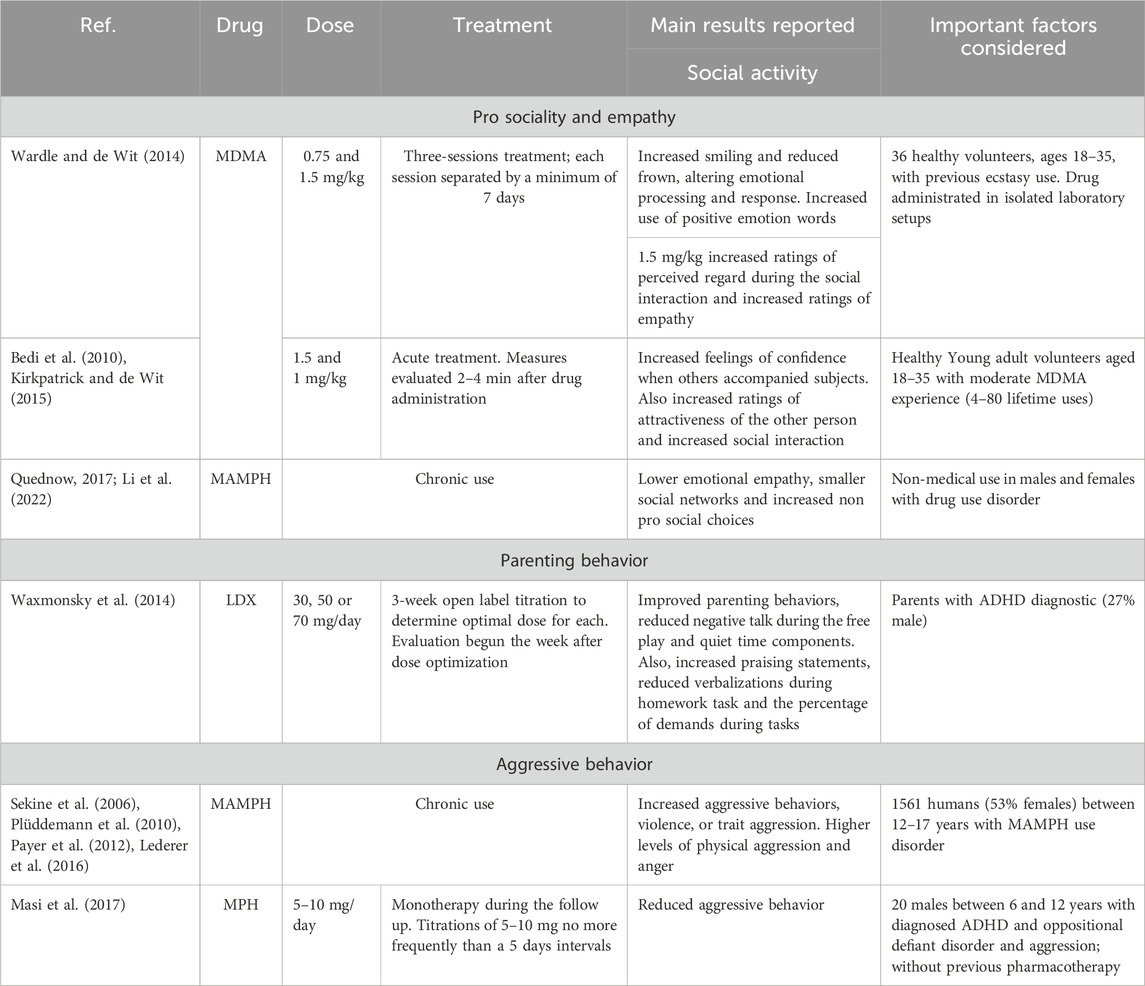
Table 2. Human research summary in social behaviors.
There is mounting evidence, both from animal models and human studies, showing that psychostimulants can significantly affect social behaviors. This is not surprising considering that the neural circuits underlying the regulation and expression of social behaviors are highly overlapped with those targeted by psychostimulants, which in most cases have strong rewarding and consequently addictive properties. Thus, it is also not surprising that the effects of psychostimulants on these behaviors can be very different depending on numerous variables such as the type of drug considered, acute versus long-term use, clinical versus recreational consumption or the presence or absence of concomitant risk factors (e.g., neuropsychiatric comorbidities, simultaneous consumption of other drugs, etc.), to name a few.
Furthermore, even though we used that terminology in this work, we think that considering psychostimulants as a relatively homogeneous drug class might be a misleading approach. Thus, the diverse effects on monoamines, i.e., more pronounced effects on DA and/or NE and/or 5-HT, as well as their differential pharmacodynamics regarding monoamines uptake (blockers, releasers, partial releasers, etc.) should be considered carefully when assessing the impact of different psychostimulants upon social behaviors.
In our opinion, psychostimulants-induced changes in social behaviors, either prosocial or antisocial, should be always monitored, particularly when using this type of drugs in therapeutic contexts. This assessment will be not only beneficial to establish some specific behavioral effects and how these can translate to neurochemistry, but also would shed light on how this dimension can be used/modified to improve the desired clinical outcomes and to reduce the risk of abuse and/or addiction. Indeed, studies in rodents have shown that the interaction with a partner during the acquisition of addictive behavior or relapse decreases the chances of developing addiction (noteworthy, opposite results were found if the partner also uses the drug) (Sampedro-Piquero et al., 2019; Venniro et al., 2022). Therefore, it is essential to consider the reward elicited by social interaction as an important factor that could improve addiction treatments.
Despite these considerations, it is clear that much more research is needed to have a better understanding of the physiological mechanisms underlying different social behaviors and how they are affected by diverse psychostimulants. This seems to be of particular relevance in a scenario of a growing consumption of this type of compounds.
VC-J: Writing–original draft, Writing–review and editing. TB: Writing–review and editing. AC: Writing–original draft. IA: Writing–original draft. MR-P: Writing–original draft, Writing–review and editing. GR: Writing–original draft, Writing–review and editing.
