 Kian Chung Chok
Kian Chung Chok Ming Guan Ng
Ming Guan Ng Khuen Yen Ng
Khuen Yen Ng Rhun Yian Koh3
Rhun Yian Koh3 Soi Moi Chye
Soi Moi Chye
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Pharmacol. , 01 October 2021
Sec. Ethnopharmacology
Volume 12 - 2021 | https://doi.org/10.3389/fphar.2021.746656
This article is part of the Research Topic Edible Birds Nest - Chemical Composition and Potential Health Efficacy and Risks View all 11 articles
Edible bird’s nest (EBN) is a traditional Chinese delicacy made of the saliva of swiftlets found in Southeast Asia. With increasing demands for EBN, quality control of EBN products is important for safe consumption. The processing steps are particularly important for efficient extraction of bioactive compounds. Geographical location, collection place, and harvesting season contribute to differences in nutritional contents in EBN. Concerns regarding presence of adulterant, chemical, and microbial contaminants in EBN as well as authentication and chemical composition measuring methods are discussed in this review. Recent discoveries of beneficial health functions of EBN in antimicrobial and antiviral actions, immunomodulation, cancer prevention and treatment, tissue regeneration, cardiometabolic maintenance, antioxidant action and neuroprotection are also reviewed. Our review provides an update on the recent research on EBN.
Consumption of edible bird’s nest could be traced back to the 7th century from China, in which EBN was classified as a high-grade health food, tonic, medicine, and a symbol of wealth (Lau and Melville, 1994). Traditional Chinese medicine states that EBN confers health benefits such as moisturizing the lung, resolving phlegm, alleviating cough, and recuperating from diseases (Lim and Earl of Cranbrook, 2002; Hobbs, 2004). EBN is usually referred to the nests of Aerodramus fuciphagus (A. fuciphagus) (white-nest swiftlet) and Aerodramus maximus (A. maximum) (black-nest swiftlet) harvested for consumption in Southeast Asia (Hao et al., 2015). The growth and reproduction of swiftlets require appropriate environmental conditions such as humidity of about 80–90%, temperature between 26 and 35°C and sufficient food sources. Hence, EBN-producing swiftlets are mainly found in Southeast Asian countries such as Malaysia, Thailand, Indonesia, Myanmar, Vietnam, The Philippines, Cambodia, and the southern region of China. However, the quality, composition, and nutritional values in EBN could be different based on the swiftlet species, geographical as well as environmental conditions (Dai et al., 2020). Increasing demands for EBN has produced economic value from US$1000.00 to US$10,000.00 per kilogram depending on its grade, shape, species, and origin. Indonesia is the largest bird’s nest producer in Southeast Asia, exporting around 2,000 tons/year, followed by Malaysia at 600 tons/year, and Thailand at 400 tons/year (Panyaarvudh, 2018). China is the largest EBN importer every year. From January to June 2021, China imported approximately 128.3 tons, 42.3 tons, and 0.1 tons from Indonesia, Malaysia, and Thailand respectively, in the form of ready-to-drink beverages and processed EBN (Development Report of Imported Bird’s Nest in 2021, 2021). Indonesia exported about US$998 million worth of EBN in 2015 and the value surged up to US$3.64 billion in 2019 (Badan Pusat Statistika, 2020). EBN production in Myanmar is comparatively lower as the industry has just begun a decade ago by building the first EBN house in Bokpyin, southern Myanmar (Big business in bird’s nest in Myanmar seaside town, 2017). In China, there are 3 well-known places with swiftlets inhabitant: Huaiji County, Guangdong province, which produces 150 kg/year; EBN collection is prohibited on Dazhou Island, Hainan because swiftlets are listed as endangered species; EBN produced from Jianshui County, Yunnan is only for the consumption by local residents (Geographical distribution of swiftlet inhabitant, 2020). Due to increasing demands for EBN, quality control should be in place to ensure safe consumption of EBN. Studies that investigated the nutritional value, adulterant content, nitrite (NO2−), nitrate (NO3−), and microbial contamination in EBN have increased in recent decade (Lee et al., 2021). Our review will discuss on EBN extraction methods, nutritional values, health benefits, and the risk of EBN consumption by including the latest findings about EBN. There is an existing review about EBN which includes publications up to 2019 indexed in Web of Science (Lee et al., 2021). Our review aims to provide an update by including those publications which were not discussed in previous reviews and discuss on additional topics including industrial EBN processing based on laboratory findings. This is a narrative review which includes latest EBN publications up to April 2021. Published data were retrieved from PubMed, Semantic Scholar, Meta, X-MOL, and Google Scholar, using search string: edible bird’s nest.
The quality of EBN is based on various factors such as species of swiftlets, habitat (man-made house or natural cave), harvesting season (rainy season or drought season), and geographical location (Malaysia, Thailand, Indonesia, Myanmar, and China). Quality of EBN is usually screened and maintained at the stage of industrial preparation. Despite the challenges and limitations, numerous methods were applied to ensure authenticity, nutritional values, and safety for EBN consumption, ranging from empirical measures to molecular biology-based techniques.
Nutritional and authenticity studies show that EBN acquired from different sources, such as swiftlet premises, natural caves, and retail stores contain significantly varied chemical compositions. The A. fuciphagus EBN from swiftlet premise has higher antioxidant activities and sialic acid content whereas the A. maximus EBN from the cave has more mineral composition of calcium and magnesium. Total amino acids in A. fuciphagus EBN was found to be 23% higher than A. maximus EBN (Quek et al., 2018a). A simple gel electrophoretic method could be employed to differentiate house and cave EBN by generating a fingerprint profile of EBN. The cave EBN produces 10 bands with 2 strong bands at 30 and 35 kDa, house EBN produces 9 bands with 2 strong bands at 120 and 140 kDa (Hun et al., 2016). These results show different protein contents in EBN from different origins and hence there is a need to further investigate this difference in protein composition. Cave EBN was also reported to have 3 times more calcium content than house EBN. It is believed that the moist limestone cave wall provides abundance Ca2+ to seep into the cave EBN whereas the house EBN is built on timber strips that have no Ca2+ to leach out. (Shim and Lee, 2020). In a field study, cave EBN collected from small islands in Indonesia and Malaysia contain higher NO3− and NO2− levels when compared to house EBN. It is probably due to the symbiotic relationship between plants and nitrogen-fixing bacteria providing space (i.e., root nodules) for the growth of nitrogen-fixing bacteria in exchange for NO3− as nutrients. It is speculated that nitrogen-fixing bacteria reaches the mouth of swiftlets when the swiftlets prey on insects or directly from plants or soil (Schultze and Kondorosi, 1998; Chan et al., 2013). The risk of NO3− and NO2− contamination will be discussed later in this review.
Nutritional compositions in EBN are known to be affected by the harvesting season. Swiftlets feed on insects from Hymenoptera (winged ants, fig wasps and bees), Coleoptera (small beetles), Homoptera (leafhoppers) and Ephemenoptera (mayflies) species. The food sources for swiftlets are more abundant during the rainy season. EBN protein content ranged from 60.3 to 63.6 g/100 g for samples from North Peninsular Malaysia and 57.9–61.2 g/100 g from the East Coast Peninsular Malaysia with the highest protein level found in samples collected during December to March. For EBN samples obtained from South Peninsular Malaysia, protein content ranged from 61.8 to 65.2 g/100 g and samples harvested from August to November had the highest protein content. However, the mineral contents found in EBN samples from all different zones are not affected by the season (Norhayati et al., 2010).
Farmed EBN along the seashores of Thailand were reported to be rich in sulphur-containing amino acids, calcium, and magnesium (Saengkrajang et al., 2013). The use of Gas Chromatography-Mass Spectrometry (GC-MS) and Liquid chromatography-Mass Spectrometry (LC-MS) together with orthogonal projection to latent square discriminant analysis (OPLS-DA) could identify the origins of EBNs accurately based on the fatty acid content in the Malaysian, Indonesian, and Thailand EBNs. Generally, the levels of fatty acids and fatty acid amides are highest in Malaysian EBNs followed by Indonesian and then Thai EBNs. Moreover, the Malaysian and Indonesian EBNs could be differentiated with distinct fatty acids profiles in which the fatty acids of Malaysian and Indonesian EBNs are separated along the PC1 axis in the loading plot. (Chua et al., 2014). Thailand has a huge land mass cultivating rice whereas Malaysia and Indonesia have large palm oil plantations (Seeboonruang, 2012; Meijaard et al., 2020). Swiftlets usually feed on insects nearby their habitats, the swiftlets in Thailand feed on insects with a rice-based diet, resulting in low fatty acids profile in Thai EBN. In contrast, the swiftlets in Malaysia and Indonesia feed on insects with a diet rich in palm oil, resulting in high fatty acids content in their EBNs (Lourie and Tompkins, 2000; Zhou et al., 2003; Tres et al., 2013). In the same study by Chua et al., the cave EBN contains higher level of ergosterol, a component of fungi and heptadecasphinganine, antifungal agent when compared to house EBN indicating higher amount of fungi in cave EBN and production of heptadecasphinganine by the swiftlets (Chua et al., 2014).
EBN is seen to be rich in protein, essential amino acids, essential trace elements, and essential sugars for human biological functions. Pure EBN collected from swiftlet premises in Batu Pahat, Malaysia contains carbohydrate (46.47%) (w/w), protein (35.80%) and a very minute fat content (1.30%). EBN contains relatively high amount of sodium (6017 mg/kg), magnesium (344 mg/kg), potassium (138 mg/kg) and calcium (68 mg/kg). Other minerals such as phosphorus, iron (Fe), chromium, and selenium are 0.037, 4.52, 0.30 and 0.14 mg/kg respectively and trace amounts of heavy metals arsenic (As) (0.0237 mg/kg), lead (Pb) (0.0203 mg/kg), copper (Cu) (0.6783 mg/kg) and zinc (1.2542 mg/kg) were found in the EBN. Malaysia Food Act 1983 (Act 281), Part VII (Incidental Constituent), Regulation 38 consider them as heavy metals. However, these heavy metals were within the infant formula specification limit as set by the Act. No mercury (Hg) and cadmium (Cd) were detected in the EBN. Fibre would only be detected in EBN if adulterant containing vegetative matter was added into EBN (Lee et al., 2015). Proximate nutritional compositions were compared between A. fuciphagus EBN from Pahang and Terengganu, Malaysia (Halimi et al., 2014).
Systematic analysis of nutritional composition of EBN collected from different regions in Indonesia was recently conducted. EBN collected from West Sumatra, South Sumatra, West Java, West Kalimantan, Central Sulawesi, and Southeast Sulawesi were analysed. The proximate nutritional compositions are protein (53.09–56.25%), carbohydrate (19.57–23.04%), moisture (17.08–21.50%), ash (5.44–6.25%), and fat (0.07–0.57%). EBN collected from different locations in Indonesia contain 18 types of amino acids, including 10 essential amino acids and 8 non-essential amino acids, ranging between 16.15 and 20.88%. NO2− contents were found to be ranging from 3.11–18.28 ppm (average 8.40 ppm) (Elfita et al., 2020).
EBN from Myanmar was found to contain high protein (53.5–59.3%), sodium (0.17%), potassium (237.1 ppm), calcium (0.71%), magnesium (361.3 ppm), and zinc (12.3 ppm). The NO2− and NO3− levels in EBN were estimated by measuring NO2− and NO3− contents in filtrate water after soaking with EBN for 4 h. The NO2− and NO3− levels in filtrate water are 0.27 and 4.0 ppm respectively (Phyu Win et al., 2020).
In the past, EBN was considered 'Caviar of the East' and was expensive (from US$1000.00 to US$10,000.00 per kilogram).There have been incidents of adulterants added into EBN during the processing to increase the final weight of EBN product. The addition of adulterants possesses risks to the health of EBN consumers as well as economical loss to the consumers when faulty goods were purchased (EBN products containing adulterants). Common adulterants such as tremella fungus (Tremella fuciformisis), jelly, fish swimming bladder, egg white, pork skin, karaya gum (Sterculia urens), and red seaweed could cause unwanted health effects and allergy if consumed unnoticeably (Thomas et al., 2019; Yeo et al., 2021). The relevant government departments and companies are responsible to authenticate and prevent consumers from consuming EBN products containing health-threatening adulterants.
