
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Pharmacol., 05 June 2024
Sec. Neuropharmacology
Volume 15 - 2024 | https://doi.org/10.3389/fphar.2024.1364353
This article is part of the Research TopicRecent advances in the treatment of epilepsyView all 13 articles
 Salim M. Bastaki1,2*
Salim M. Bastaki1,2* Yousef M. Abdulrazzaq3
Yousef M. Abdulrazzaq3 Marwan Abdelrahim Zidan4Mohamed Shafiullah1,2Saif Ghdayer Alaryani5
Marwan Abdelrahim Zidan4Mohamed Shafiullah1,2Saif Ghdayer Alaryani5 Fatima Awad Alnuaimi5
Fatima Awad Alnuaimi5 Ernest Adeghate5
Ernest Adeghate5 Sahar Mohsin5Amal Akour1
Sahar Mohsin5Amal Akour1 Agata Siwek6
Agata Siwek6 Dorota Łażewska7
Dorota Łażewska7 Katarzyna Kieć-Kononowicz7
Katarzyna Kieć-Kononowicz7 Bassem Sadek1,2*
Bassem Sadek1,2*Introduction: Brain histamine is considered an endogenous anticonvulsant and histamine H1 receptor. H1R antagonists have, in earlier studies, been found to induce convulsions. Moreover, research during the last two decades has provided more information concerning the anticonvulsant activities of histamine H3R (H3R) antagonists investigated in a variety of animal epilepsy models.
Methods: Therefore, the in vivo anticonvulsant effect of the H3R antagonist DL76, with proven high in vitro affinity, in vitro selectivity profile, and high in vivo antagonist potency in mice against maximal electroshock (MES)-induced seizures in mice, was assessed. Valproic acid (VPA) was used as a reference antiepileptic drug (AED). In addition, DL76 was tested for its reproductive and fetal toxicity in the same animal species.
Results and discussion: Our observations showed that acute systemic administration (intraperitoneal; i.p.) of DL76 (7.5 mg/kg, 15 mg/kg, 30 mg/kg, and 60 mg/kg, i.p.) provided significant and dose-dependent protection against MES-induced seizures in female and male mice. Moreover, the DL76-provided protective effects were comparable to those offered by the VPA and were reversed when animals were co-administered the CNS-penetrant selective H3R agonist R-(α)-methylhistamine (RAM, 10 mg/kg, i.p.). Furthermore, the administration of single (7.5 mg/kg, 15 mg/kg, 30 mg/kg, or 60 mg/kg, i.p.) or multiple doses (3 × 15 mg/kg, i.p.) of H3R antagonist DL76 on gestation days (GD) 8 or 13 failed to affect the maternal body weight of mice when compared with the control mice group. No significant alterations were detected in the average number of implantations and resorptions between the control and DL76-treated groups at the early stages of gestation and the organogenesis period. In addition, no significant differences in the occurrence of skeletal abnormalities, urogenital abnormalities, exencephaly, exomphalos, facial clefts, and caudal malformations were observed. The only significant abnormalities witnessed in the treated groups of mice were in the length of long bones and body length. In conclusion, the novel H3R antagonist DL76 protected test animals against MES-induced seizures and had a low incidence of reproductive and fetal malformation with decreased long bone lengths in vivo, signifying the potential therapeutic value of H3R antagonist DL76 for future preclinical as well as clinical development for use in the management of epilepsy.
Epilepsy is characterized by seizures that are unpredictable in frequency and is the second most common neurological disorder that affects people of all ages, with onset most often occurring in childhood and older adulthood (Mauritz et al., 2022; Yang et al., 2022; Asadi-Pooya et al., 2023). Epilepsy leads to abnormal behavioral paradigms, necessitating a lifelong treatment with effective antiepileptic drugs (AEDs) (Brigo et al., 2013; Mauritz et al., 2022). The anticipated prevalence rate of epilepsy in the United States and other regions is high. Approximately 1.5 million women with epilepsy give birth to 27,000 infants yearly, with an estimated prevalence of epilepsy in pregnant women of approximately 0.5% (Tomson and Battino, 2007; Thomas et al., 2012; Sadek et al., 2014a; Sadek et al., 2015; Bastaki et al., 2018; Tomson et al., 2018). Clinical observations revealed that approximately 60%–70% of patients diagnosed with epilepsy respond to available treatments with AEDs. However, resistance to monotherapy is not uncommon, and a combination of several AEDs is often inevitable with the possibility of drug–drug interactions (Brigo et al., 2013; Sadek et al., 2014a). Following chronic clinical use of several AEDs, the incidence of major malformations in infants of epileptic parents was found to be twice that of non-epileptic parents (Sadek et al., 2015; Bastaki et al., 2018). In addition, gestational epilepsy is a concern for women with a pre-existing seizure and can lead to a 17% rise in seizure episodes (Tomson et al., 2018). Accordingly, hormonal fluctuations and changes in the pharmacokinetics of AEDs play a crucial role (Thomas et al., 2012). Therapeutic drug monitoring and maintaining strict adherence to AEDs have improved therapeutic management (Pennell et al., 2020), yet the threat of complications for both mother and child persists. Epilepsy can increase maternal mortality fivefold, and in utero exposure to AEDs, notably valproic acid (VPA), increases the risk of congenital defects, which is also related to drug type and dosage and is worsened by specific drug combinations (Viale et al., 2015).
The involvement of the brain histaminergic neurotransmitter system in seizure control is well-established, as several previous observations in preclinical experiments demonstrated that brain histamine is capable of modulating seizure trends in both electrically and chemically induced seizure models in animals (Kamei et al., 1993a; Kamei et al., 1993b; Chen et al., 2002; Zhang et al., 2003; Sadek et al., 2014b). Accordingly, the precursor of brain histamine, namely, the amino acid L-histidine, was reported to decrease chemically induced seizure in experimental animals by stimulating the brain histaminergic neurotransmission and increasing the seizure threshold by interactions of brain histamine with postsynaptically located histamine H1 receptors (H1Rs) in the brain (Chen et al., 2002). Moreover, the involvement of central H1Rs in the development of seizures was confirmed by preclinical studies that showed an increased tendency of seizures in mice lacking H1Rs or histidine decarboxylase enzyme responsible for the biosynthesis of histamine in the brain (Jang et al., 2010; Miyata et al., 2011). Clinically, it has been found that the use of high doses of various centrally acting H1R antagonists belonging to old-generation antihistamines, such as diphenhydramine, as antiallergic drugs occasionally increased the risk of convulsions in healthy young children, and this risk was especially observed among long-term users (Ago et al., 2006; Jang et al., 2010; Miyata et al., 2011). Histamine interacts with four G-protein-coupled H1-4R subtypes. H3R, first described in 1983, was found to regulate histamine biosynthesis and release, acting as presynaptic auto-receptors (Arrang et al., 1983; 1987; Brown et al., 2001). Accordingly, H3R antagonists or histamine N-methyl transferase (HNMT) inhibitors (e.g., methoprene) were found to increase brain histamine levels and decrease seizures in epileptic patients through interactions with H1Rs (Witkin and Nelson, 2004; Vohora et al., 2010). Hence, the potential of H3R antagonists as future AEDs has begun to be increasingly considered, as mounting evidence from both acute and chronic experimental seizure models demonstrated the anticonvulsant efficacy of numerous imidazole- and non-imidazole-based H3R antagonists, such as thioperamide, 2–18, DL77, E159, and E169 (Brown et al., 2001; Vohora et al., 2010; Witkin and Nelson, 2004; Sadek et al., 2015; Bastaki et al., 2018).
