 Mazen Tolaymat
Mazen Tolaymat Margaret H. Sundel
Margaret H. Sundel Madeline Alizadeh
Madeline Alizadeh Guofeng Xie
Guofeng Xie Jean-Pierre Raufman
Jean-Pierre Raufman
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
MINI REVIEW article
Front. Pharmacol. , 04 November 2021
Sec. Gastrointestinal and Hepatic Pharmacology
Volume 12 - 2021 | https://doi.org/10.3389/fphar.2021.786105
This article is part of the Research Topic Insights in Gastrointestinal and Hepatic Pharmacology: 2021 View all 30 articles
Despite structural similarity, the five subtypes comprising the cholinergic muscarinic family of G protein-coupled receptors regulate remarkably diverse biological functions. This mini review focuses on the closely related and commonly co-expressed M1R and M3R muscarinic acetylcholine receptor subtypes encoded respectively by CHRM1 and CHRM3. Activated M1R and M3R signal via Gq and downstream initiate phospholipid turnover, changes in cell calcium levels, and activation of protein kinases that alter gene transcription and ultimately cell function. The unexpectedly divergent effects of M1R and M3R activation, despite similar receptor structure, distribution, and signaling, are puzzling. To explore this conundrum, we focus on the gastrointestinal (GI) tract and liver because abundant data identify opposing effects of M1R and M3R activation on the progression of gastric, pancreatic, and colon cancer, and liver injury and fibrosis. Whereas M3R activation promotes GI neoplasia, M1R activation appears protective. In contrast, in murine liver injury models, M3R activation promotes and M1R activation mitigates liver fibrosis. We analyze these findings critically, consider their therapeutic implications, and review the pharmacology and availability for research and therapeutics of M1R and M3R-selective agonists and antagonists. We conclude by considering gaps in knowledge and other factors that hinder the application of these drugs and the development of new agents to treat GI and liver diseases.
Muscarinic receptors (MRs) are class A (Rhodopsin-like) guanine nucleotide protein-coupled receptors (GPCRs) differentiated from other cholinergic receptors by preferential binding of muscarine rather than nicotine (Eglen, 2012; Tiwari et al., 2013). MRs are further subcategorized into five subtypes, designated M1R through M5R and encoded by CHRM1-CHRM5, each of which modulates a range of parasympathetic activities (Caulfield and Birdsall, 1998). These functionalities depend on tissue and membrane localization (Koenig and Edwardson, 1996; Nathanson, 2008). Like other GPCRs, MRs are characterized by seven transmembrane helices designated TM1 through TM7, forming a partially-spiral configuration within the cell membrane (Hulme et al., 2003). Acetylcholine (ACh) binds on the extracellular aspect of MRs in a pocket formed by TM3, TM6, and TM7 residues. The five MR subtypes share 82–92% transmembrane region homology, with 64–82% sequence similarity overall (Maeda et al., 2019). As GPCRs, activated MRs interact with heterotrimeric guanine nucleotide-binding proteins (G-proteins), classified by their α subunits, to activate downstream targets.
Although classically responsive to ACh, MRs, like other GPCRs, possess allosteric binding sites for naturally occurring and engineered non-ACh ligands, with varying degrees of preference; allosteric effects may result in surprising downstream actions in cell types not previously considered responsive to muscarinic signaling (Tolaymat et al., 2019). ACh and these “non-traditional” ligands provide MRs with the ability to modulate a broad repertoire of cells and biological systems including those associated with neuronal signaling, immune function, and cell trafficking, proliferation, and differentiation (Wessler and Kirkpatrick, 2008; McLean et al., 2016). Dysregulated post-MR signaling is associated with unregulated cell proliferation and cancer progression (Chen et al., 2019), an “overactive” bladder (Abrams et al., 2006), autoimmune diseases (Berg et al., 2010; Lee et al., 2013) and psychiatric disorders (Scarr, 2012; Vakalopoulos, 2014; Jeon et al., 2015). In addition to the discovery that non-traditional ligands can modify MR function, the production and release of ACh is more widespread than originally thought; a wide variety of non-neuronal cells express choline acetyltransferase (ChAT), the key enzyme needed to convert acetyl CoA and choline into ACh (Wessler and Kirkpatrick, 2012). Colon cancers, for example, express high levels of ChAT (Cheng et al., 2008). The variety of processes modulated by MRs has invited extensive research into the potential use of agonists, antagonists, and allosteric modulators for myriad disorders.
MRs are expressed by a wide variety of tissues and cell types and control key digestive and metabolic functions. Salivary gland secretion, gastric, and intestinal fluid transport, cell proliferation, mucus production, motility, and mesenteric vascular constriction and dilation are all responsive to MR signaling (Tobin et al., 2009; Muise et al., 2017). In the stomach, M3R, M4R, and M5R activation modulates hydrochloric acid secretion from parietal cells (Aihara et al., 2005), and M1R and M3R activation stimulates pepsinogen secretion from chief cells (Xie et al., 2005). GI motility, through intestinal smooth muscle cell action, involves communication between the central and enteric nervous systems. These effects are partially mediated by M1R through M3R (Moro et al., 2005), with M2R and M3R playing a role in regulating longitudinal muscle contraction, and all three MR subtypes involved in circular muscle function (Harrington et al., 2010; Tanahashi et al., 2021). MR-mediated regulation of smooth muscle function extends throughout the entire GI tract. Nonetheless, it is likely that MRs play additional roles in regulating small intestinal function; for example, M2R is expressed in the stem cell compartment and may be involved in enterocyte turnover (Muise et al., 2017). ACh has both pro- (Koyama et al., 1992; Brunn et al., 1995) and anti-inflammatory (Pavlov and Tracey, 2006) effects, the latter mediated in part by reducing systemic levels of tumor necrosis factor. While the ubiquity of MRs within the digestive tract makes them attractive therapeutic targets to modulate health and disease, this same ubiquity complicates efforts to design selective agents while minimizing off-target adverse effects.
Responses of MRs to ligand binding are subtype specific. Activation of odd-numbered MRs (M1R, M3R, and M5R) stimulates phospholipid turnover and increases intracellular calcium levels while activation of even-numbered MRs (M2R, M4R) inhibits adenylyl cyclase activity, thereby reducing levels of intracellular cAMP. M1R, M3R, and M5R (MRodd) canonically couple to Gq/11 which induces the phospholipase C-mediated hydrolysis of phosphatidylinositol (4.5)-bisphosphate into diacylglycerol and inositol (1,4,5)-trisphosphate. The latter binds an endoplasmic reticulum receptor stimulating intracellular calcium release. However, these may represent oversimplifications; experimental findings suggest differential interactions of individual MRodd and MReven with their downstream targets. For example, although both M1R and M3R signal through phospholipase C, CHO cells expressing M1R exhibited four-fold greater cAMP production in response to carbachol compared to cells expressing M3R (Burford et al., 1995). Likewise, although M2R and M4R (MReven) act primarily by binding Gi/o family proteins to alter adenylyl cyclase activity, their actions can also prolong potassium channel opening, thereby causing cellular hyperpolarization (Bubser et al., 2012).
These general principles do not tell the whole story–despite substantial sequence homology among MR subtypes they demonstrate surprising individuality in their responses to stimuli, even within the same cell and when responding to the same ligand. Pancreatic acinar cells provide a useful model to study muscarinic control of exocrine digestive function. Using acinar cells prepared from M1R- and M3R-deficient mice as well as M1/M3 chimeric receptors, Nakamura et al., demonstrated greater ACh-induced IP3 release in cells expressing only M1R compared to those expressing uniquely M3R (Nakamura et al., 2013). Moreover, in M3R-compared to M1R-expressing cells, these differences were associated respectively with oscillatory versus monotonic patterns of cytosolic calcium release. Oscillatory calcium release was a function of a C-terminal region of M3R with considerable variability among MR subtypes (Nakamura et al., 2013). In murine gastric chief cells, both M1R and M3R mediate pepsinogen secretion–deletion of either MR subtype reduces and combined M1R and M3R deficiency ablates cholinergic agonist-induced proenzyme secretion (Xie et al., 2005). Thus, in some cell types, MRodd have overlapping functions whereas in other cell types, MR subtype signaling appears divergent. In addition to the influence of their cell and tissue localization, other mechanistic differences between MR subtypes result in sometimes-opposing effects. GPCRs, including MRs, can also undergo “pre-coupling”, wherein a stable multimeric complex is present before ligand binding. Unlike other MR subtypes, M1R and M3R pre-couple with Gi/o G-proteins, their non-preferential G protein, thereby potentially altering downstream effects (Jakubík et al., 2011). Crystal structures of inactive M1-4R subtypes provide some insight into different allosteric and orthosteric binding sites (Kruse et al., 2012; Thal et al., 2016), but our understanding of the resulting functional differences between MR subtypes continues to evolve.
