 Boshra Tinoush
Boshra Tinoush Iman Shirdel
Iman Shirdel Michael Wink
Michael Wink
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Pharmacol. , 19 June 2020
Sec. Ethnopharmacology
Volume 11 - 2020 | https://doi.org/10.3389/fphar.2020.00832
This article is part of the Research Topic Natural Products as a Tool to Design New anti-MDR Lead Molecules View all 12 articles
Multidrug resistance (MDR) is one of the main impediments in the treatment of cancers. MDR cancer cells are resistant to multiple anticancer drugs. One of the major mechanisms of MDR is the efflux of anticancer drugs by ABC transporters. Increased activity and overexpression of these transporters are important causes of drug efflux and, therefore, resistance to cancer chemotherapy. Overcoming MDR is a fundamental prerequisite for developing an efficient treatment of cancer. To date, various types of ABC transporter inhibitors have been employed but no effective anticancer drug is available at present, which can completely overcome MDR. Phytochemicals can reverse MDR in cancer cells via affecting the expression or activity of ABC transporters, and also through exerting synergistic interactions with anticancer drugs by addressing additional molecular targets. We have listed numerous phytochemicals which can affect the expression and activity of ABC transporters in MDR cancer cell lines. Phytochemicals in the groups of flavonoids, alkaloids, terpenes, carotenoids, stilbenoids, lignans, polyketides, and curcuminoids have been examined for MDR-reversing activity. The use of MDR-reversing phytochemicals with low toxicity to human in combination with effective anticancer agents may result in successful treatment of chemotherapy-resistant cancer. In this review, we summarize and discuss published evidence for natural products with MDR modulation abilities.
Besides surgery and radiation, chemotherapy is one of the standard treatments of cancer. Drugs used in chemotherapy usually disturb cell division by inhibition of microtubule formation or disassembly (vinca alkaloids, paclitaxel), DNA topoisomerase (camptothecin and derivatives) or they intercalate or alkylate DNA (doxorubicin, cisplatin) (Wink, 2007; Wink et al., 2012). Furthermore, chemotherapy often causes extreme side effects as these drugs also affect the division of normal cells or they cause mutations, which can lead to secondary cancers. This additionally leads to restriction of the therapeutic applications; both dosage and application interval must be kept limited. When chemotherapeutic agents are used, it is often a matter of time before the cancer cells develop resistance against them. One of the major resistance mechanisms is the overexpression of ABC transporters, which can pump out the chemotherapeutic from cancer cells. Because these ABC transporters have a wide substrate spectrum, they not only confer resistance to a single drug but to several others, therefore, the term Multidrug Resistance (MDR). Since multiple drug resistance is a major issue in tumor therapy, new strategies are necessary to overcome this obstacle. One strategy involves the combination of anti-cancer drugs with modulators of ABC transporters. Our review presents a summary about various modulating effects of phytochemicals.
One of the major difficulties in suppressing growth and survival of cancer cells is multidrug resistance. MDR is the resistance of cancer cells to various types of anticancer drugs which may have an intrinsic or acquired origin. Acquired resistance is induced after the administration of chemotherapy, whereas, intrinsic resistance already exists prior to drug application in cancer cells (Wang et al., 2019). Several mechanisms in cancer cells can lead to MDR (Coley, 2008). These include changes in target enzymes, such as DNA topoisomerases (Brown et al., 1995), alteration in microtubule-associated proteins (Zhang et al., 1998), mutations or changes in tubulin (Kamath et al., 2005; Wang and Cabral, 2005), alteration in microtubules (Kavallaris et al., 2001), mitotic arrest (Kamath et al., 2005), mutated protein p53 (O'Connor et al., 1997), disruption in DNA repair due to the damaging effect of an anticancer drug (Bernstein et al., 2002) and the impairment of apoptosis or genes involved in apoptosis and necrosis (Tanaka et al., 2000; Simstein et al., 2003).
A widespread mechanism of MDR is drug efflux via transmembrane transporters known as ATP-binding cassette transporters (ABC transporter). Overexpression of these transporters is the most important cause of drug resistance in many cancer cells. The family of ABC transporter proteins has 48 members in humans and far more in nature (Chen et al., 2016). The most well-known and widely-studied ABC transporters include P-Glycoprotein (P-gp), multidrug resistance protein 1 (MRP1), and breast cancer resistance protein (BCRP). These transporters are expressed in healthy cells of various mammalian tissues having physiological tasks for translocating small molecules. They are found especially in intestinal epithelial cells, endothelial cells of blood capillaries and epithelia of renal proximal tubules being involved in the excretion and clearance of endogenous and exogenous cytotoxic substances (Borst et al., 2000; Durmus et al., 2015; Chen et al., 2016). ABC transporters evolved in nature millions of years ago to eliminate toxic phytochemicals that herbivores would obtain from their plant diet (Wink, 2007). Anticancer drugs also can be substrates of these transporters and if being considered as foreign they get exported to the extracellular space by cells expressing ABC transporters. Cancer cells which are not resistant to anticancer agents yet can develop the ability by overexpressing these transporters for saving themselves from the substances being toxic to them. This leads to an increased efflux, leading to low intracellular drug concentrations, insufficient to kill a cancer cell. Once this overexpression has occurred, the efflux also affects other chemotherapeutics and thus, makes the cancer cell resistant to chemotherapy (Coley, 2008; Durmus et al., 2015).
P-gp is a 170 kDa protein which is encoded by the MDR1 gene. This transporter is found in normal cells of various tissues including the brain, liver, kidney, gastrointestinal tract and pancreas. P-gp transports anticancer drugs such as paclitaxel, doxorubicin, daunorubicin, epirubicin, mitoxantrone, vincristine, and vinblastine against the concentration gradient using energy derived from hydrolysis of ATP (Chen et al., 2016). Chemotherapeutic agents can stimulate P-gp expression in cancer cells and thereby cause resistance to chemotherapy. Chemotherapy has been reported to increase the proportion of P-gp-expressing tumors by approximately 1.8-fold in breast cancer. Moreover, in patients with activated P-gp transporter in their tumors, the risk of failure of chemotherapy is 3 times higher than in patients who do not express P-gp transporter (Trock et al., 1997).
Another class of membrane transporters which causes MDR is MRPs. Nine members of this class have been identified so far (König et al., 2005; Coley, 2008). MRPs are found in normal cells of some mammalian tissues and expel drugs as a complex with glutathione, glucuronate, or sulfate (Borst et al., 2000; Coley, 2008). Among the MRP transporters, MRP1 (ABCC1) is the most important and most studied one regarding MDR. The MRP1 protein has a molecular weight of 190 kDa. Similar to P-gp, MRP1 expression has been reported to be considerably higher expressed in cancer cells after chemotherapy than before chemotherapy (Trock et al., 1997). Therefore, MRP1 enhances resistance to chemotherapy and to anticancer drugs such as doxorubicin, daunorubicin, epirubicin, vincristine, and vinblastine (Coley, 2008).
Breast cancer resistance protein, also called mitoxantrone transporter (MXR1), has a molecular weight of 72 kDa. BCRP is extensively expressed in MCF-7 breast cancer cells (Doyle et al., 1998). This protein is also expressed in other tissues including the liver, kidney, and intestine (Chen et al., 2016). The anticancer drugs doxorubicin, daunorubicin, epirubicin and mitoxantrone have been described as substrates of BCRP transporter (Coley, 2008). Thus, cancer cells overexpressing BCRP transporter become resistant to these drugs.
One of the essential requirements for developing better anti-cancer therapies is overcoming multidrug resistance. Much research has been carried out on cancer treatment and development of anticancer drugs in recent years but MDR to cytostatics is still a great impediment. Although our knowledge about the mechanisms of multidrug resistance has increased, there is no effective drug which can completely overcome or reverse resistance at non-toxic concentrations. Since ABC transporters play a fundamental role in resistance to chemotherapy, the ability to inhibit them in a combination with conventional treatments will greatly help to treat cancer (Chen et al., 2016).
Until now, different types of ABC transporter inhibitors have been examined. The use of the first generation of these compounds, including verapamil and cyclosporine A, in combination with anticancer drugs had poor clinical success and toxic effects (Daenen et al., 2004). Second generation of MDR modulators included dexverapamil, valspodar, and dexniguldipine. Even though less toxic and with a higher therapeutic index than the first generation, this group of modulators is not well suited for a therapy either, both because of its interactions with other drugs and ABC transporters, as well as due to the inhibition of enzymes like CYP3A (Wandel et al., 1999; Syed and Coumar, 2016). The third-generation ABC transporter modulators do not have the disadvantages of the first and second generation. They are potent and non-competitive inhibitors of P-gp, and also less toxic. Tariquidar (XR9576) and zosuquidar are members of the third generation of MDR modulators but unfortunately they were not efficative in clinical trials (Cripe et al., 2010; Kelly et al., 2011).
Alkaloids (Figure 1) are the most widely studied group of secondary metabolites in terms of MDR, not only because of their quantity but also because of their great diversity (Wink, 2007; Wink et al., 2012). As alkaloids have a wide distribution among angiosperms (Wink, 2020) and represent a diversity of structures, they differ in pharmacological and toxicological properties. Alkaloids contain heterocyclic nitrogen, which mostly has its origin in amino acids (Cseke et al., 2006). Alkaloids are subdivided into many subcategories of special functional groups, similarities of skeleton or biosynthetic pathways.
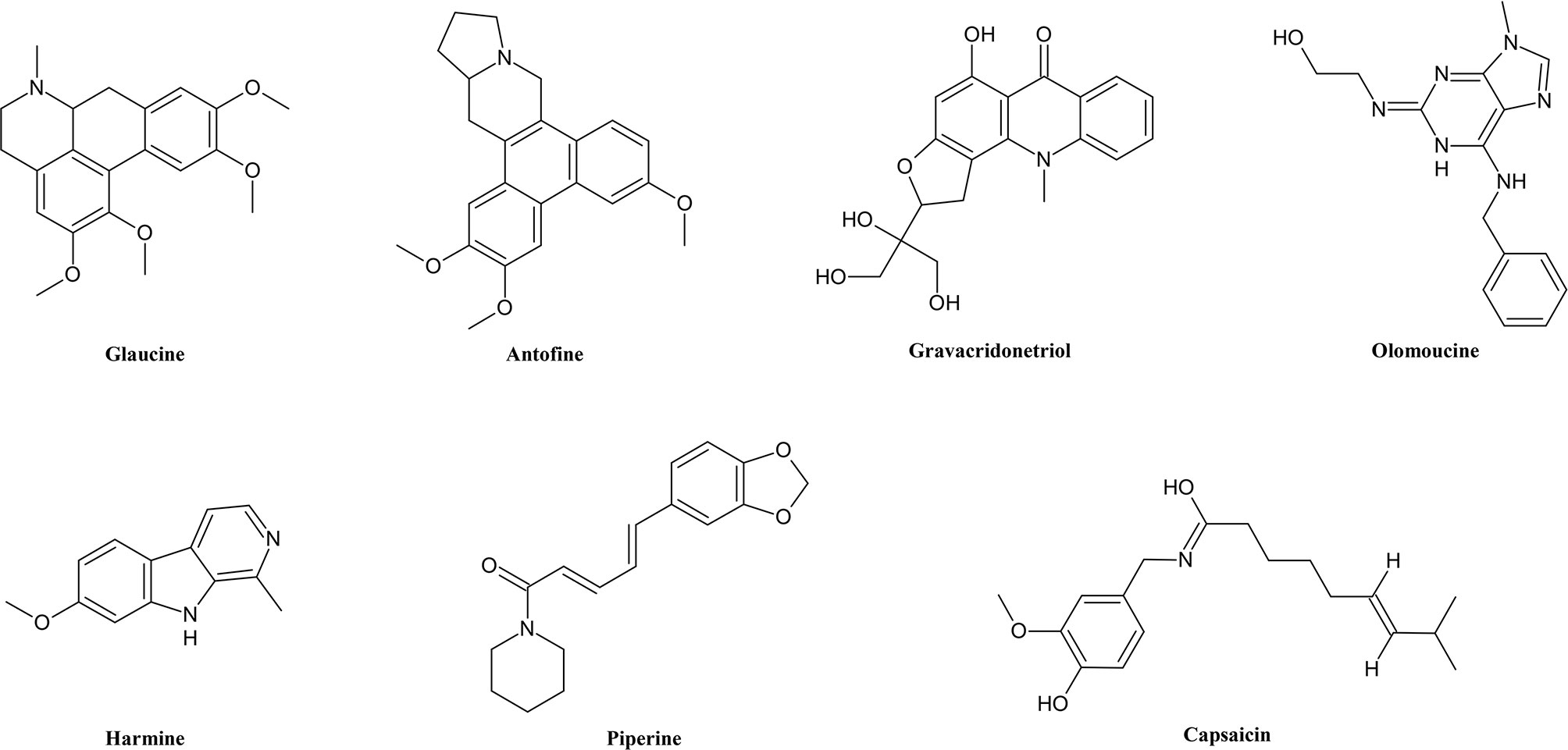
Figure 1 Chemical structures of some selected alkaloids with MDR reversal effects.
Quinoline and isoquinoline alkaloids for example have a benzopyridine ring differing in the position of their nitrogen. Quinazoline alkaloids have a similar aromatic structure but with two nitrogen atoms instead of one. Each of these structures has found several uses depending on molecular structure. There are many examples for quinoline alkaloids such as mefloquine as antimalarial agent, fluoroquinolone antibiotics and topotecan as anticancer drug, just to name a few (Collin and Höke, 2000; Tiwary et al., 2015). The latter two work as inhibitors of different DNA topoisomerases (Lemmer and Brune, 2004).
Quinolizidines also are cyclic nitrogen-containing compounds but unlike the previously mentioned subgroups they are not aromatic. A natural representative is sparteine, which is used as an antiarrhythmic agent blocking sodium channels (Ruenitz and Mokler, 1977; Körper et al., 1998; Gawali et al., 2017).
Among other groups of alkaloids, indole, monoterpene indole and β-carboline alkaloids show many pharmacological activities (Gilbert, 2001). A typical basic structure of β-carboline alkaloids consists of benzene fused with a five-membered pyrrole and is consequently similarly structured to some endogenous hormones and neurotransmitters such as serotonin and melatonin. In addition to benzene, the pyrrole ring of β-carboline is fused to a pyridine, another six-membered nitrogen-containing ring. This structure by itself is an inverse agonist of GABA-receptors, which involves psychological influence on humans (Aktories et al., 2017). Substrates among the indoles target many receptors, for example PDE-receptors e.g. by tadalafil, 5-HT receptors e.g. by naratriptane and HMG-CoA reductases e.g. by fluvastatin (Wink, 2000; de Sa et al., 2009). Several indole alkaloids have stimulant and hallucinogenic properties (Wink, 2000; Wink and van Wyk, 2008).
As the nitrogen of steroidal alkaloids does not originate from amino acids they belong to pseudoalkaloids. A member of steroidal alkaloids is the teratogenic cyclopamine, which can cause cyclopean eyes in vertebrates (Roberts and Wink, 1998; Incardona et al., 1998).
Piperidine and diketopiperazine are six-membered non-aromatic moieties in alkaloids whereby diketopiperazine is a cyclic dipeptide having two oppositely located nitrogen. Pyrazine has the same position of nitrogen, though aromatic. Here we also can find medicinal use of active ingredients such as the oxytocin antagonist retosiban and plinabulin which is still in clinical trial against multiple drug resistant non-small cell lung cancer (Borthwick and Liddle, 2011; Mohanlal et al., 2019).