Based on the afore-mentioned preclinical and clinical findings, and in search of potent and selective histamine H3R antagonists with anticonvulsant properties, our research group succeeded in developing the histamine H3R antagonist DL76 [1-(3-(4-tert-butyl-phenoxy) propyl) piperidine] (Figure 1), a non-imidazole-based H3R ligand, which proved to be a highly potent H3R antagonist (human H3R Ki = 22 nM) with good selectivity profile vs. other histamine receptors and high oral potency in mice with an ED50 value of 2.8 ± 0.4 mg/kg (Lazewska et al., 2006; Lazewska et al., 2017; Lazewska et al., 2020; Canale et al., 2021) (Figure 1). Moreover, previous research works clearly showed antiparkinsonian effects of H3R antagonist DL76 without inducing in vitro toxicity when tested in several cell lines (Canale et al., 2021). In a previous preclinical pharmacokinetic evaluation study in experimental animals, concentrations of DL76 in the cortex, hippocampus, and striatum were found to be significantly higher (44%) than in the whole brain tissues. Following administration of the doses, C0 values increased from 564 ng/mL (3 mg/kg) to 653 ng/mL (6 mg/kg), while AUC0→∞ was found to be doubled when the tested dose increased from 3 mg/kg to 6 mg/kg (298 ng◦h/mL vs. 685 ng◦h/mL) (Szafarz et al., 2015). Therefore, the H3R antagonist DL76, which has a high in vitro H3R antagonist affinity, an excellent in vitro selectivity profile, a high in vivo H3R antagonist potency, and pharmacokinetics and tissue distribution profiles, was selected for further investigations of its anticonvulsant efficacy in MES-induced seizure models in male and female adult mice. Possible reproductive toxicities in the same animal species were assessed following systemic administration of H3R antagonist DL76 in a wide range of doses.

Figure 1. Chemical structure, in vitro affinities, and in vivo H3R antagonist potency of DL76. aData from Lazewska et al. (2006). bMeasured according to a previous protocol (Lazewska et al., 2017). cRadioligand binding studies were performed commercially in Eurofins Cerep Laboratories (Celle-Lévescault, France). DData from Lazewska et al. (2020) and Canale et al. (2021). eMeasured according to a previous protocol (Canale et al., 2021).
Experiments utilized 8–10-week-old C57BL/6J mice (30–35 g) from The Jackson Laboratory, referred to as C57 mice, of both sexes. Mice were group-housed in standard Plexiglas cages with ad libitum access to food and water in a 12-h light/dark cycle (lights on at 6:00 a.m.). Procedures were conducted between 9:00 a.m. and 12:00 p.m., following guidelines of the European Communities Council Directive (86/609/EEC) and approved by the Institutional Animal Ethics Committee for epilepsy (ERA-2017-5535) and teratogenic studies (A14-14) at the College of Medicine and Health Sciences/United Arab Emirates University.
The reference AED VPA and the CNS-penetrant H3R agonist (R)-α-methylhistamine (RAM, 10 mg/kg, i.p.) were obtained from Sigma-Aldrich (St Louis, Missouri, USA). The test compound DL76 was synthesized by the Department of Technology and Biotechnology of Drugs (Kraków, Poland) according to previously described synthetic protocols (Lazewska et al., 2006). The test compound DL76, the reference drug VPA (300 mg/kg), and the CNS-penetrant H3R agonist RAM (10 mg/kg) were dissolved in isotonic saline and administered intraperitoneally (i.p.) at a volume of 1 mL/kg for all in vivo studies. DL76 was used in a dose range of 7.5 mg/kg, 15 mg/kg, 30 mg/kg, and 60 mg/kg. All doses of used drugs were expressed in terms of the free bases. Eight mice were used to assess the anticonvulsant activity of each compound.
As previously described and with a slight modification in the intensity applied, seizures were elicited in mice with a 50-Hz alternating current of 120 mA intensity (Kaminski et al., 2015; Song et al., 2020; Xiao et al., 2021; Hua et al., 2023). The current was applied through ear electrodes for 0.2 s. Protection against the spread of MES-induced convulsion was defined as the abolition of the tonic hind limb extension (THLE) component of the seizure (Kaminski et al., 2015; Song et al., 2020; Xiao et al., 2021; Hua et al., 2023). The animals were divided into the following experimental groups, with eight mice in each: the control group injected with saline, the positive control group injected with 300 mg/kg of VPA (the minimal dose of VPA that protected animals against the spread of MES-induced convulsions without mortality in mice) (Sadek et al., 2015; Bastaki et al., 2018), and four groups that were administered the test compound DL76 at doses of 7.5 mg/kg, 15 mg/kg, 30 mg/kg, or 60 mg/kg. All drugs were administered 30–45 min before the MES challenge. In a further group of eight mice, the most promising and protective dose of DL76 (60 mg/kg, i.p.) was co-injected with RAM (10 mg/kg, i.p.), 30 min apart and 15 min prior to the MES test. Protection against the spread of MES-induced convulsion was defined as the abolition of the THLE component of the convulsion (Sadek et al., 2015; Alachkar et al., 2018; Bastaki et al., 2018).
Reproductive studies were conducted in adult female mice, averaging 30 gm in weight and 6 weeks in age (Padmanabhan et al., 2006; Bastaki et al., 2018). Following successful mating, indicated by vaginal plug observation, the plug-positive day was considered gestation day 0 (GD-0). DL76 was administered at various doses (7.5 mg/kg, 15 mg/kg, 30 mg/kg, and 60 mg/kg) through single i.p. injections and three 15 mg/kg injections in 1 day to mouse groups on GD-8 and GD-13 (Tables 1, 2). The control group was injected with normal saline. Appropriate formation of different organs occurs in specific periods of the development of the mouse. These two periods were selected because GD-8 is known to be a critical period for the induction of neural tube defects, craniofacial malformations, and the development of other organs, whereas on GD-13, the retina and other parts are being formed, and most other organs have been formed. The number of implantations, fetal deaths, and resorptions was recorded. Fetal and placental weights were documented separately. After blotting dry, the fetuses were weighed and fixed in 95% ethanol. The reproductive toxicological effects of DL76 were assessed by observing gross and visceral malformations, following Sterz and Lehmann (1985), Abdulrazzaq et al. (1997), Padmanabhan et al. (2003), and Padmanabhan et al. (2006).

Table 1. Morphological effect of systemic treatment of H3R antagonist DL-76 on gestation day (GD-8) in C57/B6 mouse embryos.
