 João Victor Dutra Gomes
João Victor Dutra Gomes Corinna Herz
Corinna Herz Simone Helmig2
Simone Helmig2 Nadja Förster
Nadja Förster Inga Mewis
Inga Mewis Evelyn Lamy
Evelyn Lamy- 1Molecular Preventive Medicine, University Medical Center and Faculty of Medicine—University of Freiburg, Freiburg, Germany
- 2Institute for Occupational and Social Medicine and Department of Anesthesiology, Justus-Liebig University Giessen, Giessen, Germany
- 3Division Urban Plant Ecophysiology, Humboldt-Universität zu Berlin, Berlin, Germany
Herbal preparations of willow bark (Salix cortex) are available in many countries as non-prescription medicines for pain and inflammation, and also as dietary supplements. Currently only little information on toxicity and drug interaction potential of the extracts is available. This study now evaluated the effects of two Salix cortex extracts on human hepatocyte-like HepaRG cells, in view of clinically relevant CYP450 enzyme activity modulation, cytotoxicity and production of reactive oxygen species (ROS). Drug metabolism via the CYP450 enzyme system is considered an important parameter for the occurrence of drug-drug interactions, which can lead to toxicity, decreased pharmacological activity, and adverse drug reactions. We evaluated two different bark extracts standardized to 10 mg/ml phenolic content. Herein, extract S6 (S. pentandra, containing 8.15 mg/ml total salicylates and 0.08 mg/ml salicin) and extract B (industrial reference, containing 5.35 mg/ml total salicylates and 2.26 mg/ml salicin) were tested. Both Salix cortex extracts showed no relevant reduction in cell viability or increase in ROS production in hepatocyte-like HepaRG cells. However, they reduced CYP1A2 and CYP3A4 enzyme activity after 48 h at ≥25 μg/ml, this was statistically significant only for S6. CYP2C19 activity inhibition (0.5 h) was also observed at ≥25 μg/ml, mRNA expression inhibition by 48 h treatment with S6 at 25 μg/ml. In conclusion, at higher concentrations, the tested Salix cortex extracts showed a drug interaction potential, but with different potency. Given the high prevalence of polypharmacy, particularly in the elderly with chronic pain, further systematic studies of Salix species of medical interest should be conducted in the future to more accurately determine the risk of potential drug interactions.
Introduction
The bark of various willow (Salix cortex) varieties has long been used in medicine against inflammation, as a painkiller and against fever (Wood, 2015). Recently, our group demonstrated the anti-inflammatory potential of Salix cortex extracts in SARS-CoV-2 peptide and bacterial lipopolysaccharide (LPS)-activated human in vitro systems, providing new evidence of the potential usefulness of Salix products as therapeutics (Le et al., 2021). Salix cortex is also used in dietary supplements, e.g., for weight reduction and to enhance performance in sports (Matyjaszczyk and Schumann, 2018). Despite its long history of use, so far only little data is available on the toxicity or potential adverse effects of Salix cortex preparations (Shara and Stohs, 2015). Based on a limited number of studies, one safety report by Shara and Stohs (2015) recommended that people who 1) are allergic to aspirin, 2) suffer from pathological conditions such as gastritis, stomach ulcers, diabetes, asthma, or hemophilia; or 3) are under anticoagulant-drug therapy, as well as beta-blockers, diuretics, and non-steroidal anti-inflammatory drugs (NSAIDs), may avoid Salix cortex.
Herbal preparations including Salix cortex extracts contain hundreds of phytochemicals that can act in different ways, encompassing the risk of drug interactions that is the ability to modify the action or effect of another drug administered successively or simultaneously. Considering that the usage of pain medication plays a major role in polypharmacy (Marengoni et al., 2014; Schneider et al., 2021; Young et al., 2021), knowledge of the interaction potential of Salix species is of great importance, as this could help avoid drug-related problems that could affect patients’ safety. An important determinant in the occurrence of drug interaction is the drug metabolism via the cytochrome P450 (CYP450) system (Guengerich, 2008). This class, which is predominantly expressed in the liver, has more than 50 enzymes, but from these, only six of them (CYP1A2, CYP2C9, CYP2C19, CYP2D6, CYP3A4, and CYP3A5) metabolize 90 percent of the medications (Wilkinson, 2005). Considering the historical relevance of Salix cortex extracts in traditional medicine, their wide commercial availability, as well as their potential new pharmacological application, in the present study we investigated the potential of Salix cortex extracts for drug-drug interactions with respect to CYP450 enzymes relevant to drug metabolism. For this purpose, we used the human hepatocyte-like cell line HepaRG. As a validated in vitro model to investigate drug effects on metabolism enzymes, the HepaRG cell line is considered an alternative to primary ex vivo cultured human hepatocytes, especially in studies related to detoxification metabolism, such as CYP450 enzyme activities for predicting drug-drug interaction (Aninat et al., 2006; Anthérieu et al., 2010). Potential cytotoxicity was then assessed by measuring adenosine triphosphate (ATP) and lactate dehydrogenase (LDH), and oxygen radical formation was measured by electron magnetic resonance spectroscopy (EPR) in the cells.
Materials and Methods
Chemicals and Reagents
Fetal calf serum (FCS), L-glutamine and phosphate buffered saline (PBS, without Ca and Mg), penicillin-streptomycin (P/S) solution, L-glutamine solution, RPMI-1640, DMEM, William’s Medium E + GlutaMAX™, human recombinant insulin zinc solution (4 mg/ml), trypsin-EDTA 10x (5 mg/ml and 2.2 mg/ml), Trypsin (0.5%) solution and phosphate buffered saline (PBS, without Ca2+ and Mg2+), and penicillin/streptomycin solution (10,000 U/ml and 10,000 μg/ml) were purchased from Gibco™, Life Technologies GmbH (Darmstadt, Germany). Hydrocortisone 21-hemisuccinate sodium salt, omeprazole (≥99.0%), rifampicin (≥97%), ketoconazole (99%), naringenin (98%), menadione (≥98.0%), and acetylsalicylic acid (100%) were purchased from Sigma-Aldrich (Taufkirchen, Germany). Troglitazone (≥98%) was purchased from Santa Cruz (Heidelberg, Germany). 1-hydroxy-3-methoxy-carbonyl-2,2,5,5-tetramethylpyrolidine hydrochloride (CMH), diethyldithio-carbamate trihydrate (DETC), deferoxamine (DFO), and Krebs-HEPES buffer were purchased from Noxygen Science Transfer & Diagnostics GmbH, Elzach, Germany).
