 Yueming Chu
Yueming Chu Qiang Yuan1,2
Qiang Yuan1,2 Lin Li
Lin Li
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Pharmacol. , 20 February 2024
Sec. Pharmacology of Anti-Cancer Drugs
Volume 15 - 2024 | https://doi.org/10.3389/fphar.2024.1303412
Cancer is a globally complex disease with a plethora of genetic, physiological, metabolic, and environmental variations. With the increasing resistance to current anticancer drugs, efforts have been made to develop effective cancer treatments. Currently, natural products are considered promising cancer therapeutic agents due to their potent anticancer activity and low intrinsic toxicity. Decursin, a coumarin analog mainly derived from the roots of the medicinal plant Angelica sinensis, has a wide range of biological activities, including anti-inflammatory, antioxidant, neuroprotective, and especially anticancer activities. Existing studies indicate that decursin affects cell proliferation, apoptosis, autophagy, angiogenesis, and metastasis. It also indirectly affects the immune microenvironment and can act as a potential anticancer agent. Decursin can exert synergistic antitumor effects when used in combination with a number of common clinical anticancer drugs, enhancing chemotherapy sensitivity and reversing drug resistance in cancer cells, suggesting that decursin is a good drug combination. Second, decursin is also a promising lead compound, and compounds modifying its structure and formulation form also have good anticancer effects. In addition, decursin is not only a key ingredient in several natural herbs and dietary supplements but is also available through a biosynthetic pathway, with anticancer properties and a high degree of safety in cells, animals, and humans. Thus, it is evident that decursin is a promising natural compound, and its great potential for cancer prevention and treatment needs to be studied and explored in greater depth to support its move from the laboratory to the clinic.
To date, a large number of nations around the globe list cancer as a main cause of death. The 2020 study reports that there are currently over 19.3 million new cases of cancer, in addition to approximately 10 million cancer deaths (Sung et al., 2021). According to recent research reports, more than 1.9 million new cancer cases are expected to occur in the United States in 2023, with approximately 609,820 people dying from cancer (Siegel et al., 2023). Cancer is a diverse and mechanistically complex disease with a plethora of underlying genetic and epigenetic factors (Han et al., 2021). It is characterized by the induction of unlimited cell proliferation and epigenetic alterations through the dysfunction of many important genes encoding key proteins (e.g., growth factors, transcription factors, anti-apoptotic proteins, and tumor suppressors) (Tarver, 2012; Millimouno et al., 2014). Currently, surgery, radiation, and chemotherapy are the main treatments for cancer. However, these therapies have serious toxic side effects on healthy human tissues and can also lead to lower success rates of standard treatment regimens, metastasis of cancer cells, excessive recurrence rates, and serious side effects (Roth et al., 2000; Liang et al., 2010; Are et al., 2018). Therefore, the search for new and effective antitumor agents is of importance. In recent years, the natural bioactive ingredients of plant origin, due to their regulation of selective molecular targets, have enormous capacity and potential for different types of cancer (Shehzad et al., 2018). Therefore, the application of traditional natural medicine monomer components for cancer treatment and prevention has received much attention.
Decursin (C19H20O5, from PubChem, IUPAC Name: [(3S)-2,2-dimethyl-8-oxo-3,4-dihydropyrano[3,2-g]chromen-3-yl] 3-methylbut-2-enoate), a pyranocoumarin compound extracted from the dried roots of Angelica gigas belonging to the Umbelliferae family, has been employed in traditional folk medicine as a tonic and a remedy for anemia (Choi et al., 2012). Angelica gigas Nakai (AGN) is also marketed internationally as a functional food for healthcare (Ahn et al., 2008). Decursin has been found to have a variety of therapeutic effects in increasing numbers of studies, such as anti-cancer, anti-inflammatory, anti-angiogenesis, obesity, antibacterial, and other pharmacological effects (Shehzad et al., 2018). In addition, the derivatives with bioactivity are gradually developing, suggesting that decursin can be developed as a promising lead compound. Notably, decursin has anti-cancer potential against a variety of cancers. In addition, studies have shown that chemoprevention of natural compounds through dietary regulation is a promising and cost-effective way to reduce cancer risk (George et al., 2017). Therefore, the bioactivity of plant extracts is promising and can be developed as complementary therapeutic agents or dietary supplements.
Due to its low toxicity and anticancer potential (Ahn et al., 1996; Kim and Kim, 1999; Mahat et al., 2012; Soon-ok et al., 2012; Reddy et al., 2017), decursin has attracted increasing attention (Figure 1). Among these are influencing the immune system, limiting tumor cell growth, migration, and invasion, triggering cell death, and enhancing tumor cell sensitivity to cancer therapy. The pharmacological activity of decursin was also enhanced by modification of the active sites and the application of innovative nano-formulation methods. The anticancer potential of decursin can also be enhanced by combining it with other anticancer drugs. There are studies that have shown that the safety of dietary supplements containing decursin has been verified (Zhang et al., 2015). In this study, we first describe the natural sources, biosynthetic pathways, and active derivatives of decursin. We summarize its anticancer effects and molecular mechanisms in various cancers and update the synergistic anticancer effects of decursin with clinical drugs. It is hoped that this review will provide strong theoretical support for decursin as an anticancer drug.
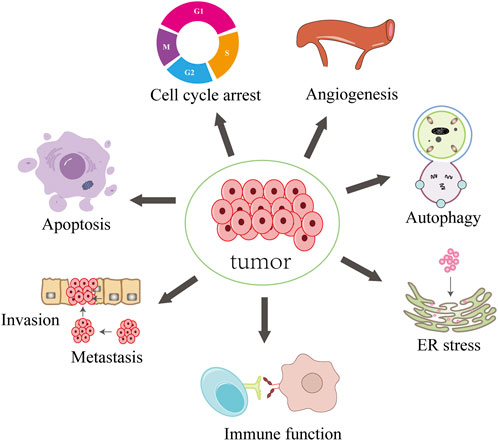
FIGURE 1. Synopsis of the mechanisms of decursin action against tumor cells.
Decursin is a novel pyranocoumarin compound, a monomeric component of traditional Chinese medicine, first isolated from the roots of the purple antebellum Angelica decursiva (Franch. et Sav.) and later from the Korean medicinal herb AGN (Hata and Sano, 1969; Ahn et al., 1996). The root of AGN is traditionally used as a folk medicine for anemia, colds, pain, and other ailments and is known to herbalists as “woman’s ginseng” (Zhang et al., 2012a). In addition, decursin was also extracted from the dried roots of the unflowered stems of Saposhnikovia divaricata (Zhao et al., 2010). Decursin is also extracted from the above-ground parts of Scutellaria lateriflora Lamiaceae (Li et al., 2009). Some other natural sources of decursin include Notopterygium incisum Ting ex H. T. Chang, Angelica dahurica, Angelica glauca Edgew, and Angelica czernaevia Kitaforma dentate Yook (Shi, 1997; Kim et al., 2006; Saeed and Sabir, 2008; Liang et al., 2018). Decursin has been extracted and isolated from a variety of herbs (Figure 2), including a study showing that in the dried roots of Korean angelica, the content has reached approximately 3% of the dried root (Ahn et al., 2008). Kweon et al. (2020) found that the ethanolic extract of AGN has good anticancer activity and confirmed decursin as the major component of AGN using an UPLC assay. Similar to AGN, the anticancer effects of Saposhnikovia divaricata, Scutellaria lateriflora Lamiaceae, Angelica glauca Edgew, Angelica dahurica, and Notopterygium incisum Ting ex H. T. Chang were also confirmed in several studies (Wu et al., 2010; Zheng et al., 2016; Matusiewicz et al., 2019; Park et al., 2021; Kumar et al., 2022). In addition, as shown in Table 1, we further summarized several natural sources of the active ingredient of decursin and found that decursin is not only present in AGN but also in the roots of most Angelica genera, such as Angelica decursiva, glauca, and dahurica. Second, coumarin analogs are widely found in a variety of natural plants and have good anticancer activity (Kupeli Akkol et al., 2020; Wang et al., 2021). As can be seen, decursin is a readily available and cost-effective pharmacologically active compound.
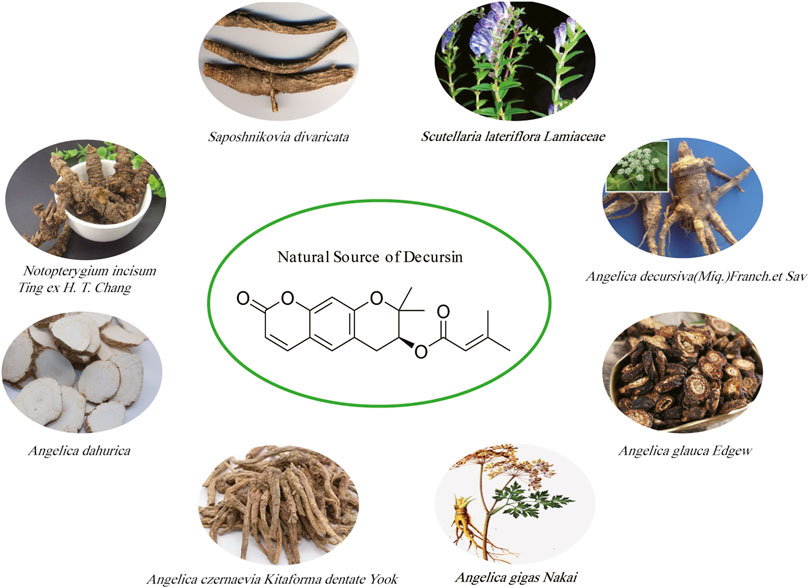
FIGURE 2. Chemical structure of decursin and its major natural sources.
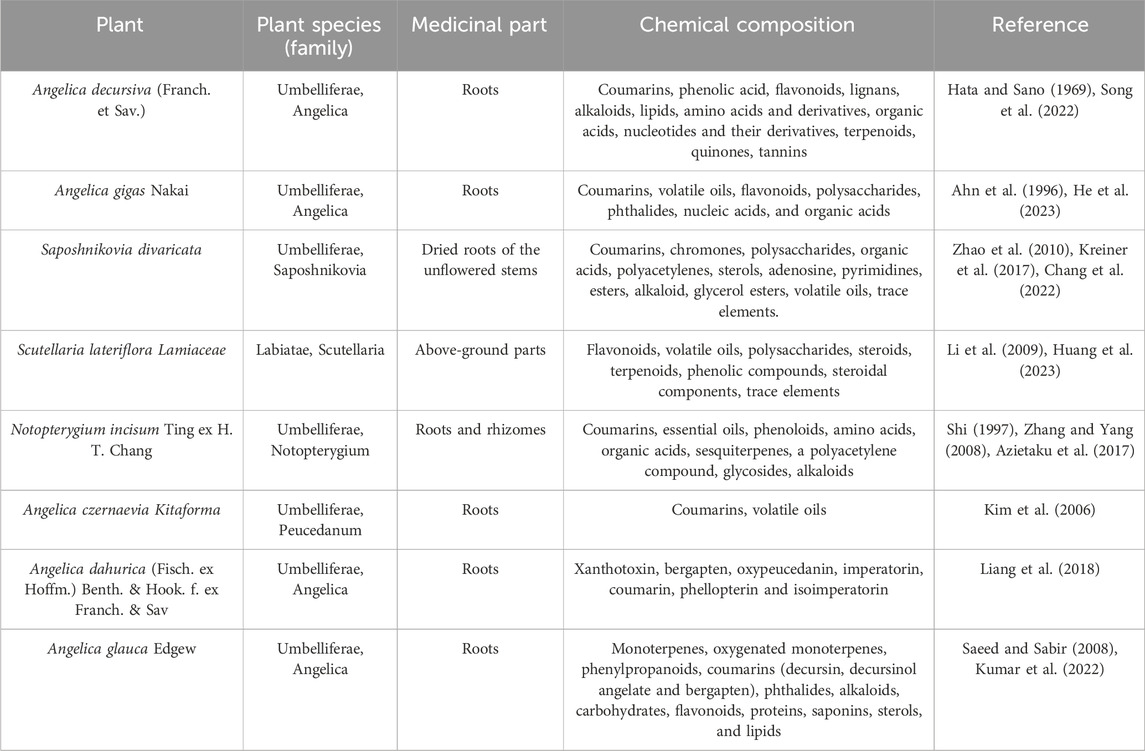
TABLE 1. Plant species with decursin constituents.
