 Kwang-Il To
Kwang-Il To Ya-Ni Wang
Ya-Ni Wang Ying-Hua Jin
Ying-Hua Jin
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Pharmacol. , 31 August 2022
Sec. Inflammation Pharmacology
Volume 13 - 2022 | https://doi.org/10.3389/fphar.2022.953871
This article is part of the Research Topic Recent Advances of Minor Saponins in Panax species View all 10 articles
Inflammation is an innate immune response to infection, and it is the main factor causing bodily injury and other complications in the pathological process. Ginsenoside Rh4 (G-Rh4), a minor ginsenoside of Panax ginseng C. A. Meyer and Panax notoginseng, has excellent pharmacological properties. However, many of its major pharmacological mechanisms, including anti-inflammatory actions, remain unrevealed. In this study, network pharmacology and an experimental approach were employed to elucidate the drug target and pathways of G-Rh4 in treating inflammation. The potential targets of G-Rh4 were selected from the multi-source databases, and 58 overlapping gene symbols related to G-Rh4 and inflammation were obtained for generating a protein–protein interaction (PPI) network. Molecular docking revealed the high affinities between key proteins and G-Rh4. Gene ontology (GO) and pathway enrichment analyses were used to analyze the screened core targets and explore the target–pathway networks. It was found that the JAK-STAT signaling pathway, TNF signaling pathway, NF-κB signaling pathway, and PI3K-Akt signaling pathway may be the key and main pathways of G-Rh4 to treat inflammation. Additionally, the potential molecular mechanisms of G-Rh4 predicted from network pharmacology analysis were validated in RAW264.7 cells. RT-PCR, Western blot, and ELISA analysis indicated that G-Rh4 significantly inhibited the production of pro-inflammatory cytokines such as TNF-α, IL-6, and IL-1β, as well as inflammation-related enzymes in lipopolysaccharide (LPS)-stimulated RAW264.7 cells. Moreover, in vitro experiments evaluated that Ginsenoside Rh4 exerts anti-inflammatory effects via the NF-κB and STAT3 signaling pathways. It is believed that our study will provide the basic scientific evidence that G-Rh4 has potential anti-inflammatory effects for further clinical studies.
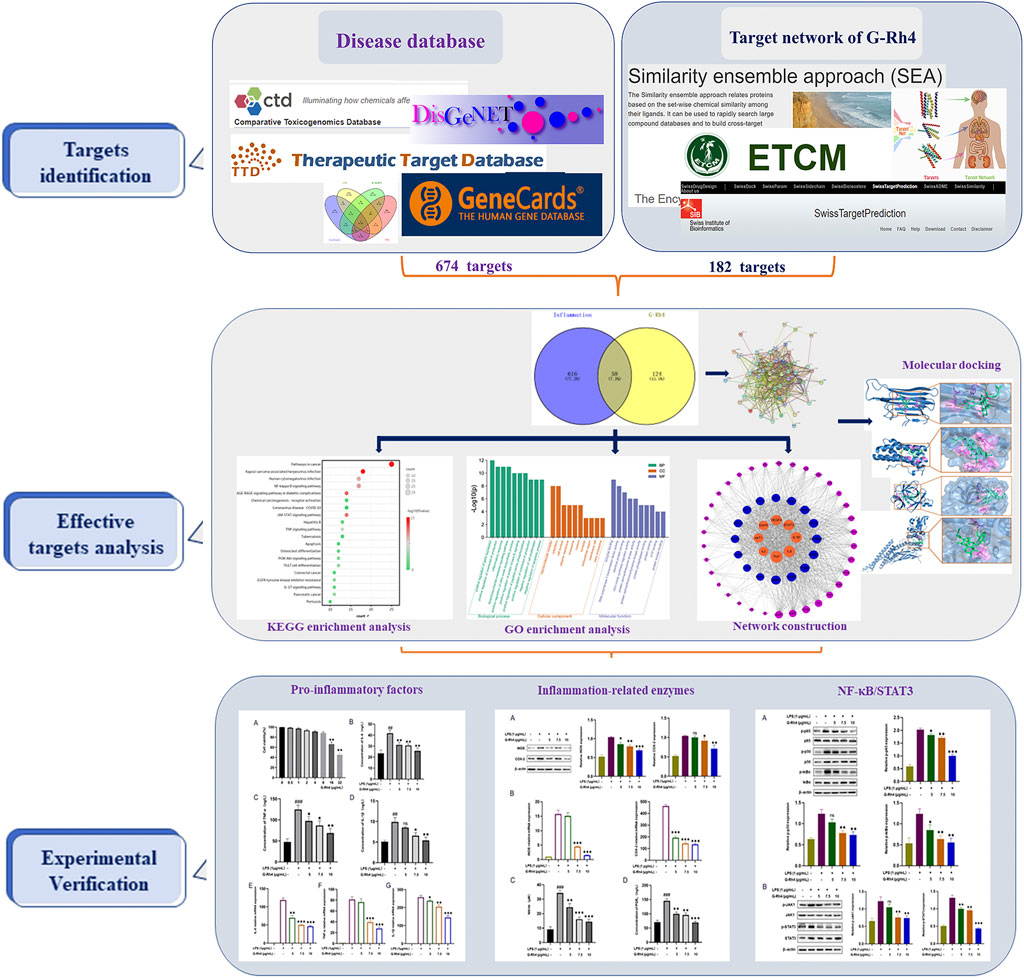
GRAPHICAL ABSTRACT
The occurrence of self-limited inflammation is critical for survival during physical injury and infection; however, the persistence of inflammation leads to several pivotal diseases that collectively represent the leading causes of disability and mortality worldwide (Furman et al., 2019; Panigrahy et al., 2020). Recent research has revealed that chronic inflammatory diseases have been recognized as the most important cause of death in the world today (Libby, 2021), and more than 50% of deaths are attributed to inflammation-related diseases such as ischemic heart disease, cancer, non-alcoholic fatty liver disease (NAFLD), and diabetes mellitus (GBD 2017 Causes of Death Collaborators, 2018). In addition, it was reported that typical biomarkers of acute inflammation can predict the incidence rate and mortality of various diseases (Arai et al., 2015), but this approach also has limitations. To address the limitations of assessment with a few selected inflammatory biomarkers, researchers have adopted a multi-dimensional approach, including analyzing a large number of inflammatory markers and then combining these markers into more reliable indicators representing high inflammatory activity (Morrisette-Thomas et al., 2014). The anti-inflammatory drugs, including non-steroidal anti-inflammatory drugs, have been developed and applied in clinics, but studies have shown that these drugs frequently cause adverse effects such as liver injury (Schmeltzer et al., 2016). Looking for more effective and safe anti-inflammatory drugs is an urgent issue in the global medical community.
