 Elena Hernández-Hernández1,2†
Elena Hernández-Hernández1,2† M. Julia García-Fuster1,2*
M. Julia García-Fuster1,2*- 1IUNICS, University of the Balearic Islands, Palma, Spain
- 2Health Research Institute of Balearic Islands (IdISBa), Palma, Spain
Aging predisposes to late-life depression and since antidepressants are known to change their efficacy with age, novel treatment options are needed for our increased aged population. In this context, the goal of the present study was to evaluate the potential antidepressant-like effect of cannabidiol in aged rats. For this purpose, 19–21-month-old Sprague–Dawley rats were treated for 7 days with cannabidiol (dose range: 3–30 mg/kg) and scored under the stress of the forced-swim test. Hippocampal cannabinoid receptors and cell proliferation were evaluated as potential molecular markers underlying cannabidiol’s actions. The main results of the present study demonstrated that cannabidiol exerted a dose-dependent antidepressant-like effect in aged rats (U-shaped, effective at the intermediate dose of 10 mg/kg as compared to the other doses tested), without affecting body weight. None of the molecular markers analyzed in the hippocampus were altered by cannabidiol’s treatment. Overall, this study demonstrated a dose-dependent antidepressant-like response for cannabidiol at this age-window (aged rats up to 21 months old) and in line with other studies suggesting a beneficial role for this drug in age-related behavioral deficits.
Introduction
Aging is the strongest risk factor for most chronic disorders (e.g., Bektas et al., 2018), and in addition to changes in cognitive performance (Rosenzweig and Barnes, 2003; Mattson and Magnus, 2006), some other key features are particularly prone to decline, such as affective-like behavior, predisposing to the development of what is called late-life depression (McKinney and Sibille, 2013). In fact, the prevalence of depression in the population aged over 80 years has been shown to be around 15–20% (Stek et al., 2004), with the corresponding increased prescription of antidepressant drugs (Sonnenberg et al., 2008). Surprisingly, antidepressant-like treatments are known to change their efficacy with age [e.g., reviewed by Felice et al. (2015)]; however, not much research is focused on characterizing classical antidepressant-like responses and/or novel therapeutical options for this age group (see some representative recent preclinical studies: Fernández-Guasti et al., 2017; Mastrodonato et al., 2022), posing a health problem for our continuously increasing elderly population. In the clinic, the only approach taken with elderly patients includes lowering the doses to adjust for a slower metabolism. Remarkably, the described state of increased negative effect emerging with age can be modeled in naïve aging rodents (e.g., from middle-age and on; Hernández-Hernández et al., 2018 and references within; also Herrera-Pérez et al., 2008), providing a preclinical platform in which to test novel antidepressant drugs (e.g., Hernández-Hernández and García-Fuster, 2021).
In the context of characterizing novel treatment options, cannabidiol, a non-psychomimetic phytocannabinoid found in Cannabis sativa, has demonstrated a valuable role in ameliorating certain stress-related psychiatric symptoms in rodent models (e.g., Marco et al., 2011; Campos et al., 2016, 2017; Haney and Evins 2016; García-Gutiérrez et al., 2020; Gonzalez-Cuevas et al., 2021; Stanciu et al., 2021), with a great safety potential (Pisani et al., 2021). Interestingly, prior preclinical studies have reported that the antidepressant- and/or anxiolytic-like effects induced by cannabidiol [reviewed recently by García-Gutiérrez et al. (2020) and Melas et al. (2021)] are sex- (Silote et al., 2021; Ledesma-Corvi and García-Fuster, 2022; Martín-Sánchez et al., 2022), stress- (Shbiro et al., 2019; Bis-Humbert et al., 2021a; Ledesma-Corvi and García-Fuster, 2022), and/or age-related (e.g., different efficacy in adolescent vs. adult rats: Bis-Humbert et al., 2020), while studies reporting the potential beneficial effects in older populations are scarce (in addition to its anti-oxidant and anti-inflammatory potential; Dash et al., 2021 and references within). Against this background, the goal of this study was to evaluate whether cannabidiol could exert an antidepressant-like response as measured in the forced-swim test in aged rats.
Several studies have been centered on elucidating cannabidiol’s actions on improving stress-related alterations, involving multiple targets (reviewed by Silote et al., 2019; García-Gutiérrez et al., 2020), such as a multimodal pharmacologic profile over the endocannabinoid system (Bisogno et al., 2001; Thomas et al., 2007; Pertwee, 2008; Campos et al., 2012; Laprairie et al., 2015; Martínez-Pinilla et al., 2017; Tham et al., 2019), an agonistic potential over TRPV1 receptors (Bisogno et al., 2001), as well as the regulation of other neurotransmitter systems [i.e., serotoninergic, opioidergic, and dopaminergic; reviewed in Silote et al. (2019)] or neuroprotective targets (i.e., hippocampal neurogenesis; Marchalant et al., 2009; Fogaça et al., 2018; Luján et al., 2018, 2020). For that reason, and since hippocampal function is altered with aging (Rosenzweig and Barnes, 2003), the current study evaluated the potential regulation of CB1 and CB2 receptors, as well as that of an early stage of neurogenesis (i.e., cell proliferation) following cannabidiol’s treatment in the hippocampus of aged rats.
Methods
Animals
For this study, 39 male Sprague–Dawley rats (bred in the animal facility at the University of the Balearic Islands) were used when they were 19–21 months old (Figure 1A). The rats were housed under standard vivarium conditions (22°C, 70% humidity, and 12-h light/dark cycle, lights on at 8:00 a.m.) with ad libitum access to a standard diet and tap water. Following size requirements (animals housed per standard cage), the rats were individually housed for several months before testing started. All procedures were performed during the light period (between 8:00 h and 15:00 h), complied with ARRIVE guidelines (Percie du Sert et al., 2020), EU Directive 2010/63/EU for animal experiments, and Spanish Royal Decree 53/2013, and were approved by the Local Bioethical Committee and the Regional Government. All efforts were made to minimize the number of rats used and their suffering.