Despite the high biological activity of natural products, their levels in organisms are still not sufficient to meet the requirements (Roberts et al., 2010). In addition, biosynthesis has been playing an increasingly significant role in the construction of these natural product molecules, such as podophyllotoxin and saframycin A (Yu et al., 2017; Tanifuji et al., 2018). Among them, the phenylpropanoid pathway is not only a secondary metabolic pathway in plant metabolites but also plays an important role in the biosynthesis of decursin (Figure 3). First, L-phenylalanine is converted to trans-cinnamic acid catalyzed by phenylalanine ammonia-lyase (PAL), and second, it undergoes para-hydroxylation under the action of cinnamic acid 4-hydroxylase (C4H), followed by the formation of p-coumaroyl CoA in the presence of 4-coumarate CoA ligase (4CL). After p-coumaroyl CoA 2′-hydroxylase (C2′H) generates 2,4-dihydroxycinnamic acid, this compound, in turn, lactonizes to form umbelliferon (Vialart et al., 2012); subsequently, umbelliferon is converted to dimethylsuberosin by the action of umbelliferone 6-prenyl transferase (U6PT). Finally, in the presence of cytochrome P-450, dimethylsuberosin is converted to decursinol and further to decursin. However, the mechanism of this last step of the biosynthetic pathway remains to be fully understood (Ji et al., 2008). Ji et al. (2008) verified the phenylpropanoid biosynthesis in AGN root cultures containing decursin using deuterium-labeled isotope data. Since PAL and C4H are important enzymes in decursin biosynthesis, transgenic hairy roots overexpressing these genes were also established (Park et al., 2010). However, this experiment does not increase decursin expression, suggesting that PAL and C4H did not affect decursin biosynthesis, whereas the effect of several other enzymes on decursin remains unknown.
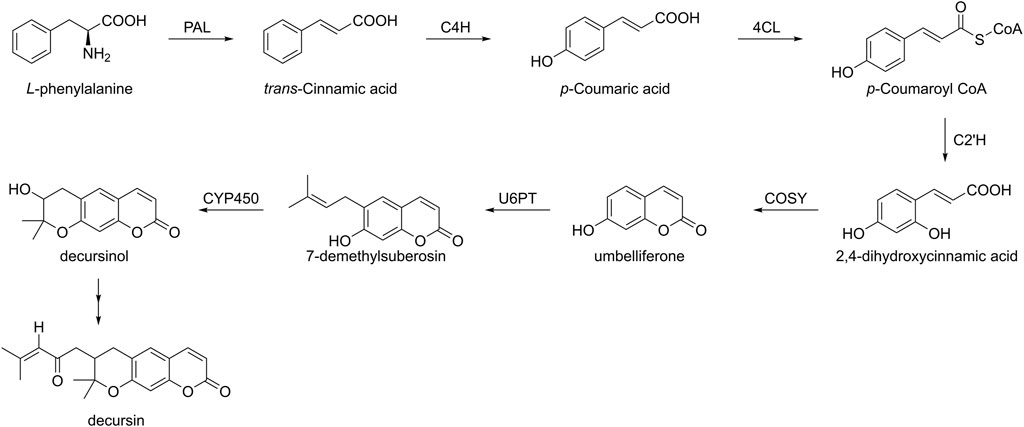
FIGURE 3. Biosynthesis pathway of decursin in plants.
Decursin’s anticancer activity has been clearly demonstrated in various forms of cancer, including prostate, sarcoma, breast, lung, colon, bladder, and blood cancers (Ahn et al., 2010; Kim et al., 2010d; Shehzad et al., 2018; Ge et al., 2020; Yang et al., 2023). Table 2 summarizes the anticancer effects of decursin on different cancer cells in vitro and the related molecular mechanisms of action. Decursin has pleiotropic effects in anticancer, including inhibition of tumor activity, cell proliferation, angiogenesis, metastasis, and invasion, induction of apoptosis and autophagy, and also affecting immune function. In this paper, we systematically summarized the anticancer effects of decursin and further analyzed the various molecular signaling pathways and molecular targets of decursin involved in tumorigenesis (Figure 4).
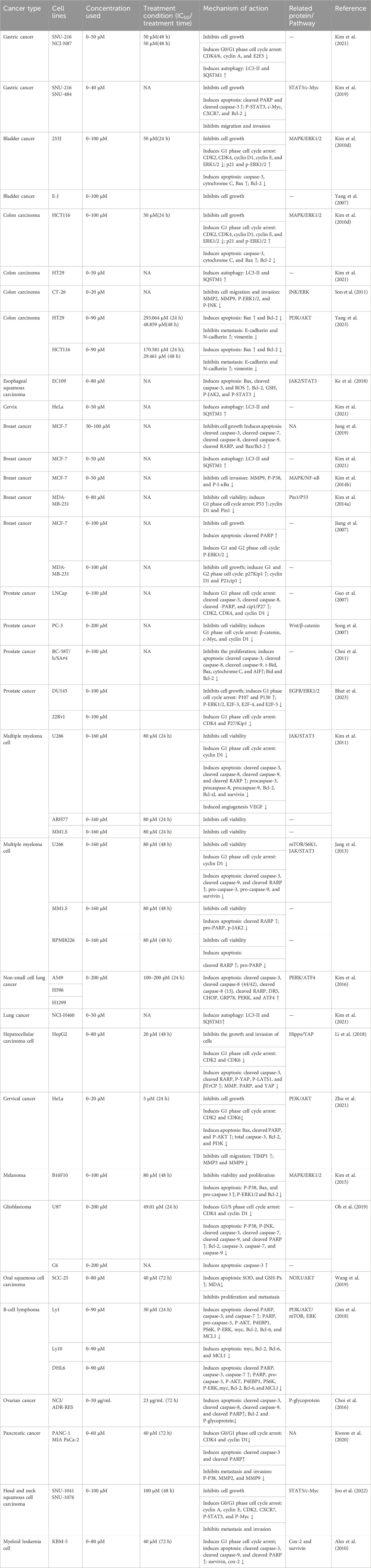
TABLE 2. Anticancer mechanisms of decursin in vitro
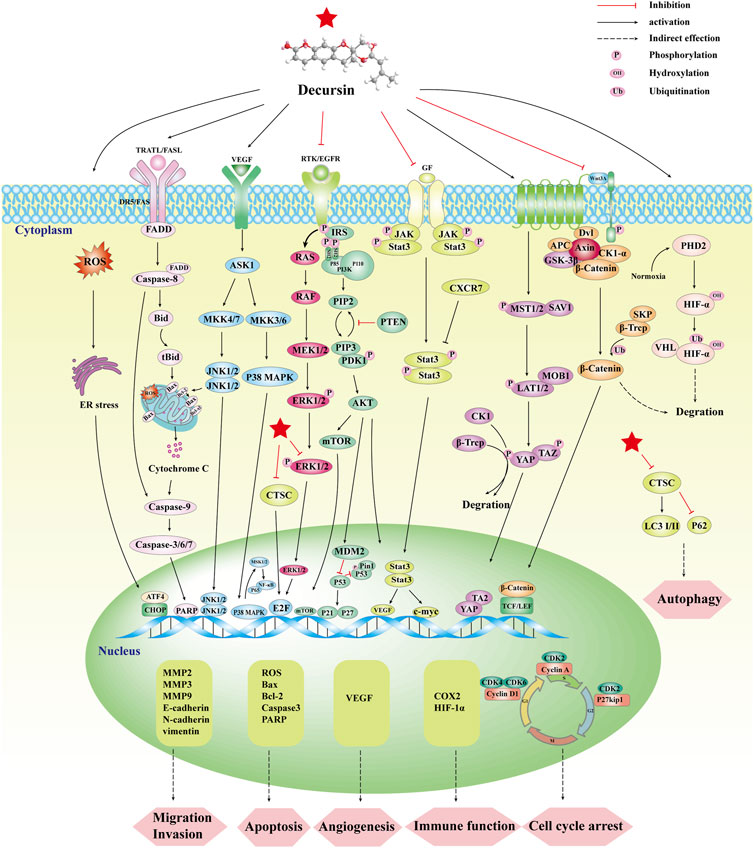
FIGURE 4. Synopsis of the mechanisms of decursin action against tumor cells.
Uncontrolled cell proliferation is a sign of cancer, and tumor cells often cause damage by directly regulating their cell cycle proteins and associated cyclin-dependent kinases (CDKs). Therefore, blocking abnormal regulation of the cell cycle is regarded as a major therapeutic method for cancer management (Sherr, 1996).
Decursin and the cell cycle arrest process are associated with the dysregulation of CDKs and the accumulation of cell cycle proteins (Kim et al., 2021; Ghafouri-Fard et al., 2022). Therefore, targeting cell cycle regulatory proteins to block cell growth provides a research direction for anticancer drug discovery. Decursin can inhibit the cell cycle at different checkpoints, blocking the G0/G1 and S phases of the cell cycle in the majority of cancers, with a small percentage of cancer cells being inhibited at the G2 phase (Jiang et al., 2007; Oh et al., 2019; Joo et al., 2022). Decursin promotes G1 phase block in 253J and HCT116 cancer cells by enhancing P21 and downregulating cyclin D1 expression through activation of the mitogen-activated protein kinase/extracellular signal-regulated kinase1/2 (MAPK/ERK1/2) signaling pathway (Kim et al., 2010d). Second, decursin induced G2 arrest at 50 μM but sparked G1 arrest at 10–20 μM in the estrogen-independent MDA-MB-231 cell line. Increased synthesis of p27Kip1 and growth-inhibitory endoplasmic reticulum (ER) protein numbers were linked to the impacts of G2 arrest (Jiang et al., 2007). However, the exact molecular mechanism by which decursin improves anticancer activity through cell cycle arrest remains unclear. Previous studies have reported that Pin1 has emerged as an essential and conserved cell cycle regulator that positively regulates cyclin D1 function in transcriptional and post-translational stabilization, causing cell cycle arrest (Liou et al., 2002). Thus, Kim et al. (2014a) are in agreement with previous studies that decursin induces G1 arrest in human breast cancer MDA-MB-231 cells mainly through Pin1/P53 signaling. Interestingly, inhibiting ERK1/2 phosphorylation prevented the G1 phase in the estrogen-dependent MCF-7 cell line (Jiang et al., 2007). Overall, decursin can target cell cycle pathways in cancer therapy.
Apoptosis is a common tumor suppression mechanism that plays an important role in cancer treatment (Morana et al., 2022). Apoptosis occurs in two typical pathways: an extrinsic pathway stimulated by death receptor activation and an intrinsic pathway activated by mitochondria-mediated endogenous stress (such as DNA damage, hypoxia, or other cellular stress), involving cysteine family proteins as well as p53 activation to initiate cell death. Fas ligand (FasL), tumor necrosis factor-alpha (TNF-α), and TNF-related apoptosis-inducing ligand (TRAIL), which function by attaching to target cell surface receptors, are the primary death ligands associated with the extrinsic route (Carneiro and El-Deiry, 2020).
Membrane proteins called death receptors, which are activated by taking in extracellular signals on the cell membrane surface, cause the external death pathway to be activated. Fas (whose ligand is FasL), the TNF receptors TNFR1 and TNFR, the TRAIL receptors death receptor 4 (DR4), and death receptor 5 (DR5) are examples of pro-apoptotic death receptors (Mohammad et al., 2015). Because of its unique capacity to kill cancer cells selectively, TRAIL is expected to be an effective anticancer therapy. Its DR5 is a transmembrane protein containing an intracellular death domain (DD), and many studies have indicated that the overexpression of DR5 leads to death receptor-induced apoptosis (Jia et al., 2012). Therefore, TRAIL resistance has been confirmed in a variety of cancers (Deng and Shah, 2020). In recent years, many natural bioactive compounds have had the potential to induce apoptosis by modulating various signaling molecules. It has been reported that decursin is able to act synergistically with TRAIL to induce apoptosis. Induction of DR5-mediated apoptosis, induction of reactive oxygen species (ROS), and selective activation of protein kinase RNA-like endoplasmic reticulum kinase (PERK)/activating transcription factor 4 (ATF4)/C/EBP homologous protein (CHOP) signaling in the ER stress pathway, which increases TRAIL sensitivity, are the primary mechanisms of action in non-small-cell lung cancer (NSCLC) cell lines (Kim et al., 2016). Consequently, decursin is an effective antitumor candidate capable of overcoming chemo-resistance in TRAIL-resistant NSCLC cells.