Ginsenoside is the main active ingredient of ginseng, which displayed a broad spectrum of activities, such as cancer cell toxicity, anti-inflammation, and enhancing immunity. G-Rh4 is a trace saponin in white ginseng, and it became the most abundant triol-type ginsenoside after the heating process. Compared with other saponins, G-Rh4 has relatively better water solubility (Wang et al., 2022), suggesting its potential clinical applications. However, due to its low content in fresh or white ginseng and consequent preparation difficulty, there are few studies on the pharmacological effects of G-Rh4; especially, its anti-inflammatory activity and underlying mechanism are largely unknown.
Network pharmacology is the construction and analysis of biological networks based on network database retrieval and computer simulation calculation to study the mechanism of drug action. In particular, in determining the pharmacological mechanism and safety of Chinese medicine, the application of network analysis is a new paradigm for traditional Chinese medicine, from empirical medicine to evidence-based medicine (Wang et al., 2021). Therefore, network pharmacology research, as a new interdisciplinary approach, is of great value to the research and development of modern Chinese medicine (Sun et al., 2020; Wang et al., 2021). In this study, we clarified the drug target and multiple pathways of G-Rh4 in the treatment of inflammation by network pharmacology and the experimental approach.
The PubChem database (https://pubchem.ncbi.nlm.nih.gov/) was searched to obtain the two-dimensional structure diagram in SDF format and the canonical smiles of G-Rh4 (Figure 1A). The main potential targets of ginsenoside-Rh4 were identified using the Swiss Target Prediction (http://swisstargetprediction.ch/), the TargetNet (http://targetnet.scbdd.com/), the Similarity Ensemble Approach (SEA) (https://sea.bkslab.org/), and the Encyclopedia of Traditional Chinese Medicine (ETCM) (http://www.tcmip.cn/ETCM/index.php/Home/Index/) database.
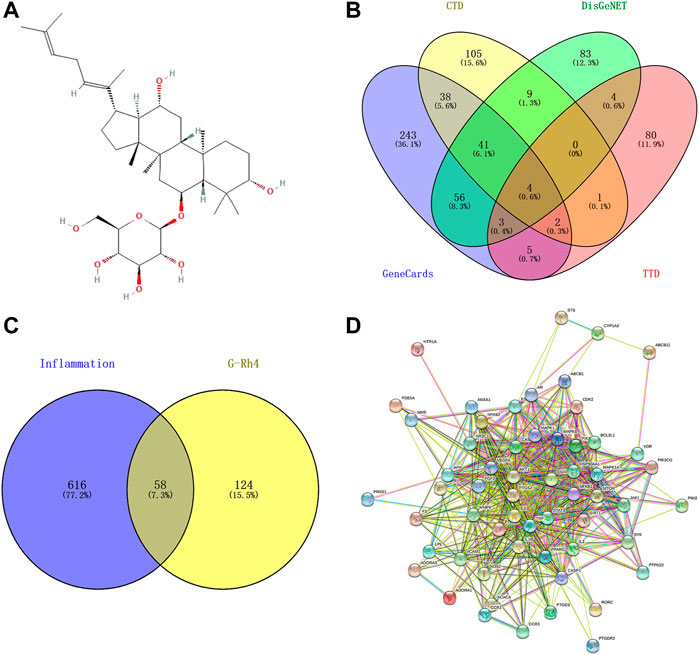
FIGURE 1. Targets screening involved in G-Rh4 for treating inflammation. (A) Molecule structure of G-Rh4. (B)Venn diagram of the potential inflammation-specific targets. (C) Fifty-eight overlapping target proteins between inflammation-related proteins and targets of G-Rh4. (D) Protein–protein networks of overlapping 58 target proteins. Edges: Interactions between protein(s) and protein(s).
The genes related to inflammation were selected from the Therapeutic Target Database (TTD, http://db.idrblab.net/ttd/) (Wang Y. et al., 2019), the GeneCards (https://www.genecards.org/) (Rappaport et al., 2017), DisGeNET (https://www.disgenet.org/search) (Piñero et al., 2019), and the Comparative Toxicogenomics Database (CTD, http://ctdbase.org/search/) (Davis et al., 2021), where the database was searched using “inflammation” and “inflammatory” as the keywords.
The target genes of G-Rh4 and the target information of inflammation were uploaded to Venny 2.1 (https://bioinfogp.cnb.csic.es/tools/venny/index.html) to obtain the target information of G-Rh4 intersecting with inflammation, and it is considered as the potential target of Rh4 in the treatment of inflammation.
The protein–protein interaction (PPI) network diagram was obtained through the STRING database (https://string-db.org/) (Szklarczyk et al., 2021). The TSV-format file was downloaded from the STRING database and imported into Cytoscape 3.8.0 for core target screening.
The candidate targets were identified by using the DAVID v6.8 (https://david.ncifcrf.gov/) (Huang et al., 2009) to conduct GO and Kyoto Encyclopedia of Genes and Genomes (KEGG) enrichment analysis on the target of G-Rh4 in the treatment of inflammation. A threshold of p < 0.05 was used to identify key GO and KEGG pathways; GraphPad Prism 8 0.1 software was used to visualize the analysis results.
Using the software Cytoscape 3 8.0, the relationship network diagram of core drug targets was constructed and topological analysis was performed, after which the importance of targets was analyzed according to the degree value (Rahimmanesh and Fatehi, 2020).
First, PDB format of the key target proteins and MOL2 format of Ginsenoside Rh4 were obtained through the RCSR protein database (https://www.rcsb.org/) and the NCBI PubChem Compound database, and PyMOL software was employed to remove the water and ligands. After that, molecular docking was conducted between the treated proteins and active components by using AutoDock Vina software, and the binding energy was evaluated, where the value of <0 indicated that the receptors and compounds can bind by themselves (Liang et al., 2021), while the value of <–5.0 kcal/mol meant that they had good binding activities. The lower the binding energy, the greater the probability of binding and the more credible the result.
RAW264.7 cells were purchased from the Chinese Academy of Sciences Stem Cell Bank. DMEM containing 10% FBS and double antibodies (penicillin 100 U/mL and streptomycin 100 μg/ml) was used as a culture medium, and cells were cultured in an incubator at 37°C and 5% CO2.
The logarithmic period RAW264.7 cells (1 × 104 cells/well) were inoculated on 96-well plates and cultured for 24 h. After treatment with different concentrations of G-Rh4 in serum-free DMEM for 24 h, 20 μL of MTT (5 mg/ml; Sigma, USA) solution was added. After incubation for another 4 h, the culture medium was discarded and 150 μL DMSO (Sigma, USA) was added to each well. The absorbance at 550 nm was measured using a TECAN microplate reader (Maennedorf, Switzerland).
