
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
MINI REVIEW article
Front. Pharmacol., 18 June 2020
Sec. Pharmacology of Ion Channels and Channelopathies
Volume 11 - 2020 | https://doi.org/10.3389/fphar.2020.00913
This article is part of the Research TopicIon Channel Signalling in Cancer: From Molecular Mechanisms to TherapeuticsView all 17 articles
 Alexis Díaz-García1,2*
Alexis Díaz-García1,2* Diego Varela2,3*
Diego Varela2,3*Ion channels have recently been recognized as novel therapeutic targets in cancer research since they are overexpressed in different histological tissues, and their activity is linked to proliferation, tumor progression, angiogenesis, metastasis, and apoptosis. Voltage gated-potassium channels (VGKC) are involved in cell proliferation, cancer progression, cell cycle transition, and apoptosis. Moreover, voltage-dependent sodium channels (VGSC) contribute to decreases in extracellular pH, which, in turn, promotes cancer cell migration and invasion. Furthermore, VGSC and VGKC modulate voltage-sensitive Ca2+ channel activity by controlling the membrane potential and regulating Ca2+ influx, which functions as a second messenger in processes related to proliferation, invasion, migration, and metastasis. The subgroup of these types of channels that have shown a high oncogenic potential have become known as “oncochannels”, and the evidence has highlighted them as key potential therapeutic targets. Scorpion venoms contain a high proportion of peptide toxins that act by modulating voltage-gated Na+/K+ channel activity. Increasing scientific data have pointed out that scorpion venoms and their toxins can affect the activity of oncochannels, thus showing their potential for anticancer therapy. In this review, we provide an update of the most relevant voltage-gated Na+\K+ ion channels as cellular targets and discuss the possibility of using scorpion venom and toxins for anticancer therapy.
Ion channels are critical regulators of cellular homeostasis in excitable and non-excitable cells, regulating vital physiological processes, such as electrical signal transmission, gene expression, cell signaling pathways, hormonal secretion, learning, and memory (Bates, 2015). During oncogenic transformation, cancer cells acquire aberrant characteristics with respect to their normal counterparts, which represent the core of cancer hallmarks, such as self-sustained proliferation, tumor progression, angiogenesis, metastasis, and apoptosis resistance (Bates, 2015; Prevarskaya et al., 2018). Many genes encoding ion channels are targets of oncogenic transformation, as previously reported (Prevarskaya et al., 2018). In turn, these gene products contribute to the development of one or more cancer hallmarks, promoting the transition to a more aggressive cancer phenotype; this is exemplified by the positive correlation between ion channel overexpression and functional dysregulation with tumor progression, invasion, and metastasis (Litan and Langhans, 2015; Prevarskaya et al., 2018). The amount of evidence showcasing abnormal ion channel activity linked to carcinogenesis, cancer migration, and invasion has led to consideration of cancer as a channelopathy (Litan and Langhans, 2015; Prevarskaya et al., 2018).
In cancer, the expression changes of ion channels can be related to early diagnosis, prediction of disease aggressiveness, or as markers that allow monitoring of the response to treatment (Lastraioli et al., 2015; Kischel et al., 2019). Different ion channel subfamilies have been associated with a great variety of cancers from different histological origins and even with particular stages of cancer initiation and progression (Rao et al., 2015; Kischel et al., 2019).
In the present article, we focus on voltage-dependent K+- and Na+-channels as these are the main targets of scorpion venom in prey capture and self-defense behaviors (Quintero-Hernández et al., 2013).
K+-channels control K+ permeability, and play crucial roles in both excitable and non-excitable cells (Kuang et al., 2015). Voltage-dependent K+-channels constitute the largest and most diverse group of voltage-gated ion channels expressed in cells and comprise a pore-forming subunit (KVα subunit) that may associate with auxiliary KVβ subunits (Tian et al., 2014; Kuang et al., 2015). The KVβ subunits modify ion channel function and/or localization and increase the diversity of physiological roles associated with these ion channels, with implications in health and disease (Tian et al., 2014; Serrano-Novillo et al., 2019). The scientific literature shows a considerable amount of information indicating the role of K+-channels in cell proliferation, cancer progression (Wulff and Castle, 2010; Ouadid-Ahidouch et al., 2016), and migration (Chow et al., 2018), and at least four different mechanisms have been proposed (Figure 1), and discussed in-depth in recent dedicated reviews (Huang and Jan, 2014; Pardo and Stühmer, 2014).
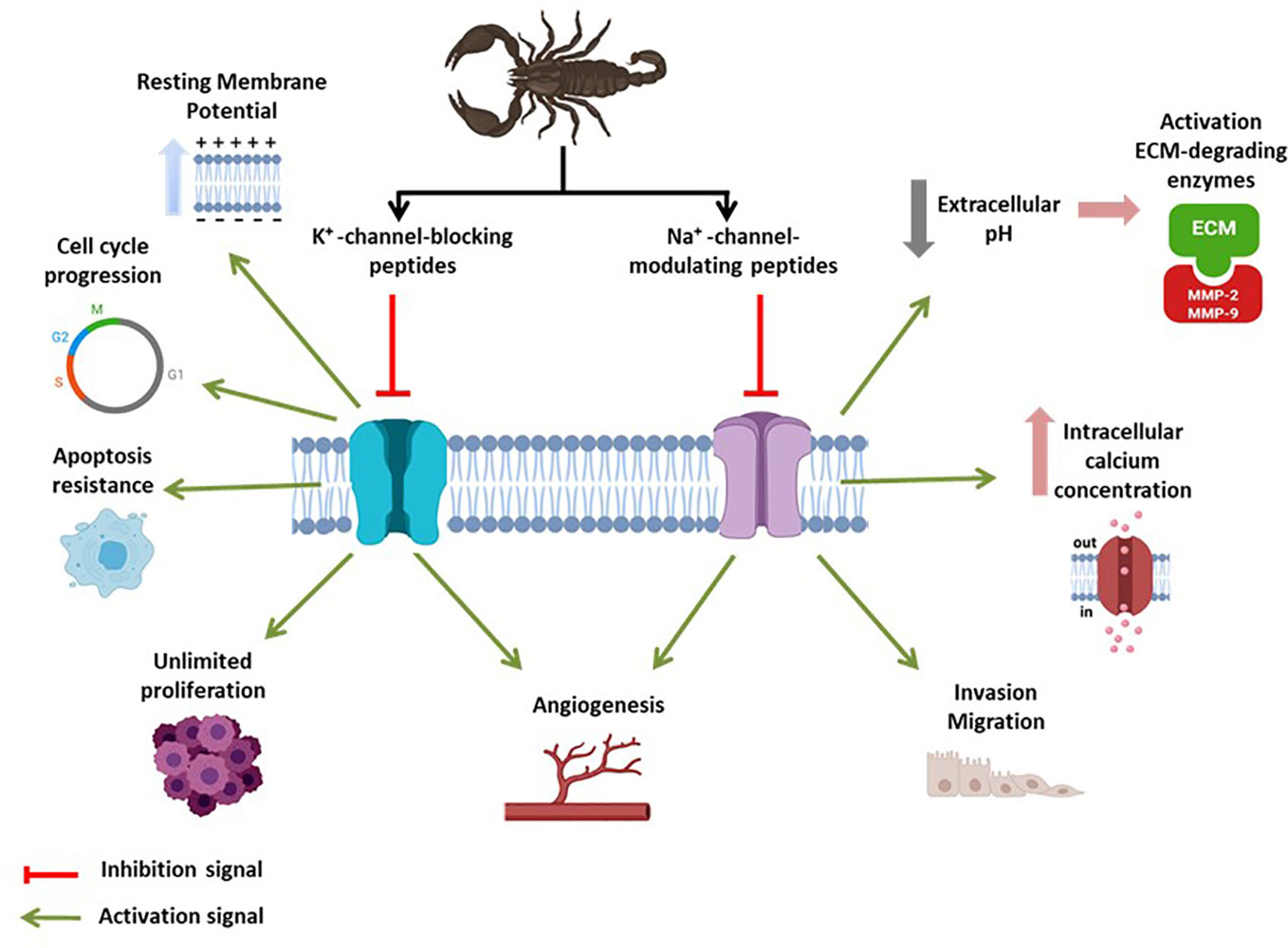
Figure 1 The global effect of scorpion toxins on cancer-related voltage-gated K+/Na+-channels. An “activation signal” (green) indicates the pathological feature of ion channel activity in the context of cancer development. An “inhibition signal” (red) indicates inhibitory action of scorpion toxins, meaning cancer-hallmark inhibition.