Members of the B-cell lymphoma-2 (Bcl-2) protein family play a major regulatory role in the endogenous pathway. Important apoptosis promoters, called BH3-only proteins, are upregulated in response to stress (such as growth factor deprivation, DNA damage, or ER stress), bind to anti-apoptotic Bcl-2 proteins with a high level of affinity, release Bcl-2-associated X protein (Bax)/Bcl-2 antagonist/killer (Bak) and form oligomers, which increase the permeability of the outer mitochondrial membrane and cause the release of apoptotic factors like cytochrome c and Smac/DIABLO from the mitochondria. This causes the cysteine cascade reaction to be activated, which causes the cleavage of hundreds of proteins and ultimately cell death (Carneiro and El-Deiry, 2020). Many natural bioactive compounds have the potential to induce internal cell death pathways by regulating various signal molecules. Studies have shown that decursin activates caspase-3 and caspase-9, members of the Caspase family proteases, by regulating circle oxymase-2 (COX-2) and survivin in leukemia KBM-5 cells and triggers the mechanism of poly ADP ribose polymerase (PARP) cutting, making human leukemia cells sensitive to cell death (Ahn et al., 2010). Further discovery that decursin induces cell death in melanoma B16F10 cells and breast cancer MCF-7 through the Bcl-2/Bax-mediated apoptosis pathway (Kim et al., 2015; Jung et al., 2019). In bladder cancer 253J and colon cancer HCT116, decursin decreases the potential of the mitochondrial membrane, makes the membranes more permeable, releases cytochrome c in the mitochondria, and activates caspase-3 to cause apoptosis (Kim et al., 2010d). However, investigations have revealed that the apoptosis-inducing factor (AIF) is released from the mitochondria and translocated to the nucleus when bladder cancer cells in RC-58T/h/SA#4 cells are in a stressful state. This causes chromosome condensation and DNA degradation, which can result in cell death (Choi et al., 2011). In multiple myeloma U266 cells, decursin regulates tumor cell viability by downregulating survivin, Bcl-2, Bcl-XL, and vascular endothelial growth factor (VEGF). It also inhibits Janus kinase 2 (JAK2) to prevent the activation of the signal transducer and activator of transcription 3 (STAT3) (Kim et al., 2011; Jang et al., 2013; Ke et al., 2018). In addition, in SNU-216, SNU-484, SNU-1041, and SNU-1076 cells of stomach cancer, decursin can also reduce c-myc expression by targeting STAT3 signals mediated by chemokine receptor 7 (CXCR7) to activate the caspase family to inhibit tumor growth and induce apoptosis (Kim et al., 2019; Joo et al., 2022). Decursin also affects the abnormal activation of the phosphoinositide 3-kinase (PI3K)/protein kinase B (Akt) signaling pathway to influence cancer growth, metabolism, and survival. It inhibits cancer cell growth and induces cysteine-dependent apoptosis in cervical cancer, B-cell lymphoma, colon cancer, oral squamous carcinoma, liver cancer, pancreatic cancer, and glioma (Kim et al., 2018; Li et al., 2018; Oh et al., 2019; Wang et al., 2019; Kweon et al., 2020; Zhu et al., 2021; Yang et al., 2023). Doxorubicin-resistant ovarian cancer cells NCI/ADR-RES induce apoptosis in the presence of decursin treatment. The main mechanism is that decursin induces apoptosis by blocking P-glycoprotein expression, activating caspase family activity, and increasing cleaved PARP levels in the presence of doxorubicin (Choi et al., 2016).
It has been established that apoptosis is a crucial intracellular process for preserving homeostasis and regulating the number of cells in the body. Decursin can promote apoptosis through multiple signaling pathways, signaling axes, or some target proteins in different cancers and is expected to be a novel anticancer agent and a potential new treatment option for multi-drug-resistant tumors.
Despite the nearly century-long development of anticancer drugs, the rate of 5-year survival for patients with metastatic cancer, especially distant metastases, is still dismal (Steeg, 2016). In addition, secondary tumors are clinically detectable in only mature stages, frequently after several metastases have occurred. It is well-recognized that the invasion and migration of tumor cells to nearby tissues or organs is a dynamic, multi-stage process known as tumor metastasis (Li et al., 2017).
A number of key molecules are involved in the adhesion, migration, and invasion of cancer cells. These include matrix metalloproteinases (MMPs), which are helpful for tumor invasion and migration because they breakdown the extracellular matrix. The extracellular matrix (ECM) is destroyed by cancer cells, allowing them to penetrate healthy tissues (Jablonska-Trypuc et al., 2016; Huang et al., 2021). By breaking down practically all ECM protein components, destroying the histological barrier to tumor cell invasion, and being a crucial factor in tumor invasion and metastasis, MMP plays a significant role in the development of cancer (Said et al., 2014). MMPs have consequently turned into desirable targets for oncology research and the creation of antitumor medications. In addition, epithelial cells are changed into mesenchymal cells by the complex biological process of epithelial–mesenchymal transition (EMT), which also gives them the ability to move and invade (Son and Moon, 2010; Ribatti et al., 2020). There is strong evidence that EMT can promote cell motility and invasiveness, thereby allowing cancer cells to detach from the primary mass and spread to secondary sites (Son and Moon, 2010; Ribatti et al., 2020). When EMT occurs, the expression of the epithelial marker E-cadherin is downregulated, whereas the mesenchymal marker N-cadherin is upregulated (Loh et al., 2019). A recent study demonstrated that decursin inhibited the proliferation and EMT of HT29 and HCT116 colon cancer cells by downregulating N-cadherin and vimentin protein expression and upregulating E-cadherin and PI3K/AKT signaling pathway expression, thereby inhibiting metastasis (Yang et al., 2023). Additionally, it was discovered that decursin inhibited lung metastasis and CT26 cell invasion in colon cancer cells, mostly via suppressing the expression of ERK/c-Jun N-terminal kinase (JNK) but not the p38 MAPK pathway (Son et al., 2011). The results showed that decursin strongly inhibited CT 26 metastasis formation and was as effective as celecoxib against invasion. In contrast, the p38-dependent pathway is associated with the expression and activation of MMPs in pancreatic cancer cells; thus, decursin inhibits migration and invasion of pancreatic cancer (PANC-1 and MIA PaCa-2) cell lines mainly through phosphorylation of p38 to control the expression of MMP-2 and MMP-9 (Kweon et al., 2020). In breast cancer MCF-7 cell lines, the anti-metastatic mechanism of decursin remained distinct. Decursin inhibits 12-O-tetradecanoylphorbol-13-acetate (TPA)-induced translocation of protein kinase C α (PKCα) from cytoplasmic lysate to the membrane rather than impacting PKCδ translocation, primarily through the MAPK/nuclear factor-κB (NF-κB) pathway, to affect MMP-9 production and cell invasion (Kim et al., 2014b). Among them, decursin increased cell proliferation, growth, migration, and invasion in gastric cancer by downregulating CXCR7 to affect the STAT3/c-Myc pathway (Kim et al., 2019). In addition, the specific mechanism of migration and invasion inhibition by decursin in head and neck squamous cell carcinoma (SNU-1041 and SNU-1076) and hepatocellular carcinoma (HepG2) is unclear and needs further insight to enable the natural product and its derivatives to play an important role in inhibiting the activity of MMP-2 and MMP-9 (Li et al., 2018; Joo et al., 2022).
Cancer cells require nutrients as well as oxygen to survive and multiply, and being close to blood vessels provides them access to the circulatory system. Additionally, the creation of blood vessels is required for tumor development. Therefore, inhibiting angiogenesis can inhibit tumor growth and spread, making cancer a manageable chronic condition (Son et al., 2009; Lugano et al., 2020). Pro-angiogenic factors and their associated receptors in abundance, primarily VEGF, fibroblast growth factor 2 (FGF-2), platelet-derived growth factor (PDGF), and angiopoietins, stimulate the formation of new blood vessels in tumors (Lugano et al., 2020). Among them, one of the most effective agents for promoting angiogenesis is VEGF (Ferrara et al., 2003). VEGF is produced and secreted by cancerous tumor cells and the stroma that surrounds them. It is linked to tumor progression, increased vascular density, invasiveness, metastasis, and tumor recurrence (Apte et al., 2019). Jung et al. (2009) revealed for the first time that decursin had excellent inhibitory effects on VEGF-induced vascular formation in human umbilical vein endothelial cells (HUVECs), fertilized eggs, and intramuscular animal models in vitro. Son et al. (2009) proceeded on to clarify that decursin had significant in vivo antiangiogenic effects, primarily via preventing the angiogenesis generated by vascular endothelial growth factor by reducing ERK and JNK activation in HUVECs. In vascular endothelial cell morphogenesis, MAPK signaling is a key molecular event in VEGF-induced proliferation, survival, and migration. Decursin inhibits VEGF-induced angiogenesis primarily by decreasing ERK and JNK activation. Consequently, decursin has the potential to be a novel angiogenesis inhibitor (Rousseau et al., 2000; Son et al., 2009). In a mouse model of oxygen-induced retinopathy, decursin also reduced blood retinal barrier breakdown, retinal angiogenesis, and migration of retinal endothelial cells via blocking the VEGFR-2 signaling pathway (Kim et al., 2009a). It was further shown that decursin may inhibit VEGFR-2 transcription or activation by directly regulating VEGFR-2 phosphorylation or by blocking VEGF binding to its receptor VEGFR-2, while the effect on endothelial progenitor cell (EPC) differentiation may be through the VEGF/VEGFR-2 and stromal cell-derived factor 1 (SDF-1)/CXCR4 signaling pathways, further demonstrating the good preventive potential of abscisin in the early stages of tumor formation in EPCs (Jung et al., 2012). The Lee et al. (2010) patent also demonstrates that decursin-containing compositions are effective in the treatment of VEGF-induced angiogenesis-related disorders. Thus, decursin primarily inhibits VEGFR2-mediated angiogenesis to prevent early tumor development.
The human immune system, with its own adaptive and innate immunity, is able to recognize and kill abnormally proliferating cells, thereby eliminating tumor cells or controlling tumor growth. Among other things, immune cells are able to infiltrate the tumor microenvironment (TME) to regulate tumor progression (Palazon et al., 2012). T cells, B cells, and natural killer (NK) cells are adaptive immune cells that can be activated to produce an immune response by exposure to specific antigens. Macrophages and neutrophils, which carry out the innate immune response, are a non-specific defense mechanism that will act within hours of the entry of a foreign antigen into the body (Hinshaw and Shevde, 2019; Anderson and Simon, 2020). Cancer cells can shape the microenvironment to help support tumorigenesis and evade the immune system through suppression (Arner and Rathmell, 2023). It has been suggested that this phenomenon develops as a result of crosstalk between cancer cells and proximal immune cells (Hinshaw and Shevde, 2019). Thus, immune cells are an important part of the matrix of the tumor microenvironment. Reports are suggesting that hypoxia-stabilized HIF-1α can mediate the tumor immune response by inducing the expression of the immune checkpoint programmed cell death ligand 1 (PD-L1), which leads to immunosuppression and evasion, ultimately leading to tumor growth (Noman et al., 2014; Palazon et al., 2014). In recent years, Ge et al. (2020) demonstrated for the first time that decursin reduces HIF-1 protein accumulation by promoting ubiquitination and proteasomal degradation. They also demonstrated that in the xenograft mouse tumor model, decursin increased infiltrating lymphocytes (CD3+), helper T cells (CD4+), and cytotoxic T cell (CD8+) accumulations while decreasing tumor expression of regulatory T cells (Foxp3) and myeloid-derived suppressor cell-mediated immunosuppressive factor (Arg1) in tissues. Therefore, decursin is a promising novel HIF-1α inhibitor that can directly inhibit the expression of PD-L1, which can help improve T-cell activation in the tumor microenvironment and inhibit the growth of tumor tissues. To confirm whether decursin may be employed as an immunotherapeutic anticancer agent, however, plenty of research is still required.
Autophagy, which is a mechanism for delivering cellular material to lysosomes for degradation, is an evolutionarily conserved catabolic process that leads to a basal turnover of cellular components and provides energy and macromolecular precursors (Levy et al., 2017). It has been demonstrated that autophagy is crucial for cancer cell survival and adaptation to changes in the tumor microenvironment under diverse stress circumstances and that autophagy is crucial for maintaining cellular homeostasis as a protein/organelle quality control process (Nazio et al., 2019). Autophagy dysregulation is frequently linked to the development of cancer. The creation of membrane-bound vacuoles, or autophagosomes, in the cytoplasm as a result of nutritional deprivation, which contain cytoplasmic organelles, such as organelles and inclusions, is one characteristic of autophagy. Autophagy-related protein (ATG), which includes Beclin1 and microtubule-associated protein light chain 3 (LC3), is the primary signaling molecule in autophagy (Chifenti et al., 2013). Currently, the most widely studied ATG8 protein in mammalian cells is LC3 (Mareninova et al., 2020). The results of existing studies show that LC3-II levels are an important indicator of cellular autophagic activity (Plaza-Zabala et al., 2020). After treatment with decursin, an increase in LC3II/LC3I and Sequestosome 1 (SQSTM1) levels was observed in a time-dependent manner, consistent with the inhibitory effect of treatment with bafilomycin A, a well-known autophagy inhibitor, suggesting that autophagic fluxes are affected by decursin (Kim et al., 2021). Second, decursin-mediated inhibition of autophagic flux was observed in gastric, colon, cervical, breast, and lung cancer cells (Kim et al., 2021). The role of autophagy in tumors is complex, and the therapeutic potential of interventions that activate or inhibit autophagy is enormous (de Souza et al., 2020; Debnath et al., 2023). Therefore, there is a need to further investigate autophagy and lysosomal proteases in decursin-mediated tumorigenesis and development and fully exploit the therapeutic potential of autophagy modulators.