Tropane alkaloids are widespread and their plants one of the oldest medicines to use because of spasmolytic, mydriatic and hallucinogenic properties (Wink and van Wyk, 2008; van Wyk and Wink, 2017). They contain a special bicyclic moiety which is made of a seven-membered ring and a nitrogen atom which is linked to its C-1 and C-5 and forms the second ring (Osman et al., 2013). Due to their spasmolytic effect nowadays we use tropane alkaloids such as scopolamine or atropine for digestive tract spastic conditions and for ophthalmological purposes (Kukula-Koch and Widelski, 2017).
The most popular alkaloid caffeine belongs to purine alkaloids, which are consumed by many people on all continents. Theobromine, theophylline and caffeine are common members found as main ingredients in chocolate, mate, cola, green and black tea or coffee (Baumann and Frischknecht, 1988). By inhibiting adenosine receptors and cAMP phosphodiesterase they can mediate a stimulant effect (van Wyk and Wink, 2017).
Flavonoids (Figure 2) are another complex but also often colored group of secondary metabolites. Unlike alkaloids, we can make a general statement about their common origin and basic skeleton. Flavonoids can be classified as polyphenols which share a common biosynthesis. They contain aromatic rings with phenolic hydroxyl groups. These phenolic hydroxyl groups can dissociate under physiological condition and form negatively charged phenolate ions. Because of these properties flavonoids and polyphenols can interact with proteins forming multiple hydrogen and ionic bonds (Wink, 2015). Flavonoids are widely distributed in plants and are responsible, inter alia, for their pollinator attracting colors, ultraviolet light protection, antioxidant and antimicrobial functions, and mediating symbiosis with bacteria. Including the main subgroups flavones, isoflavones, flavonols, flavanones, anthocyanins, chalcones and catechins, they derive from flavan, a benzopyran structure with a phenyl ring in position 2. Flavonoids with a phenyl ring in position 3 and 4 are called iso- and neoflavonoids, respectively. The variety of flavonoids comes from many functional groups and different states of oxidation of the heterocycle (Koes et al., 1994; Wink and van Wyk, 2008; Hänsel and Sticher, 2009). Although there has been a lot of research on the antioxidant capacity of flavonoids, the mechanism is not fully understood yet. Many studies have reported anti-inflammatory, anti-carcinogenic, anti-mutagenic, antiviral, anti-allergic and osteogenetic potentials of flavonoids in vitro. There is evidence, that polyphenols are also important for the pharmacological activity of many medicinal plants (van Wyk and Wink, 2017). Still there is a lack of information about how the necessary bioavailability is achieved in the human body as polyphenols are polar compounds (Panche et al., 2016). Most common representatives in food are luteolin and apigenin. Isoflavones are known for their estrogenic properties (Wink, 2015).
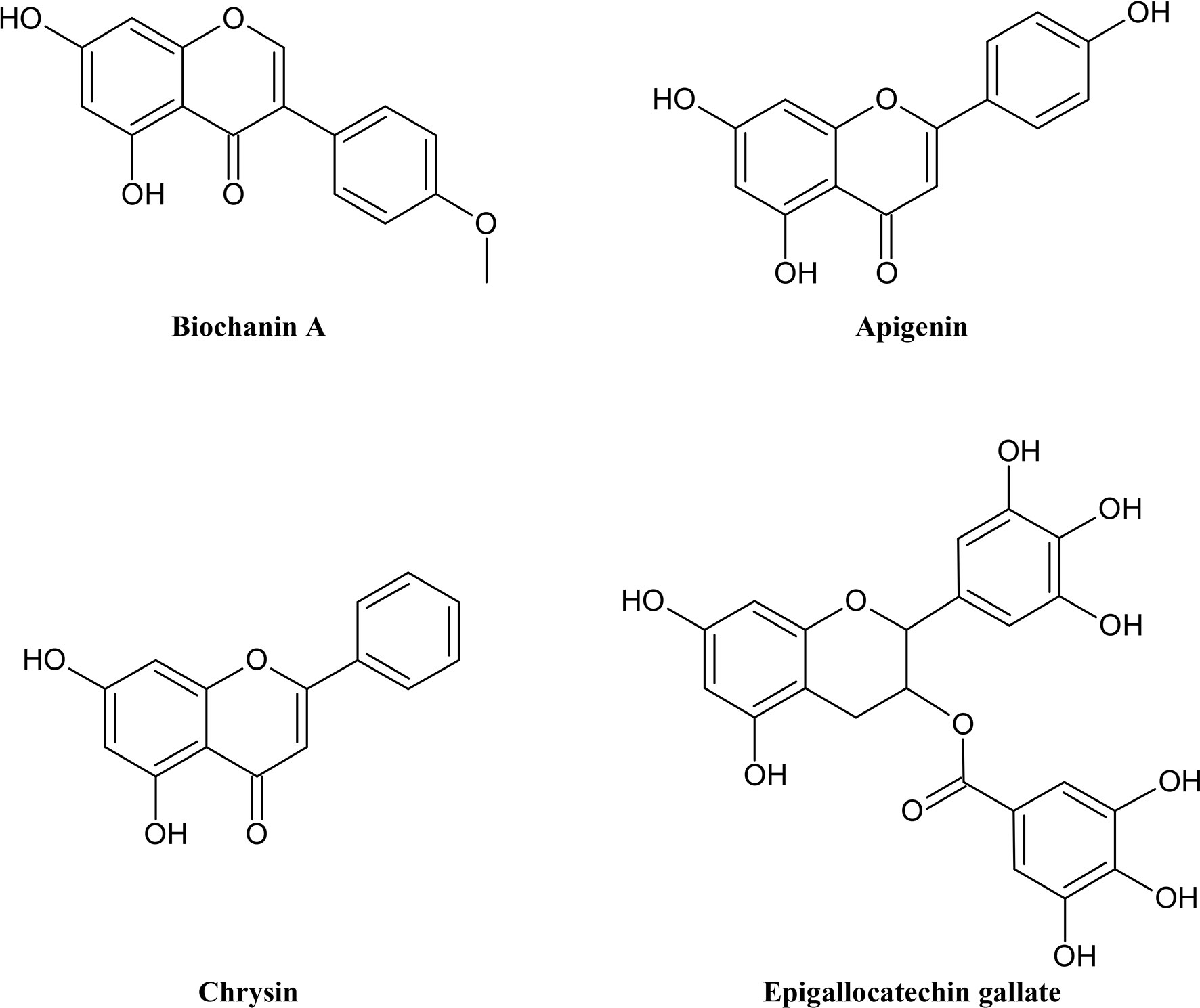
Figure 2 Chemical structures of some selected flavonoids with MDR reversal effects.
In addition to flavonoids, there are smaller but also important groups of polyphenols, for example stilbenoid with resveratrol as its main member known for its potential as anti-cancer (Huang et al., 2014), antioxidant and anti-aging agent (Alamolhodaei et al., 2017). Curcuminoids (Figure 3) have been widely studied and have been found to have many functions such as antioxidative, anti-cancer, anti-microbial and anti-inflammatory effects in humans. They can interact with many targets (Fantini et al., 2015).
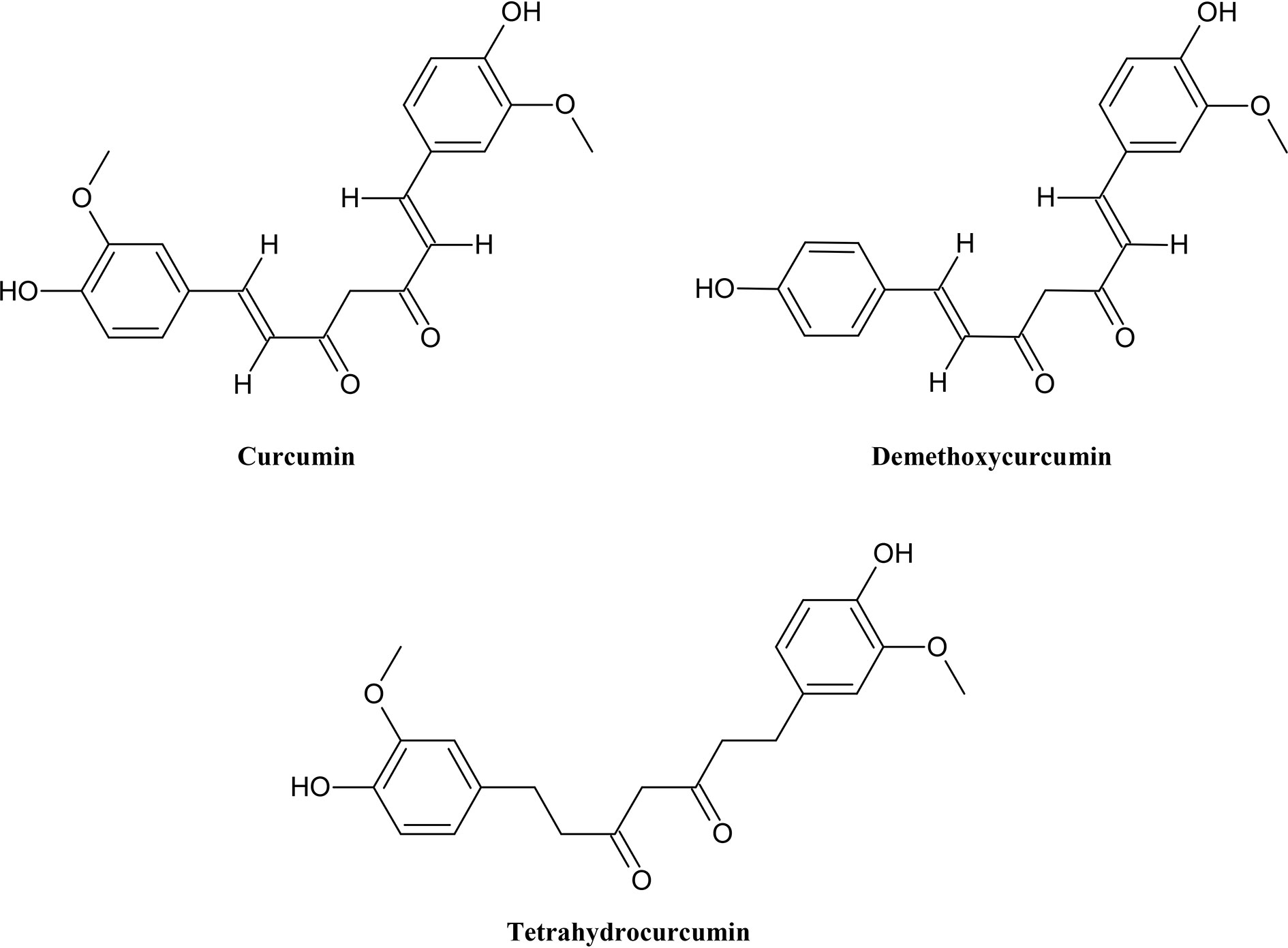
Figure 3 Chemical structures of some selected curcuminoids with MDR reversal effects.
Terpenes (Figure 4) are widely distributed in plants, fungi and animals. They are composed of different numbers of isoprene units, forming mono- (C10), di- (C20), sesqui- (C15), tetra- (C40) and triterpenes (C30). Many mono- and sequiterpenes are volatile and aromatic and typical ingredient of essential oils. These compounds are often lipophilic and can thus modulate the fluidity and permeability of biomembranes in animals and microbes. Many plants with essential oil have been used in traditional medicine for treatment of microbial infections and inflammation (Wink, 2015; van Wyk and Wink, 2017).
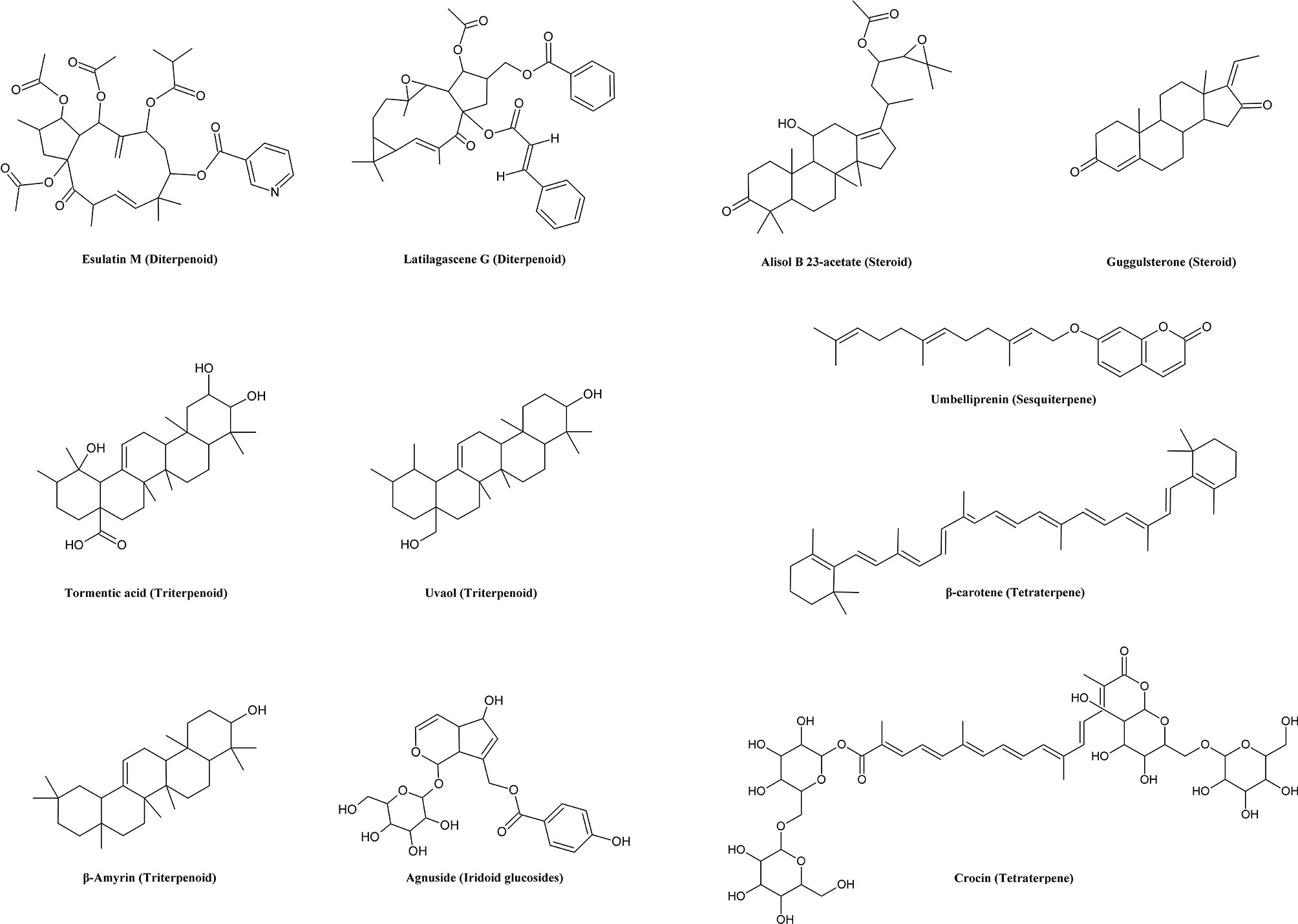
Figure 4 Chemical structures of some selected terpenes with MDR reversal effects.
Known for their skin permeation enhancing ability, terpenes have been used as moieties of synthetic structures for topical use (Smith and Maibach, 1995). In vitro studies have shown anti-cancer, antimicrobial and antioxidant activities but for practice and use in humans this class of secondary metabolites must be further investigated (Lu et al., 2012; Cör et al., 2018). Squalene is a standard triterpene produced in plants and animals, and it is the precursor for steroid synthesis. Glycosides of steroids or triterpenes, so-called saponins have one or more polysaccharides attached saponins are amphiphilic and react as a detergent. They generally form stable foams and complex cholesterol in biomembranes. As a consequence, saponins can lyse biomembranes (Bloch, 1983). In traditional medicine they have been used for example as expectorants and anti-infectants (Wink, 2015). As effective components of vaccine adjuvants they enhance the cellular immune response (Sun et al., 2009).