Cell Culture
The human hepatic cell line HepaRG was obtained from Biopredic International® (Rennes, France). The cell line was cultured in William’s Medium E + GlutaMAX™, supplemented with 10% FCS, 100 U/mlµ penicillin, and 100 μg/ml streptomycin, 50 µM hydrocortisone 21-hemisuccinate sodium salt, and 5 μg/ml human insulin. The maintenance and differentiation of the cell line was performed according to Biopredic International® instructions, as previously described (Le et al., 2021). Cells were maintained at 37°C in a humidified incubator with a 5% CO2 and 95% air atmosphere.
Salix Cortex Extract Preparations
Salix cortex extracts were prepared and standardized as previously described (Le et al., 2021). S. pentandra clone PE1 (extract S6), originally collected in 2006 in Eggersdorf (Brandenburg, Germany), was cultivated in Wriezen (in northeastern Berlin, Brandenburg, Germany). One-year-old branches of the clone were cut off in August 2016 and bark was peeled at a height from 10–100 cm. Afterwards, the bark material was frozen (−80°C) and immediately lyophilized. Hardwood cuttings of PE1 were also planted in a clone collection at Humboldt-Universität zu Berlin (Germany) to guarantee the availability and conservation of the Salix clone. The bark was extracted using a solution of 70% methanol and 0.1% formic acid. Extract B refers to a willow bark reference used for phytopharmaceutical production, which was provided from Bionorica SE (Neumarkt, Germany). Both extracts (B and S6) were standardized to 10 mg/ml phenolic content using high performance liquid chromatography (HPLC). Based on the reported pharmacological potential and knowledge of characteristic compounds in different Salix species, the following phytochemicals were used to standardize the extracts: salicylates (salicin, salicortin, 2′-O-acetylsalicin, 2′-O-acetylsalicortin, and tremulacin), flavan-3-ols (catechin and epicatechin), flavonoids (two isomers of naringenin-5-O-glucoside, naringenin-7-O-glucoside, luteolin-7-O-glucoside, quercetin-hexoside, and isosalipurposide), other phenolic compounds (triandrin, two caffeic acid derivatives, and syrengin). S6 extract contained 8.15 mg/ml total salicylates and 0.08 mg/ml salicin, and extract B contained 5.35 mg/ml total salicylates and 2.26 mg/ml salicin (Le et al., 2021).
Assessment of Cell Viability and Cytotoxicity
The enzyme lactate dehydrogenase (LDH) is involved in energy production and found in almost all cells of the human body. Upon damage, it is released from the cell in the medium and can thus be used as a marker for cytotoxicity. After 24 h extract exposure, LDH was quantified using an LDH-Glo™ Cytotoxicity Assay kit (Promega GmbH, Mannheim, Germany) according to the manufacturer’s instructions. As positive control, cell exposure to 0.2% triton-X for 15 min was used.
Adenosine triphosphate (ATP) is a key indicator of cellular activity and has been used as another marker of cytotoxicity upon 24 and 48 h treatment using the CellTiter-Glo® 2.0 Cell Viability Assay (Promega GmbH, Mannheim, Germany) according to the manufacturer’s instructions. As positive control, cell exposure to 0.2% triton-X for 24 or 48 h was used. In both assays, 0.5% distilled water was used as solvent control (SC).
Assessment of Reactive Oxygen Species (ROS) Production Using EPR
The production of reactive oxygen species (ROS) by Salix cortex extracts in hepatocyte-like HepaRG cells was detected using electron paramagnetic resonance (EPR) spectroscopy as described by Lamy et al. (2013) and adapted by Odongo et al. (2017). Differentiated HepaRG cells were treated with different concentrations of Salix cortex extracts or 0.5% distilled water (solvent control) for 1 or 24 h. Cell exposure to 200 µM menadione for 30 min was used as positive control. Afterwards, for ROS detection, 200 µM 1-hydroxy-3- methoxy-carbonyl-2,2,5,5-tetramethylpyrolidine hydrochloride (CMH, Noxygen Science Transfer & Diagnostics GmbH, Elzach, Germany), 25 µM deferoxamine (DFO), and 5 µM DETC were used in Krebs-HEPES buffer for 30 min (Odongo et al., 2017). Supernatants were then measured by EPR spectroscopy for ROS production evaluation. The instrument setting and the number of scans used were set as previously described (Lamy et al., 2013).
Cytochrome P450 Enzyme Activity Quantification
The effects of Salix cortex extracts on CYP1A2 and CYP3A4 enzyme activity were evaluated at 1 and 48 h treatment (Bernasconi et al., 2019; FDA, 2020) using the cell based P450-Glo™ Induction/Inhibition Assay Systems (Promega, Walldorf, Germany) according to the manufacturer’s protocol. In brief, HepaRG cells were differentiated as described above in white 96 well plates. After differentiation, cells were incubated for 1 or 48 h with the Salix cortex extracts, or 0.5% distilled water (solvent control). For 48 h treatment, the medium was exchanged after 24 h with the addition of fresh extract. To analyze enzyme activity of CYP2C19, the biochemical P450-Glo™ CYP2C19 Assay and Screening System (Promega, Walldorf, Germany) was used according to the manufacture’s protocol.
50 µM omeprazole (induction) and 320 µM naringenin (inhibition) were used as positive controls (PC) in the CYP1A2 assay.10 µM Rifampicin (induction) and 10 µM ketoconazole (inhibition) were used as positive control in the CYP3A4 assay. 10 μg/ml (22.6 µM) troglitazone was used as positive control for CYP2C19 inhibition (Wishart et al., 2018).
