 Raquel Jurado-Escobar1,2
Raquel Jurado-Escobar1,2 Inmaculada Doña1,3,4
Inmaculada Doña1,3,4 José Triano-Cornejo1
José Triano-Cornejo1 James R. Perkins5,6,7Natalia Pérez-Sánchez3Almudena Testera-Montes3Marina Labella3Joan Bartra4,8
James R. Perkins5,6,7Natalia Pérez-Sánchez3Almudena Testera-Montes3Marina Labella3Joan Bartra4,8 José J. Laguna4,9
José J. Laguna4,9 Miguel Estravís4,10
Miguel Estravís4,10 José A. G. Agúndez4,11
José A. G. Agúndez4,11 María J. Torres1,2,3,4,12
María J. Torres1,2,3,4,12 José A. Cornejo-García1,4*
José A. Cornejo-García1,4*- 1Allergy Research Group, Instituto De Investigación Biomédica De Málaga-IBIMA, Malaga, Spain
- 2Departamento De Medicina, Universidad De Málaga, Malaga, Spain
- 3Allergy Unit, Hospital Regional Universitario De Málaga, Malaga, Spain
- 4ARADyAL Network, Instituto De Salud Carlos III, Madrid, Spain
- 5Department of Molecular Biology and Biochemistry, University of Malaga, Malaga, Spain
- 6CIBER De Enfermedades Raras (CIBERER), ISCIII, Madrid, Spain
- 7The Biomedical Research Institute of Malaga (IBIMA), Malaga, Spain
- 8Allergy Section, Pneumology Department, Hospital Clinic, Universitat De Barcelona, Barcelona, Spain
- 9Allergy Unit, Allergo-Anaesthesia Unit, Hospital Central De La Cruz Roja, Faculty of Medicine, Alfonso X El Sabio University, Madrid, Spain
- 10Instituto De Investigación Biomédica De Salamanca (IBSAL), Salamanca, Spain
- 11Institute of Molecular Pathology Biomarkers, UEx, Cáceres, Spain
- 12Nanostructures for Diagnosing and Treatment of Allergic Diseases Laboratory, Andalusian Center for Nanomedicine and Biotechnology-BIONAND, Malaga, Spain
Nonsteroidal anti-inflammatory drugs (NSAIDs) are among the main triggers of drug hypersensitivity reactions, probably due to their high consumption worldwide. The most frequent type of NSAID hypersensitivity is NSAID cross-hypersensitivity, in which patients react to NSAIDs from different chemical groups in the absence of a specific immunological response. The underlying mechanism of NSAID cross-hypersensitivity has been linked to cyclooxygenase (COX)-1 inhibition causing an imbalance in the arachidonic acid pathway. Despite NSAID-induced acute urticaria/angioedema (NIUA) being the most frequent clinical phenotype, most studies have focused on NSAID-exacerbated respiratory disease. As NSAID cross-hypersensitivity reactions are idiosyncratic, only appearing in some subjects, it is believed that individual susceptibility is under the influence of genetic factors. Although associations with polymorphisms in genes from the AA pathway have been described, no previous study has evaluated the potential role of cytosolic phospholipase A2 (cPLA2) variants. This enzyme catalyzes the initial hydrolysis of membrane phospholipids to release AA, which can be subsequently metabolized into eicosanoids. Here, we analyzed for the first time the overall genetic variation in the cPLA2 gene (PLA2G4A) in NIUA patients. For this purpose, a set of tagging single nucleotide polymorphisms (tagSNPs) in PLA2G4A were selected using data from Europeans subjects in the 1,000 Genomes Project, and genotyped with the iPlex Sequenom MassArray technology. Two independent populations, each comprising NIUA patients and NSAID-tolerant controls, were recruited in Spain, for the purposes of discovery and replication, comprising a total of 1,128 individuals. Fifty-eight tagSNPs were successfully genotyped in the discovery cohort, of which four were significantly associated with NIUA after Bonferroni correction (rs2049963, rs2064471, rs12088010, and rs12746200). These polymorphisms were then genotyped in the replication cohort: rs2049963 was associated with increased risk for NIUA after Bonferroni correction under the dominant and additive models, whereas rs12088010 and rs12746200 were protective under these two inheritance models. Our results suggest a role for PLA2G4A polymorphisms in NIUA. However, further studies are required to replicate our findings, elucidate the mechanistic role, and evaluate the participation of PLA2G4A variants in other phenotypes induced by NSAID cross-hypersensitivity.
Introduction
Nonsteroidal anti-inflammatory drugs (NSAIDs) are among the most highly consumed medicines worldwide because of their efficiency in the treatment of pain and different inflammatory conditions (Fosbøl et al., 2008; Conaghan, 2012; Duong et al., 2014). The wide usage of NSAIDs is reflected in recent studies reporting that the 57.8% of the Danish (Fosbøl et al., 2008) and the 43.6% of the French (Duong et al., 2014) general populations filled at least one prescription for NSAIDs in the periods 1997–2005 and 2009–2010, respectively; for children, an European study reported that 16.9% have taken at least one NSAID (Valkhoff et al., 2013). However, NSAIDs also trigger the 21–25% of all adverse drug reactions, which includes hypersensitivity to drugs (Kowalski et al., 2011). The most common type of NSAID-hypersensitivity is cross-hypersensitivity, with patients suffering different reactions to chemically unrelated NSAIDs (Kowalski et al., 2011; Kowalski et al., 2013). NSAID-induced acute urticaria/angioedema (NIUA), a phenotype characterized by wheals and/or angioedema development in patients without chronic spontaneous urticaria (CSU), is the most frequent clinical entity (Dona et al., 2012; Kowalski et al., 2013; Dona et al., 2014; Doña et al., 2020).
NSAID cross-hypersensitivity reactions are not mediated by an immunological mechanism but associated with the pharmacological inhibition of cyclooxygenase-1 (COX-1) and increased of cysteinyl-leukotriene (CysLT) levels. COX-1 and the inducible isozyme COX-2 catalyze the initial steps in the biosynthesis of prostaglandins (PGs), leading to the formation of PGG2 and PGH2, which are subsequently transformed into PGE2, prostacyclins, or thromboxanes. COX-1 inhibition blocks PG synthesis, leading to an increased transformation of arachidonic acid (AA) into LTA4, which is metabolized to LTC4 and LTE4 (Doña et al., 2020). Thus, COX-1 inhibition by NSAIDs shunts the AA pathway from PG toward CysLT synthesis, eliciting a reaction in susceptible individuals (Doña et al., 2020).
The COX-1 inhibition hypothesis, first proposed for NSAID-exacerbated respiratory disease (NERD) (Szczeklik et al., 1975), has been supported by different studies showing increased LTE4 levels in urine (Christie et al., 1991; Sladek and Szczeklik, 1993), and nasal (Sladek and Szczeklik, 1993) and bronchoalveolar lavages (Sladek et al., 1994) after a positive salicylic acid (ASA) drug provocation test (ASA-DPT). Moreover, increased baseline levels of LTE4 have been reported in NERD patients (Oosaki et al., 1997; Mita et al., 2001; Higashi et al., 2004; Micheletto et al., 2006) and are associated with the severity of the respiratory reaction during ASA-DPT (Daffern et al., 1999). The COX-1 inhibition hypothesis has been further extended to NSAID-exacerbated cutaneous disease (NECD) (Zembowicz et al., 2003; Setkowicz et al., 2009) and recently to NIUA (Doña et al., 2019).