Carotenoids are tetraterpenes with many conjugated double bonds. They often exhibit yellow to purple colors and can function as anti-oxidants and precursors for vitamin A.
As the name implies polyketides contain carbonyl groups positioned between methylene groups. Still they vary in shape and volume. Although they appear with an impressive structural variety, they have their source from the same biosynthetic pathway. The most famous members among drugs may be the antibiotic erythromycin or the antifungal amphotericin B.
A considerable number of secondary metabolites, which affect ABC transporters has already been discovered (Table 1) and several of them will be discussed in the following; glaucine (an isoquinoline alkaloid) increased the efflux of substrates such as ADR and MTX in the P-gp over-expressing cell lines of MCF-7/ADR and reduced DOX resistance in Caco-2 and CEM/ADR5000 (Eid et al., 2013; Lei et al., 2013). Tetrandrine (a benzolisoquinoline alkaloid) also caused an inhibition of efflux in Caco-2 and CEM/ADR5000 cells (Sun and Wink, 2014). A 5-substituted derivative of it named PY35 was tested for the MDR reversal activity, and showed more MDR reversal than the natural compound in resistant K562/ADM and MCF-7/ADM cells (Cao et al., 2014). Hernandezine, a bisbenzyl-isoquinoline, is a potent inhibitor of P-gp in MDR19-HEK293 cells and was able to resensitize MDR19-HEK293 and KB-V-1 cells to DOX after entering the cell membrane (Hsiao et al., 2016). High MDR-reversing activities of the quinoline derivatives were linked to the presence of two aryl rings in the hydrophobic moiety, deviation of the aryl rings from a common plane, basicity of nitrogen atom in piperazine, as well as to the distance between the hydrophobic moiety and the basic nitrogen of piperazine which must be no less than 5 Å (Suzuki et al., 1997). The quinolyl group was also suggested to have a key role in the activity of quinolines, because substitution of a quinoline ring by a naphthyl ring or a phenyl ring resulted in the reduction of MDR-reversing activity of the compounds (Suzuki et al., 1997). Among indole alkaloids, antofine showed synergistic effects with PTX in A549-PA cells and overcame resistance to PTX (Kim et al., 2012). The β-carboline harmine reversed the resistance of MTX and CPT in MDA-MB-231 cells with BCRP as overexpressed transporter but it could not affect the P-gp over-expressing CEM/ADR5000 cells the same way in this study (Ma and Wink, 2010). Not only has harmine been tested in combination with DOX but also as three-drug-combination with DTN both of which showed here an increase of effect on cells and a reduction of DOX resistance in Caco-2 and CEM/ADR5000 cells (Eid et al., 2013). As an N-acylpiperidine, piperine inhibited the efflux of the tested substrate which led to an increase of its concentration in Caco-2 and CEM/ADR5000 cells (Li et al., 2018a). Tests of piperine on MCF-7/DOX and A-549/DDP cells resulted in an increase in cytotoxicity of MTX and DOX (Li et al., 2011). Gravacridonetriol, gravacridonediol and its monomethyl ether, all had an inhibitive effect on P-gp L5178/MDR1 cells, which also led to a higher cytotoxicity of DOX (Rethy et al., 2008). Substitution of methyl groups at the positions C-2 and C-4 of acridone led to the increased lipophilicity, which can enhance the binding affinity of acridone derivatives to P-gp (Mayur, 2015). Murahari et al. (2017) studied on 2,4-dimethylacridone derivatives and showed that these compounds were potential modulators of P-gp-mediated MDR. 2,4-dimethylacridones are tricyclic and hydrophobic and have methyl groups at C-2 and C-4 positions with a propyl or butyl side chain containing terminally substituted tertiary amino groups. They found that an alkyl side chain and hydroxyl substituted secondary amine are necessary for the acridones to reverse the P-gp-mediated multidrug resistance. Moreover, an alkyl side chain of length four (butyl) was found to have higher biological activity (Murahari et al., 2017). Hegde et al. (2004) reported that replacing the hydrogen atom at position C-4 by a methoxy group slightly enhanced the lipophilicity of the acridone derivatives. Furthermore, they investigated the MDR-reversing activity of N-10-substituted acridones and N-10-substituted 4-methoxyacridones relative to their corresponding unsubstituted counterparts where the C-4 positions of the acridone rings are occupied by a hydrogen atom and a methoxy group, respectively. The parent acridone and 4-methoxyacridone had the least effect in inhibiting drug efflux suggesting that N-10-substitution is necessary for an ideal activity. It was also found that 4-methoxyacridone derivatives are more efficient than their acridone derivatives' counterparts in increasing drug accumulation. Several N-10-substituted acridones and N-10-substituted-4-methoxyacridones showed MDR-reversing activity greater than the P-gp inhibitor verapamil (Hegde et al., 2004). In another study eighteen N-10-substituted-2-bromoacridones were examined for the anti-MDR activity and compared to the parent compound 2-bromo-10H-acridin-9-one. N-10-substitution was suggested to be necessary for optimal activity of 2-bromoacridones because the parent compound had the least effect in efflux inhibiting activity (Mayur et al., 2006).

Table 1 Phytochemicals modulating MDR via ABC-transporters.
Research on olomoucine, purvalanol and roscovitine revealed amongst other things the inhibitive potential of purine alkaloids on P-gp in the form of MDCKII-ABCB1 cells and human HCT-8 and HepG2 cells while olomoucine had the strongest effect of all (Cihalova et al., 2013). Several groups have run experiments with the benzazepine alkaloid capsaicin on different cell lines such as KB-C2, Caco-2 and CEM/ADR5000. The most relevant result was the inhibition of P-gp efflux in the presence of digoxin as substrate at non-toxic concentrations (Nabekura et al., 2005; Han et al., 2006; Li et al., 2018a).
Isoflavonoids and some flavonoids often act as phytoestrogens. Investigations revealed that hydroxyl groups at position C-5 and C-7 are an important property for the P-gp inhibitory effect of flavonoids, although hydrophobicity usually promotes the affinity (Sheu et al., 2010). Acacetin, genistein, kaempferol and naringenin, were examined by Imai et al. (2004) and it resulted in positive effects on K562 cells expressing BCRP. The cytotoxicity of MTX and 7-ethyl-10-hydroxycamptothecin was enhanced using phytoestrogens. In addition, genistein and naringenin enhanced the accumulation of topotecan in K562/BCRP cells. Also apigenin had strong reversal effect on BCRP-mediated MDR (Imai et al., 2004). In MCF-7 MX100 cells, which also overexpressed BCRP, apigenin had similar effect on the accumulation of the anticancer agent MTX (Zhang et al., 2004). However, naringenin, which differs from apigenin only in the saturation of C-2 and C-3, had a significant loss of potency in comparison to the latter due to the lack of a double bond (Conseil et al., 1998). Several groups experimented with the isoflavone and phytoestrogen biochanin A and its outcome on BCRP and P-gp in cell lines such as Panc-1, MCF-7 MX100, MDA435/LCC6, MCF-7/ADR, MDA435/LCCMDR1 and Caco-2. In all studies, an inhibition of drug efflux, an increase of accumulation and a potentiation of cytotoxicity were observed. A combination of biochanin A with some other flavonoids yielded additive effects (Nguyen et al., 2003; Zhang and Morris, 2003; Zhang et al., 2004; Sergent et al., 2005). Nobiletin and tangeretin prevented the P-gp mediated efflux of [3H]vincristine (Ikegawa et al., 2000). Among other flavones with methoxyls they were examined by Ohtani et al. (2007) for the uptake potential of [3H]vincristine as a P-gp substrate. The MDR-reversing effect increased with the number of methoxyl moieties but with the exception of both at C-3′ and C-5′ position on B-ring. In this case there was even a decrease of MDR-reversing potency. Chrysin is a hydroxyflavone with simple structure which has also been examined by a number of groups. The competitive inhibition of the MRP-2 pumps in Caco-2 cells and an antiproliferative effect of chrysin are certain (Gyémánt et al., 2005; Sergent et al., 2005). Moreover, a prevention of BCRP and P-gp mediated efflux took place in MCF-7 MX100 cells and L5178 cells, respectively (Zhang et al., 2004; Zhang et al., 2005; Gyémánt et al., 2005). A prenyl or geranyl group at C-6 and C-8 of chrysin, and a prenyl in both positions ensured an improvement of the binding affinity to P-gp. In addition, there was more potency for inhibition after geranylation than prenylation which could be due to its level of lipophilicity (Di Pietro et al., 2002). In contrast, glycosylation taking place at any position tested, dramatically decreased the binding affinity of flavonoids (Sheu et al., 2010). Another way to gain binding affinity toward the C-terminal nucleotide-binding domain of P-gp, were other kinds of alkylation of chrysin including methyl, benzyl, isopropyl and 3,3-dimethylallyl (Boumendjel et al., 2002).
Several experiments have been carried out for the flavan epigallocatechin gallate (EGCG), a derivative of epigallocatechin (EGC) abundant in green tea. EGCG has been tested on different cells such as liver cancer cells Bel-7404/DOX, the colon cancer cell line Caco-2, leukemia cells CEM/ADR5000, endocervical adenocarcinoma cells KB-C2 and others. An increase of drug accumulation and sensitization in these and several other cell lines was observed (Kitagawa et al., 2004; Liang et al., 2010; Eid et al., 2013; Reyes-Esparza et al., 2015). Particularly substitutions at D-ring had positive effect on EGC and derivatives. A derivative of EGC with three methoxy groups in cis-configuration at D-ring and an oxycarbonylvinyl as its connection to C-3 led to a higher potency. This structure only regulated P-gp and could not affect BCRP or MRP1 (Wong et al., 2015).
While curcumin is a promising agent for liver protection and inhibition of cancerous cell growth, other curcuminoids like demethoxycurcumin and tetrahydrocurcumin have being studied more closely. They inhibited the efflux of chemptherapeutics and consequently increased cellular accumulation of drugs (Limtrakul et al., 2007; Ampasavate et al., 2010).
Among phenylpropanoids, some lignans (Figure 5) showed an inhibitory effect on P-gp. Schisandrin A was used on Caco-2 cells resulting in an increase in apical-to-basal transport and cytotoxicity Yoo et al. (2007b). A study which examined the structure activity of lignans showed the benefit of the absence of a hydroxyl group in position C-8 as in schisandrin A and γ-schizandrin for the function as p-gp inhibitor. In addition, a higher effect was seen in R-configurated biaryl than in the S-configuration (Slaninová et al., 2009). The reversal of cytotoxicity in CEM/ADR 5000 and Caco-2 cells was moreover successful by using menthol or thymol, monoterpenes obtained from volatile oils. They could reduce the IC50 value of doxorubicin which enhances the effectiveness of the drug (Eid et al., 2013).
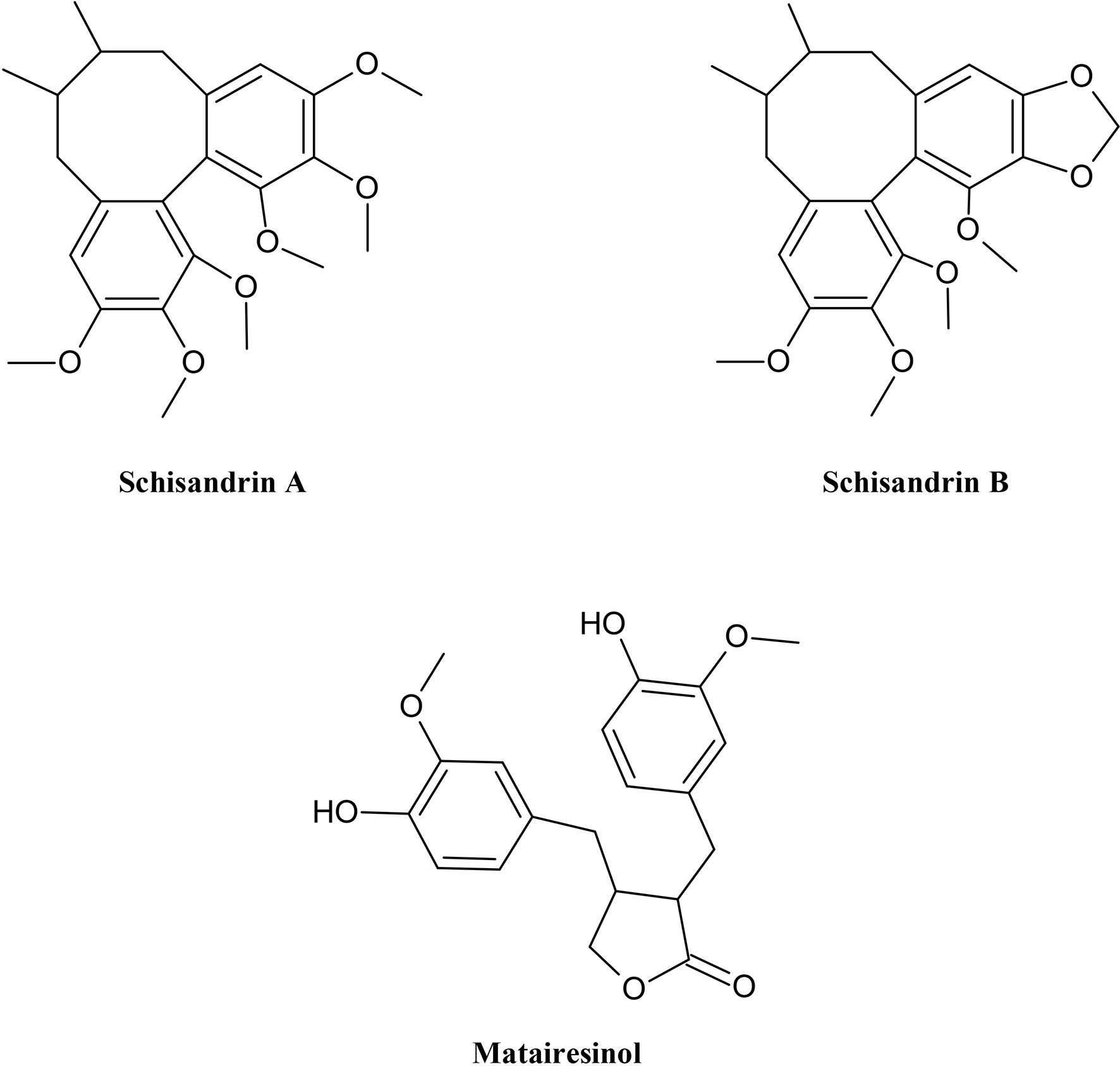
Figure 5 Chemical structures of some selected lignans with MDR reversal effects.