The author(s) declare that financial support was received for the research, authorship, and/or publication of this article. Proyecto POSTDOC_DICYT 022101RP, Vicerrectoría de Investigación, Desarrollo e Innovación, DICYT, Universidad de Santiago de Chile. FONDECYT Grants 1220656 and 1230446 (MR-P).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Achterberg, E., Trezza, V., Siviy, S. M., Schrama, L., Schoffelmeer, A. N., and Vanderschuren, L. J. (2014). Amphetamine and cocaine suppress social play behavior in rats through distinct mechanisms. Psychopharmacology 231, 1503–1515. doi:10.1007/s00213-013-3272-9
Achterberg, E., Van Kerkhof, L. W., Servadio, M., Van Swieten, M. M., Houwing, D. J., Aalderink, M., et al. (2016). Contrasting roles of dopamine and noradrenaline in the motivational properties of social play behavior in rats. Neuropsychopharmacology 41, 858–868. doi:10.1038/npp.2015.212
Achterberg, E. J., van Kerkhof, L. W., Damsteegt, R., Trezza, V., and Vanderschuren, L. J. (2015). Methylphenidate and atomoxetine inhibit social play behavior through prefrontal and subcortical limbic mechanisms in rats. J. Neurosci. 35, 161–169. doi:10.1523/JNEUROSCI.2945-14.2015
Ahloy-Dallaire, J., Espinosa, J., and Mason, G. (2018). Play and optimal welfare: does play indicate the presence of positive affective states? Behav. Process. 156, 3–15. doi:10.1016/j.beproc.2017.11.011
Ahrens, A. M., Ferguson, L. M., Robinson, T. E., and Aldridge, J. W. (2018). Dynamic encoding of incentive salience in the ventral pallidum: dependence on the form of the reward cue. eneuro 5, ENEURO.0328. doi:10.1523/ENEURO.0328-17.2018
Alkalay, S., and Dan, O. (2022). Effect of short-term methylphenidate on social impairment in children with attention deficit/hyperactivity disorder: systematic review. Child. Adolesc. Psychiatry Ment. Health 16, 93. doi:10.1186/s13034-022-00526-2
Alves, C., Magalhães, A., Melo, P., de Sousa, L., Tavares, M., Monteiro, P., et al. (2014). Long-term effects of chronic cocaine exposure throughout adolescence on anxiety and stress responsivity in a Wistar rat model. Neuroscience 277, 343–355. doi:10.1016/j.neuroscience.2014.07.008
Ambroggi, F., Ghazizadeh, A., Nicola, S. M., and Fields, H. L. (2011). Roles of nucleus accumbens core and shell in incentive-cue responding and behavioral inhibition. J. Neurosci. 31, 6820–6830. doi:10.1523/JNEUROSCI.6491-10.2011
Amiri, S., Mohammadi, M. R., Mohammadi, M., Nouroozinejad, G. H., Kahbazi, M., and Akhondzadeh, S. (2008). Modafinil as a treatment for Attention-Deficit/Hyperactivity Disorder in children and adolescents: a double blind, randomized clinical trial. Prog. Neuropsychopharmacol. Biol. Psychiatry 32, 145–149. doi:10.1016/j.pnpbp.2007.07.025
Anastasiades, P. G., and Carter, A. G. (2021). Circuit organization of the rodent medial prefrontal cortex. Trends Neurosci. 44 (7), 550–563. doi:10.1016/j.tins.2021.03.006
Anderson, S., and Pierce, R. (2005). Cocaine-induced alterations in dopamine receptor signaling: implications for reinforcement and reinstatement. Pharmacol. Ther. 106, 389–403. doi:10.1016/j.pharmthera.2004.12.004
Argue, K. J., and McCarthy, M. M. (2015a). Utilization of same-vs. mixed-sex dyads impacts the observation of sex differences in juvenile social play behavior. Curr. Neurobiol. 6, 17–23. doi:10.4172/0975-9042.000117
Argue, K. J., and McCarthy, M. M. (2015b). Characterization of juvenile play in rats: importance of sex of self and sex of partner. Biol. Sex. Differ. 6, 16. doi:10.1186/s13293-015-0034-x
Arnold, V. K., Feifel, D., Earl, C. Q., Yang, R., and Adler, L. A. (2014). A 9-week, randomized, double-blind, placebo-controlled, parallel-group, dose-finding study to evaluate the efficacy and safety of modafinil as treatment for adults with ADHD. J. Atten. Disord. 18, 133–144. doi:10.1177/1087054712441969
Aron, J. L., and Paulus, M. P. (2007). Location, location: using functional magnetic resonance imaging to pinpoint brain differences relevant to stimulant use. Addiction 102 (Suppl. 1), 33–43. doi:10.1111/j.1360-0443.2006.01778.x
Arria, A. M., and DuPont, R. L. (2010). Nonmedical prescription stimulant use among college students: why we need to do something and what we need to do. J. Addict. Dis. 29, 417–426. doi:10.1080/10550887.2010.509273
Baggott, M. J., Kirkpatrick, M. G., Bedi, G., and de Wit, H. (2015). Intimate insight: MDMA changes how people talk about significant others. J. Psychopharmacol. 29 (6), 669–677. doi:10.1177/0269881115581962
Bardo, M. T., Neisewander, J., and Kelly, T. (2013). Individual differences and social influences on the neurobehavioral pharmacology of abused drugs. Pharmacol. Rev. 65, 255–290. doi:10.1124/pr.111.005124
Bartholow, B. D. (2018). The aggressive brain: insights from neuroscience. Curr. Opin. Psychol. 19, 60–64. doi:10.1016/j.copsyc.2017.04.002
Bedi, G., Hyman, D., and de Wit, H. (2010). Is ecstasy an "empathogen"? Effects of ±3,4-methylenedioxymethamphetamine on prosocial feelings and identification of emotional states in others. Biol. Psychiatry 68 (12), 1134–1140. doi:10.1016/j.biopsych.2010.08.003