RAW264.7cells (2 × 105 cells/mL) were inoculated in 100-mm dishes and cultured for 24 h. LPS (1 μg/ml) was pretreated for 4 h and further treated with different concentrations (5, 7.5, and 10 μg/ml) of G-Rh4 for another 16 h. At the end of treatments, the total RNA of RAW264.7 cells was isolated with TRIzol (Invitrogen, Grand Island, NY, USA), and 5 µg total RNA was proceeded for cDNA synthesis with a High Capacity cDNA Reverse Transcription Kit (4368814, Applied Biosystems, Foster City, CA, USA). Real-time quantitative experiments were conducted according to the instructions of the 7,500 Real-time PCR system (Applied Biosystems, Foster City, CA, USA) to determine the mRNA expression. The primer sequences are shown in Supplementary Table S1.
RAW264.7 cells (2.5 × 105 cells/mL) were inoculated in 12-well plates and cultured for 24 h. LPS (1 μg/ml) was pretreated for 4 h and different concentrations of G-Rh4 were treated for 16 h. The levels of IL-6, TNF-α, IL-1β, and PGE2 in the supernatant were measured, respectively, according to the operation instructions of the ELISA kit (CLOUD-CLONE CORP., Wuhan, Hubei).
The logarithmic period RAW264.7 cells (1 × 104 cells/well) were inoculated on 96-well plates and cultured for 24 h. The cells were pretreated with LPS (1 μg/ml) for 4 h and then treated with different concentrations of G-Rh4 for 16 h. Nitrite levels in the cell culture supernatants were measured using Griess assay. Subsequently, 50 μL of the culture medium were mixed with 50 μL reagents of Griess A and Griess B, followed by incubation for 10 min at room temperature (light protected). A wavelength of 540 nm was selected to detect the absorbance values using a microplate reader, and nitrite levels were measured using a standard curve prepared from sodium nitrite (Park et al., 2005).
RAW264.7cells (7×105 cells/well) were seeded in 6-well plates and cultured for 24 h. The cells were pretreated with LPS (1 μg/ml) for 4 h and treated with different concentrations of G-Rh4 for 16 h. At the end of treatments, the cells were collected in a 1.5-ml centrifuge tube and then centrifuged at 10,000 r/min (rpm) for 5 min at 4°C, and the supernatant was discarded. The supernatant was washed with PBS and centrifuged at 10,000 rpm for 5 min, after which the cells were lysed with the RIPA cell lysate supplemented with 1% PMSF for 50 min and centrifuged at 12,000 rpm for 15 min to collect the supernatant. The protein concentrations were detected using BSA (bovine serum albumin, Sigma) as a standard. Equal amounts of protein (30 µg) were taken and subjected to SDS-PAGE electrophoresis. Then, the protein was transferred to the PVDF membrane and sealed with 5% (w/v) skim milk in Tris-buffered saline containing 0.1% Tween 20 (TBST). The primary antibody was incubated overnight at 4°C. After washing with TBST buffer, the membranes were incubated with the secondary antibody at room temperature for 1 h. The protein bands were quantitatively analyzed by ECL chromogenic exposure in a dark chamber.
The experimental data were obtained from independent triple-replicated experiments and were expressed as the mean ± SD. Statistical analyses were processed using GraphPad Prism 8.0, and the Student’s t-test statistical analysis method was used to compare groups. p < 0.05 showed that the difference was statistically significant.
In this experiment, network pharmacology and an experimental approach were employed to elucidate the drug target and pathways of G-Rh4 against inflammation (Figure 1). A total of 182 potential targets of ginsenoside Rh4 (G-Rh4) were collected from the databases such as Swiss Target Prediction, TargetNet, SEA, and ETCM, and 674 inflammation-related targets were screened through databases including GeneCards, DisGeNET, Therapeutic Target Database (TTD), and Comparative Toxicogenomics Database (CTD) (Figure 1B). The potential targets of G-Rh4 were intersected with inflammation-related genes, and as a result, 58 intersected genes were obtained. The Venn diagram is drawn in Figure 1C, and the names of intersection genes are listed in Supplementary Table S2.
A total of 58 intersected target genes of G-Rh4 against inflammation were uploaded to the String database. Organization was set as Homo sapiens, and obtained the key targets (Figure 1D). The PPI data were imported into Cytoscape software, and the target node degree is taken as an important parameter of topology analysis to screen out the key nodes in the network, which are shown in Figure 2, where the size of the ellipse represents the major degree of the target genes.
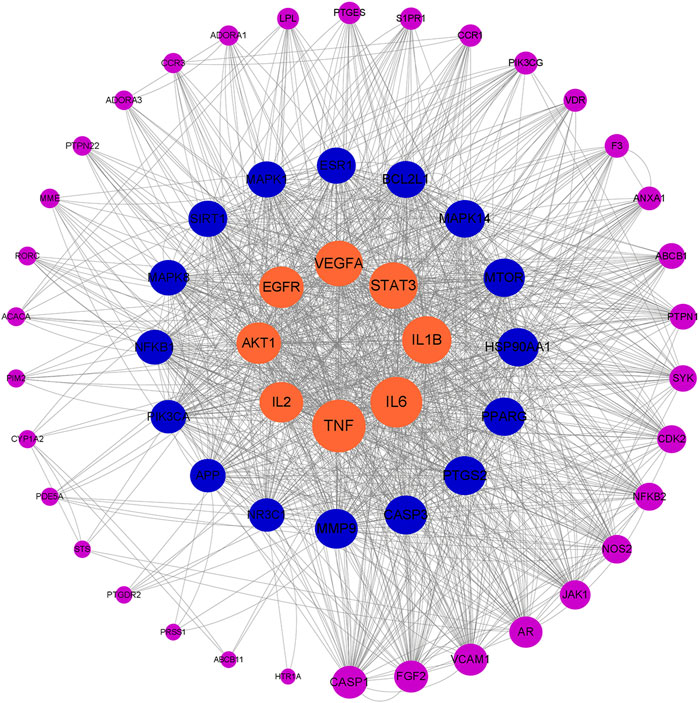
FIGURE 2. Network topology analysis of G-Rh4 treating inflammation; the target genes are sorted according to degree.