In cancer cells, there are significant alterations in the expression of K+-channels, which is manifested not only by the increase in their total expression, but also in the relative proportion of their different subtypes (Jiang et al., 2017; Zavala et al., 2019). The most prominent ion channel subfamilies present in primary tumors and metastases include Kv, Ether-à-go-go (EAG), and KCa (Tian et al., 2014; Kuang et al., 2015). Kv10.1, Kv11.1, KCa1.1, and Kv1.3 are the most investigated ion channels, due to their cancer hallmark-related properties. Their implication in preclinical and clinical behavior related to different cancer stages raises them as potential targets for therapy (Table 1) (Comes et al., 2015; Prevarskaya et al., 2018).
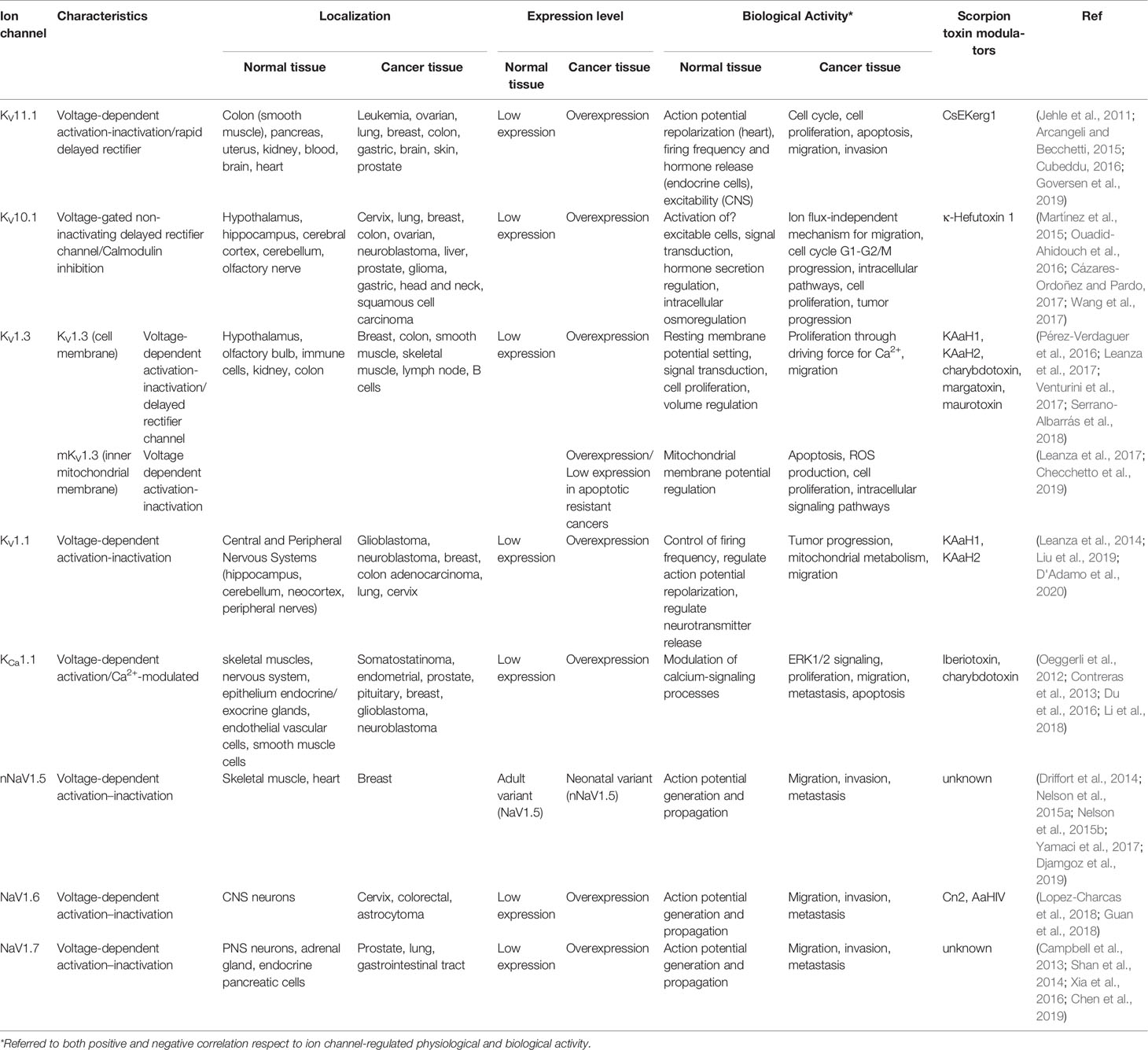
Table 1 Main characteristics of the most studied cancer-related K+/Na+-channels and their recognized modulating toxins.
KV11.1 (also known as the human Ether-à-go-go (hERG) channel) is probably the most studied ion channel in the EAG subfamily. In normal healthy tissues, its expression is usually low. In contrast, this ion channel is expressed in a higher proportion in leukemia, ovarian, lung, and breast cancer cells, among others (Jehle et al., 2011). KV11.1 channels have notable participation in the cell cycle and appear as regulators of apoptosis and cell proliferation in cancer cells (Staudacher et al., 2014; Arcangeli and Becchetti, 2015). In the heart, Kv11.1 is key for cardiac repolarization and therefore, its off-target inhibition induces long QT syndrome. Thus, safety pharmacological studies include KV11.1 channel assays as the primary test, decreasing its practical impact as an anticancer therapy-related target (Goversen et al., 2019).
KV10.1 channel is selectively expressed in brain areas (Table 1). However, this channel is overexpressed in more than 70% of tumors and in cancer cell lines from the cervix, lung, breast, ovary, neuroblast, liver, prostate, glial cells, and gastrointestinal tract (Martínez et al., 2015; Wang et al., 2017). Moreover, its crucial role in tumorigenesis, cell signaling, cell cycle, and tumor growth has been recognized (Ouadid-Ahidouch et al., 2016). Different experimental approaches have demonstrated the relationship between KV10.1 channel blockage and anticancer effects, including induction of apoptosis, inhibition of cell proliferation, and delay in tumor growth (Cázares-Ordoñez and Pardo, 2017), suggesting that this channel is a promising candidate as a tumor and therapeutic marker in oncology.
KCa1.1 channel is ubiquitously expressed in human tissues such as skeletal muscle and the nervous system, with the exception of cardiac myocytes. KCa1.1 channels regulate calcium influx into cells and thereby modulate Ca2+-signaling processes (Contreras et al., 2013). This channel is overexpressed in cancer cell lines from prostate, glia, breast, pancreas, and endometrium (Table 1) (Du et al., 2014; Du et al., 2016; Klumpp et al., 2016; Li et al., 2018; Noda et al., 2020). In the prostate, KCa1.1 channel overexpression regulates proliferation and migration (Du et al., 2016) and in breast cancer, its overexpression has been associated with advanced tumor stage, high tumor cell proliferation, and poor prognosis (Oeggerli et al., 2012).