Gel electrophoretic protein fingerprint profiling could be done to differentiate EBN from white fungus, jelly, fish swimming bladder and egg white. Cave EBN (30 and 35 kDa), house EBN (120 and 140 kDa), white fungus (22, 35, and 75 kDa), egg white (35 and 75 kDa), fish swimming bladder (streaking bands), and jelly (no band). The amino acid fingerprint profile of cave EBN, house EBN, white fungus, egg white, fish swimming bladder, and jelly are all distinct from each other and could be identified by HPLC (Hun et al., 2016). Proximate nutritional analysis shows that addition of adulterants, such as karaya gum, red seaweed, and tremella fungus, could account for 2–10% of the final weight of EBN and reduce the crude protein content of EBN up to 6.2% (Marcone, 2005). Authentic EBN contains more than 3 mg epidermal growth factor (EGF) in 1 g of protein as detected using a simple immunoblotting assay (Yang et al., 2014). Guo et al. established a TaqMan-based real-time polymerase chain reaction (PCR) assay to specifically detect EBN components and 4 common adulterants: white fungus, agar, pork skin and egg white. Five sets of primers and probes were designed for these five components. The assays were specific and reproducible even after the samples have undergone vigorous, tedious, and complicated processing. The relative detection sensitivities were 0.5% EBN, 0.001% white fungus, 0.5% agar, 0.001% fried pork skin and 1% egg white (Guo et al., 2014).
Metabolite profiling refers to holistic analysis of the molecular information of food which has recently gained attention in the food industry (Xu et al., 2006). The use of mass spectrometry coupled with chromatography resulted in improved data quality and sensitivity (Chua et al., 2014). The mass spectrometry method was further improved using GC-MS and LC-MS together with the chemometrics model, orthogonal projection to latent square discriminant analysis (OPLS-DA). These methods could classify the EBNs based on production site, colours, and countries. OPLS-DA has the advantage of being able to predict the identity of unknown samples. GC-MS coupled with OPLS-DA was discovered to be a better EBN authentication method for quality control of EBN compared to LC-MS, of which detection of the metabolite could be affected due to EBN processing (Chua et al., 2014). GC-MS was used to detect the oligosaccharides content in EBN, especially the N-Acetylneuraminic acid (NANA). By using environmental scanning electron microscopy, the micrographs of authentic raw EBN samples display the crater-like structure of irregularly shaped three-dimensional networks, measuring from less than one, up to several microns. However, the fake EBN samples made from jelly fungus, agar, and pork skin display granule- or cudgel-like structures rather than network structures. In another study, well-established gas chromatography with flame-ionisation detection (GC-FID) was employed to analyse and quantify amino acids in EBN whereas optimised liquid chromatography with tandem mass spectrometry (LC-MS-MS) was applied for monosaccharide analysis. The data generated were combined with Hotelling T2 range plot to identify EBN and non-EBN. Hotelling T2 range plot is a chemometrics plot that reduces the amino acids and monosaccharides into a T2 value. This plot simplifies data analysis process and, more importantly, combines multiple variables to create a unique fingerprint for the sample (Chua et al., 2015). Its application is often found in the quality control of products (Eppe and De Pauw, 2009). This method has since then discovered EBN to contain a group of glycoproteins that are not affected by the EBN’s colouration, country of origin, and/or the processing method of the food item (Chua et al., 2015).
Samples damaging during authentication of EBN is the major concern in the authentication process. Contamination or loss of chemical composition could result in false reporting if samples were not handled with care. Thermogravitiy (TG) and differential thermogravity (DTG) techniques require 5–10 mg of EBN samples for rapid authentication of EBN by comparing the TG and DTG curves produced from EBN and adulterated EBN. EBN was heated in the crucible from 25°C to 1000°C and 800°C for unadulterated and adulterated samples to generate the TG and DTG curves. TG is presented in the curve of mass percentage versus temperature whereas the first derivative DTG curve gave the rate of change of mass loss percentage versus temperature. Hence, adulterated EBN (e.g. glucose, sucrose, hydrolysed marine collagen, and monosodium glutamate) has distinctive TG and DTG curves which could be identified when compared to TG and DTG curves of pure EBN (Shim et al., 2017). Shi et al. (2017) designed a rapid and non-destructive method to characterise the distribution map of carbohydrates, proteins, and sialic acid in EBN using hyper-spectral imaging. The chemometrics and spectral signals of EBN were used to build calibration models which then were tested by extracting and predicting spectra of each pixel in EBN hyperspectral images. The distribution map of carbohydrates, proteins, and sialic acid in EBN were characterised with unevenly distributed carbohydrate and protein contents based on the swiftlets diet consumption: Insects and small fish consumption increased protein content whereas seaweed consumption increased carbohydrates content. However, sialic acid is evenly distributed in the entire EBN (Shi et al., 2017). Future studies on these methods to identify adulterated EBN and their potential application in industry EBN preparation is promising to be ventured.
Fourier Transform Infrared (FTIR) spectrum of the raw unprocessed EBN was identical to that of the spectra of processed EBN samples. The spectra of adulterated EBN samples were different from that of processed EBN. Fingerprint region of the spectra of pure edible bird and adulterated edible bird nest samples were different at <1700 cm−1. The NH group was absent in all adulterants except in pork skin and egg white. Interestingly, only pork skin has ester C=O stretch bond and C=CH group which were not present in EBN. Hence, this technique could be used for testing the authenticity of the EBN samples as well as detection of porcine products (Hamzah et al., 2013). Apart from that, indirect enzyme-linked immunosorbent assays (ELISAs) was developed to detect porcine gelatine adulteration using anti-peptide polyclonal antibodies. Three indirect ELISAs were developed for porcine species-specific amino acid sequences of the collagen (I) α2 chain (PAB1 and PAB2) and the collagen (I) α1 chain (PAB3), which had limits of detection of 0.12, 0.10 and 0.11 µg/g respectively (Tukiran et al., 2015).
Environmental factors, such as locations, harvesting places, and harvesting seasons could affect the quality of EBN and a standardised guideline is required for quality control of EBN products. Differences in nutritional, adulterants, and contaminants detected could be due to different EBN handling, beginning from collection, processing, delivery, and storage. The EBN industry in Malaysia and Indonesia suffered a severe blow in 2011 when China banned exportation of EBNs due to high concentrations of NO3−, Pb and As in certain products. Various regulations were rolled out in response to the ban (Yeo et al., 2021). The improved approaches for authentication and classification of EBN could benefit the whole industry by having a more economical, rapid, and streamlined industrial preparation of EBN products, as well as providing better guarantee of the quality of EBN products to meet the consumers requirements.
EBN has been a traditional, healthy food delicacy since the Tang Dynasty, however scientific research has been lacking over the last century. EBN-related publications remained scarce between 2000 and 2011, but the number of publications has increased significantly since 2012–2019. Discussions about the functions of EBN can be found in this review article by Lee et al. (2021) while our review will be focusing on recent discoveries of bioactive components in EBN and the functions of EBN that have never been discussed before.
EBN is known to exert antiviral bioactivity against influenza A virus (IAV) (Biddle and Belyavin, 1963). In another study carried out decades later, it was found that the non-pancreatin treated EBN extracts can bind to influenza viruses, but this antiviral effect was not seen in haemagglutination inhibition assay or the neutralization assay of influenza virus infected Madin-Darby Canine Kidney (MDCK) cells. The proteins or peptides which had the best anti-influenza effect were found to be sialylglycoproteins of size 10–25 kDa in the pancreatin treated EBN extract. Although proteins above 50 kDa (including a major allergen of 66 kDa) in the non-pancreatin treated EBN extract could bind to influenza A virus, neutralization against the virus was limited, indicating small molecular weight proteins of EBN extract was more favourable for its anti-viral effect. Furthermore, the EBN extract after pancreatin treatment did not trigger detrimental effects, such as hemolysis and cytolysis of erythrocytes and MDCK cells even at a higher concentration (4 mg/ml) (Guo et al., 2006). In summary, EBN extract treated with pancreatin is a more effective and safer antiviral agent when compared to non-pancreatin treated EBN extract.
Sialic acid (SA) and its derivatives found in EBN are highlighted by Guo et al., in which they proposed that these compounds potentially contribute to the anti-influenza effects of EBN (Guo et al., 2006). Indeed, sialic acid is the most studied antiviral compound in EBN. One of the identified sialic acid is 5-N-acetylneuraminic acid (Neu5Ac) (Ogura, 2011). Interestingly, the Neu5Ac content in EBN is correlated with its potency in antiviral activity. The concentration of Neu5Ac varies among EBN samples collected at different geographic locations in Malaysia. EBN from Gua Madai contained 6.7 mg/g of Neu5Ac, which is higher than EBN collected from Teluk Intan which contained 3.2 mg/g of Neu5Ac (Haghani et al., 2017). SA residues could interact with the neuraminidase, and prevent this enzyme from cleaving the SA receptor present on the host cell surface hence halting the viral spread and release (Bianco et al., 2001; Haghani et al., 2016; Benton et al., 2017). Treatment with EBN for 24 h after the MDCK cells were inoculated with IAV for 1 h had the highest antiviral activity and percentage of protection against IAV when compared to EBN pre-treatment (MDCK cells treated with EBN for 1 h before IAV inoculation) and co-treatment (MDCK cells treated with EBN and IAV at the same time for 1 h) groups. It is speculated that the antiviral action of EBNs might be related to the viral release process from the cell membrane rather than viral attachment to the host cell. In the same study, EBN reduced early endosomal trafficking of the IAV by downregulation Rab5 and RhoA GTPase proteins. EBN also inhibited the IAV life cycle by regulation of autophagy with decreased expression of LC3-II and increased lysosomal degradation (Haghani et al., 2017). Besides that, EBN also reduced viral proliferation as effective as oseltamivir phosphate (a commercial antiviral drug) as shown by its effect in suppressing the viral enzyme, neuraminidase in IAV-infected BALB/c mice. Concomitantly, EBN showed high immunomodulatory effects against IAV by significantly increasing the levels of interferon-gamma (IFN-γ), tumour necrosis factor-alpha (TNFα), nuclear factor kappa B (NF-κB), interleukin-2 (IL-2), some pro-inflammatory cytokines (IL-1β and IL-6) and other regulatory cytokines (IL-4, IL-10, IL-12, IL-27 and CCL-2) depending on the stage of infection (Haghani et al., 2016).
At the same time, EBN also possesses immune-enhancing properties by modulating the immune system. However, there are only a limited number of experiments which have studied the effects of EBN on immune regulation. Zhao et al. studied the immunomodulatory effects of EBN in immunocompromised BALB/c mice treated by cyclophosphamide. Results showed that EBN stimulated proliferation and activation of B cells. Enhanced antibody secretion from B cells and better protection of the B cells from damage were detected in experimental mice, resulting in reduced intestinal immune damage (Zhao et al., 2016). Furthermore, EBN was also proven to enhance T lymphocytes transformation and increase serum immunoglobulin M (IgM) in the mouse model (Zhang et al., 1994). In addition, Cao et al. also stated that humoral immune, cellular immune and nonspecific immune were strengthened and regulated through EBN-induced improvements on the spleen and thymus index together with the phagocytosis rate and phagocytosis index of the peritoneal macrophages (Cao, 2012). Last but not least, EBN could also potentiate the proliferation of human peripheral blood monocytes (Ng et al., 1986).
EBN has also shown the potential in curing ulcerative colitis. An in vivo study was done by administering 5 ml 2% dextran sulfate sodium (DSS) via oral route daily for 7 days to induce ulcerative colitis in the male C57BL/6J mouse model. EBN was introduced in three concentrations (0.3 g/kg, 0.7 g/kg and 1.3 g/kg). Histologically, EBN-treated mice had a lower inflammation activity and a more complete structure of the submucosa, in comparison to the positive control group (without EBN supplementation) which showed excessive inflammatory cell infiltration, exfoliated epithelium, disrupted glandular arrangement and reduction in number of goblet cells in the submucosa. EBN reduced severe inflammatory reaction by lowering myeloperoxidase (MPO) activity and production of TNF-α and IL-6. In addition, reduced level of IL-17 and increased level of TGF-β were also determined in the EBN-treated groups. The author suggested that EBN ameliorated the inflammation by restoring the balance of Th17 cells and Treg cells, which was seen to be increased in the positive control group. This was further proved by the immunohistochemistry (IHC) and western blots, in which the IL-17A and Foxp3 genes were overexpressed in the positive control group whereas the overexpression of these two genes was inhibited by EBN. Concisely, EBN was proven to alleviate DSS-induced inflammation through immunomodulation by restoring the balance of Th17 and Treg cells and their associated cytokines (Fan et al., 2021).