Berridge, K. C., and Kringelbach, M. L. (2015). Pleasure systems in the brain. Neuron 86, 646–664. doi:10.1016/j.neuron.2015.02.018
Berridge, K. C., and Robinson, T. E. (2016). Liking, wanting, and the incentive-sensitization theory of addiction. Am. Psychol. 71, 670–679. doi:10.1037/amp0000059
Bolanos, C. A., Barrot, M., Berton, O., Wallace-Black, D., and Nestler, E. J. (2003). Methylphenidate treatment during pre- and periadolescence alters behavioral responses to emotional stimuli at adulthood. Biol. Psychiatry 54, 1317–1329. doi:10.1016/s0006-3223(03)00570-5
Breckenridge, A. M., Breckenridge, R. A., and Peck, C. C. (2019). Report on the current status of the use of real-world data (RWD) and real-world evidence (RWE) in drug development and regulation. Br. J. Clin. Pharmacol. 85, 1874–1877. doi:10.1111/bcp.14026
Bucher, J. T., Vu, D. M., and Hojat, M. (2013). Psychostimulant drug abuse and personality factors in medical students. Med. Teach. 35 (1), 53–57. doi:10.3109/0142159X.2012.731099
Caldwell, J. L., Schroeder, V. M., Kunkle, C. L., and Stephenson, H. G. (2020). Differential effects of modafinil on performance of low-performing and high-performing individuals during total sleep deprivation. Pharmacol. Biochem. Behav. 196, 172968. doi:10.1016/j.pbb.2020.172968
Chen, P., and Hong, W. (2018). Neural circuit mechanisms of social behavior. Neuron 98, 16–30. doi:10.1016/j.neuron.2018.02.026
Cid-Jofre, V., Garate-Perez, M., Clark, P. J., Valero-Jara, V., Espana, R. A., Sotomayor-Zarate, R., et al. (2021). Chronic modafinil administration to preadolescent rats impairs social play behavior and dopaminergic system. Neuropharmacology 183, 108404. doi:10.1016/j.neuropharm.2020.108404
Clare, K., Pan, C., Kim, G., Park, K., Zhao, J., Volkow, N. D., et al. (2021). Cocaine reduces the neuronal population while upregulating dopamine D2-receptor-expressing neurons in brain reward regions: sex-effects. Front. Pharmacol. 12, 624127. doi:10.3389/fphar.2021.624127
d’Angelo, L. C., Savulich, G., and Sahakian, B. J. (2017). Lifestyle use of drugs by healthy people for enhancing cognition, creativity, motivation and pleasure. Br. J. Pharmacol. 174, 3257–3267. doi:10.1111/bph.13813
Dawe, S., Davis, P., Lapworth, K., and McKetin, R. (2009). Mechanisms underlying aggressive and hostile behavior in amphetamine users. Curr. Opin. Psychiatry 22, 269–273. doi:10.1097/YCO.0b013e32832a1dd4
de Wit, H., and Richards, J. B. (2004). “Dual determinants of drug use in humans: reward and impulsivity,” in Motivational factors in the etiology of drug abuse volume 50 of the Nebraska symposium on motivation. Editors R. A. Bevins, and T. B. Michael (Lincoln, NE: University of Nebraska Press), 19–55.
Douglas, H., Halverstadt, B., Reinhart-Anez, P., Webber, E., and Cromwell, H. (2018). A possible social relative reward effect: influences of outcome inequity between rats during operant responding. Behav. Process. 157, 459–469. doi:10.1016/j.beproc.2018.06.016
Douglas, L. A., Varlinskaya, E. I., and Spear, L. P. (2004). Rewarding properties of social interactions in adolescent and adult male and female rats: impact of social versus isolate housing of subjects and partners. Dev. Psychobiol. 45, 153–162. doi:10.1002/dev.20025
Dunlap, L. E., Andrews, A. M., and Olson, D. E. (2018). Dark classics in chemical neuroscience: 3,4-methylenedioxymethamphetamine. ACS Chem. Neurosci. 9 (10), 2408–2427. doi:10.1021/acschemneuro.8b00155
EllenLeslie, M. E. M., Andrew Smirnov, A., Adrian Cherney, A., Helene Wells, H., Margot Legosz, M., Robert Kemp, R., et al. (2017). Simultaneous use of alcohol with methamphetamine but not ecstasy linked with aggression among young adult stimulant users. Addict. Behav. 70, 27–34. doi:10.1016/j.addbeh.2017.01.036
Estelles, J., Rodriguez-Arias, M., Aguilar, M., and Minarro, J. (2004). Social behavioural profile of cocaine in isolated and grouped male mice. Drug alcohol dependence 76, 115–123. doi:10.1016/j.drugalcdep.2004.04.019
Fantozzi, P., Muratori, P., Caponi, M. C., Levantini, V., Nardoni, C., Pfanner, C., et al. (2021b). Treatment with methylphenidate improves affective but not cognitive empathy in youths with attention-deficit/hyperactivity disorder. Child. (Basel) 8 (7), 596. doi:10.3390/children8070596
Fantozzi, P., Sesso, G., Muratori, P., Milone, A., and Masi, G. (2021a). Biological bases of empathy and social cognition in patients with attention-deficit/hyperactivity disorder: a focus on treatment with psychostimulants. Brain Sci. 11 (11), 1399. doi:10.3390/brainsci11111399
Faraone, S. V. (2018). The pharmacology of amphetamine and methylphenidate: relevance to the neurobiology of attention-deficit/hyperactivity disorder and other psychiatric comorbidities. Neurosci. Biobehav. Rev. 87, 255–270. doi:10.1016/j.neubiorev.2018.02.001
Feduccia, A. A., Jerome, L., Yazar-Klosinski, B., Emerson, A., Mithoefer, M. C., and Doblin, R. (2023). Breakthrough for trauma treatment: safety and efficacy of MDMA-assisted psychotherapy compared to paroxetine and sertraline. Focus Am Psychiatr. Publ. 21 (3), 306–314. doi:10.1176/appi.focus.23021013