Reis et al. (2016) reported that jatrophane diterpenes including esulatin M, epoxywelwitschene, welwitschene, and euphowelwitschine A were more efficative in MDR cells than the positive control verapamil. In comparison to the known MDR modifier verapamil, euphoboetirane C, D, E, F, and G extracted from Euphorbia boetica showed multifold P-gp modulatory activity in L5178Y‐MDR cells (Neto et al., 2019). Three types of latilagascenes (G, H and I), macrocyclic diterpene esters, were effective in L5178 cells expressing P-gp as transport inhibitors (Duarte et al., 2008). All successfully tested diterpenes mentioned above had a cyclopentane moiety in their structure. An enormous increase of activity was observed due to their saturation. Moreover, there was lower MDR-reversing activity mostly when the number of three hydroxyls was exceeded, especially in position 5, 7, 9 and 12. Nevertheless hydropxyls at C-1, C-13, C-14 and C-15 had not any negative effect on their function (Zhu et al., 2016). Vasas et al. (2011) observed that the lack of oxygenation at C-2 increased the MDR-reversing activity in diterpene jatrophanes such as 3β,5α,15β-triacetoxy-7β-isobutanoyloxyjatropha-6(17),11E-diene-9,14-dione and 3β,5α,15β-triacetoxy-7β-isobutanoyloxy-9α-nicotinoyloxyjatropha-6(17),11-dien-14-one. Furthermore, they stated that the presence of benzoyl or nicotinoyl group at C-2 instead of an acetyl group increases the potency of jatrophane diterpenes. The reversal of MDR was noticed in some triterpenoids including ursolic acid tested in KB-C2 cells (Nabekura et al., 2010), and deacteylnomilin and limonin in CEM/ADR5000 cells (El-Readi et al., 2010). A study on triterpenoids such as tormentic acid and derivatives suggests that an acetylation of C-2 of a terpenoid can cause an increase of activity, while this is not the case with the acetylation of C-3 (da Graça Rocha et al., 2007). Carotenoids, a group derived from tetraterpenes, are substrates for ABC transporters and can effectively modulate MDR in cancer cells (Eid et al., 2012). A few types such as β-carotene (Eid et al., 2013) or the xanthophyll member capsorubin showed MDR reversal activity (Molnár et al., 2006). To mention another important group with aromatic members, coumarins and coumaric acids are benzopyrones which we presented in Tables 1 and 2. Auraptene for example not only caused an enhancement of drug uptake (Nabekura et al., 2008b); its effect even was of a synergistic kind with drugs such as cisplatin and VCR (Moussavi et al., 2017). Kasaian et al. (2015) examined fifteen sesquiterpene coumarins. Considering the structure-activity relationship they described that ring-opened drimane-type sesquiterpene coumarins such as lehmferin, farnesiferol B and farnesiferol C showed the best P-gp inhibitory effects. Their study also revealed that farnesiferol C inhibited the Rh-123 efflux even more than the positive control verapamil.
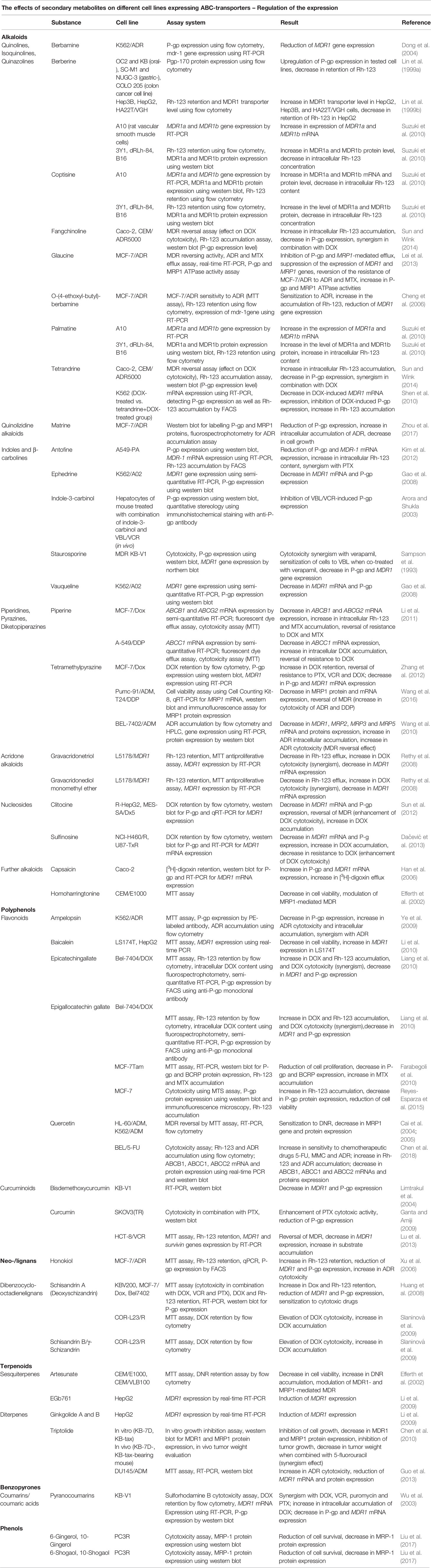
Table 2 Phytochemicals Modulating Transporter or Protein Expression.
The studies introduced phytochemicals with completely different basic structures, each showing both strong and ineffective members in targeting ABC-transporters. The molecules mentioned were a selection chosen due to their effectiveness, especially in relation to the substances to which they were compared.
The following discussion focusses on the question how far phytochemicals affect the expression of ABC transporters or proteins in cancer cell lines. A list of relevant publications is documented in Table 2. Although many investigations addressed the inhibitory effect of secondary metabolites on transporter activity, many phytochemicals regulate the expression of corresponding genes, including alkaloids, polyphenols, lignans, terpenes and benzopyrones. Berbamine caused a downregulation of Mdr-1 expression in K562/ADR cells (Dong et al., 2004), and so did its derivative O-(4-ethoxylbutyl)-berbamine (Cheng et al., 2006). Glaucine not only functioned as inhibitor of transporters but also reduced the expression of MDR1 and MRP1 (Lei et al., 2013) while the isoquinoline alkaloids berberine and coptisine, mediated the expression of P-gp (Lin et al., 1999a). Tetramethylpyrazine downregulated the expression of MDR1, MRP2, MRP3 and MRP5 in BEL-7402/ADM cells (Wang et al., 2010). It has also reduced the expression of MDR1 and MRP1 in MCF-7/Dox and Pumc-91/ADM cells, respectively (Zhang et al., 2012; Wang et al., 2016). MDR1 mRNA and P-gp expression were reduced in response to clitocine and sulfinosine in several cell lines, such as R-HepG2, MES-SA/Dx5, NCI-H460/R and U87-TxR (Sun et al., 2012; Dačević et al., 2013). Among polyphenols, EGCG mediated a reduction of P-gp and BCRP expression resulting in drug accumulation (Farabegoli et al., 2010). Curcumin enhanced the cytotoxicity of PTX and ADR, respectively against SKOV3(TR) and K562/A02 cells by reducing the expression of P-gp in both cell lines (Chang et al., 2006; Ganta and Amiji, 2009). While some diterpenes and sesquiterpenes induced the transporter expression (Li et al., 2009), others like triptolide suppressed the expression of MDR1 and MRP1 proteins in KB-7D and KB-tax cells (Chen et al., 2010). Beside the positive effect of some coumarins on the activity of ABC transporters, pyranocoumarin was tested on KB-V1 cells and showed a reduction of P-gp protein and MDR1 expression (Wu et al., 2003).
Athough no clear relationship could be established between the structure of a modulator with thre expression of a transporter protein, these findings can be relevant for clinical applications.
In addition to affecting the activity or expression of transporters, phytochemicals can also reverse MDR in cancer cells through synergism. Phytochemicals causing synergistic interactions with anticancer drugs are documented in Tables 1 and 2. The two-drug combination of harmine with DOX not only showed an enhancement of cytotoxicity but a synergistic effect. Glaucine potentiated DOX toxicity in Caco-2 and CEM/ADR5000 cells and reversed their resistance to anticancer drugs. Glaucine exerted synergistic interaction with DOX and even more in a three drug combination with DTN (Eid et al., 2013). Euphoboetirane C, D, F, G, H and I extracted from Euphorbia boetica showed strong synergistic interactions with DOX in L5178Y‐MDR cells (Neto et al., 2019). Nobiletin and antofine showed synergistic interactions with PTX (Kim et al., 2012; Ma et al., 2015). Jatrophane diterpenes including euphowelwitschine A, euphowelwitschine B, epoxywelwitschene, esulatin M isolated from Euphorbia welwitschii demonstrated synergism with DOX in L5178Y‐MDR cells (Reis et al., 2016). Fangchinoline, tetrandrine, and pyranocoumarins has been shown to synergistically increase the cytotoxicity of DOX (Wu et al., 2003; Sun and Wink, 2014). These synergistic effects are probably caused by interference of the phytochemicals with ABC transporters and additional molecular targets in cancer cells (Wink et al., 2012). These studies show that a combination of substances can have a great advantage over using single drugs against cancer cells by exploiting synergism.
In Silico experiments are promising methods for making predictions about interactions of molecules to the drug-binding site of a target such as P-gp, BCRP and MRP1. Molecular docking studies can help finding the phytochemicals with best affinity for the drug-binding site which should be used as lead molecules for further studies. The nucleotide-binding domains (NBD) of these ABC-Transporters are hydrophilic protein parts (Jones and George, 2002). In addition to essential hydrogen bonds which play a crucial role in maintaining the stability and function of biomolecules, modulators with hydrophobic moieties often show affinties to NBDs.
The benzolisoquinoline alkaloid tetrandrine was confirmed to have inhibitory capability. Its binding affinity is close to verapamil and their docking positions are similar. Main part of tetrandrine is fixed in a lipophilic pocket formed by the amino acids Ala729, Ala987, Ileu306, Ileu340, Leu339, Leu65, Leu975, Met69, Met986, Phe303, Phe336, Phe343, Phe728, Phe732, Phe983, Tyr307, Tyr310. Moreover, the positively charged methylamine moiety of tetrandrine formed a cation-π interaction with Phe343 and an aryl ether got into a π-π interaction with Phe336 (Liao et al., 2019).
Regarding in silico investigations, another subtype of alkaloids, piperine was examined by Syed et al. (2017) revealing that hydrophobic interactions with P-gp took place in following positions: Leu339, Met69, Met986, Phe72, Phe336, Phe728, Phe983, Tyr953 and Val982. Moreover, an H-bond with Tyr307 was established. Two piperine analogs were designed which showed better hydrophobic interaction with most of the amino acids mentioned (Syed et al., 2017).
Pharmacophore modeling which was carried out for acridones indicated that three aromatic rings and two H–acceptors which are given in position C-9 bearing a carbonyl group and N-10 were conducive for stable docking. Including designed analogs of acridone the oxygen of a morpholine moiety at a phenyl ring built a water bridge with Ser309 of P-gp and a carboxamide at C-4 helped docking with Phe343. This analog showed hydrophobic interactions with Ala229, Ala302, Ala342, Ile218, Ile221, Ile299, Ile306, Leu225 with even more than two interactions, Leu339, Phe343, and Val345 of P-gp (Gade et al., 2018).
Badhan and Penny (2006) worked on the main characteristics of the pharmacophore of flavonoids by in silico modeling. Flavonoids tested get located within the C-terminal NBD of P-gp binding to amino acids of the ATP pocket such as Tyr1044. The hydroxyl moiety at C-5 of natural flavonoids can form a water bridge with Lys1076 while the hydroxyl group in position 3 and the carbonyl group get into hydrogen bonds within the NBD. Hydroxyl groups at position C-7 led to important hydrogen bonds with amino acids for several flavonoids such as EGC and chrysin (Wongrattanakamon et al., 2017). Ring B gets involved in π- π interaction with Tyr1044. If benzyl, geranyl or prenyl groups added at C-6 and several substitutes added at C-8 position, the docking capability was enhanced. Molecular docking analysis demonstrated that apigenin binds to the ATP-binding site of P-gp through a hydrogen bond with LYS408 and therefore, interferes with binding and cleavage of ATP which are vital for the function of ABC transporters (Saeed et al., 2015). Comparing flavonols to their corresponding flavones such as kaempferol to apigenin, better docking properties have been found in flavonols due to the number of hydrogen bonds (Badhan and Penny, 2006). A number of naringenin derivatives such as hydrazones and azines were produced by Ferreira et al. (2018) to achieve an improved MDR reversal in P-gp and BCRP. There was a high selectivity for most members of these two groups. It has been shown that the active compounds bind to hydrophobic residues such as transmembrane helix 4 and 5 in the center of an active pocket of BCRP similar to fumitremorgin C, a common inhibitor of BCRP. The hydrazone of these derivatives, a continuation of imine, is located in a pocket in π- π interaction with Phe515 and a dipole-dipole interaction with Met541. The B-ring is in another lipophilic bag made of Phe545, Pro574 and Val516. An improvement in the interaction between the transporter MRP1 and the carbohydrazide derivatives of flavonones due to the resulting flexibility was also observed (Ferreira et al., 2018).
Due to the increase of cancer cases and restrictions in therapy owing to the degree of harmfulness, effectiveness and the associated high costs, the development of new substances with less side effects and higher efficacy is required. So far, many structures from the plant kingdom have been discovered and used, such as, diterpene derivatives as taxanes and vinca alkaloids are prime examples with regard to antitumor therapy. Therefore, numerous secondary metabolites which already had positive effect on cell lines should be taken into account for further investigations, which is why we have tried to list them extensively in our tables.
Many phytochemicals including alkaloids, flavonoids, curcuminoids, stilbenoids, terpenes, carotenoids, lignans, and polyketides were examined for their pharmacological activity against MDR. In most of the studies it was possible to differentiate between more effective and less effective molecules. As the next step these molecules must be further studied to determine molecular mechanisms and to identify the pharmacophores. Some valuable results have been provided by structure-activity relationship studies or in silico modeling as shown in this article. Subtances which were superior to others should be selected for further research, so possible lead molecules can be developed. A further step to increase efficacy could be the use of new drug delivery methods and controlled release as with nanotechnologies. As shown in this review, the combination of two and more modulators of differing structures and mode of actions together with chemotherapeutics is another interesting approach. The resulting additive, but above all synergistic effects can be of great importance. They would allow reducing the dose of chemotherapeutics resulting in less side effects and a higher compliance of patients.
Summin up, many phytochemicals in the group of alkaloids, flavonoids, curcuminoids, stilbenoids, terpenes, carotenoids, lignans, and polyketides have been investigated for MDR-reversing activity. Phytochemicals can be a promising source of adjuvant chemicals against cancer, not at least because of their generally low cytotoxicity in the human body. The adjuvant uses of MDR reversing phytochemicals in combination with anticancer drugs may improve the treatment of multidrug resistant cancer types. The present review summarized reports of several secondary metabolites that are capable of synergistically reversing MDR and inhibiting chemotherapy-resistant cancer cells by affecting transporter activity and the expression of ABC transporter genes. Synergism would allow reducing the dose of chemotherapeutics resulting in less side effects and a higher compliance of patients. The efficacy of phytochemicals needs to be confirmed clinically, but nevertheless they already can be considered as the fourth generation of ABC transporter modulators.
BT and IS performed the literature search and wrote the first draft of the manuscript. MW revised and edited the manuscript. All the authors approved the final version of the manuscript.
Ruprecht-Karls-Universität Heidelberg provided financial support within Open-Access Publishing Program.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
ADR, adriamycin; CPT, camptothecin; DOX, doxorubicin; DNR, daunorubicin; DTN, digitonin; EGC, epigallocatechin; EGCG, epigallocatechin gallate; MDR, multidrug resistance; MTX, mitoxantrone; P-gp, P-glycoprotein; PTX, paclitaxel; Rh-123, rhodamine 123; VBL, vinblastine; VCR, vincristine; BCRP, breast cancer resistance protein; MRP, multidrug resistance protein.
Aktories, K., Förstermann, U., Hofmann, F. B., Starke, K. (2017). Allgemeine und spezielle Pharmakologie und Toxikologie: Begründet von W. Forth. Eds. Henschler, D., Rummel, W. (München, Germany: Urban & Fischer Verlag/Elsevier GmbH).