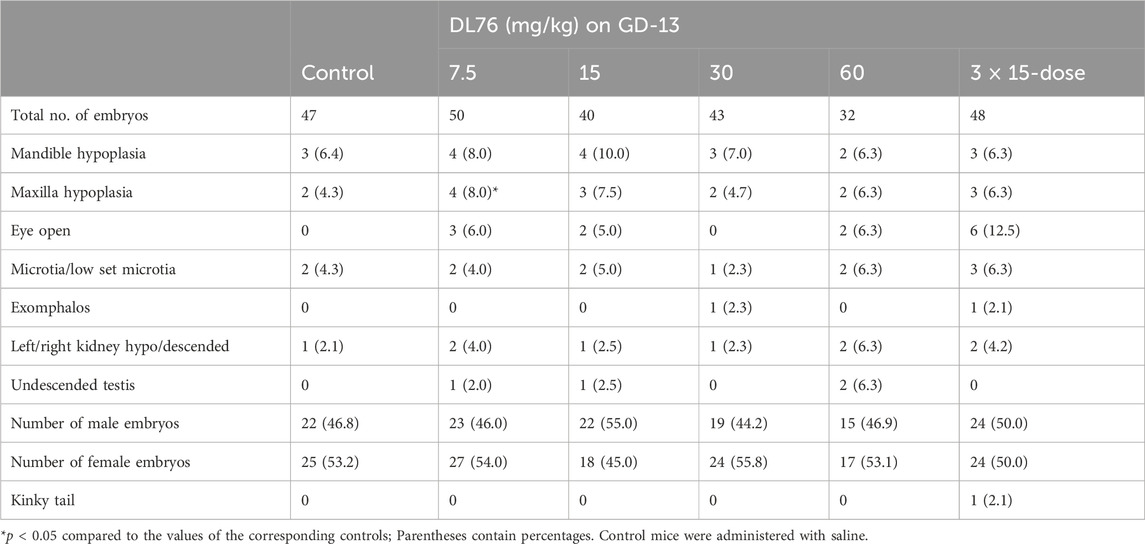
Table 2. Morphological effect of systemic treatment of drug DL-76 on gestation day (GD-13) in C57/B6 mouse embryos.
Deformities of the embryos were identified according to a modified method of Sterz and Lehmann (1985), Abdulrazzaq et al. (1997), and Bass et al. (2020). Accordingly, the embryos were removed from the 95% ethanol, and the lower part of the abdomen was cut using a razor blade. All organs were checked for abnormalities with a stereo dissecting microscope, and finally, the organs were removed by using forceps.
After isolating fetuses from viscera and skin, they underwent a fat-removal process by immersion in acetone for 1–3 days. The resulting transparent specimens underwent a sequence of glycerin solutions (50% and 80%) before being finally stored in 100% glycerin for observation of malformations using a stereo dissecting microscope (McLeod, 1980; Sterz and Lehmann, 1985; Abdulrazzaq et al., 1997; Bass et al., 2020). Following this, specimens were processed and stained with Alizarin Red-S and Alcian blue (McLeod, 1980; Sterz and Lehmann, 1985; Abdulrazzaq et al., 1997; Bass et al., 2020) to identify bone and cartilage deformities.
Statistical analyses employed SPSS 26.0 (IBM Middle East, Dubai, UAE). Data were presented as means ± SEM. Convulsion effects, measured by THLE in seconds, and reproductive toxicities were assessed using one-way ANOVA with treatment (vehicle or test compound) and dose (test compound) as between-subjects factors, followed by least significant difference (LSD) post hoc analysis. Fetal abnormality data were presented as counts and percentages. Fisher’s exact test compared the percentages of abnormalities between the control group (saline) and the different dose groups. All tests were two-tailed, and significance was set at p < 0.05.
The acute systemic administration of a dose range of DL76 (7.5 mg/kg, 15 mg/kg, 30 mg/kg, and 60 mg/kg, i.p.) exhibited a protective effect on MES-induced convulsions [F (7,56) = 16.54; p < 0.0001] (Figure 2). The results show that there are substantial protective effects of all doses of DL76 (i.e., 7.5 mg/kg, 15 mg/kg, 30 mg/kg, and 60 mg/kg when compared with saline (all p values <0.0001) (Figure 2). Among the doses tested, the 60 mg/kg dose of DL76 showed the most promising protection in an MES model in adult male mice when compared with the saline-, DL76 (7.5 mg/kg)-, DL76 (15 mg/kg)-, and DL76 (30 mg/kg)-treated groups [mean difference of THLE 6.25 s, 7.25 s, 8.88 s, and 10.75 s; respectively, all p < 0.0001] (Figure 2). Notably, the protection provided by H3R antagonist DL76 at the higher dose (60 mg/kg, i.p.) was comparable to that provided by the reference drug VPA (p = 1.000) (Figure 2) and was reversed following co-injection with the CNS-penetrant histamine H3R agonist RAM (10 mg/kg, i.p.) 15 min before the MES challenge, with a mean difference of 2.5 s (p-value = 0.094), for the comparison of saline vs. DL76+RAM (Figure 2). Interestingly, RAM (10 mg/kg, i.p.), when injected alone, did not affect MES-induced convulsions (p = 0.602 saline + saline vs. DL76 (60 mg/kg) + RAM) (Figure 2). Figure 2 represents the significant differences between all doses of DL76 and compared to VPA (300 mg/kg), RAM, and DL76 60 mg/kg + RAM observed in adult male mice.
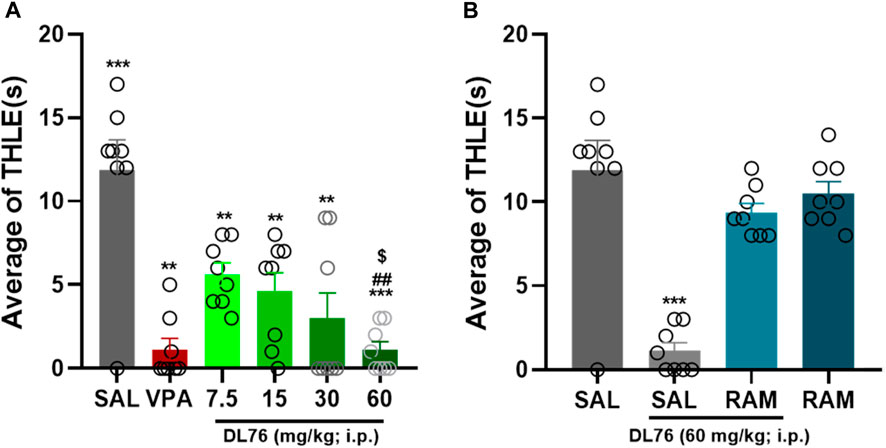
Figure 2. Anticonvulsant effect of acute systemic administration of H3R antagonist DL76 on MES-induced seizure in male adult mice. (A) Anticonvulsant effects of valproic acid (VPA, 300 mg/kg, i.p.) and test compound DL76 (7.5 mg/kg, 15 mg/kg, 30 mg/kg, and 60 mg/kg, i.p.) on the duration of tonic hind limb extension (THLE) induced in an MES model in male adult mice. (B) Effect of CNS-penetrant H3R agonist RAM (10 mg/kg, i.p.) pretreatment on the protection by H3R antagonist DL76 (60 mg/kg, i.p.) on MES-induced convulsions in male adult mice. Each value represents mean ± SEM (n = 8). ***p < 0.001 versus saline- and DL76 (7.5 mg/kg)-treated groups. ##p < 0.001 versus DL76 (7.5 mg/kg)- and DL76 (15 mg/kg)-treated groups. $p < 0.05 versus DL76 (30 mg/kg)-treated group.