Fischer, E. K., and O’Connell, L. A. (2018). Circuit architecture underlying distinct components of parental care. Trends Neurosci. 41, 334–336. doi:10.1016/j.tins.2018.04.003
Fox, H. C., Bergquist, K. L., Casey, J., Hong, K. A., and Sinha, R. (2011). Selective cocaine-related difficulties in emotional intelligence: relationship to stress and impulse control. Am. J. Addict. 20 (2), 151–160. doi:10.1111/j.1521-0391.2010.00108.x
Franklin, T. B., Silva, B. A., Perova, Z., Marrone, L., Masferrer, M. E., Zhan, Y., et al. (2017). Prefrontal cortical control of a brainstem social behavior circuit. Nat. Neurosci. 20, 260–270. doi:10.1038/nn.4470
Goez, H. R., Scott, O., Nevo, N., Bennett-Back, O., and Zelnik, N. (2012). Using the test of variables of attention to determine the effectiveness of modafinil in children with attention-deficit hyperactivity disorder (ADHD): a prospective methylphenidate-controlled trial. J. Child Neurology 27, 1547–1552. doi:10.1177/0883073812439101
Golden, S. A., Jin, M., and Shaham, Y. (2019). Animal models of (or for) aggression reward, addiction, and relapse: behavior and circuits. J. Neurosci. 39, 3996–4008. doi:10.1523/JNEUROSCI.0151-19.2019
Goodwin, J. S., Larson, G. A., Swant, J., Sen, N., Javitch, J. A., Zahniser, N. R., et al. (2009). Amphetamine and methamphetamine differentially affect dopamine transporters in vitro and in vivo. J. Biol. Chem. 284, 2978–2989. doi:10.1074/jbc.M805298200
Groppe, S. E., Gossen, A., Rademacher, L., Hahn, A., Westphal, L., Grunder, G., et al. (2013). Oxytocin influences processing of socially relevant cues in the ventral tegmental area of the human brain. Biol. Psychiatry 74, 172–179. doi:10.1016/j.biopsych.2012.12.023
Gunaydin, L. A., Grosenick, L., Finkelstein, J. C., Kauvar, I. V., Fenno, L. E., Adhikari, A., et al. (2014). Natural neural projection dynamics underlying social behavior. Cell 157, 1535–1551. doi:10.1016/j.cell.2014.05.017
Heilig, M., Epstein, D. H., Nader, M. A., and Shaham, Y. (2016). Time to connect: bringing social context into addiction neuroscience. Nat. Rev. Neurosci. 17, 592–599. doi:10.1038/nrn.2016.67
Henry, J. D., Mazur, M., and Rendell, P. G. (2009). Social-cognitive difficulties in former users of methamphetamine. Br. J. Clin. Psychol. 48 (Pt 3), 323–327. doi:10.1348/000712609X435742
Homberg, J. R., Schiepers, O. J., Schoffelmeer, A. N., Cuppen, E., and Vanderschuren, L. J. (2007). Acute and constitutive increases in central serotonin levels reduce social play behaviour in peri-adolescent rats. Psychopharmacology 195, 175–182. doi:10.1007/s00213-007-0895-8
Hoover, W. B., and Vertes, R. P. (2007). Anatomical analysis of afferent projections to the medial prefrontal cortex in the rat. Brain Struct. Funct. 212 (2), 149–179. doi:10.1007/s00429-007-0150-4
Hostetler, C. M., Phillips, T. J., and Ryabinin, A. E. (2016). Methamphetamine consumption inhibits pair bonding and hypothalamic oxytocin in prairie voles. PLoS One 11, e0158178. doi:10.1371/journal.pone.0158178
Hung, L. W., Neuner, S., Polepalli, J. S., Beier, K. T., Wright, M., Walsh, J. J., et al. (2017). Gating of social reward by oxytocin in the ventral tegmental area. Science 357, 1406–1411. doi:10.1126/science.aan4994
Hyman, S. E., Malenka, R. C., and Nestler, E. J. (2006). Neural mechanisms of addiction: the role of reward-related learning and memory. Annu. Rev. Neurosci. 29, 565–598. doi:10.1146/annurev.neuro.29.051605.113009
Jager, A., Kanters, D., Geers, F., Buitelaar, J. K., Kozicz, T., and Glennon, J. C. (2019). Methylphenidate dose-dependently affects aggression and improves fear extinction and anxiety in BALB/cJ mice. Front. Psychiatry 10, 768. doi:10.3389/fpsyt.2019.00768
Johns, J. M., McMurray, M. S., Hofler, V. E., Jarrett, T. M., Middleton, C. L., Elliott, D. L., et al. (2007). Cocaine disrupts pup-induced maternal behavior in juvenile and adult rats. Neurotoxicol Teratol. 29, 634–641. doi:10.1016/j.ntt.2007.07.003
Kangaslampi, S., and Zijlmans, J. (2023). MDMA-assisted psychotherapy for PTSD in adolescents: rationale, potential, risks, and considerations. Eur. Child. Adolesc. Psychiatry. doi:10.1007/s00787-023-02310-9
Karen Pierce, K., Frank Haist, F., Farshad Sedaghat, F., and Eric Courchesne, E. (2004). The brain response to personally familiar faces in autism: findings of fusiform activity and beyond. Brain 127 (12), 2703–2716. doi:10.1093/brain/awh289
Kim, Y. T., Lee, J. J., Song, H. J., Kim, J. H., Kwon, D. H., Kim, M. N., et al. (2010). Alterations in cortical activity of male methamphetamine abusers performing an empathy task: fMRI study. Hum. Psychopharmacol. 25 (1), 63–70. doi:10.1002/hup.1083
Kirkpatrick, M. G., and de Wit, H. (2015). MDMA: a social drug in a social context. Psychopharmacol. Berl. 232 (6), 1155–1163. doi:10.1007/s00213-014-3752-6
Knell, S. M. (2022). “Cognitive behavioral play therapy,” in Creative CBT with youth (Springer), 65–82.
