 Jing Ye1†Yuan Wang2†
Jing Ye1†Yuan Wang2† Zhen Wang1†Ling Liu3Zicong Yang3Menglong Wang1Yao Xu1Di Ye1Jishou Zhang1
Zhen Wang1†Ling Liu3Zicong Yang3Menglong Wang1Yao Xu1Di Ye1Jishou Zhang1 Yingzhong Lin3*Qingwei Ji3*
Yingzhong Lin3*Qingwei Ji3* Jun Wan1*
Jun Wan1*- 1Hubei Key Laboratory of Cardiology, Department of Cardiology, Renmin Hospital of Wuhan University, Cardiovascular Research Institute, Wuhan University, Wuhan, China
- 2Department of Thyroid Breast Surgery, Renmin Hospital of Wuhan University, Wuhan, China
- 3Department of Cardiology, the People's Hospital of Guangxi Zhuang Autonomous Region, Nanning, China
Cardiovascular diseases represent a complex group of clinical syndromes caused by a variety of interacting pathological factors. They include the most extensive disease population and rank first in all-cause mortality worldwide. Accumulating evidence demonstrates that cytokines play critical roles in the presence and development of cardiovascular diseases. Interleukin-12 family members, including IL-12, IL-23, IL-27 and IL-35, are a class of cytokines that regulate a variety of biological effects; they are closely related to the progression of various cardiovascular diseases, including atherosclerosis, hypertension, aortic dissection, cardiac hypertrophy, myocardial infarction, and acute cardiac injury. This paper mainly discusses the role of IL-12 family members in cardiovascular diseases, and the molecular and cellular mechanisms potentially involved in their action in order to identify possible intervention targets for the prevention and clinical treatment of cardiovascular diseases.
Introduction
To date, cardiovascular disease remains the leading killer worldwide, especially in less-developed areas (Leong et al., 2017). It is not only a serious threat to patients' lives, but also poses a serious psychological burden to patients and their families. Although a large number of useful drugs and new technologies have been widely used in clinical treatment over recent years and have significantly improved survival rates, the overall prognosis of cardiovascular diseases is still very poor, and the death rate related to cardiovascular diseases is far higher than that of other diseases, even malignant tumors (Donofrio et al., 2014; Dukkipati et al., 2017; Bethel et al., 2018).
There are four members of the interleukin-12 (IL-12) family, including IL-12, IL-23, IL-27, and IL-35. An obvious feature of IL-12 family members is that each consists of two heterogeneous dimers, including an α subunit (p19, p28, and p35) and a β subunit [p40 and Epstein-Barr virus-induced protein 3 (EBI3)] (Vignali and Kuchroo, 2012; Sun et al., 2015). Therefore, deletion of either an α or β subunit can cancel the biological effects of the IL-12 family cytokines. Interestingly, the receptor for IL-12 family members also consists of two protein chains. Among them, the IL-12 receptor (IL-12R) utilizes IL-12Rβ1 and IL-12Rβ2, IL-23 signaling employs IL-12Rβ1 and IL-23R, and IL-27 signals through gp130 and WSX-1; however, an exception is IL-35R, which consists of two downstream signals, including gp130-gp130 or IL-12Rβ1-IL-12Rβ1 (Presky et al., 1996; Oppmann et al., 2000; Pflanz et al., 2004; Collison et al., 2012). Molecular signaling mechanisms involving IL-12 family members are basically similar, and are all mediated by members of the Janus kinase (JAK) signal transducers and activators of transcription (STAT) family, especially JAK1/2-STAT1/3/4 (Ihle, 1995; O'Shea et al., 2002; Delgoffe et al., 2011). All IL-12 family members can be secreted by both immune and non-immune cells (Vignali and Kuchroo, 2012; Sun et al., 2015). For the immune cells, IL-12, IL-23, and IL-27 are mainly derived from effector T lymphocytes, macrophages, and dendritic cells, while IL-35 is mainly secreted by T helper cells (Tregs) (Langrish et al., 2004; Hunter, 2005; Collison et al., 2009; Collison et al., 2010; Andrews et al., 2016; Wei et al., 2017). IL-12 and IL-23 are considered to be pro-inflammatory factors that amplify downstream inflammatory signals. IL-35 plays an anti-inflammatory role and protects against tissue damage mediated by inflammatory responses, while IL-27 has a two-sided effect on the regulation of inflammation, in which it can not only play an anti-inflammatory role, but also a pro-inflammatory role, depending on the inflammatory environment (Ma and Trinchieri, 2001; Kastelein et al., 2007; Collison and Vignali, 2008; Vignali et al., 2008; Cox et al., 2011; Vignali and Kuchroo, 2012; Wojno and Hunter, 2012; Sun et al., 2015). The subunits, receptors, signaling pathways, and regulatory roles of the members of the IL-12 family in inflammation are listed in Table 1.
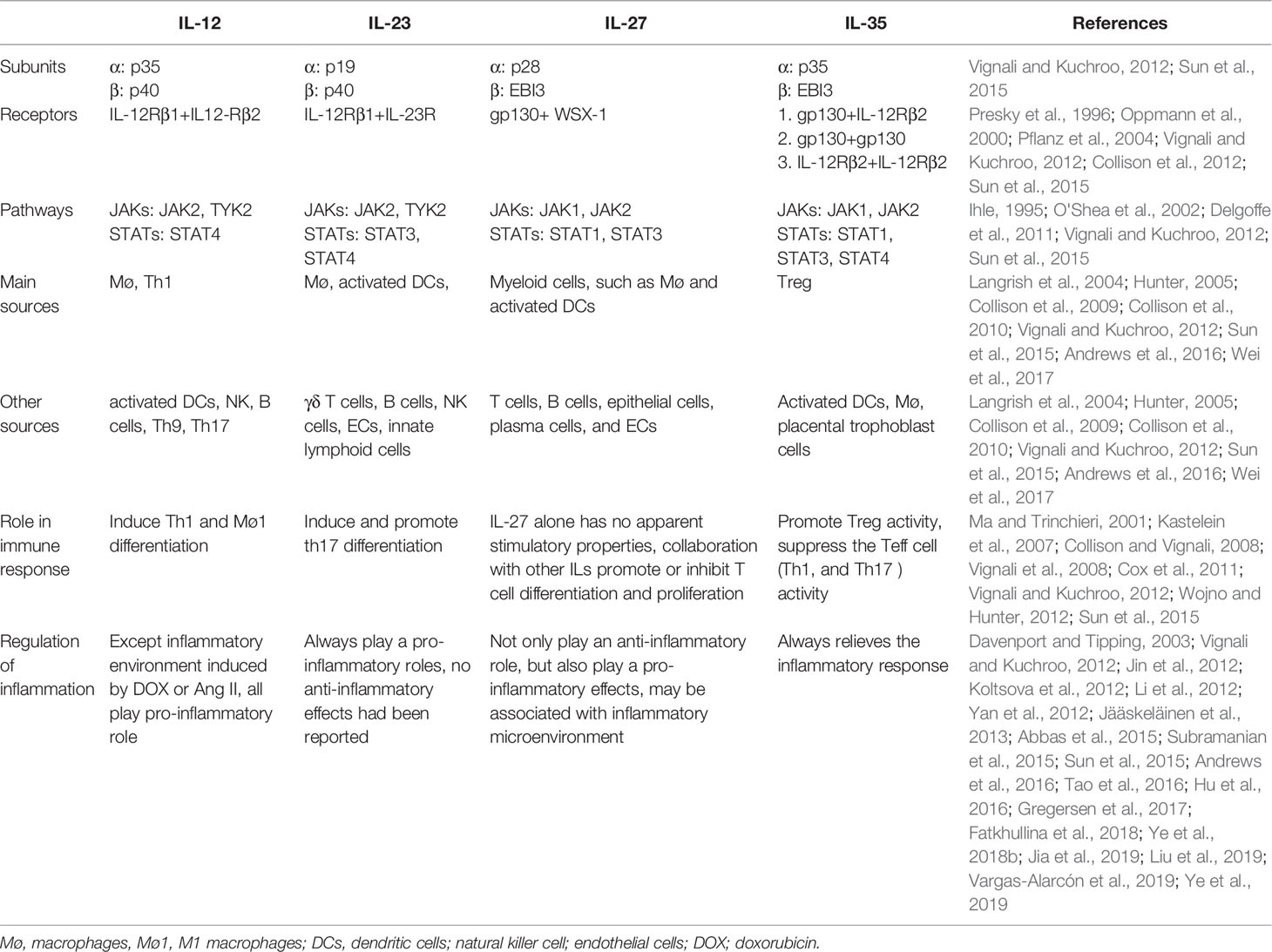
Table 1 The subunits, receptors, signaling pathways of the IL-12 family members.
Interleukin-12 Family Members and Cardiovascular Disease
Interleukin-12 Family Members and Atherosclerosis, Coronary Artery Disease
Atherosclerosis and coronary artery diseases due to atherosclerosis are chronic inflammatory disorders, and infiltration by immune cells and inflammatory factors can be observed at all stages of disease development (Peter et al., 2009; Longenecker et al., 2016; Rahman and Fisher., 2018). IL-12 family members have significantly higher levels of expression in patients with atherosclerosis and coronary artery disease, and are closely related to the progression of these diseases.
Clinical Data
Previous studies reported that plasma IL-12 concentrations are significantly increased in many types of atherosclerosis and atherosclerotic cardiovascular disease, including stable angina pectoris (SAP), non-ST segment elevation myocardial infarction (NSTEMI), ST-elevation myocardial infarction (STEMI), acute myocardial infarction (AMI), and gradually increased SAP, unstable angina pectoris (UAP), and AMI (Zhou et al., 2001; Correia et al., 2010; Lin et al., 2012; Yong et al., 2013; Chistiakov et al., 2015; Opstad et al., 2016; Zykov et al., 2016). Clinical data showed that patients with coronary artery disease exhibit higher circulating IL-23 levels (Lin et al., 2012; Abbas et al., 2015; Sun et al., 2019a). In coronary artery disease patients who underwent percutaneous coronary intervention (PCI) with drug-eluting stents (DES), subjects with in-stent restenosis show higher circulating IL-23 levels in peripheral blood mononuclear cells (PBMCs) (Khojasteh-Fard et al., 2012). Numerous studies have confirmed that IL-27 expression is increased in plasma and plaques in the coronary and carotid arteries of coronary artery disease patients (Kempe et al., 2009; Jin et al., 2012; Lin et al., 2012; A Shahi et al., 2015; Gregersen et al., 2017). Abundant evidence identifies that IL-35 expression is significantly reduced in patients with coronary artery disease; plasma IL-35 levels are gradually reduced in SAP, UAP, and AMI patients, and decreased plasma IL-35 levels are inversely correlated with the left ventricular ejection fraction (LVEF) in coronary artery diseases (Lin et al., 2012; Rasa et al., 2018; Zhu et al., 2018).