Type of solvent used for extraction may have an impact on the antimicrobial activity of EBN. A metabolite, thymol-beta-D glucopyranoside was detected in the EBN. This metabolite has been previously reported to be an effective agent against food-borne bacteria (i.e. Staphylococcus aureus (S. aureus), Escherichia coli (E. coli), Salmonella typhimurium, Shigella flexneri etc.) (Liang et al., 2007; Chua et al., 2014). Hence, it was speculated that presence of this metabolite would confer antimicrobial activity in EBN. EBN extracted with methanol showed antibacterial activities for S. aureus (100 mg/L), Candida albican (C. albican) (100 mg/L), E. coli (1,000 mg/L), and Aspergillus niger (A. niger) (>3,000 mg/L), but the EBN extract derived by soaking extraction in ethyl acetate showed no antibacterial effects. Results of the solvent extraction methods showed that the EBN extracts with ethyl acetate were more effective than using methanol with C. albican (20 mg/L) and A. niger (20 mg/L) while the solvent extraction method with methanol showed slight effects with S. aureus (20–100 mg/L) and E. coli (>100 mg/L). In another study, the antibacterial activities of EBN extracted with 0.05 M alkaline with heating were tested on two Gram-positive (Bacillus subtilis and S. aureus) and two Gram-negative (E. coli and Klebsiella pneumoniae) bacterial strains but no antibacterial effects were observed in those tests (Saengkrajang et al., 2011). It is speculated that the extraction method could significantly affect the antibacterial effects of EBN. More studies should be done to identify the optimal extraction method of EBN to retain antimicrobial functions.
In short, EBN has shown its potential in enhancing the immune system in both in vitro and in vivo models. Nevertheless, more investigations are warranted to understand the mechanisms of immuno-stimulation and antibacterial effects exhibited by EBN.
EGF were detected in the crude EBN collected from Rompin (30.7 pg/ml) and Sibu (74.5 pg/ml) from Malaysia using ELISA quantification kit. Tan et al. postulated that the presence of EGF in EBN can stimulate cancer cell growth. Four cancer cell lines (MCF-7 human breast adenocarcinoma cells, A549 human alveolar adenocarcinoma cells, Caco-2 human epithelial colorectal adenocarcinoma cells, and HCT116 human colorectal carcinoma cells) with EGF+ phenotype were selected to test for cell proliferative effects of EGF content in EBN using MTT assay. However, results showed no significant growth was observed in all 4 cell lines treated with EBN. It was thought that the EGF content in EBN could be too low to induce cancer growth (Tan et al., 2020). In another study, the cell viability of Caco-2 cell treated with two commercial EBNs were 84 and 115% respectively, while Caco-2 cells treated with unprocessed EBN from the East Coast, North and South Zones of Peninsular Malaysia were 91, 35 and 47% respectively. RAW 264.7 cell is a macrophage cell line that is usually used to study TNF-α expressions in macrophage. The EBNs collected from South and East Coast Zones of peninsular Malaysia as well as one of the commercial EBN, significantly reduced TNF-α production in RAW cells, where it was reduced to 24%, 32%, and 43% respectively. Geographical location and sources of the EBN are the factors resulting in the observed discrepancies in cancer cell proliferation rate. In the same study, commercial EBN products induced Caco-2 colorectal cancer cell growth. It is speculated that the adulterant in commercial EBN products stimulated Caco-2 cell growth, future studies are required to find out the substance responsible for promoting cancer cell growth in adulterants with introduction of better authentication procedure to detect such adulterants (Aswir and Wan Nazaimoon, 2011).
There is evidence that EBN possesses wound-healing effects. Hwang et al. studied the wound healing effects of EBN on HaCaTs cells and normal human dermal fibroblasts (NHDFs) which had been irradiated by ultraviolet B (UVB). Results showed that treatment with EBN significantly improved wound healing effects in both HaCaTs and NHDF cells. In fibroblasts, healing rate of the cells treated with 10 μg/ml increased by 39.6% after 24 h as compared to 0 h timepoint. Healing rate is faster in EBN treated cells as compared to fibroblastic self-healing (17% without any treatment) and 10 μg/ml allantoin treated group (28.2%). Hyaluronic acid, also known as hyaluronan, is a straight-chain carbohydrate extracellular matrix polymer that is a key component for the wound healing process (Aya and Stern, 2014). 10 μg/ml of EBN also significantly increased the production of hyaluronan in HaCaTs cells by 109.1%. In the study, EBN was also shown to confer anti-inflammatory effects as EBN reduced the expressions of two inflammatory cytokines, TARC/CCL17 by 89.7% and MDC/CCL22 by 46.1% in TNF-α/IFN-γ-stimulated HaCaTs cells. The author suggested that rapid healing effect induced by EBN was mediated through enhanced production of hyaluronan and the downregulation of MMP-1 and upregulation of procollagen type I expression in UVB-irradiated NHDF cells. In summary, these results indicate that EBNs have the potential to ameliorate UVB-induced skin photo ageing and TNF-α/IFN-γ-stimulated inflammation as well as wound injuries, resulting in faster healing rate (Hwang et al., 2020).
EBN also improves healing in the cornea. It was shown that rabbit corneal tissues treated with 0.05% EBN (with or without the addition of serum) resulted in increased expressions of collagen type I, aldehyde dehydrogenase (ALDH) and lumican, as compared to those treated with either serum-containing medium or serum-free medium (Zainal Abidin et al., 2011).
An in vivo study done by Albishtue et al. to study the effects of EBN on female reproductive system using adult female Sprague Dawley rats showed that oral EBN treatment at 60 and 120 mg/kg per day for 9 weeks augmented proliferation of uterine cells, including luminal epithelium, glandular epithelium, and stromal cells, resulting in better uterine and reproductive functions. Besides that, the endometrial receptivity and the number of implantation sites were also increased by EBN treatment. EBN also enhanced production of reproductive hormones, estrogen, progesterone, and prolactin, as well as expression of steroid receptors, progesterone and estrogen receptors. Upregulation of vascular endothelial growth factor (VEGF), EGF, EGF receptor, and proliferating cell nuclear antigen (PCNA) were found in the endometrial tissue. In short, EBN showed its potential to improve embryo implantation and also increased successful pregnancy rates (Albishtue et al., 2019).
Recently, it was reported that the action of Neu5Ac is not only limited to antiviral functions, but it also exhibits skin whitening and bone maintenance effects. In a skin whitening test, pepsin-digested EBN showed stronger inhibition of melanogenesis in cultured murine B16 skin cells and enzymatic activity of tyrosinase, as compared to that of undigested EBN. In addition, the pepsin-digested EBN also exhibited stronger osteogenic activity in cultured MG-63 osteoblasts cells. Neu5Ac in EBN was originally in conjugated form and was released from the conjugated form by treating EBN with pepsin in simulated gastric fluid at pH 2 condition for 48 h. This study indicated that prolonged EBN extraction in a gastric-like condition could achieve the full beneficial functions of EBN. Wong et al. tailored the method of Neu5Ac extraction from EBN (Wong et al., 2018c).
EBN also has an impact on metabolism. Western blots showed that diabetic db/db mice treated with 75 and 150 mg/kg of EBN orally had elevated insulin levels when compared to mice treated with distilled water only. Meanwhile, insulin signalling receptor (IRβ) and downstream proteins (p-IRS1, PI3K and p-Akt) were also upregulated in mice treated with 75 and 150 mg/kg of EBN. Treatment with 75 and 150 mg/kg of raw EBN also decreased expression of pro-inflammatory cytokines, IL-6 and TNF-α, and also inflammatory protein, NF-κB in these mice. Moreover, at these doses, oxidative stress was ameliorated, as shown by downregulation of NADPH oxidase 4 (NOX4) protein, a reactive oxygen species (ROS) marker, and upregulation of superoxide dismutase-1 (SOD-1) protein, an antioxidant protein. In summary, EBN improves β-cell function and insulin signalling by attenuation of oxidative stress-mediated chronic inflammation in type 2 diabetic mice (Choy et al., 2021).
In another study, EBN demonstrated its potential role in preserving endothelial function by reducing oxidative stress in both cultured cells and mouse models. In the ex vivo study using male C57BL/6J mice, aortic ring assay indicated that impaired aortic relaxation in high glucose-fed mice was reversed with treatment of raw EBN (15 and 30 μg/ml). Human umbilical vein endothelial cells (HUVECs) were also utilized to examine the effects of EBN on high glucose-induced ROS formation. Results of lucigenin-enhanced chemiluminescence assay showed that treatment with 30 μg/ml of raw EBN significantly diminished intercellular ROS level and vascular superoxide anion production in HUVECs. EBN treatment at 30 μg/ml in HUVECs or 150 mg/kg in db/db mice reversed the high glucose-induced depletion of nitric oxide (NO). Western blot analysis also showed that EBN treatment at 30 μg/ml in HUVECs or 150 mg/kg in db/db mice could significantly reduce the level of NADPH oxidase 2 (NOX-2) and nitrotyrosine proteins while increase SOD-1 and p-eNOS protein levels (Murugan et al., 2020).
EBN has antioxidant effects which could serve as a novel alternative therapy for oxidative stress-mediated neurodegenerative diseases such as Alzheimer’s disease (AD) and Parkinson’s disease (PD) (Tai et al., 2017). The antioxidant capacity of EBN is higher in extracts obtained via alkaline extraction, it is speculated that the hydrolysed proteins release amino acids such as Cys, Met, His, Try and Lys with antioxidant properties (Lee et al., 2015). Simulated human gastro-intestinal digestion study demonstrated that the antioxidant components of EBN are released after digestion in the human gut. The digested EBN was shown to enhance antioxidative activities and protected HepG2 human liver cells from hydrogen peroxide (H2O2)-induced cytotoxicity (Yida et al., 2014).
Pathogenesis of PD involves oxidative stress-induced death of the midbrain dopaminergic neurons. EBN reversed ROS and nitric oxide (NO) build-up, reduced lipid peroxidation, inhibited caspase-3 cleavage, and attenuated apoptotic cell death in an in vitro PD model, neurotoxin 6-hydroxydopamine (6-OHDA)-treated SH-SY5Y cells. EBN improved motor function and balancing in 6-OHDA-treated C57BL/6J mice in vivo PD model. Inhibition of microglia activation and enhancement of antioxidant enzyme activity was shown in EBN-treated mice (Yew et al., 2014, 2018). Neurotrophic properties of EBN were demonstrated by increased cell proliferation and migration in a neural stem cell model, embryonic mouse neuroectodermal cells (NE-4C) (Yew et al., 2019). In another study, lipopolysaccharide (LPS) elicited cognitive impairment in rats by significantly increasing the escape latency while decreasing the number of entries in the probe trial, which were coupled with increased production of proinflammatory cytokines (TNF-α, IL-1β, and IL-6) and oxidative markers (ROS and TBARS) in the hippocampus. Treatment with EBN (125 mg/kg, 250 mg/kg, and 500 mg/kg; p.o.) effectively reversed effects of LPS on escape latency and probe trial. In addition, these treatments also inhibited the LPS-induced upregulation of pro-inflammatory cytokines and oxidative markers (Careena et al., 2018).
The effects of EBN on spatial learning and memory were also examined. The results showed that EBN supplementation had a dose-dependent improvement on cognition as evidenced by significant shorter escape latency. EBN-induced improvement of spatial learning and memory was also seen in the newly born offspring mice, which was fed on maternal milk of mother mice exposed to EBN supplementation. At the molecular level, EBN also exhibited its effect on attenuating neuro-inflammation and neuro-oxidative stress through increasing expression of SOD and decreasing levels of malondialdehyde (MDA) in the newly born offspring mice fed on maternal milk of mothers exposed to EBN supplementation. Evidence showed that EBN-induced enhancement of cognitive activity was due to elevation of SIRT1 expression in the pyramidal layer and dentate gyrus of the hippocampus. Besides that, EBN exhibited neuro-protective effect through anti-apoptotic mechanism in which the caspase-3 cleavage and early apoptotic membrane phosphatidylserine externalization were inhibited. The antioxidant effect of EBN through decreasing ROS level and increasing the expression of SOD gene in hippocampal neurons (SH-SY5Y neuroblastoma cells) was another neuroprotective mechanism (Ismail et al., 2021). In a recent study, zeroth generation CJ57BL/6 mice gave birth to first and second generations of offspring after 6 weeks of EBN supplementation (10 mg/kg). Both generations of offspring showed improvement in Y-maze cognitive performance at 6 weeks of age. Brain samples of the offspring mice demonstrated upregulation of GNE, ST8SiaIV, SLC17A5, and BDNF mRNA, and increased densities of synaptic vesicles in the presynaptic terminal (Mahaq et al., 2020).