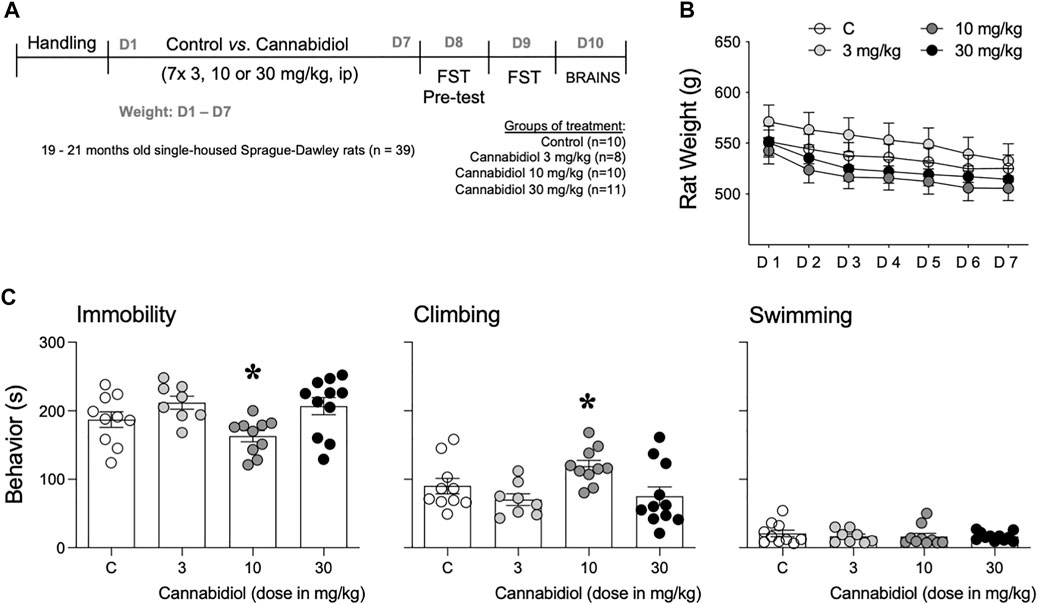
FIGURE 1. (A) Experimental timeline. (B) Monitorization of body weight (g) during the experimental treatment. Data represent mean ± SEM of the body weight (g). A two-way repeated-measures ANOVA did not detect a significant effect of treatment. (C) Antidepressant-like effect as measured in the forced-swim test. Data represent mean ± SEM of the time spent (s) immobile, climbing, or swimming. Individual values are shown for each rat (symbols). One-way ANOVAs detected significant changes for immobility and climbing. Multiple comparisons were performed with Tukey’s test: *p < 0.05 vs. the other doses tested, both 3 and 30 mg/kg of cannabidiol. Groups of treatment: control (C, n = 10) and cannabidiol (3 mg/kg, n = 8; 10 mg/kg, n = 10; and 30 mg/kg, n = 11).
Repeated Cannabidiol Treatment
All rats were handled for 5 days prior to drug treatment, during which they were separated into four groups and received either a daily dose of cannabidiol (3, 10, or 30 mg/kg, i.p.; n = 8, 10, and 11, respectively) (purity ≥98%; THC Pharm, Germany) or vehicle (1 ml/kg of DMSO, control group; n = 10) for 7 days. The doses of cannabidiol were selected based on a prior study from our research group showing age- and dose-dependent antidepressant-like responses (i.e., different dose-efficacy during adolescence vs. adulthood in male naïve rats; Bis-Humbert et al., 2020). Body weight was daily monitored through the treatment process (D1–D7).
Forced-Swim Test
To ascertain cannabidiol’s antidepressant-like response in aged rats we used the forced-swim test, since it is a standardized test for screening antidepressant-like responses under a stressful situation (Slattery and Cryan, 2012). For this purpose, and following prior well-established procedures in our group (Bis-Humbert et al., 2020; 2021b), rats were individually exposed to a 15 min pre-test on D8, and a 5-min test on D9 (Figure 1A) that was videotaped. The water tanks (41 cm high × 32 cm diameter) were filled with water up to 25 cm depth (25 ± 1°C) and were changed for each rat. Videos were coded and blindly analyzed using Behavioral Tracker Software (CA, United States) to calculate the time spent (s) immobile or active (climbing or swimming).
Hippocampal Neurochemical Correlation
All rats were sacrificed by rapid decapitation on D10 (Figure 1A), their brains were extracted, and both hemispheres were separated. On the one hand, the right hippocampus was freshly dissected and frozen in liquid nitrogen until the contents of CB receptors were evaluated in total homogenates (40 μg) by Western blot analysis with anti-CB1 (1:2000; Abcam, Cat. No. 23703, United Kingdom) or anti-CB2 (1:1000; Cayman Chemical, Cat. No. 101550, United States) primary antibodies, as previously detailed by our group (García-Cabrerizo and García-Fuster, 2016; Bis-Humbert et al., 2021a). Low quantities of total homogenates (15 μg) were loaded to detect β-actin (1:10000; Sigma-Aldrich, clone AC-15, United States), which was used as a negative loading control, since its content was not altered by any of the treatment conditions. Each sample was run at least three times in different gels, and percent changes were calculated for each rat as compared to control-treated samples loaded in the same gels. On the other hand, the left hemisphere was quickly frozen and stored until the entire hippocampus (−1.72 to −6.80 mm from Bregma) was cryostat-cut (30 μm sections) and slide mounted (8 sections/slide, 8 slides/series, and 3 series/animal from the most anterior to the posterior part of the hippocampus) to evaluate the rate of cell proliferation with the anti-Ki-67 antibody (1:40,000; kindly provided by Dr. Huda Akil and Dr. Stanley J. Watson, University of Michigan, United States) by immunohistochemistry as detailed earlier (García-Fuster et al., 2010; García-Cabrerizo et al., 2015; García-Fuster et al., 2017). The number of immunostained Ki-67 + cells was quantified using a Leica DMR light microscope (63× objective lens) in all sections by a blind experimenter to the treatment groups and as previously described in detail (e.g., García-Fuster et al., 2010, 2011, 2017).