Alamolhodaei, N. S., Tsatsakis, A. M., Ramezani, M., Hayes, A. W., Karimi, G. (2017). Resveratrol as MDR reversion molecule in breast cancer: An overview. Food Chem. Toxicol. 103, 223–232. doi: 10.1016/j.fct.2017.03.024
Ampasavate, C., Sotanaphun, U., Phattanawasin, P., Piyapolrungroj, N. (2010). Effects of Curcuma spp. on P-glycoprotein function. Phytomedicine 17, 506–512. doi: 10.1016/j.phymed.2009.09.004
Arora, A., Shukla, Y. (2003). Modulation of vinca-alkaloid induced P-glycoprotein expression by indole-3-carbinol. Canc. Lett. 189, 167–173. doi: 10.1016/S0304-3835(02)00550-5
Badhan, R., Penny, J. (2006). In silico modelling of the interaction of flavonoids with human P-glycoprotein nucleotide-binding domain. Eur. J. Med. Chem. 41, 285–295. doi: 10.1016/j.ejmech.2005.11.012
Baumann, T. W., Frischknecht, P. M. (1988). “Purines,” in Phytochemicals in Plant Cell Cultures (San Diego, USA: Academic Press), 403–417.
Bayet, C., Fazio, C., Darbour, N., Berger, O., Raad, I., Chaboud, A., et al. (2007). Modulation of P-glycoprotein activity by acridones and coumarins from Citrus sinensis. Phytother. Res. 21, 386–390. doi: 10.1002/ptr.2081
Berek, L., Szabo, D., Petri, I. B., Shoyama, Y., Lin, Y. H., Molnár, J. (2001). Effects of naturally occurring glucosides, solasodine glucosides, ginsenosides and parishin derivatives on multidrug resistance of lymphoma cells and leukocyte functions. Vivo 15, 151–156.
Bernstein, C., Bernstein, H., Payne, C. M., Garewal, H. (2002). DNA repair/pro-apoptotic dual-role proteins in five major DNA repair pathways: fail-safe protection against carcinogenesis. Mutat. Res. Rev. Mutat. 511, 145–178. doi: 10.1016/S1383-5742(02)00009-1
Bloch, K. E. (1983). Sterol, structure and membrane function. Crit. Rev. Biochem. 14, 47–92. doi: 10.3109/10409238309102790
Borst, P., Evers, R., Kool, M., Wijnholds, J. (2000). A family of drug transporters: the multidrug resistance-associated proteins. J. Natl. Canc. Inst. 92, 1295–1302. doi: 10.1093/jnci/92.16.1295
Borthwick, A. D., Liddle, J. (2011). The design of orally bioavailable 2, 5-diketopiperazine oxytocin antagonists: from concept to clinical candidate for premature labor. Med. Res. Rev. 31, 576–604. doi: 10.1002/med.20193
Boumendjel, A., Di Pietro, A., Dumontet, C., Barron, D. (2002). Recent advances in the discovery of flavonoids and analogs with high-affinity binding to P-glycoprotein responsible for cancer cell multidrug resistance. Med. Res. Rev. 22, 512–529. doi: 10.1002/med.10015
Brown, G. A., McPherson, J. P., Gu, L., Hedley, D. W., Toso, R., Deuchars, K. L., et al. (1995). Relationship of DNA topoisomerase IIα and β expression to cytotoxicity of antineoplastic agents in human acute lymphoblastic leukemia cell lines. Canc. Res. 55, 78–82.
Cai, X., Chen, F. Y., Han, J. Y., Gu, C. H., Zhong, H., Ouyang, R. R. (2004). Restorative effect of quercetin on subcellular distribution of daunorubicin in multidrug resistant leukemia cell lines K562/ADM and HL-60/ADM. Chin. J. Canc. 23, 1611–1615.
Cai, X., Chen, F. Y., Han, J. Y., Gu, C. H., Zhong, H., Teng, Y., et al. (2005). Reversal of multidrug resistance of HL-60 adriamycin resistant leukemia cell line by quercetin and its mechanisms. Chin. J. Oncol. 27, 326–329.
Cao, J., Chen, X., Liang, J., Yu, X. Q., Xu, A. L., Chan, E., et al. (2007). Role of P-glycoprotein in the intestinal absorption of glabridin, an active flavonoid from the root of Glycyrrhiza glabra. Drug Metabol. Dispos. 35, 539–553. doi: 10.1124/dmd.106.010801
Cao, Z., Wright, M., Cheng, J., Huang, X., Liu, L., Wu, L., et al. (2014). The novel bis-benzylisoquinoline PY35 reverses P-glycoprotein-mediated multidrug resistance. Oncol. Rep. 32, 1211–1217. doi: 10.3892/or.2014.3326
Chanmahasathien, W., Ampasavate, C., Greger, H., Limtrakul, P. (2011). Stemona alkaloids, from traditional Thai medicine, increase chemosensitivity via P-glycoprotein-mediated multidrug resistance. Phytomedicine 18, 199–204. doi: 10.1016/j.phymed.2010.07.014
Chang, H. Y., Pan, K. L., Ma, F. C., Jiao, X. Y., Zhu, H. F., Liu, J. H., et al. (2006). The Study on Reversing Mechanism of Multidrug Resistance of K562/A02 Cell Line by Curcumin and Erythromycin. Chin. J. Hematol. 27, 254–258.
Chen, Y. W., Lin, G. J., Chuang, Y. P., Chia, W. T., Hueng, D. Y., Lin, C. K., et al. (2010). Triptolide circumvents drug-resistant effect and enhances 5-fluorouracil antitumor effect on KB cells. Anti Canc. Drugs 21, 502–513. doi: 10.1097/CAD.0b013e328337337c
Chen, Z., Shi, T., Zhang, L., Zhu, P., Deng, M., Huang, C., et al. (2016). Mammalian drug efflux transporters of the ATP binding cassette (ABC) family in multidrug resistance: A review of the past decade. Canc. Lett. 370, 153–164. doi: 10.1016/j.canlet.2015.10.010
Chen, Z., Huang, C., Ma, T., Jiang, L., Tang, L., Shi, T., et al. (2018). Reversal effect of quercetin on multidrug resistance via FZD7/β-catenin pathway in hepatocellular carcinoma cells. Phytomedicine 43, 37–45. doi: 10.1016/j.phymed.2018.03.040
Cheng, Y. H., Qi, J., Xiong, D. S., Liu, J. W., Qi, S. L., Pan, B., et al. (2006). Reversal of multidrug resistance in drug-resistant human breast cancer cell line MCF-7/ADR by calmodulin antagonist O-(4-ethoxyl-butyl)-berbamine. Acta Academiae Medicinae Sinicae 28, 164–168.
Choi, C. H., Kang, G., Min, Y. D. (2003). Reversal of P-glycoprotein-mediated multidrug resistance by protopanaxatriol ginsenosides from Korean red ginseng. Planta Med. 69, 235–240. doi: 10.1055/s-2003-38483
Chung, S. Y., Sung, M. K., Kim, N. H., Jang, J. O., Go, E. J., Lee, H. J. (2005). Inhibition of P-glycoprotein by natural products in human breast cancer cells. Arch. Pharmacal Res. 28, 823–828. doi: 10.1007/BF02977349
Cihalova, D., Hofman, J., Ceckova, M., Staud, F. (2013). Purvalanol A, olomoucine II and roscovitine inhibit ABCB1 transporter and synergistically potentiate cytotoxic effects of daunorubicin in vitro. PloS One 8, e83467. doi: 10.1371/journal.pone.0083467
Coley, H. M. (2008). Mechanisms and strategies to overcome chemotherapy resistance in metastatic breast cancer. Canc. Treat. Rev. 34, 378–390. doi: 10.1016/j.ctrv.2008.01.007
Collin, G., Höke, H. (2000). “Quinoline and isoquinoline,” in Ullmann"s Encyclopedia of Industrial Chemistry (New Jersey, USA: Wiley-VCH).
Conseil, G., Baubichon-Cortay, H., Dayan, G., Jault, J. M., Barron, D., Di Pietro, A. (1998). Flavonoids: a class of modulators with bifunctional interactions at vicinal ATP-and steroid-binding sites on mouse P-glycoprotein. Proc. Natl. Acad. Sci. 95, 9831–9836. doi: 10.1073/pnas.95.17.9831
Cör, D., Knez, Ž., Knez Hrnčič, M. (2018). Antitumour, antimicrobial, antioxidant and antiacetylcholinesterase effect of Ganoderma lucidum terpenoids and polysaccharides: A review. Molecules 23, 649. doi: 10.3390/molecules23030649
Cripe, L. D., Uno, H., Paietta, E. M., Litzow, M. R., Ketterling, R. P., Bennett, J. M., et al. (2010). Zosuquidar, a novel modulator of P-glycoprotein, does not improve the outcome of older patients with newly diagnosed acute myeloid leukemia: a randomized, placebo-controlled trial of the Eastern Cooperative Oncology Group 3999. Blood 116, 4077–4085. doi: 10.1182/blood-2010-04-277269
Cseke, L. J., Kirakosyan, A., Kaufman, P. B., Warber, S. L., Duke, J. A., Brielmann, H. L. (2006). Natural Products from Plants. 2nd Ed (Boca Raton, USA: CRC Press), ISBN: 0-8493-2976-0.
da Graça Rocha, G., Simoes, M., Lucio, K. A., Oliveira, R. R., Kaplan, M. A. C., Gattass, C. R. (2007). Natural triterpenoids from Cecropia lyratiloba are cytotoxic to both sensitive and multidrug resistant leukemia cell lines. Bioorg. Med. Chem. 15, 7355–7360. doi: 10.1016/j.bmc.2007.07.020
Dačević, M., Isaković, A., Podolski-Renić, A., Isaković, A. M., Stanković, T., Milošević, Z., et al. (2013). Purine nucleoside analog-sulfinosine modulates diverse mechanisms of cancer progression in multi-drug resistant cancer cell lines. PloS One 8, e54044. doi: 10.1371/journal.pone.0054044
Daenen, S., van der Holt, B., Verhoef, G. E., Löwenberg, B., Wijermans, P. W., Huijgens, P. C., et al. (2004). Addition of cyclosporin A to the combination of mitoxantrone and etoposide to overcome resistance to chemotherapy in refractory or relapsing acute myeloid leukaemia: A randomised phase II trial from HOVON, the Dutch–Belgian Haemato-Oncology Working Group for adults. Leuk. Res. 28, 1057–1067. doi: 10.1016/j.leukres.2004.03.001
De Castro, W. V., Mertens-Talcott, S., Derendorf, H., Butterweck, V. (2008). Effect of grapefruit juice, naringin, naringenin, and bergamottin on the intestinal carrier-mediated transport of talinolol in rats. J. Agr. Food Chem. 56, 4840–4845. doi: 10.1021/jf0728451
de Sa, A., Fernando, R., Barreiro, E. J., Fraga, M., Alberto, C. (2009). From nature to drug discovery: the indole scaffold as a ‘privileged structure'. Mini Rev. Med. Chem. 9, 782–793. doi: 10.2174/138955709788452649
Di Pietro, A., Conseil, G., Perez-Victoria, J. M., Dayan, G., Baubichon-Cortay, H., Trompier, D., et al. (2002). Modulation by flavonoids of cell multidrug resistance mediated by P-glycoprotein and related ABC transporters. Cell. Mol. Life Sci. 59, 307–322. doi: 10.1007/s00018-002-8424-8
Dong, Q. H., Zheng, S., Xu, R. Z., Lu, Q., He, L. (2004). Study on effect of berbamine on multidrug resistance leukemia K562/Adr cells. Chin. J. Integrated Tradit. West. Med. 24, 820–822.
Doyle, L. A., Yang, W., Abruzzo, L. V., Krogmann, T., Gao, Y., Rishi, A. K., et al. (1998). A multidrug resistance transporter from human MCF-7 breast cancer cells. Proc. Natl. Acad. Sci. 95, 15665–15670. doi: 10.1073/pnas.95.26.15665
Duarte, N., Gyémánt, N., Abreu, P. M., Molnár, J., Ferreira, M. J. U. (2006). New macrocyclic lathyrane diterpenes, from Euphorbia lagascae, as inhibitors of multidrug resistance of tumour cells. Planta Med. 72, 162–168. doi: 10.1055/s-2005-873196
Duarte, N., Járdánházy, A., Molnár, J., Hilgeroth, A., Ferreira, M. J. U. (2008). Synergistic interaction between p-glycoprotein modulators and epirubicine on resistant cancer cells. Bioorg. Med. Chem. 16, 9323–9330. doi: 10.1016/j.bmc.2008.08.071
Durmus, S., Hendrikx, J. J., Schinkel, A. H. (2015). “Apical ABC transporters and cancer chemotherapeutic drug disposition,” in Advances in Cancer Research (San Diego, USA: Academic Press), 1–41.
Efferth, T., Davey, M., Olbrich, A., Rücker, G., Gebhart, E., Davey, R. (2002). Activity of drugs from traditional Chinese medicine toward sensitive and MDR1-or MRP1-overexpressing multidrug-resistant human CCRF-CEM leukemia cells. Blood Cells Mol. Dis. 28, 160–168. doi: 10.1006/bcmd.2002.0492
Eid, S. Y., El-Readi, M. Z., Wink, M. (2012). Carotenoids reverse multidrug resistance in cancer cells by interfering with ABC-transporters. Phytomedicine 19, 977–987. doi: 10.1016/J.PHYMED.2012.05.010
Eid, S. Y., El-Readi, M. Z., Eldin, E. E. M. N., Fatani, S. H., Wink, M. (2013). Influence of combinations of digitonin with selected phenolics, terpenoids, and alkaloids on the expression and activity of P-glycoprotein in leukaemia and colon cancer cells. Phytomedicine 21, 47–61. doi: 10.1016/J.PHYMED.2013.07.019
El-Readi, M. Z., Hamdan, D., Farrag, N., El-Shazly, A., Wink, M. (2010). Inhibition of P-glycoprotein activity by limonin and other secondary metabolites from Citrus species in human colon and leukaemia cell lines. Eur. J. Pharmacol. 626, 139–145. doi: 10.1016/j.ejphar.2009.09.040
El-Readi, M. Z., Eid, S., Abdelghany, A. A., Al-Amoudi, H. S., Efferth, T., Wink, M. (2019). Resveratrol mediated cancer cell apoptosis, and modulation of multidrug resistance proteins and metabolic enzymes. Phytomedicine 55, 269–281. doi: 10.1016/J.PHYMED.2018.06.046
Fantini, M., Benvenuto, M., Masuelli, L., Frajese, G. V., Tresoldi, I., Modesti, A., et al. (2015). In vitro and in vivo antitumoral effects of combinations of polyphenols, or polyphenols and anticancer drugs: Perspectives on cancer treatment. Int. J. Mol. Sci. 16, 9236–9282. doi: 10.3390/ijms16059236
Farabegoli, F., Papi, A., Bartolini, G., Ostan, R., Orlandi, M. (2010). (-)-Epigallocatechin-3-gallate downregulates P-gp and BCRP in a tamoxifen resistant MCF-7 cell line. Phytomedicine 17, 356–362. doi: 10.1016/j.phymed.2010.01.001
Ferreira, R. J., Baptista, R., Moreno, A., Madeira, P. G., Khonkarn, R., Baubichon-Cortay, H., et al. (2018). Optimizing the flavanone core toward new selective nitrogen-containing modulators of ABC transporters. Future Med. Chem. 10, 725–741. doi: 10.4155/fmc-2017-0228
Fong, W. F., Wan, C. K., Zhu, G. Y., Chattopadhyay, A., Dey, S., Zhao, Z., et al. (2007). Schisandrol A from Schisandra chinensis reverses P-glycoprotein-mediated multidrug resistance by affecting Pgp-substrate complexes. Planta Med. 73, 212–220. doi: 10.1055/s-2007-967120
Gade, D. R., Makkapati, A., Yarlagadda, R. B., Peters, G. J., Sastry, B. S., Prasad, V. R. (2018). Elucidation of chemosensitization effect of acridones in cancer cell lines: Combined pharmacophore modeling, 3D QSAR, and molecular dynamics studies. Comput. Biol. Chem. 74, 63–75. doi: 10.1016/j.compbiolchem.2018.02.014
Gan, C. Y., Low, Y. Y., Etoh, T., Hayashi, M., Komiyama, K., Kam, T. S. (2009). Leuconicines A− G and (−)-Eburnamaline, Biologically Active Strychnan and Eburnan Alkaloids from Leuconotis. J. Nat. Prod. 72, 2098–2103. doi: 10.1021/np900576b
Gan, C. Y., Yoganathan, K., Sim, K. S., Low, Y. Y., Lim, S. H., Kam, T. S. (2014). Corynanthean, eburnan, secoleuconoxine, and pauciflorine alkaloids from Kopsia pauciflora. Phytochemistry 108, 234–242. doi: 10.1016/j.phytochem.2014.09.014
Ganta, S., Amiji, M. (2009). Coadministration of paclitaxel and curcumin in nanoemulsion formulations to overcome multidrug resistance in tumor cells. Mol. Pharm. 6, 928–939. doi: 10.1021/mp800240j
Gao, N., Zhang, Y., Mao, J. Q., Li, G. Q., Zhou, W., Gao, B., et al. (2008). Effects of some traditional Chinese drugs on Mdr1 gene and its expression product in K562/A02 cells. J. Exp. Hematol. 16, 785–789.