NSAID cross-hypersensitivity susceptibility is thought to be influenced by genetic factors, as has been suggested by familial aggregation; nevertheless, the inheritance pattern does not seem to be Mendelian (Mastalerz et al., 2006; Caimmi et al., 2014). Despite NIUA being the most frequent clinical phenotype induced by drug hypersensitivity, most genetic association studies have focused on NERD (Jurado-Escobar et al., 2017). Most of these studies have focused centered on a limited number of single-nucleotide polymorphisms (SNPs), without evaluating the overall variability of genes of potential interest. In addition, most studies reporting genetic associations did not include a second population for replication (Oussalah et al., 2016; Jurado-Escobar et al., 2017).
Some genetic associations have been found for variants in enzymes and receptors from the AA pathway and cross-hypersensitivity to NSAIDs (Jurado-Escobar et al., 2017). However, no study has been made of the potential role of study of cytosolic phospholipase A2 (cPLA2 or PLA2G4A) overall genetic variability in NSAID cross-hypersensitivity. This gene encodes a member of the cytosolic phospholipase A2 group IV family, responsible for the hydrolysis of membrane phospholipids to release AA, which is subsequently metabolized to produce eicosanoids, including PGs and CysLTs. However, in a previous study from our group evaluating 9 SNPs across 5 genes, a variant on PLA2G4A was found to be associated with NIUA (Ayuso et al., 2015). Moreover, from the 8 variants on chromosome 1, we found to be marginally associated with NIUA in a GWAS, none was related with PLA2G4A (Cornejo-García et al., 2013).
In this study, we have evaluated the overall variability of the PLA2G4A gene and the potential role of its variants in the development of NIUA. For this purpose, we have selected a set of tagging SNPs (tagSNPs), which encompass the overall variability of this gene, and genotyped a group of NIUA patients and NSAID-tolerant individuals. Moreover, we have included a second unrelated population of NIUA patients and controls to further validate our initial findings. As far as we know, this is the first time such an approach has been performed in cross-hypersensitivity to NSAIDs. Our findings help to disentangle the molecular mechanisms underlying this pathology and to identify potential biomarkers for diagnosis.
Methods
Subjects
In this case–control study, two consecutively recruited, independent groups of unrelated cases and controls, considered discovery and replication populations, were included. All patients were aged 18–60 years and self-reported Spanish ancestry. The discovery population was recruited from June 2016 to December 2017 in the Allergy Unit of the Malaga Regional University Hospital (Spain), and consisted on of 269 NIUA patients and 299 NSAID-tolerant controls. The replication population was recruited from January 2018 to December 2019 in the Allergy Units of the Cruz Roja Central Hospital (Madrid, Spain) and Clinic Hospital (Barcelona, Spain), and included 181 NIUA patients and 379 controls (Table 1).
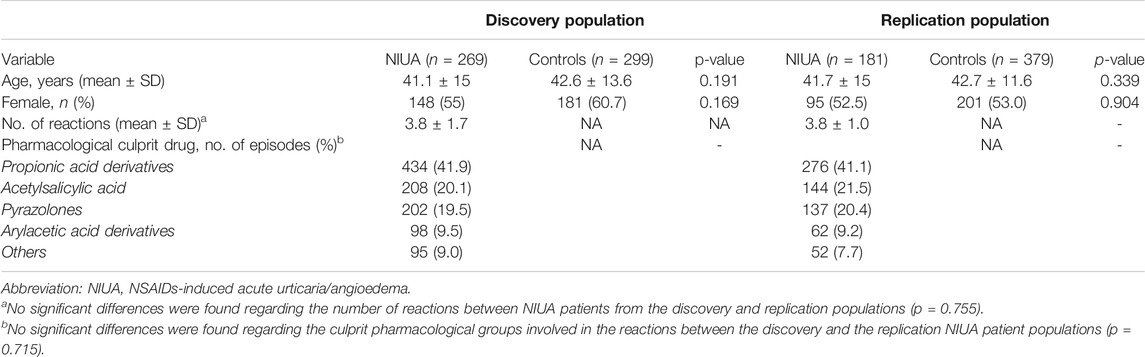
TABLE 1. Demographic and clinical data for NIUA patients and NSAID-tolerant controls.
To be included, patients had to have reported at least two episodes of acute urticaria (NIUA) induced by NSAIDs belonging to at least two different chemical groups. Patients with CSU (i.e., the reoccurrence of urticaria for more than 6 weeks with no external trigger) were ruled out according to their clinical history. NSAID cross-hypersensitivity was confirmed by ASA-DPT, as described (Doña et al., 2018).
Age- and sex-matched individuals were included as control subjects. They all reported taking NSAIDs regularly or on occasion without developing a clinical reaction, and had no history of CSU, drug hypersensitivity, rhinitis and/or asthma, or nasal polyposis.
The study was approved by the Ethics Committee of the participant centers and conducted according to the principles of the Declaration of Helsinki. All participants gave informed consent.
ASA Drug Provocation Test
ASA-DPT was performed in a single-blind manner, giving placebo capsules at different times on the 1st day, as reported previously (Doña et al., 2018). ASA and placebo were given in opaque capsules prepared by the hospital pharmacy service. Other medications were withheld before testing, according to international guidelines (Nizankowska-Mogilnicka et al., 2007).
On the 2nd day, two doses of ASA were administered orally with an interval of 3 h (50 and 100 mg). If no reaction occurred, two larger doses (250 and 500 mg) were administered on the 3rd day, with a 3-h interval. The procedure was stopped if cutaneous and/or respiratory symptoms or changes in vital signs (cardiac rhythm alterations, decrease in peak expiratory flow, or hypotension) appeared, and symptoms were evaluated and treated (Doña et al., 2018). If no symptoms appeared during these periods, this was followed by a 2-day/8-h course of the therapeutic dose (500 mg) after a gap of 24 h (Doña et al., 2018).
Genotype Determination
Data from individuals of European ancestry were downloaded from the 1,000 Genome Project website (phase 3) (https://www.internationalgenome.org/data-portal/data-collection/phase-3). The region corresponding to PLA2G4A plus 2 kb up and downstream from the gene start and end positions, respectively, were extracted using Tabix (Li, 2011) and VCFtools (Danecek et al., 2011) (Table 2). The resulting file was converted to plink format to be loaded into Haploview (Barrett et al., 2005). Non-biallelic and indel markers were not considered. Common tagSNPs (MAF≥5%) were selected based on linkage disequilibrium with Tagger (de Bakker et al., 2005), using pairwise tagging with an r2 threshold of 0.8.