The results showed that VPA (300 mg/kg) and DL76 (7.5–60 mg/kg, i.p.) significantly protected animals against MES-induced convulsions [F (7,56) = 51.78; p < 0.0001] (Figure 3). In addition, the results showed that DL76 at a dose of 60 mg/kg significantly provided the highest protection in an MES model in female adult mice when compared with the saline-, DL76 (7.5 mg/kg)-, DL76 (15 mg/kg)-, and DL76 (30 mg/kg)-treated groups [mean difference in THLE of 12.13 s, 10.63 s, and 3.13 s; respectively (p < 0.05)] (Figure 3). Similarly, the protection provided by DL76 at the highest dose used in the current study (60 mg/kg, i.p.) was comparable to that provided by the reference drug VPA (300 mg/kg) (p = 0.72) and was abrogated following acute systemic co-administration of RAM (10 mg/kg, i.p.) 15 min before the MES challenge [mean difference = 1.33; p = 0.17, for the comparison of saline + saline vs. DL76 + RAM] (Figure 3). Data showed that RAM alone in female adult mice generated results similar to those witnessed in male adult mice (10 mg/kg, i.p.; p = 0.91 saline-saline vs. saline-RAM) (Figure 3). Interestingly, no significant differences were observed for the protective effects provided in male and female mice after acute systemic administration of VPA (300 mg/kg, i. p.) or H3R antagonist DL76 at doses of 7.5 mg/kg, 15 mg/kg, 30 mg/kg, and 60 mg/kg, i.p. [F (1,14) = 2.15; p = 0.164, for the comparison of saline + VPA in male mice vs. saline + VPA in female mice], [F (1,14) = 0.156; p = 0.70, for the comparison of saline + DL76 (7.5 mg/kg) in male mice vs. saline + DL76 (7.5 mg/kg) in female mice], [F (1,14) = 0.17; p = 0.685, for the comparison of saline + DL76 (15 mg/kg) in male mice vs. saline + DL76 (15 mg/kg) in female mice], [F (1,14) = 2.31; p = 0.151, for the of comparison saline + DL76 (30 mg/kg) in male mice vs. saline + DL76 (30 mg/kg) in female mice], and [F (1,14) = 0.76; p = 0.398, for the comparison of saline + DL76 (60 mg/kg) in male mice vs. saline + DL76 (60 mg/kg) in female mice], respectively.
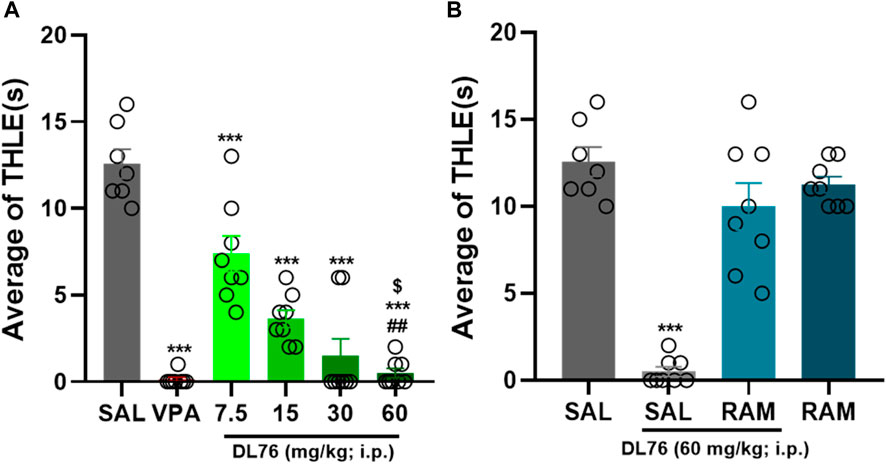
Figure 3. Anticonvulsant effect of acute systemic administration of H3R antagonist DL76 on MES-induced seizure in female adult mice. (A) Anticonvulsant effects of valproic acid (VPA, 300 mg/kg, i.p.) and test compound DL76 (7.5 mg/kg, 15 mg/kg, 30 mg/kg, and 60 mg/kg, i.p.) on the duration of tonic hind limb extension (THLE) induced in an MES model in female adult mice. (B) Effect of CNS-penetrant H3R agonist RAM (10 mg/kg, i.p.) pretreatment on the protection by H3R antagonist DL76 (60 mg/kg, i.p.) on MES-induced convulsions in female adult mice. Each value represents mean ± SEM (n = 8). ***p < 0.001 versus saline- and DL76 (7.5 mg/kg)-treated groups. ##p < 0.001 versus DL76 (7.5 mg/kg)- and DL76 (15 mg/kg)-treated groups. $p < 0.05 versus DL76 (30 mg/kg)-treated group.
There was no sign of maternal toxicity in any of the mice from all groups in both the GD-8 and GD-13 treated animals. The number of fetuses in each litter was not significantly different between all study groups, nor was the number of live fetuses. The resorption rate was not significantly different between all the groups. The mean fetal weight in each litter was not significantly different between all treatment groups and the controls. The mean placental weights were also not significantly different between the treated groups and the controls. The incidence of gross morphological anomalies in the treated fetuses of the single- and multiple-dose groups given i.p. was not significantly different from that in the control group. There was no significant difference in the incidence of exencephaly and craniofacial malformations, such as mandibular and maxillary hypoplasia, exomphalos, low set microtia, exophthalmia, eye remaining open, posterior bilateral palate, posterior unilateral palate, hydronephrosis, descended kidney, undescended testis, or kinky tail, between any of the groups studied (Tables 1, 3).

Table 3. Effect of systemic treatment of H3R antagonist DL-76 on gestation day (GD-8) in C57/B6 mouse embryos.
Observations of skeletal defects showed that mean crown-to-rump length decreased significantly in the 7.5 mg/kg-, 30 mg/kg-, and 60 mg/kg-treated groups when compared to control groups. Humerus and ulna lengths were reduced in the 30 mg/kg and 60 mg/kg groups, with only the 30 mg/kg group showing a significant decrease in radius length. Femur length significantly decreased in the 7.5 mg/kg and 60 mg/kg groups; a non-significant decrease (p = 0.06) occurred in the 30 mg/kg group. Tibia and fibula lengths significantly decreased in the 7.5 mg/kg, 30 mg/kg, and 60 mg/kg groups when compared to the controls (Tables 1, 2). No significant differences were observed in skull or facial bones (Tables 1, 2). In addition, in the vertebrae, our observed results showed that there were no differences in the number of centra or the number of unossified/hypoplastic bodies in all the DL-76-treated groups. There was no hypoplasia or poor ossification of the caudal vertebrae in any of the groups. The number of coccygeal vertebral bodies was the same in all tested mice groups (Tables 1, 2). The cervical ribs were attached to the seventh cervical vertebra and ended free anteriorly, becoming the first thoracic ribs. The lumbar ribs attached to the first lumbar vertebra were shorter than the last pair of thoracic ribs. The incidence of lumbar ribs in the DL76-treated group was not significantly different when compared to the saline-treated control mice groups. In addition, there was no fusion of ribs, reduction in number and size, or splitting and forking of ribs in either the control or the DL76-treated mice groups. Notably, 13 pairs of thoracic ribs were observed in both control and experimental embryos (Tables 1, 2). Furthermore, the sternum of the control fetuses consisted of seven sternebrae. There was no significant difference in the fifth sternebrium between the DL76-treated and control mice groups with regards to the absence or the occurrence of hypoplasia. We did not observe any malalignment, hemilateral or unilateral agenesis, scrambling, or fusion, or any cases of bifid sternebrae in any of the groups (Tables 1, 2). Figure 4 shows some of the malformations seen in the embryos.
