
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Pharmacol. , 08 March 2019
Sec. Translational Pharmacology
Volume 10 - 2019 | https://doi.org/10.3389/fphar.2019.00206
This article is part of the Research Topic Celebrating 40 Years of the Chilean Society of Pharmacology View all 34 articles
 María A. Lavanderos1
María A. Lavanderos1 Juan P. Cayún1
Juan P. Cayún1 Ángela Roco1,2Christopher Sandoval1Leslie Cerpa1
Ángela Roco1,2Christopher Sandoval1Leslie Cerpa1 Juan C. Rubilar1Roberto Cerro1Sebastián Molina-Mellico1
Juan C. Rubilar1Roberto Cerro1Sebastián Molina-Mellico1 Cesar Celedón1Berta Cerda3
Cesar Celedón1Berta Cerda3 Elena García-Martín4
Elena García-Martín4 José A. G. Agúndez4Cristián Acevedo1,5Karina Peña6
José A. G. Agúndez4Cristián Acevedo1,5Karina Peña6 Dante D. Cáceres1,7
Dante D. Cáceres1,7 Nelson M. Varela1*
Nelson M. Varela1* Luis A. Quiñones1*
Luis A. Quiñones1*Testicular cancer is one of the most commonly occurring malignant tumors in young men with fourfold higher rate of incidence and threefold higher mortality rates in Chile than the average global rates. Surgery is the initial line of treatment for testicular cancers, and is generally followed by chemotherapy, usually with combinations of bleomycin, etoposide, and cisplatin (BEP). However, the adverse effects of chemotherapy vary significantly among individuals; therefore, the present study explored the association of functionally significant allelic variations in genes related to the pharmacokinetics/pharmacodynamics of BEP and DNA repair enzymes with chemotherapy-induced toxicity in BEP-treated testicular cancer patients. We prospectively recruited 119 patients diagnosed with testicular cancer from 2010 to 2017. Genetic polymorphisms were analyzed using PCR and/or qPCR with TaqMan®probes. Toxicity was evaluated based on the Common Terminology Criteria for Adverse Events, v4.03. After univariate analyses to define more relevant genetic variants (p < 0.2) and clinical conditions in relation to severe (III–IV) adverse drug reactions (ADRs), stepwise forward multivariate logistic regression analyses were performed. As expected, the main severe ADRs associated with the non-genetic variables were hematological (neutropenia and leukopenia). Univariate statistical analyses revealed that patients with ERCC2 rs13181 T/G and/or CYP3A4 rs2740574 A/G genotypes are more likely to develop alopecia; patients with ERCC2 rs238406 C/C genotype may develop leukopenia, and patients with GSTT1-null genotype could develop lymphocytopenia (III–IV). Patients with ERCC2 rs1799793 A/A were at risk of developing severe anemia. The BLMH rs1050565 G/G genotype was found to be associated with pain, and the GSTP1 G/G genotype was linked infection (p < 0.05). Multivariate analysis showed an association between specific ERCC1/2 genotypes and cumulative dose of BEP drugs with the appearance of severe leukopenia and/or febrile neutropenia. Grades III–IV vomiting, nausea, and alopecia could be partly explained by the presence of specific ERCC1/2, MDR1, GSTP1, and BLMH genotypes (p < 0.05). Hence, we provide evidence for the usefulness of pharmacogenetics as a tool for predicting severe ADRs in testicular cancer patients treated with BEP chemotherapy.
Testicular cancers are malignant cancers that mainly affect young men. Cisplatin-based chemotherapy has been routinely used as the standard therapy for the treatment of metastatic testicular cancers. The standard treatment scheme for patients with low-risk testicular cancers involves three cycles of bleomycin, etoposide, and cisplatin (BEP) or four cycles of etoposide and cisplatin. Patients with intermediate-risk or high-risk disease are routinely treated with up to four cycles of BEP or four cycles of etoposide, ifosfamide, and cisplatin. Chemotherapy for rescuing the relapse of testicular cancers includes a standard dose of vinblastine, cisplatin and ifosfamide (Pizzocaro et al., 1985; Mezvrishvili and Managadze, 2006).