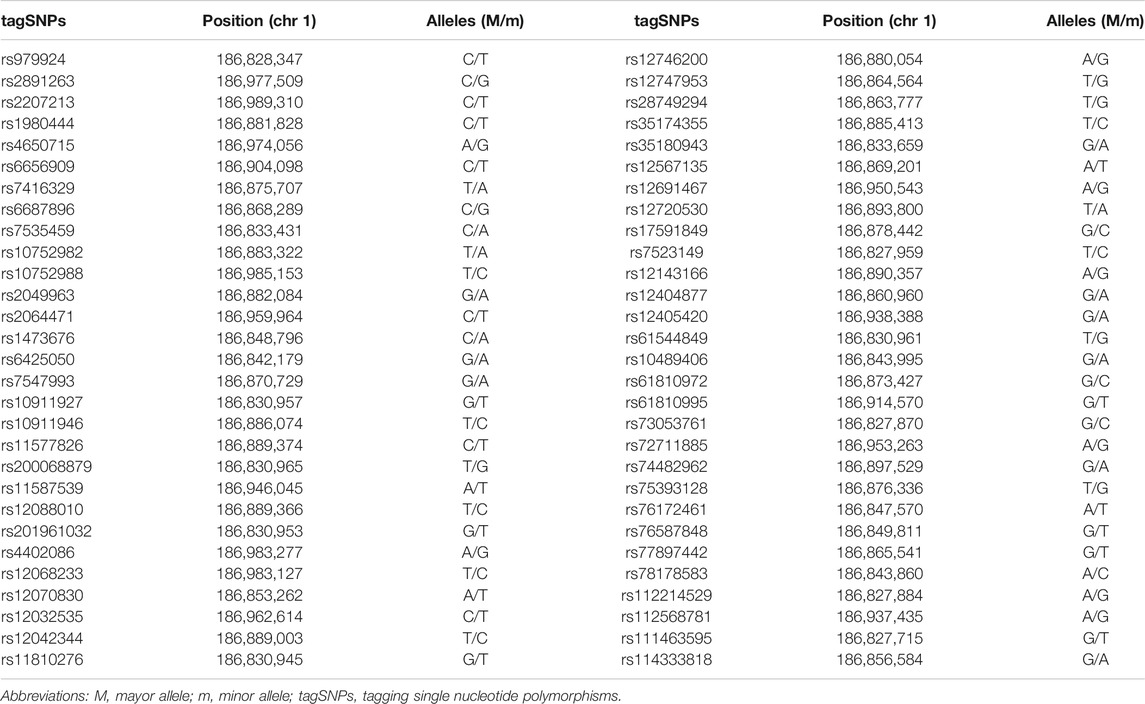
TABLE 2. Selected tagSNPs.
Genomic DNA was isolated from peripheral blood using the FlexiGene DNA Kit system (QIAGEN). Genotyping during the discovery phase was performed using the iPlex Sequenom MassArray technology, whereas Taqman probes and qPCR were used for the replication population. To ensure that the different technologies gave identical results, 10 samples from each population were genotyped using both systems, finding 100% agreement.
Assuming a disease prevalence of 0.01, a risk allele frequency of 0.2, and a relative risk of >1.5, the discovery phase of this study was estimated to have 80% power to detect an associated tagSNP with α = 0.05 (Purcell et al., 2003).
Statistical Analysis
The Mann–Whitney U test was used for continuous variables comparisons between groups, and the χ2 test for categorical variables. Hardy–Weinberg equilibrium (HWE) and individual SNP association tests with NIUA and NERD were performed using SNPassoc (Gonzalez et al., 2007), evaluating dominant, recessive, and additive models for all tagSNPs. The latter was performed using logistic regressions to calculate tagSNP effects as odd ratios with 95% confidence intervals. A p-value ≤0.05 after Bonferroni correction was considered statistically significant. Bonferroni correction thresholds were 8.62E-4 (0.05/58) for the discovery cohort, and 0.0125 (0.05/4) for the replication one.
Results
Demographic and Clinical Data
A total of 1,128 individuals were genotyped, comprising NIUA patients and controls from two unrelated, consecutively recruited populations. Table 1 shows demographic and clinical data from both the discovery and replication populations. No significant differences in sex or age between patients and controls were found in either of the considered groups (Table 1). Mean age for NIUA subjects was 41 ± 15 years and 42.6 ± 13.6 years for controls from the discovery population (p = 0.191), whereas in the replication population, mean age for NIUA was 41.7 ± 15 years and 42.7 ± 11.6 years for controls (p = 0.033).
According to their clinical history, more than 80% of NIUA patients had suffered at least three reactions of cross-hypersensitivity to NSAIDs; however, final diagnosis was established in all patients on the basis of a positive ASA-DPT, as described (Doña et al., 2018). We did not find a statistically significant difference in the total number of reactions when comparing NIUA patients from both the discovery and the replication populations (p = 0.755) (Table 1). In addition, no statistically significant difference was found between NIUA patients from the two populations included for the pharmacological drug groups inducing the reactions (p = 0.715) (Table 1). For the two patient groups, propionic derivatives were the most frequent triggers, followed by ASA and pyrazolones (Table 1).
Genetic Association Study
Fifty-eight tagSNPs were selected and genotyped. As none of them deviated significantly from HWE in the control group, they were all retained for association analyses. The genotyping call rate for all tagSNPS was >99% across all individuals.
Table 3 shows the results of genetic association testing in the discovery population under the dominant, recessive, and additive inheritance models.
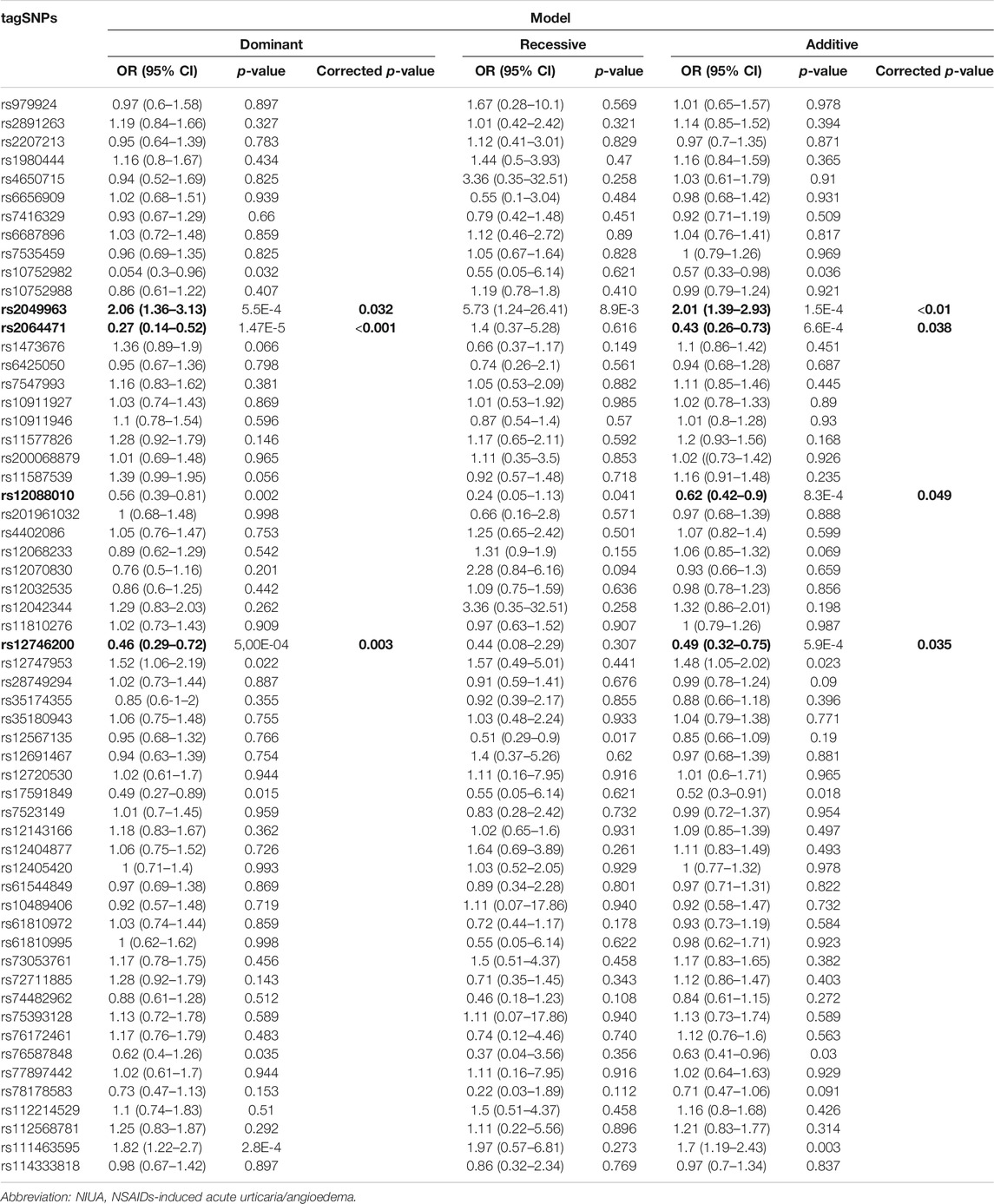
TABLE 3. Association study with the selected tagSNPs in the discovery population of NIUA patients and NSAID-tolerant controls. Statistically significant associations after Bonferroni correction are indicated in bold.