Combination therapy is a useful method for overcoming drug resistance in all varieties of malignant diseases affecting humans. Currently, molecular targeted therapy, chemotherapy, radiotherapy, and surgical resection are the primary cancer treatments (Debela et al., 2021; Saini and Twelves, 2021). However, some tumors develop drug resistance quickly, and tumor cell resistance is a significant factor in treatment failure and a poor prognosis for oncology chemotherapeutic treatments. It has been observed that using two or more medications in combination therapy can increase effectiveness and lower the occurrence of drug resistance (Vasan et al., 2019; Casak et al., 2021). Additionally, there is mounting evidence that natural compound combination therapy may be more beneficial than monotherapy, which aims to enhance effectiveness while limiting the possibility of adverse medication reactions (Zacchino et al., 2017). Decursin is a novel inhibitor of STAT3 activation, and when combined with bortezomib, it increases cytotoxicity and induces cell death in human multiple myeloma cells (Kim et al., 2011). Decursin, in combination with doxorubicin, enhances mitochondrial apoptosis in multiple myeloma cells by decreasing mitochondrial membrane potential through the mechanistic target of the rapamycin (mTOR)/STAT3 signaling pathway, decreasing cyclin D1 and survivin expression, and thereby inhibiting the negative regulation of the upstream complex phosphatase (PTP) of the STAT pathway (Jang et al., 2013). The anticancer activity of doxorubicin has reportedly been shown to inhibit DNA polymerase and topoisomerase II (Zunina et al., 1975; Tewey et al., 1984). Therefore, it may be inferred that the suppression of DNA polymerase or topoisomerase II activity may contribute to the synergistic anticancer action of decursin and doxorubicin. However, more experiments are still needed in the near future to test this hypothesis (Jang et al., 2013). Interestingly, decreased P-glycoprotein expression by decursin also increased the chemosensitivity of doxorubicin. By reducing P-glycoprotein expression, decursin and doxorubicin together cause apoptotic cell death in doxorubicin-resistant ovarian cancer cells. Although NF-κB can regulate the gene expression of P-glycoprotein, the specific mechanism by which it affects decursin synergistically with doxorubicin needs to be further explored (Choi et al., 2016). Decursin substantially recovered Cu/Zn superoxide dismutase (SOD), catalase, and glutathione peroxidase activity in cisplatin-treated human renal epithelial cells, according to Kim et al. (2010b), whereas the known antioxidant N-acetyl-L-cysteine (NAC) did not restore levels of antioxidant enzymes inhibited by cisplatin treatment. Combination treatment with decursin and cisplatin significantly protected human primary renal epithelial cells (HRCs) from cisplatin-induced cytotoxicity and apoptosis. Therefore, for multi-drug-resistant (MDR) tumor, decursin may be a viable new treatment option. However, although the combination of decursin with chemotherapeutic agents can increase efficacy and has shown some safety in in vivo trials in animals, its results in vivo need to be further studied in large samples.
The in vivo antitumor activity of decursin was significant in different types of tumors (Table 3). Among them, BALB/c nude mice, C57BL/6J mice, pathogen-free male ICR mice, and Eμ-myc transgenic mice are commonly used models for in vivo anticancer studies of decursin . Tumor models are established by subcutaneous, intraperitoneal, or oral implantation of mouse cancer cells or xenografts to form tumors (Lee et al., 2003; Jung et al., 2012; Kim et al., 2018; Kim et al., 2021). Decursin was reported to significantly suppress tumor size in gastric cancer xenograft mice by disrupting autophagy, inhibiting cathepsin C (CTSC), and downregulating CXCR7 (Kim et al., 2019; Kim et al., 2021). Decursin suppresses hepatocellular carcinoma growth in nude mice via the Hippo/YAP signaling system (Li et al., 2018). Similarly, decursin inhibited the production of cervical cancer cells in mice in vivo (Zhu et al., 2021). When compared to the effects of phosphate-buffered saline (PBS) therapy, decursin significantly decreased tumor weight and growth. Tumor tissue homogenate analysis of the B16F10 tumor revealed that the decursin-treated group was able to promote tumor apoptosis in vivo by reducing procaspase-3 protein expression in vivo (Kim et al., 2015). Furthermore, decursin suppressed the proliferation and invasion of CT-26 colon cancer cells along with lung metastases in mice by decreasing MMP-9 production via the ERK/JNK signaling pathway (Son et al., 2011). Decursin administration totally restored the normal appearance of the spleen in Eμ-myc transgenic mice, which was closely associated with a massive decrease in Myc expression, showing that decursin could be effective in the treatment of Myc-driven B-cell lymphoma (Kim et al., 2018). Decursin was also found to inhibit sarcoma size in vivo in pathogen-free male ICR mice, tumor growth in prostate cancer in C57BL/6 TRAMP mice, and angiogenesis in C57BL/6J mice in liver cancer (Lee et al., 2003; Jung et al., 2012; Tang et al., 2015). Finally, in the esophageal squamous carcinoma, decursin increased the expression of the pro-apoptotic protein Bax and the cleavage activation of caspase-3 through the JAK2/STAT3 pathway and inhibited the expression of anti-apoptotic proteins, thus promoting apoptosis in tumor cells (Ke et al., 2018). In vivo anticancer studies of decursin show the potential of decursin to treat cancer. Unfortunately, only in vivo rodent studies are insufficient to demonstrate that decursin may be used clinically as an anticancer drug. As a result, the anticancer potential of decursin has to be investigated further in vivo.
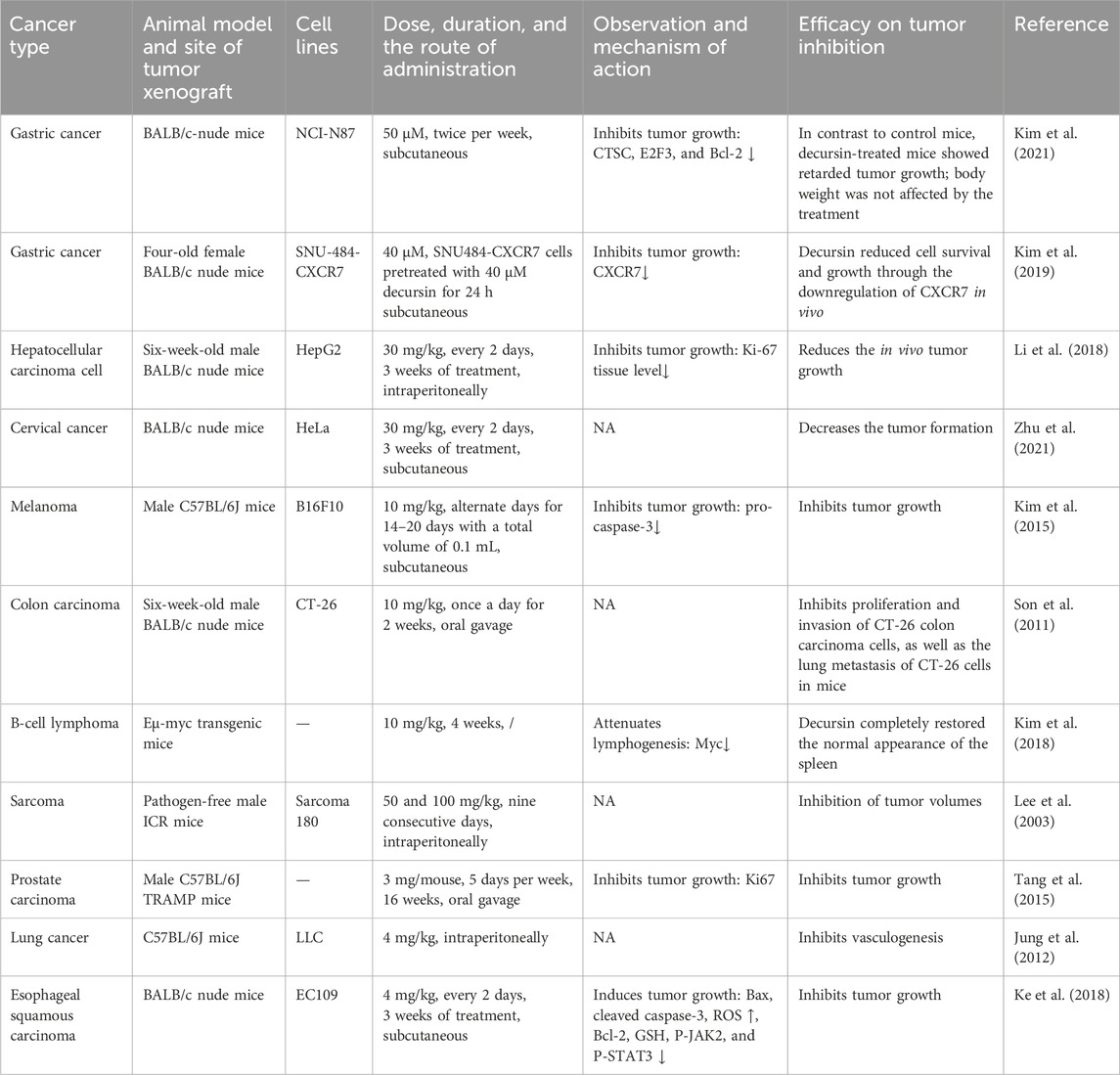
TABLE 3. In vivo anticancer properties of decursin.
Currently, there are many natural compounds that have very low potential for direct use as phytotherapeutic agents due to disadvantages such as poor water solubility and high toxicity (Atanasov et al., 2021). The most common strategy is to create analogs by modifying the structure (Maier, 2015). Although the antiproliferative effects of decursin are well known, they are not sufficient to be approved for clinical use.
Decursin is a biologically active natural compound that, due to its hydrophobic nature, is extracted using methods such as ethanol or supercritical carbon dioxide fluids (Zhang et al., 2012a). In order to further explain the effect of different structures on the antitumor activity of decursin, the corresponding structure–activity relationships were systematically summarized, and the structures of decursin and its analogs are shown in Figure 5. It was found that the C-7 position side chain of decursin was replaced by 1-phenyl acrylate to generate decursinol angelate, which showed reduced spatial site resistance and significantly increased anticancer activity. Decursinol angelate was able to significantly inhibit the invasion of the fibrosarcoma cell line HT1080 and the breast cancer cell line MDA-MB-231, as well as inhibit the increase of B16F10 melanoma cells (Kim et al., 2010c; Chang et al., 2021). Second, decursinol is not only the active derivative of decursin but also its metabolite (Yim et al., 2005; Lee et al., 2009). The side chain of decursin (CH3)2-C = CH-COO is replaced by -OH, but decursinol, which has the same coumarin ring system, is not as anticancer as decursin, suggesting that this isoprenoid structure may be important for the anticancer effect (Kim et al., 2005; Yim et al., 2005; Song et al., 2007). Interestingly, the Son team further showed that oral administration of decursinol at 10 mg/kg resulted in a significantly better reduction of lung tumor nodules in mice than decursin at the same dose (Son et al., 2011). It can be seen that although decursinol lacking a side chain is not as effective as decursin against cancer, it shows superior anti-metastatic ability in vivo than decursin. Zhang et al. (2012b) synthesized a decursin derivative, decursinol phenylthiocarbamate (DPTC), whose phenylthiocarbamate group modification of the side chain not only retained the anti-AR activity in vivo and in vitro but also exhibited greater hydrolysis resistance than decursin. Consequently, DPTC would not be converted into DOH in vivo. At the same time, its anticancer efficacy was somewhat diminished due to its low solubility and low bioavailability. Therefore, the substituent group at the C-7 position of decursin is a determinant of AR antagonist activity. In addition, the introduction of (CH3) 2-C = CH-COO at the C-7 and C-8 positions of decursin produces a more active decursin derivative, CSL-32, which inhibits the expression of pro-inflammatory mediators that invade or migrate through the extracellular matrix and human fibrosarcoma cells in the presence of TNF (Lee et al., 2012). This suggests that the increased lipid solubility of decursin at the C-8 position facilitates more favorable binding to amino acid residues of proteins, resulting in a more stable binding and a greater impact on anticancer effects. Lee et al. (2019) also introduced 3-(3-pyridyl) acrylic acid at the C-7 position and synthesized the decursin derivative JH-4, which can effectively block the progerin–lamin A/C combination and treat a variety of severe vascular inflammatory diseases. Therefore, its anticancer potential warrants further investigation. There are recently disclosed patents indicating that dihydropyranocoumarin D2 is a pharmaceutical composition with anticancer potential capable of combining with Bruton’s tyrosine kinase inhibitor (Jeon et al., 2022). In addition, it has also been shown that the derivative dihydropyranocoumarin D2 is a promising lead compound that can effectively regulate tyrosinase and thus inhibit melanogenesis due to its strong potential (Kim et al., 2010a). Furthermore, it has been shown that decursin and its derivative, decursinol angelate, inhibit melanin formation in B16 murine melanoma cells (Zhang et al., 2012b). Therefore, there is a need to further investigate whether dihydropyranocoumarin D2 has considerable potential to be effective against melanoma.