Statistics
All data were analyzed with GraphPad Prism, Version 9.3.1 (GraphPad Software, United States). The results are expressed as mean values ± standard error of the mean (SEM), with individual symbols for each rat shown within bar graphs. The changes in body weight were analyzed with two-way repeated-measures ANOVA. Potential overall changes in behavior (s) in the forced-swim test (i.e., immobility, climbing, or swimming) or in the content of brain markers (i.e., CB receptors and Ki-67 + cells) were evaluated by one-way ANOVAs followed by Tukey’s multiple comparisons test. The level of significance was set at p ≤ 0.05.
Results
Cannabidiol Did Not Alter Normal Body Weight (g) in Aged Rats
Although no changes were observed by cannabidiol’s treatment (F3,35 = 1.18, p = 0.329), there was a significant effect of day (F6,210 = 68.15, p < 0.001), probably driven by the observed moderate decreases in body weight for all groups during the course of the experimental procedure, and most probably caused by the stress of cannabidiol’s treatment (Figure 1B).
Dose-Dependent Antidepressant-Like Effects of Cannabidiol in Aged Rats
When evaluating the antidepressant-like potential of cannabidiol (3, 10, and 30 mg/kg; i.p.) in the forced-swim test, although a one-way ANOVA detected a significant difference in the time-aged rats spent immobile (F3,35 = 4.18, p = 0.012; Figure 1C), this effect was mainly driven by the dose of 10 mg/kg, which significantly reduced immobility as compared to the other doses tested (−49 ± 16 s, ∗p = 0.021 vs. 3 mg/kg; −44 ± 15 s, ∗p = 0.026 vs. 30 mg/kg; U-shaped dose response; Figure 1C). Interestingly, the decrease observed in immobility paralleled an increase in the time rats spent climbing (F3,35 = 3.89, p = 0.017), with significant changes when comparing the dose of 10 mg/kg vs. 3 mg/kg (+49 ± 16 s, ∗p = 0.026) and 30 mg/kg (+44 ± 15 s, ∗p = 0.031; Figure 1C). Finally, no changes were observed in swimming behavior (F3,35 = 0.33, p = 0.803).
Cannabidiol Did Not Modulate CB1 and CB2 Receptors or the Rate of Early Cell Proliferation in the Hippocampus
When evaluating some of the potential molecular markers regulated by cannabidiol and that could parallel its behavioral actions, the results showed no changes in CB1 (F3,35 = 1.68, p = 0.189; Figure 2A) and CB2 (F3,35 = 0.11, p = 0.105; Figure 2B) receptors, nor in the rate of cell proliferation (Ki-67 + cells: F3,35 = 1.78, p = 0.168; Figure 2C) in the hippocampus of aged rats as measured 3 days post-treatment.
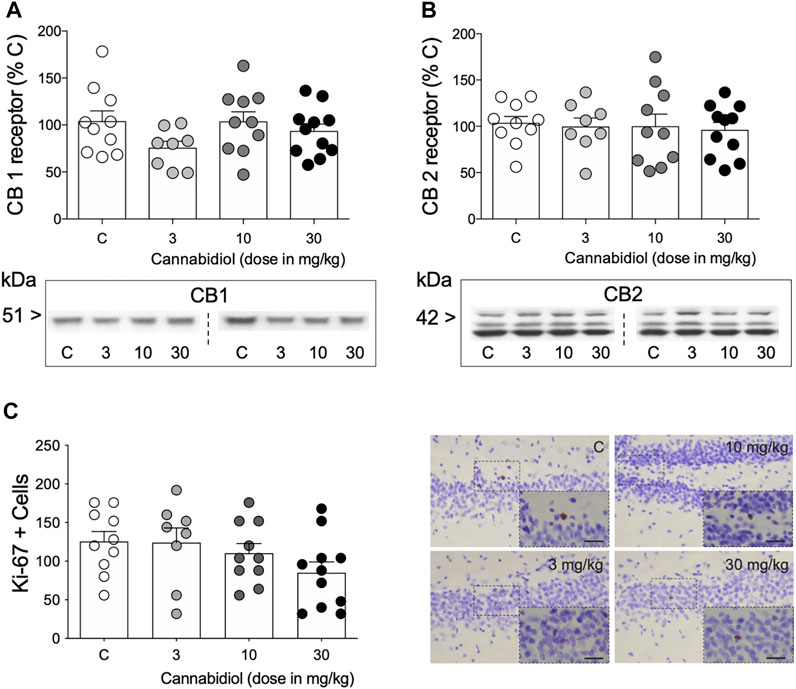
FIGURE 2. Modulation of hippocampal molecular markers by repeated cannabidiol treatment. (A) CB1 and (B) CB2 receptors as measured by Western blot analysis. Data represent mean ± SEM of CB1 and CB2 protein contents expressed as % change vs. control-treated rats. Individual values are shown for each rat (symbols). One-way ANOVAs did not detect any significant changes. Representative immunoblots are shown depicting CB1 and CB2 labeling. (C) Ki-67 + cells in the dentate gyrus as measured by immunohistochemistry analysis. Data represent mean ± SEM of Ki-67 + cells. Individual values are shown for each rat (symbols). A one-way ANOVA did not detect any significant changes. Representative images of Ki-67 + cells (brown labeling in the blue granular layer) were taken using a light microscope and quantified using a 63× objective lens. Scale bar: 30 μm.