However, chemotherapy represents a significant challenge in the day-to-day management of the patients, since the inter-individual variations in response to the chemotherapy drugs are a major cause of concern. A drug that is well-tolerated and causes a strong response in some patients could prove to be ineffective, toxic or cause adverse drug reactions (ADRs) in others; therefore, research is required to analyze the effects of genetic variations on the pharmacokinetics and/or pharmacodynamics of these drugs. Statistics indicate that 1 in 15 hospital admissions for testicular cancer in the United Kingdom was due to ADRs (Pirmohamed et al., 2004), and adverse drug effects in hospitalized patients were identified to be the fifth leading cause of death in the United States (Mancinelli et al., 2000). Other evidence suggests that the annual number of reported cases of adverse reactions is around two millions which has been reported to cost US $100 billion (Ross et al., 2011). The antineoplastic drugs have often been shown to exhibit toxicity at therapeutic concentrations; therefore, ADRs are frequently observed during chemotherapy, which has reportedly increased the total medical costs by 1.9%, and the costs of medicines by 15% (Huang and Ratain, 2009).
Response to chemotherapy may be determined by gene polymorphisms, which eventually govern the metabolism of cytotoxic drugs. The allelic variants of genes related with pharmacokinetics/pharmacodynamics processes can alter the enzyme activity of the metabolic proteins leading to changes in drug metabolism (Agundez, 2004; Quiñones et al., 2017). Therefore, the response to chemotherapy in patients may be partly determined by gene polymorphisms involved in the metabolism of these cytotoxic drugs. Most of the chemotherapeutic drugs are metabolized by phase I polymorphic Cytochrome P450 enzymes, whose variant alleles commonly affect drug effectiveness and toxicity (Kivistö et al., 1995; Quiñones et al., 2008; Božina et al., 2009; Ingelman-Sundberg and Sim, 2010; Deenen et al., 2011). Cytochrome P450 isoforms 3A4 and 3A5 (CYP3A4/5) play a role in etoposide metabolism (Zhuo et al., 2004). The CYP3A4∗1B rs2740574 genotype is associated with an increased risk of leukemia following treatment with etoposide and teniposide. This variant has been reported to contribute to higher risk of secondary cancers (Felix et al., 1998). Moreover, two studies have also suggested that CYP3A4∗1B is a risk factor allele for prostate cancer (Keshava et al., 2004).
On the other hand, polymorphisms in phase II enzymes have been reported to affect the resistance and adverse reactions to several chemotherapy regimens (Jain et al., 2007; Mazerska et al., 2016; Marchewka et al., 2017). Previous reports have shown that Glutathione-S-transferases (GSTs) are associated with resistance to cisplatin-based chemotherapy (Roco et al., 2014; Nissar et al., 2017). GSTM1-null and/or GSTT1-null genotypes are associated with the development of grades III–IV thrombocytopenia (Cho et al., 2010) upon combined chemotherapy with rituximab and cyclophosphamide/doxorubicin/vincristine/prednisone or R-CHOP. Grade ≥ III toxicity and grade ≥ III neurotoxicity has been observed in children with medulloblastoma treated with cycles of cisplatin, cyclophosphamide, and vincristine (Barahmani et al., 2009). Besides, the GSTP1 Ile105Val polymorphism has been strongly associated with progression-free survival. The T/T genotype of the −69 C > T GSTA1 polymorphism correlates with overall survival. Thrombocytopenia, anemia, and neuropathy were less frequent among patients with the GSTM1-null or GSTM3 intron 6 AGG/AGG genotypes (Khrunin et al., 2010). Moreover, the presence of UDP-Glucuronosyl-transferase Family 1 Member A1 (UGT1A1) polymorphic variants has also been associated with chemotherapy response and Gilbert Syndrome after chemotherapy (Ha et al., 2017; Negoro et al., 2018). The most studied UGT1A1 allele is UGT1A1∗28, which has mainly been associated with an increased risk of irinotecan toxicity (Lyer et al., 2002). In line with this evidence, the FDA recommended tests to detect the presence of UGT1A1∗28, to predict patients at risk of irinotecan poisoning1.