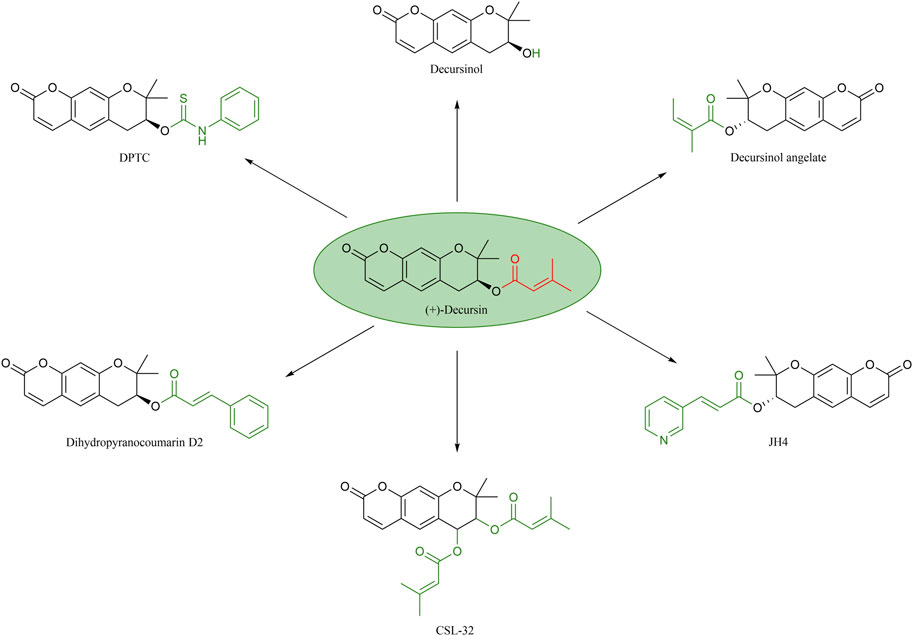
FIGURE 5. Structure–activity relationship of decursin.
In conclusion, decursin is an important prototype lead compound, and the C-7 position of decursin is an important active site that can be altered in terms of lipid solubility and bioavailability through the introduction of various groups, with a view to designing more decursin derivatives with higher stability and broader application prospects.
Decursin has limited solubility and bioavailability, which creates difficulties for drug delivery systems and targeted drug release. In recent years, nanoparticles (NPs) have been repeatedly reported to play an important role in modern medicine. The rapid growth of nanotechnology toward the development of nanomedicines holds great promise for the improvement of cancer therapeutics (Aghebati-Maleki et al., 2020). Due to the poor water solubility of decursin, in order to further improve it, Lee et al. (2017) prepared the first nanocomposites (NCs) of AGN ethanol extract (decursin as the main component) by using a modified nanocrystal preparation method and further found that the group of AGN NCs was capable of promoting apoptosis generation. The anticancer activity in breast cancer was significantly higher than that of the AGN EtOH extract group and had no effect on normal cells, suggesting that AGN nanoformulations may have tumor-selective killing efficacy. This was further validated, and polydopamine (PD)-coated NCs were investigated. It was found that both PD-AGN NCs and AGN NCs had better activity than the AGN EtOH extract. The PD layer of PD-AGN NCs appeared to have better cell adhesion in MDA-MB-231 cells. In addition, they contribute to apoptotic and antiproliferative efficacy (Nam et al., 2018b). Second, Nam et al. (2018a) further showed that AGN NPs act through endocytosis into the cytoplasm rather than the nucleus. This shows that nanoformulations improve the aqueous solubility, bioavailability, and targeting of desmoglein for better application in tumor therapy.
Decursin is crucial for the treatment of a variety of illnesses, including cancer. In recent years, it has been shown that in vitro experiments based on two-dimensional cultures do not accurately reflect the environment of the patient (Edwards et al., 2015). As a result, it is critical to assess its toxicity and safety levels before using it in cancer treatment. In recent years, it has been discovered that not all PKC activators are tumor promoters (Ruan and Zhu, 2012). Interestingly, Ahn et al. (1996) demonstrated in vitro for the first time that decursin was more effective in blocking rapidly growing cancer cells than slowly growing normal fibroblasts and that the cytotoxic activity of decursin may be related to the mechanism of PKC activation, but further exploration is still needed in vitro. Kim et al. (2021) established a spheroid 3D culture, a patient-derived organoid model, and a rodent xenograft tumor model to show that 50 μM of dexmethylphenidate is toxic to cancer cells but has a good safety profile and has no effect on body weight in mice. Similarly, decursin showed no toxic effects on normal human cell lines (Ahn et al., 2010; Kim et al., 2011; Kweon et al., 2020). By modulating COX-2 and survivin, decursin induces an apoptotic process that is safe for human peripheral blood lymphocytes but detrimental to leukemia cells (Ahn et al., 2010). Decursin, an agent used to treat variable retinopathy caused by a breakdown of the blood–retinal barrier (BRB), did not produce retinal toxicity at concentrations up to 50 mol/L, which is five times the therapeutically effective value. This implies that decursin can be used to treat this problem without putting the retina or healthy retinal vessels at risk (Kim et al., 2009b). In addition to investigating the safety of decursin in cell lines and rodents, a related clinical trial has also investigated the pharmacokinetics of a single oral dose of decursin and the decursinol angelate-enriched dietary supplement Cogni-Q. In humans, a total of 20 healthy subjects, each taking 119 mg of decursin and 77 mg of decursinol angelate, were enrolled in the pharmacokinetic study. The results of the study provide credibility to the safety data using rodent models (Zhang et al., 2015). The above studies facilitate the clinical translation of decursin against cancer and other diseases and explore relevant active molecular targets. However, further studies are still needed to assess the genotoxicity and reproductive toxicity of decursin, among others, in order to support anticancer studies of decursin.
In recent years, natural medicine therapies have shown great potential in the treatment of cancer. Decursin is a natural compound that, through abnormal signaling and epigenetics, inhibits the growth, invasion, migration, apoptosis, autophagy, and immunity of cancer cells. Although previous studies have shown that decursin has good anticancer effects and a certain degree of safety, as the current studies on its anticancer effects are still in the preclinical stage, only one of its safety studies has been marketed abroad as a dietary supplement. This shows that, due to the lack of research in the clinical stage, decursin needs to be explored comprehensively before it can be established as an anticancer agent. As a result, we propose some ideas that we hope will serve as a direction for further research. First, due to the limited solubility and bioavailability of ecdysteroids, there is a need to structurally modify their active side chain at the C-7 position and to investigate nanoformulations and other types of novel targeted delivery systems to enhance their anticancer potential. Second, decursin is able to inhibit tumor growth by targeting CXCR7, but limited research has been conducted, and the mechanism is unknown. Therefore, the potential of decursin as a CXCR7-targeting agent warrants further research. Third, chemotherapy resistance is still the key factor contributing to poor survival and prognosis in tumor patients. Whether decursin can be used as a combination of chemotherapeutic drugs to reduce chemotherapy drug resistance still needs further in-depth validation. Fourth, so as to maximize the anticancer potential of decursin, additional studies are required on the main mechanisms underlying particular forms of death (autophagy, ferroptosis, pyroptosis, and cuproptosis), which have not been conducted. Fifth, the safety of decursin has been shown in rodent in vivo studies in rodents but remains insufficient in the clinical trial part. Finally, decursin is being used as the primary active element in the nutraceutical Cogni-Q. As a result, more research is needed to discover whether decursin may be utilized as a medical herbal monomer and whether it can be employed in the prevention and treatment of cancer.
YC: data curation, formal analysis, and writing–original draft. QY: data curation, formal analysis, and writing–original draft. HJ: visualization and writing–original draft. LW: data curation and writing–original draft. YX: methodology and writing–original draft. XZ: conceptualization, funding acquisition, supervision, and writing–review and editing. LL: conceptualization, funding acquisition, supervision, and writing–review and editing.
The authors declare that financial support was received for the research, authorship, and/or publication of this article. The present study was supported by the City-school Cooperation Project of Nanchong (No. 22SXQT0071), the City-school Cooperation Project of Nanchong (No. 20SXQT0047), the Science and Technology Plan Project of Nanchong (No. 23JCYJPT0026), the Natural Science Foundation of Sichuan Province (20237F30473), the Research and Development Fund of North Sichuan Medical College (No. CBY21-QA05), and the Medical Research Project of Sichuan Medical Association (No. S23088).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors, and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Aghebati-Maleki, A., Dolati, S., Ahmadi, M., Baghbanzhadeh, A., Asadi, M., Fotouhi, A., et al. (2020). Nanoparticles and cancer therapy: perspectives for application of nanoparticles in the treatment of cancers. J. Cell. Physiol. 235 (3), 1962–1972. doi:10.1002/jcp.29126
Ahn, K. S., Sim, W. S., and Kim, I. H. (1996). Decursin: a cytotoxic agent and protein kinase C activator from the root of Angelica gigas. Planta Med. 62 (1), 7–9. doi:10.1055/s-2006-957785
Ahn, M. J., Lee, M. K., Kim, Y. C., and Sung, S. H. (2008). The simultaneous determination of coumarins in Angelica gigas root by high performance liquid chromatography-diode array detector coupled with electrospray ionization/mass spectrometry. J. Pharm. Biomed. Anal. 46 (2), 258–266. doi:10.1016/j.jpba.2007.09.020
Ahn, Q., Jeong, S. J., Lee, H. J., Kwon, H. Y., Han, I., Kim, H. S., et al. (2010). Inhibition of cyclooxygenase-2-dependent survivin mediates decursin-induced apoptosis in human KBM-5 myeloid leukemia cells. Cancer Lett. 298 (2), 212–221. doi:10.1016/j.canlet.2010.07.007
Anderson, N. M., and Simon, M. C. (2020). The tumor microenvironment. Curr. Biol. 30 (16), R921–R925. doi:10.1016/j.cub.2020.06.081
Apte, R. S., Chen, D. S., and Ferrara, N. (2019). VEGF in signaling and disease: beyond discovery and development. Cell. 176 (6), 1248–1264. doi:10.1016/j.cell.2019.01.021
Are, C., Caniglia, A., Mohammed, M., Smith, L., Cummings, C., Lecoq, C., et al. (2018). Global variations in the level of cancer-related research activity and correlation to cancer-specific mortality: proposal for a global curriculum. Ann. Surg. Oncol. 25 (3), 594–603. doi:10.1245/s10434-017-6276-6
Arner, E. N., and Rathmell, J. C. (2023). Metabolic programming and immune suppression in the tumor microenvironment. Cancer Cell. 41 (3), 421–433. doi:10.1016/j.ccell.2023.01.009
Atanasov, A. G., Zotchev, S. B., Dirsch, V. M., International Natural Product Sciences, T., and Supuran, C. T. (2021). Natural products in drug discovery: advances and opportunities. Nat. Rev. Drug Discov. 20 (3), 200–216. doi:10.1038/s41573-020-00114-z
Azietaku, J. T., Ma, H., Yu, X. A., Li, J., Oppong, M. B., Cao, J., et al. (2017). A review of the ethnopharmacology, phytochemistry and pharmacology of Notopterygium incisum. J. Ethnopharmacol. 202, 241–255. doi:10.1016/j.jep.2017.03.022
Bhat, T. A., Dheeraj, A., Nambiar, D. K., Singh, S. P., Yim, D. S., and Singh, R. P. (2023). Decursin inhibits EGFR-ERK1/2 signaling axis in advanced human prostate carcinoma cells. Prostate 83 (6), 534–546. doi:10.1002/pros.24482
Carneiro, B. A., and El-Deiry, W. S. (2020). Targeting apoptosis in cancer therapy. Nat. Rev. Clin. Oncol. 17 (7), 395–417. doi:10.1038/s41571-020-0341-y
Casak, S. J., Donoghue, M., Fashoyin-Aje, L., Jiang, X., Rodriguez, L., Shen, Y. L., et al. (2021). FDA approval summary: atezolizumab plus bevacizumab for the treatment of patients with advanced unresectable or metastatic hepatocellular carcinoma. Clin. Cancer Res. 27 (7), 1836–1841. doi:10.1158/1078-0432.CCR-20-3407
Chang, L., Jing, W., and Cheng, X. (2022). Progress of pharmacological effects of chemical constituents of Fangfeng and analysis of quality markers. Mod. Chin. Med. 24 (10), 2026–2039. doi:10.13313/j.issn.1673-4890.20211115003