KV1.3 channel is mostly expressed in neurons and immune cells (Pérez-Verdaguer et al., 2016). It is located at the plasma membrane, sets the resting membrane potential (RMP) and regulates cell proliferation and cell volume. Furthermore, this channel is also located in the inner mitochondrial membrane (mKV1.3), where it plays a role in apoptotic signaling (Teisseyre et al., 2019) (Table 1). Overexpression of KV1.3 channels is observed in breast, colon, smooth muscle, skeletal muscle, and lymph node cancers (Teisseyre et al., 2015; Teisseyre et al., 2019). Its plasma membrane expression is associated with controlling cell proliferation by inducing a transitory hyperpolarization necessary to augment the driving force for Ca2+ influx during G1/S progression (Serrano-Albarrás et al., 2018). Moreover, mKV1.3 channels play a role in drug-induced apoptosis by mechanisms that sensitize cancer cells (Pérez-Verdaguer et al., 2016). The potential role of KV1.3 channels as cancer therapy targets has been recently evidenced in in vitro and in vivo experimental models of glioblastoma, melanoma, and pancreatic adenocarcinoma, where mKV1.3 inhibition induces apoptotic cell death in vitro (Leanza et al., 2017; Venturini et al., 2017; Checchetto et al., 2019). All these pieces of evidence promoted KV1.3 channels as attractive potential molecular targets in both cancer diagnostics and therapy (Comes et al., 2015; Prevarskaya et al., 2018).
Notwithstanding that the ion channels mentioned above represent some of the most prominent ones in cancer; other voltage-gated ion channels linked to cancer proliferation and progression are upregulated in some tumors and have been described in dedicated reviews (Huang and Jan, 2014; Serrano-Novillo et al., 2019).
Voltage-dependent sodium channels (VGSC) are transmembrane proteins that are generally expressed in excitable cells, although they are also found, to a limited extent, in non-excitable cells (Catterall, 2012; Erickson et al., 2018). There are nine pore-forming α-subunits of sodium channels, NaV1.1-NaV1.9, encoded by the genes SCN1A-SCN11A. The pore-forming α-subunit comprises four highly similar transmembrane domains (I-IV), each composed of six transmembrane segments (S1–S6). The first four transmembrane segments of each domain constitute the voltage sensor domain, and the last two form the pore domain (Catterall, 2012). The α-subunit properties can be modulated in a subtype-specific manner, by association with one or more than one smaller auxiliary β-subunit (NaVβ1–4); conferring tissue-specific expression patterns, varying voltage dependent activation and inactivation, and increasing functional channel density at the plasma membrane (Catterall, 2017).
The oncogenic transformation of VGSC can contribute to the development of one or more cancer hallmarks, promoting the transition to more aggressive cancer phenotypes, as previously reported (Prevarskaya et al., 2018); this is particularly exemplified by the positive correlation between VGSC overexpression and functional dysregulation with invasion/migration and metastatic potential (Andrikopoulos et al., 2011; Djamgoz et al., 2019; Mao et al., 2019) (Table 1).
Proliferating and cancer cells show a RMP between -10 to -50 mV, compared to normal and non-proliferating cells (-50 to -90 mV) (Yang and Brackenbury, 2013). This RMP range fits with the window current range for VGSC, meaning that although the majority of VGSCs will be inactivated, the small percentage of non-inactivated channels will lead to a persistent Na+-current, increasing the [Na]i (Yang and Brackenbury, 2013). The augmented intracellular Na+ concentration leads to an increased intracellular Ca2+ concentration, either by promoting the reverse mode of the Na+/Ca2+ exchanger (NCX) or by inducing plasma membrane depolarization and consequent activation of voltage-sensitive Ca2+ channels (VGCC) (Patel and Brackenbury, 2015; Roger et al., 2015). Both mechanisms, driven directly or indirectly by VGSC, might be considered relevant for cancer migration and invasion. However, there are very few reports providing experimental evidence about the functional link between VGSC, NCX, and VGCC (Besson et al., 2015; Angus and Ruben, 2019; Rodrigues et al., 2019) and this aspect needs broader investigation.
A hallmark of a tumor´s extracellular space is a more acidic environment than in normal healthy tissues (pH 6.2–6.8 instead of pH 7.2–7.4), as a consequence of the predominant glycolytic metabolism of cancer cells; this particular extracellular environment enhances the degradation of the extracellular matrix by favoring Cathepsin B and S activation, and thus, promotes cell migration (Besson et al., 2015; Angus and Ruben, 2019). This extracellular acidification is dependent on Na+/H+ exchanger 1 (NHE1), which in turn depends on the [Na+] transmembrane gradient (Besson et al., 2015; Angus and Ruben, 2019). Given the increased [Na]i, a reduced NHE1 activity should be expected; however, two hypotheses have been suggested to explain this apparent contradiction. i) that these channels allosterically regulate NHE1 by inducing a higher rate of H+ extrusion at neutral pHi ranges, and ii) that the expression of VGSC in late endosome vesicles is responsible for the extra-acidification of these vesicles (Besson et al., 2015; Angus and Ruben, 2019). In this last scenario, the extracellular acidic environment would be a consequence of vesicle release.
Tetrodotoxin (TTX) is a toxin, mainly associated with fishes of the Tetraodontidae family, that specifically blocks a subgroup of VGSCs and inhibits the migration and invasion of cancer cells, indicating that cell motility requires Na+-channel activity (Nelson et al., 2015a) a feature mainly associated with overexpression of the neonatal variants of NaV1.5 (nNaV1.5), NaV1.6, and NaV1.7 (Roger et al., 2015; Mao et al., 2019).
nNaV1.5 overexpression was initially identified in the metastatic human breast cancer cell line MDA-MB-231 and breast biopsy samples (Table 1) (Yamaci et al., 2017). Later, the same positive correlation was found between the expression of nNaV1.5 channels and the high invasive potential of cancer cells from diverse histological origins (Djamgoz et al., 2019), suggesting that the overexpression of nNaV1.5 channel is necessary and sufficient to increase the metastatic potential of cancer cells (Nelson et al., 2015b).
NaV1.6 is overexpressed in cervical cancer biopsies, cancer cell lines, and primary cultures positive for the human papillomavirus (Table 1). In these cases, a NaV1.6 splice variant with preferential cytoplasmatic localization is expressed (Lopez-Charcas et al., 2018). Overexpression of NaV1.6 protein is associated with invasive status in cervical cancer and low-grade astrocytoma, mediated through increased MMP-2 activity (Lopez-Charcas et al., 2018; Guan et al., 2018).
NaV1.7 is ectopically expressed in particular types of cancers (Table 1) (Campbell et al., 2013; Xia et al., 2016; Chen et al., 2019). In gastric cancer, this channel is associated with poor patient outcomes by promoting cell invasion through the modulation of H+ efflux (Xia et al., 2016). In rat prostate cancer, NaV1.7 channel activity promotes the activation of p38/NF-κβ, and Rho GTPase signaling pathways as a linking node for controlling cellular motility, cell adhesion, and vesicular trafficking (Chen et al., 2019). In non-small cell lung cancer, the NaV1.7 channel is overexpressed in metastatic cells by more than 60% when compared to their non-metastatic counterparts (Campbell et al., 2013).
Independent of their function as auxiliary subunits, NaVβ1-3 are overexpressed in different cancers and have been associated with increased cellular motility, invasion, and metastasis (O'Malley and Isom, 2015; Bouza and Isom, 2018). Additionally, NaVβ1 has been linked to tumor growth, increase of vascular endothelial growth factor secretion, and angiogenesis (O'Malley and Isom, 2015; Bouza and Isom, 2018). In contrast, NaVβ3 functions as a tumor suppressor by inducing p53-dependent apoptosis when overexpressed (Bouza and Isom, 2018). Thus, the NaVβ-subunits are interesting and poorly explored potential targets for cancer therapy, needing an in-depth investigation to identify their complete clinical and physiopathological relevance.