Discussion
The main results of the present study demonstrated that cannabidiol is capable of exerting a dose-dependent antidepressant-like response in aged rats, without inducing changes in body weight. None of the molecular markers analyzed in the hippocampus (CB receptors and cell proliferation) were altered by cannabidiol treatment.
Cannabidiol induced a dose-dependent antidepressant-like effect in aged rats, as observed in the forced-swim test by a decrease in immobility paired with an increase in climbing. Interestingly, these effects were dose-dependent, being efficacious only with the intermediate dose tested (10 mg/kg) and as compared to lower (3 mg/kg) or higher (30 mg/kg) doses. In regards to the effective dose-range for cannabidiol’s antidepressant-like efficacy through its ability to reduce immobility scores under the stress of a forced-swim test, prior studies suggested that lower doses were needed for mice (3–30 mg/kg) than rats (30–60 mg/kg) as reviewed in Silote et al. (2019). However, a recent study compared cannabidiol’s antidepressant-like potential in adolescent vs. adult rats and demonstrated that while a dose of 30 mg/kg was needed in adulthood to induce changes in the forced-swim test, only the intermediate dose tested (10 mg/kg) rendered efficacious in adolescent rats (Bis-Humbert et al., 2020), demonstrating a similar U-shaped dose–response curve for adolescent and aged rats, and lowering the dose at which cannabidiol could induce an antidepressant-like effect in older rats. A similar U-shaped pattern has been previously reported for cannabidiol’s anxiolytic-like effects, with effective responses at an intermediate dose, but no change with lower or higher doses (García-Gutiérrez et al., 2020 and references within) and for cannabidiol’s medicinal usage for cocaine-related behaviors has been reported (Nedelescu et al., 2022). Interestingly, the reduced rate in immobility was paralleled by an increase in climbing behavior, as previously reported for cannabidiol (Bis-Humbert et al., 2020; Ledesma-Corvi and García-Fuster, 2022) and in line with the antidepressants that exert their actions through the modulation of the noradrenergic system (Detke et al., 1995). In conjunction, and to the best of our knowledge, this is the first study to demonstrate that cannabidiol is capable of inducing dose-dependent responses in the forced-swim test indicative of an antidepressant-like response in naïve aged rats, which physiologically show increased negative effect as compared to younger rats (Hernández-Hernández et al., 2018). These data provide a new therapeutical option for late-life depression that deserves further characterization. In this context, the observed lack of effect of cannabidiol on body weight at the doses tested suggested a good safety profile for cannabidiol in aged rats. Although some prior studies reported that repeated cannabidiol treatment could decrease normal body weight gain in adult rats (Ignatowska-Jankowska et al., 2011; Santiago et al., 2019), an effect probably mediated by CB2 receptors (Ignatowska-Jankowska et al., 2011), these effects seemed to be age dependent, since they replicated in a separate study for adult rats, but were not observed when cannabidiol was administered at the same doses during adolescence (Bis-Humbert et al., 2020) or in aged rats (present study). Moreover, one might speculate that the effects observed in the forced-swim test could be driven by changes in locomotor activity; however, several studies with doses up to 60 mg/kg of cannabidiol reported no changes in spontaneous locomotion when administered alone (e.g., Taffe et al., 2015; Anooshe et al., 2021; Ledesma et al., 2021), including our own evaluations when measuring distance traveled in the open field test (e.g., Bis-Humbert et al., 2020; Ledesma-Corvi and García-Fuster, 2022).
In an attempt to study some of the potential molecular targets and/or markers modulated by cannabidiol in aged rats, we explored the regulation of CB receptors (e.g., Fogaça et al., 2018) and a marker of an early stage of hippocampal neurogenesis (i.e., cell proliferation; Luján et al., 2018, 2020; Bis-Humbert et al., 2020). The results showed no changes in any of the markers analyzed. As for CB receptors, cannabidiol did not alter the protein content of CB receptors in the hippocampus at the time rats were sacrificed. Further studies should be carried out to evaluate the dynamics on how cannabidiol modulates CB receptors and include other brain regions. Also, other age-related changes such as the observed decrease in cannabinoid receptor binding and mRNA levels in aged rats (Romero et al., 1998) must be considered. Moreover, regarding cell proliferation, our prior study also showed a lack of regulation by cannabidiol in adolescent or adult rats at the same doses tested here and at the same specific time-points of analysis (Bis-Humbert et al., 2020), reinforcing the idea that the previously described beneficial effects of cannabidiol on improving cell genesis were observed in the context of prior exposure to a given stressor (Luján et al., 2018, 2020). In any case, these results are limited by the fact that brains were analyzed 3 days post-treatment (1 day after the observed antidepressant-like effect), with this timing being a particular photo-finish, and not necessarily correlative of the behavioral effects, and therefore conclusions should be made cautiously and in the context of this limitation. Further studies should collect brains at the specific time when the antidepressant-like effect was observed.