Chang, S. N., Khan, I., Kim, C. G., Park, S. M., Choi, D. K., Lee, H., et al. (2021). Decursinol angelate arrest melanoma cell proliferation by initiating cell death and tumor shrinkage via induction of apoptosis. Int. J. Mol. Sci. 22 (8), 4096. doi:10.3390/ijms22084096
Chifenti, B., Locci, M. T., Lazzeri, G., Guagnozzi, M., Dinucci, D., Chiellini, F., et al. (2013). Autophagy-related protein LC3 and Beclin-1 in the first trimester of pregnancy. Clin. Exp. Reprod. Med. 40 (1), 33–37. doi:10.5653/cerm.2013.40.1.33
Choi, H. S., Cho, S. G., Kim, M. K., Kim, M. S., Moon, S. H., Kim, I. H., et al. (2016). Decursin in Angelica gigas Nakai (AGN) enhances doxorubicin chemosensitivity in NCI/ADR-RES ovarian cancer cells via inhibition of P-glycoprotein expression. Phytother. Res. 30 (12), 2020–2026. doi:10.1002/ptr.5708
Choi, K. O., Lee, I., Paik, S. Y., Kim, D. E., Lim, J. D., Kang, W. S., et al. (2012). Ultrafine Angelica gigas powder normalizes ovarian hormone levels and has antiosteoporosis properties in ovariectomized rats: particle size effect. J. Med. Food 15 (10), 863–872. doi:10.1089/jmf.2011.2047
Choi, S. R., Lee, J. H., Kim, J. Y., Park, K. W., Jeong, I. Y., Shim, K. H., et al. (2011). Decursin from Angelicagigas Nakai induces apoptosis in RC-58T/h/SA#4 primary human prostate cancer cells via a mitochondria-related caspase pathway. Food Chem. Toxicol. 49 (10), 2517–2523. doi:10.1016/j.fct.2011.06.016
Debela, D. T., Muzazu, S. G., Heraro, K. D., Ndalama, M. T., Mesele, B. W., Haile, D. C., et al. (2021). New approaches and procedures for cancer treatment: current perspectives. SAGE Open Med. 9, 20503121211034366. doi:10.1177/20503121211034366
Debnath, J., Gammoh, N., and Ryan, K. M. (2023). Autophagy and autophagy-related pathways in cancer. Nat. Rev. Mol. Cell. Biol. 24 (8), 560–575. doi:10.1038/s41580-023-00585-z
Deng, D., and Shah, K. (2020). TRAIL of hope meeting resistance in cancer. Trends Cancer 6 (12), 989–1001. doi:10.1016/j.trecan.2020.06.006
de Souza, A. S. C., Goncalves, L. B., Lepique, A. P., and de Araujo-Souza, P. S. (2020). The role of autophagy in tumor immunology-complex mechanisms that may Be explored therapeutically. Front. Oncol. 10, 603661. doi:10.3389/fonc.2020.603661
Edwards, A. M., Arrowsmith, C. H., Bountra, C., Bunnage, M. E., Feldmann, M., Knight, J. C., et al. (2015). Preclinical target validation using patient-derived cells. Nat. Rev. Drug Discov. 14 (3), 149–150. doi:10.1038/nrd4565
Ferrara, N., Gerber, H. P., and LeCouter, J. (2003). The biology of VEGF and its receptors. Nat. Med. 9 (6), 669–676. doi:10.1038/nm0603-669
Ge, Y., Yoon, S. H., Jang, H., Jeong, J. H., and Lee, Y. M. (2020). Decursin promotes HIF-1α proteasomal degradation and immune responses in hypoxic tumour microenvironment. Phytomedicine 78, 153318. doi:10.1016/j.phymed.2020.153318
George, V. C., Dellaire, G., and Rupasinghe, H. P. V. (2017). Plant flavonoids in cancer chemoprevention: role in genome stability. J. Nutr. Biochem. 45, 1–14. doi:10.1016/j.jnutbio.2016.11.007
Ghafouri-Fard, S., Khoshbakht, T., Hussen, B. M., Dong, P., Gassler, N., Taheri, M., et al. (2022). A review on the role of cyclin dependent kinases in cancers. Cancer Cell. Int. 22 (1), 325. doi:10.1186/s12935-022-02747-z
Guo, J., Jiang, C., Wang, Z., Lee, H. J., Hu, H., Malewicz, B., et al. (2007). A novel class of pyranocoumarin anti-androgen receptor signaling compounds. Mol. Cancer Ther. 6 (3), 907–917. doi:10.1158/1535-7163.MCT-06-0231
Han, H., Feng, F., and Li, H. (2021). Research advances on epigenetics and cancer metabolism. Zhejiang Da Xue Xue Bao Yi Xue Ban. 50 (1), 1–16. doi:10.3724/zdxbyxb-2021-0053
Hata, K., and Sano, K. (1969). Studies on coumarins FR.om the root of Angelica decursiva FR et SAV. I. The structure of decursin and decursidin. Yakugaku Zasshi 89 (4), 549–557. doi:10.1248/yakushi1947.89.4_549
He, Z., Wang, Y., Chen, Y., Geng, F., Jiang, Z., and Li, X. (2023). Angelica gigas Nakai: an overview on its chemical composition and pharmacological activity. Biochem. Syst. Ecol. 111, 104717. doi:10.1016/j.bse.2023.104717
Hinshaw, D. C., and Shevde, L. A. (2019). The tumor microenvironment innately modulates cancer progression. Cancer Res. 79 (18), 4557–4566. doi:10.1158/0008-5472.CAN-18-3962
Huang, J., Zhang, L., Wan, D., Zhou, L., Zheng, S., Lin, S., et al. (2021). Extracellular matrix and its therapeutic potential for cancer treatment. Signal Transduct. Target Ther. 6 (1), 153. doi:10.1038/s41392-021-00544-0
Huang, Q., Wang, M., Wang, M., Lu, Y., Wang, X., Chen, X., et al. (2023). Scutellaria baicalensis: a promising natural source of antiviral compounds for the treatment of viral diseases. Chin. J. Nat. Med. 21 (8), 563–575. doi:10.1016/S1875-5364(23)60401-7
Jablonska-Trypuc, A., Matejczyk, M., and Rosochacki, S. (2016). Matrix metalloproteinases (MMPs), the main extracellular matrix (ECM) enzymes in collagen degradation, as a target for anticancer drugs. J. Enzyme Inhib. Med. Chem. 31 (Suppl. 1), 177–183. doi:10.3109/14756366.2016.1161620
Jang, J., Jeong, S. J., Kwon, H. Y., Jung, J. H., Sohn, E. J., Lee, H. J., et al. (2013). Decursin and doxorubicin are in synergy for the induction of apoptosis via STAT3 and/or mTOR pathways in human multiple myeloma cells. Evid. Based Complement. Altern. Med. 2013, 506324. doi:10.1155/2013/506324
Jeon, R., Ryu, J.-h., and Cho, H. (2022). Pharmaceutical composition of three-membered ring having anti-cancer and anti-inflammatory activities. K.R patent application KR102465988 B1. November 17, 2023.
Ji, X., Huh, B., and Kim, S. U. (2008). Determination of biosynthetic pathway of decursin in hairy root culture of Angelica gigas. J. Korean Soc. Appl. Biol. Chem. 51 (4), 258–262. doi:10.3839/jksabc.2008.045
Jia, L. T., Chen, S. Y., and Yang, A. G. (2012). Cancer gene therapy targeting cellular apoptosis machinery. Cancer Treat. Rev. 38 (7), 868–876. doi:10.1016/j.ctrv.2012.06.008
Jiang, C., Guo, J., Wang, Z., Xiao, B., Lee, H. J., Lee, E. O., et al. (2007). Decursin and decursinol angelate inhibit estrogen-stimulated and estrogen-independent growth and survival of breast cancer cells. Breast Cancer Res. 9 (6), R77. doi:10.1186/bcr1790
Joo, M., Heo, J. B., Kim, S., Kim, N., Jeon, H. J., An, Y., et al. (2022). Decursin inhibits tumor progression in head and neck squamous cell carcinoma by downregulating CXCR7 expression in vitro. Oncol. Rep. 47 (2), 39. doi:10.3892/or.2021.8250
Jung, M. H., Lee, S. H., Ahn, E. M., and Lee, Y. M. (2009). Decursin and decursinol angelate inhibit VEGF-induced angiogenesis via suppression of the VEGFR-2-signaling pathway. Carcinogenesis 30 (4), 655–661. doi:10.1093/carcin/bgp039
Jung, M. Y., Seo, C. S., Baek, S. E., Lee, J., Shin, M. S., Kang, K. S., et al. (2019). Analysis and identification of active compounds from gami-soyosan toxic to MCF-7 human breast adenocarcinoma cells. Biomolecules 9 (7), 272. doi:10.3390/biom9070272
Jung, S. Y., Choi, J. H., Kwon, S. M., Masuda, H., Asahara, T., and Lee, Y. M. (2012). Decursin inhibits vasculogenesis in early tumor progression by suppression of endothelial progenitor cell differentiation and function. J. Cell. Biochem. 113 (5), 1478–1487. doi:10.1002/jcb.24085
Ke, C., Liu, Y., Zhang, Z., Zhang, J., Zhu, Y., Liu, J., et al. (2018). Decursin inhibits ECl09 cells proliferation via suppression of JAK2/STAT3 signaling pathway. Chin. J. Thorac. Cardiovasc Surg. 34 (4), 230–236. doi:10.3760/cma.i.issn.1001-4497.2018.04.009
Kim, B. S., Seo, H., Kim, H. J., Bae, S. M., Son, H. N., Lee, Y. J., et al. (2015). Decursin from Angelica gigas Nakai inhibits B16F10 melanoma growth through induction of apoptosis. J. Med. Food 18 (10), 1121–1127. doi:10.1089/jmf.2014.3397
Kim, D. S., Park, S. H., Lee, H. K., Choo, S. J., Lee, J. H., Song, G. Y., et al. (2010a). Hypopigmentary action of dihydropyranocoumarin D2, a decursin derivative, as a MITF-degrading agent. J. Nat. Prod. 73 (5), 797–800. doi:10.1021/np900455j
Kim, E., Nam, J., Chang, W., Zulfugarov, I. S., Okhlopkova, Z. M., Olennikov, D., et al. (2018). Angelica gigas Nakai and decursin downregulate myc expression to promote cell death in B-cell lymphoma. Sci. Rep. 8 (1), 10590. doi:10.1038/s41598-018-28619-z
Kim, H. H., Sik Bang, S., Seok Choi, J., Han, H., and Kim, I. H. (2005). Involvement of PKC and ROS in the cytotoxic mechanism of anti-leukemic decursin and its derivatives and their structure-activity relationship in human K562 erythroleukemia and U937 myeloleukemia cells. Cancer Lett. 223 (2), 191–201. doi:10.1016/j.canlet.2004.10.025
Kim, H. J., Kim, S. M., Park, K. R., Jang, H. J., Na, Y. S., Ahn, K. S., et al. (2011). Decursin chemosensitizes human multiple myeloma cells through inhibition of STAT3 signaling pathway. Cancer Lett. 301 (1), 29–37. doi:10.1016/j.canlet.2010.11.002
Kim, I.-H., and Kim, K.-S. (1999). Decursin as an anti-tumour agent. K.R. patent application KR0176413. March 20, 1999.
Kim, J., Yun, M., Kim, E. O., Jung, D. B., Won, G., Kim, B., et al. (2016). Decursin enhances TRAIL-induced apoptosis through oxidative stress mediated-endoplasmic reticulum stress signalling in non-small cell lung cancers. Br. J. Pharmacol. 173 (6), 1033–1044. doi:10.1111/bph.13408
Kim, J. H., Jeong, S. J., Kwon, H. Y., Park, S. Y., Lee, H. J., Lee, H. J., et al. (2010b). Decursin prevents cisplatin-induced apoptosis via the enhancement of antioxidant enzymes in human renal epithelial cells. Biol. Pharm. Bull. 33 (8), 1279–1284. doi:10.1248/bpb.33.1279
Kim, J. H., Jung, J. H., Kim, S. H., and Jeong, S. J. (2014a). Decursin exerts anti-cancer activity in MDA-MB-231 breast cancer cells via inhibition of the Pin1 activity and enhancement of the Pin1/p53 association. Phytother. Res. 28 (2), 238–244. doi:10.1002/ptr.4986
Kim, J. H., Kim, J. H., Lee, Y. M., Ahn, E. M., Kim, K. W., and Yu, Y. S. (2009a). Decursin inhibits retinal neovascularization via suppression of VEGFR-2 activation. Mol. Vis. 15, 1868–1875. doi:10.1038/jcbfm.2009
Kim, J. H., Kim, J. H., Lee, Y. M., Ahn, E. M., Kim, K. W., and Yu, Y. S. (2009b). Decursin inhibits VEGF-mediated inner blood-retinal barrier breakdown by suppression of VEGFR-2 activation. J. Cereb. Blood Flow. Metab. 29 (9), 1559–1567. doi:10.1038/jcbfm.2009.78
Kim, J. M., Noh, E. M., Kim, M. S., Hwang, J. K., Hwang, H. Y., Ryu, D. G., et al. (2014b). Decursin prevents TPA-induced invasion through suppression of PKCα/p38/NF-κB-dependent MMP-9 expression in MCF-7 human breast carcinoma cells. Int. J. Oncol. 44 (5), 1607–1613. doi:10.3892/ijo.2014.2327
Kim, K., Liu, X., Yook, C., and Park, S. (2006). Coumarins from angelica czernaevia kita forma dentate Yook. Nat. Prod. Res. Dev. (01), 7–10. doi:10.16333/j.1001-6880.2006.01.003
Kim, S., Kim, J. E., Kim, N., Joo, M., Lee, M. W., Jeon, H. J., et al. (2019). Decursin inhibits tumor growth, migration, and invasion in gastric cancer by down-regulating CXCR7 expression. Am. J. Cancer Res. 9 (9), 2007–2018. doi:10.1093/carcin/bgp039
Kim, S., Lee, S. I., Kim, N., Joo, M., Lee, K. H., Lee, M. W., et al. (2021). Decursin inhibits cell growth and autophagic flux in gastric cancer via suppression of cathepsin C. Am. J. Cancer Res. 11 (4), 1304–1320.