Koob, G. F., and Volkow, N. D. (2010). Neurocircuitry of addiction. Neuropsychopharmacology 35, 217–238. doi:10.1038/npp.2009.110
Koob, G. F., and Volkow, N. D. (2016). Neurobiology of addiction: a neurocircuitry analysis. Lancet Psychiatry 3, 760–773. doi:10.1016/S2215-0366(16)00104-8
Koolhaas, J. M., Coppens, C. M., de Boer, S. F., Buwalda, B., Meerlo, P., and Timmermans, P. J. (2013). The resident-intruder paradigm: a standardized test for aggression, violence and social stress. JoVE J. Vis. Exp., e4367. doi:10.3791/4367
Lakhan, S. E., and Kirchgessner, A. (2012). Prescription stimulants in individuals with and without attention deficit hyperactivity disorder: misuse, cognitive impact, and adverse effects. Brain Behav. 2, 661–677. doi:10.1002/brb3.78
Lederer, K., Fouche, J. P., Wilson, D., Stein, D. J., and Uhlmann, A. (2016). Frontal white matter changes and aggression in methamphetamine dependence. Metab. Brain Dis. 31, 53–62. doi:10.1007/s11011-015-9775-9
Lei, K., Liu, Y., Smith, A. S., Lonstein, J. S., and Wang, Z. (2017). Effects of pair bonding on parental behavior and dopamine activity in the nucleus accumbens in male prairie voles. Eur. J. Neurosci. 46, 2276–2284. doi:10.1111/ejn.13673
Lesscher, H., Spoelder, M., Rotte, M. D., Janssen, M. J., Hesseling, P., Lozeman-van’t Klooster, J. G., et al. (2015). Early social isolation augments alcohol consumption in rats. Behav. Pharmacol. 26, 673–680. doi:10.1097/FBP.0000000000000165
Levi-Shachar, O., Gvirts, H. Z., Goldwin, Y., Bloch, Y., Shamay-Tsoory, S., Boyle, D., et al. (2021). The association between symptom severity and theory of mind impairment in children with attention deficit/hyperactivity disorder. Psychiatry Res. 303, 114092. doi:10.1016/j.psychres.2021.114092
Levy, D. R., Tamir, T., Kaufman, M., Parabucki, A., Weissbrod, A., Schneidman, E., et al. (2019). Dynamics of social representation in the mouse prefrontal cortex. Nat. Neurosci. 22, 2013–2022. doi:10.1038/s41593-019-0531-z
Li, M. H., Zhang, M., Yuan, T. F., and Rao, L. L. (2022). Impaired social decision-making in males with methamphetamine use disorder. Addict. Biol. 27, e13204. doi:10.1111/adb.13204
Lippard, E. T., Jarrett, T. M., McMurray, M. S., Zeskind, P. S., Garber, K. A., Zoghby, C. R., et al. (2015). Early postpartum pup preference is altered by gestational cocaine treatment: associations with infant cues and oxytocin expression in the MPOA. Behav. Brain Res. 278, 176–185. doi:10.1016/j.bbr.2014.09.045
Liu, Y., Aragona, B. J., Young, K. A., Dietz, D. M., Kabbaj, M., Mazei-Robison, M., et al. (2010). Nucleus accumbens dopamine mediates amphetamine-induced impairment of social bonding in a monogamous rodent species. Proc. Natl. Acad. Sci. 107, 1217–1222. doi:10.1073/pnas.0911998107
Liu, Y., Young, K. A., Curtis, J. T., Aragona, B. J., and Wang, Z. (2011). Social bonding decreases the rewarding properties of amphetamine through a dopamine D1 receptor-mediated mechanism. J. Neurosci. 31, 7960–7966. doi:10.1523/JNEUROSCI.1006-11.2011
Lukkes, J. L., Watt, M. J., Lowry, C. A., and Forster, G. L. (2009). Consequences of post-weaning social isolation on anxiety behavior and related neural circuits in rodents. Front. Behav. Neurosci. 3, 18. doi:10.3389/neuro.08.018.2009
Machalova, A., Slais, K., Vrskova, D., and Sulcova, A. (2012). Differential effects of modafinil, methamphetamine, and MDMA on agonistic behavior in male mice. Pharmacol. Biochem. Behav. 102, 215–223. doi:10.1016/j.pbb.2012.04.013
Mackey, S., and Paulus, M. (2013). Are there volumetric brain differences associated with the use of cocaine and amphetamine-type stimulants? Neurosci. Biobehav Rev. 37 (3), 300–316. doi:10.1016/j.neubiorev.2012.12.003
Malinová-Ševčíková, M., Hrebíčková, I., Macúchová, E., Nová, E., Pometlová, M., and Šlamberová, R. (2014). Differences in maternal behavior and development of their pups depend on the time of methamphetamine exposure during gestation period. Physiological Res. 63, S559–S572. doi:10.33549/physiolres.932925
Manduca, A., Servadio, M., Damsteegt, R., Campolongo, P., Vanderschuren, L. J., and Trezza, V. (2016). Dopaminergic neurotransmission in the nucleus accumbens modulates social play behavior in rats. Neuropsychopharmacology 41, 2215–2223. doi:10.1038/npp.2016.22
Masi, G., Manfredi, A., Nieri, G., Muratori, P., Pfanner, C., and Milone, A. (2017). A naturalistic comparison of methylphenidate and risperidone monotherapy in drug-naive youth with attention-deficit/hyperactivity disorder comorbid with oppositional defiant disorder and aggression. J. Clin. Psychopharmacol. 37 (5), 590–594. doi:10.1097/JCP.0000000000000747
McCreary, A. C., Muller, C. P., and Filip, M. (2015). Psychostimulants: basic and clinical pharmacology. Neuropsychiatric Complicat. Stimul. Abuse 120, 41–83. doi:10.1016/bs.irn.2015.02.008
McGraw, L. A., and Young, L. J. (2010). The prairie vole: an emerging model organism for understanding the social brain. Trends Neurosci. 33, 103–109. doi:10.1016/j.tins.2009.11.006
Miczek, K. A. (1974). Intraspecies aggression in rats: effects of d-amphetamine and chlordiazepoxide. Psychopharmacologia 39, 275–301. doi:10.1007/BF00422968
Miczek, K. A., and Tidey, J. W. (1989). Amphetamines: aggressive and social behavior. NIDA Res. Monogr. 94, 68–100.