Ideally, we would have included female rats to compare cannabidiol’s effects at this age range, since depression is about twice as common in women as in men (e.g., Labaka et al., 2018), and preclinical studies evaluating antidepressant-like responses in females are scarce in general, but even more so for female aged rodents (reviewed in Felice et al., 2015; Fernández-Guasti et al., 2017). Unfortunately, no female rats were available at the time of our experiments, and therefore, the effect of sex as a biological variable could not be included in the present study. In any case, prior studies suggested certain inefficacy for cannabidiol’s antidepressant-like potential in female adult rats (e.g., Silote et al., 2021), including our own (Ledesma-Corvi and García-Fuster, 2022), and thus recommending future studies to better characterize the potential of this drug in both sexes and at all age-ranges.
In conclusion, this study increased the age-window at which cannabidiol exerted dose-dependent responses in this behavioral test, to include aged rats (up to 21 months old), at which it could be considered as a potential antidepressant, and in line with other studies suggesting a beneficial role for this drug in age-related behavioral deficits. Further experiments should evaluate other parameters (i.e., a wider range of doses, longer treatment paradigms, and models of induced depression) and include female aged rats to not only characterize the behavioral potential of cannabidiol but also try to better understand the molecular mechanisms underlying its actions.
Data Availability Statement
The orginal contributions presented in this study are included in the article; further inquires can be directed to the corresponding author.
Ethics Statement
The animal study was reviewed and approved by the Local Bioethical Committee and the Regional Government, Balearic Islands, Spain.
Author Contributions
MJG-F conceived the study; MJG-F and EH-H designed the experiments; EH-H performed the experiments, analyzed hte data, and prepared the intiial figures; MJG-F revised all the data and reformatted the figures; EH-H prepared the first draft of the manuscript; MJG-F wrote the final version of the manuscript. Both authors have read, commented, edited and approved the final version of the manuscript.
Funding
This research was partly funded by Grants PID 2020-118582RB-I00 from MCIN/AEI/10.13039/501100011033 (Madrid, Spain), PDR2020/14 from Comunitat Autònoma de les Illes Balears (Direcció General de Política Universitària i Recerca with funds from the Tourist Stay Tax Law ITS 2017-006), and by Fundación Alicia Koplowitz (Madrid, Spain) to MJG-F; a predoctoral scholarship covered the salary of EH-H (FPI/2102/2018; CAIB), who is currently funded by the Margarita Salas Program (Ministerio de Universidades; Plan de Recuperación, Transformación y Resilencia; NextGenerationEU) with the participation of the University of the Balearic Islands.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors, and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
We would like to thank Professors Huda Akil and Stanley J. Watson (University of Michigan, Ann Arbor, MI, United States) for kindly providing Ki-67 antibody. Open Access was funded by the IV Convocatòria de publicacions en Accés Obert “Liberi” de l’IdISBa.
References
Anooshe, M., Nouri, K., Karimi-Haghighi, S., Mousavi, Z., and Haghparast, A. (2021). Cannabidiol Efficiently Suppressed the Acquisition and Expression of Methamphetamine-Induced Conditioned Place Preference in the Rat. Behav. Brain Res. 404, 113158. doi:10.1016/j.bbr.2021.113158
Bektas, A., Schurman, S. H., Sen, R., and Ferrucci, L. (2018). Aging, Inflammation and the Environment. Exp. Gerontol. 105, 10–18. doi:10.1016/j.exger.2017.12.015
Bis-Humbert, C., García-Cabrerizo, R., and García-Fuster, M. J. (2021a). Antidepressant-like Effects of Cannabidiol in a Rat Model of Early-Life Stress with or without Adolescent Cocaine Exposure. Pharmacol. Rep. 73, 1195–1202. doi:10.1007/s43440-021-00285-5
Bis-Humbert, C., García-Cabrerizo, R., and García-Fuster, M. J. (2020). Decreased Sensitivity in Adolescent versus Adult Rats to the Antidepressant-like Effects of Cannabidiol. Psychopharmacol. Berl. 237, 1621–1631. doi:10.1007/s00213-020-05481-4
Bis-Humbert, C., García-Cabrerizo, R., and García-Fuster, M. J. (2021b). Dose-dependent Opposite Effects of Nortriptyline on Affective-like Behavior in Adolescent Rats: Comparison with Adult Rats. Eur. J. Pharmacol. 910, 174465. doi:10.1016/j.ejphar.2021.174465
Bisogno, T., Hanus, L., De Petrocellis, L., Tchilibon, S., Ponde, D. E., Brandi, I., et al. (2001). Molecular Targets for Cannabidiol and its Synthetic Analogues: Effect on Vanilloid VR1 Receptors and on the Cellular Uptake and Enzymatic Hydrolysis of Anandamide. Br. J. Pharmacol. 134, 845–852. doi:10.1038/sj.bjp.0704327