Quantitative PCR for CYP450 mRNA Expression
CYP2C19 mRNA expression was quantified using qRT-PCR. In brief, differentiated HepaRG cells were treated with different concentrations of Salix cortex extracts or 0.5% distilled water (solvent control) for 6 or 48 h. Total RNA from HepaRG cells was isolated using the RNeasy mini Isolation kit from Qiagen (Hilden, Germany) followed by DNA purification step using the RNase-free DNase kit from Qiagen (Hilden, Germany) according to the manufacturer’s instructions. RNA quality and quantity were measured using a NanoDrop ND-1000 spectrophotometer (Thermo Scientific, Freiburg, Germany). Isolated RNA was resuspended in 10 µL of RNAse-free water. Each sample was treated twice with 2 µL RNAse-free DNAse 1unit/μL (Qiagen, Hilden, Germany) for 10 min at 37°C to eliminate remaining DNA. The prepared RNA was reverse-transcribed as previously described (Helmig et al., 2009). For quantitative comparison of CYP2B6, CYP2C19 and CYP2D6 mRNA levels real-time PCR was performed using SYBR-green fluorescence in a LightCycler® System (Roche Diagnostic GmbH). After optimization of PCR conditions, amplification efficiency was tested in standard curves using serial cDNA dilutions. The correlation coefficient had to be above 0.9 and the slope around −3.5. Amplification specificity was checked using melting curves. Gene expression was related to the mean expression of the three housekeeping genes (HSK) beta-2-microglobulin (B2M), hypoxanthine-guanine phosphoribosyltransferase (HPRT) and glycerinaldehyd-3-phosphat-dehydrogenase (GAPDH) (Vandesompele et al., 2002). Calculations of expression was performed with the 2−ΔΔCT method (Pfaffl, 2001). The sequences of the used specific primers are listed in Table 1 (Zhang et al., 2005; Chen et al., 2014).
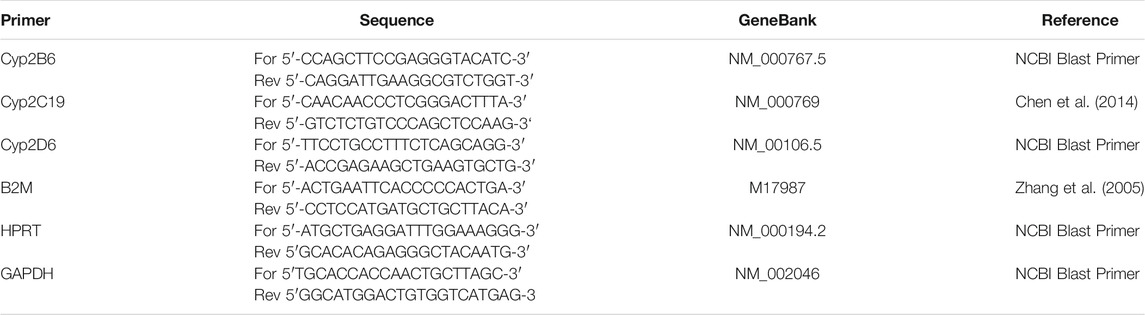
TABLE 1. Sequences of specific primers.
PCR reactions were carried out in a final volume of 20 µl using 1x ABsolute® QPCR SYBR Green Capillary Mixes (Abgene, Brumath, France), 300 nM of primers and 2 µL cDNA. The PCRs started with a 15 min denaturation phase and at the end, a melting curve was acquired form 40°C to 95°C at a thermal transition rate of 0.1°C for 40 s. Specific conditions for primers were as follows: CYP2B6 45 cycles of 95°C for 10 s, 58°C for 15 s, 72°C for 15 s; CYP2C19 45 cycles of 95°C for 15 s, 61°C for 15 s, 72°C for 20 s; CYP2D6 45 cycles of 95°C for 10 s, 61°C for 15 s, 72°C for 15 s. The PCR conditions for HSKs were as follows: B2M 55 cycles of 95°C for 10 s, 63°C for 10 s, 72°C for 25 s; HPRT 45 cycles of 95°C for 15 s, 61°C for 15 s, 72°C for 15 s; GAPDH 45 cycles of 95°C for 10 s, 61°C for 10 s, 72°C for 25 s. All measurements were made without information about the origin of the samples and were performed in duplicate.
Statistical Analysis
Data were analyzed using GraphPad Prism 6.0 software (La Jolla, CA, United States) and presented as means + SD of at least three independent experiments. When comparing multiple means, the results were analyzed either by one-way ANOVA followed by Dunnett’s multiple comparison tests or two-way ANOVA followed by Tukey’s multiple comparison test.
Results
CYP450 Enzyme Activity Quantification
The experiments tested a concentration range of the extracts that had previously shown pharmacological activity in terms of blocking LPS-induced inflammation in primary human immune cells (Le et al. 2021). To investigate potential drug interaction, CYP450 enzyme activity of three enzymes (CYP1A2, CYP3A4 and CYP2C19) was quantified upon Salix cortex extract exposure. As shown in Figure 1A, narigenin (PC1) reduced CYP1A2 enzyme activity in hepatocyte-like HepaRG cells after 1 h by 70%. Omeprazole (PC2) increased the CYP1A2 enzyme activity after 1 h by more that 2-fold (1.6 at 48 h, Figures 1A,B). The Salix cortex extracts did not affect cellular enzyme activity at that time. After 48 h treatment, both extracts reduced the enzyme activity at high concentrations (25 or 50 μg/ml), while the effect was more pronounced by S6, then. Ketoconazole (PC3) completely abolished CYP3A4 activity after 1 h exposure of HepaRG cells, while rifampicin (PC4) triggered enzyme activity induction by about 16-fold compare to control after 48 h (Figures 1C,D). Acetylsalicylic acid (ASA) did not affect CYP1A2 and CYP3A4 enzyme activity after 1 or 48 h at the tested concentrations (Figures 1A–D). For assessment of CYP2C19 enzyme activity, a cell-free assay was used. After 0.5 h, troglitazone (PC5) reduced CYP2C19 activity by 68%; at ≥25 μg/ml both Salix cortex extracts also reduced enzyme activity by 81% (extract S6) and 31% (extract B) compared to solvent control. Again, the inhibitory effect of S6 on CYP450 enzyme activity was stronger compared to extract B.