The rs20449963 variant was associated with an increased risk of NIUA. This remained statistically significant after Bonferroni multiple test correction under the dominant and additive models (OR = 2.06, 95% CI = 1.36–3.13, corrected p-value = 0.032; and OR = 2.01, 95% CI = 1.39–2.93, corrected p-value = 0.009, respectively) (Table 3). MAF for controls was 0.079, and for patients 0.152. The tagSNP rs2064471 was associated with a diminished risk of developing NIUA, and this association also was significant under the dominant and additive model when considering the multiple comparisons performed (OR = 0.27, 95% CI = 0.14–0.52, corrected p-value<0.001; and OR = 0.43, 95% CI = 0.26–0.73, corrected p-value = 0.038, respectively) (Table 3). MAF values for controls and patients were 0.082 and 0.026, respectively. In addition, the rs12746200 variant was protective after Bonferroni correction according to both the dominant and additive models (OR = 0.46, 95% CI = 0.29–0.72, corrected p-value = 0.003; and OR = 0.49, 95% CI = 0.32–0.75, corrected p-value = 0.035, respectively; MAF for controls = 0.125, MAF for patients = 0.208). Finally, the rs12088010 polymorphism was also associated with a diminished risk of NIUA under the additive inheritance model (OR = 0.62, 95% CI = 0.42–0.9; corrected p-value = 0.049; MAF values of 0.094 and of 0.119 for controls and patients, respectively) (Table 3).
Marginal associations were found between NIUA and the variants rs12747953, rs17591849, rs76587848, and rs111463595 under the dominant and additive models; however, these associations did not retain statistical significance after multiple testing correction. In addition, the rs12567135 was also marginally associated with a diminished risk of NIUA under a recessive inheritance model (Table 3).
To validate these initial findings, all four nominally associated tagSNPs found in the NIUA discovery cohort were further genotyped in an independent case–control population (replication cohort), and tested for association after verifying that none of them departed significantly from HWE in the control group.
The rs2049963 variant was associated with a statistically significant increased risk for developing NIUA under the dominant (OR = 1.79, 95% CI = 1.14–2.82; corrected p-value = 0.048) and additive models (OR = 1.79, 95% CI = 1.21–2.65; corrected p-value = 0.016), although the association of this variant with NIUA when considering the recessive model did not surpass Bonferroni correction (Table 4). The polymorphism rs1288010 showed a protective role under the dominant (OR = 0.57, 95% CI = 0.37–0.87; corrected p-value = 0.032) and additive models (OR = 0.59, 95% CI = 0.4–0.88; corrected p-value = 0.028). Similar results were found for the variant rs12746200, which was significantly associated with NIUA under these two inheritance models (OR = 0.51, 95% CI = 0.31–0.83, corrected p-value = 0.02 for the dominant model; and OR = 0.56, 95% CI = 0.36–0.87, corrected p value = 0.028 fort the additive model). Finally, marginal associations were also detected for the rs2064471 (dominant and recessive models) (Table 4).
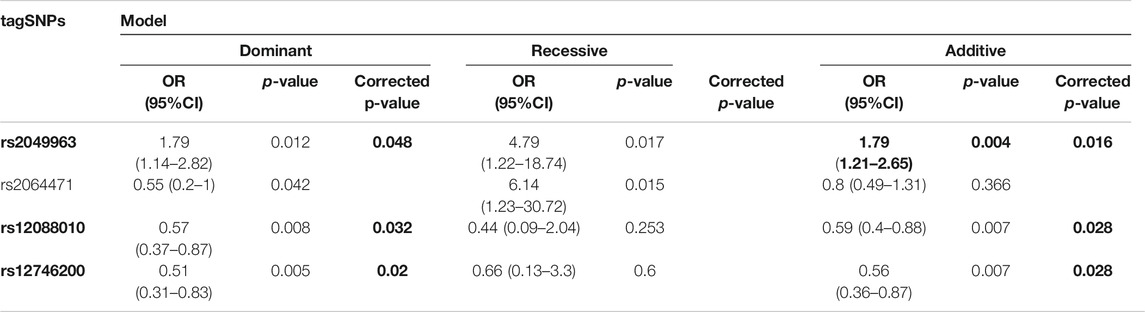
TABLE 4. Association study in the in the replication population of NIUA patients and NSAID-tolerant controls. Statistically significant associations after Bonferroni correction are indicated in bold.
Discussion
NSAIDs are widely recognized as most frequently medicines involved in drug hypersensitivity reactions, and NSAID cross-hypersensitivity as the most frequent underlying mechanism. Through the pharmacological inhibition of the COX-1 enzyme, PG production is blocked, and the AA metabolic pathway is shunted toward the biosynthesis of CysLTs. In some individuals, this results in a pathological increase of these mediators, which leads to the elicitation of a hypersensitivity response (Szczeklik et al., 1975; Doña et al., 2019). Susceptibility to develop NSAID hypersensitivity seems to be influenced by the interaction of multiple factors, with most of them still unknown. However, there is thought to be a genetic component. Although there is a lack of familial studies on NSAID hypersensitivity, some data support this idea: the familial aggregation of NIUA has been described, even though a classical Mendelian segregation pattern has not been observed (Mastalerz et al., 2006). In addition, the development of NIUA in twins has been reported recently (Caimmi et al., 2014).
In spite of NIUA being the most common NSAID-hypersensitivity phenotype (Dona et al., 2012; Dona et al., 2014; Doña et al., 2020), most genetic association studies have focused on NERD (Jurado-Escobar et al., 2017). Although they have largely investigated polymorphisms in genes related to AA metabolism, no previous work has considered overall common genetic variability in PLA2G4A. This gene encodes a cPLA2 group family member, an upstream enzyme that hydrolyzes membrane phospholipids to release AA, which is subsequently metabolized to produce CysLTs. Thus, such an enzyme may be considered the primary source of inflammatory mediators. In addition to the heterogeneity of their results, which can often not be replicated, most previous genetic studies did not take into account the overall variability in genes of potential interest, and rarely included a second population to support their findings (Jurado-Escobar et al., 2017). Here, we have evaluated the overall genetic variability of PLA2G4A and its potential association with NIUA development, through a two-stages approach, validating findings in a second population.