Interestingly, EBN has also shown to have antioxidant effect in fly model. Drosophila melanogaster was cultured in four different groups, in which they were fed by food medium supplemented with 0 g/kg (control), 1 g/kg, 3 g/kg and 9 g/kg of EBN. Results of ferric reducing antioxidant power (FRAP) showed that the EBN-treated groups had higher total antioxidant activity in a dose-dependent manner as compared to control which had low total antioxidant activity. EBN enhanced the antioxidant capacity by increasing the SOD and catalase activities (CAT) whereas the MDA level was decreased. In a nutshell, EBN could improve ageing problems by reducing oxidative stress, hence increasing the lifespan of Drosophila melanogaster (Hu et al., 2016).
Based on the Memorandum of understanding (MOU) on the Protocol of Inspection, Quarantine and Hygiene Requirements for importation of bird nest products from Indonesia and Malaysia into China, sealed and signed by Malaysia and Indonesia with China, the contaminants were assessed from the aspects of physical, microbiological, residual, heavy metals and excessive minerals, parameters and tolerance levels (Yeo et al., 2021). Bacteria, fungi, and mites are commonly reported to be found in EBN. Microbial growth in EBN might be attributed to the environment of EBN collection, such as high humidity levels and lower temperature or microbial infestation during EBN storage. The microorganisms could also be originated from saliva or feathers of swiftlets or the nest itself (Kew et al., 2014; Yeo et al., 2021). Additionally, Wong et al. revealed that both raw and commercial EBNs contain diverse types of bacteria, including Staphylococcus sp., Bacillus sp. and Acinetobacter sp. Although the double-boiling treatment could effectively kill most of the bacteria, heat-resistant species like Bacillus sp. and Brevibacillus sp. were still isolated after treatment (Wong S. F. et al., 2018). Consumption of Bacillus cereus (B. cereus)-contaminated food can lead to diarrheal and emetic syndrome, resulting from production of B. cereus toxins (Griffiths and Schraft, 2017). A promising non-thermal processing method for food preservation by polychromatic low-energy X-ray with a high linear energy transfer (LET) effect results in a high relative biological effect (RBE). Low-energy X-ray with cut-off energy of 150 KeV was applied to inactivate two of the most prevalent foodborne pathogens in dry EBN. X-ray irradiation at 350 and 400 Gy decreased E. coli O157:H7 and S. Typhimurium from 6.35 ± 0.56 and 5.84 ± 0.67 log CFU/g, respectively, to undetectable levels. Based on dose distribution in 10 stacked pieces of EBN, two-sided irradiation could effectively inactivate pathogens uniformly (Zhang et al., 2020).
Environmental fungi from soil and plants are generally detected in both raw and commercial EBN. Although most of the fungi were removed after boiling of EBN up to 100˚C for 3 h, the environmental fungal genera of Aspergillus sp. and Penicillum sp., can still be isolated in both EBN samples. It was speculated that these fungi were introduced from the EBN processing facility environment after the boiling process because these are two of the most frequently isolated environmental fungal genera. However, it is also possible that these fungal genera are thermoresistant or thermotolerant (Chen et al., 2015). Aspergillus sp. and Penicillum sp. are commonly known as food-spoilage fungi and can produce different mycotoxins (e.g. aflatoxins and ochratoxin A), which could cause various diseases in human as well as opportunistic infections in immunocompromised individuals (Greeff-Laubscher et al., 2020). Future studies are warranted for complete elimination and contamination management of these two fungal genera. Moreover, Cladosporium sp. and Eurotium sp. are detected in raw EBN samples, as these two genera of fungi are known to cause respiratory infections, swiftlet ranchers require personal protective equipment to prevent inhalation of these hazardous microbes (Chen et al., 2015). Mites are known as a source of allergens that could cause anaphylaxis (Sanchez-Borges et al., 1997). Mites, their faeces and eggs, and feather strands were observed on the surface of both raw and commercial EBN through EBN surface structural analysis under the electron microscope (SEM). These contaminants remained on the surface of EBN even after the washing and processing procedures were done (Kew et al., 2014; Tai et al., 2020). EBN is one of the causes of food-related anaphylaxis among children (Goh et al., 1999) and the pathophysiology is suggested to be related to immunoglobulin E (IgE)-mediated hypersensitivity caused by the protein existing in several isoforms seen at 66 kDa. N-terminal sequence of the major putative allergen (66 kD) showing homology to a domain of an ovoinhibitor precursor in chicken (Goh et al., 2001).
Insects consume plants inhabited by nitrogen-fixing bacteria and crops fertilized with nitrogen-based fertilizers. When swiftlets consume these insects, this results in accumulation of nitrites and nitrates in the swiftlets. As the nest is made of salivary secretion of swiftlets and embedded with droppings of swiftlets, high concentrations of NO3− and nitrogen-fixing bacteria are detected. The nitrogen-fixing bacteria converts NO3− into NO2− giving rise to redding of EBN by NO3− reductase. Although up to 98% of NO2− could be removed after commercial EBN processing, NO2− and NO3− is still found in high concentrations in EBN, exceeding the acceptable tolerance level which is ≤30 ppm (But et al., 2013; Quek et al., 2018a; Yeo et al., 2021). In fact, the daily intake limit of NO2− as advised by the World Health Organization (WHO) is between 0 and 3.7 mg/kg body weight (Yeo et al., 2021). To clear the traditional belief of red EBN being more precious, Paydar et al. (2013) discovered that the red colour in EBN is contributed by NO3− and NO2− instead of the presence of haemoglobin. NO3− is relatively stable but NO2− is active and can react with coexisting amino acids to form a carcinogenic compound called nitrosamine (Yeo et al., 2021). Semicarbazide, which originates from bleaching process used to remove impurities, was another carcinogenic compound found in EBN (Xing et al., 2012). A safer method to prevent red colouration of EBN is by using sodium tungstate, an inhibitor of NO3− reductase that suppresses formation of NO2− in cubilose (Chan et al., 2013). Nonetheless, a recent study reported that the acidic mammalian chitinase-like protein found in EBN contributes to the red colour as a noticeable increase in Fe-O bonding intensity after the colour change in EBN (Wong et al., 2018b).
Heavy metal and mineral contaminations in EBN were also reported in recent studies. Studies showed that in raw EBN, levels of Hg and Cu were higher than the permissible limits whereas the levels of Pb, As and Cd were below the limits (Chen et al., 2014; Quek et al., 2018b). A trace amount of these elements was also found in commercial EBN (Tan et al., 2020). As a matter of fact, the maximum permissible levels of each element is as follows; 1) Hg ≤0.05 ppm; 2) Cu ≤1 ppm; 3) Pb ≤2 ppm; 4) As ≤1 ppm; and 5) Cd ≤1 ppm. In addition Fe level was found to be higher than the regulatory limit (0.3 ppm) in both raw and commercial EBN (Yeo et al., 2021). All these heavy metals, if consumed in excessive amounts could react with proteins or enzymes in the human body, causing chronic heavy metal poisoning syndrome (Dai et al., 2020). For example, Hg intoxication could cause severe behavioural and cognitive changes as well as delayed development of growth and neural system in children (Ha et al., 2017). Other than that, Cd is known to be a mutagenic compound and Cd consumption could result in cancer development (Fatima et al., 2019). An excess amount of Cu is also associated with certain human disorders, such as cardiovascular diseases, neurotoxicity and hepatic disease (But et al., 2013; Paydar et al., 2013).
In short, EBN consumption undoubtedly has its safety issues due to the potential residual contaminations including microorganisms (bacteria, fungi, and mites), heavy metals (Hg, Pb, As and Cd), minerals (Fe and Cu) as well as NO2− and NO3− contents. On top of that, adulterants used in EBN production and semicarbazide used in the bleaching process could also pose hazardous effects to human health. Therefore, the quality and authentication of EBN should be strictly regulated to avoid consumers from eating inauthentic EBN and thus the risk of EBN consumption could be reduced to a greaer extent.
The study of EBN has recently gained attention from researchers in the past decade as recent laboratory studies of EBN demonstrated many optimisation steps for the production of better EBN products. We summarise the optimisation steps based on laboratory findings that could be useful references for setting up the industrial EBN processing with the aims to improve the safety, taste, and preserve the health benefits of EBN products (Table 1).

TABLE 1. Summary of EBN extraction studies.
After collection from the swiftlet premises or caves, EBN is processed following these general procedures: cleaning, drying, grinding, soaking/elution (specific for elution water extraction (eHMG)), heating, acid/alkaline treatment (specific for acid/alkaline extraction), enzymatic treatment (specific for enzymatic extraction), filter, centrifugation, freeze-drying/lyophilisation, storage. Slight changes for processing steps were made as reported in different groups for the consideration of targeted nutrients, convenience, cost-effectiveness, and instruments availability. These processing steps are critical for preservation of nutritional values of EBN (Yew et al., 2014; Gan et al., 2017b; Hong et al., 2020; Tong et al., 2020).
Firstly, the collected EBN is soaked in ultrapure water until softened to loosen the protein strands and cleaned by washing in water or ultrapure water, followed by manual removal of dirt, feathers, and egg shells using forceps (Yew et al., 2014). The dust and lighter impurities floating in the water and EBN could be easily picked with forceps as the EBN expands in size after soaking in water (Hong et al., 2020). Then, the cleaned EBN is dried with or without controlled air circulation, heat, and humidity. EBN could be dried at a faster rate with higher air circulation speed and temperature but this intervention is associated with reduced sialic acid and antioxidant retention in EBN (Gan et al., 2017a; 2017b). The dried EBN, around 10–12% moisture then undergoes grounding with pestle and mortar manually or electric blending based on preferences as no study was conducted for the comparison between manual and automatic grounding process (Gan et al., 2017a; Yew et al., 2019). The grounded EBN is sieved through a wire mesh (0.4–1.0 mm) to further separate the feathers and other impurities (Zainal Abidin et al., 2011). The next step is extraction of chemical compositions of EBN by extraction method of HMG, heat extraction, enzymatic extraction, acid extraction, alkaline extraction, and eHMG.
Grounded EBN is soaked in cold distilled water 2.5% (w/v) or deionized water at 0.2% (w/v) at 4˚C, and usually left for overnight but some studies have also left it for 16–48 h (Guo et al., 2006; Yew et al., 2014; Tong et al., 2020). After that, heating of EBN solution is carried out in the range of 40˚C to 121˚C for 15 min to 4 h (Amiza et al., 2019a; Tong et al., 2020). If it was acidic extraction, the EBN mixture is heated at 80°C with 2% (v/v) of 0.4 M sulfuric acid for 4 h subsequently allowed to cool down and centrifuged at 2,716 g (5,000 rpm) for 15 min. The pH of supernatant collected is neutralized to pH 7.0. White precipitation formed is removed through centrifugation with 2,716 g (5,000 rpm) for 15 min at 4°C. The supernatant is then collected and kept at 4°C for further analysis (Tong et al., 2020). However, if it was an alkaline extraction, 1 g of each raw sample is immersed in 30 ml of 0.1–0.4 M NaOH solution for 48 h. Then, the aliquot of each extract is immersed in the water bath at 65°C for 2 h. The extracted solutions are centrifuged and eventually the supernatant is obtained. After the extraction process, the suspensions are centrifuged at 18,000 rpm for 20 min, and the supernatants are thoroughly dialysed against distilled water (Hun et al., 2016). For enzymatic treatment, enzymes such as pancreatin, alcalase (Khushairay et al., 2014), protease from Bacillus licheniformis (Ling et al., 2020), and neuraminidase from C. perfringens (Guo et al., 2006) is used. In continuation of the heating, the enzyme is usually added to the EBN mixture at a warm temperature between 38°C-60°C, at pH 8.5–9.0 to facilitate partial protein denaturation. It is then followed by heating at 90°C for 5 min for enzyme deactivation. The EBN mixture is then filtered using filter paper, filtrate is collected and freeze-dried to obtain EBN powder (Guo et al., 2006; Yew et al., 2014). Freeze-drying process aims to produce dehydrated EBN powder while retaining maximum nutrient and bioactive compounds (Bhatta et al., 2020). Lastly, the freeze-dried EBN powder is stored at 4°C or −80°C freezers for future use. In industrial setting, freeze-dried EBN could be made into cosmetics, tablets, ready-to-drink beverages, tonics, spices and baking powder among others (Lee et al., 2021). In the industrial processing of whole raw EBN, it is started with sorting and grading of EBN based on the difficulty to clean the EBN. The cleaning step is important as EBN with more feathers and impurities are graded lower and sold at a lower price. After sorting, EBN will be soaked in clean water, so that dust and impurities will float on the water. Feathers that remained on the EBN will be hand-picked with forceps or tweezers. This is a labour-intensive process that could be time-consuming. After that, EBN is placed on a mold and left to dry by heating cabinet and fan blowing. This process could lead to nutrients loss. The dried EBNs in cup shape is sorted and graded again based on their colour, cleanliness, and shape for packing (Hong et al., 2020).