Overall, VGSCs and NaVβ are up-regulated in numerous types of metastatic cancer cells and play important roles in regulating cell migration and invasion in solid tumors. Therefore, they can be considered as key regulators of cancer development and the metastatic cascade (Mao et al., 2019). The noncanonical activity of VGSC that regulates other cancer hallmarks (i.e., cell proliferation) is scarcely understood and needs to be investigated with more detail (Black and Waxman, 2013).
Worldwide, there are more than 2,200 scorpion species, grouped in 19 families (Ward et al., 2018). The scorpion venom is a complex mixture containing a great variety of proteins with molecular weights between 3 kDa and 90 kDa, which constitute most of the components. The main biological activity of the scorpion venom is due to the presence of low molecular weight peptide toxins of basic nature, which are highly cross-linked (3–4 disulfide bridges) (Quintero-Hernández et al., 2013; Kuzmenkov et al., 2015). These peptides exhibit different pharmacological and toxicological activities (Quintero-Hernández et al., 2013; Kuzmenkov et al., 2015). Until now, only a few scorpion species have been experimentally tested as anticancer agents, mainly for cancer cells from solid tumors and to a lesser extent, for hematopoietic cancers (Raposo, 2017).
In only two cases (B. martensii and R. junceus), the scientific results correlate with the experiences in traditional medicine and with the low toxicity recognized in toxicological experiments in mice (Wang and Ji, 2005; Diaz-Garcia et al., 2019a; Díaz-García et al., 2019b). The anticancer effect of B. martensii scorpion venom has been tested successfully against human glioma U251-MG by using rodent xenograft models (Wang and Ji, 2005). Likewise, in vivo toxicological studies have been carried out, using R. junceus venom administered through intraperitoneal (10 mg/kg) or oral (2,000 mg/kg) routes, and toxic effects have not been observed (Garcia-Gomez et al., 2011; Lagarto et al., 2020). Pharmacokinetic and biodistribution studies carried out on breast tumor-bearing mice administered with a single dose (12.5 mg/kg), by intravenous or oral routes, showed that medium residence time (MRT) of venom in tumor tissue was higher than in the remaining organs tested, suggesting a high selectivity for tumor tissue, adding to their antitumor effect (Diaz-Garcia et al., 2019a). Additionally, breast tumor-bearing mice injected intraperitoneally with ten consecutive doses of R. junceus venom (3.2 mg/kg), showed reduced tumor progression and reduction of Ki67 and CD31 tumor markers, confirming its anticancer potential (Díaz-García et al., 2019b). Two additional scorpion species, Androctonus amoreuxi (Salem et al., 2016) and Leiurus quinquestriatus (Al Asmari and Khan, 2016), have been tested with some favorable in vivo anticancer effects, even though both are two of the most dangerous species (Ward et al., 2018). These overall promising results have focused the scientific research on the isolation and identification of the components responsible for the anticancer effects of scorpion venoms.
Peptides recognizing K+- and Na+-channels are prominent in scorpion venoms, constituting more than 75% of all peptide/proteins (de Oliveira et al., 2018; Cid-Uribe et al., 2019). Most peptides recognizing K+ channels are pore-blocking peptides and some of them have been studied in the context of cancer (Table 1). For example, KAaH1, a KV1.1 and KV1.3 blocker, and KAaH2, a KV1.1 blocker, both derived from the Androctonus australis Hector venom, have shown anticancer potential (Aissaoui et al., 2018). KAaH1 inhibits migration and adhesion of different cancer cells, whereas KAaH2 inhibits the proliferation of gliomas (Aissaoui et al., 2018). Evidence indicates that iberiotoxin inhibits cell proliferation, migration, and invasion in breast and endometrial cancer cell lines, due to its blocking effects on BK channels (Schickling et al., 2015; Li et al., 2018); while charybdotoxin, a known blocker of KCa3.1, KV1.3, and BK channels, inhibits proliferation and cell cycle progression in pancreatic and endometrial cancer cell lines (Jager et al., 2004; Schickling et al., 2015; Li et al., 2018). Both toxins were isolated from the Leiurus quinquestriatus scorpion. Similarly, margatoxin (MgTX), a peptide isolated from Centruroides margaritatus, is a selective KV1.3-blocker that reduces cell proliferation, and tumor progression, decreases the expression of cell cycle regulators and increases the expression level of proapoptotic proteins in cancer experimental models (Jang et al., 2011). CsEKerg1 toxin, from the Centruroides sculpturatus scorpion has been evaluated as a hERG current inhibitor in an in vitro cancer model, suggesting its potential use in Kv11.1 channel-overexpressing cancer cells (Nastainczyk et al., 2002); this result opens a window of opportunity for other Kv11.1-blocking toxins described until now (Jimenez-Vargas et al., 2012). κ-Hefutoxin 1 from Heterometrus fulvipes scorpion venom (Moreels et al., 2017) has been identified as the first toxin recognizing KV10.1 channels, without affecting other voltage-gated K+-channels (Moreels et al., 2017). Moreover, maurotoxin isolated from Scorpio maurus palmatus scorpion can block various potassium channels, including SK, IK, KV1.1, and KV1.3, some of which have been recognized as cancer-related ion channels (Castle et al., 2003). Tapamin, a toxin isolated from the Mesobuthus tamulus scorpion, can block some cancer-related ion channels, such as SK and KCa3.1, and exerts a cytotoxic effect on cancer cells (Pedarzani et al., 2002; Ramirez-Cordero et al., 2014).
Although Na+-channel-modulating peptides represent the highest percentage among all scorpion venom-derived toxins (Cid-Uribe et al., 2019), the identification of scorpion venom peptides that interact with metastatic-related Na+ channels has been difficult, and only three cases have been identified (Table 1). Cn2, a β-toxin from Centruroides noxius Hoffmann scorpion venom, modulates NaV1.6 activity in F11 neuroblastoma cells (Escalona et al., 2014). In cell culture, Cn2 reduces proliferation by increasing cells at the SubG1 and G0/G1 stages, leading to apoptosis induction (Escalona et al., 2014). This toxin binds to the receptor site 4, located in the S3–S4 and S1–S2 extracellular loops of the VGSC channel domain II, enhancing channel activation by shifting the voltage-dependence of channel activation to the left, as a consequence of voltage-sensor trapping (Cestele et al., 1998), and reducing the Na+ current peak amplitude (Pedraza Escalona and Possani, 2013). AGAP, isolated from Buthus martensii, is an α-toxin that interacts with Na+-channels. Evidence suggests that AGAP affects the translation of the NaVβ1 subunit in cancer cells and has been successfully evaluated against Ehrlich ascites tumor and S-180 fibrosarcoma models in vivo. Furthermore, this peptide can inhibit cancer cell stemness, epithelial-mesenchymal transition (EMT), migration, and invasion in MCF-7 and MDA-MB-231 human breast cancer cells in vitro and tumor growth in vivo (Guo et al., 2016; Kampo et al., 2019). Finally, AaHIV toxin, isolated from Androctonus australis venom, is a Na+ channel-modulating toxin active against cancer cells (BenAissa et al., 2019). AaHIV can interact with the extracellular loops of segments S1–S2 in the voltage sensor domain, prolonging the inactivation recovery time of Nav1.6 channels, and inhibiting cancer cell proliferation in a dose-dependent manner (BenAissa et al., 2019). Unlike anti-migratory and anti-metastatic properties, the antiproliferative properties of Na+-channel-interacting scorpion toxins represent an unexpected feature that should be deeply investigated. There is no doubt that scorpion venom peptide toxins inhibit the functional activity of voltage-gated K+/Na+-channels, reducing their impact on the hallmark of cancer (Figure 1).
It is worth mentioning that Chlorotoxin is the only toxin from scorpion venom that has been successfully evaluated in cancer preclinical and clinical trials (Cohen-Inbar and Zaaroor, 2016; Mahadevappa et al., 2017; Cohen et al., 2018). However, this toxin recognizes voltage-dependent Cl- channels (Dardevet et al., 2015), which was not within the scope of this review.