Campos, A. C., Fogaça, M. V., Scarante, F. F., Joca, S. R. L., Sales, A. J., Gomes, F. V., et al. (2017). Plastic and Neuroprotective Mechanisms Involved in the Therapeutic Effects of Cannabidiol in Psychiatric Disorders. Front. Pharmacol. 8, 269. doi:10.3389/fphar.2017.00269
Campos, A. C., Fogaça, M. V., Sonego, A. B., and Guimarães, F. S. (2016). Cannabidiol, Neuroprotection and Neuropsychiatric Disorders. Pharmacol. Res. 112, 119–127. doi:10.1016/j.phrs.2016.01.033
Campos, A. C., Moreira, F. A., Gomes, F. V., del Bel, E. A., and Guimarães, F. S. (2012). Multiple Mechanisms Involved in the Large-Spectrum Therapeutic Potential of Cannabidiol in Psychiatric Disorders. Philos. Trans. R. Soc. Lond B Biol. Sci. 367, 3364–3378. doi:10.1098/rstb.2011.0389
Dash, R., Ali, M. C., Jahan, I., Munni, Y. A., Mitra, S., Hannan, M. A., et al. (2021). Emerging Potential of Cannabidiol in Reversing Proteinopathies. Ageing Res. Rev. 65, 101209. doi:10.1016/j.arr.2020.101209
Detke, M. J., Rickels, M., and Lucki, I. (1995). Active Behaviors in the Rat Forced Swimming Test Differentially Produced by Serotonergic and Noradrenergic Antidepressants. Psychopharmacol. Berl. 121, 66–72. doi:10.1007/BF02245592
Felice, D., O'Leary, O. F., Cryan, J. F., Dinan, T. G., Gardier, A. M., Sánchez, C., et al. (2015). When Ageing Meets the Blues: Are Current Antidepressants Effective in Depressed Aged Patients? Neurosci. Biobehav Rev. 55, 478–497. doi:10.1016/j.neubiorev.2015.06.005
Fernández-Guasti, A., Olivares-Nazario, M., Reyes, R., and Martínez-Mota, L. (2017). Sex and Age Differences in the Antidepressant-like Effect of Fluoxetine in the Forced Swim Test. Pharmacol. Biochem. Behav. 152, 81–89. doi:10.1016/j.pbb.2016.01.011
Fogaça, M. V., Campos, A. C., Coelho, L. D., Duman, R. S., and Guimarães, F. S. (2018). The Anxiolytic Effects of Cannabidiol in Chronically Stressed Mice Are Mediated by the Endocannabinoid System: Role of Neurogenesis and Dendritic Remodeling. Neuropharmacology 135, 22–33. doi:10.1016/j.neuropharm.2018.03.001
García-Cabrerizo, R., and García-Fuster, M. J. (2016). Opposite Regulation of Cannabinoid CB1 and CB2 Receptors in the Prefrontal Cortex of Rats Treated with Cocaine during Adolescence. Neurosci. Lett. 615, 60–65. doi:10.1016/j.neulet.2016.01.018
García-Cabrerizo, R., Keller, B., and García-Fuster, M. J. (2015). Hippocampal Cell Fate Regulation by Chronic Cocaine during Periods of Adolescent Vulnerability: Consequences of Cocaine Exposure during Adolescence on Behavioral Despair in Adulthood. Neuroscience 304, 302–315. doi:10.1016/j.neuroscience.2015.07.040
García-Fuster, M. J., Flagel, S. B., Mahmood, S. T., Mayo, L. M., Thompson, R. C., Watson, S. J., et al. (2011). Decreased Proliferation of Adult Hippocampal Stem Cells during Cocaine Withdrawal: Possible Role of the Cell Fate Regulator FADD. Neuropsychopharmacology 36, 2303–2317. doi:10.1038/npp.2011.119
García-Fuster, M. J., Parsegian, A., Watson, S. J., Akil, H., and Flagel, S. B. (2017). Adolescent Cocaine Exposure Enhances Goal-Tracking Behavior and Impairs Hippocampal Cell Genesis Selectively in Adult Bred Low-Responder Rats. Psychopharmacol. Berl. 234, 1293–1305. doi:10.1007/s00213-017-4566-0
García-Fuster, M. J., Perez, J. A., Clinton, S. M., Watson, S. J., and Akil, H. (2010). Impact of Cocaine on Adult Hippocampal Neurogenesis in an Animal Model of Differential Propensity to Drug Abuse. Eur. J. Neurosci. 31, 79–89. doi:10.1111/j.1460-9568.2009.07045.x
García-Gutiérrez, M. S., Navarrete, F., Gasparyan, A., Austrich-Olivares, A., Sala, F., and Manzanares, J. (2020). Cannabidiol: A Potential New Alternative for the Treatment of Anxiety, Depression, and Psychotic Disorders. Biomolecules 10, 1575. doi:10.3390/biom10111575
Gonzalez-Cuevas, G., Garcia-Gutierrez, M. S., Navarrete, F., de Guglielmo, G., and Manzanares, J. (2021). Editorial: Cannabidiol Treatment in Neurotherapeutic Interventions. Front. Pharmacol. 12, 752292. doi:10.3389/fphar.2021.752292
Haney, M., and Evins, A. E. (2016). Does Cannabis Cause, Exacerbate or Ameliorate Psychiatric Disorders? an Oversimplified Debate Discussed. Neuropsychopharmacology 41, 393–401. doi:10.1038/npp.2015.251
Hernández-Hernández, E., and García-Fuster, M. J. (2021). Evaluating the Effects of 2-BFI and Tracizoline, Two Potent I2-Imidazoline Receptor Agonists, on Cognitive Performance and Affect in Middle-Aged Rats. Naunyn Schmiedeb. Arch. Pharmacol. 394, 989–996. doi:10.1007/s00210-020-02042-6
Hernández-Hernández, E., Miralles, A., Esteban, S., and García-Fuster, M. J. (2018). Improved Age-Related Deficits in Cognitive Performance and Affective-like Behavior Following Acute, but Not Repeated, 8-OH-DPAT Treatments in Rats: Regulation of Hippocampal FADD. Neurobiol. Aging 71, 115–126.