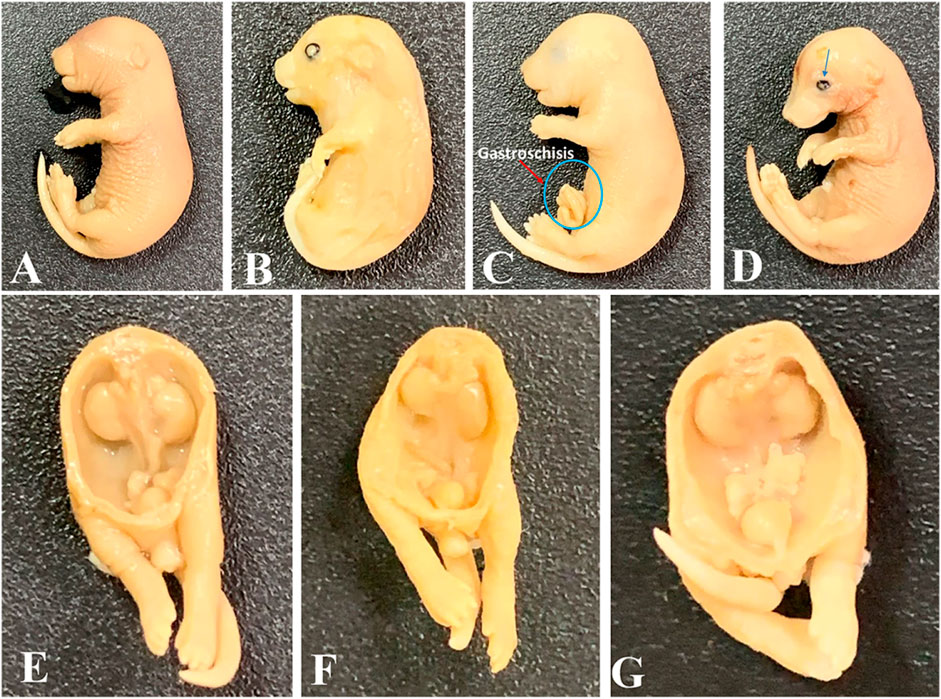
Figure 4. Some abnormalities detected in tested mice embryos at gestational day 18. (A) Normal embryo; (B) mandibular and maxillary hypoplasia with an open eye; (C) embryo with gastroschisis; (D) intrauterine growth restriction (IUGR) and open eye; (E) normal urogenital system; (F) embryo with a hypoplastic right kidney and a descended left kidney; (G) embryo with undescended testes. Magnification: ×1.25.
H3R antagonist DL76 showed very promising results in terms of its ability to exert protective effects against MES-induced convulsions in male and female adult mice, especially when mice were pretreated with 60 mg/kg i.p., as compared with the saline-treated groups or other mice groups that received lower doses (Figures 2–4). Therefore, the results demonstrated a dose–response relationship of the protective effects provided by H3R antagonist DL76 against convulsions in adult mice of both sexes (Figure 2, 3). Notably, the DL76 (60 mg/kg)-provided protective effect was similar to that observed for the reference drug VPA (300 mg/kg, i.p.) (Figure 2, 3). The latter observations are in harmony with previous preclinical studies that showed dose-dependent protective effects of a couple of H3R antagonists against MES convulsions induced in several animal models (Sadek et al., 2013; Sadek et al., 2014a; Sadek et al., 2014b; Sadek et al., 2015; Sadek et al., 2016a; Sadek et al., 2016b).
In a very recent study, a group of compounds known as 6-aminoalkoxy-3,4-dihydroquinolin-2(1H)-ones were synthesized to evaluate their potential as H3R antagonists and their ability to prevent seizures (Hua et al., 2023). The synthesized compounds exhibited strong H3R antagonism. Specifically, compounds labeled 2a, 2c, 2h, and 4a demonstrated exceptionally potent H3R antagonistic activities, with half-maximal inhibitory concentrations (IC50) of 0.52 μM, 0.47 μM, 0.12 μM, and 0.37 μM, respectively. When tested in the MES model, three compounds—2h, 4a, and 4b—were identified as having antiseizure properties. Molecular docking studies involving compounds 2h, 4a, and a reference compound PIT with the H3R protein were conducted to predict how these molecules might interact with the H3R binding site. The docking results indicated that 2h, 4a, and PIT likely share a similar binding mode to the H3R, providing insights into the molecular basis for their antagonistic effects (Hua et al., 2023).
In addition, several non-imidazole H3R antagonists have been tested and, of these, some, such as E159 and E177, emerged as particularly effective, reducing the duration of THLE in a dose-dependent manner in MES-induced seizures in rats (Sadek et al., 2014b; Alachkar et al., 2018). Previous clinical studies showed that the H3R antagonist PIT exhibited protective effects against the photosensitivity seizure model in adult patients (Lamb, 2020).
The current study showed that the protection observed for H3R antagonist DL76 was almost nullified when animals were co-injected with the CNS-penetrant histamine H3R agonist RAM (10 mg/kg i.p.) (Figure 2B, 3B). However, when injected alone, RAM (10 mg/kg) showed neither a protective nor an epileptogenic activity in mice challenged by the MES-induced convulsion (Figure 2B, 3B). These results suggest that the protective effects of H3R antagonist DL76 in the MES-induced convulsions are mediated, at least in part, through an H3R blockade, which is consistent with the formerly observed protective activities of various H3R antagonists (Brigo et al., 2013; Sadek et al., 2014b; Mauritz et al., 2022). These later effects can be devised from the inhibitory effect of H3Rs on the biosynthesis and release of histamine in the presynaptic histaminergic terminals (Sadek et al., 2014b). Subsequently, blockage of these H3 auto-receptors by selective antagonists, such as our compound H3R antagonist DL76, would result in an enhanced neuronal release of brain histamine, providing the proposed protective effect in the MES-induced convulsion status in mice. Notably, the abrogation effect on numerous previously assessed H3R antagonists brought about either by H3R agonists or by CNS-penetrant H1- or H2R antagonists was described, providing further evidence about the role of the H3R antagonism-released brain histamine (Sadek et al., 2013; Sadek et al., 2014a; Sadek et al., 2014b; Sadek et al., 2015; Sadek et al., 2016a; Sadek et al., 2016b). Interestingly, no significant differences were observed for the reference drug VPA (300 mg/kg, i.p.) and the 30 mg/kg and 60 mg/kg doses of the H3R antagonist DL76 in both sexes of adult mice, but there was a significant difference between the THLE values of VPA and the lower doses of DL76 (i.e., the 7.5 mg/kg, and 15 mg/kg, i.p.). The latter results are discrepant with previous studies in which there was a variation in seizure threshold between both sexes due to differences in levels of steroids and steroidal derivatives, such as 3α-hydroxylated pregnane steroids, which can interact with GABA receptor complex, and therefore, decrease seizure susceptibility in female adult mice (Belelli et al., 1990a; Belelli et al., 1990b; Lan et al., 1990; Maguire et al., 2005). In a further series of experiments, H3R antagonist DL76 was administered at various doses to groups of mice on gestational days 8 and 13 (Tables 1–4). The observed results showed that there was no difference in the abnormalities that occurred in the fetuses. The latter observation is crucial, as all AEDs, including the third-generation agents given to pregnant women, have been shown to have deleterious effects on fetuses in numerous worldwide studies (Abdulrazzaq et al., 1997; Holmes et al., 2001; Padmanabhan et al., 2003; Kaplan, 2004; Padmanabhan et al., 2006).