As a result of their central role in maintaining homeostasis in the GI tract, dysregulated MR signaling can be an important modifier of intestinal disease. In Hirschsprung disease, lack of mucosal cholinergic innervation in aganglionic colon segments increases the risk of postoperative enterocolitis (Keck et al., 2021). In diarrhea-predominant irritable bowel syndrome (IBS-D) without a concomitant psychiatric disorder, pyridostigmine (an acetylcholinesterase inhibitor) induces a stronger IL-6 response that is highly correlated with symptoms (Dinan et al., 2008). Given the pharmacotherapies targeting MRs already approved or being explored to treat IBS (Tables 1, 2), achieving a more precise mechanistic understanding of the role MR dysregulation plays in IBS is important.
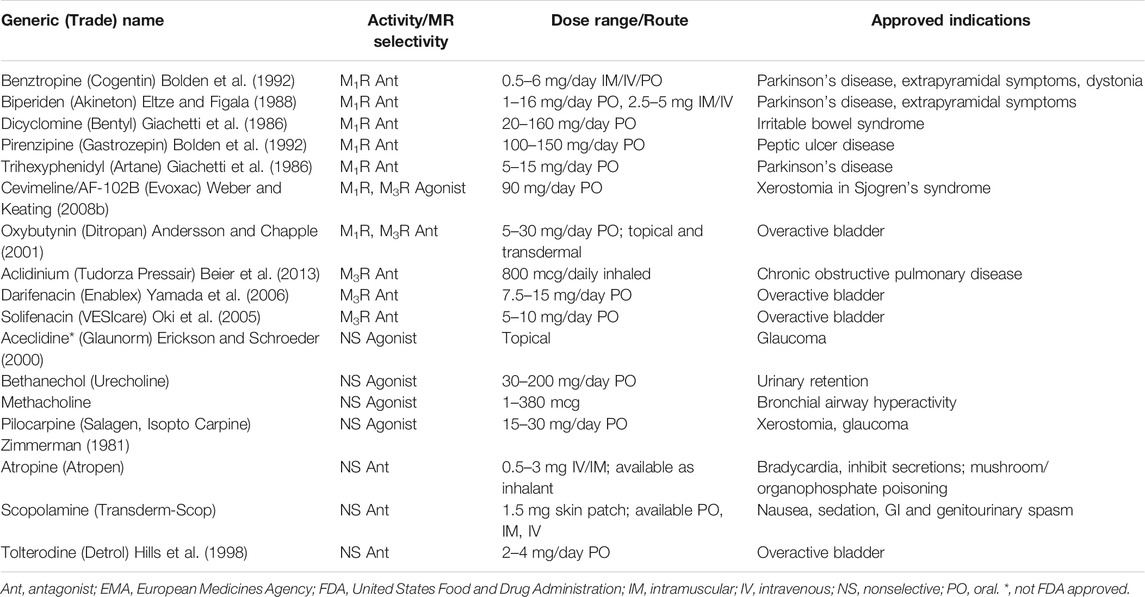
TABLE 1. FDA/EMA approved muscarinic receptor antagonists and agonists.
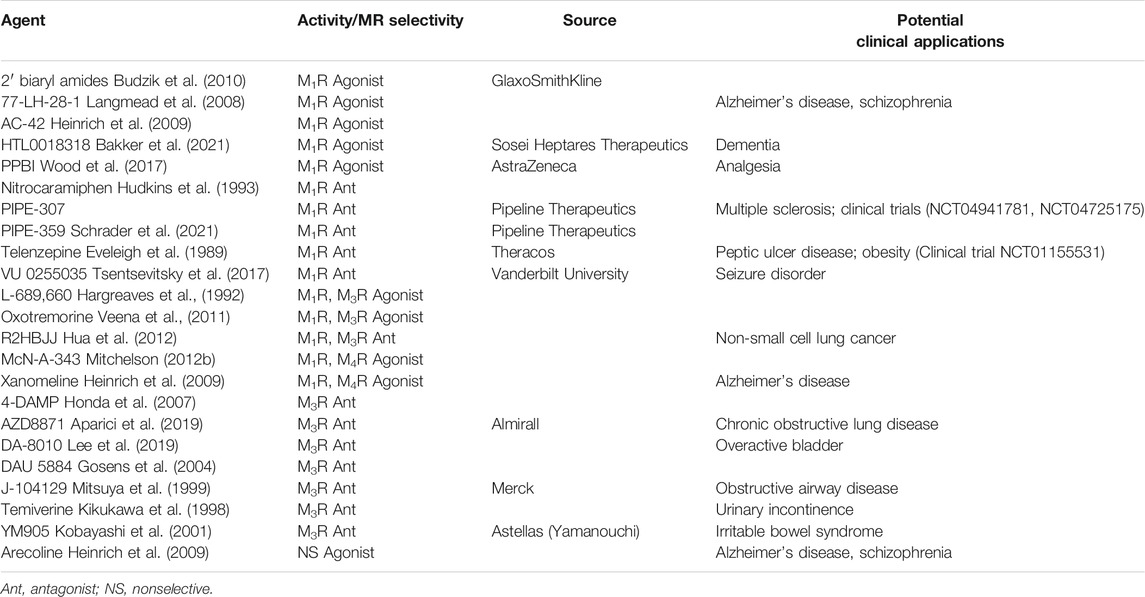
TABLE 2. Selective M1R/M3R agents used for research and under clinical investigation.
Diseases associated with MR dysregulation are not restricted to the lower GI tract. In the stomach, cholinergic signaling is balanced with histamine and gastrin release to regulate gastric acid levels; peptic ulcer disease is associated with greater MR expression in the gastric body, whereas progressive MR loss in that region is associated with chronic gastritis (Pfeiffer et al., 1995). In progressive systemic sclerosis and Sjogren’s syndrome, an autoimmune condition which impairs lacrimal and salivary function, esophageal dysmotility may be associated with anti-M3R antibodies (Goldblatt et al., 2002; Kawaguchi et al., 2009; Gyger and Baron, 2012); anti-M3R antibodies are also reported in progressive systemic sclerosis with anal dysmotility (Singh et al., 2009; Gyger and Baron, 2012). Intravenous immunoglobulin to neutralize anti-M3R antibodies may be beneficial (Smith et al., 2005).
Compared to the normal liver, individuals with primary biliary cholangitis (PBC) are more likely to have a CHRM3 single nucleotide polymorphism (rs4620530) of uncertain significance; this is not associated with baseline disease characteristics or treatment responses (Greverath et al., 2020). PBC is more commonly associated with anti-M3R antibodies than other liver diseases (Tsuboi et al., 2014); those with anti-M3R antibodies are more likely to have a benign disease course. Nonetheless, M3R antibody levels do not correlate with treatment responses or serological markers either at baseline or during the disease course (Mayer et al., 2020). A subset of patients with PBC develop Sjogren’s syndrome; the shared increase in anti-M3R antibody levels in both conditions suggests overlapping features could form the basis for a mutual treatment.
MRs play key roles in normal cell proliferation and turnover. As reviewed by Campoy et al. (2016), presumably to benefit tumor progression, neoplastic cells hijack MR-dependent proliferative signal transduction pathways. Treating neoplastic cells with exogenous ACh and inhibiting ACh hydrolysis promotes their proliferation and, conversely, reducing M3R expression and activation is anti-proliferative. Moreover, because neoplastic cells tend to lose cellular polarity, receptors normally expressed on the basolateral membrane may be expressed more diffusely around the cell membrane, thereby facilitating their access to orthosteric and allosteric ligands in the tumor microenvironment and GI lumen (Cheng et al., 2002). For example, bile acids, at concentrations achieved in stool, promote atropine-inhibitable colon cancer cell proliferation (Cheng and Raufman, 2005).
Abundant data support the conclusion that M3R plays an important role in colon cancer progression. In mouse models of sporadic and genetic colon cancer, using azoxymethane (AOM)-treated and ApcMin/+ mice, respectively, Chrm3 ablation with resulting M3R deficiency substantially reduces the intestinal tumor burden (Raufman et al., 2008; Raufman et al., 2011). As M3R deficiency primarily reduces the number of adenocarcinomas rather than adenomas, the major impact of blocking M3R activation appears to be on promotion, rather than initiation, of neoplasia. M3R activation has similar pro-proliferative effects on gastric cancer (Hayakawa et al., 2017; Wang et al., 2018). M3R expression is enhanced in cholangiocarcinoma and associated with reduced cell differentiation, perineural invasion, and metastasis (Feng et al., 2012; Feng et al., 2018).