Four intronic variants from 58 initially considered tagSNPs in PLA2G4A were associated with NIUA in the discovery population and subsequently genotyped in the replication population. Three of them retained statistical signification after Bonferroni correction. The rs2049963 variant was associated with an increased risk of developing NIUA, whereas rs12088010 and rs12746200 were linked to a diminished risk.
We have previously described rs12746200 as associated with NIUA (Ayuso et al., 2015). Although this study did not evaluate the overall variability in PLA2G4A and a second replication case-control group was not included, it provides strong support for the results presented here, suggesting a protective role for rs12746200. This variant was also correlated with the development of familial adenomatous polyposis in the single polymorphism evaluation (Umeno et al., 2010). The rs12746200 polymorphism has been also associated with a diminished risk of myocardial infarction in patients with coronary artery disease (Hartiala et al., 2011), an association which was further replicated and which appears to be modulated by dietary polyunsaturated fatty acids, with minor allele carriers showing a significantly lower PLA2G4A gene expression (Hartiala et al., 2012). Although the authors did not check if such downregulation was also accompanied by a decrease in protein levels or the effect on other molecules, it has been reported that a PLAG4A blockade may modify gene expression pattern for a variety of proteins in the asthma-associated response (Whalen et al., 2008).
There is a lack of information regarding the clinical impact of the rs2049963 and rs12088010 intronic variants. However, introns are noncoding DNA sequences located between exons (coding regions) which have been widely recognized as key regulators of gene expression and alternative splicing, as they may act as transcription factor binding sites and also modulate transcription rate, nuclear export, and transcript stability (Shaul, 2017). The development of specific clinical entities may be influenced by intron regulation. In this sense, introns have been linked to cardiovascular (Bose et al., 2017; Wang et al., 2019) and autoimmune diseases (AlFadhli, 2013) and cancer (Neves Filho et al., 2012; Blackburn et al., 2016; Tauziède-Espariat et al., 2017; Garinet et al., 2019). Deep-sequencing technologies have also highlighted the role of intronic mutations in human diseases through different molecular mechanisms (Vaz-Drago et al., 2017; Dufner-Almeida et al., 2019). Non-coding polymorphisms in the AA metabolic pathway have been associated with diabetes (Nejatian et al., 2019) and the clinical response to the CysLT receptor antagonist montelukast (Klotsman et al., 2007). We recently reported that some intronic variants in a guanine nucleotide–binding protein to be associated with NIUA (Blanca et al., 2020); moreover, several polymorphisms in 5-lipoxygenase, a central enzyme in CysLT synthesis, have been associated with NERD (Choi et al., 2004; Kim et al., 2005). We cannot exclude also the possibility that these intronic variants are involved in changes in methylation patterns that could affect gene expression, as it has been also shown for other pathologies (Beltrami et al., 2017; Ozaki et al., 2017; Prunicki et al., 2018; McGuire et al., 2019).
In summary, our results strongly suggest that PLA2G4A genetic variants play a role in NIUA development. Particular cPLA2 isozymes have been associated with different pathologies (Runarsson et al., 2007; Niknami et al., 2010), and biological processes affecting platelet, endothelial, and leukocyte eicosanoid generation (Hurley et al., 2011; Kirkby et al., 2015) and immune regulation (Rousseau et al., 2015). However, the mechanisms supporting these associations with NIUA at the molecular level are currently unknown. In addition, we cannot exclude the possibility that the tagSNPs we have found here to be associated with NIUA may not be the causal variants but merely tagging them, and that the causal ones may be functional coding variants. To our knowledge, this is the first study addressing overall common genetic variability in PLA2G4A in NSAID cross-hypersensitivity. In addition to functional studies, further research expanding the sample size to cover rare variants and including other populations will provide a better understanding of the genetics underlying NSAID cross-hypersensitivity and its different phenotypes. This will help in the search for potential biomarkers for prognostic and diagnosis purposes.
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics Statement
The studies involving human participants were reviewed and approved by Comité Ético de Investigación Ciéntifica HRUM. The patients/participants provided their written informed consent to participate in this study.
Author Contributions
ID, JP, MT, and JC-G designed the study. ID, NP-S, and MT recruited, evaluated, and diagnosed patients from the discovery population, and JL and JB from the replication population. AT-M and ML revised all clinical data and diagnosis from the two populations. RJ-E and JT-C performed experiments and data analysis, and RJ-E drafted the manuscript, and they were supervised by JC-G. ME revised all final results. JG-A and MT revised the manuscript. ID and JC-G are responsible for the final version. All authors revised and approved the submitted version of the manuscript.
Funding
This work was supported by Instituto de Salud Carlos III (ISCIII, Spanish Ministry of Science and Innovation), co-founded by Fondo Europeo de Desarrollo Regional-FEDER for Research Projects (PI17/01593, PI18/00540, and PI20/01540), GR18145 from Junta de Extremadura, the Thematic Networks and Co-operative Research Centers ARADyAL (RD16/0006), and from the Sociedad Española de Alergología e Inmunología Clínica (SEAIC; Ref. Convocatoria Ayudas 2016 and REF: 18_B02). RJ-E holds a P-FIS PhD student grant (Ref FI18/00133), AT-M and ML from the Rio Hortega Program (references CM20/00160 and CM20/00210, respectively), and JC-G is a senior researcher from the Miguel Servet Program II (Ref CPII19/00006), all of them from the ISCIII. ID is a Clinical Investigator (B-0001–2017) from Consejería de Salud of Junta de Andalucía.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank Claudia Corazza and JP for their help with the English version of the manuscript, Tatiana Diaz from the Malaga Regional Biobank (Andalusian Public Health System Biobank), and Juan Miguel Gómez Zumaquero from the Genotyping and Sequencing Unit of IBIMA, for technical support.