Gene polymorphisms in members of the IL-12 family have been reported to be associated with the occurrence or progression of coronary artery disease. An IL-23R polymorphism was observed to be related to coronary artery disease, and the IL-23R rs6682925T/C polymorphism may independently relate to the presence of coronary artery disease (Zhang et al., 2014a). IL-27 gene polymorphism had no effect on the presence of subclinical atherosclerosis, while closely related to atherosclerosis and coronary artery disease, rs26528 T and rs40837 A alleles significantly reduced the risk of coronary artery disease (Posadas-Sánchez et al., 2017; Vargas-Alarcón et al., 2019). No research about gene polymorphisms of IL-12 and IL-35 and the presence of coronary artery disease was reported yet.
Animal Studies
Elevated serum IL-12 levels are observed of atherosclerosis in ApoE-KO mice, and increased IL-12 levels are associated with the progression of atherosclerosis (Jääskeläinen et al., 2013). Accumulating animal study reports also demonstrate that treatment with exogenous recombinant murine IL-12 significantly aggravates the progression of atherosclerosis, and increases aortic atherosclerotic plaque areas in both ApoE-knockout mice and in low density lipoprotein (LDL) receptor-deficient mice, while cancelation the biological effects of IL-12 can significantly diminish such effects (Lee et al., 1999; Davenport and Tipping, 2003; Hauer et al., 2005). In a murine myocardial infarction model, canceling the biological effects of IL-12 alleviates cardiac dysfunction by promoting angiogenesis (Kan et al., 2016). In a recent study, Shi et al. reported that knockout of IL-12p35 subunit, which can cancel the biological effects of IL-12 and IL-35, significantly aggravated Th1/Th2 and Th17/Treg imbalance and increased atherosclerotic plaque areas in ApoE mice, which may suggest that the pro-atherosclerotic effects of IL-12 can be mediated by promoting the CD4+ T lymphocyte differentiation imbalance (Huang et al., 2019).
The role of IL-23 in atherosclerosis is controversial. Therapy involving IL-23p19, a subunit of IL-23, had no significant effect on atherosclerosis development in ApoE-deficient mice, although inflammatory responses were reduced (Wang et al., 2019). Another study reported that there was no significant difference in atherosclerotic area between low-density lipoprotein receptor (LDLR) knockout mice and IL-23 + LDLR double-knockout mice, after they were all fed with high-fat diet (Engelbertsen et al., 2018). A recent study reported that deficiency of IL-23 significantly decreased IL-22 expression in ApoE-knockout mice, and also reduced expression of IL-22, thereby relieving the release of inflammatory substances, and thus alleviating the process of atherosclerosis (Fatkhullina et al., 2018). Subramanian et al. reported that granulocyte-macrophage colony stimulating factor (GM-CSF) up-regulates the expression of IL-23, which further promotes the differentiation of macrophages and atherosclerosis development (Subramanian et al., 2015). These studies suggest that IL-23 has a strong regulatory effect on inflammation mediated by a high-fat diet in both ApoE-knockout mice and LDL-R-knockout mice (Subramanian et al., 2015; Engelbertsen et al., 2018; Fatkhullina et al., 2018; Wang et al., 2019), while the special role of IL-23 in atherosclerotic progression is unclear and further studies are needed to clarify this aspect.
Both the effects of IL-27R and IL-27 on atherosclerosis were reported. Koltsova et al. found that knockout of IL-27R significantly enhanced Th17 immune responses, up-regulated inflammatory responses, promoted the expression of tumor necrosis factor (TNF) and IL-17A, and further promoted the development of atherosclerosis in ApoE-deficient mice (Koltsova et al., 2012). Hirase et al. also reported that knockout of IL-27 plays similar roles in atherosclerosis development in LDLR-knockout mice; the mechanism may be related to the promotion of macrophage differentiation (Hirase et al., 2013). Ryu et al. found that in a high-fat diet-treated ApoE-knockout mouse atherosclerotic model, blockade of IL-27 signaling increased the plaque area via promotion of autoimmune follicular helper T cell responses (Ryu et al., 2018). These results suggest that IL-27 may be an important target for the treatment and prevention of atherosclerosis and coronary artery disease by inhibiting the differentiation of various immune cells and reducing inflammatory responses, thereby alleviating atherosclerotic progression. Hence, IL-27 may be an important target for the treatment and prevention of atherosclerosis and coronary artery disease.
Contrary to clinical experiments, as an anti-inflammatory cytokine, IL-35 expression in mouse atherosclerotic plaques was significantly increased (Wang et al., 2014). A small number of other studies, however, have reported increased IL-35 expression in atherosclerotic plaques and serum in ApoE mice fed with a high-fat diet, as well as in the plasma of patients with coronary artery disease (Gorzelak-Pabiś et al., 2017; Li et al., 2018). Using a mouse model of atherosclerosis, administration of recombinant mouse IL-35 significantly decreased plaque area in the aortic root, and Treg immune responses were also found to be enhanced (Tao et al., 2016). In a recent study, Shi et al. reported that knockout of IL-12p35 subunit, which can cancel the biological effects of IL-12 and IL-35, significantly aggravated Th1/Th2 and Th17/Treg imbalance and increased atherosclerotic plaque areas in ApoE mice, given that mouse IL-35 reverses Th35/Treg imbalance and up-regulates atherosclerosis development, whereas there were no effects on Th1/Th2 imbalance (Huang et al., 2019). In a recently published study, two subunits of IL-35 found in the left anterior descending branch following its ligation induced myocardial infarction in the heart tissues in mice. In addition, exogenous IL-35 treatment can significantly reduce infarct area of the left ventricle and reduce the incidence of left ventricular rupture; the mechanisms underlying this phenomenon may be related to the inhibitory role of IL-35 in the apoptosis of myocardial macrophages, thus increasing the differentiation of M2 macrophages and augmenting the expression of collagen (Jia et al., 2019). In rat models of coronary artery disease, IL-35 treatment significantly promotes early drug-eluting stent endothelialization; its mechanism may be related to the regulation of the activation of M2 macrophages (Liu et al., 2019). These studies have demonstrated that IL-35 regulates the differentiation of various immune cells involved in the progression of atherosclerotic heart disease.
Interleukin-12 Family Members and Hypertension
Hypertension is a complex group of clinical syndromes. Although the specific mechanisms remain unclear, it has been demonstrated that a variety of pathological factors are involved in the process of hypertension, among which immune responses and inflammation are most closely related to hypertension (Kirabo et al., 2014; Pober, 2014; Guzik and Touyz, 2017).
So far, little research has been conducted on IL-12 family members and hypertension. Data from clinical experiments reported that plasma IL-12 levels are significantly increased in hypertensive patients, and are positively correlated with both systolic blood pressure (SBP) and diastolic blood pressure (DBP) (Ye et al., 2019). IL-12 polymorphism is closely related to the incidence of hypertension-induced complications: hypertension patients who carry the IL12B 1159 A/A genotype exhibit a lower risk of incidence of stroke, while IL12B A/A carriers have an elevated risk of stroke (Timasheva et al., 2008).
In an animal study, angiotensin II (Ang II) infusion significantly increased aortic IL-12p35 expression and macrophages were the primary source (Ye et al., 2019). In an Ang II-induced mouse hypertension model, IL-12p35 knockout promoted M1 macrophage differentiation and elevated blood pressure, while IL-12 treatment unexpectedly lowered blood pressure (Ye et al., 2019). Another study reported that knockdown of IL-12p35 did not affect Ang II-induced hypertension (Li et al., 2012). One possible reason for this is that the IL-12p35 knockout mice in that study were treated for only a week, which is too short a period for blood pressure to change. Little research has been conducted regarding IL-23, IL-27, and IL-35 in relation to hypertension. One group reported that in deoxycorticosterone acetate and Ang II-treated mice, deficiency of IL-17 could decrease IL-23 expression and accelerate kidney injury (Krebs et al., 2014), and another study found that treatment with recombinant mouse IL-35 had no effects on blood pressure in Ang II-treated mice (Ye et al., 2019).
Interleukin-12 Family Members and Aortic Aneurysms and Aortic Dissection
Aortic aneurysms and aortic dissection are both degenerative lesions of the aorta and share the same pathological mechanisms, such as the excessive loss of aortic extracellular matrix mediated by multiple pathological factors, especially local aortic inflammation (Mallat et al., 2016; Rabkin, 2017; Raffort et al., 2017; Sherifova and Holzapfel, 2019).
There have been few studies regarding IL-12 family members, aortic aneurysms, and aortic dissection. Davis et al. reported that IL-12 levels in aortic tissue and serum were not significantly different in patients with abdominal aortic aneurysms compared to those in patients without abdominal aortic aneurysms (Davis et al., 2001). In aortic dissection patients, decreased plasma IL-35 concentrations were observed compared to non-aortic dissection patients (Ye et al., 2018a). In addition, no studies have been conducted on IL-12 family members and aortic aneurysms and aortic dissection.
Only one recent animal study reported that deletion of IL-27R reduced the formation of abdominal aortic aneurysm in ApoE deficiency mice, the mechanisms may be associated with a blunted accumulation of myeloid cells in the aorta (Peshkova et al., 2019).
Interleukin-12 Family Members and Cardiac Fibrosis
Cardiac fibrosis is a common feature of many heart diseases and is closely related to deterioration in cardiac function. The essence of cardiac fibrosis is that pathological factors activate cardiac fibroblasts, leading to abnormal deposition and increased numbers of cardiac collagen fibers (Moore-Morris et al., 2015; Gorabi et al., 2019).
All studies on IL-12 family members and cardiac fibrosis have been focused on animal studies and no related clinical studies have been reported. In an earlier report, the authors reported that infusion with Ang II increases cardiac IL-12 expression derived from cardiac macrophages; detection of IL-12 promotes the activation of CD4+ T lymphocytes and increases differentiation of M2 macrophages, thereby up-regulating the activation of the transforming growth factor-β1 (TGF-β1) signaling pathway, which then aggravates cardiac fibrosis (Li et al., 2012). In mouse models of myocardial infarction, deletion of IL-23 significantly reduces the expression of multiple fibrosis markers, including α-smooth muscle actin (α-SMA), collagen I, and collagen III (Savvatis et al., 2014). Unexpectedly, Yan et al. also reported that IL-23 deficiency amplifies the inflammatory response and promotes the release of various inflammatory factors, especially IL-17, which further promotes the infiltration and deposition of γδT cells in the left ventricle, promotes the apoptosis of cardiomyocytes, and aggravates cardiac fibrosis in a murine myocardial infarction model (Yan et al., 2012). In addition, IL-12p35 knockout increased the levels of cardiac mitochondrial reactive oxygen species (ROS) and calcium ion overload, which further aggravated mitochondrial dysfunction and energy failure, increased myocardial cell apoptosis, worsened cardiac dysfunction, and increased cardiac fibrosis in 25-month-old aging mice (Ye et al., 2020). However, how the cytokines IL-12 and IL-35 mediate these biological effects is currently unknown. Furthermore, no studies concerning IL-27 and IL-35 related to cardiac fibrosis have been reported.
Interleukin-12 Family Members and Cardiac Ischemia Reperfusion Injury
Ischemia reperfusion injury of the heart is an important issue that cannot be ignored in heart transplantation. A large number of studies have confirmed that myocardial apoptosis mediated by inflammatory responses after cardiac reperfusion is one of the most important mechanisms of ischemia reperfusion injury of the heart (Shin et al., 2017; Schanze et al., 2019).