In contrast to the impact of M3R deficiency, M1R deficiency in mice does not attenuate, and may modestly enhance, AOM-induced colon carcinogenesis. Strikingly, mice with combined M1R and M3R deficiency develop as many colon tumors as control mice (Cheng et al., 2014); that is, M1R deficiency negates the anti-neoplastic effects of M3R deficiency. Likewise, M1R agonism appears protective against pancreatic ductal adenocarcinoma (PDAC) and counteracts enhanced carcinogenesis following vagotomy (Renz et al., 2018), suggesting a potential therapeutic opportunity. In contrast, in hepatocellular and prostate carcinomas, M1R activation promotes cellular migration and invasiveness (Yin et al., 2018; Zhang et al., 2020). Notably, many of these studies are limited by using global rather than conditional knockout mouse models. Hence, it remains uncertain whether the respective MR deficiencies are due to effects on neoplastic cells versus other cellular elements in the tumor microenvironment (e.g., immunocytes). Nonetheless, these observations argue strongly for the importance of MR subtype selectivity in designing and developing therapeutics.
Branches of the vagus nerve, a major source of ACh signaling within the GI tract, innervate the liver and modulate hepatocyte regeneration by progenitor cells and fibrosis by stellate cells (Cassiman et al., 2002). The current lack of effective anti-fibrotic therapies highlights the potential of leveraging these muscarinic actions to prevent or reverse fibrosis in advanced liver disease and stimulate hepatocyte regeneration. For example, in rodents, carbon tetrachloride (CCl4)-induced hepatic fibrosis can be attenuated by vagotomy and treatment with atropine (Lam et al., 2008). M3R expression and activation protects against AOM-induced liver fibrosis (Khurana et al., 2010; Khurana et al., 2013; Rachakonda et al., 2015). Surprisingly, M1R expression and activation appears to have opposite effects, worsening AOM-induced hepatic fibrosis (Rachakonda et al., 2015). Thus, in the absence of effective anti-fibrotic therapy, manipulation of MR subtype activity to limit or reverse fibrosis may have therapeutic potential although, again, divergent effects in different tissues warrants caution.
MR subtype, tissue distribution, and off-target side effects have hindered efforts to manipulate MR activity precisely and effectively with drugs. MR antagonists are most effective in treating chronic obstructive pulmonary disease and overactive bladder (Table 1) (Eglen et al., 1999; Athanasopoulos and Giannitsas, 2011)—their utility for GI and hepatic disorders is currently limited. Cholinesterase inhibitors that increase ACh levels, also used clinically for digestive tract disorders, have similar limitations as their actions are largely non-selective. Adverse effects with these classes of drugs are attributed primarily to off-target effects on the CNS (e.g., convulsions, confusion) and other peripheral MR subtypes (e.g., sialorrhea, rhinitis, diaphoresis, diarrhea, nausea, vomiting, and bronchospasm). Novel MR agonists and antagonists are currently under investigation primarily for diseases of the central nervous system such as Alzheimer’s disease and schizophrenia (Table 2) (Davie et al., 2013).
Several MR agonists and cholinesterase inhibitors are in clinical use. Oral and topical pilocarpine. and cevimeline (Evoxac), an M3R-selective activator, augment salivary gland secretions in xerostomia due to radiation therapy and Sjogren’s syndrome (Iga et al., 1998; Fife et al., 2002; Petrone et al., 2002; Weber and Keating, 2008a; Berk, 2008; Mitchelson, 2012a; Davies and Thompson, 2015; Panarese and Moshirfar, 2021). Bethanechol, a structural analogue of ACh that resists hydrolysis by cholinesterases, has potential to treat esophageal dysmotility. Currently approved to treat urinary retention and neurogenic bladder (Gaitonde et al., 2019), bethanechol strengthens esophageal contractions in subjects with ineffective esophageal motility (Agrawal et al., 2007) and augments lower esophageal sphincter pressure in gastroesophageal reflux disease (Farrell et al., 1973). Nonetheless, in a pilot study, topical bethanechol did not significantly improve esophageal motility (O’Rourke et al., 2013). Edrophonium, a cholinesterase inhibitor used to diagnose myasthenia gravis, was used to provoke esophageal spasm in the investigation of non-cardiac chest pain, but the lack of correlation between symptoms and objective changes in esophageal manometry limited its utility (Botoman, 2002).
Gastric acid secretion is controlled by a mix of cholinergic muscarinic stimulation and hormonal signaling by gastrin and histamine; thus, only partial inhibition of acid release is achieved with anti-muscarinic agents. Consequently, histamine-2 receptor and H+-K+ATPase (proton pump) inhibitors are highly successful and MR antagonists rarely prescribed. Pirenzepine (Gastrozepin), an M1R antagonist that is not FDA approved, has limited use to treat acid-related disorders in the EU (Tryba and Cook, 1997). Scopolamine, a non-selective MR antagonist, is commonly used as a transdermal patch for nausea associated with anesthesia or motion sickness (Riad and Hithe, 2021).
Dicyclomine (Bentyl), an M1R- and M3R-selective antagonist that inhibits small and large intestinal motility, is used as an anti-spasmodic agent to treat IBS (Giachetti et al., 1986; Doods et al., 1987). Neostigmine, a cholinesterase inhibitor, is used to treat acute intestinal pseudo-obstruction associated with critical illness or opioid use, another condition of impaired smooth muscle motility. Colonic decompression may be achieved with intravenous neostigmine (De Giorgio et al., 2001), although cardiac monitoring is important and rapid administration of atropine may be required for resulting bradycardia.
Although a potential role for modulating MR activity to treat cancer was demonstrated in a variety of cell types (Shah et al., 2009), except for an ongoing trial to investigate the utility of bethanechol before surgery for resectable PDAC (U.S. National Library of Medicine, 2021), the efficacy of modulators of MR activity in digestive tract cancers has not been tested in the clinic. Moreover, anti-tumor efficacy may be limited by the inability to achieve adequate concentrations in target tissues while, at the same time, preventing off-target adverse effects. An ideal agent would exhibit target organ and MR-subtype specificity, goals hampered by the extensive similarity between orthosteric and allosteric ligand binding sites among the five MR subtypes (Liu et al., 2018). Studies of naturally occurring ligands, such as muscarinic toxins in snake venom, have provided insight into how subtype-selective agents may be formulated (Maeda et al., 2020). Such agents with potential for oncotherapy continue to be developed. For example, the M3R-specific antagonist darifenacin which is approved to treat bladder dysfunction (Yamada et al., 2006) reportedly inhibits tumor progression and invasiveness in human-derived cell lines, most recently in colorectal cancer cell lines (Hering et al., 2021). As darifenacin is in clinical use with a known safety profile, it is an attractive candidate for adjunctive therapy, especially for cancers already shown to overexpress M3R, like colon cancer cells (Frucht et al., 1999; Cheng et al., 2014), PDAC (Zhang et al., 2016), and non-small cell lung cancer (Lin et al., 2014).
Some therapeutic approaches may circumvent the need for MR subtype and tissue specificity. For example, treating colorectal cancers with poorly absorbed oral agents or drugs with extensive first-pass metabolism may target GI mucosal lesions with limited systemic side effects. However, even within a limited area of distribution, MRs are not constrained to only one downstream signaling pathway; the same receptor may have contradictory effects on neighboring cell types. Even when occupying the same binding pocket, ligands can influence the activation of pathways on other cell membrane surfaces via signaling bias and functional selectivity (Randáková and Jakubík, 2021). A ligand may bind several MR subtypes, but only activate one or a few, thereby compensating for binding pocket homogeneity. Furthermore, through selective interactions with residues in the binding pocket of a single subtype, ligands can encourage activated receptor configurations that favor interaction with certain G proteins. As an example, the MR agonist cevimeline increased intracellular calcium levels in Chinese hamster ovary (CHO) cells transfected with rat M1R but did not increase cAMP levels. In contrast, carbachol, a non-selective MR agonist, elevated both calcium and cAMP levels (Gurwitz et al., 1994). Even more intriguing, cevimeline did not activate signaling in M3R-transfected cells, contrary to its clinical use in Sjogren’s syndrome which is thought to be mediated by M3R activation. This complexity makes it difficult to predict the clinical effects of new MR agonists and antagonists but suggests highly selective agents can be developed.