Evidence indicates that upregulation of voltage-dependent K+ and Na+ channels is linked to cancer hallmarks. Thus, they have become key player as new alternatives to be used as diagnostic, prognostic, and therapeutic targets in cancer. Scorpion venoms contain small peptides acting either at the cell membrane or intracellularly, and even cross the blood-brain barrier. The mechanisms of action of scorpion venom toxins described here, related to ion channel-modulating effects, give new insights to the plethora of potential new mechanisms of action that could be discovered from scorpion venom peptides. Laboratories dedicated to scorpion venom research have usually described the anticancer effects of scorpion venom and/or components for the first time; far away from the anticancer drug development programs and their resources. There is no doubt that the inclusion of these natural products, such as plant extracts, as part of the anticancer drug discovery programs, might increase the arsenal of active components as potential new drugs against relatively new targets. Importantly, the interaction of both research areas might represent a substantial qualitative leap that could open a highway of promising alternatives to be used as adjuvant therapeutic approaches or conventional treatment in anticancer therapy.
Both authors contributed equally to the writing and preparation of the manuscript.
The Millennium Nucleus of Ion Channel-Associated Diseases (MiNICAD) is a Millennium Nucleus supported by the Iniciativa Científica Milenio of the Ministry of Economy, Development, and Tourism (Chile). This work was supported by Vicerrectoría de Investigación y Desarrollo, Universidad de Chile (VID‐Enlace, ENL24/19).
AD-G works for LifEscozul Chile SpA.
The remaining author declares that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Aissaoui, D., Mlayah-Bellalouna, S., Jebali, J., Abdelkafi-Koubaa, Z., Souid, S., Moslah, W., et al. (2018). Functional role of Kv1.1 and Kv1.3 channels in the neoplastic progression steps of three cancer cell lines, elucidated by scorpion peptides. Int. J. Biol. Macromol. 111, 1146–1155. doi: 10.1016/j.ijbiomac.2018.01.144
Al Asmari, A. K., Khan, A. Q. (2016). Investigation of in vivo potential of scorpion venom against skin tumorigenesis in mice via targeting markers associated with cancer development. Drug Des. Devel. Ther. 10, 3387–3397. doi: 10.2147/DDDT.S113171
Andrikopoulos, P., Fraser, S. P., Patterson, L., Ahmad, Z., Burcu, H., Ottaviani, D., et al. (2011). Angiogenic functions of voltage-gated Na+ Channels in human endothelial cells: modulation of vascular endothelial growth factor (VEGF) signaling. J. Biol. Chem. 286 (19), 16846–16860. doi: 10.1074/jbc.M110.187559
Angus, M., Ruben, P. (2019). Voltage gated sodium channels in cancer and their potential mechanisms of action. Channels (Austin) 13 (1), 400–409. doi: 10.1080/19336950.2019.1666455
Arcangeli, A., Becchetti, A. (2015). Novel perspectives in cancer therapy: Targeting ion channels. Drug Resist. Update 21-22, 11–19. doi: 10.1016/j.drup.2015.06.002
Bates, E. (2015). Ion channels in development and cancer. Annu. Rev. Cell Dev. Biol. 31, 231–247. doi: 10.1146/annurev-cellbio-100814-125338
BenAissa, R., Othman, H., Villard, C., Peigneur, S., Mlayah-Bellalouna, S., Abdelkafi-Koubaa, Z., et al. (2019). AaHIV a sodium channel scorpion toxin inhibits the proliferation of DU145 prostate cancer cells. Biochem. Biophys. Res. Commun. 521 (2), 340–346. doi: 10.1016/j.bbrc.2019.10.115
Besson, P., Driffort, V., Bon, E., Gradek, F., Chevalier, S., Roger, S. (2015). How do voltage-gated sodium channels enhance migration and invasiveness in cancer cells? Biochim. Biophys. Acta 1848 (10 Pt B), 2493–2501. doi: 10.1016/j.bbamem.2015.04.013
Black, J. A., Waxman, S. G. (2013). Noncanonical roles of voltage-gated sodium channels. Neuron 80 (2), 280–291. doi: 10.1016/j.neuron.2013.09.012
Bouza, A. A., Isom, L. L. (2018). Voltage-Gated Sodium Channel beta Subunits and Their Related Diseases. Handb. Exp. Pharmacol. 246, 423–450. doi: 10.1007/164_2017_48
Cázares-Ordoñez, V., Pardo, L. (2017). Kv10.1 potassium channel: from the brain to the tumors. Biochem. Cell Biol. 95 (5), 531–536. doi: 10.1139/bcb-2017-0062
Campbell, T., Main, M., Fitzgerald, E. (2013). Functional expression of the voltage-gated Na+-channel Nav1.7 is necessary for EGF-mediated invasion in human non-small cell lung cancer cells. J. Cell Sci. 126 ((Pt 21), 4939–4949. doi: 10.1242/jcs.130013
Castle, N. A., London, D. O., Creech, C., Fajloun, Z., Stocker, J. W., Sabatier, J. M. (2003). Maurotoxin: a potent inhibitor of intermediate conductance Ca2+-activated potassium channels. Mol. Pharmacol. 63 (2), 409–418. doi: 10.1124/mol.63.2.409
Catterall, W. (2012). Voltage-gated sodium channels at 60: structure, function and pathophysiology. J. Physiol. 590 (11), 2577–2589. doi: 10.1113/jphysiol.2011.224204
Catterall, W. (2017). Forty Years of Sodium Channels: Structure, Function, Pharmacology, and Epilepsy. Neurochem. Res. 42 (9), 2495–2504. doi: 10.1007/s11064-017-2314-9
Cestele, S., Qu, Y., Rogers, J. C., Rochat, H., Scheuer, T., Catterall, W. A. (1998). Voltage sensor-trapping: enhanced activation of sodium channels by beta-scorpion toxin bound to the S3-S4 loop in domain II. Neuron 21 (4), 919–931. doi: 10.1016/s0896-6273(00)80606-6
Checchetto, V., Prosdocimi, E., Leanza, L. (2019). Mitochondrial Kv1.3: a New Target in Cancer Biology? Cell Physiol. Biochem. 53 (S1), 52–62. doi: 10.33594/000000195
Chen, B., Zhang, C., Wang, Z., Chen, Y., Xie, H., Li, S., et al. (2019). Mechanistic insights into Nav1.7-dependent regulation of rat prostate cancer cell invasiveness revealed by toxin probes and proteomic analysis. FEBS J. 286 (13), 2549–2561. doi: 10.1111/febs.14823
Chow, L., Cheng, K., Wong, K., Leung, Y. (2018). Voltage-gated K+ channels promote BT-474 breast cancer cell migration. Chin. J. Cancer Res. 30 (6), 613–622. doi: 10.21147/j.issn.1000-9604.2018.06.06
Cid-Uribe, J. I., Meneses, E. P., Batista, C. V. F., Ortiz, E., Possani, L. D. (2019). Dissecting Toxicity: The Venom Gland Transcriptome and the Venom Proteome of the Highly Venomous Scorpion Centruroides limpidus (Karsch, 1879). Toxins (Basel) 11 (5), 247. doi: 10.3390/toxins11050247
