 Annagrazia Adornetto1*
Annagrazia Adornetto1* Carlo Gesualdo2
Carlo Gesualdo2 Maria Luisa Laganà1
Maria Luisa Laganà1 Maria Consiglia Trotta3
Maria Consiglia Trotta3 Settimio Rossi2
Settimio Rossi2 Rossella Russo1
Rossella Russo1- 1Preclinical and Translational Pharmacology, Department of Pharmacy, Health and Nutritional Sciences, University of Calabria, Rende, Italy
- 2Multidisciplinary Department of Medical, Surgical and Dental Sciences, University of Campania “Luigi Vanvitelli”, Naples, Italy
- 3Department of Experimental Medicine, University of Campania “Luigi Vanvitelli”, Naples, Italy
Autophagy is the major catabolic pathway involved in removing and recycling damaged macromolecules and organelles and several evidences suggest that dysfunctions of this pathway contribute to the onset and progression of central and peripheral neurodegenerative diseases. Diabetic retinopathy (DR) is a serious complication of diabetes mellitus representing the main preventable cause of acquired blindness worldwide. DR has traditionally been considered as a microvascular disease, however this concept has evolved and neurodegeneration and neuroinflammation have emerged as important determinants in the pathogenesis and evolution of the retinal pathology. Here we review the role of autophagy in experimental models of DR and explore the potential of this pathway as a target for alternative therapeutic approaches.
Introduction
Diabetic retinopathy (DR), a chronic and progressive complication of diabetes mellitus, is the main cause of legal blindness in working-age population (20–65 years old) (Ting et al., 2016; Simo-Servat et al., 2019). DR is prompted by hyperglycemia, which causes an increase of oxidative stress leading to an adaptive inflammatory response in microvasculature and neuroretinal tissue (Saxena et al., 2016; Al-Kharashi, 2018). The disease has long been considered as a microvascular disease, since loss of pericytes, damage of vascular endothelial cells and breakdown of blood-retinal barrier (BRB) are typical hallmarks of the early stage of the pathology (Beltramo and Porta, 2013; Mrugacz et al., 2021). However, recent and intensive research identified neurodegeneration and neuroinflammation as processes involved in the pathogenesis and evolution of DR (Kadlubowska et al., 2016). Furthermore, experimental and clinical studies have shown that neuronal apoptosis and reactive gliosis, with thinning of the nerve fiber layer often precede the typical vascular alterations (Barber et al., 2011; Gu et al., 2019). Importantly, DME (diabetic macular edema), which is due to an abnormal intraretinal fluid collection in the macular area, is the most common cause of vision loss in patients with DR (Romero-Aroca et al., 2016). Experimental and clinical studies have highlighted the role of inflammation in DME and OCT-imaging biomarkers of retinal inflammation have been identified (Ceravolo et al., 2020).
The mechanisms underlying the neurodegenerative and neuroinflammatory processes occurring in DR are common to other central and retinal diseases, like glaucoma and retinitis pigmentosa (Baumgartner, 2000; Barber, 2003; Gupta and Yücel, 2007). These mechanisms include oxidative stress and free radical formation, advanced glycation end products (AGEs) production, glutamate excitotoxicity, mitochondrial dysfunction, impaired bioenergetics, dysfunction of neurotrophin signals and autophagy (Dong et al., 2009; Jellinger, 2010; Rosa et al., 2016).
Autophagy is a major lysosomal pathway for the turnover of cytoplasmic organelles and long-lived proteins and, besides its homeostatic functions, it also acts as an adaptive response to cellular stresses (Mizushima, 2007). Dysfunctions of this process have been identified as recurrent events in neurodegenerative disorders (Frake et al., 2015) and, more recently, experimental and clinical data have shown that autophagy modulation also occurs in experimental models of DR and in the retina of diabetic patients, with or without retinopathy (Lopes de Faria et al., 2016; Dehdashtian et al., 2018).
However, the functional role of autophagy in DR remains unclear. Here we discuss the available literature on the role of autophagy in experimental models of DR and explore the potential of this pathway as a target for alternative therapeutic approaches.
Diabetic Retinopathy: A Neurodegenerative Retinal Disease
DR is a social disease with considerable costs, whose global incidence is strongly increasing due to the improved life expectancy and the exponential spread of diabetes (Flaxman et al., 2017). Indeed in 2015, 415 million people were affected by diabetes globally and this number is projected to reach 642 million by 2040 (Ogurtsova et al., 2017). In addition, it has been estimated that more than a third of people with diabetes worldwide have some form of DR and that nearly one in 10 develops forms of DR or complications that are particularly threatening for the sight such as proliferative DR or diabetic macular edema (Yau et al., 2012).
The diagnosis of DR is made on the bases of typical vascular abnormalities following the clinical examination of ocular fundus; it is possible to distinguish two stages: the non-proliferative DR (NPDR) and the proliferative DR (PDR). NPDR, the earliest form of DR, is divided into three stages of increasing severity, namely: 1) the mild DR characterized by rare microaneurysms and hemorrhages; 2) the moderate DR characterized by an increase in the aforementioned lesions associated with hard exudates; 3) the severe or pre-proliferating DR characterized by the coexistence of numerous microaneurysms, retinal hemorrhages, cottony nodules and venous caliber anomalies (Singh et al., 2008; Karst et al., 2018). At this stage, diabetic subjects can sometimes be asymptomatic for long time. On the other hand, in PDR, characterized by the appearance of epiretinal or epi-papillary new vessels that can sometimes invade the vitreous, patients may present a sudden vision impairment due to vitreous hemorrhages and/or tractional retinal detachment (Gotzaridis et al., 2001). Both forms of DR can be further complicated by macular edematous (DME) and/or ischemic damage, which are the main causes of severe vision impairment (Wilkinson et al., 2003; Cheung et al., 2010; Wang and Lo, 2018).
The onset of typical DR vascular changes is determined by prolonged hyperglycemic episodes resulting from suboptimal glycemic control in patients with type I or II diabetes mellitus. Elevated blood glucose levels result in aberrant regulation of a number of biochemical pathways, eventually leading to superoxide production and overload of oxidative stress in retinal tissues. Prolonged hyperglycemia has been shown to cause increased flow of the polyol pathway (Gabbay et al., 1966; Lee and Chung, 1999), increased formation of AGEs (Shinohara et al., 1998; Stitt, 2010), abnormal activation of signaling cascades like protein kinase C (PKC) pathway (Koya and King, 1998; Idris et al., 2001), increase in the flux of the hexosamine pathway (Kolm-Litty et al., 1998; Du et al., 2000) and reactive oxygen species (ROS) (Brownlee, 2005). All these changes lead to an intensification in oxidative stress and an inflammatory attack on the retina with consequent structural and functional changes (Kowluru and Chan, 2007; Hammes, 2018) (Figure 1).
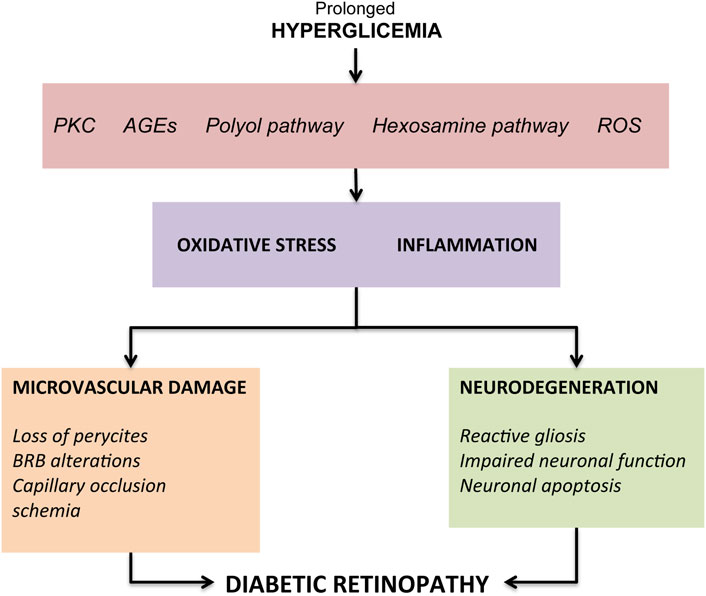
FIGURE 1. Overview of the pathogenic mechanisms leading to diabetic retinopathy.
The first responses, considered as metabolic self-regulation to increase retinal metabolism, are vessel dilation and changes in blood flow (Bek, 2017). Another hallmark of early DR is the loss of pericytes which has been demonstrated in both in vitro and in vivo studies (Romeo et al., 2002). Since pericytes provide structural support to capillaries, their loss leads to the formation of microaneurysms, which represents the first clinical characteristic sign of DR (Ejaz et al., 2008). Other pathogenetic processes found during DR include endothelial cells apoptosis and thickening of the basement membrane, which overall contribute to compromise the BRB integrity (Beltramo and Porta, 2013). Finally, the loss of pericytes and endothelial cells causes capillaries occlusion and consequent ischemia. Retinal ischemia, through the activation of hypoxia-inducible factor-1 (HIF-1) (Huang et al., 2015), determines an overproduction of vascular endothelial growth factor (VEGF), a key factor involved in both the progression of retinopathy towards PDR, and in DME development through phosphorylation of tight junction proteins such as occludin and zonula occludens-1 (Antonetti et al., 1999). Furthermore, VEGF, by the activation of the mitogen activated protein (MAP), stimulates the proliferation of endothelial cells, resulting in new vessels development (Rousseau et al., 1997). The key role of VEGF in DR has been demonstrated in multiple studies showing its increased expression in diabetic mice retina (Li et al., 2010; Rossi et al., 2016), as well as in the vitreous of patients with DME and PDR (Adamis et al., 1994). In light of these evidence, the intravitreal injection of anti-VEGF agents is currently the gold standard for both early and advanced stages DR therapy (Sun and Jampol, 2019). Other therapeutic tools aimed at managing the microvascular complications of DR are steroid intravitreal injection, laser photocoagulation and vitreous surgery (Stitt et al., 2016; Wang and Lo, 2018; Sun and Jampol, 2019). However, although these treatments demonstrate clinical benefits, no tools are effective in completely blocking clinical progression or reversing retinal damage. In fact, such therapies are often used in the more advanced stages of DR, characterized by a high risk of irreversible and severe visual impairment.
Furthermore, in many cases frequent administration of intravitreal agents is necessary with a consequent increased risk of side effects related to the injection, not to mention the high costs associated with frequent eye examinations (Donnelly et al., 2004; Gupta et al., 2013).
For many years, microangiopathic lesions were considered the exclusive cause of DR, leading to the visual loss in diabetic patients. However, the concept of DR as a microvascular disease has evolved: nowadays, it is considered a more complex diabetic complication, in which neurodegeneration has emerged as an important factor, playing a significant role in DR pathogenesis and evolution (Ola and Alhomida, 2014; Jindal, 2015). Indeed, the American Diabetes Association (ADA) recently defined DR as a diabetes neurovascular complication that involves a progressive disruption of the interdependence between multiple cell types in the retina (Solomon et al., 2017).
The hallmarks of diabetes-induced neuroglial degeneration, which include reactive gliosis, impaired retinal neuronal function and apoptosis of neural cells, have been described before typical microangiopathy in multiple experimental models of DR and also in the retina of diabetic donors (Barber et al., 1998; Lieth et al., 1998; Lung et al., 2012; Howell et al., 2013; Jindal, 2015).