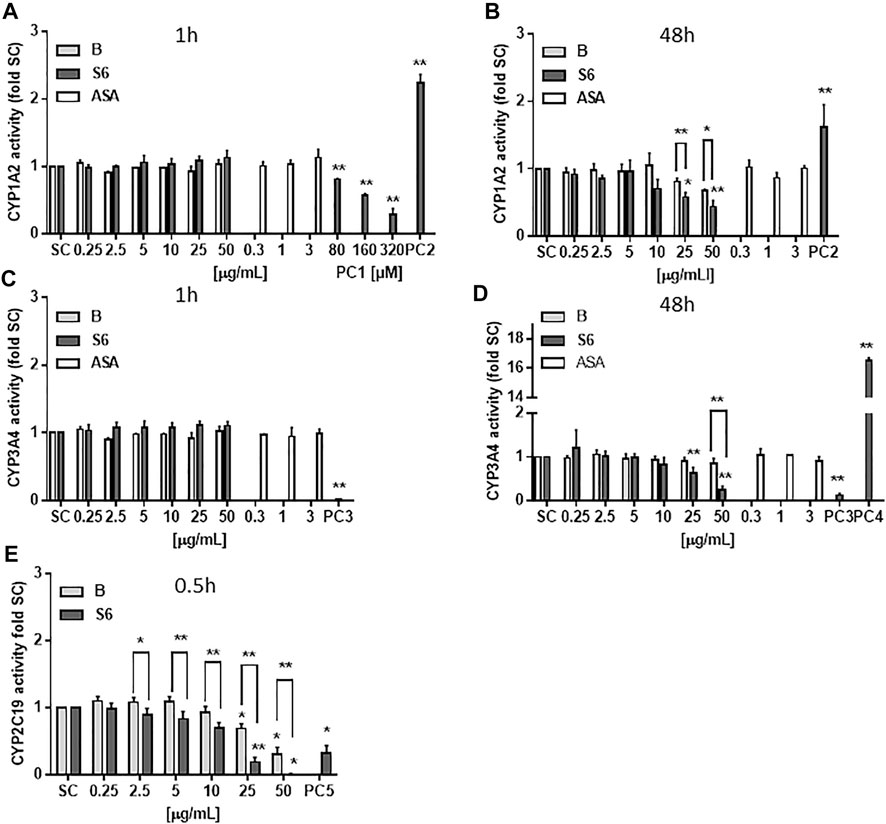
FIGURE 1. CYP450 enzyme activity quantification after treatment with Salix cortex extracts using a luminescent method. (A–D) Differentiated HepaRG cells were exposed to extracts for 1 or 48 h before analysis. (E) CYP2C19 enzyme activity was analysed in a cell-free assay after 0.5 h incubation of extracts with a human recombinant CYP2C19 enzyme, followed by analysis. Positive control (PC): 320 μM naringenin (CYP1A2 inhibition, PC1), 50 μM omeprazole (CYP1A2 induction, PC2), 10 μM rifampicin (CYP3A4 inhibition, PC3), 10 μM ketoconazole, (CYP3A4 induction, PC4), and 22.6 μM troglitazone (CYP2C19 inhibition, PC5). ASA, acetylsalicylic acid. The values are presented as means + SD (CYP1A2 1 and 48 h, n = 3; CYP3A4 1 h, n = 3, 48 h n = 4; CYP2C19 n = 3). Ordinary one-way ANOVA was used for statistical analysis, followed by a Dunnett test. Significance was evaluated between extracts and solvent control (a. d.) as well as between extract S6 and B. *p > 0.05, **p > 0.01.
CYP2C19 mRNA Expression
Differentiated HepaRG cells were exposed for 6 and 48 h to Salix cortex extracts and mRNA expression of CYP2D6, CYP2B6 and CYP2C19 quantified using qRT-PCR (Figure 2). The baseline mRNA levels of CYP2D6 and CYP2B6 were very low in HepaRG cells and no mRNA expression upon treatment could be seen (data not shown). Baseline CYP2C19 mRNA levels were not reduced after 6 h treatment with Salix cortex extracts. After 48 h treatment with 25 μg/ml extract S6, but not B significantly reduced CYP2C19 mRNA expression by 55%.
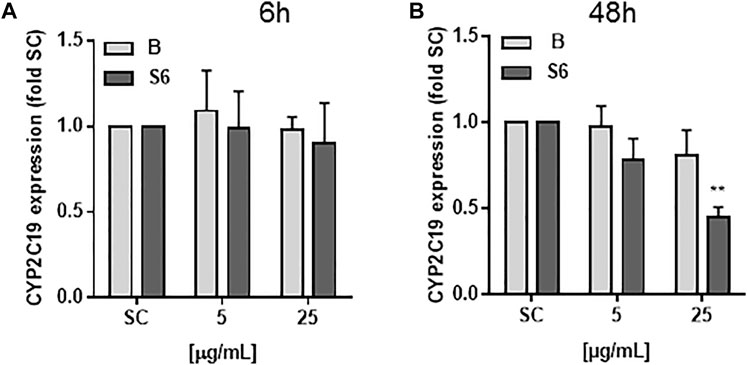
FIGURE 2. mRNA levels of CYP2C19 in differentiated HepaRG cells after Salix cortex extracts treatment. mRNA levels were quantified by qRT-PCR. Differentiated HepaRG cells were treated with Salix cortex extracts (extract B or S6) by 6 h (A) or 48 h (B). CYP2C19 expression levels were expressed as mean + SD; (n = 3; S6, 48 h n = 2). Ordinary one-way ANOVA was used for statistical analysis, followed by a Dunnett test versus solvent control (SC: 0.5% destilled water) group. **p < 0.001.
Cytotoxicity and ROS Production
As given in Figures 3A–C, neither of the two extracts affected intracellular ATP levels or triggered LDH release in hepatocyte-like HepaRG cells at the tested concentrations (0.25–50 μg/ml). We also tested whether the extracts could elevate the level of intracellular ROS in the cells, which in turn could cause damage to lipids, proteins and DNA. From Figures 3D,E it can be seen that after treatment with Salix cortex extracts for 1 or 48 h, no increase in ROS production could be detected.
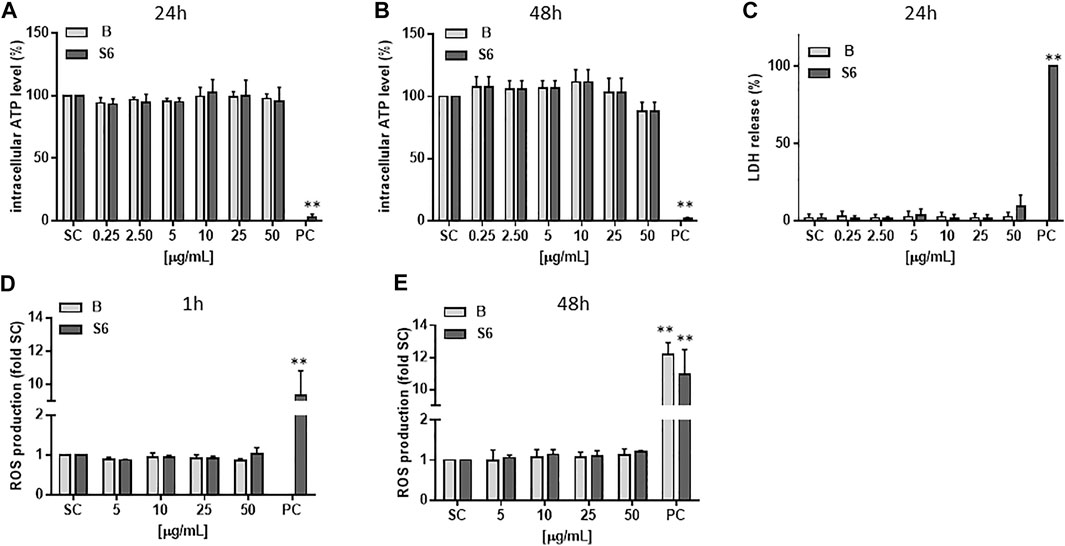
FIGURE 3. Cytotoxicity and ROS production of Salix cortex extracts in differentiated HepaRG cells. ATP levels were analysed after (A) 24 h (B) 48 h of extract treatment. n = 3 (C) LDH cell release was measured after 24 h extract treatment. 0.2% Triton-X was used as positive control (PC). n = 4 (D, E) ROS production was measured by EPR spectroscopy after (D) 1 h or (E) 24 h of extract treatment. 200 μM menadione for 30 min. was used as positive control (PC). n = 3. The values are presented as means + SD. Ordinary one-way ANOVA was used for statistical analysis, followed by a Dunnett test versus solvent control (SC: 0.5% destilled water) group. **p < 0.01.