References
AlFadhli, S. (2013). Influence of Endothelial Nitric Oxide Synthase Gene Intron-4 27bp Repeat Polymorphism on its Expression in Autoimmune Diseases. Dis. Markers 34 (5), 349–356. doi:10.3233/DMA-130983
Ayuso, P., Plaza-Serón, M. d. C., Doña, I., Blanca-López, N., Campo, P., Cornejo-García, J. A., et al. (2015). Association Study of Genetic Variants in PLA2G4A, PLCG1, LAT, SYK, and TNFRS11A Genes in NSAIDs-Induced Urticaria And/or Angioedema Patients. Pharmacogenet Genomics 25 (12), 618–621. doi:10.1097/FPC.0000000000000179
Barrett, J. C., Fry, B., Maller, J., and Daly, M. J. (2005). Haploview: Analysis and Visualization of LD and Haplotype Maps. Bioinformatics 21 (2), 263–265. doi:10.1093/bioinformatics/bth457
Beltrami, C. M., Dos Reis, M. B., Barros-Filho, M. C., Marchi, F. A., Kuasne, H., Pinto, C. A. L., et al. (2017). Integrated Data Analysis Reveals Potential Drivers and Pathways Disrupted by DNA Methylation in Papillary Thyroid Carcinomas. Clin. Epigenet 9, 45. doi:10.1186/s13148-017-0346-2
Blackburn, J., Roden, D. L., Ng, R., Wu, J., Bosman, A., and Epstein, R. J. (2016). Damage‐inducible Intragenic Demethylation of the Human TP53 Tumor Suppressor Gene Is Associated with Transcription from an Alternative Intronic Promoter. Mol. Carcinog. 55 (12), 1940–1951. doi:10.1002/mc.22441
Blanca, M., Oussalah, A., Cornejo‐García, J. A., Blanca‐López, N., Guéant‐Rodriguez, R. M., Doña, I., et al. (2020). GNAI2 Variants Predict Nonsteroidal Anti‐inflammatory Drug Hypersensitivity in a Genome‐wide Study. Allergy 75 (5), 1250–1253. doi:10.1111/all.14100
Bose, D., D, V., Shetty, M., J, K., and Kutty, A. V. M. (2017). Identification of Intronic-Splice Site Mutations in GATA4 Gene in Indian Patients with Congenital Heart Disease. Mutat. Research/Fundamental Mol. Mech. Mutagenesis 803-805, 26–34. doi:10.1016/j.mrfmmm.2017.08.001
Caimmi, S. M. E., Manca, E., Caimmi, D., Marseglia, G. L., and Demoly, P. (2014). NSAID Hypersensitivity in Twins. Pediatr. Allergy Immunol. 25 (8), 828–829. doi:10.1111/pai.12300
Choi, J.-H., Park, H.-S., Oh, H.-B., Lee, J.-H., Suh, Y.-J., Park, C.-S., et al. (2004). Leukotriene-related Gene Polymorphisms in ASA-Intolerant Asthma: an Association with a Haplotype of 5-lipoxygenase. Hum. Genet. 114 (4), 337–344. doi:10.1007/s00439-004-1082-1
Christie, P. E., Tagari, P., Ford-Hutchinson, A. W., Charlesson, S., Chee, P., Arm, J. P., et al. (1991). Urinary Leukotriene E4Concentrations Increase after Aspirin Challenge in Aspirin-Sensitive Asthmatic Subjects. Am. Rev. Respir. Dis. 143 (5 Pt 1), 1025–1029. doi:10.1164/ajrccm/143.5_Pt_1.1025
Conaghan, P. G. (2012). A Turbulent Decade for NSAIDs: Update on Current Concepts of Classification, Epidemiology, Comparative Efficacy, and Toxicity. Rheumatol. Int. 32 (6), 1491–1502. doi:10.1007/s00296-011-2263-6
Cornejo-García, J. A., Liou, L.-B., Blanca-López, N., Doña, I., Chen, C.-H., Chou, Y.-C., et al. (2013). Genome-wide Association Study in NSAID-Induced Acute Urticaria/angioedema in Spanish and Han Chinese Populations. Pharmacogenomics 14 (15), 1857–1869. doi:10.2217/pgs.13.166
Daffern, P. J., Muilenburg, D., Hugli, T. E., and Stevenson, D. D. (1999). Association of Urinary Leukotriene E4 Excretion during Aspirin Challenges with Severity of Respiratory Responses. J. Allergy Clin. Immunol. 104 (3 Pt 1), 559–564. doi:10.1016/s0091-6749(99)70324-6
Danecek, P., Auton, A., Abecasis, G., Albers, C. A., Banks, E., DePristo, M. A., et al. (2011). The Variant Call Format and VCFtools. Bioinformatics 27 (15), 2156–2158. doi:10.1093/bioinformatics/btr330
de Bakker, P. I. W., Yelensky, R., Pe'er, I., Gabriel, S. B., Daly, M. J., and Altshuler, D. (2005). Efficiency and Power in Genetic Association Studies. Nat. Genet. 37 (11), 1217–1223. doi:10.1038/ng1669
Dona, I., Barrionuevo, E., Blanca-Lopez, N., Torres, M. J., Fernandez, T. D., Mayorga, C., et al. (2014). Trends in Hypersensitivity Drug Reactions: More Drugs, More Response Patterns, More Heterogeneity. J. Investig. Allergol. Clin. Immunol. 24 (3), 143–153. quiz 1 p following 153.
Doña, I., Barrionuevo, E., Salas, M., Laguna, J. J., Agúndez, J., García-Martín, E., et al. (2018). NSAIDs-hypersensitivity Often Induces a Blended Reaction Pattern Involving Multiple Organs. Sci. Rep. 8 (1), 16710. doi:10.1038/s41598-018-34668-1
Dona, I., Blanca-Lopez, N., Torres, M. J., Garcia-Campos, J., Garcia-Nunez, I., Gomez, F., et al. (2012). Drug Hypersensitivity Reactions: Response Patterns, Drug Involved, and Temporal Variations in a Large Series of Patients. J. Investig. Allergol. Clin. Immunol. 22 (5), 363–371.
Doña, I., Jurado‐Escobar, R., Perkins, J. R., Ayuso, P., Plaza‐Serón, M. C., Pérez‐Sánchez, N., et al. (2019). Eicosanoid Mediator Profiles in Different Phenotypes of Nonsteroidal Anti‐inflammatory Drug‐induced Urticaria. Allergy 74 (6), 1135–1144. doi:10.1111/all.13725
Doña, I., Pérez‐Sánchez, N., Eguiluz‐Gracia, I., Muñoz-Cano, R., Bartra, J., Torres, M. J., et al. (2020). Progress in Understanding Hypersensitivity Reactions to Nonsteroidal Anti‐inflammatory Drugs. Allergy 75 (3), 561–575. doi:10.1111/all.14032
Dufner-Almeida, L. G., do Carmo, R. T., Masotti, C., and Haddad, L. A. (2019). Understanding Human DNA Variants Affecting Pre-mRNA Splicing in the NGS Era. Adv. Genet. 103, 39–90. doi:10.1016/bs.adgen.2018.09.002
Duong, M., Salvo, F., Pariente, A., Abouelfath, A., Lassalle, R., Droz, C., et al. (2014). Usage Patterns of 'over-The-Counter'vs. Prescription-Strength Nonsteroidal Anti-inflammatory Drugs in France. Br. J. Clin. Pharmacol. 77 (5), 887–895. doi:10.1111/bcp.12239
Fosbøl, E. L., Gislason, G. H., Jacobsen, S., Abildstrom, S. Z., Hansen, M. L., Schramm, T. K., et al. (2008). The Pattern of Use of Non-steroidal Anti-inflammatory Drugs (NSAIDs) from 1997 to 2005: a Nationwide Study on 4.6 Million People. Pharmacoepidem. Drug Safe. 17 (8), 822–833. doi:10.1002/pds.1592