Numerous animal studies have reported that members of the IL-12 family are involved in cardiac ischemia-reperfusion injury. In a recent study, Yan found that dectin-2 deficiency could protect against cardiac ischemia-reperfusion injury via alleviating Th1 immune responses and further decreasing IL-12 expression (Yan et al., 2017). An earlier study reported that high-mobility group box 1 (Hmgb-1) promoted ischemia-reperfusion injury in a mouse cardiac transplantation model (Zhu et al., 2013). In subsequent studies, pentraxin-3 and necrostatin-1 were also found to attenuate ischemic reperfusion injury by decreasing the expression of IL-23 (Zhang et al., 2014b; Zhu et al., 2014). Hu et al. reported that administration of mouse anti-IL-23 neutralizing antibody significantly reduced the expression of inflammatory markers such as IL-6, tumor necrosis factor α (TNF-α), and pro-oxidant markers such as malondialdehyde (MDA), and decreased the levels of superoxide dismutase (SOD), thereby relieving cardiac ischemia reperfusion injury (Hu et al., 2016). Up-regulation of cardiac IL-23 expression by adenovirus significantly increased the expression of serum lactate dehydrogenase (LDH) and creatine kinase myocardial band (CK-MB), elevated the expression of apoptosis-related proteins and infarcted areas, and these effects could be reversed by AG490, an inhibitor of the JAK2-STAT pathway (Liao et al., 2017). However, the roles of both IL-27 and IL-35 in cardiac ischemia and reperfusion injury have not been studied to date.
Interleukin-12 Family Members and Atrial Fibrillation
Atrial fibrillation is one of the most common arrhythmias and can lead to vascular embolizations, the most serious of which is cerebral artery embolization. Literature reports confirm that the mechanism of atrial fibrillation may be closely related to the occurrence of atrial fibrosis (Jalife and Kaur, 2015; Nattel, 2017).
Previous clinical studies have reported that elevated IL-12 expression is observed in left atrial tissues of atrial fibrillation patients (Stein et al., 2008; Lappegård et al., 2013). Chen et al. found that IL-27 genetic variants, including the rs153109 G allele and GG genotype, increased the occurrence of atrial fibrillation in the Chinese Han population (Chen et al., 2017a).
Recent animal studies have reported that inhibition of Ang II-induced M1 macrophage differentiation and reduction of IL-12 release can reduce the occurrence of atrial fibrosis and atrial fibrillation (Sun et al., 2019b). No studies on the expression and mechanisms of the involvement of IL-23 and IL-35 in atrial fibrillation have been reported.
Interleukin-12 Family Members and Viral Myocarditis
Viral myocarditis is an uncommon heart disease. The death rate involving severe myocarditis exceeds that of AMI. Immune responses induced by viral infection are an important cause of myocardial injury in viral myocarditis (Chen et al., 2013; Pollack et al., 2015).
So far, although no clinical experiments have been reported on IL-12 family members and viral myocarditis, a large number of animal experiments have confirmed that all IL-12 family members are associated with viral myocarditis. Substantial evidence indicates that IL-12 expression is increased in both plasma and heart tissue of coxsackievirus B3-induced viral myocarditis in mice. In addition, elevated IL-12R levels were also found in heart tissue of mice with viral myocarditis (Fairweather et al., 2003; Nyland et al., 2012; Jenke et al., 2014; Zha et al., 2015; Miteva et al., 2017; Zhang et al., 2017). In an earlier study, the authors found that treatment with coxsackievirus B3 significantly increased both cardiac IL-12p35 and IL-12p40 expression, and treatment with recombinant mouse IL-12 and anti-IL-12 neutralizing antibodies reduced and increased mortality, respectively, in mice with viral myocarditis (Shioi et al., 1997). In a subsequent study, Nishio et al. reported that carvedilol treatment increases both IL-12 and interferon-γ (IFN-γ) expression, thereby reducing virus replication and thus improving survival rates in viral myocarditis mice (Nishio et al., 2003). In another study, Fairweather et al. demonstrated that the protective effect of IL-12 in viral myocarditis is mediated by activation of the STAT4 pathway and promotion of IFN-γ release. Knockout of the STAT4 pathway and IFN-γ can significantly reverse the protective effects of IL-12 and aggravate myocardial cell injury and mortality (Fairweather et al., 2005). Similarly, circulating IL-23 levels were also observed to be increased in coxsackievirus B3-induced mouse viral myocarditis (Yang et al., 2011; Sesti-Costa et al., 2017). Although there are no direct reports concerning the effects of IL-23 on viral myocarditis, emodin can reduce myocardial injury and mortality mediated by viral myocarditis by reducing the expression of IL-23, indicating that IL-23 can aggravate myocardial injury in viral myocarditis (Jiang et al., 2014). In an initial study, Kong et al. found that IL-27 levels were elevated in mice with viral myocarditis, and regulated IL-17 expression, suggesting that IL-27 may be involved in the development of viral myocarditis (Kong et al., 2014). In a follow-up study, Zhu et al. found that IL-27 inhibited immune responses to Th17 and reduced the expression of IL-17, thereby protecting against coxsackievirus B3-induced viral myocarditis (Zhu et al., 2015). Unlike other members of the IL-12 family, IL-35 levels were found to be reduced in a mouse model of viral myocarditis, and were negatively correlated with the severity of viral myocarditis, as was the frequency of Tregs (Hu et al., 2014; Ouyang et al., 2017; Xu et al., 2018). In addition, up-regulation of IL-35 expression can significantly reduce Th17-mediated immune responses, and decrease IL-17 expression, thereby alleviating cardiac injury caused by viral myocarditis (Hu et al., 2014). These studies have confirmed that all IL-12 family members are involved in the course of viral myocarditis, and the mechanisms involved in their action are related to the regulation of Th1 and Th17 immune responses. Whether other immune cells are involved needs further confirmation.
Interleukin-12 Family Members and Cardiomyopathy
Cardiomyopathy is a rare heart disease characterized by enlarged ventricular spaces with unknown etiology. Its pathological mechanisms are very complex, and many factors, including genetic variation, can induce its occurrence (Heinig et al., 2017).
Data from previous clinical experiment reported that IL-12 expression was found to be unchanged in patients with idiopathic dilated cardiomyopathy and their relatives (Marriott et al., 1996), whereas IL-12 expression was found to be elevated in patients with autoimmune cardiomyopathy or alcoholic cardiomyopathy (Izumi et al., 2000; Jenke et al., 2013; Panchenko et al., 2015). IL-12R gene polymorphisms, including IL-12B 3' UTR C and IL-12B 3' UTR CC, result in significantly higher gene expression, and may increase the incidence of Chagas cardiomyopathy (Zafra et al., 2007). Similar to the IL-12 expression trends, circulating or cardiac IL-23 levels were found to be increased in patients with dilated cardiomyopathy, and with idiopathic dilated cardiomyopathy (Yi et al., 2009; Li et al., 2010; Myers et al., 2016). Individuals with IL-12, IL-23R polymorphisms, such as SNP rs10889677, are more susceptible to dilated cardiomyopathy among the Chinese Han population, rather than those with rs1884444 and rs11465817 (Chen et al., 2009). Elevated IL-27 mRNA levels were observed in the heart tissue of human dilated cardiomyopathy patients, and an IL-27 gene polymorphism involving SNP rs153109, rather than SNP rs17855750, predisposes to dilated cardiomyopathy in the Chinese Han population (Noutsias et al., 2011; Chen et al., 2017b). The expression of IL-35 in human cardiomyopathy has not been reported.
In a mouse model of cardiac myosin immunized-mice, the absence of IL-12R significantly reduced cardiac immune responses and delayed the progression of autoimmune cardiomyopathy, whereas knockout of the STAT4 pathway and IFN-γ significantly reversed the protective effect of IL-12 in autoimmune cardiomyopathy (Afanasyeva et al., 2001). In another study, Fairweather et al. found that knockout of IL-12R significantly slowed the progression of dilated cardiomyopathy in murine chronic viral myocarditis (Fairweather et al., 2004). Using IL-12p35 and IL-12p40 knockout mice and anti-IL-23 neutralizing antibodies, other researchers found that IL-23, rather than IL-12, exacerbated the progression of a localized underwear purchase response in the heart and autoimmune myocarditis, which could be blocked by anti-IL-17 neutralizing antibodies (Sonderegger et al., 2006). In a recent study, Wu et al. demonstrated that IL-23 is necessary to initiate cardiac autoimmunity by stimulating the activation and differentiation of CD4+ T lymphocytes (Wu et al., 2016). In contrast there have been no studies regarding cardiomyopathy and involvement of IL-27 and IL-35.
Interleukin-12 Family Members and Other Cardiovascular Diseases
IL-12 family members are also implicated in other cardiovascular diseases that are less common, such as congenital heart disease, ventricular fibrillation, and rejection after cardiac transplantation.
In earlier studies, it was reported that in young children with congenital heart disease, circulating IL-12 levels did not exhibit significant change after surgery (Madhok et al., 2006). Furthermore, the IL-27 gene polymorphism, SNP rs153109, rather than rs17855750, is associated with congenital atrial septal defects and congenital ventricular septal defects (Zhang et al., 2016). In addition, IL-12 levels in plasma and brain tissue are significantly increased in an animal model of cardiac arrest after ventricular fibrillation. (Janata et al., 2014; Heo et al., 2017). In a mouse model of heart transplantation, administration of an anti-IL-12p40 antibody significantly reduced invasion by γδT cells, reduced the expression of various inflammatory factors, and greatly improved the survival of mice (Wang et al., 2012). In an animal model of acute myocardial injury induced by the chemotherapeutic drug doxorubicin, deletion of IL-12p35 significantly increased cardiac injury, which was associated with increased inflammatory responses, oxidative stress, apoptosis, and autophagy. Treatment with recombinant mouse IL-12 significantly reversed these effects, suggesting that both IL-12 and IL-35 may play protective roles in cardiac injury induced by doxorubicin (Jia, 2018; Ye et al., 2018b; Ye et al., 2018c).
Conclusions
The current review sought to describe the composition, structure, molecular receptors, signaling pathways, and regulatory roles of each IL-12 family member. The expression of IL-12 family members in different cardiovascular diseases in humans and animals, and the regulatory effect of IL-12 family members on inflammatory response in different cardiovascular models are also summarized in this paper in Tables 2 and 3. In addition, we also described the roles and possible mechanisms of involvement of IL-12 members in different cardiovascular diseases. Among these IL-12 family members, IL-12 can aggravate a variety of cardiovascular diseases, in addition to acute cardiac injury induced by doxorubicin, and hypertension prompted by Ang II. IL-23 mostly plays a role in injury. IL-27 has a two-sided regulatory effect in cardiovascular disease, with both protective and damaging effects; while IL-35 has been found to play a protective role in all cardiovascular diseases. Just as Table 4. Although IL-12 family members are involved in various biological effects such as inflammatory responses, oxidative stress, and apoptosis, the regulation of immune cell differentiation and inflammation is still the most important mechanism for the involvement of IL-12 in the development of cardiovascular diseases. In view of this, IL-12 family members may be potential targets for clinical prevention, intervention, and treatment of cardiovascular diseases. Hence, when considering IL-12 family members as potential targets for cardiovascular disease therapy, the influence of other cytokines and interactions involving interleukin family members should be considered.