The first retinal neurons affected are retinal ganglion cells (RGCs) and amacrine cells, however photoreceptors also show an increased apoptosis (Lynch and Abràmoff, 2017). The structural consequence of this early death is a reduction in the ganglion cell layer (GCL) thinning and corresponding loss of nerve fiber layer (NFL) thickness, detected by optical coherence tomography (OCT) (van Dijk et al., 2010; Sohn et al., 2016). Moreover, functional studies performed with multifocal electroretinography (mfERG) have shown a delayed implicit time P1 and a reduction in the trace’s amplitude as a consequence of the early neurodegenerative process (Simão et al., 2017). These structural and functional alterations lead to reduced contrast sensitivity, delayed dark adaptation and altered visual fields, resulting overall in reduced vision-related quality of life, despite the absence of clinically detectable vascular anomalies (Wolff et al., 2015; Trento et al., 2017).
Müller cells and retinal astrocytes play an important role in the damage to retinal neurons and in linking the neurodegenerative process with vascular disease. Indeed, gliosis is associated with higher expression of VEGF and hyper-activation of pro-inflammatory pathways, with consequent overexpression of pro-inflammatory cytokines and dysfunction of the BRB (Bringmann and Wiedemann, 2012). Diabetes-induced subclinical inflammation is further amplified by the activation of immune cells resident in the retina, namely microglial cells. This microglial activation is accompanied by a phenotypic shift from the anti-inflammatory (M2) towards a pro-inflammatory amoeboid (M1) form (Coorey et al., 2012; Arroba and Valverde Á, 2017). This shift results in transcriptional changes mediated by nuclear factor-kappa B (NF-κB) and extracellular signaling mechanisms of the signal-regulated kinase (ERK) responsible for the release or activation of pro-inflammatory and neurotoxic molecules (i.e. cytokines, chemokines, glutamate) which contributes to the disruption of BRB and neuronal death (Altmann and Schmidt, 2018).
Autophagy: Mechanisms and Functions
Autophagy is a highly conserved catabolic pathway by which cells remove misfolded or aggregated proteins and damaged organelles (Klionsky et al., 2021). This process regulates essential biological functions such as cell survival, cell metabolism, development, aging, and immunity. It also represents an adaptive response to different forms of stresses, like nutrient deprivation, growth factor depletion, infection, hypoxia, ischemia/reperfusion injury, oxidative stress, endoplasmic reticulum (ER) stress and mitochondrial damage (Glick et al., 2010; Dikic and Elazar, 2018).
In mammalian cells, there are three primary types of autophagy: microautophagy, macroautophagy, and chaperone-mediated autophagy (CMA) (Yang and Klionsky, 2010). Furthermore, different selective forms of autophagy, such as mitophagy, ribophagy or aggrephagy, have also been identified (Menzies et al., 2017).
In microautophagy, cytosolic components are directly taken up by lysosomes through the invagination of their membrane (Li et al., 2012). CMA, involves the formation of a complex between target proteins (identified by bearing a CMA targeting motif) and chaperones of the Hsp70 family; these complexes are recognized by the lysosome-associated membrane protein type-2A (LAMP-2A) at the lysosomal membrane where the substrate proteins unfold and translocate in the lumen for degradation by lysosomal hydrolase (Itakura and Mizushima, 2010).
Macroautophagy (hereafter referred to as autophagy) involves the formation of a cup-shaped membrane structure, the phagophore, that elongates and closes around the cytosolic cargo; the resulted double-membrane vesicle is called autophagosome and it is selectively associated with this pathway (Yang and Klionsky, 2010). Autophagosomes are transported, along the microtubules, to the perinuclear region where they fuse with lysosomes; here the autophagic content is degraded and released for recycling into the cytoplasm (Parzych and Klionsky, 2014). Autophagy and its regulatory mechanisms are evolutionarily conserved among eukaryotic cells even if the level of complexity of the process may differ (Yang and Klionsky, 2009).
Autophagosome biogenesis is orchestrated by the sequential action of autophagy-related (Atg) proteins; most of them were originally identified in yeast but have their homologs in mammalian cells (Mizushima et al., 2011). The ULK1 complex, formed by the serine/threonine protein kinase Atg1/(unc-51-like kinase 1), FIP200 (focal adhesion kinase family interacting protein of 200 kDa), Atg13 and Atg101 is involved in the initiation of autophagy (Hara et al., 2008). Upon autophagy induction, the mammalian target of rapamycin (mTOR), one of the main negative regulators of the process, is inactivated resulting in upregulation of ULK1 kinase activity and consequent phosphorylation of Atg13 and FIP200 (Noda, 2017). ULK1 complex gathers to specific ER region engaged in autophagosome formation (Itakura and Mizushima, 2010) and regulates the recruitment of a second kinase complex, the vacuolar protein sorting 34 (Vps34) complex formed by Beclin-1, AMBRA, Vps34, Vps15 and Atg14 (Glick et al., 2010). Vps34 participates in various membrane-sorting processes but it is selectively involved in autophagy when complexed to Beclin-1 (Backer, 2008). At variance with the other PI3-kinases, Vps34 only uses phosphatidylinositol (PI) as substrate to generate phosphatidyl inositol triphosphate (PI3P), which is therefore essential for phagophore nucleation, elongation and recruitment of other Atg proteins (Xie and Klionsky, 2007). The interaction of Beclin-1 with Vps34 promotes its catalytic activity and increases PI3P levels (Glick et al., 2010).
Following the initiation step, the elongation process is undertaken by two ubiquitin like proteins: Atg12 and Atg8/LC3. In this system, the E1-like enzyme Atg7 and E2-like enzyme Atg10 catalyze the formation of the Atg12-Atg5 complex that allows the formation of the Atg12-Atg5-Atg16 (L1) complex. The latter is crucial for autophagosome formation and for efficient promotion of the microtubule-associated protein light chain 3 (LC3) lipidation (Otomo et al., 2013).
Several experimental evidences demonstrate that LC3 is involved in the selective identification of autophagy substrates (Yoshii and Mizushima, 2017; Mizushima and Murphy, 2020). Indeed, LC3-II interacts with the constitutively expressed adaptor molecule p62 (or sequestosome-1, SQSTM1) that contains both a ubiquitin binding domain and a LC3-interacting (LIR) domain to deliver sequestered proteins to the autophagosomes (Zhang et al., 2015).
The fusion of the autophagosomal membrane with lysosome results in the release of a single-membrane autophagic body into the lysosomal lumen, which is followed by the degradation of the autophagic cargo by the lysosomal acid proteases (Dikic and Elazar, 2018).
Cellular homeostasis depends on the balance between the production and removal of macromolecules and organelles. In this context, basal autophagy activity plays a key role in the maintenance of cellular integrity (Chun and Kim, 2018). As a quality control mechanism, the process is fundamental for every cell, but it is particularly important in neurons. Indeed, neuronal cells are metabolically highly active and, being post mitotic cells, cannot dilute damaged or aged organelles and misfolded proteins by cell division (Mariño et al., 2011; Russo et al., 2013). Therefore, not surprisingly, accumulation of these altered components, due to autophagy inefficiency, has been associated with neurotoxicity and neurodegeneration.
Autophagy disruption or insufficiency has been reported in a number of different ocular diseases and pathological conditions like: retinal injury (Besirli et al., 2011), retinal degenerations (Punzo et al., 2009; Rodríguez-Muela et al., 2015), light-induced stress (Kunchithapautham et al., 2011; Chen et al., 2014), hyperglycemia (Lopes de Faria et al., 2016) and hypoglycemia (Zhou et al., 2015).
In this context, autophagy is becoming an attractive target to treat neurodegenerative disorders (Zhu et al., 2013), including the ones affecting the retina (Russo et al., 2013).
The Role of Autophagy in Animal Models of Diabetic Retinopathy
Several groups have reported a modulation of Atg proteins in animal models of TD1 and TD2 diabetes (Table 1). In C57BL/6J mice, induction of type1 (T1D) diabetes by administration of streptozotocin (STZ) was associated with increased LC3-II immunoreactivity in the outer plexiform layer (OPL) and upregulation of the Atg related proteins, Beclin-1 and Atg5 (Piano et al., 2016). These changes occurred within the same time frame of outer retinal damage and might take part to the process of photoreceptors loss in the early phase of DR, before the appearance of evident signs of vascular damage (Piano et al., 2016). Accordingly, upregulation of Beclin-1, LC3-II, Atg12-Atg5 and Atg3 was reported in STZ-diabetic rats and Ins2Akita mice, a spontaneous T1D mouse model (Wang et al., 2017). In this study knockdown of Hist1h1c, a gene that encoded for Histone H1.2 protein, significantly reduced both basal and high-glucose-induced autophagy, attenuated inflammation and cell toxicity. Conversely, adeno-associated virus (AAV)-mediated histone HIST1H1C overexpression led to increased autophagy, glial activation and neuronal loss which are pathological changes identified in the early stages of DR (Wang et al., 2017). These findings suggest that over-stimulation of autophagy is associated with increased retinal cell death and takes part to the progression of DR through advanced stages.
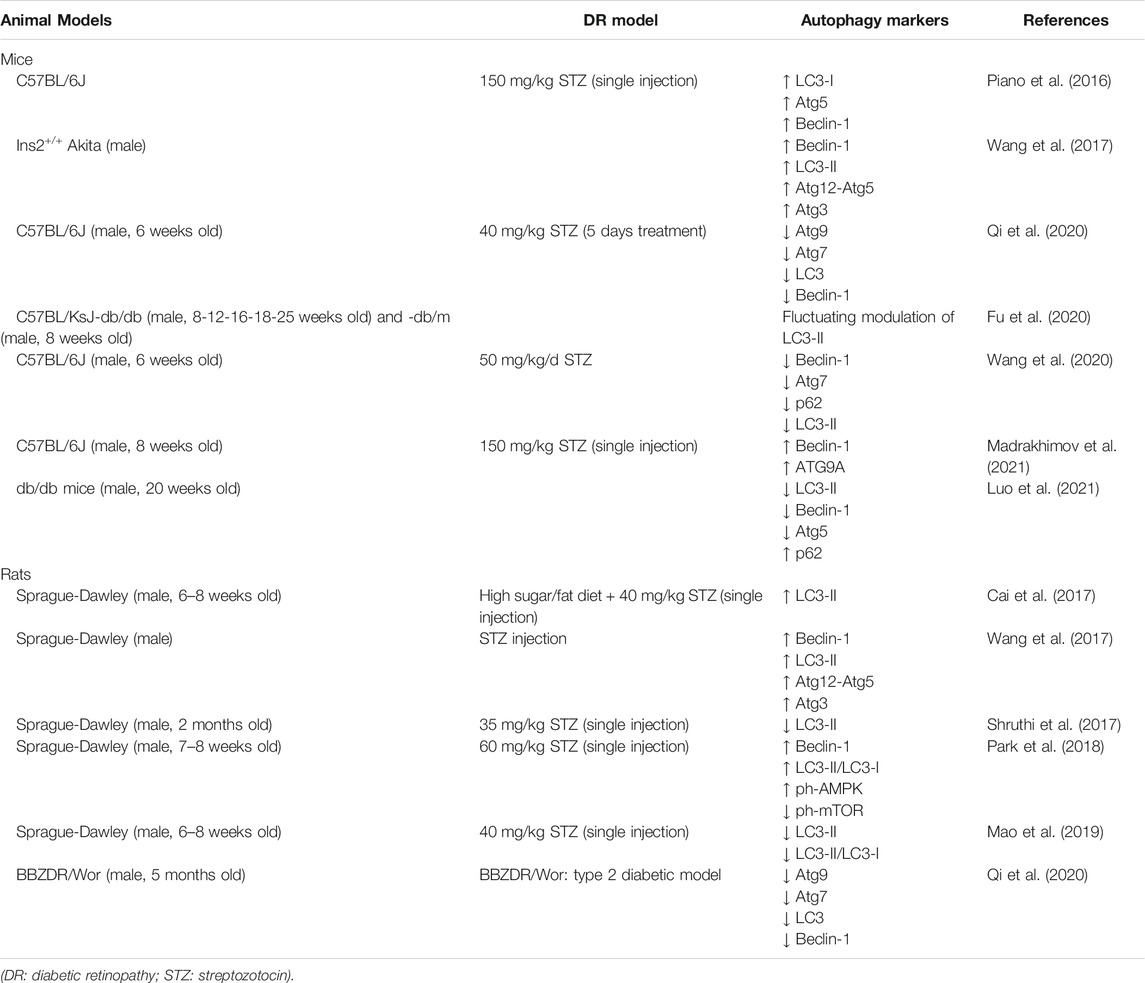
TABLE 1. Autophagy modulation in animal models of diabetic retinopathy.