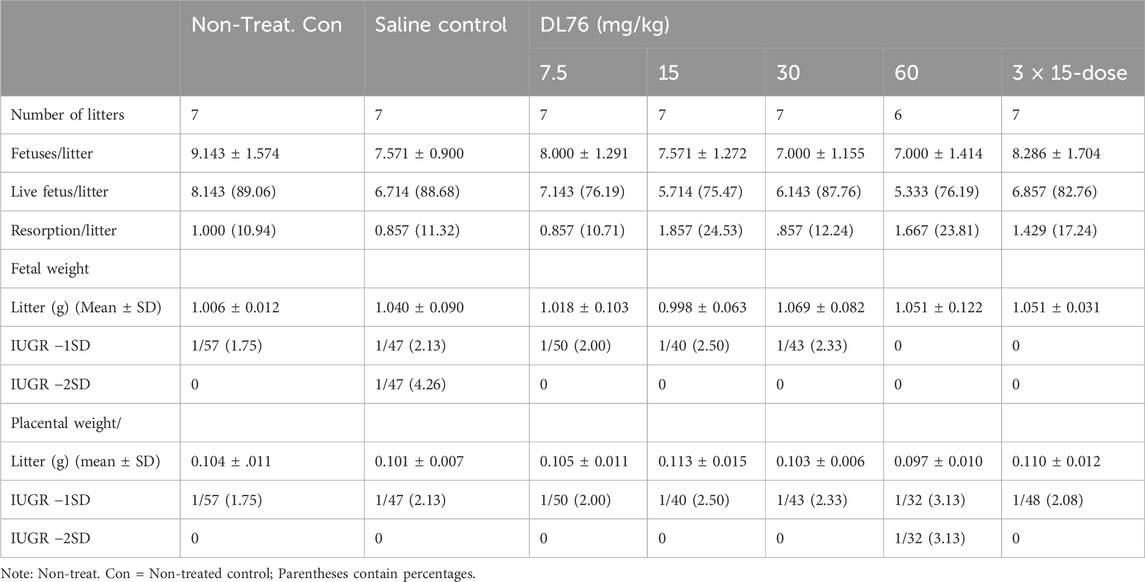
Table 4. Effect of systemic treatment of H3R antagonist DL-76 on gestation day (GD-13) in C57/B6 mouse embryos.
Although some abnormalities were detected in some of the embryos, their numbers were not different than those in the control groups, except for the crown–rump length and lengths of the long bones of the extremities, which seemed to be affected at the higher doses of 30 mg/kg, 60 mg/kg, and 15 mg/kg × 3. No significant signs of maternal toxicity or fetal resorption were seen in the treated groups (Figure 4). The absence of significant abnormalities in the embryos of mice treated with H3R antagonist DL76 in this study is similar to the lack of teratogenic effects of H3R antagonist 2–18 on mice; the only difference is the adverse effects on the bones of the extremities, which seemed to be shorter in the higher dose groups than the controls (Bastaki et al., 2018). Many women abandon AEDs during pregnancy for fear of their effects on the fetus and are, therefore, at great risk of seizure occurrence and morbidity in both the mother and fetus. The H3R antagonist DL76 appears to be relatively safe except for its effects on the length of the long bones of the extremities at high doses. One of the limitations of this study was that it should have compared the antiseizure and anti-toxicity activities of DL76 to a reference H3R antagonist drug. This will be considered in future experiments.
Unfortunately, it is difficult to extrapolate these findings to humans, and therefore, it is difficult to select a dose that would be safe in humans. Considering these findings, it is reasonable to conclude that the safety profile of H3R antagonist DL76 is obviously higher than that of the other AEDs and comes close to the safety profile of other H3R antagonists like 2–18 (Bastaki et al., 2018).
In conclusion, the H3R antagonist DL76 shows the prospect of being a useful protective anticonvulsant drug with potential use during pregnancy, given the fact that AED VPA was described to induce malformations in C57BL/6 mice (Chateauvieux et al., 2010). Nevertheless, further preclinical studies are necessary to determine if these findings can be replicated when administering the H3R antagonist DL76 throughout the entire gestation period, both in the same species and in other animal models.
The original contributions presented in the study are included in the article/Supplementary Material; further inquiries can be directed to the corresponding authors.
The animal study procedures were conducted following guidelines of the European Communities Council Directive (86/609/EEC) and approved by the Institutional Animal Ethics Committee for epilepsy (ERA-2017-5535) and teratogenic studies (A14-14) at the College of Medicine and Health Sciences/United Arab Emirates University. The study was conducted in accordance with the local legislation and institutional requirements.
SB: conceptualization, investigation, supervision, writing–original draft, writing–review and editing, data curation, formal analysis, methodology, software, and validation. YA: investigation, writing–original draft, and writing–review and editing. MZ: formal analysis, investigation, validation, writing–original draft, and writing–review and editing. MS: investigation, writing–original draft, and writing–review and editing. SA: investigation, writing–original draft, and writing–review and editing. FA: investigation, writing–original draft, and writing–review and editing. EA: investigation, writing–original draft, and writing–review and editing. SM: investigation, writing–original draft, and writing–review and editing. AA: writing–original draft and writing–review and editing. AS: investigation, writing–original draft, and writing–review and editing. DL: investigation, writing–original draft, and writing–review and editing. KK-K: investigation, writing–original draft, and writing–review and editing. BS: conceptualization, data curation, formal analysis, funding acquisition, investigation, methodology, project administration, resources, software, supervision, validation, visualization, writing–original draft, and writing–review and editing.
The author(s) declare that financial support was received for the research, authorship, and/or publication of this article. The Office of Graduate Studies and Research of UAE University is thanked for the support provided to BS with funds (12M099, 12R207). This study was also supported by statutory funds from the Faculty of Pharmacy Jagiellonian University Medical College, Kraków, Poland [N42/DBS/000300 (DL)].
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors, and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Abdulrazzaq, Y. M., Bastaki, S. M., and Padmanabhan, R. (1997). Teratogenic effects of vigabatrin in TO mouse fetuses. Teratology 55, 165–176. doi:10.1002/(SICI)1096-9926(199703)55:3<165::AID-TERA1>3.0.CO;2-1
Ago, J., Ishikawa, T., Matsumoto, N., Ashequr Rahman, M., and Kamei, C. (2006). Mechanism of imipramine-induced seizures in amygdala-kindled rats. Epilepsy Res. 72, 1–9. doi:10.1016/j.eplepsyres.2006.06.022
Alachkar, A., Lazewska, D., Latacz, G., Frank, A., Siwek, A., Lubelska, A., et al. (2018). Studies on anticonvulsant effects of novel histamine H3R antagonists in electrically and chemically induced seizures in rats. Int. J. Mol. Sci. 19, 3386. doi:10.3390/ijms19113386
Arrang, J. M., Garbarg, M., and Schwartz, J. C. (1983). Auto-inhibition of brain histamine release mediated by a novel class (H3) of histamine receptor. Nature 302, 832–837. doi:10.1038/302832a0
Arrang, J. M., Garbarg, M., and Schwartz, J. C. (1987). Autoinhibition of histamine synthesis mediated by presynaptic H3-receptors. Neuroscience 23, 149–157. doi:10.1016/0306-4522(87)90279-x
Asadi-Pooya, A. A., Brigo, F., Lattanzi, S., and Blumcke, I. (2023). Adult epilepsy. Lancet 402, 412–424. doi:10.1016/S0140-6736(23)01048-6
Bass, J. S., Tuo, A. H., Ton, L. T., Jankovic, M. J., Kapadia, P. K., Schirmer, C., et al. (2020). On the digital psychopharmacology of valproic acid in mice. Front. Neurosci. 14, 594612. doi:10.3389/fnins.2020.594612
Bastaki, S. M., Abdulrazzaq, Y. M., Shafiullah, M., Wiecek, M., Kiec-Kononowicz, K., and Sadek, B. (2018). Anticonvulsant and reproductive toxicological studies of the imidazole-based histamine H3R antagonist 2-18 in mice. Drug Des. Devel Ther. 12, 179–194. doi:10.2147/DDDT.S144730
Belelli, D., Lan, N. C., and Gee, K. W. (1990a). Anticonvulsant steroids and the GABA/benzodiazepine receptor-chloride ionophore complex. Neurosci. Biobehav Rev. 14, 315–322. doi:10.1016/s0149-7634(05)80041-7
Belelli, D., Mccauley, L., and Gee, K. W. (1990b). Heterotropic cooperativity between putative recognition sites for progesterone metabolites and the atypical benzodiazepine Ro 5-4864. J. Neurochem. 55, 83–87. doi:10.1111/j.1471-4159.1990.tb08824.x
Brigo, F., Ausserer, H., Tezzon, F., and Nardone, R. (2013). When one plus one makes three: the quest for rational antiepileptic polytherapy with supraadditive anticonvulsant efficacy. Epilepsy Behav. 27, 439–442. doi:10.1016/j.yebeh.2013.03.010
Brown, R. E., Stevens, D. R., and Haas, H. L. (2001). The physiology of brain histamine. Prog. Neurobiol. 63, 637–672. doi:10.1016/s0301-0082(00)00039-3
Canale, V., Kotanska, M., Dziubina, A., Stefaniak, M., Siwek, A., Starowicz, G., et al. (2021). Design, sustainable synthesis and biological evaluation of a novel dual α2a/5-HT7 receptor antagonist with antidepressant-like properties. Molecules 26, 3828. doi:10.3390/molecules26133828
Chateauvieux, S., Morceau, F., Dicato, M., and Diederich, M. (2010). Molecular and therapeutic potential and toxicity of valproic acid. J. Biomed. Biotechnol. 2010, 479364. doi:10.1155/2010/479364
Chen, Z., Li, W. D., Zhu, L. J., Shen, Y. J., and Wei, E. Q. (2002). Effects of histidine, a precursor of histamine, on pentylenetetrazole-induced seizures in rats. Acta Pharmacol. Sin. 23, 361–366.