Miller, E. K., and Cohen, J. D. (2001). An integrative theory of prefrontal cortex function. Annu. Rev. Neurosci. 24, 167–202. doi:10.1146/annurev.neuro.24.1.167
Moeller, F. G., Dougherty, D. M., Barratt, E. S., Oderinde, V., Mathias, C. W., Harper, R. A., et al. (2002). Increased impulsivity in cocaine dependent subjects independent of antisocial personality disorder and aggression. Drug Alcohol Depend. 68, 105–111. doi:10.1016/s0376-8716(02)00106-0
Monterosso, J. R., Ainslie, G., Xu, J., Cordova, X., Domier, C. P., and London, E. D. (2007). Frontoparietal cortical activity of methamphetamine-dependent and comparison subjects performing a delay discounting task. Hum. Brain Mapp. 28 (5), 383–393. doi:10.1002/hbm.20281
Morgan, M. J., and Marshall, J. P. (2013). Deficient fear recognition in regular cocaine users is not attributable to elevated impulsivity or conduct disorder prior to cocaine use. J. Psychopharmacol. 27 (6), 526–532. doi:10.1177/0269881113477708
Nephew, B. C., and Febo, M. (2010). Effect of cocaine sensitization prior to pregnancy on maternal care and aggression in the rat. Psychopharmacol. Berl. 209, 127–135. doi:10.1007/s00213-010-1777-z
Nichols, D. E. (2022). Entactogens: how the name for a novel class of psychoactive agents originated. Front. Psychiatry 13, 863088. doi:10.3389/fpsyt.2022.863088
O’Connell, L. A., and Hofmann, H. A. (2011). Genes, hormones, and circuits: an integrative approach to study the evolution of social behavior. Front. Neuroendocrinol. 32, 320–335. doi:10.1016/j.yfrne.2010.12.004
Olivier, B., and Young, L. J. (2002). “Animal models of aggression,” in Neuropsychopharmacology: the fifth generation of progress Vol. 118, 1699–1708.
O’Malley, K. Y., Hart, C. L., Casey, S., and Downey, L. A. (2022). Methamphetamine, amphetamine, and aggression in humans: a systematic review of drug administration studies. Neurosci. Biobehav. Rev. 141, 104805. doi:10.1016/j.neubiorev.2022.104805
Ottenheimer, D. J., Wang, K., Tong, X., Fraser, K. M., Richard, J. M., and Janak, P. H. (2020). Reward activity in ventral pallidum tracks satiety-sensitive preference and drives choice behavior. Sci. Adv. 6, eabc9321. doi:10.1126/sciadv.abc9321
Payer, D., Nurmi, E., Wilson, S., McCracken, J. T., and London, E. D. (2012). Effects of methamphetamine abuse and serotonin transporter gene variants on aggression and emotion-processing neurocircuitry. Transl. Psychiatry 2, e80. doi:10.1038/tp.2011.73
Pellis, S. M., and Pellis, V. C. (1987). Play-fighting differs from serious fighting in both target of attack and tactics of fighting in the laboratory rat Rattus norvegicus. Aggress. Behav. 13, 227–242. doi:10.1002/1098-2337(1987)13:4<227::aid-ab2480130406>3.0.co;2-c
Pellis, S. M., and Pellis, V. C. (1990). Differential rates of attack, defense, and counterattack during the developmental decrease in play fighting by male and female rats. Dev. Psychobiol. 23, 215–231. doi:10.1002/dev.420230303
Perry, A. N., Ortiz, R. J., Hernandez, K. R., and Cushing, B. S. (2019). Effects of methamphetamine on alloparental behavior in male and female prairie voles. Physiol. Behav. 203, 128–134. doi:10.1016/j.physbeh.2017.09.012
Piantadosi, P. T., Yeates, D. C., and Floresco, S. B. (2018). Cooperative and dissociable involvement of the nucleus accumbens core and shell in the promotion and inhibition of actions during active and inhibitory avoidance. Neuropharmacology 138, 57–71. doi:10.1016/j.neuropharm.2018.05.028
Plüddemann, A., Flisher, A. J., McKetin, R., Parry, C., and Lombard, C. (2010). Methamphetamine use, aggressive behavior and other mental health issues among high-school students in Cape Town, South Africa. Drug Alcohol Depend. 109 (1-3), 14–19. doi:10.1016/j.drugalcdep.2009.11.021
Ponchio, R., Teodorov, E., Kirsten, T. B., Coelho, C. P., Oshiro, A., Florio, J. C., et al. (2015). Repeated methylphenidate administration during lactation reduces maternal behavior, induces maternal tolerance, and increases anxiety-like behavior in pups in adulthood. Neurotoxicology Teratol. 50, 64–72. doi:10.1016/j.ntt.2015.05.008
Preller, K. H., Hulka, L. M., Vonmoos, M., Jenni, D., Baumgartner, M. R., Seifritz, E., et al. (2014). Impaired emotional empathy and related social network deficits in cocaine users. Addict. Biol. 19 (3), 452–466. doi:10.1111/adb.12070
Quednow, B. B. (2017). Social cognition and interaction in stimulant use disorders. Curr. Opin. Behav. Sci. 13, 55–62. doi:10.1016/j.cobeha.2016.10.001
Rademacher, D. J., Schuyler, A. L., Kruschel, C. K., and Steinpreis, R. E. (2002). Effects of cocaine and putative atypical antipsychotics on rat social behavior: an ethopharmacological study. Pharmacol. Biochem. Behav. 73, 769–778. doi:10.1016/s0091-3057(02)00904-8
Rigney, N., de Vries, G. J., Petrulis, A., and Young, L. J. (2022). Oxytocin, vasopressin, and social behavior: from neural circuits to clinical opportunities. Endocrinology 163 (9), bqac111. doi:10.1210/endocr/bqac111
Roozen, H. G., van der Kroft, P., van Marle, H. J., and Franken, I. H. (2011). The impact of craving and impulsivity on aggression in detoxified cocaine-dependent patients. J. Subst. Abuse Treat. 40 (4), 414–418. doi:10.1016/j.jsat.2010.12.003
Rudebeck, P. H., Walton, M. E., Millette, B. H. P., Shirley, E., Rushworth, M. F. S., and Bannerman, D. M. (2007). Distinct contributions of frontal areas to emotion and social behaviour in the rat. Eur. J. Neurosci. 26 (8), 2315–2326. doi:10.1111/j.1460-9568.2007.05844.x
Rugino, T. (2007). A review of modafinil film-coated tablets for attention-deficit/hyperactivity disorder in children and adolescents. Neuropsychiatr. Dis. Treat. 3 (3), 293–301.