Gawali, V. S., Simeonov, S., Drescher, M., Knott, T., Scheel, O., Kudolo, J., et al. (2017). C2-modified sparteine derivatives are a new class of potentially long-acting sodium channel blockers. ChemMedChem 12, 1819–1822. doi: 10.1002/cmdc.201700568
Gilbert, K. (2001). Review of Plant Secondary Metabolism, by David S. Seigler. Plant Growth Regul. 34, 149. doi: 10.1023/A:1013354907356
Guo, Q., Nan, X. X., Yang, J. R., Yi, L., Liang, B. L., Wei, Y. B., et al. (2013). Triptolide inhibits the multidrug resistance in prostate cancer cells via the downregulation of MDR1 expression. Neoplasma 60, 598–604. doi: 10.4149/neo_2013_077
Gyémánt, N., Tanaka, M., Antus, S., Hohmann, J., Csuka, O., Mándoky, L., et al. (2005). andIn vitro search for synergy between flavonoids and epirubicin on multidrug-resistant cancer cells. Vivo 19, 367–374.
Gyémánt, N., Tanaka, M., Molnár, P., Deli, J., Mándoky, L., Molnár, J. (2006). Reversal of multidrug resistance of cancer cells in vitro: modification of drug resistance by selected carotenoids. Anticancer Res. 26, 367–374.
Hänsel, R., Sticher, O. (Eds.) (2009). Pharmakognosie-Phytopharmazie (Heidelberg, Germany: Springer-Verlag).
Han, Y., Tan, T. M. C., Lim, L. Y. (2006). Effects of capsaicin on P-gp function and expression in Caco-2 cells. Biochem. Pharmacol. 71, 1727–1734. doi: 10.1016/j.bcp.2006.03.024
He, L., Zhao, C., Yan, M., Zhang, L. Y., Xia, Y. Z. (2009). Inhibition of P-glycoprotein function by procyanidine on blood–brain barrier. Phytother. Res. 23, 933–937. doi: 10.1002/ptr.2781
He, L., Yang, J., Hu, L. (2010). Transmembrane transport activity of paclitaxel regulated by fangchinoline in MDR1-mDCK II cells. China J. Chin. Materia Med. 35, 1478–1481.
Hegde, R., Thimmaiah, P., Yerigeri, M. C., Krishnegowda, G., Thimmaiah, K. N., Houghton, P. J. (2004). Anti-calmodulin acridone derivatives modulate vinblastine resistance in multidrug resistant (MDR) cancer cells. Eur. J. Med. Chem. 39, 161–177. doi: 10.1016/j.ejmech.2003.12.001
Hsiao, S. H., Lu, Y. J., Yang, C. C., Tuo, W. C., Li, Y. Q., Huang, Y. H., et al. (2016). Hernandezine, a bisbenzylisoquinoline alkaloid with selective inhibitory activity against multidrug-resistance-linked ATP-binding cassette drug transporter ABCB1. J. Nat. Prod. 79, 2135–2142. doi: 10.1021/acs.jnatprod.6b00597
Hu, Y. J., Shen, X. L., Lu, H. L., Zhang, Y. H., Huang, X. A., Fu, L. C., et al. (2008). Tenacigenin B derivatives reverse P-glycoprotein-mediated multidrug resistance in HepG2/Dox cells. J. Nat. Prod. 71, 1049–1051. doi: 10.1021/np070458f
Huang, M., Jin, J., Sun, H., Liu, G. T. (2008). Reversal of P-glycoprotein-mediated multidrug resistance of cancer cells by five schizandrins isolated from the Chinese herb Fructus schizandrae. Canc. Chemother. Pharmacol. 62, 1015–1026. doi: 10.1007/s00280-008-0691-0
Huang, F., Wu, X. N., Chen, J. I. E., Wang, W. X., Lu, Z. F. (2014). Resveratrol reverses multidrug resistance in human breast cancer doxorubicin-resistant cells. Exp. Ther. Med. 7, 1611–1616. doi: 10.3892/etm.2014.1662
Ikegawa, T., Ushigome, F., Koyabu, N., Morimoto, S., Shoyama, Y., Naito, M., et al. (2000). Inhibition of P-glycoprotein by orange juice components, polymethoxyflavones in adriamycin-resistant human myelogenous leukemia (K562/ADM) cells. Canc. Lett. 160, 21–28. doi: 10.1016/S0304-3835(00)00549-8
Imai, Y., Tsukahara, S., Asada, S., Sugimoto, Y. (2004). Phytoestrogens/flavonoids reverse breast cancer resistance protein/ABCG2-mediated multidrug resistance. Canc. Res. 64, 4346–4352. doi: 10.1158/0008-5472.CAN-04-0078
Incardona, J. P., Gaffield, W., Kapur, R. P., Roelink, H. (1998). The teratogenic Veratrum alkaloid cyclopamine inhibits sonic hedgehog signal transduction. Development 125, 3553–3562.
Ivanova, A., Serly, J., Dinchev, D., Ocsovszki, I., Kostova, I., Molnár, J. (2009). Screening of some saponins and phenolic components of Tribulus terrestris and Smilax excelsa as MDR modulators. Vivo 23, 545–550.
Ivanova, A., Serly, J., Christov, V., Stamboliyska, B., Molnaár, J. (2011). Alkaloids derived from genus Veratrum and Peganum of Mongolian origin as multidrug resistance inhibitors of cancer cells. Fitoterapia 82, 570–575. doi: 10.1016/J.FITOTE.2011.01.015
Jones, P. M., George, A. M. (2002). Mechanism of ABC transporters: a molecular dynamics simulation of a well characterized nucleotide-binding subunit. Proc. Natl. Acad. Sci. 99, 12639–12644. doi: 10.1073/pnas.152439599
Kam, T. S., Subramaniam, G., Sim, K. M., Yoganathan, K., Koyano, T., Toyoshima, M., et al. (1998). Reversal of multidrug resistance (MDR) by aspidofractinine-type indole alkaloids. Bioorg. Med. Chem. Lett. 8, 2769–2772. doi: 10.1016/S0960-894X(98)00486-7
Kamath, K., Wilson, L., Cabral, F., Jordan, M. A. (2005). βIII-tubulin induces paclitaxel resistance in association with reduced effects on microtubule dynamic instability. J. Biol. Chem. 280, 12902–12907. doi: 10.1074/jbc.M414477200
Kasaian, J., Mosaffa, F., Behravan, J., Masullo, M., Piacente, S., Ghandadi, M., et al. (2015). Reversal of P-glycoprotein-mediated multidrug resistance in MCF-7/Adr cancer cells by sesquiterpene coumarins. Fitoterapia 103, 149–154. doi: 10.1016/j.fitote.2015.03.025
Kashiwada, Y., Nishimura, K., Kurimoto, S., Takaishi, Y. (2011). New 29-nor-cycloartanes with a 3,4-seco- and a novel 2,3-seco-structure from the leaves of Sinocalycanthus chinensis. Bioorg. Med. Chem. 19, 2790–2796. doi: 10.1016/j.bmc.2011.03.055
Kavallaris, M., Tait, A. S., Walsh, B. J., He, L., Horwitz, S. B., Norris, M. D., et al. (2001). Multiple microtubule alterations are associated with Vinca alkaloid resistance in human leukemia cells. Canc. Res. 61, 5803–5809.
Kelly, R. J., Draper, D., Chen, C. C., Robey, R. W., Figg, W. D., Piekarz, R. L., et al. (2011). A pharmacodynamic study of docetaxel in combination with the P-glycoprotein antagonist tariquidar (XR9576) in patients with lung, ovarian, and cervical cancer. Clin. Canc. Res. 17, 569–580. doi: 10.1158/1078-0432.CCR-10-1725
Kim, S. W., Kwon, H. Y., Chi, D. W., Shim, J. H., Park, J. D., Lee, Y. H., et al. (2003). Reversal of P-glycoprotein-mediated multidrug resistance by ginsenoside Rg3. Biochem. Pharmacol. 65, 75–82. doi: 10.1016/S0006-2952(02)01446-6
Kim, E.-H., Min, H.-Y., Chung, H.-J., Song, J., Park, H.-J., Kim, S., et al. (2012). Anti-proliferative activity and suppression of P-glycoprotein by (–)-antofine, a natural phenanthroindolizidine alkaloid, in paclitaxel-resistant human lung cancer cells. Food Chem. Toxicol. 50, 1060–1065. doi: 10.1016/J.FCT.2011.11.008
Kitagawa, S., Nabekura, T., Kamiyama, S. (2004). Inhibition of P-glycoprotein function by tea catechins in KB-C2 cells. J. Pharm. Pharmacol. 56, 1001–1005. doi: 10.1211/0022357044003
Kitagawa, S., Nabekura, T., Takahashi, T., Nakamura, Y., Sakamoto, H., Tano, H., et al. (2005). Structure–activity relationships of the inhibitory effects of flavonoids on P-glycoprotein-mediated transport in KB-C2 cells. Biol. Pharm. Bull. 28, 2274–2278. doi: 10.1248/bpb.28.2274
Kitagawa, S., Takahashi, T., Nabekura, T., Tachikawa, E., Hasegawa, H. (2007). Inhibitory effects of ginsenosides and their hydrolyzed metabolites on daunorubicin transport in KB-C2 cells. Biol. Pharm. Bull. 30, 1979–1981. doi: 10.1248/bpb.30.1979
Koes, R. E., Quattrocchio, F., Mol, J. N. (1994). The flavonoid biosynthetic pathway in plants: function and evolution. BioEssays 16, 123–132. doi: 10.1002/bies.950160209
König, J., Hartel, M., Nies, A. T., Martignoni, M. E., Guo, J., Büchler, M. W., et al. (2005). Expression and localization of human multidrug resistance protein (ABCC) family members in pancreatic carcinoma. Int. J. Canc. 115, 359–367. doi: 10.1002/ijc.20831
Körper, S., Wink, M., Fink, R. H. (1998). Differential effects of alkaloids on sodium currents of isolated single skeletal muscle fibers. FEBS Lett. 436, 251–255. doi: 10.1016/S0014-5793(98)01135-1
Kukula-Koch, W. A., Widelski, J. (2017). “Alkaloids,” in Pharmacognosy (San Diego, USA: Academic Press), 163–198.
Kurimoto, S. I., Kashiwada, Y., Lee, K. H., Takaishi, Y. (2011a). Triterpenes and a triterpene glucoside from Dysoxylum cumingianum. Phytochemistry 72, 2205–2211. doi: 10.1016/j.phytochem.2011.08.002
Kurimoto, S. I., Kashiwada, Y., Morris-Natschke, S. L., Lee, K. H., Takaishi, Y. (2011b). Dyscusins A—C, three new steroids from the leaves of Dysoxylum cumingianum. Chem. Pharm. Bull. 59, 1303–1306. doi: 10.1248/cpb.59.1303
Lei, Y., Tan, J., Wink, M., Ma, Y., Li, N., Su, G. (2013). An isoquinoline alkaloid from the Chinese herbal plant Corydalis yanhusuo W.T. Wang inhibits P-glycoprotein and multidrug resistance-associate protein 1. Food Chem. 136, 1117–1121. doi: 10.1016/j.foodchem.2012.09.059
Li, L., Stanton, J. D., Tolson, A. H., Luo, Y., Wang, H. (2009). Bioactive terpenoids and flavonoids from Ginkgo biloba extract induce the expression of hepatic drug-metabolizing enzymes through pregnane X receptor, constitutive androstane receptor, and aryl hydrocarbon receptor-mediated pathways. Pharmaceut. Res. 26, 872. doi: 10.1007/s11095-008-9788-8
Li, Y., Wang, Q., Yao, X., Li, Y. (2010). Induction of CYP3A4 and MDR1 gene expression by baicalin, baicalein, chlorogenic acid, and ginsenoside Rf through constitutive androstane receptor-and pregnane X receptor-mediated pathways. Eur. J. Pharmacol. 640, 46–54. doi: 10.1016/j.ejphar.2010.05.017
Li, S., Lei, Y., Jia, Y., Li, N., Wink, M., Ma, Y. (2011). Piperine, a piperidine alkaloid from Piper nigrum re-sensitizes P-gp, MRP1 and BCRP dependent multidrug resistant cancer cells. Phytomedicine 19, 83–87. doi: 10.1016/j.phymed.2011.06.031
Li, H., Krstin, S., Wang, S., Wink, M. (2018a). Capsaicin and piperine can overcome multidrug resistance in cancer cells to doxorubicin. Molecules 23, 557. doi: 10.3390/molecules23030557
Li, H., Krstin, S., Wink, M. (2018b). Modulation of multidrug resistant in cancer cells by EGCG, tannic acid and curcumin. Phytomedicine 50, 213–222. doi: 10.1016/j.phymed.2018.09.169
Liang, G., Tang, A., Lin, X., Li, L., Zhang, S., Huang, Z., et al. (2010). Green tea catechins augment the antitumor activity of doxorubicin in an in vivo mouse model for chemoresistant liver cancer. Int. J. Oncol. 37, 111–123. doi: 10.3892/ijo_00000659
Liao, D., Zhang, W., Gupta, P., Lei, Z. N., Wang, J. Q., Cai, C. Y., et al. (2019). Tetrandrine Interaction with ABCB1 Reverses Multidrug Resistance in Cancer Cells Through Competition with Anti-Cancer Drugs Followed by Downregulation of ABCB1 Expression. Molecules 24, 4383. doi: 10.3390/molecules24234383
Limtrakul, P., Anuchapreeda, S., Buddhasukh, D. (2004). Modulation of human multidrug-resistance MDR-1 gene by natural curcuminoids. BMC Canc. 4, 13. doi: 10.1186/1471-2407-4-13
Limtrakul, P., Chearwae, W., Shukla, S., Phisalphong, C., Ambudkar, S. V. (2007). Modulation of function of three ABC drug transporters, P-glycoprotein (ABCB1), mitoxantrone resistance protein (ABCG2) and multidrug resistance protein 1 (ABCC1) by tetrahydrocurcumin, a major metabolite of curcumin. Mol. Cell. Biochem. 296, 85–95. doi: 10.1007/s11010-006-9302-8
Lin, H. L., Liu, T. Y., Lui, W. Y., Chi, C. W. (1999a). Up-regulation of multidrug resistance transporter expression by berberine in human and murine hepatoma cells. Canc. Interdiscipl. Int. J. Am. Canc. Soc 85, 1937–1942. doi: 10.1002/(SICI)1097-0142(19990501)85:9<1937::AID-CNCR9>3.0.CO;2-F
Lin, H.-L., Liu, T.-Y., Wu, C.-W., Chi, C.-W. (1999b). Berberine modulates expression of MDR1 gene product and the responses of digestive track cancer cells to paclitaxel. Br. J. Canc. 81, 416–422. doi: 10.1038/sj.bjc.6690710
Liu, C. M., Kao, C. L., Tseng, Y. T., Lo, Y. C., Chen, C. Y. (2017). Ginger phytochemicals inhibit cell growth and modulate drug resistance factors in docetaxel resistant prostate cancer cell. Molecules 22, 1477. doi: 10.3390/molecules22091477
Lu, J. J., Dang, Y. Y., Huang, M., Xu, W. S., Chen, X. P., Wang, Y. T. (2012). Anti-cancer properties of terpenoids isolated from Rhizoma curcumae – a review. J. Ethnopharmacol. 143, 406–411. doi: 10.1016/j.jep.2012.07.009
Lu, W. D., Qin, Y., Yang, C., Li, L. (2013). Effect of curcumin on human colon cancer multidrug resistance in vitro and in vivo. Clinics 68, 694–701. doi: 10.6061/clinics/2013(05)18
Ma, Y., Wink, M. (2008). Lobeline, a piperidine alkaloid from Lobelia can reverse P-gp dependent multidrug resistance in tumor cells. Phytomedicine 15, 754–758. doi: 10.1016/j.phymed.2007.11.028
Ma, Y., Wink, M. (2010). The beta-carboline alkaloid harmine inhibits BCRP and can reverse resistance to the anticancer drugs mitoxantrone and camptothecin in breast cancer cells. Phytother. Res. 24, 146–149. doi: 10.1002/ptr.2860
Ma, W., Feng, S., Yao, X., Yuan, Z., Liu, L., Xie, Y. (2015). Nobiletin enhances the efficacy of chemotherapeutic agents in ABCB1 overexpression cancer cells. Sci. Rep. 5, 18789. doi: 10.1038/srep18789
Martins, A., Vasas, A., Schelz, Z., Viveiros, M., Molnár, J., Hohmann, J., et al. (2010). Constituents of Carpobrotus edulis inhibit P-glycoprotein of MDR1-transfected mouse lymphoma cells. Anticancer Res. 30, 829–835.