Garinet, S., Pignot, G., Vacher, S., Le Goux, C., Schnitzler, A., Chemlali, W., et al. (2019). High Prevalence of a Hotspot of Noncoding Somatic Mutations in Intron 6 of GPR126 in Bladder Cancer. Mol. Cancer Res. 17 (2), 469–475. doi:10.1158/1541-7786.MCR-18-0363
Gonzalez, J. R., Armengol, L., Sole, X., Guino, E., Mercader, J. M., Estivill, X., et al. (2007). SNPassoc: an R Package to Perform Whole Genome Association Studies. Bioinformatics 23 (5), 654–655. doi:10.1093/bioinformatics/btm025
Hartiala, J., Gilliam, E., Vikman, S., Campos, H., and Allayee, H. (2012). Association of PLA2G4A with Myocardial Infarction Is Modulated by Dietary PUFAs. Am. J. Clin. Nutr. 95 (4), 959–965. doi:10.3945/ajcn.111.032094
Hartiala, J., Li, D., Conti, D. V., Vikman, S., Patel, Y., Wilson Tang, W. H., et al. (2011). Genetic Contribution of the Leukotriene Pathway to Coronary Artery Disease. Hum. Genet. 129 (6), 617–627. doi:10.1007/s00439-011-0963-3
Higashi, N., Taniguchi, M., Mita, H., Kawagishi, Y., Ishii, T., Higashi, A., et al. (2004). Clinical Features of Asthmatic Patients with Increased Urinary Leukotriene E4 Excretion (hyperleukotrienuria)Involvement of Chronic Hyperplastic Rhinosinusitis with Nasal Polyposis. J. Allergy Clin. Immunol. 113 (2), 277–283. doi:10.1016/j.jaci.2003.10.068
Hurley, B. P., Pirzai, W., Mumy, K. L., Gronert, K., and McCormick, B. A. (2011). Selective Eicosanoid-Generating Capacity of Cytoplasmic Phospholipase A2 in Pseudomonas Aeruginosa-Infected Epithelial Cells. Am. J. Physiology-Lung Cell Mol. Physiol. 300 (2), L286–L294. doi:10.1152/ajplung.00147.2010
Jurado-Escobar, R., Perkins, J., García-Martín, E., Isidoro-García, M., Doña, I., Torres, M., et al. (2017). Update on the Genetic Basis of Drug Hypersensitivity Reactions. J. Investig. Allergol. Clin. Immunol. 27 (6), 336–345. doi:10.18176/jiaci.0199
Kim, S.-H., Choi, J.-H., Holloway, J. W., Suh, C.-H., Nahm, D.-H., Ha, E.-H., et al. (2005). Leukotriene-related Gene Polymorphisms in Patients with Aspirin-Intolerant Urticaria and Aspirin-Intolerant Asthma: Differing Contributions of ALOX5 Polymorphism in Korean Population. J. Korean Med. Sci. 20 (6), 926–931. doi:10.3346/jkms.2005.20.6.926
Kirkby, N. S., Reed, D. M., Edin, M. L., Rauzi, F., Mataragka, S., Vojnovic, I., et al. (2015). Inherited Human Group IVA Cytosolic Phospholipase A 2 Deficiency Abolishes Platelet, Endothelial, and Leucocyte Eicosanoid Generation. FASEB j. 29 (11), 4568–4578. doi:10.1096/fj.15-275065
Klotsman, M., York, T. P., Pillai, S. G., Vargas-Irwin, C., Sharma, S. S., van den Oord, E. J. C. G., et al. (2007). Pharmacogenetics of the 5-lipoxygenase Biosynthetic Pathway and Variable Clinical Response to Montelukast. Pharmacogenet Genomics 17 (3), 189–196. doi:10.1097/FPC.0b013e3280120043
Kowalski, M. L., Asero, R., Bavbek, S., Blanca, M., Blanca-Lopez, N., Bochenek, G., et al. (2013). Classification and Practical Approach to the Diagnosis and Management of Hypersensitivity to Nonsteroidal Anti-inflammatory Drugs. Allergy 68 (10), 1219–1232. doi:10.1111/all.12260
Kowalski, M. L., Makowska, J. S., Blanca, M., Bavbek, S., Bochenek, G., Bousquet, J., et al. (2011). Hypersensitivity to Nonsteroidal Anti-inflammatory Drugs (NSAIDs) - Classification, Diagnosis and Management: Review of the EAACI/ENDA# and GA2LEN/HANNA*. Allergy 66 (7), 818–829. doi:10.1111/j.1398-9995.2011.02557.x
Li, H. (2011). Tabix: Fast Retrieval of Sequence Features from Generic TAB-Delimited Files. Bioinformatics 27 (5), 718–719. doi:10.1093/bioinformatics/btq671
Mastalerz, L., Setkowicz, M., Sanak, M., Rybarczyk, H., and Szczeklik, A. (2006). Familial Aggregation of Aspirin-Induced Urticaria and Leukotriene C4 Synthase Allelic Variant. Br. J. Dermatol. 154 (2), 256–260. doi:10.1111/j.1365-2133.2005.06851.x
McGuire, M. H., Herbrich, S. M., Dasari, S. K., Wu, S. Y., Wang, Y., Rupaimoole, R., et al. (2019). Pan-cancer Genomic Analysis Links 3'UTR DNA Methylation with Increased Gene Expression in T Cells. EBioMedicine 43, 127–137. doi:10.1016/j.ebiom.2019.04.045
Micheletto, C., Visconti, M., Tognella, S., Facchini, F. M., and Dal Negro, R. W. (2006). Aspirin Induced Asthma (AIA) with Nasal Polyps Has the Highest Basal LTE4 Excretion: a Study vs AIA without Polyps, Mild Topic Asthma, and Normal Controls. Eur. Ann. Allergy Clin. Immunol. 38 (1), 20–23.
Mita, H., Endoh, S., Kudoh, M., Kawagishi, Y., Kobayashi, M., Taniguchi, M., et al. (2001). Possible Involvement of Mast-Cell Activation in Aspirin Provocation of Aspirin-Induced Asthma. Allergy 56 (11), 1061–1067. doi:10.1111/j.1398-9995.2001.00913.x
Nejatian, N., Häfner, A.-K., Shoghi, F., Badenhoop, K., and Penna-Martinez, M. (2019). 5-Lipoxygenase (ALOX5): Genetic Susceptibility to Type 2 Diabetes and Vitamin D Effects on Monocytes. J. Steroid Biochem. Mol. Biol. 187, 52–57. doi:10.1016/j.jsbmb.2018.10.022
Neves Filho, E. H. C., Cordeiro, D. E. F., Vieira, A. P. F., and Rabenhorst, S. H. B. (2012). TP53 Codon 72 and Intron 3 Polymorphisms and Mutational Status in Gastric Cancer: an Association with Tumor Onset and Prognosis. Pathobiology 79 (6), 323–328. doi:10.1159/000338436
Niknami, M., Vignarajan, S., Yao, M., Hua, S., Witting, P. K., Kita, Y., et al. (2010). Decrease in Expression or Activity of Cytosolic Phospholipase A2α Increases Cyclooxygenase-1 Action: A Cross-Talk between Key Enzymes in Arachidonic Acid Pathway in Prostate Cancer Cells. Biochim. Biophys. Acta (Bba) - Mol. Cell Biol. Lipids 1801 (7), 731–737. doi:10.1016/j.bbalip.2010.03.003
Nizankowska-Mogilnicka, E., Bochenek, G., Mastalerz, L., Swierczynska, M., Picado, C., Scadding, G., et al. (2007). EAACI/GA2LEN Guideline: Aspirin Provocation Tests for Diagnosis of Aspirin Hypersensitivity. Allergy 62 (10), 1111–1118. doi:10.1111/j.1398-9995.2007.01409.x
Oosaki, R., Mizushima, Y., Mita, H., Shida, T., Akiyama, K., and Kobayashi, M. (1997). Urinary Leukotriene E4and 11-dehydrothromboxane B2in Patients with Aspirin-Sensitive Asthma. Allergy 52 (4), 470–473. doi:10.1111/j.1398-9995.1997.tb01032.x