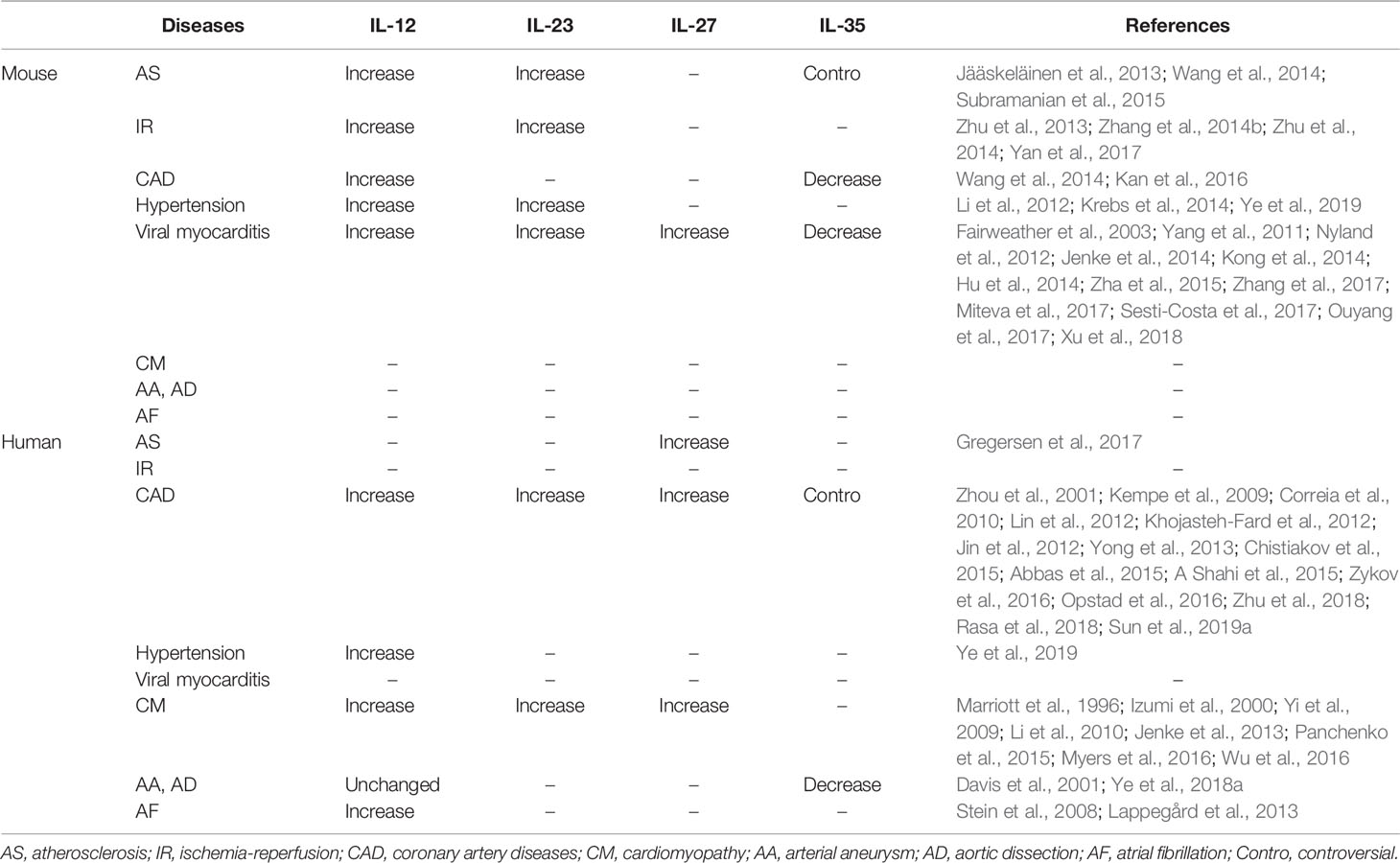
Table 2 Expression of IL-12 family members in cardiovascular diseases.
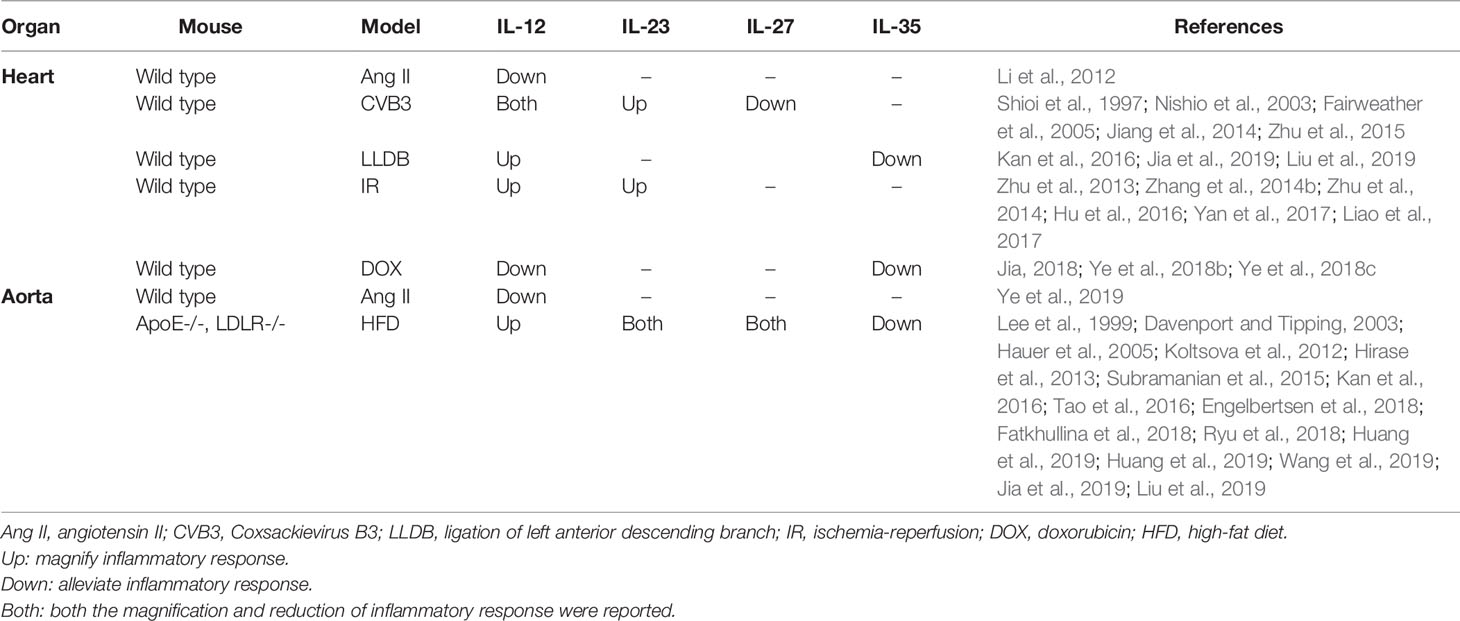
Table 3 Regulation of different inflammatory environments by members of the interleukin-12 family.
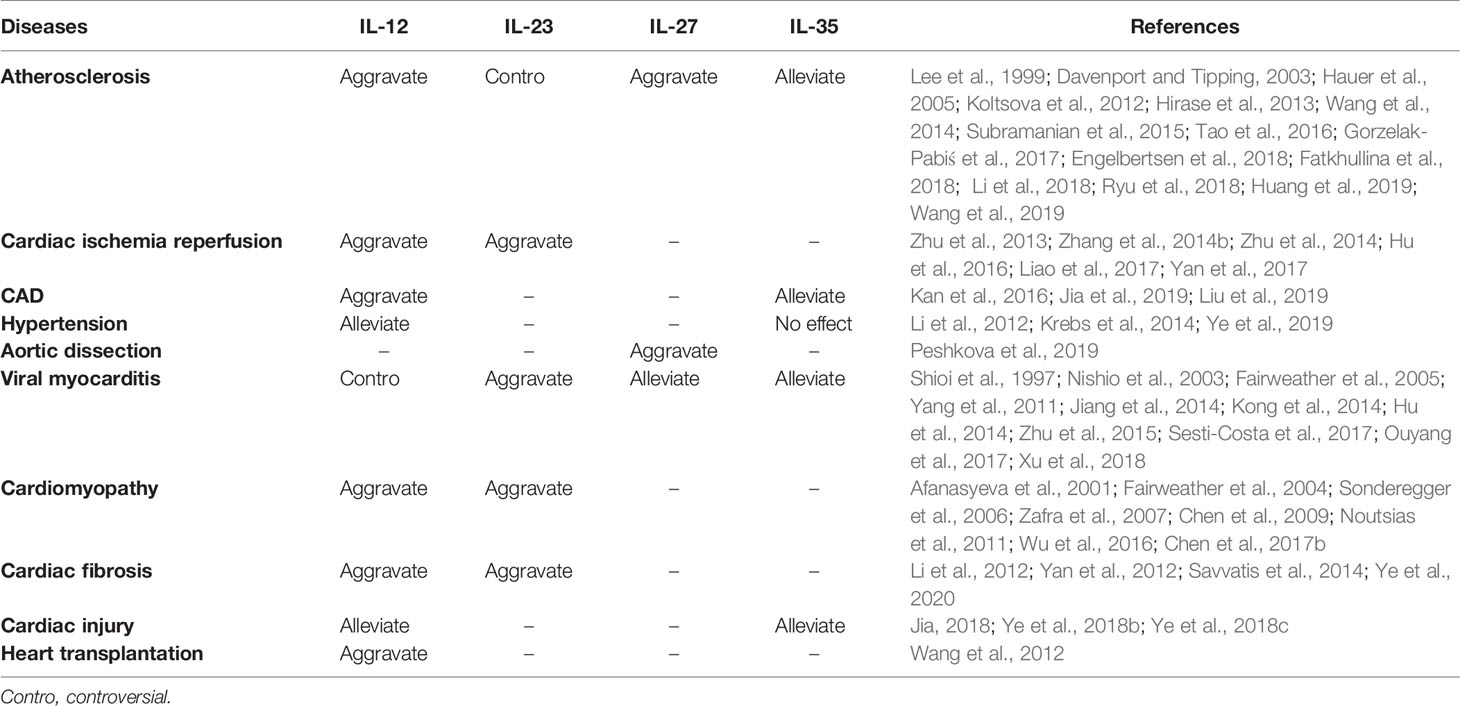
Table 4 Regulation of IL-12 family members on cardiovascular diseases.
Author Contributions
JY, YW, and ZW wrote this article. LL, ZY, MW, YX, DY, and JZ searched literatures. YL, QJ, and JW provided ideas and financial support.