As mentioned earlier, EBN is required to be free from avian influenza. The MOU on the Protocol of Inspection, Quarantine and Hygiene Requirements demands EBN products from Indonesia and Malaysia to be free from avian influenza to be imported into China (Badan Karantina Indonesia, 2014; Rohaizan, 2017). Only EBN obtained from farms registered with the Department of Veterinary Services, Malaysia is authenticated. The Chinese government has imposed stringent procedures for quality assurance by tracing the entire supply chain from EBN harvesting to exporting to China (Thorburn, 2015). Additionally, EBN processing facilities must be inspected and approved by Certification and Accreditation Administration of the People’s Republic of China (CNCA) (Minister Department, 2012). According to the information provided by the CAIQ official website http://ebn.caiq.org.cn/overseasRegist, Indonesia has 23 EBN processing facilities and 436 swiftlet houses approved to be exported to China; Malaysia has 41 EBN processing facilities, but there is no information about the registered swiftlet houses; Thailand has 2 EBN processing facilities and 21 registered cave EBN sources (Development Report of Imported Bird’s Nest in 2021, 2021).
Department of Veterinary Services of the Ministry of Agriculture and Food Safety and Quality Division, Ministry of Health, Malaysia, implements and enforces that EBN products must be processed with heat treatment, with core temperature of the products shall be higher than 70°C and retained for at least 3.5 s to effectively kill the avian influenza virus (Yeo et al., 2021). Apart from that, environmental fungi in raw EBN should be removed before consumption. Heating EBN to ≥100°C for at least 3 h could remove the environmental fungi in EBN (Chen et al., 2015). The heating temperature and time possess critical effects on extraction of bioactive compounds from EBN as well as retention its nutritional values. Excessive heat treatment is known to cause alteration in protein native structure resulting in unfolding of protein and altered protein surface exposure (Teodorowicz et al., 2017). As the proteins denature, bioactive properties are lost and this would affect its nutritional value.
Furthermore, differential scanning calorimetry have often detected loosely bound water (dehydrates from EBN below 110˚C) and tightly bound water (dehydrates from EBN between 100˚C and 200˚C) in EBN samples. In fact, it implies the wide use of H2O2, as a bactericide and bleaching agent in bird nest cleaning industry to make the EBN more whitish, so that it could be graded as high-grade EBN. Some of the H2O2 may be tightly hydrogen-bond in coils of the glycoprotein chains, just like the tightly bound water, and may remain trapped in the EBN on drying typically below 60°C, and this could present as a food hazard. It may be necessary to heat dry the cleaned EBN at above 100°C to dislodge and decompose H2O2, or to reduce it in the moist EBN with permitted reducing agents like ascorbic acid, or by enzymatic decomposition with catalase, before drying (Shim et al., 2017). This study further supports the importance of heat treatment for EBN. Future studies focusing on the optimum heating condition for pathogenic microorganisms removal and nutrients retention should be carried out. The heating step should be introduced into industrial EBN products processing, especially for those ready-to-drink beverages to prevent potential avian influenza virus infection and food poisoning.
Mites and fungal spores are contaminants widely found in EBN and more mite faecal pellets and eggshells were found in raw EBN compared to commercially processed EBN (Kew et al., 2014). Certain mites are heat-resistant and can survive the cooking process to elicit allergic reactions in consumers (Yan et al., 2008). Bromelain, an enzyme derived from pineapple showed promising results in killing mites and fungi found in raw EBN (Tai et al., 2020). Using bromelain to remove microbial contaminants could be a safer choice than bleaching agents like H2O2 and semicarbazide. A study showed that semicarbazide detected in commercial EBN products originated from bleaching of EBN. As semicarbazide is carcinogenic, use of semicarbazide to bleach EBN should be strictly prohibited (Xing et al., 2012). Moreover, the packaging of EBN products should avoid using azodicarbonamide-containing packaging as it could produce semicarbazide due to thermal decomposition (Stadler et al., 2004). Most importantly, adulterants should not be added to EBN products and storage should be kept clean from potential contaminants for safe consumption of EBN. Hypothetically, simultaneous applications of both heat and enzymatic EBN extraction could provide additional advantages after optimisation.
Apart from using heat and enzymatic treatments to remove contaminants, these steps could improve bioavailability of nutrients in EBN. The extracted product compositions are unique for each extraction method due to their physicochemical properties (Tong et al., 2020). Protein functionalities based on its physical and chemical properties including size, shape, amino acid composition, sequence, net charge and distribution, hydrophobicity/hydrophilic ratio, secondary, tertiary, and quaternary structures, molecular flexibility/rigidity, and ability to interact with other components. However, extensive denaturation of proteins often results in insolubilisation, which can consequently impair functional properties that are dependent on solubility. Ideally, partial denaturation of protein often improves digestibility and biological availability of essential amino acids (Damodaran and Parkin, 2017). Enzymatic protein hydrolysis is commonly used to modify the nutritional, physicochemical, functional, digestibility, sensory, and bioactive properties of the protein, as well as to reduce allergenic and anti-nutritional compounds (Tavano, 2013).
A widely studied enzyme, pancreatin is a mixture of digestive enzymes that is secreted from the pancreas with proteolytic, amylolytic and lipolytic activities. Treatment with pancreatin significantly increases the protein’s solubility, degree of hydrolysis and bioactive peptides concentration of the mixture (Khushairay et al., 2014). For example, pancreatin-treated EBN has higher efficiency compared to non-pancreatin treated EBN in reducing cell death in 6-OHDA-challenged SH-SY5Y cells. However, water extract exhibited higher efficacy in ameliorating ROS build up, early apoptotic membrane phosphatidylserine externalization as well as inhibition of caspase-3 cleavage (Yew et al., 2014). In in vitro assay, EBN treated with pancreatin had superior anti-viral effect and showed no side effects, rendering the pancreatin treated EBN a safer and more effective antiviral agent as compared to EBN extracted with water (Guo et al., 2006).
The choice of EBN extraction should be made based on desired biomedical application because bioactive compounds could be best extracted by a particular extraction method. Alcalase is an enzyme that has broad specificity and has been employed to modify the functional properties of a range of proteins. It is used mainly to cleave the carboxyl side of the hydrophobic amino acid (Adler-Nissen and Olsen, 1979). It is speculated that combined applications of heat treatment and enzymatic protein hydrolysis may help to maximise bioactive components functionality, digestibility, and bioactivity. When compared to a variety of heating modalities and heating plus alcalase treatment: slow cook with or without alcalase treatment (85–96˚C, 60–120 min), autoclave with or without alcalase (121˚C, 15 min), and raw (without heating and alcalase treatment), EBN heated at 100˚C for 60 and 30 min followed by alcalase treatment gave highest degree of hydrolysis, 22.59 and 20.42% respectively. The degree of hydrolysis is positively correlated with solubility where high solubility of normal boiled, enzymatically hydrolysed EBN possess the potential of EBN being used for food formulation, nutraceutical, and cosmetic products (Amiza et al., 2019a). On top of that, alcalase treatment also helped in removal of impurities (for example ash content), and improves the aesthetics of commercially available EBN (Noor et al., 2018).
Neu5Ac in EBN is known for its antiviral, skin whitening, and bone maintenance functions. The EBN treated with pepsin in simulated gastric fluid at pH 2 for 48 h showed higher potency for the functions as mentioned than those EBN samples without extraction process under such condition. Hence, prolonged treatment of EBN in gastric-like conditions could retain better Neu5Ac bioactivity (Wong et al., 2018c). The application of enzymes in EBN preparation have shown promising results and it is anticipated that these procedures should be incorporated into industrial EBN processing. Treating EBN with enzymes for elimination of contaminants, extraction of bioactive compounds, and allergen deactivation could provide consumers with better and safer EBN products.
The pH value during EBN extraction is one of the important factors to retain maximum bioactivity. Both acid and alkaline extraction methods including hydrochloric acid (Quek et al., 2018a) and sodium hydroxide (Hun et al., 2016) were studied. Extracting bioactive compounds from EBN using acid was shown to be inefficient when compared to acidic enzymatic extraction. Hence, supplementing enzymatic extraction (e.g. pepsin) with addition of acids facilitates protein denaturation and the EBN extract will be neutralized to achieve a pH of 7.0 with alkaline to halt the enzyme activity (Tong et al., 2020). In contrast, Amiza et al. (2019b) demonstrated that the optimum conditions for EBN hydrolysis is at pH of 9.46, Alcalase to substrate concentration of 2%, hydrolysis time of 179.55 min, and 64.99°C, can achieve 37.9% degree of hydrolysis. Alcalase require pH 7.5–9.5 and neutralized with acids after the extraction (Amiza et al., 2019b).
The fatty acids content of EBN is relatively understudied but recently Lee et al. reported that EBN contains 48.43% of poly-unsaturated fatty acids (PUFA), 25.35% of saturated fatty acids (SFA), and 24.74% mono-unsaturated fats (MUFA). These fatty acids explain the antioxidative activity of EBN. Briefly, after HMG was done, the triglyceride fraction from EBN was extracted with acetone water mixture (ratio of 75:25) at room temperature for 2 days. The aqueous solution was then obtained after solvent evaporation under a vacuum that was successively partitioned with ethyl acetate (Hun et al., 2020). One of the important factors for EBN nutrients retention is the pH value during acid extraction (Hun et al., 2020).
Recent studies have reported new extraction methods with high efficiency in extracting the highest amount of nutrients from EBN. The EBN eHMG extraction was first demonstrated by Oda et al. and remained a non-disclosure procedure to extract EBN (Oda et al., 1998). Recently, eHMG was found to be the more effective than acid extraction, enzyme extraction, and HMG in terms of EBN extraction, with the highest number of water-soluble (193) metabolites and extra sialic acids extracted (Tong et al., 2020). A recent study showed that the EBN processed with dynamic high-pressure micro-fluidisation (DHPM) at various pressure (range from 20–200 MPa) displayed a remarkable increase in protein solubility. DHPM treatment significantly improved the solubility accompanied by changes in the structural properties of EBN insoluble proteins, which may be caused by intense shear, turbulence force, and cavitation effect of DHPM treatment. Confirmational changes such as the increase in α-helix and β-turn, reduced particle size and de-aggregation of large proteins in EBN water-insoluble fraction. Hence, optimized DHPM could be an alternative method to traditional stew to release the bioactive compound from EBN (Fan et al., 2020). Mass eHMG and DHPM EBN processing could be developed and tested in the future.
Based on the common practice of current whole EBN processing, whole EBN is dried on a mold after soaking and cleaning, skipping extraction as shown in Figure 1 (Hong et al., 2020). The drying process after cleaning of EBN was discovered to be critical for sialic acids and antioxidants retention, in turn providing benefit to the quality and grade of EBN. Higher drying temperature causes degradation of sialic acids and antioxidants in EBN, exhibiting first-order kinetics. EBN dried at 25°C had 83.9 and 96.6% retention of sialic acids and antioxidants, 78.7 and 91.5% at 40°C, 42.5 and 38.7% at 70°C respectively. Longer drying time is required for drying at lower temperature to achieve similar level of dryness in the EBN as the processing of EBN would be slowed down but the retention of nutritional value is significantly better. When compared to convective hot air drying, heat pump drying at a temperature lower than 40°C, with forced air in the initial stage of drying and intermittent forced air supply in the later stage could efficiently retain colour and sialic acid in EBN. Traditional EBN drying step could be improved as suggested by Gan et al. to preserve the highest possible amount of nutrients in EBN. Temperature as low as 25°C could be used to best preserve the colour, antioxidants, and sialic acids in EBN although it will take a longer time to dry and therefore affects the turnaround time for the industrial preparation process. In summary, heat pump drying at a temperature lower than 40°C, with forced-air in the initial stage of drying and intermittent forced air supply in the later stage could efficiently retain colour, antioxidants, and sialic acids in EBN (Gan et al., 2017a; 2017b).