Discussion
The biggest consumers of prescription and over-the-counter medicines are older adults (Qato et al., 2008; Qato et al., 2016), and self-medication (Vacas Rodilla et al., 2009; Jerez-Roig et al., 2014) as well as consumption of non-prescription medicines, herbal and other dietary supplements in the first place, is widespread among them (Izzo and Ernst, 2009; de Souza Silva et al., 2014; Raji et al., 2017). Conditions of chronic pain, or other chronic conditions, such as diabetes, heart disease, stroke, or cancer, may experience concurrent use of multiple medications (Lunsky and Modi, 2018; Schneider et al., 2021). With the number of drugs, the risk of drug interactions increases exponentially while many drug interactions can be explained by changes in metabolic enzymes in the liver and other extrahepatic tissues. Interaction with hepatic CYP450 enzymes in terms of induction or inhibition is here one of the most important causes after co-administration of medications (Cadieux, 1989; Johnell and Klarin, 2007). CYP450 enzyme induction usually leads to accelerated biotransformation of a drug. For most drugs, accelerated metabolism results in decreased efficacy, but if a pro-drug is activated by CYP450 enzymes, its efficacy and/or toxicity may increase. When two drugs compete over the same enzyme receptor site, an enzyme inhibition occurs. The stronger inhibitor predominates, resulting in decreased metabolism of the competing drug. This can result in increased serum levels of the unmetabolized drug and thus greater potential for toxicity. For drugs whose pharmacological activity requires biotransformation from a pro-drug form, inhibition may result in decreased efficacy. Besides substrate competition, a drug can also reduce enzyme activity due to direct interaction or mRNA inhibition (Lee et al., 2009).
The drug interaction experiments reported in the present study were carried out using hepatocyte-like HepaRG cells. Excluding CYP2A6 and CYP2E1, the HepaRG cell line has been reported to express high functional levels of most of the major xenobiotic metabolizing CYP450 enzymes. These activities were then found to be inhibited and induced by prototypical compounds at comparable levels to primary hepatocytes (Turpeinen et al., 2009). With these characteristics, HepaRG cells have been proposed to be a more physiologically relevant pre-clinical platform for drug–drug interaction studies and safety pharmacology compared to e.g., the pre-clinically widely used cell line HepG2. Even though HepG2 cells are inexpensive and convenient, they lack a substantial set of liver-specific functions, particularly CYP450 activity (Gómez-Lechón et al., 2010). So far, there are only few reports on Salix cortex extracts investigating a CYP450 interaction potential. Using a cell-free fluorimetric in vitro assay, an ethanolic extract of Salix planifolia was found to inhibit CYP2C8 (60.9%), CYP2C19 (48.5%), CYP3A4 (92.3%), CYP3A5 (73.9%), and CYP3A7 (71.4%) at 10 μg/ml concentration. All other investigated enzymes were inhibited by less than 30.0%, which includes CYP1A2 (Tam et al., 2009). In HepaRG cells, we observed a low CYP1A2 and CYP3A4 interference potential of the tested Salix cortex extracts at a concentration which was about 5-fold higher as compared to an effective anti-inflammatory concentration reported earlier by us (Le et al., 2021). This effect was evident only after 48 h, which argues against a direct CYP enzyme activity interaction potential. CYP2C19 metabolizes important drugs in clinical practice, such as proton pump inhibitors (esomeprazole, lansoprazole, omeprazole, pantoprazole, rabeprazole), clopidogrel, tamoxifen, diazepam, citalopram, or escitalopram (Sienkiewicz-Oleszkiewicz and Wiela-Hojeńska, 2018). For this enzyme, our data suggest that both Salix cortex extracts have the potential to interfere with drug metabolism, as they both reduced CYP2C19 enzyme activity in a concentration-dependent manner after 30 min incubation. As with the other enzymes investigated, extract S6 was more potent in enzyme inhibition than extract B. On mRNA level, only S6 significantly reduced CYP2C19 expression in HepaRG cells. It is certain that none of the observed effects on CYP450 enzymes can be attributed to cytotoxic effects, since there was no reduction in ATP levels, increase in LDH or ROS production upon Salix cortex extract treatment in HepaRG cells. The two extracts differed in their salicylate content, which might account for the observed differences, but information on CYP450 regulation by e.g., acetylsalicortin or acetylsalicin, which were both present solely in extract S6, does not exist so far. In contrast to extract B, extract S6 also contained the flavonoids catechin (0.78 mg/ml) and epicatechin (0.03 mg/ml) (Le et al., 2021). For both compounds no relevant inhibition of CYP1A2, CYP2C9, CYP2D6, and CYP3A4 could be detected in a study on human liver microsomes (Satoh et al., 2016), which confirmed previous data (Muto et al., 2001). Thus, it is unlikely that the presence of these flavonoids add to the observed effects. For the aglycone of quercetin, some weak CYP450 activity inhibition has been described (Mohos et al., 2020). Quercetin-hexoside (but not the aglycone) is present in S6 at a 3-fold higher concentration compared to extract B. In contrast, extract B contains some O-glucosides of naringenin. For the aglycone CYP1A2 inhibition has been reported by Fuhr et al. (1993) and this was confirmed in the present study (20% at 80 µM) (Fuhr et al., 1993). However, extract B contained naringenin glucosides only at about 8 µM in total. Even if the glucosides were as potent as the aglycone of naringenin, this concentration would have been too low to inhibit CYP1A2. Taken together, at present, too little information is available to explain the observed differences between the extracts or to attribute the effects to individual extract constituents. Both salicylates and flavonoids as well as other phenolic compounds, such as syrengin, or yet unidentified compounds in the extracts and possible additivity between the compounds need to be investigated with respect to CYP450 inhibition and their role further elucidated in the future.