A very recent study by Madrakhimov and colleagues demonstrated that long-term hyperglycemia causes mTOR inhibition leading to autophagy dysregulation (Madrakhimov et al., 2021). Indeed, inhibition of the mTORC1 pathway in STZ-induced diabetic mice was associated with upregulation of Beclin-1 in the entire inner retina and ATG9A in NeuN (Neuronal Nuclei) positive RGCs. These changes were accompanied with signs of neuronal cell damage, such as activation of cleaved caspase three and decrease of the total number of cells in the GCL. Interestingly, blockade of autophagy by mTOR activator-MHY1485 injections in diabetic mice resulted in a prominent rescue of neuronal cells (Madrakhimov et al., 2021).
Increased LC3-II expression was also reported by Cai and colleagues (2017) in male rats fed with sugar, high fat diet followed by STZ injection; in this model treatment with Glucagon-like peptide-1 (GLP-1), reduced oxidative stress and reverted the upregulation of LC3-II expression (Cai et al., 2017).
At variance with the previous reported results, in the retina of STZ-induced diabetic mice a decrease of Beclin-1, Atg7, p62 and LC3-II expression was reported as compared to control group; treatment with the heparanase inhibitor PG545 promoted autophagy and inhibited the secretion of pro-inflammatory cytokines alleviating diabetic retinopathy (Wang et al., 2020).Similarly, in STZ-induced diabetic C57BL/6J mice, as well as in Bio-Breeding Zucker diabetic (BBZDR/Wor) rats that spontaneously develop a T2D, Qi and colleagues reported a dramatic reduction of Atg7, Atg9, LC3 and Beclin-1 in diabetic retina as compared to controls (Qi et al., 2020).
Interestingly, in this same study the authors showed a diurnal rhythmicity of Atg proteins levels. Under basal conditions Atg9 and LC3 expression showed a biphasic diurnal cycle with two peaks of highest and lowest levels, respectively, while Atg7 and Beclin-1 had a monophasic 24 h cycle. In the retinas from both T1D and T2D mice a significant impairment of Atg proteins diurnal rhythmicity was reported (Qi et al., 2020). This suggests that in diabetic retina the molecular circuit regulating basal autophagy, in terms of intensity and duration, might be altered.
Suppression of LC3-II expression was also reported by Mao and colleagues in STZ-induced diabetic rats; the reduced level of LC3-II correlated with a significant upregulation of a specific microRNA (miRNA), miR-204-5p. Indeed, anti-miR-204-5p treatment enhanced the expression of LC3-II and increased LC3-II/LC3-I ratio, while miR-204-5p mimic treatment was associated with opposite effects thus suggesting that in DR miR-204-5p is responsible for the inhibition of the autophagy pathway (Mao et al., 2019).
The modulation of autophagy in diabetic models may vary depending on the progression of the disease and therefore on the time points analyzed. In RGCs of C57BL/KsJ-db/db mouse, a rodent model of spontaneous diabetes, Fu and co-workers (2020) observed a fluctuating modulation of LC3-II protein levels depending on the age of diabetic mice without identifying a clear trend (Fu et al., 2020). In db/db mice Luo and colleagues reported a downregulation of pro-autophagy proteins like LC3-II, Beclin-1 and Atg5 and a significant upregulation of p62 (Luo et al., 2021). More interestingly, in STZ-induced diabetic rats Shruthi and collaborators (2017) observed a biphasic modulation of LC3-II retinal expression characterized by an increase in 2 months old followed by a significant decrease in 4 months old diabetic rats when compared to control animals. The initial upregulation of the pathway could be part of the adaptive response to the damage induced by hyperglycemia. On the other hand, the later impairment of autophagy might be the consequence of the system overload due to the prolonged diabetic-related damage and contribute to the apoptotic retinal cell death (Shruthi et al., 2017).
A recent study by Park and collaborators (2018) focused on the role of autophagy on RGC survival depending on the type of triggering injury (Park et al., 2018). Autophagy was upregulated in both diabetic and glaucomatous retinas, however while autophagy inhibition, by 3-methyladenine (3-MA), an inhibitor of phosphatidylinositol 3-kinases (PI3K), decreased the apoptosis of RGCs in glaucomatous retina, it failed in rescuing RGCs in diabetic retina. The work by Park and collaborators suggests that, depending on the type of injury and the intracellular pathway engaged for cell death, autophagy could either promote RGC survival or death (Park et al., 2018).
Interestingly in a drosophila model of hyperglycemia developed by raising adult fruit flies under high-sucrose regimens, signs of autophagy deregulation, such as significant and progressive increase of LC3 and p62 staining, with accumulation of autophagosomes were observed in eye sections (Catalani et al., 2021).
In murine retinal explant, exposure to HG was associated with reduced LC3-II levels and upregulation of the cargo-protein p62. Treatment with octreotide, an analogue of somatostatin, prevented the autophagy changes induced by HG, and exerted anti-apoptotic effects. Co-treatment with the autophagy inhibitor chloroquine (CQ) reverted the neuroprotective effects of octreotide suggesting that a cross talk between autophagy and apoptosis occurs in the injured retina (Amato et al., 2018).
The Role of Autophagy in In Vitro Models of Diabetic Retinopathy
Retinal lesions observed over the course of DR are initially characterized by pericyte cell death, which generates ischemia and promotes the extravasation of plasma constituents such as low-density lipoproteins (LDLs). This generates the damage of RPE and activation of microglial and Müller cells (Fu et al., 2012). On the other hand, DR induced neuronal dysfunction, with RGCs death, apoptosis of amacrine cells in INL, loss of synapses and dendrites and alteration of synaptic activity (Ozcan et al., 2006; Oshitari et al., 2011). It is clear that a large number of cellular elements in the retina are affected by DR (Yang et al., 2020) and therefore, several in vitro studies have focused on the modulation of autophagy in the different cell types exposed to diabetes-related insults (Table 2).
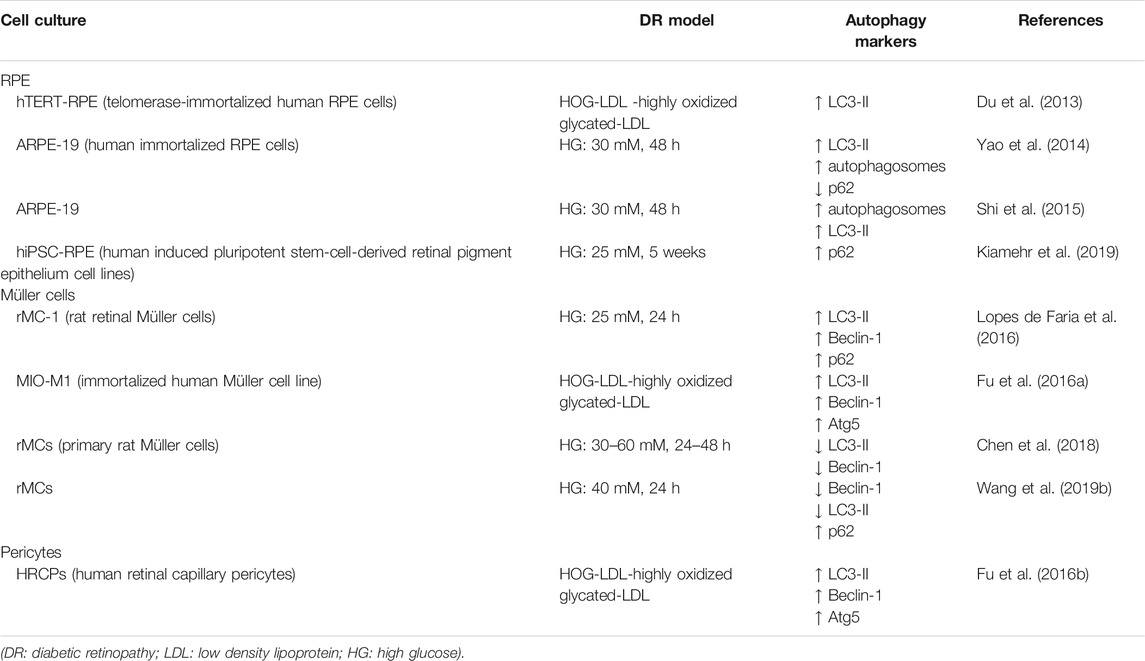
TABLE 2. Autophagy modulation in cell culture models of diabetic retinopathy.
Retinal Pigment Epithelial Cells
Exposure of human immortalized RPE cell, ARPE-19, to high glucose (HG) induced a significant upregulation of autophagy flux. Compared to cells cultured under normal glucose condition, cultures exposed to HG showed increased autophagosome formation, upregulation and changes in the expression pattern of LC3-II and reduction of p62 levels. HG-induced autophagy was mainly regulated through the ROS-mediated ER stress and independent of mTOR signaling pathway (Yao et al., 2014). Similarly, in the same cell line exposed to HG, Shi and colleagues (2015), showed activation of autophagy by reporting an increase of autophagosome number and upregulation of LC3-II protein expression. Under these experimental conditions, inhibition of autophagy obtained by pre-treatment with 3-MA, induced accumulation of damaged-mitochondria, increased the activity of interleukin-1β (IL-1β) and NLRP3 (a NOD-like receptor family pyrin domain containing three inflammasome responsible for the processing of pro-IL1β to the active form of IL-1β) and reduced cell survival (Shi et al., 2015). Altogether, these experimental observations would suggest that in RPE cells exposed to HG stress induction of autophagy represents a cytoprotective response.
Accordingly, treatment with fenofibrate, a peroxisome proliferator-activated receptor alpha (PPARα) agonist by preventing ER-stress and inducing autophagy, exhibited a protective effect in RPE cells exposed to hyperglycemia (25 mM, 18 days) and hypoxia (1% oxygen, for 6 h or 24 h), two components of the diabetic milieu (Miranda et al., 2012; Lazzara et al., 2020).