ABC-drug transporters are also implicated in the metabolic response to chemotherapy (Domenichini et al., 2019). P-glycoprotein 1 (P-gp1), also known as multidrug resistance protein 1 (MDR1) or ATP- binding cassette sub-family B member 1, is highly polymorphic and several studies have reported that carriers of the T-allele for the genetic variation C3435T (rs1045642) have an increased risk of colon, breast, and renal cancer (Phuthong et al., 2017). However, Cizmarikova et al. (2010), found no significant differences in hematological toxicities in the groups with the MDR1 C3435T polymorphism in breast cancer.
On the other hand, bleomycin (BLM) is metabolically inactivated by the action of cysteine peptidase bleomycin hydrolase (BLMH) (Schwartz et al., 1999; Deenen et al., 2011). de Haas et al. (2008), showed that BLMH A/G genotype was related with reduced survival and higher prevalence of early relapses in testicular cancer patients. Recently, Jóna et al. (2016), showed lower rate of pulmonary complications in the A/A genotype group than those in the group containing the mutated allele: A/G+G/G in ABVD (doxorubicin, bleomycin, vinblastine, and dacarbazine)-treated Hodgkin lymphoma patients.
Several antineoplastic drugs have been reported to cause DNA damage. Numerous studies have investigated the association between single nucleotide polymorphisms (SNPs) in DNA repair enzymes, clinical outcomes, and resistance to chemotherapy (Zamble and Lippard, 1995; Gossage and Madhusudan, 2007; Frosina, 2009; Zhang et al., 2017) indicating that reduced activity of DNA repair enzymes may lead to an improved response to chemotherapy. However, compromised repair may also lead to accumulation of DNA damage in normal cells, leading to adverse side effects in normal tissues, thereby predisposing them toward secondary cancers. Due to these side effects, commonly used chemotherapeutic agents, including alkylating agents (cisplatin), inhibitors of DNA topoisomerase II (etoposide), and BLM have proven to be toxic to the patients.
Excision repair cross-complementary 1 (ERCC1) is a key protein involved in nucleotide excision repair (NER), and ERCC1-Xeroderma pigmentosum (ERCC1-XPF) catalyzes the incision on the site of DNA damage (Park et al., 1995). Elevated ERCC1 expression in cancers is associated with resistance to DNA damage-based chemotherapy (Chiu et al., 2011; Tsai et al., 2011). On the other hand, Xeroderma pigmentosum complementary group D (XPD/ERCC2) encodes a helicase which participates in both NER and basal transcription as part of the transcription factor IIH. Mutations abolishing the enzymatic function of the ERCC2 protein are manifested clinically in combinations of three severe syndromes, including Xeroderma pigmentosum (Lehmann, 2001; Clarkson and Wood, 2005). Polymorphisms in these enzymes further affect DNA repair and are involved in resistance to chemotherapy, survival, and cancer manifestation (Benhamou and Sarasin, 2002; Giovannetti et al., 2011).
Based on the accumulated scientific evidence about BEP chemotherapy, we here focused on functionally significant gene polymorphisms in proteins that control metabolism, uptake, and response to BEP drugs (2Roco et al., 2014; Chen et al., 2017). Mainly, the allelic variants of CYP3A4 rs2740574 (∗1B), CYP3A4 rs35599367 (∗22), CYP3A5 rs776746 (∗3), GSTM1-null, GSTP1 rs1695, GSTT1- null, UGT1A1 rs8175347 (∗28), BLMH rs1050565, ERCC1 rs11615, ERCC1 rs3212986, ERCC2 rs13181, ERCC2 rs1799793, ERCC2 rs238406, and MDR1 rs1045642 were analyzed with non-genetic factors to validate their association with ADRs in testicular cancer patients treated with BEP schedule.