From the about 450 Salix species which are known (Lauron-Moreau et al., 2015), only few of them are of medical interest so far according to the guidelines of EMA and the United States Pharmacopeia (EMA, 2017; Oketch-Rabah et al., 2019). However, Salix species show huge differences in their phytochemical content, depending on the genotype (Förster et al., 2008; Förster et al., 2010; Gawlik-Dziki et al., 2014; Gligorić et al., 2019), and also other factors such as the plant part used as a source material for medical products (Gawlik-Dziki et al., 2014; Sugier et al., 2013). This currently complicates a reliable therapeutic efficacy of the product. Based on the present data, it also calls for further systematic and more detailed studies on possible drug interactions. For the moment, a potential interaction with drugs that are metabolized by CYP2C19, CYP1A2, as well as CYP3A4 with Salix cortex containing formulations cannot be excluded in common dosages. Although after oral intake, the amount of most phytochemicals in Salix species that becomes accessible for absorption through the epithelial layer of the gastrointestinal tract is currently not known (Schmid et al., 2001), it must be considered that e.g., CYP3A4 is not only the most abundant CYP in the liver but also the wall of the small intestine. There, before absorption into the blood stream occurs, it plays a major role in the metabolism of many different drugs such as calcium channel blocker, lovastatin or diazepam (Vuppalanchi and Saxena, 2011), which either limits or increases the amount of bioavailable active drug. Especially people that have an inherent risk of polypharmacy and consider long-term use of Salix products (ESCOP, 2017) should be aware of this.
Data Availability Statement
The original contributions presented in the study are included in the article, further inquiries can be directed to the corresponding author.
Author Contributions
EL conceived and designed the study and experiments. JG, CH, and SH designed and carried out the experiments. CH and JG prepared the graphs and analyzed the data. JG, EL, CH, and SH wrote the paper. NF and IM prepared the Salix extracts and performed chemical analysis of the extracts. All authors have given approval to the final version of the manuscript.
Funding
This project was funded by the Federal Ministry of Education and Research (BMBF), grant numbers: 031B0349A, B. The article processing charge was partly funded by the Baden-Wuerttemberg Ministry of Science, Research and Art and the University of Freiburg in the funding program Open Access Publishing.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
The authors are grateful to Monika Phillip for experimental support. The authors would also like to thank Christiane Guguen-Guillouzo, Phillipe Gripon, and Christian Trepo for the opportunity to use the HepaRG cell line in our study. We further would like to acknowledge Bionorica SE (Neumarkt, Germany) for providing plant extract B.
References
Aninat, C., Piton, A., Glaise, D., Le Charpentier, T., Langouët, S., Morel, F., et al. (2006). Expression of Cytochromes P450, Conjugating Enzymes and Nuclear Receptors in Human Hepatoma HepaRG Cells. Drug Metab. Dispos. 34, 75–83. doi:10.1124/dmd.105.006759
Anthérieu, S., Chesné, C., Li, R., Camus, S., Lahoz, A., Picazo, L., et al. (2010). Stable Expression, Activity, and Inducibility of Cytochromes P450 in Differentiated HepaRG Cells. Drug Metab. Dispos. 38, 516–525. doi:10.1124/dmd.109.030197
Bernasconi, C., Pelkonen, O., Andersson, T. B., Strickland, J., Wilk-Zasadna, I., Asturiol, D., et al. (2019). Validation of In Vitro Methods for Human Cytochrome P450 Enzyme Induction: Outcome of a Multi-Laboratory Study. Toxicol. Vitro 60, 212–228. doi:10.1016/j.tiv.2019.05.019
Cadieux, R. J. (1989). Drug Interactions in the Elderly. How Multiple Drug Use Increases Risk Exponentially. Postgrad. Med. 86, 179–186. doi:10.1080/00325481.1989.11704506
Chen, H., Shen, Z. Y., Xu, W., Fan, T. Y., Li, J., Lu, Y. F., et al. (2014). Expression of P450 and Nuclear Receptors in normal and End-Stage Chinese Livers. World J. Gastroenterol. 20, 8681–8690. doi:10.3748/wjg.v20.i26.8681
de Souza Silva, J. E., Santos Souza, C. A., da Silva, T. B., Gomes, I. A., Brito, Gde. C., de Souza Araújo, A. A., et al. (2014). Use of Herbal Medicines by Elderly Patients: A Systematic Review. Arch. Gerontol. Geriatr. 59, 227–233. doi:10.1016/j.archger.2014.06.002
EMA (2017). Assessment Report on Salix [various Species Including S. Purpurea L., S. Daphnoides Vill., S. Fragilis L.], Cortex. European Medicines Agency, Committee on Herbal Medicinal Products (HMPC). Avaiable at: https://www.ema.europa.eu/en/documents/herbal-report/final-assessment-report-salix-various-species-including-s-purpurea-l-s-daphnoides-vill-s-fragilis-l_en.pdf.
ESCOP (2017). Salicis Cortex, Willow Bark. European Scientific Cooperative on Phytotherapy. Online Series. Available at: https://escop.com/wp-content/uploads/edd/2017/09/Salix.pdf.
FDA (2020). Vitro Drug Interaction Studies — Cytochrome P450 Enzyme- and Transporter-Mediated Drug Interactions Guidance for Industry. Center for Drug Evaluation and Research (CDER), Food and Drug Administration. Available at: https://www.fda.gov/regulatory-information/search-fda-guidance-documents/vitro-drug-interaction-studies-cytochrome-p450-enzyme-and-transporter-mediated-drug-interactions.
Förster, N., Ulrichs, C., Zander, M., Kätzel, R., and Mewis, I. (2010). Factors Influencing the Variability of Antioxidative Phenolic Glycosides in Salix Species. J. Agric. Food Chem. 58, 8205–8210. doi:10.1021/jf100887v
Förster, N., Ulrichs, C., Zander, M., Kätzel, R., and Mewis, I. (2008). Influence of the Season on the Salicylate and Phenolic Glycoside Contents in the Bark of Salix Daphnoides, Salix Pentandra, and Salix Purpurea. J. Appl. Bot. Food Qual. 82, 99–102.