More recently, Kiamehr and co-workers (2019) using human induced pluripotent stem-cell-derived retinal pigment epithelium (hiPSC-RPE) cell lines, obtained from T2D and healthy control patients, evaluated the effects of hyperglycemia, in the presence of absence of added insulin, on cellular functionality and autophagy (Kiamehr et al., 2019). The authors did not detect any differences in LC3-II expression between diabetic or healthy control hiPSC-RPEs, whereas they observed a significant p62 accumulation in T2D hiPSC-RPE as compared to healthy control (Kiamehr et al., 2019). This change in p62 expression might be unrelated to the autophagy pathway, since p62 is involved in several other functions; one possible hypothesis is that this upregulation of p62 is linked to the antioxidative NRF-2ARE pathway (nuclear factor erythroid-2 related factor/antioxidant response elements) evoked by the energy depletion in diabetic cells (Jain et al., 2010; Felszeghy et al., 2019).
In addition to hyperglycemia, extravasation of plasma lipoproteins modified by oxidation and glycation are important factors driving DR and leading to cytotoxicity (Yu and Lyons, 2013). In telomerase-immortalized human RPE (hTERT-RPE) cells treated with in vitro-modified highly oxidized glycated- (HOG-) LDL, reduced viability was accompanied by the induction of LC3-II expression with no changes in Beclin-1 protein level (Du et al., 2013). Pre-treatment with either native-High-density lipoprotein (N-HDL) or HOG-HDL inhibited HOG-LDL-induced LC3-II expression and partially mitigated RPE cell death (Du et al., 2013).
Retinal Müller Cells
Retinal Müller cells (rMCs), the primary retinal glial cells, make contacts with every cell type in the retina and are necessary for both neuronal and vascular function and viability (Shen et al., 2014).
The role of autophagy in modulating rMCs response to HG was investigated by Lopes de Faria and collaborators (Lopes de Faria et al., 2016). The study showed that rMCs exposed to HG upregulated the initial steps of autophagy, as shown by increase of Beclin-1 and LC3-II protein expression levels; however, the process of cargo degradation could not be completed due to the overcome of lysosomal dysfunction. The latter caused accumulation of p62 that, in turn, led to VEGF release and rMCs apoptosis. Inhibition of the initial stage of autophagy with 3-MA or the final stage with Bafilomycin A1 (a vacuolar-type H+-ATPase inhibitor) increased the number of apoptotic rMCs under either normal condition or following exposure to diabetic milieu conditions. On the contrary, induction of autophagy by rapamycin, a mTOR inhibitor, upregulated Beclin-1 expression, prevented p62 accumulation by restoring autophagy cargo degradation and protected cells from apoptosis (Lopes de Faria et al., 2016).
Comparable results were reported by Wang and collaborators (2019) in a similar cell culture model of primary rat rMCs. Following exposure of rMCs to HG, the authors detected a downregulation of autophagy with reduction of Beclin-1 and LC3-II expression and accumulation of p62 (L. Wang et al., 2019a). Treatment with epigallocatechin gallate (EGCG), a polyphenol present in green tea, protected cells from apoptosis by activating autophagy and reestablishing cargo degradation (L. Wang et al., 2019a). Accordingly, in rat primary rMCs the number of autophagic/lysosomal vacuoles was reduced after exposure to HG; this observation, together with the reported decrease of LC3-II and Beclin-1 protein expression suggested that autophagy activity in rMCs was inhibited by HG conditions. Under these experimental conditions, treatments with berberine reduced HG-induced rMCs apoptosis at least in part by enhancing autophagy (Chen et al., 2018).
Upregulation of Atg5, Beclin-1 and LC3-II proteins were reported in spontaneously immortalized human Müller cell line (MIO-M1) exposed to in vitro-modified HOG-LDL. Müller cell death was partially prevented by inhibiting autophagy with 3-MA or by knocking down Atg5 and Beclin-1 suggesting that autophagy takes part to the apoptosis induced by HOG-LDL (Fu et al., 2016a).
Pericytes
Pericyte cell death is one of the early features of DR (Hammes et al., 2002). Fu and colleagues (2012, 2016) investigated the modulation of autophagy in human retinal capillary pericytes (HRCP) exposed to HOG-LDL (Fu et al., 2012; Fu et al., 2016b) showing a significant dose-dependent increase of LC3-II, Atg5 and Beclin-1 (Fu et al., 2016b). In this study, autophagy appeared to play a dual role depending of the HOG-LDL concentrations: exposure to low levels of HOG-LDL was associated with a pro-survival autophagy response, on the contrary, when the cells were exposed to higher HOG-LDL concentration autophagy promoted cell death (Fu et al., 2016b).
Mitophagy and Diabetic Retinopathy
Mitophagy is a specialized form of autophagy responsible for the quality and quantity control of mitochondria (Pickles et al., 2018). These organelles are the primary source of cellular energy (ATP production), involved in respiration and metabolic processes (Kowluru, 2005) and a key source of ROS in diabetes (Sivitz and Yorek, 2010; Hammes, 2018). Oxidative stress originating in mitochondria from endothelial cell has been reported to alter several independent pathways, each contributing to the development of microvascular complications in DR (Du et al., 2000; Nishikawa et al., 2000). Furthermore, the increase of oxidative stress during hyperglycemia damages itself mitochondria function and structure (Madsen-Bouterse et al., 2010). Indeed, retina of diabetic patients and diabetic rodents showed accumulation of damaged and dysfunctional mitochondria (Masser et al., 2017; Kowluru and Mishra, 2018).
Recently, Zhou and co-workers (2019) showed activation of mitophagy in the retinas of diabetic (db/db) mice (Zhou et al., 2019). Indeed, a significant increase of mitophagy associated protein, PINK-1 and Parkin, was reported in the retinas of db/db mice as compared with non-diabetic (db/m) mice together with the upregulation of LC3-II/LC3-I ratio and reduction of p62. PINK1 (PTEN induces putative kinase protein 1) is a mitochondrially localized serine/threonine protein kinase (Valente et al., 2004) responsible for activation and translocation of Parkin, an E3 ubiquitin-ligase (Kitada et al., 1998), from the cytoplasm to damaged mitochondria (Matsuda et al., 2010). Parkin then marks damaged mitochondria with ubiquitin chains targeting them to mitophagy (Bingol et al., 2014). Accordingly, to the in vivo data, rMC-1 cells exposed to HG displayed significant increase of PINK1, Parkin and LC3-II/LC3-I expression as compared to cells exposed to normal glucose (Zhou et al., 2019).
Zhang and collaborators (2019) demonstrated that while the exposure of ARPE-19 cell cultures to low glucose (LG) (15 mM) induced autophagy, treatment with HG (50 mM) was associated with ROS mediated inhibition of mitophagy and reduced proliferative abilities (Zhang et al., 2019). Under HG conditions PINK1 and Parkin were downregulated and exogenous overexpression of these proteins, which reestablished mitophagy, reduced apoptosis and promoted cellular proliferation (Zhang et al., 2019). Intriguingly, the study by Zhang et al. (2019) showed that LG recruit LC3 to mitochondrial fraction suggesting that this condition may specifically induce mitophagy in RPE cells (Zhang et al., 2019).
Devi and colleagues reported induction of mitochondrial damage and mitophagy in rMCs exposed to HG (Devi et al., 2017) that were mediated by the upregulation of thioredoxin-interacting protein (TXNIP), a pro-oxidative stress and pro-apoptotic protein strongly induced by diabetes and HG conditions (Singh, 2013).
In a recent study, Taki and co-workers (2020), using 661W cells, a transformed murine cone cell line, observed that HG treatment (25 mM, 48 h) induced changes in mitophagy and autophagy with mitochondria accumulation and upregulation of p62. Treatment with 3-MA caused a greater increase of p62, superoxides and caspase 3/7 activation suggesting that impairment of the autophagy pathway correlates with superoxide formation and induction of apoptosis (Taki et al., 2020).
In spontaneous Ins2Akita diabetic mouse model, Hombrebueno and colleagues (2019), showed a time-dependent modulation of mitophagy (Hombrebueno et al., 2019). Indeed PINK1-dependent mitophagy in both Müller cells and photoreceptors was exacerbated within the first 2 months of diabetes, while a significant impairment of the pathway was reported in the advanced stages of neurovascular dysfunction (8 months of diabetes). Furthermore, during prolonged diabetes, impairment of mitophagy correlated with the development of premature outer retina senescence (Hombrebueno et al., 2019).
Concluding Remarks
Autophagy in DR has become an area of intense research, however, despite the studies currently available, the question of whether autophagy is counteracting or favoring the evolution of DR remains unclear. Furthermore, controversial results have often been reported in terms of the type of autophagy modulation induced by hyperglycemia (induction vs impairment), in both in vitro and in vivo models (Gong et al., 2021). However, most evidence suggests that autophagy may act with a damage/time-dependent double action (Figure 2). Under mild stress or during the initial phase of DR, autophagy acts as an adaptative response with pro-survival and anti-apoptotic effects (Dehdashtian et al., 2018); on the other hand, under severe stress and in the later phase of DR, dysregulated autophagy, as a consequence of the system overload due to the prolonged damage, contributes to the apoptotic retinal cell death exacerbating the damage (Fu et al., 2016b; Shruthi et al., 2016).
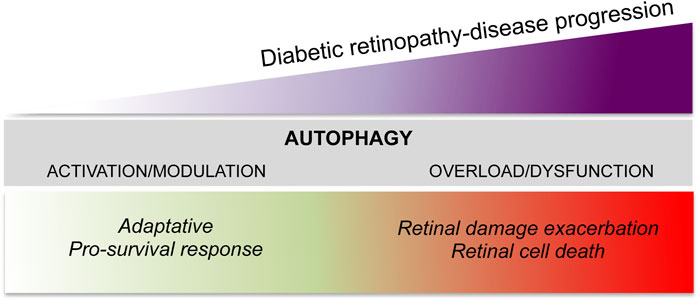
FIGURE 2. The dual role of autophagy in diabetic retinopathy.
In must also be stressed that the claim of some studies related to the induction or inhibition of autophagy are often not supported by the data. Indeed, being a dynamic process, autophagy should be studied in terms of flux. A simply increased number of autophagosomes, either by LC3 immunofluorescent staining or by transmission electron microscope (TEM), as well as changes in LC3-II/LC3-I ratio detected by western blot are not enough to drawing conclusion on the kind of autophagy activity modulation (Abudu and Acevedo-Arozena, 2021; Klionsky et al., 2021). Therefore, the use of more specific experimental settings, i.e. measurement of autophagosome substrates degradation, comparison of LC3-II accumulation in the absence or presence of lysosomal enzymatic activity inhibitors, should be performed to be able to state the occurrence of an autophagic process (Rubinsztein et al., 2009; Klionsky et al., 2021). It should be also taken into consideration that autophagy activity varies with animal age, sex or strain background and it also undergoes a diurnal rhythmicity. All these factors might affect the final results introducing a complex variability among the different experimental settings and making difficult a direct comparison of the different studies.
Based on the data accumulated so far, interpreting the contribution of autophagy in DR is still difficult and further studies are guaranteed in order to unravel the possibility that pharmacological modulation of the pathway could be exploited for DR supportive therapies.
Author Contributions
Data collection and writing: AA, CG, ML, and RR; manuscript revising SR, MT, and RR; editing and figure preparation RR.
Funding
This work is supported by grants from the Italian Ministry of Education, University and Research: PRIN 2017 protocol number “2017TSHBXZ_002.”