To further understand the biological processes involved in the screened key candidate targets above and their correlation with “inflammation,” we conducted analyses of GO and KEGG signaling pathways. The DAVID database was used to classify and count the GO function of 58 intersected genes of G-Rh4 in the treatment of inflammation (Supplementary Table S3). The pathways were visually analyzed to understand the functional distribution characteristics of different genes. The histogram for the pathway enrichment analysis was shown in Figure 3A. The GO analysis results indicated that the biological process (BP) terms were mainly associated with the positive regulation of the nitric oxide biosynthetic process, inflammatory response, positive regulation of interleukin-6 production, the lipopolysaccharide-mediated signaling pathway, and negative regulation of cell proliferation; the main cellular component (CC) terms were the macromolecular complex, cytoplasm, cell surface, plasma membrane, and endosome. Their primary molecular function (MF) was concentrated on enzyme binding, RNA polymerase II transcription factor activity, identical protein binding, nitric-oxide synthase regulator activity, and protein serine/threonine kinase activity.
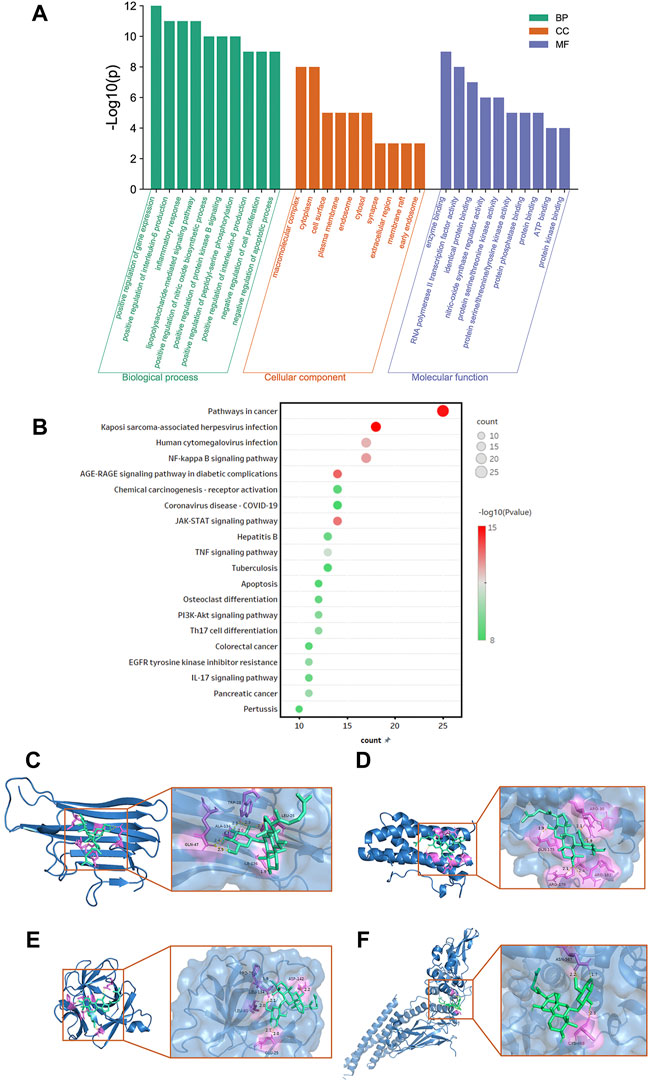
FIGURE 3. Functional enrichment analysis and docking simulation. Gene Ontology(A) and KEGG pathway enrichment (B) analysis of the target genes of G-Rh4 against inflammation. (C–F) The docking mode of G-Rh4 binding with target proteins; (C) TNF-α, (D) IL-6, (E) IL-1β, and (F) STAT3.
In addition, to clarify the underlying involved pathway of inflammation targets treated by G-Rh4, the pathway enrichment analysis on target genes was conducted through the KEGG public database (Supplementary Table S4). A total of 58 intersected genes were imported into the DAVID database for KEGG pathway annotation analysis, after which the top 20 pathways were visually analyzed to obtain the pathway enrichment analysis bubble diagram (Figure 3B). It could be seen that common targets were mainly concentrated in the JAK-STAT signaling pathway, TNF signaling pathway, NF-κB signaling pathway, and PI3K-Akt signaling pathway.
According to hub gene analysis, TNF-α, IL-6, IL-1β, and STAT3 are the key targets of G-Rh4 in the treatment of inflammation. Molecular docking simulation was applied to predict the binding ability between G-Rh4 and hub targets. The molecular docking of G-Rh4 with key targets was performed using AutoDock Vina software, and the binding energy was evaluated. The binding energy between G-Rh4 and TNF-α and IL-6 was –6.79 and –7.25 kcal/mol, respectively. The binding energy between G-Rh4 and IL-1β and STAT3 was –7.99 and –5.15 kcal/mol, respectively, suggesting that they have good binding activities (Table 1). The docking results were visualized using PyMOL software, as shown in Figures 3C–F.
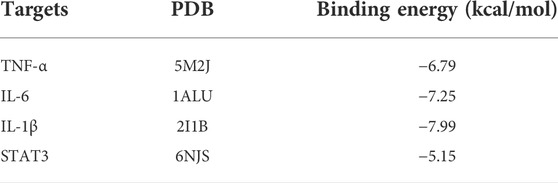
TABLE 1. Result of molecular docking.
To study the effect of G-Rh4 on the viability of RAW264.7 macrophages, cells were treated with different concentrations of G-Rh4 for 24 h, and the cell activity was detected by MTT assay. As shown in Figure 4A, compared with the control group, G-Rh4 treatment showed no significant effect on the viability of macrophages at the concentration of 0.5–8 μg/ml (p > 0.05), indicating that G-Rh4 has no toxicity to RAW264.7 cells at this concentration.
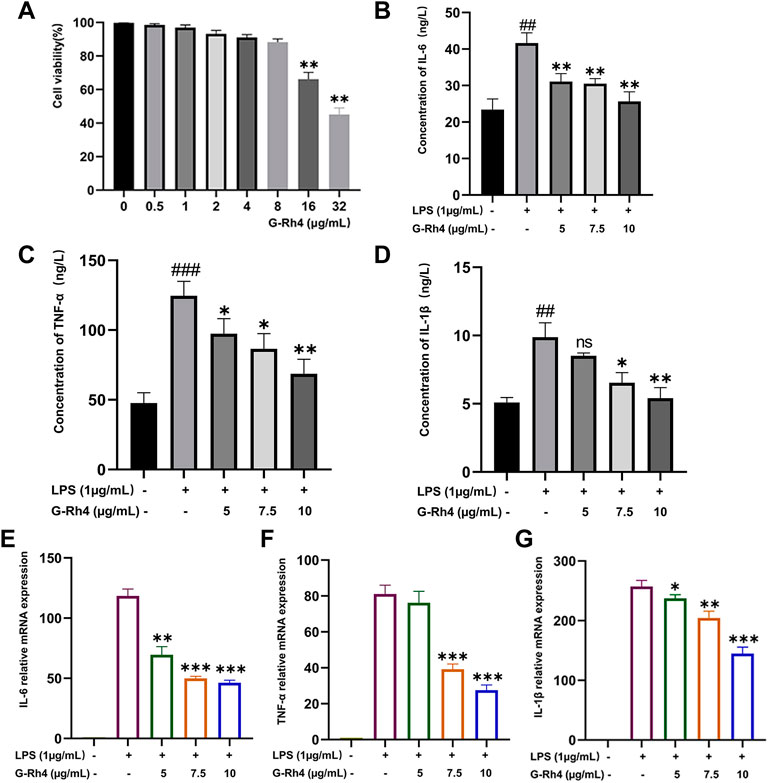
FIGURE 4. Effect of G-Rh4 on RAW264.7 cells. (A) Effect of G-Rh4 on cell viability. Cells were treated with different concentrations of G-Rh4 for 24 h. (B–D) Production of IL-6, TNF-α, and IL-1β in LPS-stimulated RAW264.7 cells was determined by ELISAs. (E–G) Effect of G-Rh4 on TNF-α, IL-6, and IL-1β mRNA levels in RAW264.7 cells. Data were presented as the mean ± SD values performed in triplicate (##p < 0.01 and ###p < 0.001 vs. control group, *p < 0.05, **p < 0.01, ***p < 0.001 vs. LPS group).