Fuhr, U., Klittich, K., and Staib, A. H. (1993). Inhibitory Effect of Grapefruit Juice and its Bitter Principal, Naringenin, on CYP1A2 Dependent Metabolism of Caffeine in Man. Br. J. Clin. Pharmacol. 35, 431–436. doi:10.1111/j.1365-2125.1993.tb04162.x
Gawlik-Dziki, U., Sugier, D., Dziki, D., and Sugier, P. (2014). Bioaccessibility In Vitro of Nutraceuticals from Bark of Selected Salix Species. ScientificWorldJournal 2014, 782763. doi:10.1155/2014/782763
Gligorić, E., Igić, R., Suvajdžić, L., and Grujić-Letić, N. (2019). Species of the Genus Salix L.: Biochemical Screening and Molecular Docking Approach to Potential Acetylcholinesterase Inhibitors. Appl. Sci. 9, 1842. doi:10.3390/app9091842
Gómez-Lechón, M. J., Lahoz, A., Gombau, L., Castell, J. V., and Donato, M. T. (2010). In Vitro evaluation of Potential Hepatotoxicity Induced by Drugs. Curr. Pharm. Des. 16, 1963–1977. doi:10.2174/138161210791208910
Guengerich, F. P. (2008). Cytochrome P450 and Chemical Toxicology. Chem. Res. Toxicol. 21, 70–83. doi:10.1021/tx700079z
Helmig, S., Hadzaad, B., Döhrel, J., and Schneider, J. (2009). Influence of the Cyp1B1 L432V Gene Polymorphism and Exposure to Tobacco Smoke on Cyp1B1 mRNA Expression in Human Leukocytes. Drug Metab. Dispos. 37, 1490–1495. doi:10.1124/dmd.109.027060
Izzo, A. A., and Ernst, E. (2009). Interactions between Herbal Medicines and Prescribed Drugs: an Updated Systematic Review. Drugs 69, 1777–1798. doi:10.2165/11317010-000000000-00000
Jerez-Roig, J., Medeiros, L. F., Silva, V. A., Bezerra, C. L., Cavalcante, L. A., Piuvezam, G., et al. (2014). Prevalence of Self-Medication and Associated Factors in an Elderly Population: a Systematic Review. Drugs Aging 31, 883–896. doi:10.1007/s40266-014-0217-x
Johnell, K., and Klarin, I. (2007). The Relationship between Number of Drugs and Potential Drug-Drug Interactions in the Elderly: a Study of over 600,000 Elderly Patients from the Swedish Prescribed Drug Register. Drug Saf. 30, 911–918. doi:10.2165/00002018-200730100-00009
Lamy, E., Herz, C., Lutz-Bonengel, S., Hertrampf, A., Márton, M. R., and Mersch-Sundermann, V. (2013). The MAPK Pathway Signals Telomerase Modulation in Response to Isothiocyanate-Induced DNA Damage of Human Liver Cancer Cells. PLoS One 8, e53240. doi:10.1371/journal.pone.0053240
Lauron-Moreau, A., Pitre, F. E., Argus, G. W., Labrecque, M., and Brouillet, L. (2015). Correction: Phylogenetic Relationships of American Willows (Salix L., Salicaceae). PLoS One 10, e0138963. doi:10.1371/journal.pone.0138963
Le, N. P. K., Herz, C., Gomes, J. V. D., Förster, N., Antoniadou, K., Mittermeier-Klessinger, V. K., et al. (2021). Comparative Anti-inflammatory Effects of Salix Cortex Extracts and Acetylsalicylic Acid in SARS-CoV-2 Peptide and LPS-Activated Human In Vitro Systems. Int. J. Mol. Sci. 22, 6766. doi:10.3390/ijms22136766
Lee, M. D., Ayanoglu, E., and Gong, L. (2009). Drug-induced Changes in P450 Enzyme Expression at the Gene Expression Level: a New Dimension to the Analysis of Drug-Drug Interactions. Xenobiotica 36, 1013–1080. doi:10.1080/00498250600861785
Lunsky, Y., and Modi, M. (2018). Predictors of Psychotropic Polypharmacy Among Outpatients with Psychiatric Disorders and Intellectual Disability. Psychiatr. Serv. 69, 242–246. doi:10.1176/appi.ps.201700032
Marengoni, A., Pasina, L., Concoreggi, C., Martini, G., Brognoli, F., Nobili, A., et al. (2014). Understanding Adverse Drug Reactions in Older Adults through Drug-Drug Interactions. Eur. J. Intern. Med. 25, 843–846. doi:10.1016/j.ejim.2014.10.001
Matyjaszczyk, E., and Schumann, R. (2018). Risk Assessment of white Willow (Salix alba) in Food. EFSA J. 16, e16081. doi:10.2903/j.efsa.2018.e16081
Mohos, V., Fliszár-Nyúl, E., Ungvári, O., Kuffa, K., Needs, P. W., Kroon, P. A., et al. (2020). Inhibitory Effects of Quercetin and its Main Methyl, Sulfate, and Glucuronic Acid Conjugates on Cytochrome P450 Enzymes, and on OATP, BCRP and MRP2 Transporters. Nutrients 12, 2306. doi:10.3390/nu12082306
Muto, S., Fujita, K., Yamazaki, Y., and Kamataki, T. (2001). Inhibition by green tea Catechins of Metabolic Activation of Procarcinogens by Human Cytochrome P450. Mutat. Res. 479, 197–206. doi:10.1016/S0027-5107(01)00204-4
Odongo, G. A., Schlotz, N., Herz, C., Hanschen, F. S., Baldermann, S., Neugart, S., et al. (2017). The Role of Plant Processing for the Cancer Preventive Potential of Ethiopian Kale (Brassica Carinata). Food Nutr. Res. 61, 1271527. doi:10.1080/16546628.2017.1271527
Oketch-Rabah, H. A., Marles, R. J., Jordan, S. A., and Low Dog, T. (2019). Low Dog TUnited States Pharmacopeia Safety Review of Willow Bark. Planta Med. 85, 1192–1202. doi:10.1055/a-1007-5206
Pfaffl, M. W. (2001). A New Mathematical Model for Relative Quantification in Real-Time RT-PCR. Nucleic Acids Res. 29, e45–2007. doi:10.1093/nar/29.9.e45