One hundred nineteen (119) patients with germinal (seminoma or non-seminoma) testicular cancer confirmed histologically, >18 years old, without chronic unbalanced or systemic pathology or other active cancers and without being included in the interventional study conducted 3 months before, were enrolled prospectively in this study. The enrollment was carried out from December 2010 – December 2017 at the Polyclinic of Hematology-oncology of Hospital San Juan de Dios, and the Polyclinic of Oncology of the National institute of Cancer. All the patients signed a written consent and an agreement to be included in this study. The study was carried out under strict ethical procedures recommended by the Ethics Committee of the University of Chile (August 17, 2010), and the Northern Metropolitan Health Service, National Cancer Institute (April 4, 2015), in accordance with the procedures suggested in the Declaration of Helsinki (Declaration of Helsinki, 1964), and according to Chilean Laws 20.120, 20.584, and 19.628, and the guidelines of the Good Clinical Practices. Chemotherapy regimen for all patients involved the administration of cisplatin and etoposide in combination with bleomycin for up to four cycles (BEP schedule), which is the standard treatment for patients with low or intermediate risk testicular cancers, all over the world (Pizzocaro et al., 1985; Mezvrishvili and Managadze, 2006).
This regimen was applied in conjunction with a rigorous and standardized hydration regimen for all the patients. The clinical variables were obtained from patients’ clinical files and recorded in proper case report forms (CRFs). Laboratory assessment was performed after each cycle of chemotherapy. Treatment-related toxicity was graded according to the terminological common criteria for adverse reactions (v4.03), of the U.S. Department of Health and Human Services with a follow-up after 6 months of the last cycle of chemotherapy. The association study included age, height, weight, body surface area, and cumulative doses for the statistical analyses. Table 1 shows the general characteristics of the studied patients.