Funding
This work was supported by the National Natural Science Foundation of China (No. 81770472 and No. 81560085 to QJ; No. 81760051 to YL).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
A Shahi, H., Shimada, K., Miyauchi, K., Yoshihara, T., Sai, E., Shiozawa, T., et al. (2015). Elevated circulating levels of inflammatory markers in patients with acute coronary syndrome. Int. J. Vasc. Med. 2015, 805375. doi: 10.1155/2015/805375
Abbas, A., Gregersen, I., Holm, S., Daissormont, I., Bjerkeli, V., Krohg-Sørensen, K., et al. (2015). Interleukin 23 levels are increased in carotid atherosclerosis: possible role for the interleukin 23/interleukin 17 axis. Stroke 46 (3), 793–799. doi: 10.1161/STROKEAHA.114.006516
Afanasyeva, M., Wang, Y., Kaya, Z., Stafford, E. A., Dohmen, K. M., Sadighi Akha, A. A., et al. (2001). Interleukin-12 receptor/STAT4 signaling is required for the development of autoimmune myocarditis in mice by an interferon-gamma-independent pathway. Circulation 18104 (25), 3145–3151. doi: 10.1161/hc5001.100629
Andrews, C., McLean, M. H., Durum, S. K. (2016). Interleukin-27 as a novel therapy for inflammatory bowel disease: a critical review of the literature. Inflamma. Bowel Dis. 22 (9), 2255–2264. doi: 10.1097/MIB.0000000000000818
Bethel, M. A., Patel, R. A., Merrill, P., Lokhnygina, Y., Buse, J. B., Mentz, R. J., et al. (2018). Cardiovascular outcomes with glucagon-like peptide-1 receptor agonists in patients with type 2 diabetes: a meta-analysis. Lancet Diabetes Endocrinol. 6 (2), 105–113. doi: 10.1016/S2213-8587(17)30412-6
Chen, Y., Zhou, B., Peng, Y., Wang, Y., Li, C., Ding, X., et al. (2009). Interleukin-23 receptor gene polymorphisms is associated with dilated cardiomyopathy in Chinese Han population. Tissue Antigens 73 (4), 330–334. doi: 10.1111/j.1399-0039.2009.01217.x
Chen, H. S., Wang, W., Wu, S. N., Liu, J. P. (2013). Corticosteroids for viral myocarditis. Cochrane Database Syst. Rev. 10, CD004471. doi: 10.1002/14651858.CD004471.pub3
Chen, Y., Zeng, J., Zhang, R., Zeng, L., Li, Y., Wei, H., et al. (2017a). Effect of interleukin-27 genetic variants on atrial fibrillation susceptibility. Genet. Test Mol. Biomarkers 21 (2), 97–101. doi: 10.1089/gtmb.2016.0219
Chen, Y., Zhang, R., Zeng, L., Wei, H., Chen, Y., Zeng, J. (2017b). IL-27 genetic variation and susceptibility of dilated cardiomyopathy in Chinese Han population. Per. Med. 14 (5), 401–408. doi: 10.2217/pme-2017-0013
Chistiakov, D. A., Bobryshev, Y. V., Orekhov, A. N. (2015). Heterogeneity of Tregs and the complexity in the IL-12 cytokine family signaling in driving T-cell immune responses in atherosclerotic vessels. Mol. Immunol. 65 (1), 133–138. doi: 10.1016/j.molimm.2015.01.013
Collison, L. W., Vignali, D. A. (2008). Interleukin-35: odd one out or part of the family? Immunol. Rev. 226, 248–262. doi: 10.1111/j.1600-065X.2008.00704.x
Collison, L. W., Pillai, M. R., Chaturvedi, V., Vignali, D. A. (2009). Regulatory T cell suppression is potentiated by target T cells in a cell contact, IL-35- and IL-10-dependent manner. J. Immunol. 182 (10), 6121–6128. doi: 10.4049/jimmunol.0803646
Collison, L. W., Chaturvedi, V., Henderson, A. L., Giacomin, P. R., Guy, C., Bankoti, J., et al. (2010). IL-35-mediated induction of a potent regulatory T cell population. Nat. Immunol. 11 (12), 1093–1101. doi: 10.1038/ni.1952
Collison, L. W., Delgoffe, G. M., Guy, C. S., Vignali, K. M., Chaturvedi, V., Fairweather, D., et al. (2012). The composition and signaling of the IL-35 receptor are unconventional. Nat. Immunol. 13 (3), 290–299. doi: 10.1038/ni.2227
Correia, L. C., Andrade, B. B., Borges, V. M., Clarêncio, J., Bittencourt, A. P., Freitas, R., et al. (2010). Prognostic value of cytokines and chemokines in addition to the GRACE Score in non-ST-elevation acute coronary syndromes. Clin. Chim. Acta 411 (7-8), 540–545. doi: 10.1016/j.cca.2010.01.011
Cox, J. H., Kljavin, N. M., Ramamoorthi, N., Diehl, L., Batten, M., Ghilardi, N. (2011). IL-27 promotes T cell-dependent colitis through multiple mechanisms. J. Exp. Med. 208 (1), 115–1123. doi: 10.1084/jem.20100410
Davenport, P., Tipping, P. G. (2003). The role of interleukin-4 and interleukin-12 in the progression of atherosclerosis in apolipoprotein E-deficient mice. Am. J. Pathol. 163 (3), 1117–1125. doi: 10.1016/S0002-9440(10)63471-2
Davis, V. A., Persidskaia, R. N., Baca-Regen, L. M., Fiotti, N., Halloran, B. G., Baxter, B. T. (2001). Cytokine pattern in aneurysmal and occlusive disease of the aorta. J. Surg. Res. 101 (2), 152–156. doi: 10.1006/jsre.2001.6281
Delgoffe, G. M., Murray, P. J., Vignali, D. A. (2011). Interpreting mixed signals: the cell's cytokine conundrum. Curr. Opin. Immunol. 23 (5), 632–638. doi: 10.1016/j.coi.2011.07.013
Donofrio, M. T., Moon-Grady, A. J., Hornberger, L. K., Copel, J. A., Sklansky, M. S., Abuhamad, A., et al. (2014). Diagnosis and treatment of fetal cardiac disease: a scientific statement from the American Heart Association. Circulation 129 (21), 2183–2242. doi: 10.1161/01.cir.0000437597.44550.5d
Dukkipati, S. R., Koruth, J. S., Choudry, S., Miller, M. A., Whang, W., Reddy, V. Y. (2017). Catheter ablation of ventricular tachycardia in structural heart disease: indications, strategies, and outcomes-part II. J. Am. Coll. Cardiol. 70 (23), 2924–2941. doi: 10.1016/j.jacc.2017.10.030
Engelbertsen, D., Depuydt, M. A. C., Verwilligen, R. A. F., Rattik, S., Levinsohn, E., Edsfeldt, A., et al. (2018). IL-23R deficiency does not impact atherosclerotic plaque development in mice. J. Am. Heart Assoc. 7 (8). doi: 10.1161/JAHA.117.008257
Fairweather, D., Yusung, S., Frisancho, S., Barrett, M., Gatewood, S., Steele, R., et al. (2003). IL-12 receptor beta 1 and Toll-like receptor 4 increase IL-1 beta- and IL-18-associated myocarditis and coxsackievirus replication. J. Immunol. 170 (9), 4731–4737. doi: 10.4049/jimmunol.170.9.4731
Fairweather, D., Frisancho-Kiss, S., Yusung, S. A., Barrett, M. A., Davis, S. E., Gatewood, S. J., et al. (2004). Interferon-gamma protects against chronic viral myocarditis by reducing mast cell degranulation, fibrosis, and the pro-fibrotic cytokines transforming growth factor-beta 1, interleukin-1 beta, and interleukin-4 in the heart. Am. J. Pathol. 165 (6), 1883–1894. doi: 10.1016/S0002-9440(10)63241-5
Fairweather, D., Frisancho-Kiss, S., Yusung, S. A., Barrett, M. A., Davis, S. E., Steele, R. A., et al. (2005). IL-12 protects against coxsackievirus B3-induced myocarditis by increasing IFN-gamma and macrophage and neutrophil populations in the heart. J. Immunol. 174 (1), 261–269. doi: 10.4049/jimmunol.174.1.261
Fatkhullina, A. R., Peshkova, I. O., Dzutsev, A., Aghayev, T., McCulloch, J. A., Thovarai, V., et al. (2018). An interleukin-23-interleukin-22 axis regulates intestinal microbial homeostasis to protect from diet-induced atherosclerosis. Immunity 49 (5), 943–957. doi: 10.1016/j.immuni.2018.09.011
Gorabi, A. M., Hajighasemi, S., Kiaie, N., Rosano, G. M. C., Sathyapalan, T., Al-Rasadi, K., et al. (2019). Anti-fibrotic effects of curcumin and some of its analogues in the heart. Heart Fail. Rev. doi: 10.1007/s10741-019-09854-6
Gorzelak-Pabiś, P., Chalubinski, M., Wojdan, K., Luczak, E., Duraj, I., Mozdzan, M., et al. (2017). Increased plasma concentrations of interleukin 35 in patients with coronary artery disease. Arch. Med. Sci. 13 (4), 778–784. doi: 10.5114/aoms.2016.63751
Gregersen, I., Sandanger, Ø., Askevold, E. T., Sagen, E. L., Yang, K., Holm, S., et al. (2017). Interleukin 27 is increased in carotid atherosclerosis and promotes NLRP3 inflammasome activation. PloS One 12 (11), e0188387. doi: 10.1371/journal.pone.0188387
Guzik, T. J., Touyz, R. M. (2017). Oxidative Stress, inflammation, and vascular aging in hypertension. Hypertension 70 (4), 660–667. doi: 10.1161/HYPERTENSIONAHA.117.07802
Hauer, A. D., Uyttenhove, C., de Vos, P., Stroobant, V., Renauld, J. C., van Berkel, T. J., et al. (2005). Blockade of interleukin-12 function by protein vaccination attenuates atherosclerosis. Circulation 112 (7), 1054–1062. doi: 10.1161/CIRCULATIONAHA.104.533463
Heinig, M., Adriaens, M. E., Schafer, S., van Deutekom, H. W. M., Lodder, E. M., Ware, J. S., et al. (2017). Natural genetic variation of the cardiac transcriptome in non-diseased donors and patients with dilated cardiomyopathy. Genome Biol. 18 (1), 170. doi: 10.1186/s13059-017-1286-z
Heo, J. C., Kim, B., Kim, Y. N., Kim, D. K., Lee, J. H. (2017). Induction of inflammation in vivo by electrocardiogram sensor operation using wireless power transmission. Sens. (Basel) 17 (12). doi: 10.3390/s17122905
Hirase, T., Hara, H., Miyazaki, Y., Ide, N., Nishimoto-Hazuku, A., Fujimoto, H., et al. (2013). Interleukin 27 inhibits atherosclerosis via immune regulation of macrophages in mice. Am. J. Physiol. Heart Circ. Physiol. 305 (3), H420–H429. doi: 10.1152/ajpheart.00198.2013
Hu, Y., Dong, C., Yue, Y., Xiong, S. (2014). In vivo delivery of interleukin-35 relieves coxsackievirus-B3-induced viral myocarditis by inhibiting Th17 cells. Arch. Virol. 159 (9), 2411–2419. doi: 10.1007/s00705-014-2098-z