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
Abudu, Y. P., and Acevedo-Arozena, A. (2021). Guidelines for the Use and Interpretation of Assays for Monitoring Autophagy (4th Edition)(1). Autophagy 17 (1), 1–382. doi:10.1080/15548627.2020.1797280
Adamis, A. P., Miller, J. W., Bernal, M. T., D'Amico, D. J., Folkman, J., Yeo, T. K., et al. (1994). Increased Vascular Endothelial Growth Factor Levels in the Vitreous of Eyes with Proliferative Diabetic Retinopathy. Am. J. Ophthalmol. 118 (4), 445–450. doi:10.1016/s0002-9394(14)75794-0
Al-Kharashi, A. S. (2018). Role of Oxidative Stress, Inflammation, Hypoxia and Angiogenesis in the Development of Diabetic Retinopathy. Saudi J. Ophthalmol. 32 (4), 318–323. doi:10.1016/j.sjopt.2018.05.002
Altmann, C., and Schmidt, M. H. H. (2018). The Role of Microglia in Diabetic Retinopathy: Inflammation, Microvasculature Defects and Neurodegeneration. Int. J. Mol. Sci. 19 (1). doi:10.3390/ijms19010110
Amato, R., Catalani, E., Dal Monte, M., Cammalleri, M., Di Renzo, I., Perrotta, C., et al. (2018). Autophagy-mediated Neuroprotection Induced by Octreotide in an Ex Vivo Model of Early Diabetic Retinopathy. Pharmacol. Res. 128, 167–178. doi:10.1016/j.phrs.2017.09.022
Antonetti, D. A., Barber, A. J., Hollinger, L. A., Wolpert, E. B., and Gardner, T. W. (1999). Vascular Endothelial Growth Factor Induces Rapid Phosphorylation of Tight junction Proteins Occludin and Zonula Occluden 1. A Potential Mechanism for Vascular Permeability in Diabetic Retinopathy and Tumors. J. Biol. Chem. 274 (33), 23463–23467. doi:10.1074/jbc.274.33.23463
Arroba, A. I., and Valverde Á, M. (2017). Modulation of Microglia in the Retina: New Insights into Diabetic Retinopathy. Acta Diabetol. 54 (6), 527–533. doi:10.1007/s00592-017-0984-z
Backer, J. M. (2008). The Regulation and Function of Class III PI3Ks: Novel Roles for Vps34. Biochem. J. 410 (1), 1–17. doi:10.1042/bj20071427
Barber, A. J. (2003). A New View of Diabetic Retinopathy: a Neurodegenerative Disease of the Eye. Prog. Neuropsychopharmacol. Biol. Psychiatry 27 (2), 283–290. doi:10.1016/s0278-5846(03)00023-x
Barber, A. J., Gardner, T. W., and Abcouwer, S. F. (2011). The Significance of Vascular and Neural Apoptosis to the Pathology of Diabetic Retinopathy. Invest. Ophthalmol. Vis. Sci. 52 (2), 1156–1163. doi:10.1167/iovs.10-6293
Barber, A. J., Lieth, E., Khin, S. A., Antonetti, D. A., Buchanan, A. G., and Gardner, T. W. (1998). Neural Apoptosis in the Retina during Experimental and Human Diabetes. Early Onset and Effect of Insulin. J. Clin. Invest. 102 (4), 783–791. doi:10.1172/jci2425
Baumgartner, W. A. (2000). Etiology, Pathogenesis, and Experimental Treatment of Retinitis Pigmentosa. Med. Hypotheses 54 (5), 814–824. doi:10.1054/mehy.1999.0957
Bek, T. (2017). Diameter Changes of Retinal Vessels in Diabetic Retinopathy. Curr. Diab Rep. 17 (10), 82. doi:10.1007/s11892-017-0909-9
Beltramo, E., and Porta, M. (2013). Pericyte Loss in Diabetic Retinopathy: Mechanisms and Consequences. Curr. Med. Chem. 20 (26), 3218–3225. doi:10.2174/09298673113209990022
Besirli, C. G., Chinskey, N. D., Zheng, Q. D., and Zacks, D. N. (2011). Autophagy Activation in the Injured Photoreceptor Inhibits Fas-Mediated Apoptosis. Invest. Ophthalmol. Vis. Sci. 52 (7), 4193–4199. doi:10.1167/iovs.10-7090
Bingol, B., Tea, J. S., Phu, L., Reichelt, M., Bakalarski, C. E., Song, Q., et al. (2014). The Mitochondrial Deubiquitinase USP30 Opposes Parkin-Mediated Mitophagy. Nature 510 (7505), 370–375. doi:10.1038/nature13418
Bringmann, A., and Wiedemann, P. (2012). Müller Glial Cells in Retinal Disease. Ophthalmologica 227 (1), 1–19. doi:10.1159/000328979
Brownlee, M. (2005). The Pathobiology of Diabetic Complications: a Unifying Mechanism. Diabetes 54 (6), 1615–1625. doi:10.2337/diabetes.54.6.1615
Cai, X., Li, J., Wang, M., She, M., Tang, Y., Li, J., et al. (2017). GLP-1 Treatment Improves Diabetic Retinopathy by Alleviating Autophagy through GLP-1r-Erk1/2-HDAC6 Signaling Pathway. Int. J. Med. Sci. 14 (12), 1203–1212. doi:10.7150/ijms.20962
Catalani, E., Silvestri, F., Bongiorni, S., Taddei, A. R., Fanelli, G., Rinalducci, S., et al. (2021). Retinal Damage in a New Model of Hyperglycemia Induced by High-Sucrose Diets. Pharmacol. Res. 166, 105488. doi:10.1016/j.phrs.2021.105488
Ceravolo, I., Oliverio, G. W., Alibrandi, A., Bhatti, A., Trombetta, L., Rejdak, R., et al. (2020). The Application of Structural Retinal Biomarkers to Evaluate the Effect of Intravitreal Ranibizumab and Dexamethasone Intravitreal Implant on Treatment of Diabetic Macular Edema. Diagnostics (Basel) 10 (6), 413. doi:10.3390/diagnostics10060413
Chen, P., Cescon, M., and Bonaldo, P. (2014). Autophagy-mediated Regulation of Macrophages and its Applications for Cancer. Autophagy 10 (2), 192–200. doi:10.4161/auto.26927
Chen, Z., Li, Y., Jiang, G., Yang, C., Wang, Y., Wang, X., et al. (2018). Knockdown of LRP6 Activates Drp1 to Inhibit Survival of Cardiomyocytes during Glucose Deprivation. Biomed. Pharmacother. 103, 1408–1414. doi:10.1016/j.biopha.2018.04.134
Cheung, N., Mitchell, P., and Wong, T. Y. (2010). Diabetic Retinopathy. Lancet 376 (9735), 124–136. doi:10.1016/s0140-6736(09)62124-3
Chun, Y., and Kim, J. (2018). Autophagy: An Essential Degradation Program for Cellular Homeostasis and Life. Cells 7 (12), 278. doi:10.3390/cells7120278
Coorey, N. J., Shen, W., Chung, S. H., Zhu, L., and Gillies, M. C. (2012). The Role of Glia in Retinal Vascular Disease. Clin. Exp. Optom. 95 (3), 266–281. doi:10.1111/j.1444-0938.2012.00741.x
Dehdashtian, E., Mehrzadi, S., Yousefi, B., Hosseinzadeh, A., Reiter, R. J., Safa, M., et al. (2018). Diabetic Retinopathy Pathogenesis and the Ameliorating Effects of Melatonin; Involvement of Autophagy, Inflammation and Oxidative Stress. Life Sci. 193, 20–33. doi:10.1016/j.lfs.2017.12.001
Devi, T. S., Somayajulu, M., Kowluru, R. A., and Singh, L. P. (2017). TXNIP Regulates Mitophagy in Retinal Müller Cells under High-Glucose Conditions: Implications for Diabetic Retinopathy. Cell Death Dis 8 (5), e2777. doi:10.1038/cddis.2017.190
Dikic, I., and Elazar, Z. (2018). Mechanism and Medical Implications of Mammalian Autophagy. Nat. Rev. Mol. Cel Biol 19 (6), 349–364. doi:10.1038/s41580-018-0003-4
Dong, X. X., Wang, Y., and Qin, Z. H. (2009). Molecular Mechanisms of Excitotoxicity and Their Relevance to Pathogenesis of Neurodegenerative Diseases. Acta Pharmacol. Sin 30 (4), 379–387. doi:10.1038/aps.2009.24
Donnelly, R., Idris, I., and Forrester, J. V. (2004). Protein Kinase C Inhibition and Diabetic Retinopathy: a Shot in the Dark at Translational Research. Br. J. Ophthalmol. 88 (1), 145–151. doi:10.1136/bjo.88.1.145
Du, M., Wu, M., Fu, D., Yang, S., Chen, J., Wilson, K., et al. (2013). Effects of Modified LDL and HDL on Retinal Pigment Epithelial Cells: a Role in Diabetic Retinopathy? Diabetologia 56 (10), 2318–2328. doi:10.1007/s00125-013-2986-x
Du, X. L., Edelstein, D., Rossetti, L., Fantus, I. G., Goldberg, H., Ziyadeh, F., et al. (2000). Hyperglycemia-induced Mitochondrial Superoxide Overproduction Activates the Hexosamine Pathway and Induces Plasminogen Activator Inhibitor-1 Expression by Increasing Sp1 Glycosylation. Proc. Natl. Acad. Sci. U S A. 97 (22), 12222–12226. doi:10.1073/pnas.97.22.12222
Ejaz, S., Chekarova, I., Ejaz, A., Sohail, A., and Lim, C. W. (2008). Importance of Pericytes and Mechanisms of Pericyte Loss during Diabetes Retinopathy. Diabetes Obes. Metab. 10 (1), 53–63. doi:10.1111/j.1463-1326.2007.00795.x
Felszeghy, S., Viiri, J., Paterno, J. J., Hyttinen, J. M. T., Koskela, A., Chen, M., et al. (2019). Loss of NRF-2 and PGC-1α Genes Leads to Retinal Pigment Epithelium Damage Resembling Dry Age-Related Macular Degeneration. Redox Biol. 20, 1–12. doi:10.1016/j.redox.2018.09.011
Flaxman, S. R., Bourne, R. R. A., Resnikoff, S., Ackland, P., Braithwaite, T., Cicinelli, M. V., et al. (2017). Global Causes of Blindness and Distance Vision Impairment 1990-2020: a Systematic Review and Meta-Analysis. Lancet Glob. Health 5 (12), e1221–e1234. doi:10.1016/s2214-109x(17)30393-5
Frake, R. A., Ricketts, T., Menzies, F. M., and Rubinsztein, D. C. (2015). Autophagy and Neurodegeneration. J. Clin. Invest. 125 (1), 65–74. doi:10.1172/jci73944
Fu, D., Wu, M., Zhang, J., Du, M., Yang, S., Hammad, S. M., et al. (2012). Mechanisms of Modified LDL-Induced Pericyte Loss and Retinal Injury in Diabetic Retinopathy. Diabetologia 55 (11), 3128–3140. doi:10.1007/s00125-012-2692-0
Fu, D., Yu, J. Y., Connell, A. R., Yang, S., Hookham, M. B., McLeese, R., et al. (2016a). Beneficial Effects of Berberine on Oxidized LDL-Induced Cytotoxicity to Human Retinal Müller Cells. Invest. Ophthalmol. Vis. Sci. 57 (7), 3369–3379. doi:10.1167/iovs.16-19291