MR activation via the vagus nerve, the longest and most complex cranial nerve, and within the enteric nervous system, is a major modifier of normal and pathological GI and hepatic function. As reviewed here, MRs and the machinery needed to produce their ligands are not limited to neuronal cells. Abundant evidence exists that “non-traditional” ligands (e.g., other than ACh) mediate paracrine and autocrine signaling by orthosteric and allosteric interactions with MR subtypes. These findings highlight the potential for treating a broad range of physiological and disease processes with MR subtype-selective agents. Numerous non-selective and subtype-selective orthosteric ligands that modify MR signaling have been developed and investigated to treat a variety of digestive diseases (Tables 1, 2); allosteric regulation of MR activity represents a presently untapped reservoir of agents that can be designed or repurposed to alter cell function. Overall, there has been limited clinical use of both orthosteric and allosteric modifiers of MR function. Despite more than 20 years of evidence supporting an important role for MR activation in GI cancer progression, currently only one clinical trial is investigating the efficacy of a drug to modulate MR activity as adjunctive treatment for a digestive tract cancer, PDAC (ClinicalTrials.gov Identifier: NCT03572283).
Extensive sequence homology between the five MR subtypes hampers efforts to create agents with sufficiently selective actions and, thereby, limited off-target toxicity. Adding to this complexity is the observation that a receptor subtype on one cell type may activate different downstream signaling pathways, depending on the interaction between ligand and receptor and the conformational changes instigated by this interaction. In addition to subtype-specificity, ideal agents must possess sufficient tissue specificity to prevent deleterious action on neighboring and distant tissues. In this regard, targeting diseases involving intestinal mucosa, e.g., neoplasia, may be advantaged by developing agents with limited GI absorption or extensive first-pass metabolism. Current gaps in knowledge include a better understanding of subtype-selective allosteric modulation of MR function, an area in its infancy.
Lastly, several observations reviewed above suggest great potential for leveraging the divergent actions of M1R and M3R activation to treat GI cancers. Thus, a drug design challenge is to develop a molecule with dual functionality as an M1R agonist and M3R antagonist. Moreover, it has not escaped our attention that developing an agent with the opposite properties may be useful to prevent or reverse hepatic fibrosis. Success at creating dual agonists for different bile acid receptors in the gut suggests that although the challenge is formidable, it can be overcome (Ito et al., 2021). As our understanding of these complex signaling mechanisms evolves and the medicinal chemistry needed to develop MR subtype-specific agents progresses, targeting MR subtypes is likely to become a valuable adjunct for treating a variety of digestive tract disorders, including cancer.
MT and JR conceptualized and wrote the initial draft. MS, MA, and GX proofread, edited, and contributed additional material. MT and JR completed the final draft.
This work was funded by the United States (U.S.) Department of Veterans Affairs Biomedical Laboratory Research and Development Program, VA Merit Award grant numbers BX002777 and BX004890. MT, MS, and MA were supported by the U.S. National Institutes of Health, grant number T32 DK067872. The contents do not represent the views of the U.S. Department of Veterans Affairs or the U.S. Government.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Abrams, P., Andersson, K. E., Buccafusco, J. J., Chapple, C., de Groat, W. C., Fryer, A. D., et al. (2006). Muscarinic Receptors: Their Distribution and Function in Body Systems, and the Implications for Treating Overactive Bladder. Br. J. Pharmacol. 148 (5), 565–578. doi:10.1038/sj.bjp.0706780
Agrawal, A., Hila, A., Tutuian, R., Mainie, I., and Castell, D. O. (2007). Bethanechol Improves Smooth Muscle Function in Patients with Severe Ineffective Esophageal Motility. J. Clin. Gastroenterol. 41 (4), 366–370. doi:10.1097/01.mcg.0000225542.03880.68
Aihara, T., Nakamura, Y., Taketo, M. M., Matsui, M., and Okabe, S. (2005). Cholinergically Stimulated Gastric Acid Secretion Is Mediated by M(3) and M(5) but Not M(1) Muscarinic Acetylcholine Receptors in Mice. Am. J. Physiol. Gastrointest. Liver Physiol. 288 (6), G1199–G1207. doi:10.1152/ajpgi.00514.2004
Andersson, K. E., and Chapple, C. R. (2001). Oxybutynin and the Overactive Bladder. World J. Urol. 19 (5), 319–323. doi:10.1007/pl00007103
Aparici, M., Carcasona, C., Ramos, I., Montero, JL, Otal, R, Ortiz, JL, et al. (2019). Pharmacological Profile of AZD8871 (LAS191351), a Novel Inhaled Dual M(3) Receptor Antagonist/β (2)-Adrenoceptor Agonist Molecule with Long-Lasting Effects and Favorable Safety Profile. J. Pharmacol. Exp. Ther. 370 (1), 127–136. doi:10.1124/jpet.118.255620
Athanasopoulos, A., and Giannitsas, K. (2011). An Overview of the Clinical Use of Antimuscarinics in the Treatment of Overactive Bladder. Adv. Urol. 2011, 820816. doi:10.1155/2011/820816
Bakker, C., Tasker, T., Liptrot, J., and Hart, EP, Klaassen, ES, Prins, S, et al. (2021). First-in-man Study to Investigate Safety, Pharmacokinetics and Exploratory Pharmacodynamics of HTL0018318, a Novel M(1) -receptor Partial Agonist for the Treatment of Dementias. Br. J. Clin. Pharmacol. 87 (7), 2945–2955. doi:10.1111/bcp.14710
Beier, J., Kirsten, A. M., Mróz, R., and Segarra, R, Chuecos, F, Caracta, C, et al. (2013). Efficacy and Safety of Aclidinium Bromide Compared with Placebo and Tiotropium in Patients with Moderate-To-Severe Chronic Obstructive Pulmonary Disease: Results from a 6-week, Randomized, Controlled Phase IIIb Study. Copd. 10 (4), 511–522. doi:10.3109/15412555.2013.814626
Berg, C. P., Blume, K., Lauber, K., Gregor, M., Berg, P. A., Wesselborg, S., et al. (2010). Autoantibodies to Muscarinic Acetylcholine Receptors Found in Patients with Primary Biliary Cirrhosis. BMC Gastroenterol. 10, 120. doi:10.1186/1471-230x-10-120
Berk, L. (2008). Systemic Pilocarpine for Treatment of Xerostomia. Expert Opin. Drug Metab. Toxicol. 4 (10), 1333–1340. doi:10.1517/17425255.4.10.1333
Bolden, C., Cusack, B., and Richelson, E. (1992). Antagonism by Antimuscarinic and Neuroleptic Compounds at the Five Cloned Human Muscarinic Cholinergic Receptors Expressed in Chinese Hamster Ovary Cells. J. Pharmacol. Exp. Ther. 260 (2), 576–580.
Botoman, V. A. (2002). Noncardiac Chest Pain. J. Clin. Gastroenterol. 34 (1), 6–14. doi:10.1097/00004836-200201000-00004
Brunn, G., Wessler, I., and Racké, K. (1995). Mucosa-dependent Muscarinic Liberation of Prostaglandins from Rat Isolated Trachea. Br. J. Pharmacol. 116 (3), 1991–1998. doi:10.1111/j.1476-5381.1995.tb16403.x
Bubser, M., Byun, N., Wood, M. R., and Jones, C. K. (2012). “Muscarinic Receptor Pharmacology and Circuitry for the Modulation of Cognition,” in Muscarinic Receptors. Editors A D Fryer, A Christopoulos, and NM Nathanson (Springer, Berlin Heidelberg), 121–166. doi:10.1007/978-3-642-23274-9_7
Budzik, B., Garzya, V., Shi, D., and Walker, G, Lauchart, Y, Lucas, AJ, et al. (2010). 2' Biaryl Amides as Novel and Subtype Selective M1 Agonists. Part II: Further Optimization and Profiling. Bioorg. Med. Chem. Lett. 20 (12), 3545–3549. doi:10.1016/j.bmcl.2010.04.127
Burford, N. T., Tobin, A. B., and Nahorski, S. R. (1995). Differential Coupling of M1, M2 and M3 Muscarinic Receptor Subtypes to Inositol 1,4,5-trisphosphate and Adenosine 3',5'-cyclic Monophosphate Accumulation in Chinese Hamster Ovary Cells. J. Pharmacol. Exp. Ther. 274 (1), 134–142.
Campoy, F. J., Vidal, C. J., Muñoz-Delgado, E., Montenegro, M. F., Cabezas-Herrera, J., and Nieto-Cerón, S. (2016). Cholinergic System and Cell Proliferation. Chem. Biol. Interact 259 (Pt B), 257–265. doi:10.1016/j.cbi.2016.04.014
Cassiman, D., Libbrecht, L., Sinelli, N., Desmet, V., Denef, C., and Roskams, T. (2002). The Vagal Nerve Stimulates Activation of the Hepatic Progenitor Cell Compartment via Muscarinic Acetylcholine Receptor Type 3. Am. J. Pathol. 161 (2), 521–530. doi:10.1016/s0002-9440(10)64208-3
Caulfield, M. P., and Birdsall, N. J. (1998). International Union of Pharmacology. XVII. Classification of Muscarinic Acetylcholine Receptors. Pharmacol. Rev. 50 (2), 279–290.