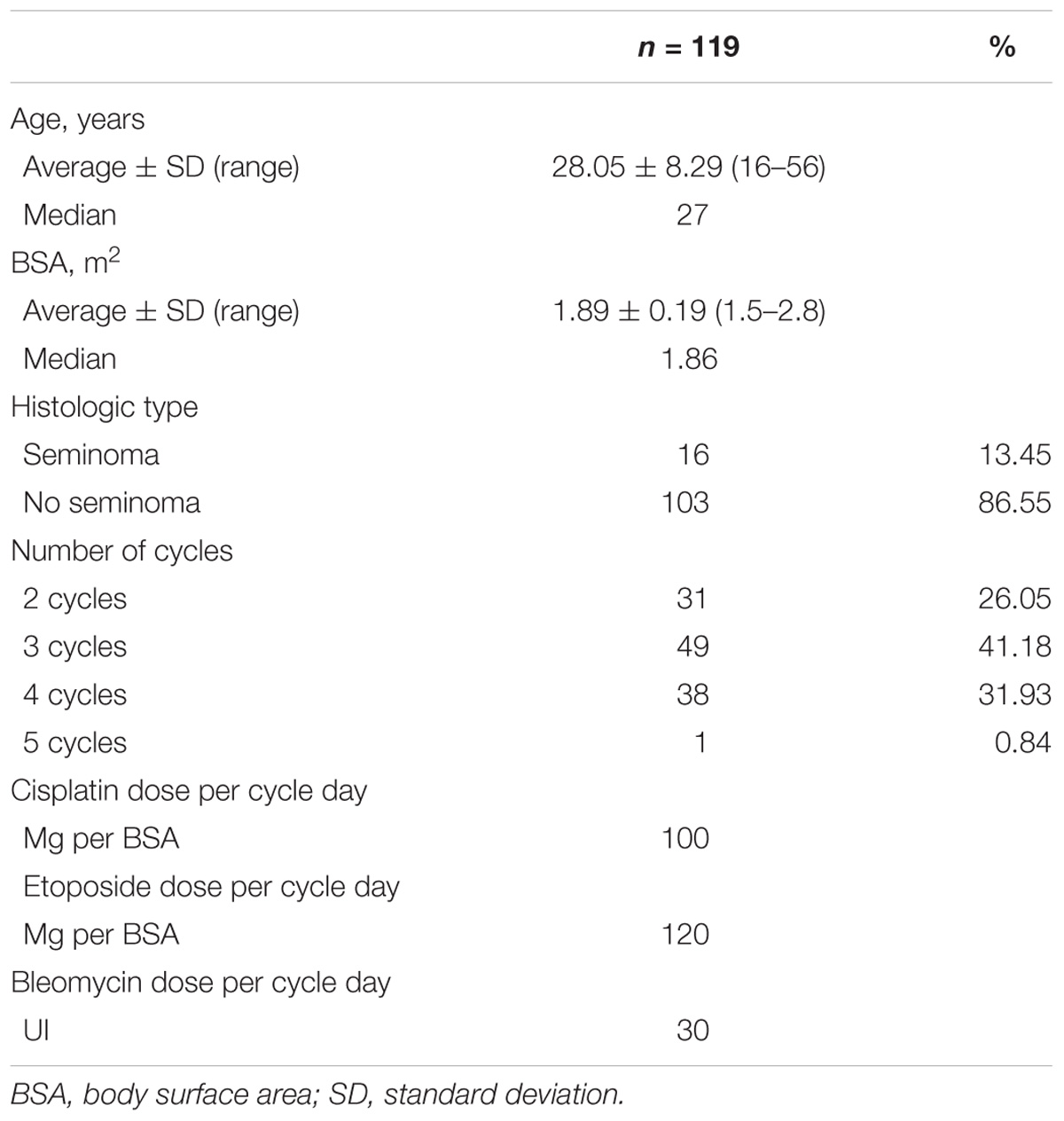
Table 1. Baseline characteristics of patients.
Potentially functional SNPs encoding the proteins related to BEP response were obtained from the NCBI dbSNP database3, and the SNPinfo Web Server4 based on the level of evidence for each SNP (Supplementary Table S1). Genomic DNA was isolated from the peripheral blood samples of the subjects using High Pure PCR Template Preparation Kit (Catalog Number, 11796828001; Roche Diagnostics GmbH, Mannheim, Germany). CYP3A4∗1B rs2740574, CYP3A4∗22 rs35599367, CYP3A5∗3 rs776746, UGT1A1∗28 rs8175347, BLMH rs1050565, GSTP1 rs1695, ERCC1 rs11615, ERCC1 rs321986, ERCC2 rs13181, ERCC2 rs238406, ERCC2 rs1799793, and MDR1 rs1045642 were analyzed using TaqMan®SNP Genotyping Assay (Catalog number, 4362691; Thermo Fisher Scientific, Waltham, MA, United States), in an Stratagene Mx3000p real-time PCR system (Agilent Technologies, Santa Clara, CA, United States). The presence of the GSTM1-null genotype was determined by the absence of a 273 bp fragment product in a 2% agarose gel (Bio-Rad Laboratories, Hercules, CA, United States). The presence of the GSTT1-null genotype was determined by the absence of a 268 bp fragment. Amplification fragment for β-globin was used as the internal control (Quiñones et al., 1999; Roco et al., 2012). Heterozygous and homozygous non-null individuals could not be differentiated, therefore double null genotypes (−/−) are the null genotypes reported. For Quality Assurance purposes we randomly choose 20% of the samples for (a) repetition of the analysis and (b) PCR-RFLP analysis for coincidence. When analyses were not coincident we excluded the samples. The sequences for TaqMan®probes and primers for PCR are listed in Supplementary Tables S2, S3 enlists the description of each polymorphism.