LPS is a component of the cell wall of gram-negative bacteria, which can stimulate the activation of immune cells such as macrophages and cause a systemic inflammatory response. To study the effect of G-Rh4 on major inflammatory factors, the protein and mRNA levels of IL-6, TNF-α, and IL-1β in LPS-stimulated RAW264.7 cells were evaluated by ELISAs and RT-PCR. The results are shown in Figures 4B–D. The contents of TNF–α, IL-6, and IL-1β in LPS-stimulated RAW264.7 were significantly higher than those in the normal group (p < 0.01, p < 0.001), but after G-Rh4 treatment, they decreased significantly in a concentration-dependent manner (p < 0.05, p < 0.01). Also, the increased mRNA expression of the main target factor after stimulation of LPS decreased significantly with the addition of G-Rh4 (Figures 4E–G), which seems to support the experimental results of ELISAs.
The expression levels of iNOS and COX-2 in RAW264.7 cells were detected by Western blot and RT-PCR analysis. Compared with the LPS treatment group, a 5 μg/ml dose of G-Rh4 had no significant difference in inhibiting the expression of COX-2. But it decreased significantly at a dose of 7.5 μg/ml and iNOS levels were markedly reduced from those at the 5 μg/ml dose of G-Rh4 (Figure 5A). The mRNA expressions of iNOS and COX-2 were also decreased significantly at the 7.5 μg/ml dose of G-Rh4 compared with those in the LPS treatment group (p < 0.001) (Figure 5B). These results showed that G-Rh4 meaningfully inhibited the expression of inflammatory promoting enzymes in the inflammatory response. iNOS and COX-2 are the key enzymes involved in the synthesis of NO and PGE2, which are also the key factors causing inflammatory responses and individual pathological status. We investigated whether G-Rh4 regulates NO and PGE2 synthesis in LPS-stimulated RAW264.7 cells. LPS stimulation increased the production of NO and PGE2, and the release of NO and PGE2 was significantly reduced in a dose-dependent manner upon G-Rh4 treatment (Figures 5C,D).
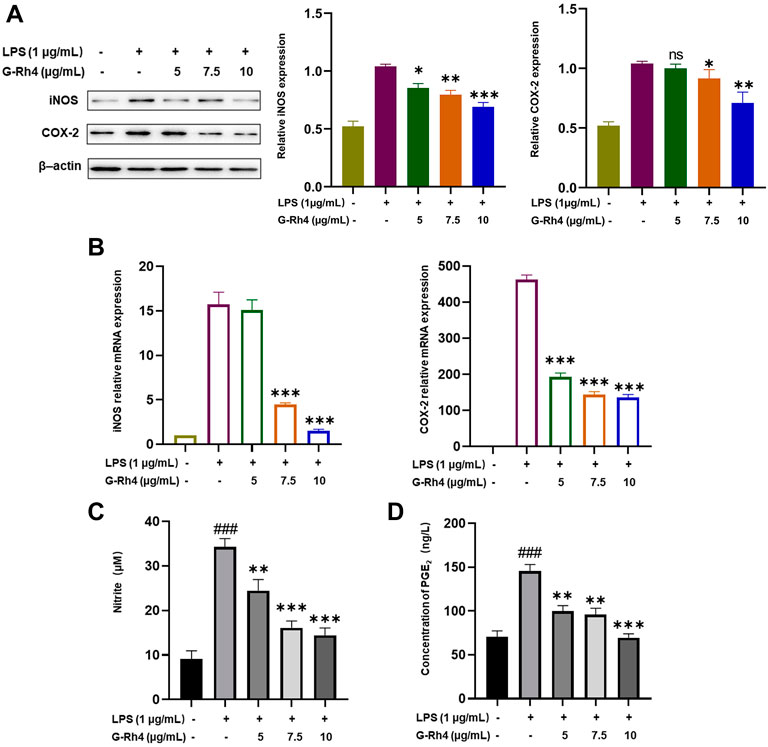
FIGURE 5. Inhibitory effects of G-Rh4 on NO/PGE2 production and iNOS/COX-2 expression in LPS-stimulated RAW264.7 cells. (A) Effect of G-Rh4 on iNOS and COX-2 protein levels in RAW264.7 cells. (B) Determination of iNOS and COX-2 mRNA levels in LPS-stimulated RAW264.7 cells. (C) NO levels in the culture medium evaluated by Griess reagent. (D) Effect of G-Rh4 on the release of PGE2 in RAW264.7 cells. Data were presented as the mean ± SD values performed in triplicate (##p < 0.01 and ###p < 0.001 vs. control group, *p < 0.05, **p < 0.01, ***p < 0.001 vs. LPS group).
NF-κB and STAT3 are transcriptional regulators involved in a series of critical signaling pathways, which plays a key role in the inflammatory response. Nuclear translocation of the NF-κB p50-p65 heterodimer is essential for NF-κB signaling. To further reveal the action mechanisms of G-Rh4, we investigated the effect of G-Rh4 on the expression and activation of p50-p65 and IκBα by Western blot analysis (Figure 6A). The results showed that G-Rh4 effectively inhibited LPS-induced phosphorylation of p50, p65, and IκBα in RAW264.7 cells. According to our network pharmacological analysis, STAT3 is also the main hub gene of G-Rh4 against inflammation, like the main inflammatory factors previously investigated. The Western blot analysis showed that the levels of p-JAK1/JAK1 and p-STAT3/STAT3 were significantly downregulated with the treatment of G-Rh4 (Figure 6B). With the increase of G-Rh4 concentration (5, 7.5, and 10 μg/ml), the phosphorylation levels of JAK1 and STAT3 protein decreased evidently (p < 0.01, p < 0.001). These results suggest that the anti-inflammatory effect of G-Rh4 can be mediated by the regulation of the NF-kB and STAT3 pathway.