Sampedro-Piquero, P., Ávila-Gámiz, F., Moreno Fernández, R. D., Castilla-Ortega, E., and Santín, L. J. (2019). The presence of a social stimulus reduces cocaine-seeking in a place preference conditioning paradigm. J. Psychopharmacol. 33 (12), 1501–1511. doi:10.1177/0269881119874414
Sekine, Y., Ouchi, Y., Takei, N., Yoshikawa, E., Nakamura, K., Futatsubashi, M., et al. (2006). Brain serotonin transporter density and aggression in abstinent methamphetamine abusers. Arch. Gen. Psychiatry 63 (1), 90–100. doi:10.1001/archpsyc.63.1.90
Shreffler, J., Thomas, J. J., McGee, S., Ferguson, B., Kelley, J., Cales, R., et al. (2022). Self-compassion in individuals with substance use disorder: the association with personal growth and well-being. J. Addict. Dis. 40 (3), 366–372. doi:10.1080/10550887.2021.2005382
Soares-Cunha, C., and Heinsbroek, J. A. (2023). Ventral pallidal regulation of motivated behaviors and reinforcement. Front. Neural Circuits 17, 1086053. doi:10.3389/fncir.2023.1086053
Sofuoglu, M., and Sewell, R. A. (2009). Norepinephrine and stimulant addiction. Addict. Biol. 14, 119–129. doi:10.1111/j.1369-1600.2008.00138.x
Solié, C., Contestabile, A., Espinosa, P., Musardo, S., Bariselli, S., Huber, C., et al. (2022). Superior Colliculus to VTA pathway controls orienting response and influences social interaction in mice. Nat. Commun. 13, 817. doi:10.1038/s41467-022-28512-4
Song, Z., Borland, J. M., Larkin, T. E., O'Malley, M., and Albers, H. E. (2016). Activation of oxytocin receptors, but not arginine-vasopressin V1a receptors, in the ventral tegmental area of male Syrian hamsters is essential for the reward-like properties of social interactions. Psychoneuroendocrinology 74, 164–172. doi:10.1016/j.psyneuen.2016.09.001
Stolzenberg, D. S., and Mayer, H. S. (2019). Experience-dependent mechanisms in the regulation of parental care. Front. Neuroendocrinol. 54, 100745. doi:10.1016/j.yfrne.2019.04.002
Substance Abuse and Mental Health Services Administration (2014). Results from the 2013 national survey on drug use and health: summary of national findings, NSDUH series H-48, HHS publication No. (SMA) 14-4863. Rockville, MD: substance abuse and mental health Services administration.
Sun, F., Zeng, J., Jing, M., Zhou, J., Feng, J., Owen, S. F., et al. (2018). A genetically encoded fluorescent sensor enables rapid and specific detection of dopamine in flies, fish, and mice. Cell 174, 481–496. doi:10.1016/j.cell.2018.06.042
Sun, F., Zhou, J., Dai, B., Qian, T., Zeng, J., Li, X., et al. (2020). Next-generation GRAB sensors for monitoring dopaminergic activity in vivo. Nat. Methods 17, 1156–1166. doi:10.1038/s41592-020-00981-9
Thiel, K. J., Okun, A. C., and Neisewander, J. L. (2008). Social reward-conditioned place preference: a model revealing an interaction between cocaine and social context rewards in rats. Drug alcohol dependence 96, 202–212. doi:10.1016/j.drugalcdep.2008.02.013
Trezza, V., Baarendse, P. J., and Vanderschuren, L. J. (2010). The pleasures of play: pharmacological insights into social reward mechanisms. Trends Pharmacol. Sci. 31, 463–469. doi:10.1016/j.tips.2010.06.008
Trezza, V., Baarendse, P. J., and Vanderschuren, L. J. (2014). On the interaction between drugs of abuse and adolescent social behavior. Psychopharmacology 231, 1715–1729. doi:10.1007/s00213-014-3471-z
Uhlmann, A., Ipser, J. C., Wilson, D., and Stein, D. J. (2018). Social cognition and aggression in methamphetamine dependence with and without a history of psychosis. Metab. Brain Dis. 33, 559–568. doi:10.1007/s11011-017-0157-3
Underhill, S. M., Colt, M. S., and Amara, S. G. (2020). Amphetamine stimulates endocytosis of the norepinephrine and neuronal glutamate transporters in cultured locus coeruleus neurons. Neurochem. Res. 45, 1410–1419. doi:10.1007/s11064-019-02939-6
Vanderschuren, L. J., Achterberg, E. M., and Trezza, V. (2016). The neurobiology of social play and its rewarding value in rats. Neurosci. Biobehav. Rev. 70, 86–105. doi:10.1016/j.neubiorev.2016.07.025
Vanderschuren, L. J., Niesink, R. J., and Van Pee, J. M. (1997). The neurobiology of social play behavior in rats. Neurosci. Biobehav. Rev. 21, 309–326. doi:10.1016/s0149-7634(96)00020-6
Vanderschuren, L. J., Trezza, V., Griffioen-Roose, S., Schiepers, O. J., Van Leeuwen, N., De Vries, T. J., et al. (2008). Methylphenidate disrupts social play behavior in adolescent rats. Neuropsychopharmacology 33, 2946–2956. doi:10.1038/npp.2008.10
van Kerkhof, L. W. M., Damsteegt, R., Trezza, V., Voorn, P., and Vanderschuren, LJMJ (2013). Social play behavior in adolescent rats is mediated by functional activity in medial prefrontal cortex and striatum. Neuropsychopharmacology 38, 1899–1909. doi:10.1038/npp.2013.83
van Kerkhof, L. W. M., Trezza, V., Mulder, T., Gao, P., Voorn, P., and Vanderschuren, LJMJ (2014). Cellular activation in limbic brain systems during social play behaviour in rats. Brain Struct. Funct. 219, 1181–1211. doi:10.1007/s00429-013-0558-y