Herrera-Pérez, J. J., Martínez-Mota, L., and Fernández-Guasti, A. (2008). Aging Increases the Susceptibility to Develop Anhedonia in Male Rats. Prog. Neuropsychopharmacol. Biol. Psychiatry 32, 1798–1803. doi:10.1016/j.pnpbp.2008.07.020
Ignatowska-Jankowska, B., Jankowski, M. M., and Swiergiel, A. H. (2011). Cannabidiol Decreases Body Weight Gain in Rats: Involvement of CB2 Receptors. Neurosci. Lett. 490, 82–84. doi:10.1016/j.neulet.2010.12.031
Labaka, A., Goñi-Balentziaga, O., Lebeña, A., and Pérez-Tejada, J. (2018). Biological Sex Differences in Depression: a Systematic Review. Biol. Res. Nurs. 20, 383–392. doi:10.1177/1099800418776082
Laprairie, R. B., Bagher, A. M., Kelly, M. E., and Denovan-Wright, E. M. (2015). Cannabidiol Is a Negative Allosteric Modulator of the Cannabinoid CB1 Receptor. Br. J. Pharmacol. 172, 4790–4805. doi:10.1111/bph.13250
Ledesma, J. C., Manzanedo, C., and Aguilar, M. A. (2021). Cannabidiol Prevents Several of the Behavioral Alterations Related to Cocaine Addiction in Mice. Prog. Neuropsychopharmacol. Biol. Psychiatry 111, 110390. doi:10.1016/j.pnpbp.2021.110390
Ledesma-Corvi, S., and García-Fuster, M. J. (2022). “Antidepresssant-like Effects of Cannabidiol in a Rat Model of Early-Life Stress: Sex- and Age-dependent Efficacy,” in European College of Neuropsychopharmacology Workshop, Nice, France, 17-20 March, 2022.
Luján, M. Á., Castro-Zavala, A., Alegre-Zurano, L., and Valverde, O. (2018). Repeated Cannabidiol Treatment Reduces Cocaine Intake and Modulates Neural Proliferation and CB1R Expression in the Mouse hippocampus. Neuropharmacology 143, 163–175. doi:10.1016/j.neuropharm.2018.09.043
Luján, M. A., Cartacorps, L., and Valverde, O. (2020). The Pharmacological Reduction of Hippocampal Neurogenesis Attenuates the Protective Effects of Cannabidiol on Cocaine Voluntary Intake. Addict. Biol. 25, e12778. doi:10.1111/adb.12778
Marchalant, Y., Brothers, H. M., Norman, G. J., Karelina, K., DeVries, A. C., and Wenk, G. L. (2009). Cannabinoids Attenuate the Effects of Aging upon Neuroinflammation and Neurogenesis. Neurobiol. Dis. 34, 300–307. doi:10.1016/j.nbd.2009.01.014
Marco, E. M., García-Gutiérrez, M. S., Bermúdez-Silva, F. J., Moreira, F. A., Guimarães, F., Manzanares, J., et al. (2011). Endocannabinoid System and Psychiatry: in Search of a Neurobiological Basis for Detrimental and Potential Therapeutic Effects. Front. Behav. Neurosci. 5, 63. doi:10.3389/fnbeh.2011.00063
Martín-Sánchez, A., González-Pardo, H., Alegre-Zurano, L., Castro-Zavala, A., López-Taboada, I., Valverde, O., et al. (2022). Early-life Stress Induces Emotional and Molecular Alterations in Female Mice that Are Partially Reversed by Cannabidiol. Prog. Neuropharmacol. Biol. Psychiatry 115, 110508. doi:10.1016/j.pnpbp.2021.110508
Martínez-Pinilla, E., Varani, K., Reyes-Resina, I., Angelats, E., Vincenzi, F., Ferreiro-Vera, C., et al. (2017). Binding and Signaling Studies Disclose a Potential Allosteric Site for Cannabidiol in Cannabinoid CB2 Receptors. Front. Pharmacol. 8, 744. doi:10.3389/fphar.2017.00744
Mastrodonato, A., Pavlova, I., Kee, N. C., Pham, V. A., McGowan, J. C., Mann, J. J., et al. (2022). Prophylactic (R,S)-ketamine Is Effective against Stress-Induced Behaviors in Adolescent but Not Aged Mice. Int. J. Neuropsychopharmacol. 2022, 20. doi:10.1093/ijnp/pyac020
Mattson, M. P., and Magnus, T. (2006). Ageing and Neuronal Vulnerability. Nat. Rev. Neurosci. 7, 278–294. doi:10.1038/nrn1886
McKinney, B. C., and Sibille, E. (2013). The Age-By-Disease Interaction Hypothesis of Late-Life Depression. Am. J. Geriatr. Psychiatry 21, 418–432. doi:10.1016/j.jagp.2013.01.053
Melas, P. A., Scherma, M., Fratta, W., Cifani, C., and Fadda, P. (2021). Cannabidiol as a Potential Treatment for Anxiety and Mood Disorders: Molecular Targets and Epigenetic Insights from Preclinical Research. Int. J. Mol. Sci. 22, 1863. doi:10.3390/ijms22041863
Nedelescu, H., Wagner, G. E., de Ness, G. L., Carroll, A., Kerr, T. M., Wang, J., et al. (2022). Cannabidiol Produces Distinct U-Shaped Dose-Response Effects on Cocaine-Induced Conditioned Place Preference and Associated Recruitment of Prelimbic Neurons in Male Rats. Biol. Psychiatry Glob. Open Sci. 2, 70–78. doi:10.1016/j.bpsgos.2021.06.014
Percie du Sert, N., Ahluwalia, A., Alam, S., Avey, M. T., Baker, M., Browne, W. J., et al. (2020). Reporting Animal Research: Explanation and Elaboration for the ARRIVE Guidelines 2.0. PLoS Biol. 18, e3000411. doi:10.1371/journal.pbio.3000411