Hu, X., Ma, R., Lu, J., Zhang, K., Xu, W., Jiang, H., et al. (2016). IL-23 promotes myocardial I/R injury by increasing the inflammatory responses and oxidative stress reactions. Cell Physiol. Biochem. 38 (6), 2163–2172. doi: 10.1159/000445572
Huang, Y., Hu, H., Liu, L., Ye, J., Wang, Z., Que, B., et al. (2019). Interleukin-12p35 deficiency reverses the Th1/Th2 imbalance, aggravates the Th17/Treg imbalance, and ameliorates atherosclerosis in ApoE-/- mice. Mediators Inflamm. 2019, 3152040. doi: 10.1155/2019/3152040
Hunter, C. A. (2005). New IL-12-family members: IL-23 and IL-27, cytokines with divergent functions. Nat. Rev. Immunol. 5 (7), 521–531. doi: 10.1038/nri1648
Ihle, J. N. (1995). The Janus protein tyrosine kinase family and its role in cytokine signaling. Adv. Immunol. 60, 1–35. doi: 10.1016/S0065-2776(08)60582-9
Izumi, T., Takehana, H., Matsuda, C., Yokoyama, H., Kohno, K., Suzuki, K., et al. (2000). Experimental autoimmune myocarditis and its pathomechanism. Herz 25 (3), 274–278. doi: 10.1007/s000590050020
Jääskeläinen, A. E., Seppälä, S., Kakko, T., Jaakkola, U., Kallio, J. (2013). Systemic treatment with neuropeptide Y receptor Y1-antagonist enhances atherosclerosis and stimulates IL-12 expression in ApoE deficient mice. Neuropeptides 47 (2), 67–73. doi: 10.1016/j.npep.2012.11.001
Jalife, J., Kaur, K. (2015). Atrial remodeling, fibrosis, and atrial fibrillation. Trends Cardiovasc. Med. 25 (6), 475–484. doi: 10.1016/j.tcm.2014.12.015
Janata, A., Magnet, I. A., Uray, T., Stezoski, J. P., Janesko-Feldman, K., Tisherman, S. A., et al. (2014). Regional TNFα mapping in the brain reveals the striatum as a neuroinflammatory target after ventricular fibrillation cardiac arrest in rats. Resuscitation 85 (5), 694–701. doi: 10.1016/j.resuscitation.2014.01.033
Jenke, A., Wilk, S., Poller, W., Eriksson, U., Valaperti, A., Rauch, B. H., et al. (2013). Adiponectin protects against Toll-like receptor 4-mediated cardiac inflammation and injury. Cardiovasc. Res. 99 (3), 422–431. doi: 10.1093/cvr/cvt118
Jenke, A., Holzhauser, L., Löbel, M., Savvatis, K., Wilk, S., Weithäuser, A., et al. (2014). Adiponectin promotes coxsackievirus B3 myocarditis by suppression of acute anti-viral immune responses. Basic Res. Cardiol. 109 (3), 408. doi: 10.1007/s00395-014-0408-y
Jia, D., Jiang, H., Weng, X., Wu, J., Bai, P., Yang, W., et al. (2019). Interleukin-35 promotes macrophage survival and improves wound healing after myocardial infarction in mice. Circ. Res. 124 (9), 1323–1336. doi: 10.1161/CIRCRESAHA.118.314569
Jia, Z. (2018). The Clinical potential of IL-12/IL-35 in treating chemotherapy drug-induced cardiac injury. EBioMedicine 35, 2–3. doi: 10.1016/j.ebiom.2018.08.018
Jiang, N., Liao, W., Kuang, X. (2014). Effects of emodin on IL-23/IL-17 inflammatory axis, Th17 cells and viral replication in mice with viral myocarditis. Nan Fang Yi Ke Da Xue Xue Bao. 34 (3), 373–378. doi: 10.3969/j.issn.1673-4254.2014.03.17
Jin, W., Zhao, Y., Yan, W., Cao, L., Zhang, W., Wang, M., et al. (2012). Elevated circulating interleukin-27 in patients with coronary artery disease is associated with dendritic cells, oxidized low-density lipoprotein, and severity of coronary artery stenosis. Mediators Inflamm. 2012, 506283. doi: 10.1155/2012/506283
Kan, X., Wu, Y., Ma, Y., Zhang, C., Li, P., Wu, L., et al. (2016). Deficiency of IL-12p35 improves cardiac repair after myocardial infarction by promoting angiogenesis. Cardiovasc. Res. 109 (2), 249–259. doi: 10.1093/cvr/cvv255
Kastelein, R. A., Hunter, C. A., Cua, D. J. (2007). Discovery and biology of IL-23 and IL-27: related but functionally distinct regulators of inflammation. Annu. Rev. Immunol. 25, 221–242. doi: 10.1146/annurev.immunol.22.012703.104758
Kempe, S., Heinz, P., Kokai, E., Devergne, O., Marx, N., Wirth, T. (2009). Epstein-barr virus-induced gene-3 is expressed in human atheroma plaques. Am. J. Pathol. 175 (1), 440–447. doi: 10.2353/ajpath.2009.080752
Khojasteh-Fard, M., Abolhalaj, M., Amiri, P., Zaki, M., Taheri, Z., Qorbani, M., et al. (2012). IL-23 gene expression in PBMCs of patients with coronary artery disease. Dis. Markers 33 (6), 289–293. doi: 10.1155/2012/629814
Kirabo, A., Fontana, V., de Faria, A. P., Loperena, R., Galindo, C. L., Wu, J., et al. (2014). DC isoketal-modified proteins activate T cells and promote hypertension. J. Clin. Invest. 124 (10), 4642–4656. doi: 10.1172/JCI74084
Koltsova, E. K., Kim, G., Lloyd, K. M., Saris, C. J., von Vietinghoff, S., Kronenberg, M., et al. (2012). Interleukin-27 receptor limits atherosclerosis in Ldlr-/- mice. Circ. Res. 111 (10), 1274–1285. doi: 10.1161/CIRCRESAHA.112.277525
Kong, Q., Gao, M., Xue, Y., Pan, X., Lai, W., Wu, W. (2014). Interleukin-17 contributes to the macrophage secretion of interleukin-27 in a murine model of viral myocarditis. Zhonghua Xin Xue Guan Bing Za Zhi 42 (5), 428–432. doi: 10.3760/cma.j.issn.0253–3758.2014.05.017
Krebs, C. F., Lange, S., Niemann, G., Rosendahl, A., Lehners, A., Meyer-Schwesinger, C., et al. (2014). Deficiency of the interleukin 17/23 axis accelerates renal injury in mice with deoxycorticosterone acetate+angiotensin II-induced hypertension. Hypertension 63 (3), 565–571. doi: 10.1161/HYPERTENSIONAHA.113.02620
Langrish, C. L., McKenzie, B. S., Wilson, N. J., de Waal Malefyt, R., Kastelein, R. A., Cua, D. J. (2004). IL-12 and IL-23: master regulators of innate and adaptive immunity. Immunol. Rev. 202, 96–105. doi: 10.1111/j.0105-2896.2004.00214.x
Lappegård, K. T., Pop-Purceleanu, M., van Heerde, W., Sexton, J., Tendolkar, I., Pop, G. (2013). Improved neurocognitive functions correlate with reduced inflammatory burden in atrial fibrillation patients treated with intensive cholesterol lowering therapy. J. Neuroinflammation 10, 78. doi: 10.1186/1742-2094-10-78
Lee, T. S., Yen, H. C., Pan, C. C., Chau, L. Y. (1999). The role of interleukin 12 in the development of atherosclerosis in ApoE-deficient mice. Arterioscler. Thromb. Vasc. Biol. 19 (3), 734–742. doi: 10.1161/01.ATV.19.3.734
Leong, D. P., Joseph, P. G., McKee, M., Anand, S. S., Teo, K. K., Schwalm, J. D., et al. (2017). Reducing the global burden of cardiovascular disease, part 2: prevention and treatment of cardiovascular disease. Circ. Res. 121 (6), 695–710. doi: 10.1161/CIRCRESAHA.117.311849
Li, J., Wang, L., Wang, S., Zhu, H., Ye, P., Xie, A., et al. (2010). The Treg/Th17 imbalance in patients with idiopathic dilated cardiomyopathy. Scand. J. Immunol. 71 (4), 298–303. doi: 10.1111/j.1365-3083.2010.02374.x
Li, Y., Zhang, C., Wu, Y., Han, Y., Cui, W., Jia, L., et al. (2012). Interleukin-12p35 deletion promotes CD4 T-cell-dependent macrophage differentiation and enhances angiotensin II-Induced cardiac fibrosis. Arterioscler. Thromb. Vasc. Biol. 32 (7), 1662–1674. doi: 10.1161/ATVBAHA.112.249706
Li, X., Shao, Y., Sha, X., Fang, P., Kuo, Y. M., Andrews, A. J., et al. (2018). IL-35 (interleukin-35) suppresses endothelial cell activation by inhibiting mitochondrial reactive oxygen species-mediated site-specific acetylation of H3K14 (Histone 3 Lysine 14). Arterioscler. Thromb. Vasc. Biol. 38 (3), 599–609. doi: 10.1161/ATVBAHA.117.310626
Liao, Y., Hu, X., Guo, X., Zhang, B., Xu, W., Jiang, H. (2017). Promoting effects of IL−23 on myocardial ischemia and reperfusion are associated with increased expression of IL−17A and up-regulation of the JAK2−STAT3 signaling pathway. Mol. Med. Rep. 16 (6), 9309–9316. doi: 10.3892/mmr.2017.7771
Lin, Y., Huang, Y., Lu, Z., Luo, C., Shi, Y., Zeng, Q., et al. (2012). Decreased plasma IL-35 levels are related to the left ventricular ejection fraction in coronary artery diseases. PloS One 7 (12), e52490. doi: 10.1371/journal.pone.0052490
Liu, X., Zhang, R., Hou, J., Wu, J., Zhang, M., Fang, S., et al. (2019). Interleukin-35 promotes early endothelialization after stent implantation by regulating macrophage activation. Clin. Sci. (Lond.) 133 (7), 869–884. doi: 10.1042/CS20180879
Longenecker, C. T., Sullivan, C., Baker, J. V. (2016). Immune activation and cardiovascular disease in chronic HIV infection. Curr. Opin. HIV AIDS 11 (2), 216–225. doi: 10.1097/COH.0000000000000227
Ma, X., Trinchieri, G. (2001). Regulation of interleukin-12 production in antigen-presenting cells. Adv. Immunol. 79, 55–92. doi: 10.1016/S0065-2776(01)79002-5
Madhok, A. B., Ojamaa, K., Haridas, V., Parnell, V. A., Pahwa, S., Chowdhury, D. (2006). Cytokine response in children undergoing surgery for congenital heart disease. Pediatr. Cardiol. 27 (4), 408–413. doi: 10.1007/s00246-006-0934-y
Mallat, Z., Tedgui, A., Henrion, D. (2016). Role of microvascular tone and extracellular matrix contraction in the regulation of interstitial fluid: implications for aortic dissection. Arterioscler. Thromb. Vasc. Biol. 36 (9), 1742–1747. doi: 10.1161/ATVBAHA.116.307909