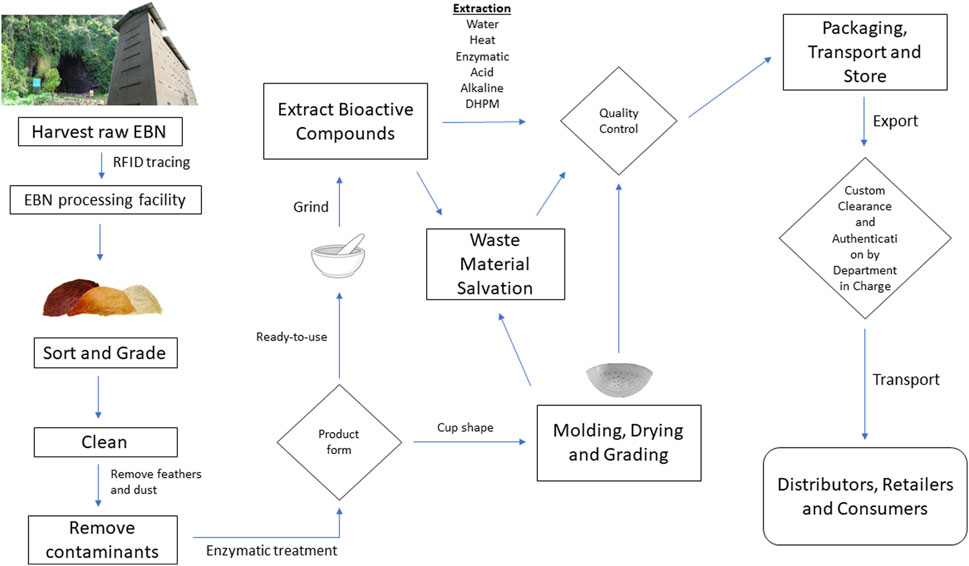
FIGURE 1. Schematic flowchart of EBN industry.
Surprisingly, around 50% of glycopeptide can be recovered from the waste material of EBN, particularly N-acetylneuraminic acid, which is 229% higher than EBN hydrolysate after enzymatic extraction. The antioxidant activities found in waste material after enzymatic extraction was higher than the EBN extract, suggesting a significant bioactive portion of EBN was wasted during the extraction process. The cleaning procedure is a time-consuming process that includes immersion, swelling, separation of impurities, sterilization, molding, drying and secondary sterilization. The cost of processed clean EBN was approximately 2200–2500 USD kg−1 and unprocessed EBN was 1200–1500 USD kg−1 in 2017, and this value is expected to increase in the future. The salvation step should be included in enzymatic extraction procedure to prevent economical and nutritional value loss (Ling et al., 2020).
This review compiled recent investigations of EBN and provided insight into various topics related to EBN products. Increasing studies have proven beneficial health effects of EBN consumption but factors, such as geographical location, harvesting place, harvesting season, and processing influences the nutritional contents of EBN. Hence, standardised EBN processing methods are required to preserve the bioactive effects discussed in this review. The health concerns of EBN consumption caused by adulterants, chemical and microbial contamination should be addressed by strict adherence to guidelines. Several studies demonstrated plausible authentication methods could be translated into industrial settings, but cost-effectiveness and equipment availability required further analysis and comparison. Numerous in vivo animal studies showed promising health-improving functions of EBN with discoveries of bioactive compounds. Future studies to fill the knowledge gap of EBN in promoting human health is promising.
KCC and MGN wrote the manuscript, and YLT, KYN and RYK critically reviewed the manuscript. SMC formulated the entire concept and reviewed the manuscript.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Adler-Nissen, J., and Olsen, H. S. (1979). The Influence of Peptide Chain Length on Taste and Functional Properties of Enzymatically Modified Soy Protein. ACS Symp. Ser. 7, 125–146. doi:10.1021/bk-1979-0092.ch007
Albishtue, A. A., Yimer, N., Zakaria, M. Z. A., Haron, A. W., Babji, A. S., Abubakar, A. A., et al. (2019). Effects of EBN on Embryo Implantation, Plasma Concentrations of Reproductive Hormones, and Uterine Expressions of Genes of PCNA, Steroids, Growth Factors and Their Receptors in Rats. Theriogenology 126, 310–319. doi:10.1016/j.theriogenology.2018.12.026
Amiza, M. A., Khuzma, D., Liew, P. S., Salma Malihah, M., and Sarbon, N. M. (2019a). Effect of Heat Treatment and Enzymatic Protein Hydrolysis on the Degree of Hydrolysis and Physicochemical Properties of Edible Bird’s Nest. Food Res. 3 (6), 149. doi:10.26656/fr.2017.3
Amiza, M. A., Oon, X. X., and Norizah, M. S. (2019b). Optimization of Enzymatic Hydrolysis Conditions on the Degree of Hydrolysis of Edible Bird’s Nest Using Alcalase® and Nutritional Composition of the Hydrolysate. Food Res. 3, 570–580. doi:10.26656/fr.2017.3(5).120
Aswir, A. R., and Wan Nazaimoon, W. M. (2011). Effect of Edible Bird’s Nest on Cell Proliferation and Tumor Necrosis Factor- Alpha (TNF-α) Release In Vitro. Int. Food Res. J. 18, 3. doi:10.3923/jftech.2010.126.130
Aya, K. L., and Stern, R. (2014). Hyaluronan in Wound Healing: Rediscovering a Major Player. Wound Repair Regen. 22, 579–593. doi:10.1111/wrr.12214
Badan Karantina Indonesia (2014). Pedoman Pemanasan Sarang Burung Walet Untuk Pengeluaran ke Negara Republik Rakyat Tiongkok. Available at: https://bbkpsoetta.com/images/Karantina/perundangan/KEPBARANTAN/SK_KaBarantan_2014_No_406.pdf.1–33.
Badan Pusat Statistika (2020). Statistik Indonesia 2020 Statistical Yearbook of Indonesia 2020. BPS-Statistics Indonesia. Available at: https://www.bps.go.id/publication/2020/04/29/e9011b3155d45d70823c141f/statistik-indonesia-2020.html.
Benton, D. J., Wharton, S. A., Martin, S. R., and McCauley, J. W. (2017). Role of Neuraminidase in Influenza A(H7N9) Virus Receptor Binding. J. Virol. 91. doi:10.1128/jvi.02293-16
Bhatta, S., Stevanovic Janezic, T., and Ratti, C. (2020). Freeze-drying of Plant-Based Foods. Foods 9, 87. doi:10.3390/foods9010087
Bianco, A., Brufani, M., Manna, F., and Melchioni, C. (2001). Synthesis of a Carbocyclic Sialic Acid Analogue for the Inhibition of Influenza Virus Neuraminidase. Carbohydr. Res. 332, 23–31. doi:10.1016/S0008-6215(01)00079-9
Biddle, F., and Belyavin, G. (1963). The Haemagglutination Inhibitor in Edible Bird-Nest: its Biological and Physical Properties. J. Gen. Microbiol. 31, 31–44. doi:10.1099/00221287-31-1-31
Big business in bird’s nest in Myanmar seaside town (2017). Big Business in Bird’s Nest in Myanmar Seaside Town. Available at: https://www.thestar.com.my/lifestyle/living/2017/07/01/bird-nest-myanmar.
But, P. P., Jiang, R. W., and Shaw, P. C. (2013). Edible Bird's Nests-Hhow Do the Red Ones Get Red? J. Ethnopharmacol. 145, 378–380. doi:10.1016/j.jep.2012.10.050
Cao (2012). Studies on Immunomodulation Function of Indonesia White Edible Bird’s Nest on Hypoimmune Mice. Acta Nutr. Sin. 34, 168–171. doi:10.13325/j.cnki.acta.nutr.sin.2012.02.018
Careena, S., Sani, D., Tan, S. N., Lim, C. W., Hassan, S., Norhafizah, M., et al. (2018). Effect of Edible Bird's Nest Extract on Lipopolysaccharide-Induced Impairment of Learning and Memory in Wistar Rats. Evidence-Based Complement. Altern. Med. 2018, 1–7. doi:10.1155/2018/9318789
Chan, G. K. L., Zhu, K. Y., Chou, D. J. Y., Guo, A. J. Y., Dong, T. T. X., and Tsim, K. W. K. (2013). Surveillance of Nitrite Level in Cubilose: Evaluation of Removal Method and Proposed Origin of Contamination. Food Control 34, 637–644. doi:10.1016/j.foodcont.2013.06.010
Chen, J. X. J., Lim, P. K. C., Wong, S. F., and Mak, J. W. (2014). Determination of the Presence and Levels of Heavy Metals and Other Elements in Raw and Commercial Edible Bird Nests. Malays. J. Nutr. 20, 377–391. doi:10.1111/anu.12085
Chen, J. X., Wong, S. F., Lim, P. K., and Mak, J. W. (2015). Culture and Molecular Identification of Fungal Contaminants in Edible Bird Nests. Food Addit Contam. Part. A. Chem. Anal. Control. Expo. Risk Assess. 32, 2138–2147. doi:10.1080/19440049.2015.1101494
Choy, K. W., Zain, Z. M., Murugan, D. D., Giribabu, N., Zamakshshari, N. H., Lim, Y. M., et al. (2021). Effect of Hydrolyzed Bird's Nest on β-Cell Function and Insulin Signaling in Type 2 Diabetic Mice. Front. Pharmacol. 12, 632169. doi:10.3389/fphar.2021.632169
Chua, Y. G., Bloodworth, B. C., Leong, L. P., and Li, S. F. (2014). Metabolite Profiling of Edible Bird's Nest Using Gas Chromatography/mass Spectrometry and Liquid Chromatography/mass Spectrometry. Rapid Commun. Mass. Spectrom. 28, 1387–1400. doi:10.1002/rcm.6914
Chua, Y. G., Chan, S. H., Bloodworth, B. C., Li, S. F., and Leong, L. P. (2015). Identification of Edible Bird’s Nest with Amino Acid and Monosaccharide Analysis. J. Agric. Food Chem. 63, 279–289. doi:10.1021/jf503157n
Dai, Y., Cao, J., Wang, Y., Chen, Y., and Jiang, L. (2021). A Comprehensive Review of Edible Bird's Nest. Food Res. Int. 140, 109875. doi:10.1016/j.foodres.2020.109875
Damodaran, S., and Parkin, K. L. (2017). Fennema’s Food Chemistry. fifth edition. Boca Raton, FL: CRC Press. doi:10.1201/9781315372914
Elfita, L., Ietje Wientarsih, I., Dondin Sajuthi, D., Indra Bachtiar, I., and Huda Shalahudin Darusman, H. S. (2020). The Diversity in Nutritional Profile of Farmed Edible Bird's Nests from Several Regions in Indonesia. Biodiversitas 21, 2362–2368. doi:10.13057/biodiv/d210604
Eppe, G., and De Pauw, E. (2009). Advances in Quality Control for Dioxins Monitoring and Evaluation of Measurement Uncertainty from Quality Control Data. J. Chromatogr. B Analyt Technol. Biomed. Life Sci. 877, 2380–2387. doi:10.1016/j.jchromb.2009.05.009
Fan, Q., Wang, P., Zheng, X., Hamzah, S. S., Zeng, H., Zhang, Y., et al. (2020). Effect of Dynamic High Pressure Microfluidization on the Solubility Properties and Structure Profiles of Proteins in Water-Insoluble Fraction of Edible Bird's Nests. Lwt 132, 109923. doi:10.1016/j.lwt.2020.109923
Fan, Y., Fan, Y., Liu, K., Lonan, P., Liao, F., Huo, Y., et al. (2021). Edible Bird's Nest Ameliorates Dextran Sulfate Sodium-Induced Ulcerative Colitis in C57BL/6J Mice by Restoring the Th17/Treg Cell Balance. Front. Pharmacol. 12, 1. doi:10.3389/fphar.2021.632602
Fatima, G., Raza, A. M., Hadi, N., Nigam, N., and Mahdi, A. A. (2019). Cadmium in Human Diseases: It's More Than Just a Mere Metal. Indian J. Clin. Biochem. 34, 371–378. doi:10.1007/s12291-019-00839-8
Gan, S. H., Ong, S. P., Chin, N. L., and Law, C. L. (2017a). Kinetic Retention of Sialic Acid and Antioxidants in Malaysian Edible Bird's Nest during Low-Temperature Drying. Drying Tech. 35, 827–837. doi:10.1080/07373937.2016.1219741
Gan, S. H., Ong, S. P., Chin, N. L., and Law, C. L. (2017b). Retention of Sialic Acid Content in Malaysian Edible Bird’s Nest by Heat Pump Drying. Malaysian J. Vet. Res. 8, 139–153.