We performed a logistic regression analysis using Stata software, version 12.0 (Copyright 1985–2011 StataCorp., LP, College Station, TX, United States). A p-value of ≤ 0.05 was considered statistically significant. The odds ratio (OR) and 95% confidence intervals (CI) were reported in the univariate and multivariate logistic regression models. The logistic multivariate models were adjusted stepwise using a forward procedure with p-value ≤ 0.2 to include potentially relevant variables in order to derive statistical association models, characterized by Pseudo R2. All association studies were assayed by testing three genetic models of inheritance, i.e., dominant, codominant and recessive models, and choosing parameters with better statistical association for each analysis.
For the univariate and multivariate analyses, we define several alternatives for dosage: Ranges for dose: to get the ranges we used quartiles (Q) to divide data in four groups, with lower range comprising Q0 to the average between Q1 and Q2, intermediate range comprising of the average between Q1 and Q2 to the average between Q2 and Q3 and the largest range comprising of the average between Q2 and Q3 to Q4. Cumulative dose was defined as the total dose administered to the patient during all the cycles of chemotherapy. Cumulative dose by average: the patients were divided in two groups according to their were lower/equal in relation the average cumulative dose (<average) or higher than the average cumulative dose (>average). Similar procedure was performed for weight, height, body surface and age and for chemotherapy cycles with frequency of 1–2, 3 and 4–5 cycles, getting dummy variables for the statistical analyses.
We did not check Hardy–Weinberg equilibrium (HWE) of our sample because it does not accomplish the conditions for HWE. This is not a random sampling in a random-mating population, a control or general population (Namipashaki et al., 2015) and is a group with a selection bias by the disease (i.e., SNPs can also be related to the cancer).