Oussalah, A., Mayorga, C., Blanca, M., Barbaud, A., Nakonechna, A., Cernadas, J., et al. (2016). Genetic Variants Associated with Drugs-Induced Immediate Hypersensitivity Reactions: a PRISMA-Compliant Systematic Review. Allergy 71 (4), 443–462. doi:10.1111/all.12821
Ozaki, Y., Yoshino, Y., Yamazaki, K., Sao, T., Mori, Y., Ochi, S., et al. (2017). DNA Methylation Changes at TREM2 Intron 1 and TREM2 mRNA Expression in Patients with Alzheimer's Disease. J. Psychiatr. Res. 92, 74–80. doi:10.1016/j.jpsychires.2017.04.003
Prunicki, M., Stell, L., Dinakarpandian, D., de Planell-Saguer, M., Lucas, R. W., Hammond, S. K., et al. (2018). Exposure to NO2, CO, and PM2.5 Is Linked to Regional DNA Methylation Differences in Asthma. Clin. Epigenet 10, 2. doi:10.1186/s13148-017-0433-4
Purcell, S., Cherny, S. S., and Sham, P. C. (2003). Genetic Power Calculator: Design of Linkage and Association Genetic Mapping Studies of Complex Traits. Bioinformatics 19 (1), 149–150. doi:10.1093/bioinformatics/19.1.149
Rousseau, M., Naika, G. S., Perron, J., Jacques, F., Gelb, M. H., and Boilard, E. (2015). Study of the Role of Cytosolic Phospholipase A2 Alpha in Eicosanoid Generation and Thymocyte Maturation in the Thymus. PLoS One 10 (5), e0126204. doi:10.1371/journal.pone.0126204
Runarsson, G., Feltenmark, S., Forsell, P. K. A., Sjöberg, J., Björkholm, M., and Claesson, H. E. (2007). The Expression of Cytosolic Phospholipase A2and Biosynthesis of Leukotriene B4in Acute Myeloid Leukemia Cells. Eur. J. Haematol. 79 (6), 468–476. doi:10.1111/j.1600-0609.2007.00967.x
Setkowicz, M., Mastalerz, L., Podolec-Rubis, M., Sanak, M., and Szczeklik, A. (2009). Clinical Course and Urinary Eicosanoids in Patients with Aspirin-Induced Urticaria Followed up for 4 Years. J. Allergy Clin. Immunol. 123 (1), 174–178. doi:10.1016/j.jaci.2008.09.005
Shaul, O. (2017). How Introns Enhance Gene Expression. Int. J. Biochem. Cell Biol. 91 (Pt B), 145–155. doi:10.1016/j.biocel.2017.06.016
Sladek, K., Dworski, R., Soja, J., Sheller, J. R., Nizankowska, E., Oates, J. A., et al. (1994). Eicosanoids in Bronchoalveolar Lavage Fluid of Aspirin-Intolerant Patients with Asthma after Aspirin Challenge. Am. J. Respir. Crit. Care Med. 149 (4 Pt 1), 940–946. doi:10.1164/ajrccm.149.4.8143059
Sladek, K., and Szczeklik, A. (1993). Cysteinyl Leukotrienes Overproduction and Mast Cell Activation in Aspirin-Provoked Bronchospasm in Asthma. Eur. Respir. J. 6 (3), 391–399.
Szczeklik, A., Gryglewski, R. J., and Czerniawska-Mysik, G. (1975). Relationship of Inhibition of Prostaglandin Biosynthesis by Analgesics to Asthma Attacks in Aspirin-Sensitive Patients. Bmj 1 (5949), 67–69. doi:10.1136/bmj.1.5949.67
Tauziède-Espariat, A., Masliah-Planchon, J., Brugières, L., Puget, S., Dufour, C., Schneider, P., et al. (2017). Deep Intronic Hotspot Variant Explaining Rhabdoid Tumor Predisposition Syndrome in Two Patients with Atypical Teratoid and Rhabdoid Tumor. Eur. J. Hum. Genet. 25 (10), 1170–1172. doi:10.1038/ejhg.2017.115
Umeno, J., Matsumoto, T., Esaki, M., Kukita, Y., Tahira, T., Yanaru-Fujisawa, R., et al. (2010). Impact of Group IVA Cytosolic Phospholipase A 2 Gene Polymorphisms on Phenotypic Features of Patients with Familial Adenomatous Polyposis. Int. J. Colorectal Dis. 25 (3), 293–301. doi:10.1007/s00384-009-0808-x
Valkhoff, V. E., Schade, R., ‘t Jong, G. W., Romio, S., Schuemie, M. J., Arfe, A., et al. (2013). Population-based Analysis of Non-steroidal Anti-inflammatory Drug Use Among Children in Four European Countries in the SOS Project: what Size of Data Platforms and Which Study Designs Do We Need to Assess Safety Issues? BMC Pediatr. 13, 192. doi:10.1186/1471-2431-13-192
Vaz-Drago, R., Custódio, N., and Carmo-Fonseca, M. (2017). Deep Intronic Mutations and Human Disease. Hum. Genet. 136 (9), 1093–1111. doi:10.1007/s00439-017-1809-4
Wang, E., Nie, Y., Fan, X., Zheng, Z., and Hu, S. (2019). Intronic Polymorphisms in Gene of Second Heart Field as Risk Factors for Human Congenital Heart Disease in a Chinese Population. DNA Cell Biol. 38 (6), 521–531. doi:10.1089/dna.2018.4254
Whalen, K. A., Legault, H., Hang, C., Hill, A., Kasaian, M., Donaldson, D., et al. (2008). In vitro allergen Challenge of Peripheral Blood Induces Differential Gene Expression in Mononuclear Cells of Asthmatic Patients: Inhibition of Cytosolic Phospholipase A2α Overcomes the Asthma-Associated Response. Clin. Exp. Allergy 38 (10), 1590–1605. doi:10.1111/j.1365-2222.2008.03059.x
Zembowicz, A., Mastalerz, L., Setkowicz, M., Radziszewski, W., and Szczeklik, A. (2003). Safety of Cyclooxygenase 2 Inhibitors and Increased Leukotriene Synthesis in Chronic Idiopathic Urticaria with Sensitivity to Nonsteroidal Anti-inflammatory Drugs. Arch. Dermatol. 139 (12), 1577–1582. doi:10.1001/archderm.139.12.1577
Keywords: NSAID cross-hypersensitivity, urticaria/angioedema, cytosolic phospholipase A2, polymorphisms, arachidomic acid
Citation: Jurado-Escobar R, Doña I, Triano-Cornejo J, Perkins JR, Pérez-Sánchez N, Testera-Montes A, Labella M, Bartra J, Laguna JJ, Estravís M, Agúndez JAG, Torres MJ and Cornejo-García JA (2021) Genetic Variants in Cytosolic Phospholipase A2 Associated With Nonsteroidal Anti-Inflammatory Drug–Induced Acute Urticaria/Angioedema. Front. Pharmacol. 12:667824. doi: 10.3389/fphar.2021.667824
Received: 14 February 2021; Accepted: 15 April 2021;
Published: 30 April 2021.
Edited by:
Marcelo Rizzatti Luizon, Federal University of Minas Gerais, BrazilReviewed by:
Sarah E. Kleinstein, Empirico, United StatesRichard Kulmacz, University of Texas Health Science Center at Houston, United States
Copyright © 2021 Jurado-Escobar, Doña, Triano-Cornejo, Perkins, Pérez-Sánchez, Testera-Montes, Labella, Bartra, Laguna, Estravís, Agúndez, Torres and Cornejo-García. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: José A. Cornejo-García, am9zZWEuY29ybmVqb0BnbWFpbC5jb20=