Geographical distribution of swiftlet inhabitant (2020). Geographical Distribution of Swiftlet Inhabitant. Available at: https://www.51yanwo.com/jinsiyan/686.html.
Goh, D., Lau, Y., Chew, F., Shek, L., and Lee, B. (1999). Pattern of Food-Induced Anaphylaxis in Children of an Asian Community. Allergy 54, 84–86. doi:10.1034/j.1398-9995.1999.00925.x
Goh, D. L. M., Chua, K. Y., Chew, F. T., Liang, R. C. M. Y., Seow, T. K., Ou, K. L., et al. (2001). Immunochemical Characterization of Edible Bird's Nest Allergens. J. Allergy Clin. Immunol. 107, 1082–1088. doi:10.1067/mai.2001.114342
Greeff-Laubscher, M. R., Beukes, I., Marais, G. J., and Jacobs, K. (2020). Mycotoxin Production by Three Different Toxigenic Fungi Genera on Formulated Abalone Feed and the Effect of an Aquatic Environment on Fumonisins. Mycology 11, 105–117. doi:10.1080/21501203.2019.1604575
Griffiths, M. W., and Schraft, H. (2017). “Bacillus Cereus Food Poisoning,” in Foodborne Diseases. Third Edition. Elsevier, 395–405. doi:10.1016/B978-0-12-385007-2.00020-6
Guo, C.-T., Takahashi, T., Bukawa, W., Takahashi, N., Yagi, H., Kato, K., et al. (2006). Edible Bird's Nest Extract Inhibits Influenza Virus Infection. Antiviral Res. 70, 140–146. doi:10.1016/j.antiviral.2006.02.005
Guo, L., Wu, Y., Liu, M., Wang, B., Ge, Y., and Chen, Y. (2014). Authentication of Edible Bird’s Nests by TaqMan-Based Real-Time PCR. Food Control 44, 220–226. doi:10.1016/j.foodcont.2014.04.006
Ha, E., Basu, N., Bose-O'Reilly, S., Dórea, J. G., McSorley, E., Sakamoto, M., et al. (2017). Current Progress on Understanding the Impact of Mercury on Human Health. Environ. Res. 152, 419–433. doi:10.1016/j.envres.2016.06.042
Haghani, A., Mehrbod, P., Safi, N., Aminuddin, N. A., Bahadoran, A., Omar, A. R., et al. (2016). In Vitro and In Vivo Mechanism of Immunomodulatory and Antiviral Activity of Edible Bird's Nest (EBN) against Influenza A Virus (IAV) Infection. J. Ethnopharmacol. 185, 327–340. doi:10.1016/j.jep.2016.03.020
Haghani, A., Mehrbod, P., Safi, N., Kadir, F. A. i. A., Omar, A. R., and Ideris, A. (2017). Edible Bird's Nest Modulate Intracellular Molecular Pathways of Influenza A Virus Infected Cells. BMC Complement. Altern. Med. 17, 1. doi:10.1186/s12906-016-1498-x
Halimi, N. M., Kasim, Z. M., and Babji, A. S. (2014). Nutritional Composition and Solubility of Edible Bird Nest (Aerodramus Fuchiphagus). AIP Conf. Proc. 1614, 476–481. doi:10.1063/1.4895243
Hamzah, Z., Jeyaraman, S., Ibrahim, N. H., Hashim, O., Lee, B. B., and Hussin, K. (2013). A Rapid Technique to Determine Purity of Edible Bird Nest. Adv. Environ. Biol. 7, 3758–3765.
Hao, L. Q., Ideris, A., Abu Bakar, M. Z., and Omar, A. R. (2015). Morphology Comparison of Swiftlet Species from Natural and Man-Made Habitats in Malaysia. Jsm 44, 497–502. doi:10.17576/jsm-2015-4404-03
Hobbs, J. J. (2004). Problems in the Harvest of Edible Birds' Nests in Sarawak and Sabah, Malaysian Borneo. Biodiversity and Conservation 13, 2209–2226. doi:10.1023/B:BIOC.0000047905.79709.7f
Hong, T. K., Chia Fah, C., and Ong Han, A. K. (2020). Approach to Improve Edible Bird Nest Quality & Establishing Better Bird Nest Cleaning Process Facility through Best Value Approach. japiv 10, 38–50. doi:10.37265/japiv.v10i1.21
Hu, Q., Li, G., Yao, H., He, S., Li, H., Liu, S., et al. (2016). Edible Bird's Nest Enhances Antioxidant Capacity and Increases Lifespan in Drosophila Melanogaster. Cel Mol Biol (Noisy-le-grand) 62, 116–122. doi:10.14715/cmb/2016.62.4.2010.1093/cid/ciw150
Hun, L. T., Hau Lee, C., Alia Azmi, N., Kavita, S., Wong, S., Znati, M., et al. (2020). Characterization of Polar and Non-polar Compounds of House Edible Bird's Nest (EBN) from Johor, Malaysia. Chem. Biodivers. 17, e1900419. doi:10.1002/cbdv.201900419
Hun, L. T., Wani, W. A., Poh, H. Y., Baig, U., Ti Tjih, E. T., Nashiruddin, N. I., et al. (2016). Gel Electrophoretic and Liquid Chromatographic Methods for the Identification and Authentication of Cave and House Edible Bird's Nests from Common Adulterants. Anal. Methods 8, 526–536. doi:10.1039/c5ay02170g
Hwang, E., Park, S. W., and Yang, J.-E. (2020). Anti-aging, Anti-inflammatory, and Wound-Healing Activities of Edible Bird’s Nest in Human Skin Keratinocytes and Fibroblasts. Pharmacogn. Mag. 16, 336–342. doi:10.4103/pm.pm_185_19
Ismail, M., Alsalahi, A., Aljaberi, M. A., Ibrahim, R. M., Bakar, F. A., and Ideris, A. (2021). Efficacy of Edible Bird's Nest on Cognitive Functions in Experimental Animal Models: A Systematic Review. Nutrients 13, 1. doi:10.3390/nu13031028
Kew, P. E., Wong, S. F., Lim, P. K. C., and Mak, J. W. (2014). Structural Analysis of Raw and Commercial Farm Edible Bird Nests. Trop. Biomed. 31, 63’76.
Khushairay, E. S. I., Ayub, M. K., and Babji, A. S. (2014). Effect of Enzymatic Hydrolysis of Pancreatin and Alcalase Enzyme on Some Properties of Edible Bird’s Nest Hydrolysate. AIP Conf. Proc., 427–432. doi:10.1063/1.4895235
Lau, A. S. M., and Melville, D. S. (1994). International Trade in Swiftlet Nests with Special Reference to Hong Kong. Hong Kong: Species in Danger.
Lee, T. H., Wani, W. A., Lee, C. H., Cheng, K. K., Shreaz, S., Wong, S., et al. (2021). Edible Bird’s Nest: The Functional Values of the Prized Animal-Based Bioproduct from Southeast Asia-A Review. Front. Pharmacol. 12, 626233. doi:10.3389/fphar.2021.626233
Lee, T. H., Waseem, W. A., Tan, E. T. T., Nur Ardawati, A., Yong, L. L., and Ramlan, A. A. (2015). Research Article Investigations into the physicochemical , biochemical and antibacterial properties of Edible Bird ’ s Nest. J. Chem. Pharm. Res. 7, 228–247. doi:10.1201/b18814-10
Liang, H., Bao, F., Dong, X., Tan, R., Zhang, C., Lu, Q., et al. (2007). Antibacterial Thymol Derivatives Isolated from Centipeda Minima. Molecules 12, 1606–1613. doi:10.3390/12081606
Lim, C. K., and Earl of Cranbrook, G. G. H. (2002). Swiftlets of Borneo: Builders of Edible Nests. Nat. Hist. Publ. Borneo, 171. Available at: http://myagric.upm.edu.my/id/eprint/10324/.
Ling, J. W. A., Chang, L. S., Babji, A. S., and Lim, S. J. (2020). Recovery of Value‐added Glycopeptides from Edible Bird's Nest ( EBN ) Co‐products: Enzymatic Hydrolysis, Physicochemical Characteristics and Bioactivity. J. Sci. Food Agric. 100, 4714–4722. doi:10.1002/jsfa.10530
Lourie, S. A., and Tompkins, D. M. (2000). The Diets of Malaysian Swiftlets. Ibis (Lond. 1859) 142, 596–602. doi:10.1111/j.1474-919x.2000.tb04459.x
Mahaq, O., P Rameli, M. A., Jaoi Edward, M., Mohd Hanafi, N., Abdul Aziz, S., Abu Hassim, H., et al. (2020). The Effects of Dietary Edible Bird Nest Supplementation on Learning and Memory Functions of Multigenerational Mice. Brain Behav. 10, e01817. doi:10.1002/brb3.1817
Marcone, M. F. (2005). Characterization of the Edible Bird’s Nest the "Caviar of the East". Food Res. Int. 38, 1125–1134. doi:10.1016/j.foodres.2005.02.008
Meijaard, E., Brooks, T. M., Carlson, K. M., Slade, E. M., Garcia-Ulloa, J., Gaveau, D. L. A., et al. (2020). The Environmental Impacts of palm Oil in Context. Nat. Plants 6, 1418–1426. doi:10.1038/s41477-020-00813-w
Minister Department, P. (2012). Economic Transformation Programme Annual Report 2011. Available at: www.pemandu.gov.my.
Moi Chye, S., Koh, R. Y., Kin Tai, S., Yian Koh, R., and Yen Ng, K. (2017). A Mini Review on Medicinal Effects of Edible Bird's Nest. Lhbs 2, 1–3. doi:10.15436/2475-6245.17.016
Murugan, D. D., Md Zain, Z., Choy, K. W., Zamakshshari, N. H., Choong, M. J., Lim, Y. M., et al. (2020). Edible Bird's Nest Protects against Hyperglycemia-Induced Oxidative Stress and Endothelial Dysfunction. Front. Pharmacol. 10, 1624. doi:10.3389/fphar.2019.01624
Ng, M. H., Chan, K. H., and Kong, Y. C. (1986). Potentiation of Mitogenic Response by Extracts of the Swiftlet's (Collocalia) Nest. Biochem. Int. 13, 521–531.
Noor, H. S. M., Babji, A. S., and Lim, S. J. (2018). Nutritional Composition of Different Grades of Edible Bird's Nest and its Enzymatic Hydrolysis. AIP Conf. Proc. 1940, 020088. doi:10.1063/1.5028003
Norhayati, M. K., Azman, O., and Wan Nazaimoon, W. (2010). Preliminary Study of the Nutritional Content of Malaysian Edible Bird's Nest. Malays. J. Nutr. 16, 389–396.
Oda, M., Ohta, S., Suga, T., and Aoki, T. (1998). Study on Food Components: The Structure ofN-Linked Asialo Carbohydrate from the Edible Bird's Nest Built byCollocalia Fuciphaga. J. Agric. Food Chem. 46, 3047–3053. doi:10.1021/jf980094k
Ogura, H. (2011). Development of Miracle Medicines from Sialic Acids. Proc. Jpn. Acad. Ser. B: Phys. Biol. Sci. 87, 328–361. doi:10.2183/pjab.87.328
Panyaarvudh, J. (2018). An Economic Nesting Ground. The Nation. Available at: https://www.nationthailand.com/business/30356222.