Qato, D. M., Alexander, G. C., Conti, R. M., Johnson, M., Schumm, P., and Lindau, S. T. (2008). Use of Prescription and Over-the-counter Medications and Dietary Supplements Among Older Adults in the United States. JAMA 300, 2867–2878. doi:10.1001/jama.2008.892
Qato, D. M., Wilder, J., Schumm, L. P., Gillet, V., and Alexander, G. C. (2016). Changes in Prescription and Over-the-counter Medication and Dietary Supplement Use Among Older Adults in the United States, 2005 vs 2011. JAMA Intern. Med. 176, 473–482. doi:10.1001/jamainternmed.2015.8581
Raji, M. A., Kuo, Y. F., Snih, S. A., Sharaf, B. M., and Loera, J. A. (2017). Ethnic Differences in Herb and Vitamin/Mineral Use in the Elderly. Ann. Pharmacother. 39, 1019–1023. doi:10.1345/aph.1E506
Satoh, T., Fujisawa, H., Nakamura, A., Takahashi, N., and Watanabe, K. (2016). Inhibitory Effects of Eight Green Tea Catechins on Cytochrome P450 1A2, 2C9, 2D6, and 3A4 Activities. J. Pharm. Pharm. Sci. 19, 188–197. doi:10.18433/J3MS5C
Schmid, B., Kötter, I., and Heide, L. (2001). Pharmacokinetics of Salicin after Oral Administration of a Standardised Willow Bark Extract. Eur. J. Clin. Pharmacol. 57, 387–391. doi:10.1007/s002280100325
Schneider, J., Algharably, E. A. E., Budnick, A., Wenzel, A., Dräger, D., and Kreutz, R. (2021). High Prevalence of Multimorbidity and Polypharmacy in Elderly Patients with Chronic Pain Receiving Home Care Are Associated with Multiple Medication-Related Problems. Front. Pharmacol. 12, 686990. doi:10.3389/fphar.2021.686990
Shara, M., and Stohs, S. J. (2015). Efficacy and Safety of White Willow Bark (Salix alba) Extracts. Phytother. Res. 29, 1112–1116. doi:10.1002/ptr.5377
Sienkiewicz-Oleszkiewicz, B., and Wiela-Hojeńska, A. (2018). CYP2C19 Polymorphism in Relation to the Pharmacotherapy Optimization of Commonly Used Drugs. Pharmazie 73, 619–624. doi:10.1691/ph.2018.8689
Sugier, D., Sugier, P., Banas, A., and Szewczuk, C. (2013). The Content of Phenolic Glycosides and Macroelements (K, Ca, Mg) in the Bark of Herbal willows. Acta Sci. Pol. Hortorum Cultus. 12, 31–41.
Tam, T. W., Liu, R., Arnason, J. T., Krantis, A., Staines, W. A., Haddad, P. S., et al. (2009). Actions of Ethnobotanically Selected Cree Anti-diabetic Plants on Human Cytochrome P450 Isoforms and Flavin-Containing Monooxygenase 3. J. Ethnopharmacol. 126, 119–126. doi:10.1016/j.jep.2009.07.036
Turpeinen, M., Tolonen, A., Chesne, C., Guillouzo, A., Uusitalo, J., and Pelkonen, O. (2009). Functional Expression, Inhibition and Induction of CYP Enzymes in HepaRG Cells. Toxicol. Vitro 23, 748–753. doi:10.1016/j.tiv.2009.03.008
Vacas Rodilla, E., Castellà Dagà, I., Sánchez Giralt, M., Pujol Algué, A., Pallarés Comalada, M. C., and Balagué Corbera, M. (2009). Self-medication and the Elderly. The Reality of the home Medicine Cabinet. Aten. Primaria. 41, 269–274. doi:10.1016/j.aprim.2008.09.018
Vandesompele, J., De Preter, K., Pattyn, F., Poppe, B., Van Roy, N., De Paepe, A., et al. (2002). Accurate Normalization of Real-Time Quantitative RT-PCR Data by Geometric Averaging of Multiple Internal Control Genes. Genome Biol. 3, research0034. doi:10.1186/gb-2002-3-7-research0034
Vuppalanchi, R. (2011). “Metabolism of Drugs and Xenobiotics,” in Practical Hepatic Pathology: A Diagnostic Approach. Editor R. Saxena (Philadelphia: Elsevier Saunders), 45–52. doi:10.1016/B978-0-443-06803-4.00004-6
Wilkinson, G. R. (2005). Drug Metabolism and Variability Among Patients in Drug Response. N. Engl. J. Med. 352, 2211–2221. doi:10.1056/NEJMra032424
Wishart, D. S., Feunang, Y. D., Guo, A. C., Lo, E. J., Marcu, A., Grant, J. R., et al. (2018). DrugBank 5.0: a Major Update to the DrugBank Database for 2018. Nucleic Acids Res. 46, D1074–D1082. doi:10.1093/nar/gkx1037
Wood, J. N. (2015). From Plant Extract to Molecular Panacea: a Commentary on Stone (1763) 'An Account of the success of the Bark of the Willow in the Cure of the Agues'. Philos. Trans. R. Soc. Lond. B Biol. Sci. 370, 20140317. doi:10.1098/rstb.2014.0317
Young, E. H., Pan, S., Yap, A. G., Reveles, K. R., and Bhakta, K. (2021). Polypharmacy Prevalence in Older Adults Seen in United States Physician Offices from 2009 to 2016. PLoS One 16, e0255642. doi:10.1371/journal.pone.0255642
Keywords: Salix species, willow bark, CYP450 enzymes, drug interaction, herb-drug interaction
Citation: Gomes JVD, Herz C, Helmig S, Förster N, Mewis I and Lamy E (2021) Drug-Drug Interaction Potential, Cytotoxicity, and Reactive Oxygen Species Production of Salix Cortex Extracts Using Human Hepatocyte-Like HepaRG Cells. Front. Pharmacol. 12:779801. doi: 10.3389/fphar.2021.779801
Received: 19 September 2021; Accepted: 29 October 2021;
Published: 18 November 2021.
Edited by:
Myrna Deciga Campos, Instituto Politécnico Nacional (IPN), MexicoReviewed by:
Elwira Sieniawska, Medical University of Lublin, PolandGabino Garrido, Catholic University of the North, Chile
Copyright © 2021 Gomes, Herz, Helmig, Förster, Mewis and Lamy. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Evelyn Lamy, ZXZlbHluLmxhbXlAdW5pa2xpbmlrLWZyZWlidXJnLmRl