Cohen, G., Burks, S. R., Frank, J. A. (2018). Chlorotoxin-A Multimodal Imaging Platform for Targeting Glioma Tumors. Toxins (Basel) 10 (12), 496. doi: 10.3390/toxins10120496
Cohen-Inbar, O., Zaaroor, M. (2016). Glioblastoma multiforme targeted therapy: The Chlorotoxin story. J. Clin. Neurosci. 33, 52–58. doi: 10.1016/j.jocn.2016.04.012
Comes, N., Serrano-Albarrás, A., Capera, J., Serrano-Novillo, C., Condom, E., Ramón, Y., et al. (2015). Involvement of potassium channels in the progression of cancer to a more malignant phenotype. Biochim. Biophys. Acta 1848 (10), 2477–2492. doi: 10.1016/j.bbamem.2014.12.008
Contreras, G. F., Castillo, K., Enrique, N., Carrasquel-Ursulaez, W., Castillo, J. P., Milesi, V., et al. (2013). A BK (Slo1) channel journey from molecule to physiology. Channels (Austin) 7 (6), 442–458. doi: 10.4161/chan.26242
Cubeddu, L. (2016). Drug-induced Inhibition and Trafficking Disruption of ion Channels: Pathogenesis of QT Abnormalities and Drug-induced Fatal Arrhythmias. Curr. Cardiol. Rev. 12 (2), 141–154. doi: 10.2174/1573403X12666160301120217
D'Adamo, M. C., Liantonio, A., Rolland, J. F., Pessia, M., Imbrici, P. (2020). Kv1.1 Channelopathies: Pathophysiological Mechanisms and Therapeutic Approaches. Int. J. Mol. Sci. 21 (8), 2935. doi: 10.3390/ijms21082935
Dardevet, L., Rani, D., Aziz, T. A., Bazin, I., Sabatier, J. M., Fadl, M., et al. (2015). Chlorotoxin: a helpful natural scorpion peptide to diagnose glioma and fight tumor invasion. Toxins (Basel) 7 (4), 1079–1101. doi: 10.3390/toxins7041079
de Oliveira, U. C., Nishiyama, M. Y., Jr., Dos Santos, M. B. V., Santos-da-Silva, A. P., Chalkidis, H. M., Souza-Imberg, A., et al. (2018). Proteomic endorsed transcriptomic profiles of venom glands from Tityus obscurus and T. serrulatus scorpions. PloS One 13 (3), e0193739. doi: 10.1371/journal.pone.0193739
Diaz-Garcia, A., Pimentel Gonzalez, G., Basaco Bernabeu, T., Rodriguez Aurrecochea, J. C., Rodriguez Sanchez, H., Sanchez Monzon, I., et al. (2019a). Pharmacokinetics and Biodistribution of Rhopalurus junceus Scorpion Venom in Tumor-Bearing Mice after Intravenous and Oral Administration. Iran BioMed. J. 23 (4), 287–296. doi: 10.29252/ibj.23.4.287
Díaz-García, A., Ruiz-Fuentes, J. L., Frión-Herrera, Y., Yglesias-Rivera, A., Riquenez Garlobo, Y., Rodríguez Sánchez, H., et al. (2019b). Rhopalurus junceus scorpion venom induces antitumor effect in vitro and in vivo against a murine mammary adenocarcinoma model. Iranian J. Basic Med. Sci. 22 (7), 759–765. doi: 10.22038/ijbms.2019.33308.7956
Djamgoz, M., Fraser, S., Brackenbury, W. (2019). In Vivo Evidence for Voltage-Gated Sodium Channel Expression in Carcinomas and Potentiation of Metastasis. Cancers (Basel) 11 (11), E1675. doi: 10.3390/cancers11111675
Driffort, V., Gillet, L., Bon, E., Marionneau-Lambot, S., Oullier, T., Joulin, V., et al. (2014). Ranolazine inhibits NaV1.5-mediated breast cancer cell invasiveness and lung colonization. Mol. Cancer 13, 264. doi: 10.1186/1476-4598-13-264
Du, C., Chen, L., Zhang, H., Wang, Z., Liu, W., Xie, X., et al. (2014). Caveolin-1 limits the contribution of BKCa channel to MCF-7 breast cancer cell proliferation and invasion. Int. J. Mol. Sci. 15 (11), 20706–20722. doi: 10.3390/ijms151120706
Du, C., Zheng, Z., Li, D., Chen, L., Li, N., Yi, X., et al. (2016). BKCa promotes growth and metastasis of prostate cancer through facilitating the coupling between alphavbeta3 integrin and FAK. Oncotarget 7 (26), 40174–40188. doi: 10.18632/oncotarget.9559
Erickson, A., Deiteren, A., Harrington, A., Garcia-Caraballo, S., Castro, J., Caldwell, A., et al. (2018). Voltage-gated sodium channels: (NaV )igating the field to determine their contribution to visceral nociception. J. Physiol. 596 (5), 785–807. doi: 10.1113/JP273461
Escalona, M. P., Batista, C. V., Cassulini, R. R., Rios, M. S., Coronas, F. I., Possani, L. D. (2014). A proteomic analysis of the early secondary molecular effects caused by Cn2 scorpion toxin on neuroblastoma cells. J. Proteom. 111, 212–223. doi: 10.1016/j.jprot.2014.04.035
Garcia-Gomez, B. I., Coronas, F. I., Restano-Cassulini, R., Rodriguez, R. R., Possani, L. D. (2011). Biochemical and molecular characterization of the venom from the Cuban scorpion Rhopalurus junceus. Toxicon 58 (1), 18–27. doi: 10.1016/j.toxicon.2011.04.011
Goversen, B., Jonsson, M., van den Heuvel, N., Rijken, R., Vos, M., van Veen, T., et al. (2019). The influence of hERG1a and hERG1b isoforms on drug safety screening in iPSC-CMs. Prog. Biophys. Mol. Biol. S0079-6107 (18), 30263–30263. doi: 10.1016/j.pbiomolbio.2019.02.003
Guan, G., Zhao, M., Xu, X., Boczek, T., Mao, X., Li, Z., et al. (2018). Abnormal changes in voltage-gated sodium channels subtypes NaV1.1, NaV1.2, NaV1.3, NaV1.6 and CaM/CaMKII pathway in low-grade astrocytoma. Neurosci. Lett. 674, 148–155. doi: 10.1016/j.neulet.2018.03.047
Guo, G., Cui, Y., Chen, H., Zhang, L., Zhao, M., Chen, B., et al. (2016). Analgesic-antitumor peptide inhibits the migration and invasion of HepG2 cells by an upregulated VGSC beta1 subunit. Tumour Biol. 37 (3), 3033–3041. doi: 10.1007/s13277-015-4067-x
Huang, X., Jan, L. (2014). Targeting potassium channels in cancer. J. Cell Biol. 206 (2), 151–162. doi: 10.1083/jcb.201404136
Jager, H., Dreker, T., Buck, A., Giehl, K., Gress, T., Grissmer, S. (2004). Blockage of intermediate-conductance Ca2+-activated K+ channels inhibit human pancreatic cancer cell growth in vitro. Mol. Pharmacol. 65 (3), 630–638. doi: 10.1124/mol.65.3.630
Jang, S. H., Choi, S. Y., Ryu, P. D., Lee, S. Y. (2011). Anti-proliferative effect of Kv1.3 blockers in A549 human lung adenocarcinoma in vitro and in vivo. Eur. J. Pharmacol. 651 (1-3), 26–32. doi: 10.1016/j.ejphar.2010.10.066
Jehle, J., Schweizer, P., Katus, H., Thomas, D. (2011). Novel roles for hERG K(+) channels in cell proliferation and apoptosis. Cell Death Dis. 18 (2), e193. doi: 10.1038/cddis.2011.77
Jiang, S., Zhu, L., Yang, J., Hu, L., Gu, J., Xing, X., et al. (2017). Integrated expression profiling of potassium channels identifys KCNN4 as a prognostic biomarker of pancreatic cancer. Biochem. Biophys. Res. Commun. 494 (1-2), 113–119. doi: 10.1016/j.bbrc.2017.10.072