Varlinskaya, E. I., and Spear, L. P. (2008). Social interactions in adolescent and adult Sprague-Dawley rats: impact of social deprivation and test context familiarity. Behav. Brain Res. 188, 398–405. doi:10.1016/j.bbr.2007.11.024
Venniro, M., Marino, R. A. M., Chow, J. J., Caprioli, D., Epstein, D. H., Ramsey, L. A., et al. (2022). The protective effect of social reward on opioid and psychostimulant reward and relapse: behavior, pharmacology, and brain regions. J. Neurosci. 42 (50), 9298–9314. doi:10.1523/JNEUROSCI.0931-22.2022
Volkow, N. D., Koob, G. F., and McLellan, A. T. (2016). Neurobiologic advances from the brain disease model of addiction. N. Engl. J. Med. 374, 363–371. doi:10.1056/NEJMra1511480
Volkow, N. D., and Morales, M. (2015). The brain on drugs: from reward to addiction. Cell 162, 712–725. doi:10.1016/j.cell.2015.07.046
von dem Hagen, E. A. H., Stoyanova, R. S., Baron-Cohen, S., and Calder, A. J. (2013). Reduced functional connectivity within and between 'social' resting state networks in autism spectrum conditions. Soc. Cognitive Affect. Neurosci. 8 (6), 694–701. doi:10.1093/scan/nss053
Wahlstrom, L. C., Scott, J. P., Tuliao, A. P., DiLillo, D., and McChargue, D. E. (2015). Posttraumatic stress disorder symptoms, emotion dysregulation, and aggressive behavior among incarcerated methamphetamine users. J. Dual Diagn 11 (2), 118–127. doi:10.1080/15504263.2015.1025026
Wall, V. L., Fischer, E. K., and Bland, S. T. (2012). Isolation rearing attenuates social interaction-induced expression of immediate early gene protein products in the medial prefrontal cortex of male and female rats. Physiol. Behav. 107 (3), 440–450. doi:10.1016/j.physbeh.2012.09.002
Wang, F., Zhang, J., Yuan, Y., Chen, M., Gao, Z., Zhan, S., et al. (2020). Salience processing by glutamatergic neurons in the ventral pallidum. Sci. Bull. 65, 389–401. doi:10.1016/j.scib.2019.11.029
Wardle, M. C., and de Wit, H. (2014). MDMA alters emotional processing and facilitates positive social interaction. Psychopharmacol. Berl. 231 (21), 4219–4229. doi:10.1007/s00213-014-3570-x
Waxmonsky, J. G., Waschbusch, D., Babinski, D., Humphrey, H., Alfonso, A., Crum, K., et al. (2014). Does pharmacological treatment of ADHD in adults enhance parenting performance? Results of a double-blind randomized trial. Cns Drugs 28, 665–677. doi:10.1007/s40263-014-0165-3
West, E. A., and Carelli, R. M. (2016). Nucleus accumbens core and shell differentially encode reward-associated cues after reinforcer devaluation. J. Neurosci. 36, 1128–1139. doi:10.1523/JNEUROSCI.2976-15.2016
Whitaker, L. R., Degoulet, M., and Morikawa, H. (2013). Social deprivation enhances VTA synaptic plasticity and drug-induced contextual learning. Neuron 77, 335–345. doi:10.1016/j.neuron.2012.11.022
Yan, Z., and Rein, B. (2022). Mechanisms of synaptic transmission dysregulation in the prefrontal cortex: pathophysiological implications. Mol. Psychiatry 27, 445–465. doi:10.1038/s41380-021-01092-3
Young, K. A., Liu, Y., Gobrogge, K. L., Dietz, D. M., Wang, H., Kabbaj, M., et al. (2011). Amphetamine alters behavior and mesocorticolimbic dopamine receptor expression in the monogamous female prairie vole. Brain Res. 1367, 213–222. doi:10.1016/j.brainres.2010.09.109
Young, K. A., Liu, Y., Gobrogge, K. L., Wang, H., and Wang, Z. (2014). Oxytocin reverses amphetamine-induced deficits in social bonding: evidence for an interaction with nucleus accumbens dopamine. J. Neurosci. 34, 8499–8506. doi:10.1523/JNEUROSCI.4275-13.2014
Zahed, G., Roozbakhsh, M., Ashtiani, R. D., and Razjouyan, K. (2022). The effect of long-acting methylphenidate and modafinil on attention and impulsivity of children with ADHD using a continuous performance test: a comparative study. Iran. J. Child Neurology 16, 67–77. doi:10.22037/ijcn.v16i2.32541
Zell, V., Steinkellner, T., Hollon, N. G., Warlow, S. M., Souter, E., Faget, L., et al. (2020). VTA glutamate neuron activity drives positive reinforcement absent dopamine Co-release. Neuron 107, 864–873. doi:10.1016/j.neuron.2020.06.011
Keywords: psychostimulants, social behavior, reward, amphetamine, methamphetamine, cocaine, modafinil
Citation: Cid-Jofré V, Bahamondes T, Zúñiga Correa A, Ahumada Arias I, Reyes-Parada M and Renard GM (2024) Psychostimulants and social behaviors. Front. Pharmacol. 15:1364630. doi: 10.3389/fphar.2024.1364630
Received: 02 January 2024; Accepted: 01 April 2024;
Published: 25 April 2024.
Edited by:
Mónica Moreira-Rodrigues, School of Medicine and Biomedical Sciences (ICBAS), PortugalReviewed by:
Alexies Dagnino-Subiabre, Universidad de Valparaiso, ChileCopyright © 2024 Cid-Jofré, Bahamondes, Zúñiga Correa, Ahumada Arias, Reyes-Parada and Renard. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Miguel Reyes-Parada, bWlndWVsLnJleWVzQHVzYWNoLmNs; Georgina M. Renard, Z2VvcmdpbmEucmVuYXJkQHVzYWNoLmNs
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.