Marriott, J. B., Goldman, J. H., Keeling, P. J., Baig, M. K., Dalgleish, A. G., McKenna, W. J. (1996). Abnormal cytokine profiles in patients with idiopathic dilated cardiomyopathy and their asymptomatic relatives. Heart 75 (3), 287–290. doi: 10.1136/hrt.75.3.287
Miteva, K., Pappritz, K., El-Shafeey, M., Dong, F., Ringe, J., Tschöpe, C., et al. (2017). Mesenchymal stromal cells modulate monocytes trafficking in coxsackievirus B3-induced myocarditis. Stem Cells Transl. Med. 6 (4), 1249–1261. doi: 10.1002/sctm.16-0353
Moore-Morris, T., Guimarães-Camboa, N., Yutzey, K. E., Pucéat, M., Evans, S. M. (2015). Cardiac fibroblasts: from development to heart failure. J. Mol. Med. (Berl.) 93 (8), 823–830. doi: 10.1007/s00109-015-1314-y
Myers, J. M., Cooper, L. T., Kem, D. C., Stavrakis, S., Kosanke, S. D., Shevach, E. M., et al. (2016). Cardiac myosin-Th17 responses promote heart failure in human myocarditis. JCI Insight 1 (9). doi: 10.1172/jci.insight.85851
Nattel, S. (2017). Molecular and cellular mechanisms of atrial fibrosis in atrial fibrillation. JACC Clin. Electrophysiol. 3 (5), 425–435. doi: 10.1016/j.jacep.2017.03.002
Nishio, R., Shioi, T., Sasayama, S., Matsumori, A. (2003). Carvedilol increases the production of interleukin-12 and interferon-gamma and improves the survival of mice infected with the encephalomyocarditis virus. J. Am. Coll. Cardiol. 41 (2), 340–345. doi: 10.1016/S0735-1097(02)02711-0
Noutsias, M., Rohde, M., Göldner, K., Block, A., Blunert, K., Hemaidan, L., et al. (2011). Expression of functional T-cell markers and T-cell receptor Vbeta repertoire in endomyocardial biopsies from patients presenting with acute myocarditis and dilated cardiomyopathy. Eur. J. Heart Fail. 13 (6), 611–618. doi: 10.1093/eurjhf/hfr014
Nyland, J. F., Fairweather, D., Shirley, D. L., Davis, S. E., Rose, N. R., Silbergeld, E. K. (2012). Low-dose inorganic mercury increases severity and frequency of chronic coxsackievirus-induced autoimmune myocarditis in mice. Toxicol. Sci. 125 (1), 134–143. doi: 10.1093/toxsci/kfr264
O'Shea, J. J., Gadina, M., Schreiber, R. D. (2002). Cytokine signaling in 2002: new surprises in the Jak/Stat pathway. Cell 109, S121–S131. doi: 10.1016/s0092-8674(02)00701-8
Oppmann, B., Lesley, R., Blom, B., Timans, J. C., Xu, Y., Hunte, B., et al. (2000). Novel p19 protein engages IL-12p40 to form a cytokine, IL-23, with biological activities similar as well as distinct from IL-12. Immunity 13 (5), 715–725. doi: 10.1016/S1074-7613(00)00070-4
Opstad, T. B., Arnesen, H., Pettersen, A. Å., Seljeflot, I. (2016). Combined elevated levels of the pro-inflammatory cytokines IL-18 and IL-12 are associated with clinical events in patients with coronary artery disease: an observational study. Metab. Syndr. Relat. Disord. 14 (5), 242–248. doi: 10.1089/met.2015.0130
Ouyang, H., Xiang, L., Chen, J., Xu, W., Jiao, Y., Shen, H. (2017). Significant reduction of peripheral blood interleukin-35 and CD4+EBI3+ T cells, which are negatively correlated with an increase in the plasma IL-17 and cTnI level, in viral myocarditis patients. Cent. Eur. J. Immunol. 42 (1), 91–96. doi: 10.5114/ceji.2016.65892
Panchenko, L. F., Moiseev, V. S., Pirozhkov, S. V., Terebilina, N. N., Naumova, T. A., Baronets, V., et al. (2015). Blood content of markers of inflammation and cytokines in patients with alcoholic cardiomyopathy and ischemic heart disease at various stages of heart failure. Kardiologiia 55 (3), 41–48. doi: 10.18565/cardio.2015.3.41-48
Peshkova, I. O., Aghayev, T., Fatkhullina, A. R., Makhov, P., Titerina, E. K., Eguchi, S., et al. (2019). IL-27 receptor-regulated stress myelopoiesis drives abdominal aortic aneurysm development. Nat. Commun. 10 (1), 5046. doi: 10.1038/s41467-019-13017-4
Peter, L., Paul, M. R., Göran, K. H. (2009). Inflammation in atherosclerosis: from pathophysiology to practice. J. Am. Coll. Cardiol. 54 (23), 2129–2138. doi: 10.1016/j.jacc.2009.09.009
Pflanz, S., Hibbert, L., Mattson, J., Rosales, R., Vaisberg, E., Bazan, J. F., et al. (2004). WSX-1 and glycoprotein 130 constitute a signal-transducing receptor for IL-27. J. Immunol. 172 (4), 2225–2231. doi: 10.4049/jimmunol.172.4.2225
Pober, J. S. (2014). Is hypertension an autoimmune disease? J. Clin. Invest. 124 (10), 4234–4236. doi: 10.1172/JCI77766
Pollack, A., Kontorovich, A. R., Fuster, V., Dec, G. W. (2015). Viral myocarditis–diagnosis, treatment options, and current controversies. Nat. Rev. Cardiol. 12 (11), 670–680. doi: 10.1038/nrcardio.2015.108
Posadas-Sánchez, R., Pérez-Hernández, N., Rodríguez-Pérez, J. M., Coral-Vázquez, R. M., Roque-Ramírez, B., Llorente, L., et al. (2017). Interleukin-27 polymorphisms are associated with premature coronary artery disease and metabolic parameters in the Mexican population: the genetics of atherosclerotic disease (GEA) Mexican study. Oncotarget 8 (38), 64459–64470. doi: 10.18632/oncotarget.16223
Presky, D. H., Yang, H., Minetti, L. J., Chua, A. O., Nabavi, N., Wu, C. Y., et al. (1996). A functional interleukin 12 receptor complex is composed of two beta-type cytokine receptor subunits. Proc. Natl. Acad. Sci. U. S. A. 93 (24), 14002–14007. doi: 10.1073/pnas.93.24.14002
Rabkin, S. W. (2017). The role matrix metalloproteinases in the production of aortic aneurysm. Prog. Mol. Biol. Transl. Sci. 147, 239–265. doi: 10.1016/bs.pmbts.2017.02.002
Raffort, J., Lareyre, F., Clément, M., Hassen-Khodja, R., Chinetti, G., Mallat, Z. (2017). Monocytes and macrophages in abdominal aortic aneurysm. Nat. Rev. Cardiol. 14 (8), 457–471. doi: 10.1038/nrcardio.2017.52
Rahman, K., Fisher, E. A. (2018). Insights from pre-clinical and clinical studies on the role of innate inflammation in atherosclerosis regression. Front. Cardiovasc. Med. 5, 32. doi: 10.3389/fcvm.2018.00032
Rasa, F., Naderi, N., Eftekhar, E., Mansoori, E., Rahimzadeh, M. (2018). Vitamin D status in coronary artery disease: association with IL-35 and TGF-β1 and disease severity. Endocr. Metab. Immune Disord. Drug Targets 18 (5), 522–529. doi: 10.2174/1871530318666180426101756
Ryu, H., Lim, H., Choi, G., Park, Y. J., Cho, M., Na, H., et al. (2018). Atherogenic dyslipidemia promotes autoimmune follicular helper T cell responses via IL-27. Nat. Immunol. 19 (6), 583–593. doi: 10.1038/s41590-018-0102-6
Savvatis, K., Pappritz, K., Becher, P. M., Lindner, D., Zietsch, C., Volk, H. D., et al. (2014). Interleukin-23 deficiency leads to impaired wound healing and adverse prognosis after myocardial infarction. Circ. Heart Fail. 7 (1), 161–171. doi: 10.1161/CIRCHEARTFAILURE.113.000604
Schanze, N., Bode, C., Duerschmied, D. (2019). Platelet contributions to myocardial ischemia/reperfusion Injury. Front. Immunol. 10, 1260. doi: 10.3389/fimmu.2019.01260
Sesti-Costa, R., Françozo, M. C. S., Silva, G. K., Proenca-Modena, J. L., Silva, J. S. (2017). TLR3 is required for survival following Coxsackievirus B3 infection by driving T lymphocyte activation and polarization: the role of dendritic cells. PloS One 12 (10), e0185819. doi: 10.1371/journal.pone.0185819
Sherifova, S., Holzapfel, G. A. (2019). Biomechanics of aortic wall failure with a focus on dissection and aneurysm: a review. Acta Biomater. 99, 1–17. doi: 10.1016/j.actbio.2019.08.017
Shin, B., Cowan, D. B., Emani, S. M., Del Nido, P. J., McCully, J. D. (2017). Mitochondrial transplantation in myocardial ischemia and reperfusion injury. Adv. Exp. Med. Biol. 982, 595–619. doi: 10.1007/978-3-319-55330-6_31
Shioi, T., Matsumori, A., Nishio, R., Ono, K., Kakio, T., Sasayama, S. (1997). Protective role of interleukin-12 in viral myocarditis. J. Mol. Cell Cardiol. 29 (9), 2327–2334. doi: 10.1006/jmcc.1997.0445
Sonderegger, I., Röhn, T. A., Kurrer, M. O., Iezzi, G., Zou, Y., Kastelein, R. A., et al. (2006). Neutralization of IL-17 by active vaccination inhibits IL-23-dependent autoimmune myocarditis. Eur. J. Immunol. 36 (11), 2849–2856. doi: 10.1002/eji.200636484
Stein, A., Wessling, G., Deisenhofer, I., Busch, G., Steppich, B., Estner, H., et al. (2008). Systemic inflammatory changes after pulmonary vein radiofrequency ablation do not alter stem cell mobilization. Europace 10 (4), 444–449. doi: 10.1093/europace/eun041
Subramanian, M., Thorp, E., Tabas, I. (2015). Identification of a non-growth factor role for GM-CSF in advanced atherosclerosis: promotion of macrophage apoptosis and plaque necrosis through IL-23 signaling. Circ. Res. 116 (2), e13–e24. doi: 10.1161/CIRCRESAHA.116.304794
Sun, L., He, C., Nair, L., Yeung, J., Egwuagu, C. E. (2015). Interleukin 12 (IL-12) family cytokines: role in immune pathogenesis and treatment of CNS autoimmune disease. Cytokine 75 (2), 249–255. doi: 10.1016/j.cyto.2015.01.030
Sun, J., Yu, H., Liu, H., Pu, D., Gao, J., Jin, X., et al. (2019a). Correlation of pre-operative circulating inflammatory cytokines with restenosis and rapid angiographic stenotic progression risk in coronary artery disease patients underwent percutaneous coronary intervention with drug-eluting stents. J. Clin. Lab. Anal., e23108. doi: 10.1002/jcla.23108