Paydar, M., Wong, Y. L., Wong, W. F., Hamdi, O. A., Kadir, N. A., and Looi, C. Y. (2013). Prevalence of Nitrite and Nitrate Contents and its Effect on Edible Bird Nest's Color. J. Food Sci. 78, T1940–T1947. doi:10.1111/1750-3841.12313
Phyu Win, P., Chaw Su, M., Yi Linn, W., Yi Myint, Y., and Myo Win, M. (2020). Investigations into the Physicochemical Properties of Raw Edible Bird's Nest and Preparation of Double Boiled Soup. Ijrp 62, 1. doi:10.47119/ijrp1006211020201467
Quek, M. C., Chin, N. L., Yusof, Y. A., Law, C. L., and Tan, S. W. (2018a). Characterization of Edible Bird's Nest of Different Production, Species and Geographical Origins Using Nutritional Composition, Physicochemical Properties and Antioxidant Activities. Food Res. Int. 109, 35–43. doi:10.1016/j.foodres.2018.03.078
Quek, M. C., Chin, N. L., Yusof, Y. A., Law, C. L., and Tan, S. W. (2018b). Pattern Recognition Analysis on Nutritional Profile and Chemical Composition of Edible Bird's Nest for its Origin and Authentication. Int. J. Food Properties 21, 1680–1696. doi:10.1080/10942912.2018.1503303
Rohaizan, M. A. (2017). Application for export of Bird Nest Products to P. R. China Putrajaya. Malaysia: Bird Nest Exporters.
Saengkrajang, W., Matan, N., and Matan, N. (2011). Antimicrobial Activities of the Edible Bird’s Nest Extracts against Food-Borne Pathogens. Thai J. Agric. Sci. 44, 326–330. doi:10.1016/j.jfca.2013.05.001
Saengkrajang, W., Matan, N., and Matan, N. (2013). Nutritional Composition of the Farmed Edible Bird's Nest (Collocalia Fuciphaga) in Thailand. J. Food Compost. Anal. 31, 41–45. doi:10.1016/j.jfca.2013.05.001
Sanchez-Borges, M., Capriles-Hulett, A., Fernandez-Caldas, E., Suarez-Chacon, R., Caballero, F., Castillo, S., et al. (1997). Mite-contaminated Foods as a Cause of Anaphylaxis. J. Allergy Clin. Immunol. 99, 738–743. doi:10.1016/S0091-6749(97)80005-X
Schultze, M., and Kondorosi, A. (1998). Regulation of Symbiotic Root Nodule Development. Annu. Rev. Genet. 32, 33–57. doi:10.1146/annurev.genet.32.1.33
Seeboonruang, U. (2012). A Statistical Assessment of the Impact of Land Uses on Surface Water Quality Indexes. J. Environ. Manage. 101, 134–142. doi:10.1016/j.jenvman.2011.10.019
Shi, J., Hu, X., Zou, X., Zhao, J., Zhang, W., Holmes, M., et al. (2017). A Rapid and Nondestructive Method to Determine the Distribution Map of Protein, Carbohydrate and Sialic Acid on Edible Bird's Nest by Hyper-Spectral Imaging and Chemometrics. Food Chem. 229, 235–241. doi:10.1016/j.foodchem.2017.02.075
Shim, E. K., Chandra, G. F., and Lee, S. Y. (2017). Thermal Analysis Methods for the Rapid Identification and Authentication of Swiftlet (Aerodramus Fuciphagus) Edible Bird's Nest - A Mucin Glycoprotein. Food Res. Int. 95, 9–18. doi:10.1016/j.foodres.2017.02.018
Shim, E. K., and Lee, S. Y. (2020). Calcite Deposits Differentiate Cave from House-Farmed Edible Bird's Nest as Shown by SEM-EDX, ATR-FTIR and Raman Microspectroscopy. Chem. Asian J. 15, 2487–2492. doi:10.1002/asia.202000520
Stadler, R. H., Mottier, P., Guy, P., Gremaud, E., Varga, N., Lalljie, S., et al. (2004). Semicarbazide Is a Minor thermal Decomposition Product of Azodicarbonamide Used in the Gaskets of Certain Food Jars. Analyst 129, 276–281. doi:10.1039/b314206j
Tai, S. K., Hamzah, Z., Ng, Q. H., and Tan, C. S. (2020). Surface Morphology Study on Unclean, Commercial and Bromelain Treated Edible Bird Nest (EBN) Using Scanning Electron Microscope. IOP Conf. Ser. Mater. Sci. Eng. 932, 012013. doi:10.1088/1757-899X/932/1/012013
Tan, S. N., Sani, D., Lim, C. W., Ideris, A., Stanslas, J., and Lim, C. T. S. (2020). Proximate Analysis and Safety Profile of Farmed Edible Bird's Nest in Malaysia and its Effect on Cancer Cells. Evid. Based Complement. Alternat Med. 2020, 8068797. doi:10.1155/2020/8068797
Tavano, O. L. (2013). Protein Hydrolysis Using Proteases: An Important Tool for Food Biotechnology. J. Mol. Catal. B: Enzymatic 90, 1–11. doi:10.1016/j.molcatb.2013.01.011
Teodorowicz, M., Van Neerven, J., and Savelkoul, H. (2017). Food Processing: The Influence of the Maillard Reaction on Immunogenicity and Allergenicity of Food Proteins. Nutrients 9, 835. doi:10.3390/nu9080835
Thomas, I., Siew, L. Q. C., Watts, T. J., and Haque, R. (2019). Seaweed Allergy. J. Allergy Clin. Immunol. Pract. 7, 714–715. doi:10.1016/j.jaip.2018.11.009
Thorburn, C. C. (2015). The Edible Nest Swiftlet Industry in Southeast Asia: Capitalism Meets Commensalism. Hum. Ecol. 43, 179–184. doi:10.1007/s10745-014-9713-1
Tong, S. R., Lee, T. H., Cheong, S. K., and Lim, Y. M. (2020). Untargeted Metabolite Profiling on the Water-Soluble Metabolites of Edible Bird's Nest through Liquid Chromatography-Mass Spectrometry. Vet. World 13, 304–316. doi:10.14202/vetworld.2020.304-316
Tres, A., Ruiz-Samblas, C., Van Der Veer, G., and Van Ruth, S. M. (2013). Geographical Provenance of palm Oil by Fatty Acid and Volatile Compound Fingerprinting Techniques. Food Chem. 137, 142–150. doi:10.1016/j.foodchem.2012.09.094
Tukiran, N. A., Ismail, A., Mustafa, S., and Hamid, M. (2015). Enzyme Immunoassay for the Detection of Porcine Gelatine in Edible Bird's Nests. Food Addit Contam. Part. A. Chem. Anal. Control. Expo. Risk Assess. 32, 1023–1028. doi:10.1080/19440049.2015.1039605
Wong, S. F., Lim, P. K. C., Mak, J. W., Ooi, S. S., and Chen, D. K. F. (2018a). Molecular Characterization of Culturable Bacteria in Raw and Commercial Edible Bird Nests (EBNs). Int. Food Res. J. 25, 966–974.
Wong, Z. C. F., Chan, G. K. L., Dong, T. T. X., and Tsim, K. W. K. (2018b). Origin of Red Color in Edible Bird's Nests Directed by the Binding of Fe Ions to Acidic Mammalian Chitinase-like Protein. J. Agric. Food Chem. 66, 5644–5653. doi:10.1021/acs.jafc.8b01500
Wong, Z. C. F., Chan, G. K. L., Wu, K. Q. Y., Poon, K. K. M., Chen, Y., Dong, T. T. X., et al. (2018c). Complete Digestion of Edible Bird's Nest Releases Free N-Acetylneuraminic Acid and Small Peptides: an Efficient Method to Improve Functional Properties. Food Funct. 9, 5139–5149. doi:10.1039/c8fo00991k
Xing, Y. N., Ni, H. G., and Chen, Z. Y. (2012). Semicarbazide in Selected Bird's Nest Products. J. Food Prot. 75, 1654–1659. doi:10.4315/0362-028X.12-065
Xu, C. J., Liang, Y. Z., Chau, F. T., and Heyden, Y. V. (2006). Pretreatments of Chromatographic Fingerprints for Quality Control of Herbal Medicines. J. Chromatogr. A. 1134, 253–259. doi:10.1016/j.chroma.2006.08.060
Yan, T. S., Elizabeth, T., Tzien, Y. C., Cheng, Y. F., Jiayi, C., Nge, C., et al. (2008). Anaphylaxis Following the Ingestion of Flour Contaminated by House Dust Mites - A Report of Two Cases from Singapore. Asian Pac. J. Allergy Immunol. 26, 165–170.
Yang, M., Cheung, S. H., Li, S. C., and Cheung, H. Y. (2014). Establishment of a Holistic and Scientific Protocol for the Authentication and Quality Assurance of Edible Bird's Nest. Food Chem. 151, 271–278. doi:10.1016/j.foodchem.2013.11.007
Yeo, B.-H., Tang, T.-K., Wong, S.-F., Tan, C.-P., Wang, Y., Cheong, L.-Z., et al. (2021). Potential Residual Contaminants in Edible Bird's Nest. Front. Pharmacol. 12, 1–15. doi:10.3389/fphar.2021.631136
Yew, M. Y., Koh, R. Y., Chye, S. M., Othman, I., and Ng, K. Y. (2014). Edible Bird's Nest Ameliorates Oxidative Stress-Induced Apoptosis in SH-Sy5y Human Neuroblastoma Cells. BMC Complement. Altern. Med. 14, 1. doi:10.1186/1472-6882-14-391
Yew, M. Y., Koh, R. Y., Chye, S. M., Othman, I., Soga, T., Parhar, I., et al. (2018). Edible Bird's Nest Improves Motor Behavior and Protects Dopaminergic Neuron against Oxidative and Nitrosative Stress in Parkinson’s Disease Mouse Model. J. Funct. Foods 48, 576–585. doi:10.1016/j.jff.2018.07.058
Yew, M. Y., Koh, R. Y., Chye, S. M., Zainal Abidin, S. A., Othman, I., and Ng, K. Y. (2019). Neurotrophic Properties and the De Novo Peptide Sequencing of Edible Bird's Nest Extracts. Food Biosci. 32, 100466. doi:10.1016/j.fbio.2019.100466
Yida, Z., Imam, M. U., and Ismail, M. (2014). In Vitro bioaccessibility and Antioxidant Properties of Edible Bird's Nest Following Simulated Human Gastro-Intestinal Digestion. BMC Complement. Altern. Med. 14, 468. doi:10.1186/1472-6882-14-468
Zainal Abidin, F., Hui, C. K., Luan, N. S., Mohd Ramli, E. S., Hun, L. T., and Abd Ghafar, N. (2011). Effects of Edible Bird's Nest (EBN) on Cultured Rabbit Corneal Keratocytes. BMC Complement. Altern. Med. 11, 1. doi:10.1186/1472-6882-11-94
Zhang, H., Ha, T. M. H., Seck, H. L., and Zhou, W. (2020). Inactivation of Escherichia coli O157:H7 and Salmonella Typhimurium in Edible Bird's Nest by Low-Energy X-ray Irradiation. Food Control 110, 107031. doi:10.1016/j.foodcont.2019.107031
Zhang, M., Wang, D. S., and Wang, J. (1994). The Effect of the ZHENZHU-YANWO Extracts on Animal Function. Pharm. Biotechnol. 1, 49–51.
Zhao, R., Li, G., Kong, X. J., Huang, X. Y., Li, W., Zeng, Y. Y., et al. (2016). The Improvement Effects of Edible Bird's Nest on Proliferation and Activation of B Lymphocyte and its Antagonistic Effects on Immunosuppression Induced by Cyclophosphamide. Drug Des. Devel. Ther. 10, 371–381. doi:10.2147/DDDT.S88193
Keywords: edible bird’s nest, industry insights, laboratory findings, saliva of swiftlets, bioactive compounds, nutritional contents
Citation: Chok KC, Ng MG, Ng KY, Koh RY, Tiong YL and Chye SM (2021) Edible Bird’s Nest: Recent Updates and Industry Insights Based On Laboratory Findings. Front. Pharmacol. 12:746656. doi: 10.3389/fphar.2021.746656
Received: 24 July 2021; Accepted: 20 September 2021;
Published: 01 October 2021.
Edited by:
Ting Hun Lee, University of Technology Malaysia, MalaysiaReviewed by:
Aline Priscilla Gomes Da Silva, Michigan State University, United StatesCopyright © 2021 Chok, Ng, Ng, Koh, Tiong and Chye. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Soi Moi Chye, Y2h5ZV9zb2ltb2lAaW11LmVkdS5teQ==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.