Chen, J., Cheuk, I. W. Y., Shin, V. Y., and Kwong, A. (2019). Acetylcholine Receptors: Key Players in Cancer Development. Surg. Oncol. 31, 46–53. doi:10.1016/j.suronc.2019.09.003
Cheng, K., Khurana, S., Chen, Y., Kennedy, R. H., Zimniak, P., and Raufman, J. P. (2002). Lithocholylcholine, a Bile Acid/acetylcholine Hybrid, Is a Muscarinic Receptor Antagonist. J. Pharmacol. Exp. Ther. 303 (1), 29–35. doi:10.1124/jpet.102.036376
Cheng, K., and Raufman, J. P. (2005). Bile Acid-Induced Proliferation of a Human colon Cancer Cell Line Is Mediated by Transactivation of Epidermal Growth Factor Receptors. Biochem. Pharmacol. 70 (7), 1035–1047. doi:10.1016/j.bcp.2005.07.023
Cheng, K., Samimi, R., Xie, G., Shant, J., Drachenberg, C., Wade, M., et al. (2008). Acetylcholine Release by Human colon Cancer Cells Mediates Autocrine Stimulation of Cell Proliferation. Am. J. Physiol. Gastrointest. Liver Physiol. 295 (3), G591–G597. doi:10.1152/ajpgi.00055.2008
Cheng, K., Xie, G., Khurana, S., Heath, J., Drachenberg, C. B., Timmons, J., et al. (2014). Divergent Effects of Muscarinic Receptor Subtype Gene Ablation on Murine colon Tumorigenesis Reveals Association of M3R and Zinc finger Protein 277 Expression in colon Neoplasia. Mol. Cancer 13, 77. doi:10.1186/1476-4598-13-77
Davie, B. J., Christopoulos, A., and Scammells, P. J. (2013). Development of M1 mAChR Allosteric and Bitopic Ligands: Prospective Therapeutics for the Treatment of Cognitive Deficits. ACS Chem. Neurosci. 4 (7), 1026–1048. doi:10.1021/cn400086m
Davies, A. N., and Thompson, J. (2015). Parasympathomimetic Drugs for the Treatment of Salivary Gland Dysfunction Due to Radiotherapy. Cochrane Database Syst. Rev. 2020 (10), Cd003782. doi:10.1002/14651858.CD003782.pub3
De Giorgio, R., Barbara, G., Stanghellini, V., Tonini, M., Vasina, V., Cola, B., et al. (2001). Review Article: the Pharmacological Treatment of Acute Colonic Pseudo-obstruction. Aliment. Pharmacol. Ther. 15 (11), 1717–1727. doi:10.1046/j.1365-2036.2001.01088.x
Dinan, T. G., Clarke, G., Quigley, E. M., Scott, L. V., Shanahan, F., Cryan, J., et al. (2008). Enhanced Cholinergic-Mediated Increase in the Pro-inflammatory Cytokine IL-6 in Irritable Bowel Syndrome: Role of Muscarinic Receptors. Am. J. Gastroenterol. 103 (10), 2570–2576. doi:10.1111/j.1572-0241.2008.01871.x
Doods, H. N., Mathy, M. J., Davidesko, D., van Charldorp, K. J., de Jonge, A., and van Zwieten, P. A. (1987). Selectivity of Muscarinic Antagonists in Radioligand and In Vivo Experiments for the Putative M1, M2 and M3 Receptors. J. Pharmacol. Exp. Ther. 242 (1), 257–262.
Eglen, R. M., Choppin, A., Dillon, M. P., and Hegde, S. (1999). Muscarinic Receptor Ligands and Their Therapeutic Potential. Curr. Opin. Chem. Biol. 3 (4), 426–432. doi:10.1016/s1367-5931(99)80063-5
Eglen, R. M. (2012). Overview of Muscarinic Receptor Subtypes. Handb Exp. Pharmacol. 208, 3–28. doi:10.1007/978-3-642-23274-9_1
Eltze, M., and Figala, V. (1988). Affinity and Selectivity of Biperiden Enantiomers for Muscarinic Receptor Subtypes. Eur. J. Pharmacol. 158 (1-2), 11–19. doi:10.1016/0014-2999(88)90247-6
Erickson, K. A., and Schroeder, A. (2000). Direct Effects of Muscarinic Agents on the Outflow Pathways in Human Eyes. Invest. Ophthalmol. Vis. Sci. 41 (7), 1743–1748.
Eveleigh, P., Hulme, E. C., Schudt, C., and Birdsall, N. J. (1989). The Existence of Stable Enantiomers of Telenzepine and Their Stereoselective Interaction with Muscarinic Receptor Subtypes. Mol. Pharmacol. 35 (4), 477–483.
Farrell, R. L., Roling, G. T., and Castell, D. O. (1973). Stimulation of the Incompetent Lower Esophageal Sphincter. A Possible advance in Therapy of Heartburn. Am. J. Dig. Dis. 18 (8), 646–650. doi:10.1007/bf01072035
Feng, Y., Hu, X., Liu, G., Lu, L., Zhao, W., Shen, F., et al. (2018). M3 Muscarinic Acetylcholine Receptors Regulate Epithelial-Mesenchymal Transition, Perineural Invasion, and Migration/metastasis in Cholangiocarcinoma through the AKT Pathway. Cancer Cel Int 18, 173. doi:10.1186/s12935-018-0667-z
Feng, Y. J., Zhang, B. Y., Yao, R. Y., and Lu, Y. (2012). Muscarinic Acetylcholine Receptor M3 in Proliferation and Perineural Invasion of Cholangiocarcinoma Cells. Hepatobiliary Pancreat. Dis. Int. 11 (4), 418–423. doi:10.1016/s1499-3872(12)60201-x
Fife, R. S., Chase, W. F., Dore, R. K., Wiesenhutter, C. W., Lockhart, P. B., Tindall, E., et al. (2002). Cevimeline for the Treatment of Xerostomia in Patients with Sjögren Syndrome: a Randomized Trial. Arch. Intern. Med. 162 (11), 1293–1300. doi:10.1001/archinte.162.11.1293
Frucht, H., Jensen, R. T., Dexter, D., Yang, W. L., and Xiao, Y. (1999). Human colon Cancer Cell Proliferation Mediated by the M3 Muscarinic Cholinergic Receptor. Clin. Cancer Res. 5 (9), 2532–2539.