Sun, F., Guo, Z., Zhang, C., Che, H., Gong, W., Shen, Z., et al. (2019b). LncRNA NRON alleviates atrial fibrosis through suppression of M1 macrophages activated by atrial myocytes. Biosci. Rep. 39 (11). doi: 10.1042/BSR20192215
Tao, L., Zhu, J., Chen, Y., Wang, Q., Pan, Y., Yu, Q., et al. (2016). IL-35 improves Treg-mediated immune suppression in atherosclerotic mice. Exp. Ther. Med. 12 (4), 2469–2476. doi: 10.3892/etm.2016.3649
Timasheva, Y. R., Nasibullin, T. R., Zakirova, A. N., Mustafina, O. E. (2008). Association of interleukin-6, interleukin-12, and interleukin-10 gene polymorphisms with essential hypertension in Tatars from Russia. Biochem. Genet. 46 (1-2), 64–74. doi: 10.1007/s10528-007-9130-x
Vargas-Alarcón, G., Pérez-Hernández, N., Rodríguez-Pérez, J. M., Fragoso, J. M., Posadas-Romero, C., López-Bautista, F., et al. (2019). Interleukin 27 polymorphisms, their association with insulin resistance and their contribution to subclinical atherosclerosis. The GEA Mexican study. Cytokine 114, 32–37. doi: 10.1016/j.cyto.2018.11.028
Vignali, D. A., Kuchroo, V. K. (2012). IL-12 family cytokines: immunological playmakers. Nat. Immunol. 13 (8), 722–728. doi: 10.1038/ni.2366
Vignali, D. A. A., Collison, L. W., Workman, C. J. (2008). How regulatory T cells work. Nat. Rev. Immunol. 8, 523–532. doi: 10.1038/nri2343
Wang, S., Xu, X., Xie, A., Li, J., Ye, P., Liu, Z., et al. (2012). Anti-interleukin-12/23p40 antibody attenuates chronic rejection of cardiac allografts partly via inhibition γδT cells. Clin. Exp. Immunol. 169 (3), 320–329. doi: 10.1111/j.1365-2249.2012.04612.x
Wang, B., Dai, S., Dong, Z., Sun, Y., Song, X., Guo, C., et al. (2014). The modulation of endoplasmic reticulum stress by chemical chaperone up-regulates immune negative cytokine IL-35 in apolipoprotein E-deficient mice. PloS One 9 (1), e87787. doi: 10.1371/journal.pone.0087787
Wang, J., Zhao, P., Gao, Y., Zhang, F., Yuan, X., Jiao, Y., et al. (2019). The effects of anti-IL-23p19 therapy on atherosclerosis development in ApoE-/- mice. J. Interferon Cytokine Res. 39 (9), 564–571. doi: 10.1089/jir.2019.0050
Wei, X., Zhang, J., Gu, Q., Huang, M., Zhang, W., Guo, J., et al. (2017). Reciprocal expression of IL-35 and IL-10 defines two distinct effector Treg subsets that are required for maintenance of immune tolerance. Cell Rep. 21 (7), 1853–1869. doi: 10.1016/j.celrep.2017.10.090
Wojno, E. D., Hunter, C. A. (2012). New directions in the basic and translational biology of interleukin-27. Trends Immunol. 33, 91–97. doi: 10.1016/j.it.2011.11.003
Wu, L., Diny, N. L., Ong, S., Barin, J. G., Hou, X., Rose, N. R., et al. (2016). Pathogenic IL-23 signaling is required to initiate GM-CSF-driven autoimmune myocarditis in mice. Eur. J. Immunol. 46 (3), 582–592. doi: 10.1002/eji.201545924
Xu, P., Ji, L., Tian, S., Li, F. (2018). Clinical effects of tanshinone IIA sodium sulfonate combined with trimetazidine and levocarnitine in the treatment of AVMC and its effects on serum TNF-α, IL-18 and IL-35. Exp. Ther. Med. 16 (5), 4070–4074. doi: 10.13194/j.issn.1673-842x.2016.02.064
Yan, X., Shichita, T., Katsumata, Y., Matsuhashi, T., Ito, H., Ito, K., et al. (2012). Deleterious effect of the IL-23/IL-17A axis and γδT cells on left ventricular remodeling after myocardial infarction. J. Am. Heart Assoc. 1 (5), e004408. doi: 10.1161/JAHA.112.004408
Yan, X., Zhang, H., Fan, Q., Hu, J., Tao, R., Chen, Q., et al. (2017). Dectin-2 deficiency modulates Th1 differentiation and improves wound healing after myocardial infarction. Circ. Res. 120 (7), 1116–1129. doi: 10.1161/CIRCRESAHA.116.310260
Yang, F., Wu, W. F., Yan, Y. L., Pang, Y., Kong, Q., Huang, Y. L. (2011). Expression of IL-23/Th17 pathway in a murine model of Coxsackie virus B3-induced viral myocarditis. Virol. J. 8, 301. doi: 10.1186/1743-422X-8-301
Ye, J., Wang, Y., Wang, Z., Ji, Q., Huang, Y., Zeng, T., et al. (2018a). Circulating Th1, Th2, Th9, Th17, Th22, and Treg levels in aortic dissection patients. Mediators Inflamm. 2018, 5697149. doi: 10.1155/2018/5697149
Ye, J., Huang, Y., Que, B., Chang, C., Liu, W., Hu, H., et al. (2018b). Interleukin-12p35 knock out aggravates doxorubicin-induced cardiac injury and dysfunction by aggravating the inflammatory response, oxidative stress, apoptosis and autophagy in mice. EBioMedicine 35, 29–39. doi: 10.1016/j.ebiom.2018.06.009
Ye, J., Wang, Z., Huang, Y., Wang, M., Que, B., Ye, D., et al. (2018c). The clinical potential of IL-12/IL-35 in treating chemotherapy drug-induced cardiac injury - authors' reply. EBioMedicine 35, 4–5. doi: 10.1016/j.ebiom.2018.08.049
Ye, J., Que, B., Huang, Y., Lin, Y., Chen, J., Liu, L., et al. (2019). Interleukin-12p35 knockout promotes macrophage differentiation, aggravates vascular dysfunction, and elevates blood pressure in angiotensin II-infused mice. Cardiovasc. Res. 115 (6), 1102–1113. doi: 10.1093/cvr/cvy263
Ye, J., Wang, Y., Wang, Z., Liu, L., Yang, Z., Ye, D., et al. (2020). Interleukin-12p35 deficiency enhances mitochondrial dysfunction and aggravates cardiac remodeling in aging mice. Aging (Albany NY). 11. doi: 10.18632/aging.102609
Yi, A., Jian, L., Xiaojing, H., Hui, X. (2009). The prevalence of Th17 cells in patients with dilated cardiomyopathy. Clin. Invest. Med. 32 (2), E144–E150. doi: 10.25011/cim.v32i2.6032
Yong, K., Dogra, G., Boudville, N., Chan, D., Adams, L., Ching, H., et al. (2013). Interleukin-12 is associated with arterial stiffness in healthy individuals. Am. J. Hypertens. 26 (2), 159–162. doi: 10.1093/ajh/hps032
Zafra, G., Morillo, C., Martín, J., González, A., González, C. I. (2007). Polymorphism in the 3' UTR of the IL12B gene is associated with Chagas' disease cardiomyopathy. Microbes Infect. 9 (9), 1049–1052. doi: 10.1016/j.micinf.2007.04.010
Zha, X., Yue, Y., Dong, N., Xiong, S. (2015). Endoplasmic reticulum stress aggravates viral myocarditis by raising inflammation through the IRE1-associated NF-κB pathway. Can. J. Cardiol. 31 (8), 1032–1040. doi: 10.1016/j.cjca.2015.03.003
Zhang, M., Cai, Z. R., Zhang, B., Cai, X., Li, W., Guo, Z., et al. (2014a). Functional polymorphisms in interleukin-23 receptor and susceptibility to coronary artery disease. DNA Cell Biol. 33 (12), 891–897. doi: 10.1089/dna.2014.2573
Zhang, A., Mao, X., Li, L., Tong, Y., Huang, Y., Lan, Y., et al. (2014b). Necrostatin-1 inhibits Hmgb1-IL-23/IL-17 pathway and attenuates cardiac ischemia reperfusion injury. Transpl. Int. 27 (10), 1077–1085. doi: 10.1111/tri.12349
Zhang, D., Ma, M., Yang, Y., Wan, L., Yang, Z., Lv, J., et al. (2016). Association between polymorphisms in IL27 and risk for CHD in a Chinese population. Cardiol. Young. 26 (2), 237–243. doi: 10.1017/S1047951115000037
Zhang, H., Yue, Y., Sun, T., Wu, X., Xiong, S. (2017). Transmissible endoplasmic reticulum stress from myocardiocytes to macrophages is pivotal for the pathogenesis of CVB3-induced viral myocarditis. Sci. Rep. 7, 42162. doi: 10.1038/srep42162
Zhou, R. H., Shi, Q., Gao, H. Q., Shen, B. J. (2001). Changes in serum interleukin-8 and interleukin-12 levels in patients with ischemic heart disease in a Chinese population. J. Atheroscler. Thromb. 8 (1), 30–32. doi: 10.5551/jat1994.8.30
Zhu, H., Li, J., Wang, S., Liu, K., Wang, L., Huang, L. (2013). Hmgb1-TLR4-IL-23-IL-17A axis promote ischemia-reperfusion injury in a cardiac transplantation model. Transplantation 95 (12), 1448–1454. doi: 10.1097/TP.0b013e318293b7e1
Zhu, H., Cui, D., Liu, K., Wang, L., Huang, L., Li, J. (2014). Long pentraxin PTX3 attenuates ischemia reperfusion injury in a cardiac transplantation model. Transpl. Int. 27 (1), 87–95. doi: 10.1111/tri.12197
Zhu, H., Lou, C., Liu, P. (2015). Interleukin-27 ameliorates coxsackievirus-B3-induced viral myocarditis by inhibiting Th17 cells. Virol. J. 12, 189. doi: 10.1186/s12985-015-0418-x
Zhu, Z., Zhang, Y., Ye, J., Wang, X., Fu, X., Yin, Y., et al. (2018). IL-35 promoted STAT3 phosphorylation and IL-10 production in B cells, but its production was reduced in patients with coronary artery diseases. Hum. Immunol. 79 (12), 869–875. doi: 10.1016/j.humimm.2018.10.009
Keywords: cardiovascular diseases, IL-12 family members, atherosclerosis, coronary artery disease, hypertension, aortic dissection, viral myocarditis
Citation: Ye J, Wang Y, Wang Z, Liu L, Yang Z, Wang M, Xu Y, Ye D, Zhang J, Lin Y, Ji Q and Wan J (2020) Roles and Mechanisms of Interleukin-12 Family Members in Cardiovascular Diseases: Opportunities and Challenges. Front. Pharmacol. 11:129. doi: 10.3389/fphar.2020.00129
Received: 28 November 2019; Accepted: 30 January 2020;
Published: 04 March 2020.
Edited by:
Chen Huei Leo, Singapore University of Technology and Design, SingaporeReviewed by:
Carlos F. Sánchez-Ferrer, Autonomous University of Madrid, SpainCees Korstanje, KorstanjePharmaConsultancy, Netherlands
Copyright © 2020 Ye, Wang, Wang, Liu, Yang, Wang, Xu, Ye, Zhang, Lin, Ji and Wan. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Yingzhong Lin, eWluZ3pob25nbGluQDEyNi5jb20=; Qingwei Ji, anF3MTI0QDE2My5jb20=; Jun Wan, d2h1d2FuanVuQDE2My5jb20=; d2FuanVuQHdodS5lZHUuY24=
†These authors have contributed equally to this work