Mayur, Y. C., Padma, T., Parimala, B. H., Chandramouli, K. H., Jagadeesh, S., Gowda, N. M., et al. (2006). Sensitization of multidrug resistant (MDR) cancer cells to vinblastine by novel acridones: correlation between anti-calmodulin activity and anti-MDR activity. Med. Chem. 2, 63–77. doi: 10.2174/157340606775197732
Mayur, Y. C. (2015). “Designing of drug molecules for reversing P-glycoprotein (Pgp) mediated drug resistance in cancer cells,” in Frontiers in Anti-Cancer Drug Discovery. Eds. Ata-ur-, R., Chaudhary, M. I. (Sharjah, UAE: Bentham Science Publishers), 157–198.
Mi, Q., Cui, B., Silva, G. L., Lantvit, D., Lim, E., Chai, H., et al. (2001). Pervilleine A, a novel tropane alkaloid that reverses the multidrug-resistance phenotype. Canc. Res. 61, 4030–4037.
Mi, Q., Cui, B., Silva, G. L., Lantvit, D., Lim, E., Chai, H., et al. (2002). Pervilleines B and C, new tropane alkaloid aromatic esters that reverse the multidrug-resistance in the hollow fiber assay. Canc. Lett. 184, 13–20. doi: 10.1016/S0304-3835(02)00202-1
Min, Y. D., Choi, S. U., Lee, K. R. (2006). Aporphine alkaloids and their reversal activity of multidrug resistance (MDR) from the stems and rhizomes of Sinomenium acutum. Arch. Pharm. Res. 29, 627–632. doi: 10.1007/BF02968246
Min, Y. D., Kwon, H. C., Yang, M. C., Lee, K. H., Choi, S. U., Lee, K. R. (2007). Isolation of limonoids and alkaloids from Phellodendron amurense and their multidrug resistance (MDR) reversal activity. Arch. Pharmacal Res. 30, 58–63. doi: 10.1007/BF02977779
Mitani, Y., Satake, K., Tsukamoto, M., Nakamura, I., Kadioglu, O., Teruya, T., et al. (2018). Epimagnolin A, a tetrahydrofurofuranoid lignan from Magnolia fargesii, reverses ABCB1-mediated drug resistance. Phytomedicine 51, 112–119. doi: 10.1016/j.phymed.2018.06.030
Mohanlal, R., Sun, Y., Kloecker, G., Feinstein, T., Shi, Y., Han, B., et al. (2019). P2. 01-23 DUBLIN-3, a phase (Ph) III trial comparing the plinabulin (P)/docetaxel (D) combination with D alone in stage IIIb/IV NSCLC. J. Thorac. Oncol. 14, S647–S648. doi: 10.1016/j.jtho.2019.08.1367
Molnár, J., Szabo, D., Pusztai, R., Mucsi, I., Berek, L., Ocsovszki, I., et al. (2000). Membrane associated antitumor effects of crocine-, ginsenoside-and cannabinoid derivates. Anticancer Res. 20, 861–867.
Molnár, J., Gyémánt, N., Tanaka, M., Hohmann, J., Bergmann-Leitner, E., Molnár, P., et al. (2006). Inhibition of multidrug resistance of cancer cells by natural diterpenes, triterpenes and carotenoids. Curr. Pharmaceut. Des. 12, 287–311. doi: 10.2174/138161206775201893
Molnár, J., Engi, H., Hohmann, J., Molnár, P., Deli, J., Wesolowska, O., et al. (2010). Reversal of multidrug resistance by natural substances from plants. Curr. Top. Med. Chem. 10, 1757–1768. doi: 10.2174/156802610792928103
Moussavi, M., Haddad, F., Rassouli, F. B., Iranshahi, M., Soleymanifard, S. (2017). Synergy between auraptene, ionizing radiation, and anticancer drugs in colon adenocarcinoma cells. Phytother. Res. 31, 1369–1375. doi: 10.1002/ptr.5863
Murahari, M., Kharkar, P. S., Lonikar, N., Mayur, Y. C. (2017). Design, synthesis, biological evaluation, molecular docking and QSAR studies of 2, 4-dimethylacridones as anticancer agents. Eur. J. Med. Chem. 130, 154–170. doi: 10.1016/j.ejmech.2017.02.022
Nabekura, T., Kamiyama, S., Kitagawa, S. (2005). Effects of dietary chemopreventive phytochemicals on P-glycoprotein function. Biochem. Biophys. Res. Commun. 327, 866–870. doi: 10.1016/j.bbrc.2004.12.081
Nabekura, T., Yamaki, T., Ueno, K., Kitagawa, S. (2008a). Inhibition of P-glycoprotein and multidrug resistance protein 1 by dietary phytochemicals. Canc. Chemother. Pharmacol. 62, 867–873. doi: 10.1007/s00280-007-0676-4
Nabekura, T., Yamaki, T., Kitagawa, S. (2008b). Effects of chemopreventive citrus phytochemicals on human P-glycoprotein and multidrug resistance protein 1. Eur. J. Pharmacol. 600, 45–49. doi: 10.1016/j.ejphar.2008.10.025
Nabekura, T., Yamaki, T., Ueno, K., Kitagawa, S. (2008c). Effects of plant sterols on human multidrug transporters ABCB1 and ABCC1. Biochem. Biophys. Res. Commun. 369, 363–368. doi: 10.1016/j.bbrc.2008.02.026
Nabekura, T., Yamaki, T., Hiroi, T., Ueno, K., Kitagawa, S. (2010). Inhibition of anticancer drug efflux transporter P-glycoprotein by rosemary phytochemicals. Pharmacol. Res. 61, 259–263. doi: 10.1016/j.phrs.2009.11.010
Najar, I. A., Sachin, B. S., Sharma, S. C., Satti, N. K., Suri, K. A., Johri, R. K. (2010). Modulation of P-glycoprotein ATPase activity by some phytoconstituents. Phytother. Res. 24, 454–458. doi: 10.1002/ptr.2951
Namanja, H. A., Emmert, D., Pires, M. M., Hrycyna, C. A., Chmielewski, J. (2009). Inhibition of human P-glycoprotein transport and substrate binding using a galantamine dimer. Biochem. Biophys. Res. Commun. 388, 672–676. doi: 10.1016/J.BBRC.2009.08.056
Neto, S., Duarte, N., Pedro, C., Spengler, G., Molnár, J., Ferreira, M. J. U. (2019). Effective MDR reversers through phytochemical study of Euphorbia boetica. Phytochem. Anal. 30, 498–511. doi: 10.1002/pca.2841
Nguyen, H., Zhang, S., Morris, M. E. (2003). Effect of flavonoids on MRP1-mediated transport in Panc-1 cells. J. Pharmaceut. Sci. 92, 250–257. doi: 10.1002/jps.10283
Nieri, P., Romiti, N., Adinolfi, B., Chicca, A., Massarelli, I., Chieli, E. (2006). Modulation of P-glycoprotein activity by cannabinoid molecules in HK-2 renal cells. Br. J. Pharmacol. 148, 682. doi: 10.1038/sj.bjp.0706778
O'Connor, P. M., Jackman, J., Bae, I., Myers, T. G., Fan, S., Mutoh, M., et al. (1997). Characterization of the p53 tumor suppressor pathway in cell lines of the National Cancer Institute anticancer drug screen and correlations with the growth-inhibitory potency of 123 anticancer agents. Canc. Res. 57, 4285–4300.
Ohtani, H., Ikegawa, T., Honda, Y., Kohyama, N., Morimoto, S., Shoyama, Y., et al. (2007). Effects of various methoxyflavones on vincristine uptake and multidrug resistance to vincristine in P-gp-overexpressing K562/ADM cells. Pharmaceut. Res. 24, 1936–1943. doi: 10.1007/s11095-007-9320-6
Osman, A. M. G., Chittiboyina, A. G., Khan, I. A. (2013). “Foodborne infections and intoxications: Chapter 32,” in Plant Toxins (San Diego, USA: Academic Press).
Panche, A. N., Diwan, A. D., Chandra, S. R. (2016). Flavonoids: an overview. J. Nutr. Sci. 5 (E47), 1–15. doi: 10.1017/jns.2016.41
Pearce, H. L., Safa, A. R., Bach, N. J., Winter, M. A., Cirtain, M. C., Beck, W. T. (1989). Essential features of the P-glycoprotein pharmacophore as defined by a series of reserpine analogs that modulate multidrug resistance. Proc. Natl. Acad. Sci. U.S.A. 86, 5128–5132. doi: 10.1073/PNAS.86.13.5128
Pires, M. M., Emmert, D., Hrycyna, C. A., Chmielewski, J. (2009). Inhibition of P-glycoprotein-mediated paclitaxel resistance by reversibly linked quinine homodimers. Mol. Pharmacol. 75, 92–100. doi: 10.1124/mol.108.050492
Reis, M. A., Ahmed, O. B., Spengler, G., Molnár, J., Lage, H., Ferreira, M. J. U. (2016). Jatrophane diterpenes and cancer multidrug resistance–ABCB1 efflux modulation and selective cell death induction. Phytomedicine 23, 968–978. doi: 10.1016/j.phymed.2016.05.007
Rethy, B., Hohmann, J., Minorics, R., Varga, A., Ocsovszki, I., Molnár, J., et al. (2008). Antitumour properties of acridone alkaloids on a murine lymphoma cell line. Anticancer Res. 28, 2737–2743.
Reyes-Esparza, J., Morales, A. I. G., González-Maya, L., Rodríguez-Fragoso, L. (2015). (-)-Epigallocatechin-3-gallate modulates the activity and expression of P-glycoprotein in breast cancer cells. J. Pharmacol. Clin. Toxicol. 3, 1044.
Roberts, M. F., Wink, M. (1998). Alkaloids: Biochemistry, Ecology, and Medicinal Applications (New York, USA: Plenum Press).
Ruenitz, P. C., Mokler, C. M. (1977). Analogs of sparteine. 5. antiarrhythmic activity of selected N, N'-disubstituted bispidines. J. Med. Chem. 20, 1668–1671. doi: 10.1021/jm00222a026
Saeed, M., Kadioglu, O., Khalid, H., Sugimoto, Y., Efferth, T. (2015). Activity of the dietary flavonoid, apigenin, against multidrug-resistant tumor cells as determined by pharmacogenomics and molecular docking. J. Nutr. Biochem. 26, 44–56. doi: 10.1016/j.jnutbio.2014.09.008
Sampson, K. E., Wolf, C. L., Abraham, I. (1993). Staurosporine reduces P-glycoprotein expression and modulates multidrug resistance. Canc. Lett. 68, 7–14. doi: 10.1016/0304-3835(93)90213-S
Sergent, T., Garsou, S., Schaut, A., De Saeger, S., Pussemier, L., Van Peteghem, C., et al. (2005). Differential modulation of ochratoxin A absorption across Caco-2 cells by dietary polyphenols, used at realistic intestinal concentrations. Toxicol. Lett. 159, 60–70. doi: 10.1016/j.toxlet.2005.04.013
Shen, X., Chen, G., Zhu, G., Fong, W. F. (2006). (±)-3′-O, 4′-O-dicynnamoyl-cis-khellactone, a derivative of (±)-praeruptorin A, reverses P-glycoprotein mediated multidrug resistance in cancer cells. Bioorg. Med. Chem. 14, 7138–7145. doi: 10.1016/j.bmc.2006.06.066
Shen, H., Xu, W., Chen, Q., Wu, Z., Tang, H., Wang, F. (2010). Tetrandrine prevents acquired drug resistance of K562 cells through inhibition of MDR1 gene transcription. J. Canc. Res. Clin. Oncol. 136, 659–665. doi: 10.1007/s00432-009-0704-3
Sheu, M. T., Liou, Y. B., Kao, Y. H., Lin, Y. K., Ho, H. O. (2010). A quantitative structure–activity relationship for the modulation effects of flavonoids on P-glycoprotein-mediated transport. Chem. Pharmaceut. Bull. 58, 1187–1194. doi: 10.1248/cpb.58.1187
Simstein, R., Burow, M., Parker, A., Weldon, C., Beckman, B. (2003). Apoptosis, chemoresistance, and breast cancer: insights from the MCF-7 cell model system. Exp. Biol. Med. 228, 995–1003. doi: 10.1177/153537020322800903
Slaninová, I., Březinová, L., Koubíková, L., Slanina, J. (2009). Dibenzocyclooctadiene lignans overcome drug resistance in lung cancer cells–study of structure–activity relationship. Toxicol. Vitro 23, 1047–1054. doi: 10.1016/j.tiv.2009.06.008
Smith, E. W., Maibach, H. I. (1995). Percutaneous Penetration Enhancers (Boca Raton, USA: CRC Press).