Fu, D., Yu, J. Y., Yang, S., Wu, M., Hammad, S. M., Connell, A. R., et al. (2016b). Survival or Death: a Dual Role for Autophagy in Stress-Induced Pericyte Loss in Diabetic Retinopathy. Diabetologia 59 (10), 2251–2261. doi:10.1007/s00125-016-4058-5
Fu, Y., Wang, Y., Gao, X., Li, H., and Yuan, Y. (2020). Dynamic Expression of HDAC3 in Db/db Mouse RGCs and its Relationship with Apoptosis and Autophagy. J. Diabetes Res. 2020, 6086780. doi:10.1155/2020/6086780
Gabbay, K. H., Merola, L. O., and Field, R. A. (1966). Sorbitol Pathway: Presence in Nerve and Cord with Substrate Accumulation in Diabetes. Science 151 (3707), 209–210. doi:10.1126/science.151.3707.209
Glick, D., Barth, S., and Macleod, K. F. (2010). Autophagy: Cellular and Molecular Mechanisms. J. Pathol. 221 (1), 3–12. doi:10.1002/path.2697
Gong, Q., Wang, H., Yu, P., Qian, T., and Xu, X. (2021). Protective or Harmful: The Dual Roles of Autophagy in Diabetic Retinopathy. Front. Med. (Lausanne) 8, 644121. doi:10.3389/fmed.2021.644121
Gotzaridis, E. V., Lit, E. S., and D'Amico, D. J. (2001). Progress in Vitreoretinal Surgery for Proliferative Diabetic Retinopathy. Semin. Ophthalmol. 16 (1), 31–40. doi:10.1076/soph.16.1.31.4218
Gu, L., Zhu, Y., Zhu, X., and Li, J. (2019). [Genetic Study of a Pedigree Affected with Oculodentodigital Dysplasia]. Zhonghua Yi Xue Yi Chuan Xue Za Zhi 36 (12), 1191–1194. doi:10.3760/cma.j.issn.1003-9406.2019.12.010
Gupta, N., Mansoor, S., Sharma, A., Sapkal, A., Sheth, J., Falatoonzadeh, P., et al. (2013). Diabetic Retinopathy and VEGF. Open Ophthalmol. J. 7, 4–10. doi:10.2174/1874364101307010004
Gupta, N., and Yücel, Y. H. (2007). Glaucoma as a Neurodegenerative Disease. Curr. Opin. Ophthalmol. 18 (2), 110–114. doi:10.1097/ICU.0b013e3280895aea
Hammes, H. P. (2018). Diabetic Retinopathy: Hyperglycaemia, Oxidative Stress and beyond. Diabetologia 61 (1), 29–38. doi:10.1007/s00125-017-4435-8
Hammes, H. P., Lin, J., Renner, O., Shani, M., Lundqvist, A., Betsholtz, C., et al. (2002). Pericytes and the Pathogenesis of Diabetic Retinopathy. Diabetes 51 (10), 3107–3112. doi:10.2337/diabetes.51.10.3107
Hara, T., Takamura, A., Kishi, C., Iemura, S., Natsume, T., Guan, J. L., et al. (2008). FIP200, a ULK-Interacting Protein, Is Required for Autophagosome Formation in Mammalian Cells. J. Cel Biol 181 (3), 497–510. doi:10.1083/jcb.200712064
Hombrebueno, J. R., Cairns, L., Dutton, L. R., Lyons, T. J., Brazil, D. P., Moynagh, P., et al. (2019). Uncoupled Turnover Disrupts Mitochondrial Quality Control in Diabetic Retinopathy. JCI Insight 4 (23), e129760. doi:10.1172/jci.insight.129760
Howell, S. J., Mekhail, M. N., Azem, R., Ward, N. L., and Kern, T. S. (2013). Degeneration of Retinal Ganglion Cells in Diabetic Dogs and Mice: Relationship to Glycemic Control and Retinal Capillary Degeneration. Mol. Vis. 19, 1413–1421.
Huang, H., He, J., Johnson, D., Wei, Y., Liu, Y., Wang, S., et al. (2015). Deletion of Placental Growth Factor Prevents Diabetic Retinopathy and Is Associated with Akt Activation and HIF1α-VEGF Pathway Inhibition. diabetes 64 (3), 200–212. doi:10.2337/db15-er03
Idris, I., Gray, S., and Donnelly, R. (2001). Protein Kinase C Activation: Isozyme-specific Effects on Metabolism and Cardiovascular Complications in Diabetes. Diabetologia 44 (6), 659–673. doi:10.1007/s001250051675
Itakura, E., and Mizushima, N. (2010). Characterization of Autophagosome Formation Site by a Hierarchical Analysis of Mammalian Atg Proteins. Autophagy 6 (6), 764–776. doi:10.4161/auto.6.6.12709
Jain, A., Lamark, T., Sjøttem, E., Larsen, K. B., Awuh, J. A., Øvervatn, A., et al. (2010). p62/SQSTM1 Is a Target Gene for Transcription Factor NRF2 and Creates a Positive Feedback Loop by Inducing Antioxidant Response Element-Driven Gene Transcription. J. Biol. Chem. 285 (29), 22576–22591. doi:10.1074/jbc.M110.118976
Jellinger, K. A. (2010). Should the Word 'dementia' Be Forgotten? J. Cel Mol Med 14 (10), 2415–2416. doi:10.1111/j.1582-4934.2010.01159.x
Jindal, V. (2015). Neurodegeneration as a Primary Change and Role of Neuroprotection in Diabetic Retinopathy. Mol. Neurobiol. 51 (3), 878–884. doi:10.1007/s12035-014-8732-7
Kadlubowska, J., Malaguarnera, L., Waz, P., and Zorena, K. (2016). Neurodegeneration and Neuroinflammation in Diabetic Retinopathy: Potential Approaches to Delay Neuronal Loss. Curr. Neuropharmacol 14 (8), 831–839. doi:10.2174/1570159x14666160614095559
Karst, S. G., Lammer, J., Radwan, S. H., Kwak, H., Silva, P. S., Burns, S. A., et al. (2018). Characterization of In Vivo Retinal Lesions of Diabetic Retinopathy Using Adaptive Optics Scanning Laser Ophthalmoscopy. Int. J. Endocrinol. 2018, 7492946. doi:10.1155/2018/7492946
Kiamehr, M., Klettner, A., Richert, E., Koskela, A., Koistinen, A., Skottman, H., et al. (2019). Compromised Barrier Function in Human Induced Pluripotent Stem-Cell-Derived Retinal Pigment Epithelial Cells from Type 2 Diabetic Patients. Int. J. Mol. Sci. 20 (15). 3773. doi:10.3390/ijms20153773
Kitada, T., Asakawa, S., Hattori, N., Matsumine, H., Yamamura, Y., Minoshima, S., et al. (1998). Mutations in the Parkin Gene Cause Autosomal Recessive Juvenile Parkinsonism. Nature 392 (6676), 605–608. doi:10.1038/33416
Klionsky, D. J., Abdel-Aziz, A. K., Abdelfatah, S., Abdellatif, M., Abdoli, A., Abel, S., et al. (2021). Guidelines for the Use and Interpretation of Assays for Monitoring Autophagy (4th Edition)(1). Autophagy 17(1), 1–382. doi:10.1080/15548627.2020.1797280
Kolm-Litty, V., Sauer, U., Nerlich, A., Lehmann, R., and Schleicher, E. D. (1998). High Glucose-Induced Transforming Growth Factor Beta1 Production Is Mediated by the Hexosamine Pathway in Porcine Glomerular Mesangial Cells. J. Clin. Invest. 101 (1), 160–169. doi:10.1172/jci119875
Kowluru, R. A., and Chan, P. S. (2007). Oxidative Stress and Diabetic Retinopathy. Exp. Diabetes Res. 2007, 43603. doi:10.1155/2007/43603
Kowluru, R. A. (2005). Diabetic Retinopathy: Mitochondrial Dysfunction and Retinal Capillary Cell Death. Antioxid. Redox Signal. 7 (11-12), 1581–1587. doi:10.1089/ars.2005.7.1581
Kowluru, R. A., and Mishra, M. (2018). Therapeutic Targets for Altering Mitochondrial Dysfunction Associated with Diabetic Retinopathy. Expert Opin. Ther. Targets 22 (3), 233–245. doi:10.1080/14728222.2018.1439921
Koya, D., and King, G. L. (1998). Protein Kinase C Activation and the Development of Diabetic Complications. Diabetes 47 (6), 859–866. doi:10.2337/diabetes.47.6.859
Kunchithapautham, K., Coughlin, B., Lemasters, J. J., and Rohrer, B. (2011). Differential Effects of Rapamycin on Rods and Cones during Light-Induced Stress in Albino Mice. Invest. Ophthalmol. Vis. Sci. 52 (6), 2967–2975. doi:10.1167/iovs.10-6278
Lazzara, F., Trotta, M. C., Platania, C. B. M., D'Amico, M., Petrillo, F., Galdiero, M., et al. (2020). Stabilization of HIF-1α in Human Retinal Endothelial Cells Modulates Expression of miRNAs and Proangiogenic Growth Factors. Front. Pharmacol. 11, 1063. doi:10.3389/fphar.2020.01063
Lee, A. Y., and Chung, S. S. (1999). Contributions of Polyol Pathway to Oxidative Stress in Diabetic Cataract. Faseb j 13 (1), 23–30. doi:10.1096/fasebj.13.1.23
Li, J., Wang, J. J., Yu, Q., Chen, K., Mahadev, K., and Zhang, S. X. (2010). Inhibition of Reactive Oxygen Species by Lovastatin Downregulates Vascular Endothelial Growth Factor Expression and Ameliorates Blood-Retinal Barrier Breakdown in Db/db Mice: Role of NADPH Oxidase 4. Diabetes 59 (6), 1528–1538. doi:10.2337/db09-1057
Li, W. W., Li, J., and Bao, J. K. (2012). Microautophagy: Lesser-Known Self-Eating. Cell Mol Life Sci 69 (7), 1125–1136. doi:10.1007/s00018-011-0865-5
Lieth, E., Barber, A. J., Xu, B., Dice, C., Ratz, M. J., Tanase, D., et al. (1998). Glial Reactivity and Impaired Glutamate Metabolism in Short-Term Experimental Diabetic Retinopathy. Penn State Retina Research Group. Diabetes 47 (5), 815–820. doi:10.2337/diabetes.47.5.815
Lopes de Faria, J. M., Duarte, D. A., Montemurro, C., Papadimitriou, A., Consonni, S. R., and Lopes de Faria, J. B. (2016). Defective Autophagy in Diabetic Retinopathy. Invest. Ophthalmol. Vis. Sci. 57 (10), 4356–4366. doi:10.1167/iovs.16-19197
Lung, J. C., Swann, P. G., Wong, D. S., and Chan, H. H. (2012). Global Flash Multifocal Electroretinogram: Early Detection of Local Functional Changes and its Correlations with Optical Coherence Tomography and Visual Field Tests in Diabetic Eyes. Doc Ophthalmol. 125 (2), 123–135. doi:10.1007/s10633-012-9343-0
Luo, Y., Dong, X., Lu, S., Gao, Y., Sun, G., and Sun, X. (2021). Gypenoside XVII Alleviates Early Diabetic Retinopathy by Regulating Müller Cell Apoptosis and Autophagy in Db/db Mice. Eur. J. Pharmacol. 895, 173893. doi:10.1016/j.ejphar.2021.173893