Gaitonde, S., Malik, R. D., Christie, A. L., and Zimmern, P. E. (2019). Bethanechol: Is it Still Being Prescribed for Bladder Dysfunction in Women. Int. J. Clin. Pract. 73 (8), e13248. doi:10.1111/ijcp.13248
Giachetti, A., Giraldo, E., Ladinsky, H., and Montagna, E. (1986). Binding and Functional Profiles of the Selective M1 Muscarinic Receptor Antagonists Trihexyphenidyl and Dicyclomine. Br. J. Pharmacol. 89 (1), 83–90. doi:10.1111/j.1476-5381.1986.tb11123.x
Goldblatt, F., Gordon, T. P., and Waterman, S. A. (2002). Antibody-mediated Gastrointestinal Dysmotility in Scleroderma. Gastroenterology 123 (4), 1144–1150. doi:10.1053/gast.2002.36057
Gosens, R., Bromhaar, M. M., Tonkes, A., and Schaafsma, D, Zaagsma, J, Nelemans, SA, et al. (2004). Muscarinic M(3) Receptor-dependent Regulation of Airway Smooth Muscle Contractile Phenotype. Br. J. Pharmacol. 141 (6), 943–950. doi:10.1038/sj.bjp.0705709
Greverath, L. M., Leicht, E., Wald de Chamorro, N., Wilde, A. B., Steinhagen, L. M., Lieb, C., et al. (2020). Evaluation of Muscarinic Acetylcholine Receptor Type 3 Gene Polymorphisms in Patients with Primary Biliary Cholangitis and Primary Sclerosing Cholangitis. Hepatol. Res. 50 (3), 321–329. doi:10.1111/hepr.13455
Gurwitz, D., Haring, R., Heldman, E., Fraser, C. M., Manor, D., and Fisher, A. (1994). Discrete Activation of Transduction Pathways Associated with Acetylcholine M1 Receptor by Several Muscarinic Ligands. Eur. J. Pharmacol. Mol. Pharmacol. 267 (1), 21–31. doi:10.1016/0922-4106(94)90220-8
Gyger, G., and Baron, M. (2012). Gastrointestinal Manifestations of Scleroderma: Recent Progress in Evaluation, Pathogenesis, and Management. Curr. Rheumatol. Rep. 14 (1), 22–29. doi:10.1007/s11926-011-0217-3
Hargreaves, R. J., McKnight, A. T., Scholey, K., and Newberry, NR, Street, LJ, Hutson, PH, et al. (1992). 689,660, a Novel Cholinomimetic with Functional Selectivity for M1 and M3 Muscarinic Receptors. Br. J. Pharmacol. 107 (2), 494–501. doi:10.1111/j.1476-5381.1992.tb12773.x
Harrington, A. M., Peck, C. J., Liu, L., Burcher, E., Hutson, J. M., and Southwell, B. R. (2010). Localization of Muscarinic Receptors M1R, M2R and M3R in the Human colon. Neurogastroenterol Motil. 22 (9), 999–1008. doi:10.1111/j.1365-2982.2009.01456.x
Hayakawa, Y., Sakitani, K., Konishi, M., Asfaha, S., Niikura, R., Tomita, H., et al. (2017). Nerve Growth Factor Promotes Gastric Tumorigenesis through Aberrant Cholinergic Signaling. Cancer Cell 31 (1), 21–34. doi:10.1016/j.ccell.2016.11.005
Heinrich, J. N., Butera, J. A., Carrick, T., and Kramer, A, Kowal, D, Lock, T, et al. (2009). Pharmacological Comparison of Muscarinic Ligands: Historical versus More Recent Muscarinic M1-Preferring Receptor Agonists. Eur. J. Pharmacol. 605 (1-3), 53–56. doi:10.1016/j.ejphar.2008.12.044
Hering, N. A., Liu, V., Kim, R., and Weixler, B, Droeser, RA, Arndt, M, et al. (2021). Blockage of Cholinergic Signaling via Muscarinic Acetylcholine Receptor 3 Inhibits Tumor Growth in Human Colorectal Adenocarcinoma. Cancers (Basel) 13 (13). doi:10.3390/cancers13133220
Hills, C. J., Winter, S. A., and Balfour, J. A. (1998). Tolterodine. Drugs. 55 (6), 813–820. doi:10.2165/00003495-199855060-00008
Honda, H., Tomizawa, M., and Casida, J. E. (2007). Insect Muscarinic Acetylcholine Receptor: Pharmacological and Toxicological Profiles of Antagonists and Agonists. J. Agric. Food Chem. 55 (6), 2276–2281. doi:10.1021/jf0631934
Hua, N., Wei, X., Liu, X., and Ma, X, He, X, Zhuo, R, et al. (2012). A Novel Muscarinic Antagonist R2HBJJ Inhibits Non-small Cell Lung Cancer Cell Growth and Arrests the Cell Cycle in G0/G1. PLoS One 7 (12), e53170. doi:10.1371/journal.pone.0053170
Hudkins, R. L., Stubbins, J. F., and DeHaven-Hudkins, D. L. (1993). Caramiphen, Iodocaramiphen and Nitrocaramiphen Are Potent, Competitive, Muscarinic M1 Receptor-Selective Agents. Eur. J. Pharmacol. 231 (3), 485–488. doi:10.1016/0014-2999(93)90130-a
Hulme, E. C., Lu, Z. L., Saldanha, J. W., and Bee, M. S. (2003). Structure and Activation of Muscarinic Acetylcholine Receptors. Biochem. Soc. Trans. 31 (Pt 1), 29–34. doi:10.1042/bst0310029
Iga, Y., Arisawa, H., Ogane, N., Saito, Y., Tomizuka, T., Nakagawa-Yagi, Y., et al. (1998). (+/-)-cis-2-methylspiro[1,3-oxathiolane-5,3'-quinuclidine] Hydrochloride, Hemihydrate (SNI-2011, Cevimeline Hydrochloride) Induces Saliva and Tear Secretions in Rats and Mice: the Role of Muscarinic Acetylcholine Receptors. Jpn. J. Pharmacol. 78 (3), 373–380. doi:10.1254/jjp.78.373
Ito, K., Okumura, A., Takeuchi, J. S., and Watashi, K, Inoue, R, Yamauchi, T, et al. (2021). Dual Agonist of Farnesoid X Receptor and Takeda G Protein-Coupled Receptor 5 Inhibits Hepatitis B Virus Infection In Vitro and In Vivo. Hepatology 74 (1), 83–98. doi:10.1002/hep.31712
Jakubík, J., Janíčková, H., Randáková, A., El-Fakahany, E. E., and Doležal, V. (2011). Subtype Differences in Pre-coupling of Muscarinic Acetylcholine Receptors. PLoS One 6 (11), e27732. doi:10.1371/journal.pone.0027732
Jeon, W. J., Dean, B., Scarr, E., and Gibbons, A. (2015). The Role of Muscarinic Receptors in the Pathophysiology of Mood Disorders: A Potential Novel Treatment. Curr. Neuropharmacol 13 (6), 739–749. doi:10.2174/1570159x13666150612230045
Kawaguchi, Y., Nakamura, Y., Matsumoto, I., Nishimagi, E., Satoh, T., Kuwana, M., et al. (2009). Muscarinic-3 Acetylcholine Receptor Autoantibody in Patients with Systemic Sclerosis: Contribution to Severe Gastrointestinal Tract Dysmotility. Ann. Rheum. Dis. 68 (5), 710–714. doi:10.1136/ard.2008.096545
Keck, S., Galati-Fournier, V., Kym, U., Moesch, M., Usemann, J., Müller, I., et al. (2021). Lack of Mucosal Cholinergic Innervation Is Associated with Increased Risk of Enterocolitis in Hirschsprung's Disease. Cell Mol Gastroenterol Hepatol 12 (2), 507–545. doi:10.1016/j.jcmgh.2021.03.004
Khurana, S., Jadeja, R., Twaddell, W., Cheng, K., Rachakonda, V., Saxena, N., et al. (2013). Effects of Modulating M3 Muscarinic Receptor Activity on Azoxymethane-Induced Liver Injury in Mice. Biochem. Pharmacol. 86 (2), 329–338. doi:10.1016/j.bcp.2013.05.010
Khurana, S., Shah, N., Cheng, K., Shiu, B., Samimi, R., Belo, A., et al. (2010). Scopolamine Treatment and Muscarinic Receptor Subtype-3 Gene Ablation Augment Azoxymethane-Induced Murine Liver Injury. J. Pharmacol. Exp. Ther. 333 (3), 639–649. doi:10.1124/jpet.109.165118
Kikukawa, H., Yoshida, M., Wada, Y., Nishi, K., and Ueda, S. (1998). Pharmacologic Actions of Temiverine (P-INN) and its Active Metabolite, RCC-36, on Isolated Human Urinary Bladder Muscle. Int. J. Urol. 5 (3), 268–275. doi:10.1111/j.1442-2042.1998.tb00602.x
Kobayashi, S., Ikeda, K., Suzuki, M., Yamada, T., and Miyata, K. (2001). Effects of YM905, a Novel Muscarinic M3-Receptor Antagonist, on Experimental Models of Bowel Dysfunction In Vivo. Jpn. J. Pharmacol. 86 (3), 281–288. doi:10.1254/jjp.86.281
Koenig, J. A., and Edwardson, J. M. (1996). Intracellular Trafficking of the Muscarinic Acetylcholine Receptor: Importance of Subtype and Cell Type. Mol. Pharmacol. 49 (2), 351–359.