Kim, W. J., Lee, M. Y., Kim, J. H., Suk, K., and Lee, W. H. (2010c). Decursinol angelate blocks transmigration and inflammatory activation of cancer cells through inhibition of PI3K, ERK and NF-kappaB activation. Cancer Lett. 296 (1), 35–42. doi:10.1016/j.canlet.2010.03.012
Kim, W. J., Lee, S. J., Choi, Y. D., and Moon, S. K. (2010d). Decursin inhibits growth of human bladder and colon cancer cells via apoptosis, G1-phase cell cycle arrest and extracellular signal-regulated kinase activation. Int. J. Mol. Med. 25 (4), 635–641. doi:10.3892/ijmm_00000386
Kreiner, J., Pang, E., Lenon, G. B., and Yang, A. W. H. (2017). Saposhnikoviae divaricata: a phytochemical, pharmacological, and pharmacokinetic review. Chin. J. Nat. Med. 15 (4), 255–264. doi:10.1016/S1875-5364(17)30042-0
Kumar, P., Rana, V., and Singh, A. N. (2022). Angelica glauca Edgew. – a comprehensive review. J. Appl. Res. Med. Aromatic Plants 31, 100397. doi:10.1016/j.jarmap.2022.100397
Kupeli Akkol, E., Genc, Y., Karpuz, B., Sobarzo-Sanchez, E., and Capasso, R. (2020). Coumarins and coumarin-related compounds in pharmacotherapy of cancer. Cancers (Basel) 12 (7), 1959. doi:10.3390/cancers12071959
Kweon, B., Han, Y. H., Kee, J. Y., Mun, J. G., Jeon, H. D., Yoon, D. H., et al. (2020). Effect of Angelica gigas Nakai ethanol extract and decursin on human pancreatic cancer cells. Molecules 25 (9), 2028. doi:10.3390/molecules25092028
Lee, H. J., Lee, H. J., Lee, E. O., Lee, J. H., Lee, K. S., Kim, K. H., et al. (2009). In vivo anti-cancer activity of Korean Angelica gigas and its major pyranocoumarin decursin. Am. J. Chin. Med. 37 (1), 127–142. doi:10.1142/S0192415X09006722
Lee, S., Lee, Y. S., Jung, S. H., Shin, K. H., Kim, B. K., and Kang, S. S. (2003). Anti-tumor activities of decursinol angelate and decursin from Angelica gigas. Arch. Pharm. Res. 26 (9), 727–730. doi:10.1007/BF02976682
Lee, S. H., Lee, J. H., Kim, E. J., Kim, W. J., Suk, K., Kim, J. H., et al. (2012). A novel derivative of decursin, CSL-32, blocks migration and production of inflammatory mediators and modulates PI3K and NF-κB activities in HT1080 cells. Cell. Biol. Int. 36 (7), 683–688. doi:10.1042/CBI20110257
Lee, S. Y., Lee, J. J., Nam, S., Kang, W. S., Yoon, I. S., and Cho, H. J. (2017). Fabrication of polymer matrix-free nanocomposites based on Angelica gigas Nakai extract and their application to breast cancer therapy. Colloids Surf. B Biointerfaces 159, 781–790. doi:10.1016/j.colsurfb.2017.08.040
Lee, W., Yuseok, O., Yang, S., Lee, B. S., Lee, J. H., Park, E. K., et al. (2019). JH-4 reduces HMGB1-mediated septic responses and improves survival rate in septic mice. J. Cell. Biochem. 120 (4), 6277–6289. doi:10.1002/jcb.27914
Lee, Y.-m., Ahn, E.-m., and Jung, M. h. (2010). Composition of treatment for VEGF-induced angiogenesis-related diseases containing decursin and decursinol angelate. K.R Pat. Appl. KR20100009295. January 27, 2010.
Levy, J. M. M., Towers, C. G., and Thorburn, A. (2017). Targeting autophagy in cancer. Nat. Rev. Cancer 17 (9), 528–542. doi:10.1038/nrc.2017.53
Li, C. C., Yu, F. S., Fan, M. J., Chen, Y. Y., Lien, J. C., Chou, Y. C., et al. (2017). Anticancer effects of cantharidin in A431 human skin cancer (Epidermoid carcinoma) cells in vitro and in vivo. Environ. Toxicol. 32 (3), 723–738. doi:10.1002/tox.22273
Li, J., Ding, Y., Li, X. C., Ferreira, D., Khan, S., Smillie, T., et al. (2009). Scuteflorins A and B, dihydropyranocoumarins from Scutellaria lateriflora. J. Nat. Prod. 72 (6), 983–987. doi:10.1021/np900068t
Li, J., Wang, H., Wang, L., Tan, R., Zhu, M., Zhong, X., et al. (2018). Decursin inhibits the growth of HepG2 hepatocellular carcinoma cells via Hippo/YAP signaling pathway. Phytother. Res. 32 (12), 2456–2465. doi:10.1002/ptr.6184
Liang, W. H., Chang, T. W., and Charng, Y. C. (2018). Effects of drying methods on contents of bioactive compounds and antioxidant activities of Angelica dahurica. Food Sci. Biotechnol. 27 (4), 1085–1092. doi:10.1007/s10068-018-0359-4
Liang, X. J., Chen, C., Zhao, Y., and Wang, P. C. (2010). Circumventing tumor resistance to chemotherapy by nanotechnology. Methods Mol. Biol. 596, 467–488. doi:10.1007/978-1-60761-416-6_21
Liou, Y. C., Ryo, A., Huang, H. K., Lu, P. J., Bronson, R., Fujimori, F., et al. (2002). Loss of Pin1 function in the mouse causes phenotypes resembling cyclin D1-null phenotypes. Proc. Natl. Acad. Sci. U. S. A. 99 (3), 1335–1340. doi:10.1073/pnas.032404099
Loh, C. Y., Chai, J. Y., Tang, T. F., Wong, W. F., Sethi, G., Shanmugam, M. K., et al. (2019). The E-cadherin and N-cadherin switch in epithelial-to-mesenchymal transition: signaling, therapeutic implications, and challenges. Cells 8 (10), 1118. doi:10.3390/cells8101118
Lugano, R., Ramachandran, M., and Dimberg, A. (2020). Tumor angiogenesis: causes, consequences, challenges and opportunities. Cell. Mol. Life Sci. 77 (9), 1745–1770. doi:10.1007/s00018-019-03351-7
Mahat, B., Chae, J. W., Baek, I. H., Song, G. Y., Song, J. S., Cho, S. K., et al. (2012). Physicochemical characterization and toxicity of decursin and their derivatives from Angelica gigas. Biol. Pharm. Bull. 35 (7), 1084–1090. doi:10.1248/bpb.b12-00046
Maier, M. E. (2015). Design and synthesis of analogues of natural products. Org. Biomol. Chem. 13 (19), 5302–5343. doi:10.1039/c5ob00169b
Mareninova, O. A., Jia, W., Gretler, S. R., Holthaus, C. L., Thomas, D. D. H., Pimienta, M., et al. (2020). Transgenic expression of GFP-LC3 perturbs autophagy in exocrine pancreas and acute pancreatitis responses in mice. Autophagy 16 (11), 2084–2097. doi:10.1080/15548627.2020.1715047
Matusiewicz, M., Baczek, K. B., Kosieradzka, I., Niemiec, T., Grodzik, M., Szczepaniak, J., et al. (2019). Effect of juice and extracts from Saposhnikovia divaricata root on the colon cancer cells caco-2. Int. J. Mol. Sci. 20 (18), 4526. doi:10.3390/ijms20184526
Millimouno, F. M., Dong, J., Yang, L., Li, J., and Li, X. (2014). Targeting apoptosis pathways in cancer and perspectives with natural compounds from mother nature. Cancer Prev. Res. (Phila) 7 (11), 1081–1107. doi:10.1158/1940-6207.CAPR-14-0136
Mohammad, R. M., Muqbil, I., Lowe, L., Yedjou, C., Hsu, H. Y., Lin, L. T., et al. (2015). Broad targeting of resistance to apoptosis in cancer. Semin. Cancer Biol. 35, S78–S103. doi:10.1016/j.semcancer.2015.03.001
Morana, O., Wood, W., and Gregory, C. D. (2022). The apoptosis paradox in cancer. Int. J. Mol. Sci. 23 (3), 1328. doi:10.3390/ijms23031328
Nam, S., Lee, S. Y., Kang, W. S., and Cho, H. J. (2018a). Development of resveratrol-loaded herbal extract-based nanocomposites and their application to the therapy of ovarian cancer. Nanomater. (Basel) 8 (6), 384. doi:10.3390/nano8060384
Nam, S., Lee, S. Y., Kim, J. J., Kang, W. S., Yoon, I. S., and Cho, H. J. (2018b). Polydopamine-coated nanocomposites of Angelica gigas Nakai extract and their therapeutic potential for triple-negative breast cancer cells. Colloids Surf. B Biointerfaces 165, 74–82. doi:10.1016/j.colsurfb.2018.02.014
Nazio, F., Bordi, M., Cianfanelli, V., Locatelli, F., and Cecconi, F. (2019). Autophagy and cancer stem cells: molecular mechanisms and therapeutic applications. Cell. Death Differ. 26 (4), 690–702. doi:10.1038/s41418-019-0292-y
Noman, M. Z., Desantis, G., Janji, B., Hasmim, M., Karray, S., Dessen, P., et al. (2014). PD-L1 is a novel direct target of HIF-1α, and its blockade under hypoxia enhanced MDSC-mediated T cell activation. J. Exp. Med. 211 (5), 781–790. doi:10.1084/jem.20131916
Oh, S. T., Lee, S., Hua, C., Koo, B. S., Pak, S. C., Kim, D. I., et al. (2019). Decursin induces apoptosis in glioblastoma cells, but not in glial cells via a mitochondria-related caspase pathway. Korean J. Physiol. Pharmacol. 23 (1), 29–35. doi:10.4196/kjpp.2019.23.1.29
Palazon, A., Aragones, J., Morales-Kastresana, A., de Landazuri, M. O., and Melero, I. (2012). Molecular pathways: hypoxia response in immune cells fighting or promoting cancer. Clin. Cancer Res. 18 (5), 1207–1213. doi:10.1158/1078-0432.CCR-11-1591
Palazon, A., Goldrath, A. W., Nizet, V., and Johnson, R. S. (2014). HIF transcription factors, inflammation, and immunity. Immunity 41 (4), 518–528. doi:10.1016/j.immuni.2014.09.008
Park, H. J., Park, S. H., Choi, Y. H., and Chi, G. Y. (2021). The root extract of Scutellaria baicalensis induces apoptosis in EGFR TKI-resistant human lung cancer cells by inactivation of STAT3. Int. J. Mol. Sci. 22 (10), 5181. doi:10.3390/ijms22105181
Park, J. H., Park, N. I., Xu, H., and Park, S. U. (2010). Cloning and characterization of phenylalanine ammonia-lyase and cinnamate 4-hydroxylase and pyranocoumarin biosynthesis in Angelica gigas. J. Nat. Prod. 73 (8), 1394–1397. doi:10.1021/np1003356
Plaza-Zabala, A., Sierra-Torre, V., and Sierra, A. (2020). Assessing autophagy in microglia: a two-step model to determine autophagosome formation, degradation, and net turnover. Front. Immunol. 11, 620602. doi:10.3389/fimmu.2020.620602
Reddy, C. S., Kim, S. C., Hur, M., Kim, Y. B., Park, C. G., Lee, W. M., et al. (2017). Natural Korean medicine dang-gui: biosynthesis, effective extraction and formulations of major active pyranocoumarins, their molecular action mechanism in cancer, and other biological activities. Molecules 22 (12), 2170. doi:10.3390/molecules22122170
Ribatti, D., Tamma, R., and Annese, T. (2020). Epithelial-mesenchymal transition in cancer: a historical overview. Transl. Oncol. 13 (6), 100773. doi:10.1016/j.tranon.2020.100773
Roberts, A. A., Ryan, K. S., Moore, B. S., and Gulder, T. A. (2010). Total (bio)synthesis: strategies of nature and of chemists. Top. Curr. Chem. 297, 149–203. doi:10.1007/128_2010_79
Roth, A. D., Maibach, R., Martinelli, G., Fazio, N., Aapro, M. S., Pagani, O., et al. (2000). Docetaxel (Taxotere)-cisplatin (TC): an effective drug combination in gastric carcinoma Swiss group for clinical cancer research (SAKK), and the European institute of oncology (EIO). Ann. Oncol. 11 (3), 301–306. doi:10.1023/a:1008342013224
Rousseau, S., Houle, F., Kotanides, H., Witte, L., Waltenberger, J., Landry, J., et al. (2000). Vascular endothelial growth factor (VEGF)-driven actin-based motility is mediated by VEGFR2 and requires concerted activation of stress-activated protein kinase 2 (SAPK2/p38) and geldanamycin-sensitive phosphorylation of focal adhesion kinase. J. Biol. Chem. 275 (14), 10661–10672. doi:10.1074/jbc.275.14.10661
Ruan, B. F., and Zhu, H. L. (2012). The chemistry and biology of the bryostatins: potential PKC inhibitors in clinical development. Curr. Med. Chem. 19 (16), 2652–2664. doi:10.2174/092986712800493020
Saeed, M. A., and Sabir, A. W. (2008). Irritant and cytotoxic coumarins from Angelica glauca Edgew roots. J. Asian Nat. Prod. Res. 10 (1-2), 49–58. doi:10.1080/10286020701273759
Said, A. H., Raufman, J. P., and Xie, G. (2014). The role of matrix metalloproteinases in colorectal cancer. Cancers (Basel) 6 (1), 366–375. doi:10.3390/cancers6010366
Saini, K. S., and Twelves, C. (2021). Determining lines of therapy in patients with solid cancers: a proposed new systematic and comprehensive framework. Br. J. Cancer 125 (2), 155–163. doi:10.1038/s41416-021-01319-8
Shehzad, A., Parveen, S., Qureshi, M., Subhan, F., and Lee, Y. S. (2018). Decursin and decursinol angelate: molecular mechanism and therapeutic potential in inflammatory diseases. Inflamm. Res. 67 (3), 209–218. doi:10.1007/s00011-017-1114-7
Sherr, C. J. (1996). Cancer cell cycles. Science 274 (5293), 1672–1677. doi:10.1126/science.274.5293.1672
Shi, Y. (1997). Study on the chemical composition of qiangwu in xinjiang Province (China: China Academy of Chinese Medical Sciences). master's thesis.