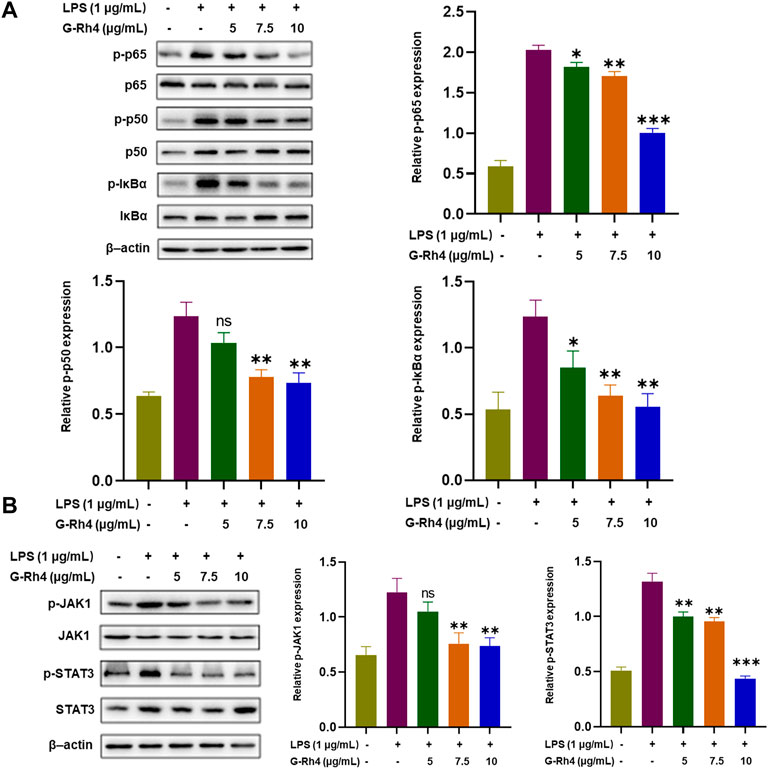
FIGURE 6. Effect of G-Rh4 on the NF-κB and STAT-3 activation in RAW264.7 cells. (A) Determination of the p-IκBα (Ser32), p-p65 (ser536), and p-p50 (ser337) protein using Western blot analysis. (B) Effect of G-Rh4 on the STAT-3 activation in RAW264.7 cells. Data were presented as the mean ± SD values performed in triplicate (*p < 0.05, **p < 0.01, ***p < 0.001 vs. LPS group).
Inflammation is a specific defense response to physical injury or infection. The persistence of inflammation, however, leads to the occurrence and development of several refractory diseases, such as cardiovascular disease, cancer, diabetes mellitus, chronic kidney disease, non-alcoholic fatty liver disease, and autoimmune and neurodegenerative disorders. Therefore, the extensive study of the inflammation pathways and the development of effective inflammation-modulating drugs are the most challenging area in the field of life science and pharmacology. At present, traditional Chinese medicine has become an emerging candidate for the treatment of inflammation because of its potent efficacy and fewer side effects (Fan et al., 2021).
Network pharmacology, from the perspective of system-level and biological networks, analyzes the molecular relationship between drugs and targets, reveals the systematic pharmacological mechanism of drugs, and guides the development of new drugs and clinical diagnosis and treatment. It is reported that the existing differences in data collection and target prediction in network pharmacology analysis result in the loss of some target genes (Luo et al., 2021). Since some target genes may not be included in the public database, we explored multiple databases for G-Rh4 and inflammation-related target genes. A total of 58 anti-inflammatory common targets of G-Rh4 were collected to analyze these common targets by hub gene analysis and GO and KEGG enrichment analysis. Our results showed that the anti-inflammatory effect of G-Rh4 was deeply related to the main target factors including TNF-α, IL-6, IL-1β, and STAT-3 and their related metabolic pathways, and the molecular docking analysis confirmed that G-Rh4 had a high binding possibility with these target factors. VEGFA is also the main target factor related to the anti-inflammatory effect in the network pharmacological analysis, but the possibility of binding with G-Rh4 was not recognized in molecular docking simulation (the experimental result has not shown).
It is reported that a prolonged inflammatory state is harmful to health, and long-term macrophages in tissues activate the abnormal expression of inflammatory factors such as TNF-α, IL-6, IL-1β, NO, and Prostaglandin E2, which can stimulate the inflammatory signaling pathway, eventually leading to chronic low-grade inflammation (Huang et al., 2017). These mediators stimulate the innate immune response, but their overexpression may cause endotoxemia, leading to tissue injury, organ failure, shock, and even death. Regulating the expression of these inflammatory mediators plays a key role in the treatment of inflammatory diseases (Joh et al., 2011). We confirmed the inhibitory effect of G-Rh4 on the expression of the primary target factors through LPS-stimulated mouse RAW264.7 cells. The in vitro inflammatory response model established by LPS-stimulated RAW264.7 macrophages is a widely used cell model for screening anti-inflammatory drugs. LPS mainly induces the synthesis and release of a variety of inflammatory mediators through the activation of nuclear transcription factors κB (NF-κB), JAK/STAT, MAPKs, phosphorylated ERK, phosphorylated JNK, and phosphorylated p38 signaling pathways (Liu et al., 2019). Our results indicated that LPS significantly increased mRNA and protein levels of TNF-α, IL-6, and IL-1β. In addition, we also confirmed using ELISAs and RT-PCR methods that G-Rh4 brings about the anti-inflammatory effects by decreasing TNF-α, IL-6, and IL-1β expression levels in LPS-activated RAW264.7 cells.