Koyama, S., Rennard, S. I., and Robbins, R. A. (1992). Acetylcholine Stimulates Bronchial Epithelial Cells to Release Neutrophil and Monocyte Chemotactic Activity. Am. J. Physiol. 262 (4 Pt 1), L466–L471. doi:10.1152/ajplung.1992.262.4.L466
Kruse, A. C., Hu, J., Pan, A. C., Arlow, D. H., Rosenbaum, D. M., Rosemond, E., et al. (2012). Structure and Dynamics of the M3 Muscarinic Acetylcholine Receptor. Nature 482 (7386), 552–556. doi:10.1038/nature10867
Lam, H. B., Yeh, C. H., Cheng, K. C., Hsu, C. T., and Cheng, J. T. (2008). Effect of Cholinergic Denervation on Hepatic Fibrosis Induced by Carbon Tetrachloride in Rats. Neurosci. Lett. 438 (1), 90–95. doi:10.1016/j.neulet.2008.04.048
Langmead, C. J., Austin, N. E., Branch, C. L., and Brown, JT, Buchanan, KA, Davies, CH, et al. (2008). Characterization of a CNS Penetrant, Selective M1 Muscarinic Receptor Agonist, 77-LH-28-1. Br. J. Pharmacol. 154 (5), 1104–1115. doi:10.1038/bjp.2008.152
Lee, B. H., Gauna, A. E., Perez, G., Park, Y. J., Pauley, K. M., Kawai, T., et al. (2013). Autoantibodies against Muscarinic Type 3 Receptor in Sjögren's Syndrome Inhibit Aquaporin 5 Trafficking. PLoS One 8 (1), e53113. doi:10.1371/journal.pone.0053113
Lee, M. J., Moon, J. H., Lee, H. K., Cho, C. H., Choi, S. H., and Im, W. B. (2019). Pharmacological Characterization of DA-8010, a Novel Muscarinic Receptor Antagonist Selective for Urinary Bladder over Salivary Gland. Eur. J. Pharmacol. 843, 240–250. doi:10.1016/j.ejphar.2018.11.037
Lin, G., Sun, L., Wang, R., Guo, Y., and Xie, C. (2014). Overexpression of Muscarinic Receptor 3 Promotes Metastasis and Predicts Poor Prognosis in Non-small-cell Lung Cancer. J. Thorac. Oncol. 9 (2), 170–178. doi:10.1097/JTO.0000000000000066
Liu, H., Hofmann, J., Fish, I., Schaake, B., Eitel, K., Bartuschat, A., et al. (2018). Structure-guided Development of Selective M3 Muscarinic Acetylcholine Receptor Antagonists. Proc. Natl. Acad. Sci. U S A. 115 (47), 12046–12050. doi:10.1073/pnas.1813988115
Maeda, S., Qu, Q., Robertson, M. J., Skiniotis, G., and Kobilka, B. K. (2019). Structures of the M1 and M2 Muscarinic Acetylcholine receptor/G-Protein Complexes. Science 364 (6440), 552–557. doi:10.1126/science.aaw5188
Maeda, S., Xu, J., Kadji, F. M. N., Clark, M. J., Zhao, J., Tsutsumi, N., et al. (2020). Structure and Selectivity Engineering of the M1 Muscarinic Receptor Toxin Complex. Science 369 (6500), 161–167. doi:10.1126/science.aax2517
Mayer, C., Preuss, B., Grottenthaler, J., Berg, C., and Klein, R. (2020). Antibodies to the Muscarinic Acetylcholine Receptor M3 in Primary Biliary Cholangitis Inhibit Receptor Function on Cholangiocytes. Front. Immunol. 11, 1151. doi:10.3389/fimmu.2020.01151
McLean, L. P., Smith, A., Cheung, L., Urban, J. F., Sun, R., Grinchuk, V., et al. (2016). Type 3 Muscarinic Receptors Contribute to Intestinal Mucosal Homeostasis and Clearance of Nippostrongylus Brasiliensis through Induction of TH2 Cytokines. Am. J. Physiol. Gastrointest. Liver Physiol. 311 (1), G130–G141. doi:10.1152/ajpgi.00461.2014
U.S. National Library of Medicine (2021). Bethanechol Prior to Pancreatic Surgery. Available at : https://clinicaltrials.gov/ct2/show/NCT03572283.
Mitchelson, F. (2012). Muscarinic Receptor Agonists and Antagonists: Effects on Ocular Function. Handb Exp. Pharmacol. 208, 263–298. doi:10.1007/978-3-642-23274-9_12
Mitchelson, F. J. (2012). The Pharmacology of McN-A-343. Pharmacol. Ther. 135 (2), 216–245. doi:10.1016/j.pharmthera.2012.05.008
Mitsuya, M., Mase, T., Tsuchiya, Y., and Kawakami, K, Hattori, H, Kobayashi, K, et al. (1999). J-104129, a Novel Muscarinic M3 Receptor Antagonist with High Selectivity for M3 over M2 Receptors. Bioorg. Med. Chem. 7 (11), 2555–2567. doi:10.1016/s0968-0896(99)00177-7
Moro, E., Crema, F., Dandolo, C., De Ponti, F., and Frigo, G. (2005). Effect of Muscarinic Receptor Blockade on Canine Gastric Tone and Compliance In Vivo. Pharmacol. Res. 51 (4), 289–296. doi:10.1016/j.phrs.2004.09.007
Muise, E. D., Gandotra, N., Tackett, J. J., Bamdad, M. C., and Cowles, R. A. (2017). Distribution of Muscarinic Acetylcholine Receptor Subtypes in the Murine Small Intestine. Life Sci. 169, 6–10. doi:10.1016/j.lfs.2016.10.030
Nakamura, K., Hamada, K., Terauchi, A., Matsui, M., Nakamura, T., Okada, T., et al. (2013). Distinct Roles of M1 and M3 Muscarinic Acetylcholine Receptors Controlling Oscillatory and Non-oscillatory [Ca2+]i Increase. Cell Calcium 54 (2), 111–119. doi:10.1016/j.ceca.2013.05.004
Nathanson, N. M. (2008). Synthesis, Trafficking, and Localization of Muscarinic Acetylcholine Receptors. Pharmacol. Ther. 119 (1), 33–43. doi:10.1016/j.pharmthera.2008.04.006
O'Rourke, A., Weinberger, P., Morrison, M., Conklin, J., and Postma, G. (2013). Topical Bethanechol for the Improvement of Esophageal Dysmotility: a Pilot Study. Ann. Otol Rhinol Laryngol. 122 (8), 481–486. doi:10.1177/000348941312200801
Oki, T., Sato, S., Miyata, K., and Yamada, S. (2005). Muscarinic Receptor Binding, Plasma Concentration and Inhibition of Salivation after Oral Administration of a Novel Antimuscarinic Agent, Solifenacin Succinate in Mice. Br. J. Pharmacol. 145 (2), 219–227. doi:10.1038/sj.bjp.0706184
Panarese, V., and Moshirfar, M. (2021). Pilocarpine. StatPearls. StatPearls Publishing Copyright © 2021. Treasure Island, FA: StatPearls Publishing LLC.
Pavlov, V. A., and Tracey, K. J. (2006). Controlling Inflammation: the Cholinergic Anti-inflammatory Pathway. Biochem. Soc. Trans. 34 (Pt 6), 1037–1040. doi:10.1042/bst0341037
Petrone, D., Condemi, J. J., Fife, R., Gluck, O., Cohen, S., and Dalgin, P. (2002). A Double-Blind, Randomized, Placebo-Controlled Study of Cevimeline in Sjögren's Syndrome Patients with Xerostomia and Keratoconjunctivitis Sicca. Arthritis Rheum. 46 (3), 748–754. doi:10.1002/art.510
Pfeiffer, A., Krömer, W., Friemann, J., Ruge, M., Herawi, M., Schätzl, M., et al. (1995). Muscarinic Receptors in Gastric Mucosa Are Increased in Peptic Ulcer Disease. Gut 36 (6), 813–818. doi:10.1136/gut.36.6.813
Rachakonda, V., Jadeja, R. N., Urrunaga, N. H., Shah, N., Ahmad, D., Cheng, K., et al. (2015). M1 Muscarinic Receptor Deficiency Attenuates Azoxymethane-Induced Chronic Liver Injury in Mice. Sci. Rep. 5, 14110. doi:10.1038/srep14110
Randáková, A., and Jakubík, J. (2021). Functionally Selective and Biased Agonists of Muscarinic Receptors. Pharmacol. Res. 169, 105641. doi:10.1016/j.phrs.2021.105641
Raufman, J. P., Samimi, R., Shah, N., Khurana, S., Shant, J., Drachenberg, C., et al. (2008). Genetic Ablation of M3 Muscarinic Receptors Attenuates Murine colon Epithelial Cell Proliferation and Neoplasia. Cancer Res. 68 (10), 3573–3578. doi:10.1158/0008-5472.Can-07-6810
Raufman, J. P., Shant, J., Xie, G., Cheng, K., Gao, X. M., Shiu, B., et al. (2011). Muscarinic Receptor Subtype-3 Gene Ablation and Scopolamine Butylbromide Treatment Attenuate Small Intestinal Neoplasia in Apcmin/+ Mice. Carcinogenesis 32 (9), 1396–1402. doi:10.1093/carcin/bgr118
Renz, B. W., Tanaka, T., Sunagawa, M., Takahashi, R., Jiang, Z., Macchini, M., et al. (2018). Cholinergic Signaling via Muscarinic Receptors Directly and Indirectly Suppresses Pancreatic Tumorigenesis and Cancer Stemness. Cancer Discov. 8 (11), 1458–1473. doi:10.1158/2159-8290.Cd-18-0046
Riad, M., and Hithe, C. C. (2021). Scopolamine. StatPearls. StatPearls Publishing. Copyright © 2021. Treasure Island, FA: StatPearls Publishing LLC.