Lynch, S. K., and Abràmoff, M. D. (2017). Diabetic Retinopathy Is a Neurodegenerative Disorder. Vis. Res 139, 101–107. doi:10.1016/j.visres.2017.03.003
Madrakhimov, S. B., Yang, J. Y., Kim, J. H., Han, J. W., and Park, T. K. (2021). mTOR-dependent Dysregulation of Autophagy Contributes to the Retinal Ganglion Cell Loss in Streptozotocin-Induced Diabetic Retinopathy. Cell Commun Signal 19 (1), 29. doi:10.1186/s12964-020-00698-4
Madsen-Bouterse, S. A., Mohammad, G., Kanwar, M., and Kowluru, R. A. (2010). Role of Mitochondrial DNA Damage in the Development of Diabetic Retinopathy, and the Metabolic Memory Phenomenon Associated with its Progression. Antioxid. Redox Signal. 13 (6), 797–805. doi:10.1089/ars.2009.2932
Mao, X. B., Cheng, Y. H., and Xu, Y. Y. (2019). miR-204-5p Promotes Diabetic Retinopathy Development via Downregulation of Microtubule-Associated Protein 1 Light Chain 3. Exp. Ther. Med. 17 (4), 2945–2952. doi:10.3892/etm.2019.7327
Mariño, G., Madeo, F., and Kroemer, G. (2011). Autophagy for Tissue Homeostasis and Neuroprotection. Curr. Opin. Cel Biol 23 (2), 198–206. doi:10.1016/j.ceb.2010.10.001
Masser, D. R., Otalora, L., Clark, N. W., Kinter, M. T., Elliott, M. H., and Freeman, W. M. (2017). Functional Changes in the Neural Retina Occur in the Absence of Mitochondrial Dysfunction in a Rodent Model of Diabetic Retinopathy. J. Neurochem. 143 (5), 595–608. doi:10.1111/jnc.14216
Matsuda, N., Sato, S., Shiba, K., Okatsu, K., Saisho, K., Gautier, C. A., et al. (2010). PINK1 Stabilized by Mitochondrial Depolarization Recruits Parkin to Damaged Mitochondria and Activates Latent Parkin for Mitophagy. J. Cel Biol 189 (2), 211–221. doi:10.1083/jcb.200910140
Menzies, F. M., Fleming, A., Caricasole, A., Bento, C. F., Andrews, S. P., Ashkenazi, A., et al. (2017). Autophagy and Neurodegeneration: Pathogenic Mechanisms and Therapeutic Opportunities. Neuron 93 (5), 1015–1034. doi:10.1016/j.neuron.2017.01.022
Miranda, S., González-Rodríguez, Á., García-Ramírez, M., Revuelta-Cervantes, J., Hernández, C., Simó, R., et al. (2012). Beneficial Effects of Fenofibrate in Retinal Pigment Epithelium by the Modulation of Stress and Survival Signaling under Diabetic Conditions. J. Cel Physiol 227 (6), 2352–2362. doi:10.1002/jcp.22970
Mizushima, N. (2007). Autophagy: Process and Function. Genes Dev. 21 (22), 2861–2873. doi:10.1101/gad.1599207
Mizushima, N., and Murphy, L. O. (2020). Autophagy Assays for Biological Discovery and Therapeutic Development. Trends Biochem. Sci. 45 (12), 1080–1093. doi:10.1016/j.tibs.2020.07.006
Mizushima, N., Yoshimori, T., and Ohsumi, Y. (2011). The Role of Atg Proteins in Autophagosome Formation. Annu. Rev. Cel Dev Biol 27, 107–132. doi:10.1146/annurev-cellbio-092910-154005
Mrugacz, M., Bryl, A., and Zorena, K. (2021). Retinal Vascular Endothelial Cell Dysfunction and Neuroretinal Degeneration in Diabetic Patients. J. Clin. Med. 10 (3), 458. doi:10.3390/jcm10030458
Nishikawa, T., Edelstein, D., Du, X. L., Yamagishi, S., Matsumura, T., Kaneda, Y., et al. (2000). Normalizing Mitochondrial Superoxide Production Blocks Three Pathways of Hyperglycaemic Damage. Nature 404 (6779), 787–790. doi:10.1038/35008121
Noda, T. (2017). Regulation of Autophagy through TORC1 and mTORC1. Biomolecules 7 (3), 52. doi:10.3390/biom7030052
Ogurtsova, K., da Rocha Fernandes, J. D., Huang, Y., Linnenkamp, U., Guariguata, L., Cho, N. H., et al. (2017). IDF Diabetes Atlas: Global Estimates for the Prevalence of Diabetes for 2015 and 2040. Diabetes Res. Clin. Pract. 128, 40–50. doi:10.1016/j.diabres.2017.03.024
Ola, M. S., and Alhomida, A. S. (2014). Neurodegeneration in Diabetic Retina and its Potential Drug Targets. Curr. Neuropharmacol 12 (4), 380–386. doi:10.2174/1570159x12666140619205024
Oshitari, T., Yoshida-Hata, N., and Yamamoto, S. (2011). Effect of Neurotrophin-4 on Endoplasmic Reticulum Stress-Related Neuronal Apoptosis in Diabetic and High Glucose Exposed Rat Retinas. Neurosci. Lett. 501 (2), 102–106. doi:10.1016/j.neulet.2011.06.057
Otomo, C., Metlagel, Z., Takaesu, G., and Otomo, T. (2013). Structure of the Human ATG12∼ATG5 Conjugate Required for LC3 Lipidation in Autophagy. Nat. Struct. Mol. Biol. 20 (1), 59–66. doi:10.1038/nsmb.2431
Ozcan, U., Yilmaz, E., Ozcan, L., Furuhashi, M., Vaillancourt, E., Smith, R. O., et al. (2006). Chemical Chaperones Reduce ER Stress and Restore Glucose Homeostasis in a Mouse Model of Type 2 Diabetes. Science 313 (5790), 1137–1140. doi:10.1126/science.1128294
Park, H. L., Kim, J. H., and Park, C. K. (2018). Different Contributions of Autophagy to Retinal Ganglion Cell Death in the Diabetic and Glaucomatous Retinas. Sci. Rep. 8 (1), 13321. doi:10.1038/s41598-018-30165-7
Parzych, K. R., and Klionsky, D. J. (2014). An Overview of Autophagy: Morphology, Mechanism, and Regulation. Antioxid. Redox Signal. 20 (3), 460–473. doi:10.1089/ars.2013.5371
Piano, I., Novelli, E., Della Santina, L., Strettoi, E., Cervetto, L., and Gargini, C. (2016). Involvement of Autophagic Pathway in the Progression of Retinal Degeneration in a Mouse Model of Diabetes. Front Cel Neurosci 10, 42. doi:10.3389/fncel.2016.00042
Pickles, S., Vigié, P., and Youle, R. J. (2018). Mitophagy and Quality Control Mechanisms in Mitochondrial Maintenance. Curr. Biol. 28 (4), R170–r185. doi:10.1016/j.cub.2018.01.004
Punzo, C., Kornacker, K., and Cepko, C. L. (2009). Stimulation of the insulin/mTOR Pathway Delays Cone Death in a Mouse Model of Retinitis Pigmentosa. Nat. Neurosci. 12 (1), 44–52. doi:10.1038/nn.2234
Qi, X., Mitter, S. K., Yan, Y., Busik, J. V., Grant, M. B., and Boulton, M. E. (2020). Diurnal Rhythmicity of Autophagy Is Impaired in the Diabetic Retina. Cells 9 (4), 905. doi:10.3390/cells9040905
Rodríguez-Muela, N., Hernández-Pinto, A. M., Serrano-Puebla, A., García-Ledo, L., Latorre, S. H., de la Rosa, E. J., et al. (2015). Lysosomal Membrane Permeabilization and Autophagy Blockade Contribute to Photoreceptor Cell Death in a Mouse Model of Retinitis Pigmentosa. Cell Death Differ 22 (3), 476–487. doi:10.1038/cdd.2014.203
Romeo, G., Liu, W. H., Asnaghi, V., Kern, T. S., and Lorenzi, M. (2002). Activation of Nuclear Factor-kappaB Induced by Diabetes and High Glucose Regulates a Proapoptotic Program in Retinal Pericytes. Diabetes 51 (7), 2241–2248. doi:10.2337/diabetes.51.7.2241
Romero-Aroca, P., Baget-Bernaldiz, M., Pareja-Rios, A., Lopez-Galvez, M., Navarro-Gil, R., and Verges, R. (2016). Diabetic Macular Edema Pathophysiology: Vasogenic versus Inflammatory. J. Diabetes Res. 2016, 2156273. doi:10.1155/2016/2156273
Rosa, M. D., Distefano, G., Gagliano, C., Rusciano, D., and Malaguarnera, L. (2016). Autophagy in Diabetic Retinopathy. Curr. Neuropharmacol 14 (8), 810–825. doi:10.2174/1570159x14666160321122900
Rossi, S., Maisto, R., Gesualdo, C., Trotta, M. C., Ferraraccio, F., Kaneva, M. K., et al. (2016). Activation of Melanocortin Receptors MC 1 and MC 5 Attenuates Retinal Damage in Experimental Diabetic Retinopathy. Mediators Inflamm., 7368389. doi:10.1155/2016/73683892016
Rousseau, S., Houle, F., Landry, J., and Huot, J. (1997). p38 MAP Kinase Activation by Vascular Endothelial Growth Factor Mediates Actin Reorganization and Cell Migration in Human Endothelial Cells. Oncogene 15 (18), 2169–2177. doi:10.1038/sj.onc.1201380
Rubinsztein, D. C., Cuervo, A. M., Ravikumar, B., Sarkar, S., Korolchuk, V., Kaushik, S., et al. (2009). In Search of an "autophagomometer. Autophagy 5 (5), 585–589. doi:10.4161/auto.5.5.8823
Russo, R., Berliocchi, L., Adornetto, A., Amantea, D., Nucci, C., Tassorelli, C., et al. (2013). In Search of New Targets for Retinal Neuroprotection: Is There a Role for Autophagy? Curr. Opin. Pharmacol. 13 (1), 72–77. doi:10.1016/j.coph.2012.09.004
Saxena, R., Singh, D., Saklani, R., and Gupta, S. K. (2016). Clinical Biomarkers and Molecular Basis for Optimized Treatment of Diabetic Retinopathy: Current Status and Future Prospects. Eye Brain 8, 1–13. doi:10.2147/EB.S69185
Shen, W., Lee, S. R., Araujo, J., Chung, S. H., Zhu, L., and Gillies, M. C. (2014). Effect of Glucocorticoids on Neuronal and Vascular Pathology in a Transgenic Model of Selective Müller Cell Ablation. Glia 62 (7), 1110–1124. doi:10.1002/glia.22666
Shi, H., Zhang, Z., Wang, X., Li, R., Hou, W., Bi, W., et al. (2015). Inhibition of Autophagy Induces IL-1β Release from ARPE-19 Cells via ROS Mediated NLRP3 Inflammasome Activation under High Glucose Stress. Biochem. Biophys. Res. Commun. 463 (4), 1071–1076. doi:10.1016/j.bbrc.2015.06.060
Shinohara, M., Thornalley, P. J., Giardino, I., Beisswenger, P., Thorpe, S. R., Onorato, J., et al. (1998). Overexpression of Glyoxalase-I in Bovine Endothelial Cells Inhibits Intracellular Advanced Glycation Endproduct Formation and Prevents Hyperglycemia-Induced Increases in Macromolecular Endocytosis. J. Clin. Invest. 101 (5), 1142–1147. doi:10.1172/jci119885