The baseline characteristics of patients are shown in Table 1. A total of 119 Chilean patients from two hospitals in Chile were included and analyzed. Most patients were young (average age: 28.05 years), and were administered 2–5 cycles of BEP and predominantly showed no seminoma (86.55%). The genotypic and allelic frequencies for the analyzed polymorphisms are shown in Supplementary Table S4.
Adverse drug reactions represented in Table 2 were recorded to determine the acute toxicity in patients with testicular cancer treated with BEP chemotherapy. The ADRs are shown in two columns, any grades (I–IV) column and severe or high-grade toxicities (III–IV) column. The most frequent toxicities observed included vomiting (82.35%), nausea (79.83%), anemia (60.68%), neutropenia (53.45%), and alopecia (52.94%). Most frequently observed high-grade toxicities included neutropenia (39.66%), leukopenia (12.71%), febrile neutropenia (12.61%), and vomiting (9.24%).
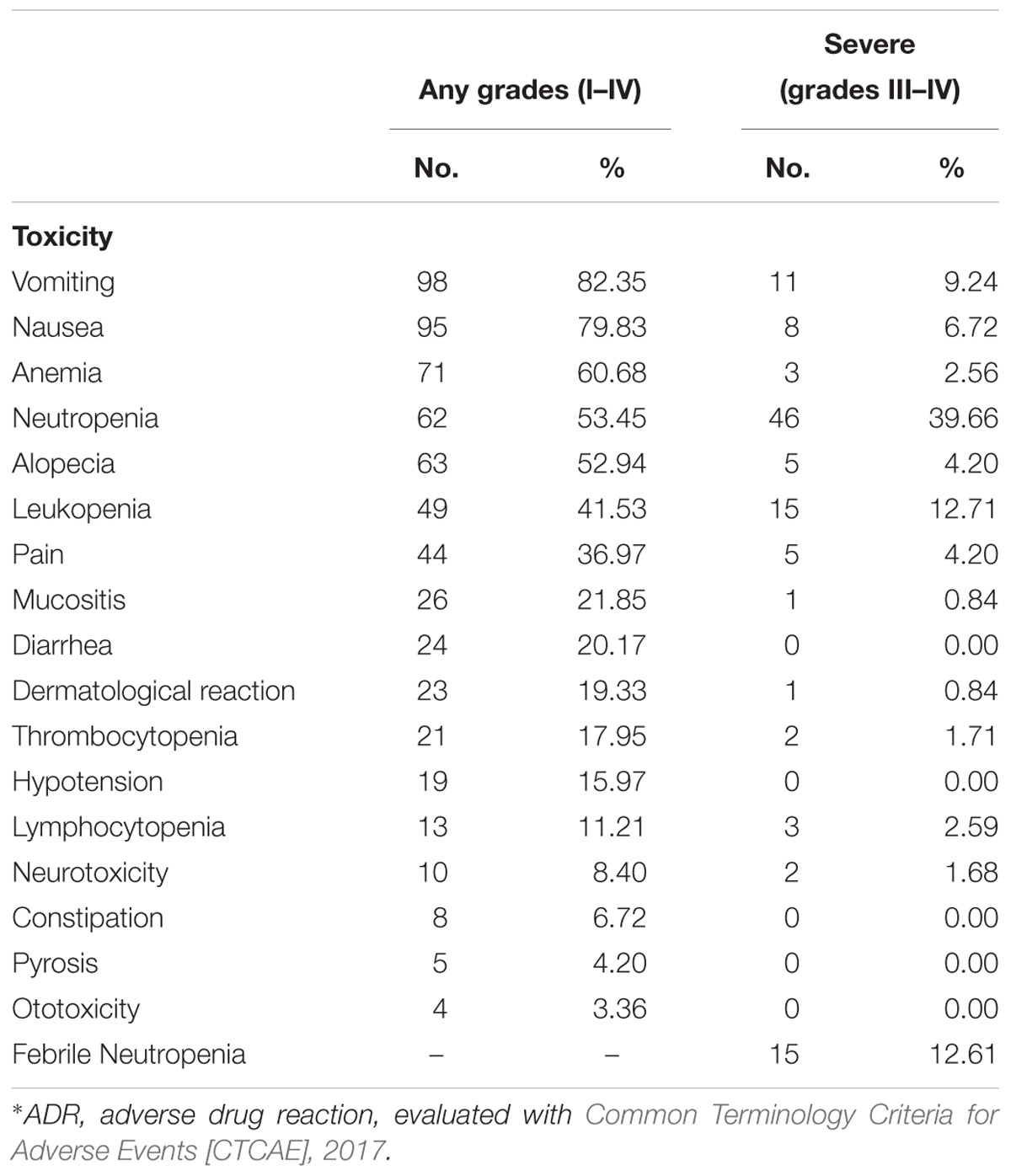
Table 2. Adverse drug reactions (ADRs) according degree of severity∗.
We performed univariate logistic regression of risk for severe (III–IV) ADRs in association with genotypes, in three models of inheritance namely, recessive, codominant, and dominant. The results are shown in Supplementary Table S5 where only results with p-value ≤ 0.2 are included for the stepwise forward procedure for multivariate analysis. In Table 3, only statistically significant results for the univariate logistic regression analysis of risk of severe ADRs (III–IV), according to genotypes are shown. These results show that ERCC2 rs1799793 A/A genotype was associated with anemia in a recessive model of inheritance, ERCC2 rs13181 T/G and CYP3A4 rs2740574 A/G genotypes were associated with alopecia in a codominant model of inheritance, and ERCC2 rs238406 A/A genotype was associated with leukopenia, both in codominant and recessive models of inheritance. GSTT1-null genotype was associated with lymphocytopenia, BLMH rs1050565 G/G genotype was linked with pain in a recessive model of inheritance and GSTP1 rs1695 G/G genotype was associated with infections in a recessive model of inheritance.