Smith, E. C., Kaatz, G. W., Seo, S. M., Wareham, N., Williamson, E. M., Gibbons, S. (2007). The phenolic diterpene totarol inhibits multidrug efflux pump activity in Staphylococcus aureus. Antimicrob. Agents Chemother. 51, 4480–4483. doi: 10.1128/AAC.00216-07
Stermitz, F. R., Scriven, L. N., Tegos, G., Lewis, K. (2002). Two flavonols from Artemisa annua which potentiate the activity of berberine and norfloxacin against a resistant strain of Staphylococcus aureus. Planta Med. 68, 1140–1141. doi: 10.1055/s-2002-36347
Su, S., Cheng, X., Wink, M. (2015). Natural lignans from Arctium lappa modulate P-glycoprotein efflux function in multidrug resistant cancer cells. Phytomedicine 22, 301–307. doi: 10.1016/J.PHYMED.2014.12.009
Sun, Y. F., Wink, M. (2014). Tetrandrine and fangchinoline, bisbenzylisoquinoline alkaloids from Stephania tetrandra can reverse multidrug resistance by inhibiting P-glycoprotein activity in multidrug resistant human cancer cells. Phytomedicine 21, 1110–1119. doi: 10.1016/J.PHYMED.2014.04.029
Sun, H. X., Xie, Y., Ye, Y. P. (2009). Advances in saponin-based adjuvants. Vaccine 27, 1787–1796. doi: 10.1016/j.vaccine.2009.01.091
Sun, J., Yeung, C. A., Tsang, T. Y., Yau, E., Luo, K., Wu, P., et al. (2012). Clitocine reversal of P-glycoprotein associated multi-drug resistance through down-regulation of transcription factor NF-κB in R-HepG2 cell line. PloS One 7, e40720. doi: 10.1371/journal.pone.0040720
Suzuki, T., Fukazawa, N., San-nohe, K., Sato, W., Yano, O., Tsuruo, T. (1997). Structure– activity relationship of newly synthesized quinoline derivatives for reversal of multidrug resistance in cancer. J. Med. Chem. 40, 2047–2052. doi: 10.1021/jm960869l
Suzuki, H., Tanabe, H., Mizukami, H., Inoue, M. (2010). Selective regulation of multidrug resistance protein in vascular smooth muscle cells by the isoquinoline alkaloid coptisine. Biol. Pharm. Bull. 33, 677–682. doi: 10.1248/bpb.33.677
Syed, S. B., Coumar, M. S. (2016). P-Glycoprotein mediated multidrug resistance reversal by phytochemicals: a review of SAR & future perspective for drug design. Curr. Top. Med. Chem. 16, 2484–2508. doi: 10.2174/1568026616666160212123814
Syed, S. B., Arya, H., Fu, I. H., Yeh, T. K., Periyasamy, L., Hsieh, H. P., et al. (2017). Targeting P-glycoprotein: Investigation of piperine analogs for overcoming drug resistance in cancer. Sci. Rep. 7, 1–18. doi: 10.1038/s41598-017-08062-2
Tanaka, K., Iwamoto, S., Gon, G., Nohara, T., Iwamoto, M., Tanigawa, N. (2000). Expression of survivin and its relationship to loss of apoptosis in breast carcinomas. Clin. Canc. Res. 6, 127–134.
Tang, X., Bi, H., Feng, J., Cao, J. (2005). Effect of curcumin on multidrug resistance in resistant human gastric carcinoma cell line SGC7901/VCR. Acta Pharmacol. Sin. 26, 1009–1016. doi: 10.1111/j.1745-7254.2005.00149.x
Teng, Y. N., Sheu, M. J., Hsieh, Y. W., Wang, R. Y., Chiang, Y. C., Hung, C. C. (2016). β-carotene reverses multidrug resistant cancer cells by selectively modulating human P-glycoprotein function. Phytomedicine 23, 316–323. doi: 10.1016/j.phymed.2016.01.008
Tiwary, B. K., Pradhan, K., Nanda, A. K., Chakraborty, R. (2015). Implication of quinazoline-4(3H)-ones in medicinal chemistry: a brief review. J. Chem. Biol. Ther. 1, 104. doi: 10.4172/2572-0406.1000104
To, K. K., Wu, X., Yin, C., Chai, S., Yao, S., Kadioglu, O. (2017). Reversal of multidrug resistance by Marsdenia tenacissima and its main active ingredients polyoxypregnanes. J. Ethnopharmacol. 203, 110–119. doi: 10.1016/j.jep.2017.03.051
Trock, B. J., Leonessa, F., Clarke, R. (1997). Multidrug resistance in breast cancer: A Meta-analysis of MDR1/gp170 expression and its possible functional significance. J. Natl. Canc. Inst. 89, 917–931. doi: 10.1093/jnci/89.13.917
Um, Y., Cho, S., Woo, H. B., Kim, Y. K., Kim, H., Ham, J., et al. (2008). Synthesis of curcumin mimics with multidrug resistance reversal activities. Bioorg. Med. Chem. 16, 3608–3615. doi: 10.1016/j.bmc.2008.02.012
Umsumarng, S., Pitchakarn, P., Yodkeeree, S., Punfa, W., Mapoung, S., Ramli, R. A. (2017). Modulation of P-glycoprotein by Stemona alkaloids in human multidrug resistance leukemic cells and structural relationships. Phytomedicine 34, 182–190. doi: 10.1016/j.phymed.2017.08.004
van Zanden, J. J., Wortelboer, H. M., Bijlsma, S., Punt, A., Usta, M., Bladeren, v., et al. (2005). Quantitative structure activity relationship studies on the flavonoid mediated inhibition of multidrug resistance proteins 1 and 2. Biochem. Pharmacol. 69, 699–708. doi: 10.1016/j.bcp.2004.11.002
Vasas, A., Sulyok, E., Rédei, D., Forgo, P., Szabó, P., Zupkó, I., et al. (2011). Jatrophane diterpenes from Euphorbia esula as antiproliferative agents and potent chemosensitizers to overcome multidrug resistance. J. Nat. Prod. 74, 1453–1461. doi: 10.1021/np200202h
Wan, C. K., Zhu, G. Y., Shen, X. L., Chattopadhyay, A., Dey, S., Fong, W. F. (2006). Gomisin A alters substrate interaction and reverses P-glycoprotein-mediated multidrug resistance in HepG2-DR cells. Biochem. Pharmacol. 72, 824–837. doi: 10.1016/j.bcp.2006.06.036
Wandel, C., Kim, R. B., Kajiji, S., Guengerich, F. P., Wilkinson, G. R., Wood, A. J. (1999). P-glycoprotein and cytochrome P-450 3A inhibition: dissociation of inhibitory potencies. Canc. Res. 59, 3944–3948.
Wang, Y., Cabral, F. (2005). Paclitaxel resistance in cells with reduced β-tubulin. Biochim. Biophys. Acta Mol. Cell Res. 1744, 245–255. doi: 10.1016/j.bbamcr.2004.12.003
Wang, E. J., Barecki-Roach, M., Johnson, W. W. (2002). Elevation of P-glycoprotein function by a catechin in green tea. Biochem. Biophys. Res. Commun. 297, 412–418. doi: 10.1016/S0006-291X(02)02219-2
Wang, C., Zhang, J. X., Shen, X. L., Wan, C. K., Tse, A. K. W., Fong, W. F. (2004). Reversal of P-glycoprotein-mediated multidrug resistance by Alisol B 23-acetate. Biochem. Pharmacol. 68, 843–855. doi: 10.1016/j.bcp.2004.05.021
Wang, X. B., Wang, S. S., Zhang, Q. F., Liu, M., Li, H. L., Liu, Y., et al. (2010). Inhibition of tetramethylpyrazine on P-gp, MRP2, MRP3 and MRP5 in multidrug resistant human hepatocellular carcinoma cells. Oncol. Rep. 23, 211–215. doi: 10.3892/or_00000625
Wang, S., Lei, T., Zhang, M. (2016). The reversal effect and its mechanisms of tetramethylpyrazine on multidrug resistance in human bladder cancer. PloS One 11, e0157759. doi: 10.1371/journal.pone.0157759
Wang, X., Zhang, H., Chen, X. (2019). Drug resistance and combating drug resistance in cancer. Canc. Drug Resist. 2, 141–160. doi: 10.20517/cdr.2019.10
Wesołowska, O., Hendrich, A. B., Łania-Pietrzak, B., Wiśniewski, J., Molnár, J., Ocsovszki, I., et al. (2009). Perturbation of the lipid phase of a membrane is not involved in the modulation of MRP1 transport activity by flavonoids. Cell. Mol. Biol. Lett. 14, 199. doi: 10.2478/s11658-008-0044-3
Wink, M., van Wyk, B. E. (2008). Mind-Altering and Poisonous Plants of the World (Portland, USA: Timber Press).
Wink, M., Ashour, M. L., El-Readi, M. Z. (2012). Secondary metabolites inhibiting ABC transporters and reversing resistance of cancer cells and fungi to cytotoxic and antimicrobial agents. Front. Microbiol. 3, 130. doi: 10.3389/fmicb.2012.00130
Wink, M. (2000). “Interference of alkaloids with neuroreceptors and ion channels,” in Studies in Natural Products Chemistry. Ed. Ata-ur-, R. (Amsterdam, Netherlands: Elsevier), 3–122.
Wink, M. (2007). “Molecular modes of action of cytotoxic alkaloids: from DNA intercalation, spindle poisoning, topoisomerase inhibition to apoptosis and multiple drug resistance,” in The Alkaloids: Chemistry and Biology, vol. 64. (San Diego, USA: Academic Press), 1–47.
Wink, M. (2015). Modes of action of herbal medicines and plant secondary metabolites. Medicines 2, 251–286. doi: 10.3390/medicines2030251
Wink, M. (2020). Evolution of the angiosperms and co-evolution of secondary metabolites, especially of Alkaloids. Reference Series in Phytochemistry. Co-Evolution of Secondary Metabolites (Cham, Switzerland: Springer Nature).
Wong, I. L., Wang, B. C., Yuan, J., Duan, L. X., Liu, Z., Liu, T., et al. (2015). Potent and nontoxic chemosensitizer of P-glycoprotein-mediated multidrug resistance in cancer: synthesis and evaluation of methylated epigallocatechin, gallocatechin, and dihydromyricetin derivatives. J. Med. Chem. 58, 4529–4549. doi: 10.1021/acs.jmedchem.5b00085
Wongrattanakamon, P., Lee, V. S., Nimmanpipug, P., Sirithunyalug, B., Chansakaow, S., Jiranusornkul, S. (2017). Insight into the molecular mechanism of p-glycoprotein mediated drug toxicity induced by bioflavonoids: An integrated computational approach. Toxicol. Mech. Meth. 27, 253–271. doi: 10.1080/15376516.2016.1273428
Wu, J. Y. C., Fong, W. F., Zhang, J. X., Leung, C. H., Kwong, H. L., Yang, M. S., et al. (2003). Reversal of multidrug resistance in cancer cells by pyranocoumarins isolated from Radix peucedani. Eur. J. Pharmacol. 473, 9–17. doi: 10.1016/S0014-2999(03)01946-0
Xu, D., Lu, Q., Hu, X. (2006). Down-regulation of P-glycoprotein expression in MDR breast cancer cell MCF-7/ADR by honokiol. Canc. Lett. 243, 274–280. doi: 10.1016/j.canlet.2005.11.031
Ye, J., Zheng, Y., Liu, D. (2009). Reversal effect and its mechanism of ampelopsin on multidrug resistance in K562/ADR cells. China J. Chin. Materia Med. 34, 761–765.
Yoo, H. H., Lee, M., Chung, H. J., Lee, S. K., Kim, D. H. (2007a). Effects of diosmin, a flavonoid glycoside in citrus fruits, on P-glycoprotein-mediated drug efflux in human intestinal Caco-2 cells. J. Agr. Food Chem. 55, 7620–7625. doi: 10.1021/jf070893f
Yoo, H. H., Lee, M., Lee, M. W., Lim, S. Y., Shin, J., Kim, D. H. (2007b). Effects of Schisandra lignans on P-glycoprotein-mediated drug efflux in human intestinal Caco-2 cells. Planta Med. 53, 444–450. doi: 10.1055/s-2007-967178
Yoshida, N., Takagi, A., Kitazawa, H., Kawakami, J., Adachi, I. (2005). Inhibition of P-glycoprotein-mediated transport by extracts of and monoterpenoids contained in Zanthoxyli fructus. Toxicol. Appl. Pharmacol. 209, 167–173. doi: 10.1016/j.taap.2005.04.001
You, M., Ma, X., Mukherjee, R., Farnsworth, N. R., Cordell, G. A., Kinghorn, A. D., et al. (1994). Indole alkaloids from Peschiera laeta that enhance vinblastine-mediated cytotoxicity with multidrug-resistant cells. J. Nat. Prod. 57, 1517–1522. doi: 10.1021/np50113a007
You, M., Wickramaratne, D. M., Silva, G. L., Chai, H., Chagwedera, T. E., Farnsworth, N. R., et al. (1995). (-)-Roemerine, an aporphine alkaloid from Annona senegalensis that reverses the multidrug-resistance phenotype with cultured cells. J. Nat. Prod. 58, 598–604. doi: 10.1021/np50118a021
Zhang, S., Morris, M. E. (2003). Effects of the flavonoids biochanin A, morin, phloretin, and silymarin on P-glycoprotein-mediated transport. J. Pharmacol. Exp. Therapeut. 304, 1258–1267. doi: 10.1124/jpet.102.044412
Zhang, C. C., Yang, J. M., White, E., Murphy, M., Levine, A., Hait, W. N. (1998). The role of MAP4 expression in the sensitivity to paclitaxel and resistance to vinca alkaloids in p53 mutant cells. Oncogene 16, 1617–1624. doi: 10.1038/sj.onc.1201658
Zhang, S., Yang, X., Morris, M. E. (2004). Combined effects of multiple flavonoids on breast cancer resistance protein (ABCG2)-mediated transport. Pharm. Res. 21, 1263–1273. doi: 10.1023/B:PHAM.0000033015.84146.4c
Zhang, S., Wang, X., Sagawa, K., Morris, M. E. (2005). Flavonoids chrysin and benzoflavone, potent breast cancer resistance protein inhibitors, have no significant effect on topotecan pharmacokinetics in rats or mdr1a/1b (–/–) mice. Drug Metabol. Dispos. 33, 341–348. doi: 10.1124/dmd.104.002501
Zhang, Y., Liu, X., Zuo, T., Liu, Y., Zhang, J. H. (2012). Tetramethylpyrazine reverses multidrug resistance in breast cancer cells through regulating the expression and function of P-glycoprotein. Med. Oncol. 29, 534–538. doi: 10.1007/s12032-011-9950-8
Zhou, N., Li, J., Li, T., Chen, G., Zhang, Z., Si, Z. (2017). Matrine-induced apoptosis in Hep3B cells via the inhibition of MDM2. Mol. Med. Rep. 15, 442–450. doi: 10.3892/mmr.2016.5999
Zhu, X., Zhang, X., Ma, G., Yan, J., Wang, H., Yang, Q. (2011). Transport characteristics of tryptanthrin and its inhibitory effect on P-gp and MRP2 in Caco-2 cells. J. Pharm. Pharmaceut. Sci. 14, 325–335. doi: 10.18433/J3501W
Zhu, J., Wang, R., Lou, L., Li, W., Tang, G., Bu, X., et al. (2016). Jatrophane diterpenoids as modulators of P-glycoprotein-dependent multidrug resistance (MDR): advances of structure–activity relationships and discovery of promising MDR reversal agents. J. Med. Chem. 59, 6353–6369. doi: 10.1021/acs.jmedchem.6b00605
Keywords: cancer, ABC-transporter, drug efflux, multidrug resistance, secondary metabolites, synergism
Citation: Tinoush B, Shirdel I and Wink M (2020) Phytochemicals: Potential Lead Molecules for MDR Reversal. Front. Pharmacol. 11:832. doi: 10.3389/fphar.2020.00832
Received: 20 December 2019; Accepted: 20 May 2020;
Published: 19 June 2020.
Edited by:
Maria José U. Ferreira, University of Lisbon, PortugalReviewed by:
Olga Wesolowska, Wroclaw Medical University, PolandCopyright © 2020 Tinoush, Shirdel and Wink. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Michael Wink, d2lua0B1bmktaGVpZGVsYmVyZy5kZQ==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.