Shruthi, K., Reddy, S. S., and Reddy, G. B. (2017). Ubiquitin-proteasome System and ER Stress in the Retina of Diabetic Rats. Arch. Biochem. Biophys. 627, 10–20. doi:10.1016/j.abb.2017.06.006
Shruthi, K., Reddy, S. S., Reddy, P. Y., Shivalingam, P., Harishankar, N., and Reddy, G. B. (2016). Amelioration of Neuronal Cell Death in a Spontaneous Obese Rat Model by Dietary Restriction through Modulation of Ubiquitin Proteasome System. J. Nutr. Biochem. 33, 73–81. doi:10.1016/j.jnutbio.2016.03.008
Simão, S., Costa, M., Sun, J. K., Cunha-Vaz, J., and Simó, R. (2017). Development of a Normative Database for Multifocal Electroretinography in the Context of a Multicenter Clinical Trial. Ophthalmic Res. 57 (2), 107–117. doi:10.1159/000450958
Simo-Servat, O., Hernandez, C., and Simo, R. (2019). Diabetic Retinopathy in the Context of Patients with Diabetes. Ophthalmic Res. 62 (4), 211–217. doi:10.1159/000499541
Singh, L. P. (2013). Thioredoxin Interacting Protein (TXNIP) and Pathogenesis of Diabetic Retinopathy. J. Clin. Exp. Ophthalmol. 4. doi:10.4172/2155-9570.1000287
Singh, R., Ramasamy, K., Abraham, C., Gupta, V., and Gupta, A. (2008). Diabetic Retinopathy: an Update. Indian J. Ophthalmol. 56 (3), 178–188. doi:10.4103/0301-4738.40355
Sivitz, W. I., and Yorek, M. A. (2010). Mitochondrial Dysfunction in Diabetes: from Molecular Mechanisms to Functional Significance and Therapeutic Opportunities. Antioxid. Redox Signal. 12 (4), 537–577. doi:10.1089/ars.2009.2531
Sohn, E. H., van Dijk, H. W., Jiao, C., Kok, P. H., Jeong, W., Demirkaya, N., et al. (2016). Retinal Neurodegeneration May Precede Microvascular Changes Characteristic of Diabetic Retinopathy in Diabetes Mellitus. Proc. Natl. Acad. Sci. U S A. 113 (19), E2655–E2664. doi:10.1073/pnas.1522014113
Solomon, S. D., Chew, E., Duh, E. J., Sobrin, L., Sun, J. K., VanderBeek, B. L., et al. (2017). Diabetic Retinopathy: A Position Statement by the American Diabetes Association. Diabetes Care 40 (3), 412–418. doi:10.2337/dc16-2641
Stitt, A. W. (2010). AGEs and Diabetic Retinopathy. Invest. Ophthalmol. Vis. Sci. 51 (10), 4867–4874. doi:10.1167/iovs.10-5881
Stitt, A. W., Curtis, T. M., Chen, M., Medina, R. J., McKay, G. J., Jenkins, A., et al. (2016). The Progress in Understanding and Treatment of Diabetic Retinopathy. Prog. Retin. Eye Res. 51, 156–186. doi:10.1016/j.preteyeres.2015.08.001
Sun, J. K., and Jampol, L. M. (2019). The Diabetic Retinopathy Clinical Research Network (DRCR.Net) and its Contributions to the Treatment of Diabetic Retinopathy. Ophthalmic Res. 62 (4), 225–230. doi:10.1159/000502779
Taki, K., Horie, T., Kida, T., Mimura, M., Ikeda, T., and Oku, H. (2020). Impairment of Autophagy Causes Superoxide Formation and Caspase Activation in 661 W Cells, a Cell Line for Cone Photoreceptors, under Hyperglycemic Conditions. Int. J. Mol. Sci. 21 (12). 4240. doi:10.3390/ijms21124240
Ting, D. S., Cheung, G. C., and Wong, T. Y. (2016). Diabetic Retinopathy: Global Prevalence, Major Risk Factors, Screening Practices and Public Health Challenges: a Review. Clin. Exp. Ophthalmol. 44 (4), 260–277. doi:10.1111/ceo.12696
Trento, M., Durando, O., Lavecchia, S., Charrier, L., Cavallo, F., Costa, M. A., et al. (2017). Vision Related Quality of Life in Patients with Type 2 Diabetes in the EUROCONDOR Trial. Endocrine 57 (1), 83–88. doi:10.1007/s12020-016-1097-0
Valente, E. M., Abou-Sleiman, P. M., Caputo, V., Muqit, M. M., Harvey, K., Gispert, S., et al. (2004). Hereditary Early-Onset Parkinson's Disease Caused by Mutations in PINK1. Science 304 (5674), 1158–1160. doi:10.1126/science.1096284
van Dijk, H. W., Verbraak, F. D., Kok, P. H., Garvin, M. K., Sonka, M., Lee, K., et al. (2010). Decreased Retinal Ganglion Cell Layer Thickness in Patients with Type 1 Diabetes. Invest. Ophthalmol. Vis. Sci. 51 (7), 3660–3665. doi:10.1167/iovs.09-5041
Wang, L., Sun, X., Zhu, M., Du, J., Xu, J., Qin, X., et al. (2019a). Epigallocatechin-3-gallate Stimulates Autophagy and Reduces Apoptosis Levels in Retinal Müller Cells under High-Glucose Conditions. Exp. Cel Res 380 (2), 149–158. doi:10.1016/j.yexcr.2019.04.014
Wang, S., Ji, L. Y., Li, L., and Li, J. M. (2019b). Oxidative Stress, Autophagy and Pyroptosis in the Neovascularization of Oxygen-Induced Retinopathy in Mice. Mol. Med. Rep. 19 (2), 927–934. doi:10.3892/mmr.2018.9759
Wang, W., and Lo, A. C. Y. (2018). Diabetic Retinopathy: Pathophysiology and Treatments. Int. J. Mol. Sci. 19 (6), 1816. doi:10.3390/ijms19061816
Wang, W., Wang, Q., Wan, D., Sun, Y., Wang, L., Chen, H., et al. (2017). Histone HIST1H1C/H1.2 Regulates Autophagy in the Development of Diabetic Retinopathy. Autophagy 13 (5), 941–954. doi:10.1080/15548627.2017.1293768
Wang, Y., Liu, X., Zhu, L., Li, W., Li, Z., Lu, X., et al. (2020). PG545 Alleviates Diabetic Retinopathy by Promoting Retinal Müller Cell Autophagy to Inhibit the Inflammatory Response. Biochem. Biophys. Res. Commun. 531 (4), 452–458. doi:10.1016/j.bbrc.2020.07.134
Wilkinson, C. P., Ferris, F. L., Klein, R. E., Lee, P. P., Agardh, C. D., Davis, M., et al. (2003). Proposed International Clinical Diabetic Retinopathy and Diabetic Macular Edema Disease Severity Scales. Ophthalmology 110 (9), 1677–1682. doi:10.1016/s0161-6420(03)00475-5
Wolff, B. E., Bearse, M. A., Schneck, M. E., Dhamdhere, K., Harrison, W. W., Barez, S., et al. (2015). Color Vision and Neuroretinal Function in Diabetes. Doc Ophthalmol. 130 (2), 131–139. doi:10.1007/s10633-014-9476-4
Xie, Z., and Klionsky, D. J. (2007). Autophagosome Formation: Core Machinery and Adaptations. Nat. Cel Biol 9 (10), 1102–1109. doi:10.1038/ncb1007-1102
Yang, S., Zhang, J., and Chen, L. (2020). The Cells Involved in the Pathological Process of Diabetic Retinopathy. Biomed. Pharmacother. 132, 110818. doi:10.1016/j.biopha.2020.110818
Yang, Z., and Klionsky, D. J. (2009). An Overview of the Molecular Mechanism of Autophagy. Curr. Top. Microbiol. Immunol. 335, 1–32. doi:10.1007/978-3-642-00302-8_1
Yang, Z., and Klionsky, D. J. (2010). Mammalian Autophagy: Core Molecular Machinery and Signaling Regulation. Curr. Opin. Cel Biol 22 (2), 124–131. doi:10.1016/j.ceb.2009.11.014
Yao, J., Tao, Z. F., Li, C. P., Li, X. M., Cao, G. F., Jiang, Q., et al. (2014). Regulation of Autophagy by High Glucose in Human Retinal Pigment Epithelium. Cell Physiol Biochem 33 (1), 107–116. doi:10.1159/000356654
Yau, J. W., Rogers, S. L., Kawasaki, R., Lamoureux, E. L., Kowalski, J. W., Bek, T., et al. (2012). Global Prevalence and Major Risk Factors of Diabetic Retinopathy. Diabetes Care 35 (3), 556–564. doi:10.2337/dc11-1909
Yoshii, S. R., and Mizushima, N. (2017). Monitoring and Measuring Autophagy. Int. J. Mol. Sci. 18 (9). 1865. doi:10.3390/ijms18091865
Yu, J. Y., and Lyons, T. J. (2013). Modified Lipoproteins in Diabetic Retinopathy: A Local Action in the Retina. J. Clin. Exp. Ophthalmol. 4 (6). 314. doi:10.4172/2155-9570.1000314
Zhang, H., Chang, J. T., Guo, B., Hansen, M., Jia, K., Kovács, A. L., et al. (2015). Guidelines for Monitoring Autophagy in Caenorhabditis elegans. Autophagy 11 (1), 9–27. doi:10.1080/15548627.2014.1003478
Zhang, Y., Xi, X., Mei, Y., Zhao, X., Zhou, L., Ma, M., et al. (2019). High-glucose Induces Retinal Pigment Epithelium Mitochondrial Pathways of Apoptosis and Inhibits Mitophagy by Regulating ROS/PINK1/Parkin Signal Pathway. Biomed. Pharmacother. 111, 1315–1325. doi:10.1016/j.biopha.2019.01.034
Zhou, K. L., Zhou, Y. F., Wu, K., Tian, N. F., Wu, Y. S., Wang, Y. L., et al. (2015). Stimulation of Autophagy Promotes Functional Recovery in Diabetic Rats with Spinal Cord Injury. Sci. Rep. 5, 17130. doi:10.1038/srep17130
Zhou, P., Xie, W., Meng, X., Zhai, Y., Dong, X., Zhang, X., et al. (2019). Notoginsenoside R1 Ameliorates Diabetic Retinopathy through PINK1-dependent Activation of Mitophagy. Cells 8 (3), 213. doi:10.3390/cells8030213
Keywords: autophagy, diabetic retinopathy, LC3, autophagosomes, retinal degeneration, hyperglycemia
Citation: Adornetto A, Gesualdo C, Laganà ML, Trotta MC, Rossi S and Russo R (2021) Autophagy: A Novel Pharmacological Target in Diabetic Retinopathy. Front. Pharmacol. 12:695267. doi: 10.3389/fphar.2021.695267
Received: 14 April 2021; Accepted: 09 June 2021;
Published: 21 June 2021.
Edited by:
Salvatore Salomone, University of Catania, ItalyReviewed by:
Elisabetta Catalani, University of Tuscia, ItalyGiovanni Casini, University of Pisa, Italy
Copyright © 2021 Adornetto, Gesualdo, Laganà, Trotta, Rossi and Russo. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Annagrazia Adornetto, YW5uYWdyYXppYS5hZG9ybmV0dG9AdW5pY2FsLml0