Pertwee, R. G. (2008). The Diverse CB1 and CB2 Receptor Pharmacology of Three Plant Cannabinoids: Delta9-Tetrahydrocannabinol, Cannabidiol and Delta9-Tetrahydrocannabivarin. Br. J. Pharmacol. 153, 199–215. doi:10.1038/sj.bjp.0707442
Pisani, S., McGoohan, K., Velayudhan, L., and Bhattacharyya, S. (2021). Safety and Tolerability of Natural and Synthetic Cannabinoids in Older Adults: a Systematic Review and Meta-Analysis of Open-Label Trials and Observational Studies. Drugs Aging 38, 887–910. doi:10.1007/s40266-021-00882-2
Romero, J., Berrendero, F., Garcia-Gil, L., de la Cruz, P., Ramos, J. A., and Fernández-Ruiz, J. J. (1998). Loss of Cannabinoid Receptor Binding and Messenger RNA Levels and Cannabinoid Agonist-Stimulated [35S]guanylyl-5'O-(thio)-triphosphate Binding in the Basal Ganglia of Aged Rats. Neuroscience 84, 1075–1083. doi:10.1016/s0306-4522(97)00552-6
Rosenzweig, E. S., and Barnes, C. A. (2003). Impact of Aging on Hippocampal Function: Plasticity, Network Dynamics, and Cognition. Prog. Neurobiol. 69, 143–179. doi:10.1016/s0301-0082(02)00126-0
Santiago, A. N., Mori, M. A., Guimarães, F. S., Milani, H., and Weffort de Oliveira, R. M. (2019). Effects of Cannabidiol on Diabetes Outcomes and Chronic Cerebral Hypoperfusion Comorbidities in Middle-Aged Rats. Neurotox. Res. 35, 463–474. doi:10.1007/s12640-018-9972-5
Shbiro, L., Hen-Shoval, D., Hazut, N., Rapps, K., Dar, S., Zalsman, G., et al. (2019). Effects of Cannabidiol in Males and Females in Two Different Rat Models of Depression. Physiol. Behav. 201, 59–63. doi:10.1016/j.physbeh.2018.12.019
Silote, G. P., Gatto, M. C., Eskelund, A., Guimarães, F. S., Wegener, G., and Joca, S. R. L. (2021). Strain-, Sex-, and Time-dependent Antidepressant-like Effects of Cannabidiol. Pharm. (Basel) 14, 1269. doi:10.3390/ph14121269
Silote, G. P., Sartim, A., Sales, A., Eskelund, A., Guimarães, F. S., Wegener, G., et al. (2019). Emerging Evidence for the Antidepressant Effect of Cannabidiol and the Underlying Molecular Mechanisms. J. Chem. Neuroanat. 98, 104–116. doi:10.1016/j.jchemneu.2019.04.006
Slattery, D. A., and Cryan, J. F. (2012). Using the Rat Forced Swim Test to Assess Antidepressant-like Activity in Rodents. Nat. Protoc. 7, 1009–1014. doi:10.1038/nprot.2012.044
Sonnenberg, C. M., Deeg, D. J., Comijs, H. C., van Tilburg, W., and Beekman, A. T. (2008). Trends in Antidepressant Use in the Older Population: Results from the LASA-Study over a Period of 10 Years. J. Affect Disord. 111, 299–305. doi:10.1016/j.jad.2008.03.009
Stanciu, C. N., Brunette, M. F., Teja, N., and Budney, A. J. (2021). Evidence for Use of Cannabinoids in Mood Disorders, Anxiety Disorders, and PTSD: A Systematic Review. Psychiatr. Serv. 72, 429–436. doi:10.1176/appi.ps.202000189
Stek, M. L., Gussekloo, J., Beekman, A. T., van Tilburg, W., and Westendorp, R. G. (2004). Prevalence, Correlates and Recognition of Depression in the Oldest Old: the Leiden 85-plus Study. J. Affect Disord. 78, 193–200. doi:10.1016/S0165-0327(02)00310-5
Taffe, M. A., Creehan, K. M., and Vandewater, S. A. (2015). Cannabidiol Fails to Reverse Hypothermia or Locomotor Suppression Induced by Δ(9) -tetrahydrocannabinol in Sprague-Dawley Rats. Br. J. Pharmacol. 172, 1783–1791. doi:10.1111/bph.13024
Tham, M., Yilmaz, O., Alaverdashvili, M., Kelly, M. E. M., Denovan-Wright, E. M., and Laprairie, R. B. (2019). Allosteric and Orthosteric Pharmacology of Cannabidiol and Cannabidiol-Dimethylheptyl at the Type 1 and Type 2 Cannabinoid Receptors. Br. J. Pharmacol. 176, 1455–1469. doi:10.1111/bph.14440
Keywords: aging, late-life depression, antidepressants, rat, forced-swim test, hippocampus, CB receptors, neurogenesis
Citation: Hernández-Hernández E and García-Fuster MJ (2022) Dose-Dependent Antidepressant-Like Effects of Cannabidiol in Aged Rats. Front. Pharmacol. 13:891842. doi: 10.3389/fphar.2022.891842
Received: 08 March 2022; Accepted: 20 May 2022;
Published: 01 July 2022.
Edited by:
Maria S. Garcia-Gutierrez, Miguel Hernández University of Elche, SpainReviewed by:
Katarzyna Socała, Marie Curie-Sklodowska University, PolandF. Khakpai, Islamic Azad University, Iran
Copyright © 2022 Hernández-Hernández and García-Fuster. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: M. Julia García-Fuster, ai5nYXJjaWFAdWliLmVz
†Present Address: Department of Pharmacology, University of the Basque Country (EHU/UPV), Leioa, Spain