As reported previously, NO is an important inflammatory signal molecule in the pathogenesis of inflammation, which reacts with superoxide free radicals to produce peroxynitrite ions, resulting in various inflammatory states (Theofilis et al., 2021). Inducible nitric oxide synthase (iNOS) is a regulator of NO synthesis, and it plays a regulatory role in the production of proinflammatory mediators. In addition, inflammation is directly related to arachidonic acid metabolism (Wang T. et al., 2019), under the action of COX-2, and arachidonic acid is transformed into prostaglandin E2(PGE 2) through several enzymatic reactions. PGE2 can not only induce the inflammatory cells to release the chemokines and recruit the inflammatory cell movement but also cooperate with lipopolysaccharide to induce the expression of IL-6 and IL-1 in macrophages (Oshima et al., 2011). It can also cooperate with IL-12 to promote the differentiation of naive T cells into helper T cell 1 (Yao et al., 2009). Since COX-2 can quickly respond to a series of proinflammatory mediators and cytokines, it has been considered for a long time to play an important role in the pathological process of inflammation. Moreover, GO analysis of network pharmacology revealed that the target genes of G-Rh4 against inflammation were involved in the “nitric oxide synthase regulator activity” in both biological process (BP) analysis and Molecular Function (MF) analysis. The results in LPS-activated RAW264.7 cells showed that G-Rh4 significantly reduced the release of NO and PGE2 as well as the expression of elevated iNOS and COX-2, indicating that network pharmacology might be an effective approach to identify the key target and action pathway of G-Rh4 against inflammation.
In addition, KEGG pathway enrichment analysis pointed out that the intersection target genes of G-Rh4 and inflammation were closely related to the NF-κB and JAK-STAT signaling pathways, the including TNF signaling pathway. Nuclear factor-kappa B(NF-κB) is a protein complex responsible for DNA transcription, cytokine secretion, and cell survival (Wu et al., 2016), and its abnormal signaling pathway is associated with some chronic inflammatory diseases such as inflammatory bowel disease, sepsis, arthritis, and atherosclerosis (Theofilis et al., 2021). NF-κB (p50-p65) is constitutively present in the cytoplasm of RAW264.7cells and is sequestered by inhibitory protein IκBα. In the inflammatory state, the activation phosphorylation of p65 (ser536) and p50 (ser337) leads to the activation of NF-κB and the expression of a series of downstream inflammatory factors, such as IL-1β and IL-6. STAT3 (signal transducer and activator of transcription 3) is a key regulator of the inflammatory response induced by lipopolysaccharide (LPS) (Li et al., 2021), and the JAK-STAT (Janus kinase-signal transducer and activator of transcription) pathway is one of the important inflammatory signal transduction pathways, which can mediate the immune response (Zhao et al., 2021). Taken together, this study reveals that G-Rh4 might exert its anti-inflammatory effects through both NF-κB and STAT3 pathways.
Our study clarified, for the first time, the targets and working pathways of G-Rh4 in the inflammation process through network pharmacological analysis and experimental approaches and puts forward that G-Rh4 can directly bind with key pro-inflammatory cytokines including TNF-α, IL-6, and IL-1β to execute its anti-inflammatory activity. The further study is necessary to evaluate the effectiveness of G-Rh4 and explore the extensive working mechanism of in vivo inflammation models. This study provides a scientific basis for further pharmacological research on ginsenoside Rh4, as well as the development of novel anti-inflammatory drugs with high efficacy and few side effects and clinical application.
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found in the article/Supplementary Material.
Study design: Y-HJ and K-IT; network pharmacology analysis: Z-XZ and K-IT; RAW264.7 cell culture: G-AL and Y-MS; molecular docking: Y-NW; drafting the manuscript: K-IT; revision of the manuscript: YL and K-IT; supervision: Y-HJ; project administration: Y-HJ; funding acquisition: Y-HJ; All authors have read and agreed to the published version of the manuscript.
This research was supported by the Specific Funding of Development and Reform Commission of Jilin Province (2021FGWCXNLJSSZ01) and Science and Technology Development Program of Jilin Province (YDZJ202101ZYTS087).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors, and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fphar.2022.953871/full#supplementary-material
Arai, Y., Martin-Ruiz, C. M., Takayama, M., Abe, Y., Takebayashi, T., Koyasu, S., et al. (2015). Inflammation, but not telomere length, predicts successful ageing at extreme old age: A longitudinal study of semi-supercentenarians. EBioMedicine 2, 1549–1558. doi:10.1016/j.ebiom.2015.07.029
Davis, A. P., Grondin, C. J., Johnson, R. J., Sciaky, D., Wiegers, J., Wiegers, T. C., et al. (2021). Comparative Toxicogenomics database (CTD): Update 2021. Nucleic Acids Res. 49, D1138–D1143. doi:10.1093/nar/gkaa891
Fan, Y., Wang, Y., Yu, S., Chang, J., Yan, Y., Wang, Y., et al. (2021). Natural products provide a new perspective for anti-complement treatment of severe COVID-19: A review. Chin. Med. 16, 67. doi:10.1186/s13020-021-00478-3
Furman, D., Campisi, J., Verdin, E., Carrera-Bastos, P., Targ, S., Franceschi, C., et al. (2019). Chronic inflammation in the etiology of disease across the life span. Nat. Med. 25, 1822–1832. doi:10.1038/s41591-019-0675-0
GBD 2017 Causes of Death Collaborators (2018). Global, regional, and national age-sex-specific mortality for 282 causes of death in 195 countries and territories, 1980-2017: A systematic analysis for the global burden of disease study 2017. Lancet 392, 1736–1788. doi:10.1016/S0140-6736(18)32203-7
Huang, D. W., Sherman, B. T., and Lempicki, R. A. (2009). Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat. Protoc. 4, 44–57. doi:10.1038/nprot.2008.211
Huang, Q., Wang, T., and Wang, H. (2017). Ginsenoside Rb2 enhances the anti-inflammatory effect of ω-3 fatty acid in LPS-stimulated RAW264.7 macrophages by upregulating GPR120 expression. Acta Pharmacol. Sin. 38, 192–200. doi:10.1038/aps.2016.135