Holmes, L. B., Harvey, E. A., Coull, B. A., Huntington, K. B., Khoshbin, S., Hayes, A. M., et al. (2001). The teratogenicity of anticonvulsant drugs. N. Engl. J. Med. 344, 1132–1138. doi:10.1056/NEJM200104123441504
Holmes, L. B., and Hernandez-Diaz, S. (2012). Newer anticonvulsants: lamotrigine, topiramate and gabapentin. Birth Defects Res. A Clin. Mol. Teratol. 94, 599–606. doi:10.1002/bdra.23028
Hua, Y., Song, M., Guo, Q., Luo, Y., Deng, X., and Huang, Y. (2023). Antiseizure properties of histamine H(3) receptor antagonists belonging 3,4-dihydroquinolin-2(1H)-Ones. Molecules 28, 3408. doi:10.3390/molecules28083408
Jang, D. H., Manini, A. F., Trueger, N. S., Duque, D., Nestor, N. B., Nelson, L. S., et al. (2010). Status epilepticus and wide-complex tachycardia secondary to diphenhydramine overdose. Clin. Toxicol. (Phila) 48, 945–948. doi:10.3109/15563650.2010.527850
Kamei, C., Okumura, Y., and Tasaka, K. (1993a). Influence of histamine depletion on learning and memory recollection in rats. Psychopharmacol. Berl. 111, 376–382. doi:10.1007/BF02244955
Kamei, C., Okumura, Y., Tsujimoto, S., and Tasaka, K. (1993b). Role of hypothalamic histamine in stimulating the corticosterone release in rats. Arch. Int. Pharmacodyn. Ther. 325, 35–50.
Kaminski, K., Rapacz, A., Luszczki, J. J., Latacz, G., Obniska, J., Kiec-Kononowicz, K., et al. (2015). Design, synthesis and biological evaluation of new hybrid anticonvulsants derived from N-benzyl-2-(2,5-dioxopyrrolidin-1-yl)propanamide and 2-(2,5-dioxopyrrolidin-1-yl)butanamide derivatives. Bioorg Med. Chem. 23, 2548–2561. doi:10.1016/j.bmc.2015.03.038
Kaplan, P. W. (2004). Reproductive health effects and teratogenicity of antiepileptic drugs. Neurology 63, S13–S23. doi:10.1212/wnl.63.10_suppl_4.s13
Kasteleijn-Nolst Trenite, D., Parain, D., Genton, P., Masnou, P., Schwartz, J. C., and Hirsch, E. (2013). Efficacy of the histamine 3 receptor (H3R) antagonist pitolisant (formerly known as tiprolisant; BF2.649) in epilepsy: dose-dependent effects in the human photosensitivity model. Epilepsy Behav. 28, 66–70. doi:10.1016/j.yebeh.2013.03.018
Lamb, Y. N. (2020). Pitolisant: a review in narcolepsy with or without cataplexy. CNS Drugs 34, 207–218. doi:10.1007/s40263-020-00703-x
Lan, N. C., Chen, J. S., Belelli, D., Pritchett, D. B., Seeburg, P. H., and Gee, K. W. (1990). A steroid recognition site is functionally coupled to an expressed GABA(A)-benzodiazepine receptor. Eur. J. Pharmacol. 188, 403–406. doi:10.1016/0922-4106(90)90201-8
Lazewska, D., Kaleta, M., Schwed, J. S., Karcz, T., Mogilski, S., Latacz, G., et al. (2017). Biphenyloxy-alkyl-piperidine and azepane derivatives as histamine H(3) receptor ligands. Bioorg Med. Chem. 25, 5341–5354. doi:10.1016/j.bmc.2017.07.058
Lazewska, D., Ligneau, X., Schwartz, J. C., Schunack, W., Stark, H., and Kiec-Kononowicz, K. (2006). Ether derivatives of 3-piperidinopropan-1-ol as non-imidazole histamine H3 receptor antagonists. Bioorg Med. Chem. 14, 3522–3529. doi:10.1016/j.bmc.2006.01.013
Lazewska, D., Olejarz-Maciej, A., Reiner, D., Kaleta, M., Latacz, G., Zygmunt, M., et al. (2020). Dual target ligands with 4-tert-Butylphenoxy scaffold as histamine H(3) receptor antagonists and monoamine oxidase B inhibitors. Int. J. Mol. Sci. 21, 3411. doi:10.3390/ijms21103411
Maguire, J. L., Stell, B. M., Rafizadeh, M., and Mody, I. (2005). Ovarian cycle-linked changes in GABA(A) receptors mediating tonic inhibition alter seizure susceptibility and anxiety. Nat. Neurosci. 8, 797–804. doi:10.1038/nn1469
Mauritz, M., Hirsch, L. J., Camfield, P., Chin, R., Nardone, R., Lattanzi, S., et al. (2022). Acute symptomatic seizures: an educational, evidence-based review. Epileptic Disord. 24, 26–49. doi:10.1684/epd.2021.1376
Mcleod, M. J. (1980). Differential staining of cartilage and bone in whole mouse fetuses by alcian blue and alizarin red S. Teratology 22, 299–301. doi:10.1002/tera.1420220306
Miyata, I., Saegusa, H., and Sakurai, M. (2011). Seizure-modifying potential of histamine H1 antagonists: a clinical observation. Pediatr. Int. 53, 706–708. doi:10.1111/j.1442-200X.2011.03328.x
Padmanabhan, R., Abdulrazzaq, Y. M., Bastaki, S. M., Shafiullah, M., and Chandranath, S. I. (2003). Experimental studies on reproductive toxicologic effects of lamotrigine in mice. Birth Defects Res. B Dev. Reprod. Toxicol. 68, 428–438. doi:10.1002/bdrb.10042
Padmanabhan, R., Shafiullah, M., Benedict, S., and Nagelkerke, N. (2006). Effect of maternal exposure to homocystine on sodium valproate-induced neural tube defects in the mouse embryos. Eur. J. Nutr. 45, 311–319. doi:10.1007/s00394-006-0600-4
Pennell, P. B., French, J. A., May, R. C., Gerard, E., Kalayjian, L., Penovich, P., et al. (2020). Changes in seizure frequency and antiepileptic therapy during pregnancy. N. Engl. J. Med. 383, 2547–2556. doi:10.1056/NEJMoa2008663
Sadek, B., Kuder, K., Subramanian, D., Shafiullah, M., Stark, H., Lazewska, D., et al. (2014a). Anticonvulsive effect of nonimidazole histamine H3 receptor antagonists. Behav. Pharmacol. 25, 245–252. doi:10.1097/FBP.0000000000000042
Sadek, B., Saad, A., Latacz, G., Kuder, K., Olejarz, A., Karcz, T., et al. (2016a). Non-imidazole-based histamine H3 receptor antagonists with anticonvulsant activity in different seizure models in male adult rats. Drug Des. Devel Ther. 10, 3879–3898. doi:10.2147/DDDT.S116192