Scarr, E. (2012). Muscarinic Receptors: Their Roles in Disorders of the central Nervous System and Potential as Therapeutic Targets. CNS Neurosci. Ther. 18 (5), 369–379. doi:10.1111/j.1755-5949.2011.00249.x
Schrader, T. O., Xiong, Y., Lorenzana, A. O., and Broadhead, A, Stebbins, KJ, Poon, MM, et al. (2021). Discovery of PIPE-359, a Brain-Penetrant, Selective M(1) Receptor Antagonist with Robust Efficacy in Murine MOG-EAE. ACS Med. Chem. Lett. 12 (1), 155–161. doi:10.1021/acsmedchemlett.0c00626
Shah, N., Khurana, S., Cheng, K., and Raufman, J. P. (2009). Muscarinic Receptors and Ligands in Cancer. Am. J. Physiol. Cel Physiol 296 (2), C221–C232. doi:10.1152/ajpcell.00514.2008
Singh, J., Mehendiratta, V., Del Galdo, F., Jimenez, S. A., Cohen, S., DiMarino, A. J., et al. (2009). Immunoglobulins from Scleroderma Patients Inhibit the Muscarinic Receptor Activation in Internal Anal Sphincter Smooth Muscle Cells. Am. J. Physiol. Gastrointest. Liver Physiol. 297 (6), G1206–G1213. doi:10.1152/ajpgi.00286.2009
Smith, A. J., Jackson, M. W., Wang, F., Cavill, D., Rischmueller, M., and Gordon, T. P. (2005). Neutralization of Muscarinic Receptor Autoantibodies by Intravenous Immunoglobulin in Sjögren Syndrome. Hum. Immunol. 66 (4), 411–416. doi:10.1016/j.humimm.2005.01.020
Tanahashi, Y., Komori, S., Matsuyama, H., Kitazawa, T., and Unno, T. (2021). Functions of Muscarinic Receptor Subtypes in Gastrointestinal Smooth Muscle: A Review of Studies with Receptor-Knockout Mice. Ijms 22 (2), 926. doi:10.3390/ijms22020926
Thal, D. M., Sun, B., Feng, D., Nawaratne, V., Leach, K., Felder, C. C., et al. (2016). Crystal Structures of the M1 and M4 Muscarinic Acetylcholine Receptors. Nature 531 (7594), 335–340. doi:10.1038/nature17188
Tiwari, P., Dwivedi, S., Singh, M. P., Mishra, R., and Chandy, A. (2013). Basic and Modern Concepts on Cholinergic Receptor: A Review. Asian Pac. J. Trop. Dis. 3 (5), 413–420. doi:10.1016/S2222-1808(13)60094-8
Tobin, G., Giglio, D., and Lundgren, O. (2009). Muscarinic Receptor Subtypes in the Alimentary Tract. J. Physiol. Pharmacol. 60 (1), 3–21.
Tolaymat, M., Larabee, S. M., Hu, S., Xie, G., and Raufman, J. P. (2019). The Role of M3 Muscarinic Receptor Ligand-Induced Kinase Signaling in Colon Cancer Progression. Cancers (Basel) 11 (3). doi:10.3390/cancers11030308
Tryba, M., and Cook, D. (1997). Current Guidelines on Stress Ulcer Prophylaxis. Drugs 54 (4), 581–596. doi:10.2165/00003495-199754040-00005
Tsentsevitsky, A. N., Kovyazina, I. V., Nurullin, L. F., and Nikolsky, E. E. (2017). Muscarinic Cholinoreceptors (M1-, M2-, M3- and M4-type) Modulate the Acetylcholine Secretion in the Frog Neuromuscular junction. Neurosci. Lett. 649, 62–69. doi:10.1016/j.neulet.2017.04.015
Tsuboi, H., Ohira, H., Asashima, H., Tsuzuki, S., Iizuka, M., Matsuo, N., et al. (2014). Anti-M3 Muscarinic Acetylcholine Receptor Antibodies in Patients with Primary Biliary Cirrhosis. Hepatol. Res. 44 (14), E471–E479. doi:10.1111/hepr.12346
Vakalopoulos, C. (2014). The Effect of Deficient Muscarinic Signaling on Commonly Reported Biochemical Effects in Schizophrenia and Convergence with Genetic Susceptibility Loci in Explaining Symptom Dimensions of Psychosis. Front. Pharmacol. 5, 277. doi:10.3389/fphar.2014.00277
Veena, J., Srikumar, B. N., Mahati, K., Raju, T. R., and Shankaranarayana Rao, B. S. (2011). Oxotremorine Treatment Restores Hippocampal Neurogenesis and Ameliorates Depression-like Behaviour in Chronically Stressed Rats. Psychopharmacology (Berl). 217 (2), 239–253. doi:10.1007/s00213-011-2279-3
Wang, L., Xu, J., Xia, Y., Yin, K., Li, Z., Li, B., et al. (2018). Muscarinic Acetylcholine Receptor 3 Mediates Vagus Nerve-Induced Gastric Cancer. Oncogenesis 217 (11), 88. doi:10.1038/s41389-018-0099-6
Weber, J., and Keating, G. M. (2008). Cevimeline. Drugs. 68 (12), 1691–1698. doi:10.2165/00003495-200868120-00006
Weber, J., and Keating, G. M. (2008). Cevimeline. Drugs 68 (12), 1691–1698. doi:10.2165/00003495-200868120-00006
Wessler, I., and Kirkpatrick, C. J. (2008). Acetylcholine beyond Neurons: the Non-neuronal Cholinergic System in Humans. Br. J. Pharmacol. 154 (8), 1558–1571. doi:10.1038/bjp.2008.185
Wessler, I. K., and Kirkpatrick, C. J. (2012). Activation of Muscarinic Receptors by Non-neuronal Acetylcholine. Handb Exp. Pharmacol. 208, 469–491. doi:10.1007/978-3-642-23274-9_20
Wood, M. W., Martino, G., Coupal, M., and Lindberg, M, Schroeder, P, Santhakumar, V, et al. (2017). Broad Analgesic Activity of a Novel, Selective M1 Agonist. Neuropharmacology 123, 233–241. doi:10.1016/j.neuropharm.2017.06.010
Xie, G., Drachenberg, C., Yamada, M., Wess, J., and Raufman, J. P. (2005). Cholinergic Agonist-Induced Pepsinogen Secretion from Murine Gastric Chief Cells Is Mediated by M1 and M3 Muscarinic Receptors. Am. J. Physiol. Gastrointest. Liver Physiol. 289 (3), G521–G529. doi:10.1152/ajpgi.00105.2004
Yamada, S., Maruyama, S., Takagi, Y., Uchida, S., and Oki, T. (2006). In Vivo demonstration of M3 Muscarinic Receptor Subtype Selectivity of Darifenacin in Mice. Life Sci. 80 (2), 127–132. doi:10.1016/j.lfs.2006.08.028
Yin, Q. Q., Xu, L. H., Zhang, M., and Xu, C. (2018). Muscarinic Acetylcholine Receptor M1 Mediates Prostate Cancer Cell Migration and Invasion through Hedgehog Signaling. Asian J. Androl. 20 (6), 608–614. doi:10.4103/aja.aja_55_18
Zhang, L., Wu, L. L., Huan, H. B., Wen, X. D., Yang, D. P., Chen, D. F., et al. (2020). Activation of Muscarinic Acetylcholine Receptor 1 Promotes Invasion of Hepatocellular Carcinoma by Inducing Epithelial-Mesenchymal Transition. Anticancer Drugs 31 (9), 908–917. doi:10.1097/cad.0000000000000907
Zhang, L., Xiu, D., Zhan, J., and He, X, Guo, L, Wang, J, et al. (2016). High Expression of Muscarinic Acetylcholine Receptor 3 Predicts Poor Prognosis in Patients with Pancreatic Ductal Adenocarcinoma. Onco Targets Ther. 9, 6719–6726. doi:10.2147/OTT.S111382
Keywords: muscarinic receptors, G protein-coupled receptors, gastrointestinal physiology, gastrointestinal disease, liver disease, cancer
Citation: Tolaymat M, Sundel MH, Alizadeh M, Xie G and Raufman J-P (2021) Potential Role for Combined Subtype-Selective Targeting of M1 and M3 Muscarinic Receptors in Gastrointestinal and Liver Diseases. Front. Pharmacol. 12:786105. doi: 10.3389/fphar.2021.786105
Received: 29 September 2021; Accepted: 19 October 2021;
Published: 04 November 2021.
Edited by:
Ralf Weiskirchen, RWTH Aachen University, GermanyReviewed by:
Ritva Tikkanen, University of Giessen, GermanyCopyright © 2021 Tolaymat, Sundel, Alizadeh, Xie and Raufman. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jean-Pierre Raufman, anJhdWZtYW5Ac29tLnVtYXJ5bGFuZC5lZHU=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.