Joh, E.-H., Lee, I.-A., Jung, I.-H., and Kim, D.-H. (2011). Ginsenoside Rb1 and its metabolite compound K inhibit IRAK-1 activation—the key step of inflammation. Biochem. Pharmacol. 82, 278–286. doi:10.1016/j.bcp.2011.05.003
Li, S., Hu, K., Li, L., Shen, Y., Huang, J., Tang, L., et al. (2021). Stattic alleviates acute hepatic damage induced by LPS/ d -galactosamine in mice. Innate Immun. 27, 201–209. doi:10.1177/1753425920988330
Liang, Y., Liang, B., Chen, W., Wu, X.-R., Liu-Huo, W.-S., Zhao, L.-Z., et al. (2021). Potential mechanism of dingji fumai decoction against atrial fibrillation based on network pharmacology, molecular docking, and experimental verification integration strategy. Front. Cardiovasc. Med. 8, 712398. doi:10.3389/fcvm.2021.712398
Libby, P. (2021). The changing landscape of atherosclerosis. Nature 592, 524–533. doi:10.1038/s41586-021-03392-8
Liu, M., Fang, G., Yin, S., Zhao, X., Zhang, C., Li, J., et al. (2019). Caffeic acid prevented LPS-induced injury of primary bovine mammary epithelial cells through inhibiting NF- κ B and MAPK activation. Mediat. Inflamm. 2019, 1897820. doi:10.1155/2019/1897820
Luo, Z., Liu, W., Sun, P., Wang, F., and Feng, X. (2021). Pan-cancer analyses reveal regulation and clinical outcome association of the shelterin complex in cancer. Brief. Bioinform. 22, bbaa441. doi:10.1093/bib/bbaa441
Morrisette-Thomas, V., Cohen, A. A., Fülöp, T., Riesco, É., Legault, V., Li, Q., et al. (2014). Inflamm-aging does not simply reflect increases in pro-inflammatory markers. Mech. Ageing Dev. 139, 49–57. doi:10.1016/j.mad.2014.06.005
Oshima, H., Hioki, K., Popivanova, B. K., Oguma, K., Van Rooijen, N., Ishikawa, T., et al. (2011). Prostaglandin E2 signaling and bacterial infection recruit tumor-promoting macrophages to mouse gastric tumors. Gastroenterology 140, 596–607. e7. doi:10.1053/j.gastro.2010.11.007
Panigrahy, D., Gilligan, M. M., Huang, S., Gartung, A., Cortés-Puch, I., Sime, P. J., et al. (2020). Inflammation resolution: A dual-pronged approach to averting cytokine storms in COVID-19? Cancer Metastasis Rev. 39, 337–340. doi:10.1007/s10555-020-09889-4
Park, E.-K., Shin, Y.-W., Lee, H.-U., Kim, S.-S., Lee, Y.-C., Lee, B.-Y., et al. (2005). Inhibitory effect of ginsenoside Rb1 and compound K on NO and prostaglandin E2 biosyntheses of RAW264.7 cells induced by lipopolysaccharide. Biol. Pharm. Bull. 28, 652–656. doi:10.1248/bpb.28.652
Piñero, J., Ramírez-Anguita, J. M., Saüch-Pitarch, J., Ronzano, F., Centeno, E., Sanz, F., et al. (2019). The DisGeNET knowledge platform for disease genomics: 2019 update. Nucleic Acids Res. 48, D845–D855. gkz1021. doi:10.1093/nar/gkz1021
Rahimmanesh, I., and Fatehi, R. (2020). Systems biology approaches toward autosomal dominant polycystic kidney disease (ADPKD). Clin. Transl. Med. 9, 1. doi:10.1186/s40169-019-0254-5
Rappaport, N., Twik, M., Plaschkes, I., Nudel, R., Iny Stein, T., Levitt, J., et al. (2017). MalaCards: An amalgamated human disease compendium with diverse clinical and genetic annotation and structured search. Nucleic Acids Res. 45, D877–D887. doi:10.1093/nar/gkw1012
Schmeltzer, P. A., Kosinski, A. S., Kleiner, D. E., Hoofnagle, J. H., Stolz, A., Fontana, R. J., et al. (2016). Liver injury from nonsteroidal anti-inflammatory drugs in the United States. Liver Int. 36, 603–609. doi:10.1111/liv.13032
Sun, J. H., Sun, F., Yan, B., Li, J. Y., and Xin, D. L. (2020). Data mining and systematic pharmacology to reveal the mechanisms of traditional Chinese medicine in Mycoplasma pneumoniae pneumonia treatment. Biomed. Pharmacother. 125, 109900. doi:10.1016/j.biopha.2020.109900
Szklarczyk, D., Gable, A. L., Nastou, K. C., Lyon, D., Kirsch, R., Pyysalo, S., et al. (2021). The STRING database in 2021: Customizable protein–protein networks, and functional characterization of user-uploaded gene/measurement sets. Nucleic Acids Res. 49, D605–D612. doi:10.1093/nar/gkaa1074
Theofilis, P., Sagris, M., Oikonomou, E., Antonopoulos, A. S., Siasos, G., Tsioufis, C., et al. (2021). Inflammatory mechanisms contributing to endothelial dysfunction. Biomedicines 9, 781. doi:10.3390/biomedicines9070781
Wang, C., Pang, X., Zhu, T., Ma, S., Liang, Y., Zhang, Y., et al. (2022). Rapid discovery of potential ADR compounds from injection of total saponins from Panax notoginseng using data-independent acquisition untargeted metabolomics. Anal. Bioanal. Chem. 414, 1081–1093. doi:10.1007/s00216-021-03734-5
Wang T, T., Fu, X., Chen, Q., Patra, J. K., Wang, D., Wang, Z., et al. (2019). Arachidonic acid metabolism and kidney inflammation. Int. J. Mol. Sci. 20, 3683. doi:10.3390/ijms20153683
Wang, Y., Yang, H., Chen, L., Jafari, M., and Tang, J. (2021). Network-based modeling of herb combinations in traditional Chinese medicine. Brief. Bioinform. 22, bbab106. doi:10.1093/bib/bbab106
Wang Y, Y., Zhang, S., Li, F., Zhou, Y., Zhang, Y., Wang, Z., et al. (2019). Therapeutic target database 2020: Enriched resource for facilitating research and early development of targeted therapeutics. Nucleic Acids Res. 48, D1031–D1041. gkz981. doi:10.1093/nar/gkz981
Wu, J., Yang, H., Zhao, Q., Zhang, X., and Lou, Y. (2016). Ginsenoside Rg1 exerts a protective effect against Aβ25–35-induced toxicity in primary cultured rat cortical neurons through the NF-κB/NO pathway. Int. J. Mol. Med. 37, 781–788. doi:10.3892/ijmm.2016.2485
Yao, C., Sakata, D., Esaki, Y., Li, Y., Matsuoka, T., Kuroiwa, K., et al. (2009). Prostaglandin E2–EP4 signaling promotes immune inflammation through TH1 cell differentiation and TH17 cell expansion. Nat. Med. 15, 633–640. doi:10.1038/nm.1968
Keywords: network pharmacology, ginsenoside, inflammation, proinflamamtory cytokines, pharmacological mechanism
Citation: To K-I, Zhu Z-X, Wang Y-N, Li G-A, Sun Y-M, Li Y and Jin Y-H (2022) Integrative network pharmacology and experimental verification to reveal the anti-inflammatory mechanism of ginsenoside Rh4. Front. Pharmacol. 13:953871. doi: 10.3389/fphar.2022.953871
Received: 26 May 2022; Accepted: 14 July 2022;
Published: 31 August 2022.
Edited by:
Weicheng Hu, Huaiyin Normal University, ChinaReviewed by:
Di Wang, Jilin Agriculture University, ChinaCopyright © 2022 To, Zhu, Wang, Li, Sun, Li and Jin. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Ying-Hua Jin, eWhqaW5Aamx1LmVkdS5jbg==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.