Jimenez-Vargas, J. M., Restano-Cassulini, R., Possani, L. D. (2012). Toxin modulators and blockers of hERG K(+) channels. Toxicon 60 (4), 492–501. doi: 10.1016/j.toxicon.2012.03.024
Kampo, S., Ahmmed, B., Zhou, T., Owusu, L., Anabah, T. W., Doudou, N. R., et al. (2019). Scorpion Venom Analgesic Peptide, BmK AGAP Inhibits Stemness, and Epithelial-Mesenchymal Transition by Down-Regulating PTX3 in Breast Cancer. Front. Oncol. 9, 21. doi: 10.3389/fonc.2019.00021
Kischel, P., Girault, A., Rodat-Despoix, L., Chamlali, M., Radoslavova, S., Abou Daya, H., et al. (2019). Ion Channels: New Actors Playing in Chemotherapeutic Resistance. Cancers (Basel) 11 (3), E376. doi: 10.3390/cancers11030376
Klumpp, L., Sezgin, E. C., Eckert, F., Huber, S. M. (2016). Ion Channels in Brain Metastasis. Int. J. Mol. Sci. 17 (9), 1513. doi: 10.3390/ijms17091513
Kuang, Q., Purhonen, P., Hebert, H. (2015). Structure of potassium channels. Cell Mol. Life Sci. 72 (19), 3677–3693. doi: 10.1007/s00018-015-1948-5
Kuzmenkov, A., Grishin, E., Vassilevski, A. (2015). Diversity of Potassium Channel Ligands: Focus on Scorpion Toxins. Biochem. (Mosc) 80 (13), 1764–1799. doi: 10.1134/S0006297915130118
Lagarto, A., Bueno, V., Perez, M. R., Rodriguez, C. C., Guevara, I., Valdes, O., et al. (2020). Safety evaluation of the venom from scorpion Rhopalurus junceus: Assessment of oral short term, subchronic toxicity and teratogenic effect. Toxicon 176, 59–66. doi: 10.1016/j.toxicon.2020.02.002
Lastraioli, E., Iorio, J., Arcangeli, A. (2015). Ion channel expression as promising cancer biomarker. Biochim. Biophys. Acta 1848 (10), 2685–2702. doi: 10.1016/j.bbamem.2014.12.016
Leanza, L., O'Reilly, P., Doyle, A., Venturini, E., Zoratti, M., Szegezdi, E., et al. (2014). Correlation between potassium channel expression and sensitivity to drug-induced cell death in tumor cell lines. Curr. Pharm. Des. 20 (2), 189–200. doi: 10.2174/13816128113199990032
Leanza, L., Romio, M., Becker, K., Azzolini, M., Trentin, L., Managò, A., et al. (2017). Direct Pharmacological Targeting of a Mitochondrial Ion Channel Selectively Kills Tumor Cells In Vivo. Cancer Cell 31 (4), 516–531. doi: 10.1016/j.ccell.2017.03.003
Li, N., Liu, L., Li, G., Xia, M., Du, C., Zheng, Z. (2018). The role of BKCa in endometrial cancer HEC-1-B cell proliferation and migration. Gene 655, 42–47. doi: 10.1016/j.gene.2018.02.055
Litan, A., Langhans, S. A. (2015). Cancer as a channelopathy: ion channels and pumps in tumor development and progression. Front. Cell Neurosci. 9, 86. doi: 10.3389/fncel.2015.00086
Liu, L., Chen, Y., Zhang, Q., Li, C. (2019). Silencing of KCNA1 suppresses the cervical cancer development via mitochondria damage. Channels (Austin) 13 (1), 321–330. doi: 10.1080/19336950.2019.1648627
Lopez-Charcas, O., Espinosa, A., Alfaro, A., Herrera-Carrillo, Z., Ramirez-Cordero, B., Cortes-Reynosa, P., et al. (2018). The invasiveness of human cervical cancer associated to the function of NaV1.6 channels is mediated by MMP-2 activity. Sci. Rep. 8 (1), 12995. doi: 10.1038/s41598-018-31364-y
Mahadevappa, R., Ma, R., Kwok, H. F. (2017). Venom Peptides: Improving Specificity in Cancer Therapy. Trends Cancer 3 (9), 611–614. doi: 10.1016/j.trecan.2017.07.004
Mao, W., Zhang, J., Körner, H., Jiang, Y., Ying, S. (2019). The Emerging Role of Voltage-Gated Sodium Channels in Tumor Biology. Front. Oncol. 9, 124. doi: 10.3389/fonc.2019.00124
Martínez, R., Stühmer, W., Martin, S., Schell, J., Reichmann, A., Rohde, V., et al. (2015). Analysis of the expression of Kv10.1 potassium channel in patients with brain metastases and glioblastoma multiforme: impact on survival. BMC Cancer 15, 839. doi: 10.1186/s12885-015-1848-y
Moreels, L., Peigneur, S., Yamaguchi, Y., Vriens, K., Waelkens, E., Zhu, S., et al. (2017). Expanding the pharmacological profile of kappa-hefutoxin 1 and analogues: A focus on the inhibitory effect on the oncogenic channel Kv10.1. Peptides 98, 43–50. doi: 10.1016/j.peptides.2016.08.008
Nastainczyk, W., Meves, H., Watt, D. D. (2002). A short-chain peptide toxin isolated from Centruroides sculpturatus scorpion venom inhibits ether-a-go-go-related gene K(+) channels. Toxicon 40 (7), 1053–1058. doi: 10.1016/S0041-0101(02)00100-9
Nelson, M., Yang, M., Millican-Slater, R., Brackenbury, W. (2015a). Nav1.5 regulates breast tumor growth and metastatic dissemination in vivo. Oncotarget 6 (32), 32914–32929. doi: 10.18632/oncotarget.5441
Nelson, M., Yang, M., Dowle, A., Thomas, J., Brackenbury, W. (2015b). The sodium channel-blocking antiepileptic drug phenytoin inhibits breast tumour growth and metastasis. Mol. Cancer 14, 13. doi: 10.1186/s12943-014-0277-x
Noda, S., Chikazawa, K., Suzuki, Y., Imaizumi, Y., Yamamura, H. (2020). Involvement of the gamma1 subunit of the large-conductance Ca(2+)-activated K(+) channel in the proliferation of human somatostatinoma cells. Biochem. Biophys. Res. Commun. 525 (4), 1032–1037. doi: 10.1016/j.bbrc.2020.02.176
O'Malley, H. A., Isom, L. L. (2015). Sodium channel beta subunits: emerging targets in channelopathies. Annu. Rev. Physiol. 77, 481–504. doi: 10.1146/annurev-physiol-021014-071846
Oeggerli, M., Tian, Y., Ruiz, C., Wijker, B., Sauter, G., Obermann, E., et al. (2012). Role of KCNMA1 in breast cancer. PloS One 7 (8), e41664. doi: 10.1371/journal.pone.0041664
Ouadid-Ahidouch, H., Ahidouch, A., Pardo, L. (2016). Kv10.1 K(+) channel: from physiology to cancer. Pflugers Arch. 468 (5), 751–762. doi: 10.1007/s00424-015-1784-3
Pérez-Verdaguer, M., Capera, J., Serrano-Novillo, C., Estadella, I., Sastre, D., Felipe, A. (2016). The voltage-gated potassium channel Kv1.3 is a promising multitherapeutic target against human pathologies. Expert Opin. Ther. Targets. 20 (5), 577–591. doi: 10.1517/14728222.2016.1112792
Pardo, L., Stühmer, W. (2014). The roles of K(+) channels in cancer. Nat. Rev. Cancer 14 (1), 39–48. doi: 10.1038/nrc3635
Patel, F., Brackenbury, W. J. (2015). Dual roles of voltage-gated sodium channels in development and cancer. Int. J. Dev. Biol. 59 (7-9), 357–366. doi: 10.1387/ijdb.150171wb
Pedarzani, P., D'Hoedt, D., Doorty, K. B., Wadsworth, J. D., Joseph, J. S., Jeyaseelan, K., et al. (2002). Tamapin, a venom peptide from the Indian red scorpion (Mesobuthus tamulus) that targets small conductance Ca2+-activated K+ channels and afterhyperpolarization currents in central neurons. J. Biol. Chem. 277 (48), 46101–46109. doi: 10.1074/jbc.M206465200