Siegel, R. L., Miller, K. D., Wagle, N. S., and Jemal, A. (2023). Cancer statistics, 2023. CA Cancer J. Clin. 73 (1), 17–48. doi:10.3322/caac.21763
Son, H., and Moon, A. (2010). Epithelial-mesenchymal transition and cell invasion. Toxicol. Res. 26 (4), 245–252. doi:10.5487/TR.2010.26.4.245
Son, S. H., Kim, M. J., Chung, W. Y., Son, J. A., Kim, Y. S., Kim, Y. C., et al. (2009). Decursin and decursinol inhibit VEGF-induced angiogenesis by blocking the activation of extracellular signal-regulated kinase and c-Jun N-terminal kinase. Cancer Lett. 280 (1), 86–92. doi:10.1016/j.canlet.2009.02.012
Son, S. H., Park, K. K., Park, S. K., Kim, Y. C., Kim, Y. S., Lee, S. K., et al. (2011). Decursin and decursinol from Angelica gigas inhibit the lung metastasis of murine colon carcinoma. Phytother. Res. 25 (7), 959–964. doi:10.1002/ptr.3372
Song, G. Y., Lee, J. H., Cho, M., Park, B. S., Kim, D. E., and Oh, S. (2007). Decursin suppresses human androgen-independent PC3 prostate cancer cell proliferation by promoting the degradation of beta-catenin. Mol. Pharmacol. 72 (6), 1599–1606. doi:10.1124/mol.107.040253
Song, Z., Li, B., and Tian, K. (2022). Progress in the study of chemical composition and pharmacological effects of Radix et Rhizoma Pinelliae and Radix et Rhizoma Pinelliae. Chin. Traditional Herb. Drugs 53 (03), 948–964. doi:10.7501/j.issn.0253-2670.2022.03.035
Soon-ok, C., Yoong-chan, K., Hee-jun, L., Young-deuk, C., Ik-hwan, K., Eui-Jae, Y., et al. (2012). Compositions supportive for anticancers containing decursin and/or decursinol angelate, or angelica extract containing decursin and/or decursinol angelate. K.R Pat. Appl. 2012, KR20120078691.
Steeg, P. S. (2016). Targeting metastasis. Nat. Rev. Cancer 16 (4), 201–218. doi:10.1038/nrc.2016.25
Sung, H., Ferlay, J., Siegel, R. L., Laversanne, M., Soerjomataram, I., Jemal, A., et al. (2021). Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 71 (3), 209–249. doi:10.3322/caac.21660
Tang, S. N., Zhang, J., Wu, W., Jiang, P., Puppala, M., Zhang, Y., et al. (2015). Chemopreventive effects of Korean angelica versus its major pyranocoumarins on two lineages of transgenic adenocarcinoma of mouse prostate carcinogenesis. Cancer Prev. Res. (Phila) 8 (9), 835–844. doi:10.1158/1940-6207.CAPR-15-0051
Tanifuji, R., Koketsu, K., Takakura, M., Asano, R., Minami, A., Oikawa, H., et al. (2018). Chemo-enzymatic total syntheses of jorunnamycin A, Saframycin A, and N-fmoc Saframycin Y3. J. Am. Chem. Soc. 140 (34), 10705–10709. doi:10.1021/jacs.8b07161
Tarver, T. (2012). Cancer facts & figures 2012. American cancer society (ACS). J. Consumer Health Internet 16 (3), 366–367. doi:10.1080/15398285.2012.701177
Tewey, K. M., Rowe, T. C., Yang, L., Halligan, B. D., and Liu, L. F. (1984). Adriamycin-induced DNA damage mediated by mammalian DNA topoisomerase II. Science 226 (4673), 466–468. doi:10.1126/science.6093249
Vasan, N., Baselga, J., and Hyman, D. M. (2019). A view on drug resistance in cancer. Nature 575 (7782), 299–309. doi:10.1038/s41586-019-1730-1
Vialart, G., Hehn, A., Olry, A., Ito, K., Krieger, C., Larbat, R., et al. (2012). A 2-oxoglutarate-dependent dioxygenase from Ruta graveolens L. exhibits p-coumaroyl CoA 2'-hydroxylase activity (C2'H): a missing step in the synthesis of umbelliferone in plants. Plant J. 70 (3), 460–470. doi:10.1111/j.1365-313X.2011.04879.x
Wang, G., Sun, S., Wu, B., and Liu, J. (2021). Coumarins as potential anti-drug resistant cancer agents: a mini review. Curr. Top. Med. Chem. 21 (19), 1725–1736. doi:10.2174/1568026620999201113110041
Wang, H., Shao, Y., Ruan, Z., and Mao, M. (2019). Inhibition effect of decursin on proliferation and invasion of oral squamous cell carcinoma cells via Nox1/akt signaling pathway. China Pharm. 28 (22), 10–13. doi:10.3969/j.issn.1006-4931.2019.22.004
Wu, S. B., Pang, F., Wen, Y., Zhang, H. F., Zhao, Z., and Hu, J. F. (2010). Antiproliferative and apoptotic activities of linear furocoumarins from Notopterygium incisum on cancer cell lines. Planta Med. 76 (1), 82–85. doi:10.1055/s-0029-1185971
Yang, X., Xu, B., Ran, F., Wang, R., Wu, J., and Cui, J. (2007). Inhibitory effects of 11 coumarin compounds against growth of human bladder carcinoma cell line E-J in vitro. J. Chin. Integr. Med. 5 (1), 56–60. doi:10.3736/jcim20070111
Yang, Y., Hu, Y., Jiang, Y., Fu, X., and You, F. (2023). Decursin affects proliferation, apoptosis, and migration of colorectal cancer cells through PI3K/Akt signaling pathway. China J. Chin. Materia Medica 48 (09), 2334–2342. doi:10.19540/j.cnki.cjcmm.20230117.703
Yim, D., Singh, R. P., Agarwal, C., Lee, S., Chi, H., and Agarwal, R. (2005). A novel anticancer agent, decursin, induces G1 arrest and apoptosis in human prostate carcinoma cells. Cancer Res. 65 (3), 1035–1044. doi:10.1158/0008-5472.1035.65.3
Yu, X., Che, Z., and Xu, H. (2017). Recent advances in the chemistry and biology of podophyllotoxins. Chemistry 23 (19), 4467–4526. doi:10.1002/chem.201602472
Zacchino, S. A., Butassi, E., Cordisco, E., and Svetaz, L. A. (2017). Hybrid combinations containing natural products and antimicrobial drugs that interfere with bacterial and fungal biofilms. Phytomedicine 37, 14–26. doi:10.1016/j.phymed.2017.10.021
Zhang, J., Li, L., Hale, T. W., Chee, W., Xing, C., Jiang, C., et al. (2015). Single oral dose pharmacokinetics of decursin and decursinol angelate in healthy adult men and women. PLoS One 10 (2), e0114992. doi:10.1371/journal.pone.0114992
Zhang, J., Li, L., Jiang, C., Xing, C., Kim, S. H., and Lu, J. (2012a). Anti-cancer and other bioactivities of Korean Angelica gigas Nakai (AGN) and its major pyranocoumarin compounds. Anticancer Agents Med. Chem. 12 (10), 1239–1254. doi:10.2174/187152012803833071
Zhang, P., and Yang, X. W. (2008). Studies on chemical constituents in roots and rhizomes of Notopterygium incisum. Chin. J. Chin. Mater Med. 3, 2918–2921. doi:10.3321/j.issn:1001-5302.2008.24.012
Zhang, Y., Shaik, A. A., Xing, C., Chai, Y., Li, L., Zhang, J., et al. (2012b). A synthetic decursin analog with increased in vivo stability suppresses androgen receptor signaling in vitro and in vivo. Investig. New Drugs 30 (5), 1820–1829. doi:10.1007/s10637-011-9738-x
Zhao, B., Yang, X., Yang, X., and Zhang, L. (2010). Study on the chemical composition of Fangfeng. China J. Chin. Materia Medica 35 (12), 1569–1572. doi:10.4268/cjcmm20101214
Zheng, Y. M., Shen, J. Z., Wang, Y., Lu, A. X., and Ho, W. S. (2016). Anti-oxidant and anti-cancer activities of Angelica dahurica extract via induction of apoptosis in colon cancer cells. Phytomedicine 23 (11), 1267–1274. doi:10.1016/j.phymed.2015.11.008
Zhu, M. L., Li, J. C., Wang, L., Zhong, X., Zhang, Y. W., Tan, R. Z., et al. (2021). Decursin inhibits the growth of HeLa cervical cancer cells through PI3K/Akt signaling. J. Asian Nat. Prod. Res. 23 (6), 584–595. doi:10.1080/10286020.2020.1821669
Zunina, F., Gambetta, R., and Di Marco, A. (1975). The inhibition in vitro of DNA polymerase and RNA polymerases by daunomycin and adriamycin. Biochem. Pharmacol. 24 (2), 309–311. doi:10.1016/0006-2952(75)90300-7

Keywords: decursin, sources, derivatives, anticancer activity, mechanisms, safety
Citation: Chu Y, Yuan Q, Jiang H, Wu L, Xie Y, Zhang X and Li L (2024) A comprehensive review of the anticancer effects of decursin. Front. Pharmacol. 15:1303412. doi: 10.3389/fphar.2024.1303412
Received: 28 September 2023; Accepted: 02 February 2024;
Published: 20 February 2024.
Edited by:
Zhaofeng Liang, Jiangsu University, ChinaReviewed by:
Ebtesam Abdullah Al-Suhaimi, King Abdulaziz and His Companions Foundation for Giftedness and Creativity, Saudi ArabiaCopyright © 2024 Chu, Yuan, Jiang, Wu, Xie, Zhang and Li. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Xiaofen Zhang, c3BhY2V3YWxrZXJAdG9tLmNvbQ==; Lin Li, bGlubGk4MkBjcXUuZWR1LmNuMg==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.