Sadek, B., Saad, A., Schwed, J. S., Weizel, L., Walter, M., and Stark, H. (2016b). Anticonvulsant effects of isomeric nonimidazole histamine H3 receptor antagonists. Drug Des. Devel Ther. 10, 3633–3651. doi:10.2147/DDDT.S114147
Sadek, B., Saad, A., Subramanian, D., Shafiullah, M., Lazewska, D., and Kiec-Kononowiczc, K. (2015). Anticonvulsant and procognitive properties of the non-imidazole histamine H3 receptor antagonist DL77 in male adult rats. Neuropharmacology 106, 46–55. doi:10.1016/j.neuropharm.2015.10.023
Sadek, B., Schwed, J. S., Subramanian, D., Weizel, L., Walter, M., Adem, A., et al. (2014b). Non-imidazole histamine H3 receptor ligands incorporating antiepileptic moieties. Eur. J. Med. Chem. 77, 269–279. doi:10.1016/j.ejmech.2014.03.014
Sadek, B., Shehab, S., Wiecek, M., Subramanian, D., Shafiullah, M., Kiec-Kononowicz, K., et al. (2013). Anticonvulsant properties of histamine H3 receptor ligands belonging to N-substituted carbamates of imidazopropanol. Bioorg Med. Chem. Lett. 23, 4886–4891. doi:10.1016/j.bmcl.2013.06.075
Schwartz, J. C. (2011). The histamine H3 receptor: from discovery to clinical trials with pitolisant. Br. J. Pharmacol. 163, 713–721. doi:10.1111/j.1476-5381.2011.01286.x
Song, M., Yan, R., Zhang, Y., Guo, D., Zhou, N., and Deng, X. (2020). Design, synthesis, and anticonvulsant effects evaluation of nonimidazole histamine H(3) receptor antagonists/inverse agonists containing triazole moiety. J. Enzyme Inhib. Med. Chem. 35, 1310–1321. doi:10.1080/14756366.2020.1774573
Sterz, H., and Lehmann, H. (1985). A critical comparison of the freehand razor-blade dissection method according to Wilson with an in situ sectioning method for rat fetuses. Teratog. Carcinog. Mutagen 5, 347–354. doi:10.1002/tcm.1770050505
Szafarz, M., Kryczyk, A., Lazewska, D., Kiec-Kononowicz, K., and Wyska, E. (2015). Pharmacokinetics and tissue distribution of the new non-imidazole histamine H3 receptor antagonist 1-[3-(4-tert-butylphenoxy) propyl]piperidine in rats. Xenobiotica 45, 912–920. doi:10.3109/00498254.2015.1025117
Thomas, S. V., Syam, U., and Devi, J. S. (2012). Predictors of seizures during pregnancy in women with epilepsy. Epilepsia 53, e85–e88. doi:10.1111/j.1528-1167.2012.03439.x
Tomson, T., and Battino, D. (2007). Pharmacokinetics and therapeutic drug monitoring of newer antiepileptic drugs during pregnancy and the puerperium. Clin. Pharmacokinet. 46, 209–219. doi:10.2165/00003088-200746030-00002
Tomson, T., Battino, D., Bonizzoni, E., Craig, J., Lindhout, D., Perucca, E., et al. (2018). Comparative risk of major congenital malformations with eight different antiepileptic drugs: a prospective cohort study of the EURAP registry. Lancet Neurol. 17, 530–538. doi:10.1016/S1474-4422(18)30107-8
Veroniki, A. A., Rios, P., Cogo, E., Straus, S. E., Finkelstein, Y., Kealey, R., et al. (2017). Comparative safety of antiepileptic drugs for neurological development in children exposed during pregnancy and breast feeding: a systematic review and network meta-analysis. BMJ Open 7, e017248. doi:10.1136/bmjopen-2017-017248
Viale, L., Allotey, J., Cheong-See, F., Arroyo-Manzano, D., Mccorry, D., Bagary, M., et al. (2015). Epilepsy in pregnancy and reproductive outcomes: a systematic review and meta-analysis. Lancet 386, 1845–1852. doi:10.1016/S0140-6736(15)00045-8
Vohora, D., Saraogi, P., Yazdani, M. A., Bhowmik, M., Khanam, R., and Pillai, K. K. (2010). Recent advances in adjunctive therapy for epilepsy: focus on sodium channel blockers as third-generation antiepileptic drugs. Drugs Today (Barc) 46, 265–277. doi:10.1358/dot.2010.46.4.1445795
Witkin, J. M., and Nelson, D. L. (2004). Selective histamine H3 receptor antagonists for treatment of cognitive deficiencies and other disorders of the central nervous system. Pharmacol. Ther. 103, 1–20. doi:10.1016/j.pharmthera.2004.05.001
Xiao, F., Yan, R., Zhang, Y., Wang, S., Chen, S., Zhou, N., et al. (2021). Synthesis and antiseizure effect evaluation of nonimidazole histamine H(3) receptor antagonists containing the oxazole moiety. Arch. Pharm. Weinh. 354, e2000298. doi:10.1002/ardp.202000298
Yang, L., Wang, Y., and Chen, Z. (2022). Central histaminergic signalling, neural excitability and epilepsy. Br. J. Pharmacol. 179, 3–22. doi:10.1111/bph.15692
Keywords: histamine H3 receptors, antagonist DL76, maximal electroshock, seizures, anticonvulsant, malformation, gestation, mice
Citation: Bastaki SM, Abdulrazzaq YM, Zidan MA, Shafiullah M, Alaryani SG, Alnuaimi FA, Adeghate E, Mohsin S, Akour A, Siwek A, Łażewska D, Kieć-Kononowicz K and Sadek B (2024) Reproductive and fetal toxicity studies of histamine H3 receptor antagonist DL76 used in mice to prevent maximal electroshock-induced seizure. Front. Pharmacol. 15:1364353. doi: 10.3389/fphar.2024.1364353
Received: 02 January 2024; Accepted: 06 May 2024;
Published: 05 June 2024.
Edited by:
Antonella Riva, University of Genoa, ItalyReviewed by:
Nicola Forte, National Research Council (CNR), ItalyCopyright © 2024 Bastaki, Abdulrazzaq, Zidan, Shafiullah, Alaryani, Alnuaimi, Adeghate, Mohsin, Akour, Siwek, Łażewska, Kieć-Kononowicz and Sadek. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Bassem Sadek, YmFzc2VtLnNhZGVrQHVhZXUuYWMuYWU=; Salim M. Bastaki, c2Jhc3Rha2lAdWFldS5hYy5hZQ==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.