Pedraza Escalona, M., Possani, L. D. (2013). Scorpion beta-toxins and voltage-gated sodium channels: interactions and effects. Front. Biosci. (Landmark Ed.) 18, 572–587. doi: 10.2741/4121
Prevarskaya, N., Skryma, R., Shuba, Y. (2018). Ion Channels in Cancer: Are Cancer Hallmarks Oncochannelopathies? Physiol. Rev. 98 (2), 559–621. doi: 10.1152/physrev.00044.2016
Quintero-Hernández, V., Jiménez-Vargas, J., Gurrola, G., Valdivia, H., Possani, L. (2013). Scorpion venom components that affect ion-channels function. Toxicon 76, 328–342. doi: 10.1016/j.toxicon.2013.07.012
Ramirez-Cordero, B., Toledano, Y., Cano-Sanchez, P., Hernandez-Lopez, R., Flores-Solis, D., Saucedo-Yanez, A. L., et al. (2014). Cytotoxicity of recombinant tamapin and related toxin-like peptides on model cell lines. Chem. Res. Toxicol. 27 (6), 960–967. doi: 10.1021/tx4004193
Rao, V., Perez-Neut, M., Kaja, S., Gentile, S. (2015). Voltage-gated ion channels in cancer cell proliferation. Cancers (Basel) 7 (2), 849–875. doi: 10.3390/cancers7020813
Raposo, C. (2017). Scorpion and spider venoms in cancer treatment: state of the art, challenges, and perspectives. J. Clin. Transl. Res. 3 (2), 233–249. doi: 10.18053/jctres.03.201702.002
Rodrigues, T., Estevez, G. N. N., Tersariol, I. (2019). Na(+)/Ca(2+) exchangers: Unexploited opportunities for cancer therapy? Biochem. Pharmacol. 163, 357–361. doi: 10.1016/j.bcp.2019.02.032
Roger, S., Gillet, L., Le Guennec, J. Y., Besson, P. (2015). Voltage-gated sodium channels and cancer: is excitability their primary role? Front. Pharmacol. 6, 152. doi: 10.3389/fphar.2015.00152
Salem, M. L., Shoukry, N. M., Teleb, W. K., Abdel-Daim, M. M., Abdel-Rahman, M. A. (2016). In vitro and in vivo antitumor effects of the Egyptian scorpion Androctonus amoreuxi venom in an Ehrlich ascites tumor model. Springerplus 5, 570. doi: 10.1186/s40064-016-2269-3
Schickling, B. M., England, S. K., Aykin-Burns, N., Norian, L. A., Leslie, K. K., Frieden-Korovkina, V. P. (2015). BKCa channel inhibitor modulates the tumorigenic ability of hormone-independent breast cancer cells via the Wnt pathway. Oncol. Rep. 33 (2), 533–538. doi: 10.3892/or.2014.3617
Serrano-Albarrás, A., Estadella, I., Cirera-Rocosa, S., Navarro-Pérez, M., Felipe, A. (2018). Kv1.3: a multifunctional channel with many pathological implications. Expert Opin. Ther. Targets. 22 (2), 101–105. doi: 10.1080/14728222.2017.1420170
Serrano-Novillo, C., Capera, J., Colomer-Molera, M., Condom, E., Ferreres, J., Felipe, A. (2019). Implication of Voltage-Gated Potassium Channels in Neoplastic Cell Proliferation. Cancers (Basel) 11 (3), E287. doi: 10.3390/cancers11030287
Shan, B., Dong, M., Tang, H., Wang, N., Zhang, J., Yan, C., et al. (2014). Voltage-gated sodium channels were differentially expressed in human normal prostate, benign prostatic hyperplasia and prostate cancer cells. Oncol. Lett. 8 (1), 345–350. doi: 10.3892/ol.2014.2110
Staudacher, I., Jehle, J., Staudacher, K., Pledl, H., Lemke, D., Schweizer, P., et al. (2014). HERG K+ channel-dependent apoptosis and cell cycle arrest in human glioblastoma cells. PloS One 9 (2), e88164. doi: 10.1371/journal.pone.0088164
Teisseyre, A., Gasiorowska, J., Michalak, K. (2015). Voltage-Gated Potassium Channels Kv1.3–Potentially New Molecular Target in Cancer Diagnostics and Therapy. Adv. Clin. Exp. Med. 24 (3), 517–524. doi: 10.17219/acem/22339
Teisseyre, A., Palko-Labuz, A., Sroda-Pomianek, K., Michalak, K. (2019). Voltage-Gated Potassium Channel Kv1.3 as a Target in Therapy of Cancer. Front. Oncol. 9, 933. doi: 10.3389/fonc.2019.00933
Tian, C., Zhu, R., Zhu, L., Qiu, T., Cao, Z., Kang, T. (2014). Potassium channels: structures, diseases, and modulators. Chem. Biol. Drug Des. 83 (1), 1–26. doi: 10.1111/cbdd.12237
Venturini, E., Leanza, L., Azzolini, M., Kadow, S., Mattarei, A., Weller, M., et al. (2017). Targeting the Potassium Channel Kv1.3 Kills Glioblastoma Cells. Neurosignals 25 (1), 26–38. doi: 10.1159/000480643
Wang, W. X., Ji, Y. H. (2005). Scorpion venom induces glioma cell apoptosis in vivo and inhibits glioma tumor growth in vitro. J. Neurooncol. 73 (1), 1–7. doi: 10.1007/s11060-004-4205-6
Wang, X., Chen, Y., Zhang, Y., Guo, S., Mo, L., An, H., et al. (2017). Eag1 Voltage-Dependent Potassium Channels: Structure, Electrophysiological Characteristics, and Function in Cancer. J. Membr. Biol. 250 (2), 123–132. doi: 10.1007/s00232-016-9944-8
Ward, M., Ellsworth, S., Nystrom, G. (2018). A global accounting of medically significant scorpions: Epidemiology, major toxins, and comparative resources in harmless counterparts. Toxicon 151, 137–155. doi: 10.1016/j.toxicon.2018.07.007
Wulff, H., Castle, N. (2010). Therapeutic potential of KCa3.1 blockers: recent advances and promising trends. Expert Rev. Clin. Pharmacol. 3 (3), 385–396. doi: 10.1586/ecp.10.11
Xia, J., Huang, N., Huang, H., Sun, L., Dong, S., Su, J., et al. (2016). Voltage-gated sodium channel Nav 1.7 promotes gastric cancer progression through MACC1-mediated upregulation of NHE1. Int. J. Cancer 139 (11), 2553–2369. doi: 10.1002/ijc.30381
Yamaci, R., Fraser, S., Battaloglu, E., Kaya, H., Erguler, K., Foster, C., et al. (2017). Neonatal Nav1.5 protein expression in normal adult human tissues and breast cancer. Pathol. Res. Pract. 213 (8), 900–907. doi: 10.1016/j.prp.2017.06.003
Yang, M., Brackenbury, W. (2013). Membrane potential and cancer progression. Front. Physiol. 4, 185. doi: 10.3389/fphys.2013.00185
Keywords: cancer, ion channels, scorpion venom, toxins, voltage-dependent
Citation: Díaz-García A and Varela D (2020) Voltage-Gated K+/Na+ Channels and Scorpion Venom Toxins in Cancer. Front. Pharmacol. 11:913. doi: 10.3389/fphar.2020.00913
Received: 02 January 2020; Accepted: 04 June 2020;
Published: 18 June 2020.
Edited by:
Sébastien Roger, Université de Tours, FranceReviewed by:
Christian Legros, Université d'Angers, FranceCopyright © 2020 Díaz-García and Varela. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Alexis Díaz-García, YWxleGlzZGc3NkBnbWFpbC5jb20=; Diego Varela, ZHZhcmVsYUB1Y2hpbGUuY2w=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.