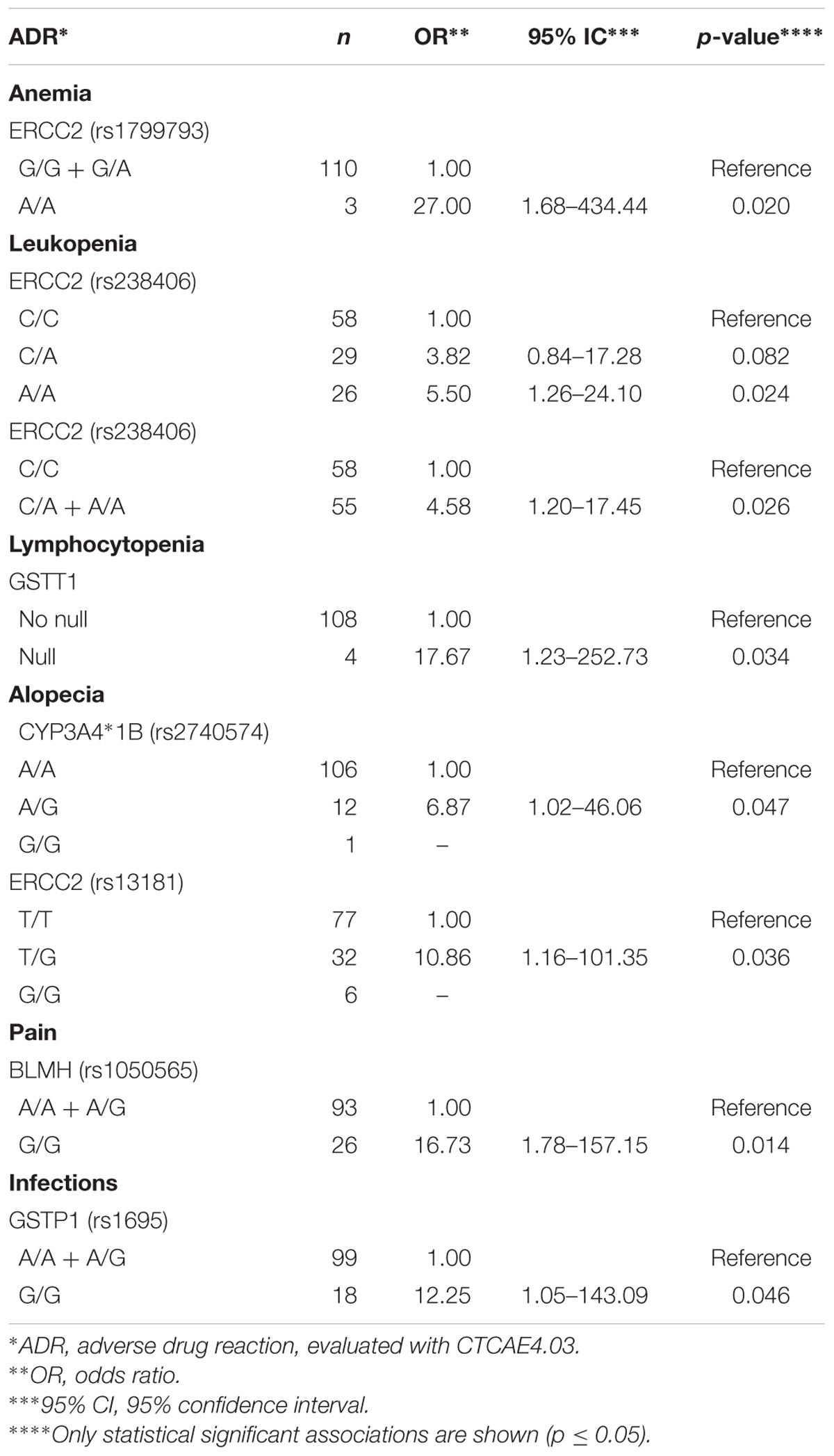
Table 3. Univariate logistic regression analysis of risk of severe ADRs (III–IV) according to genotypes.
The same analysis was performed for non-genetic factors (e.g., age, sex, weight, height, body surface, cycles, and cumulative dose) (Supplementary Table S6). Table 4 shows only the statistically significant results obtained from the univariate logistic regression analysis of risk of severe ADRs (III–IV), analyzed according to non-genetic factors. We observed that the cumulative dose of bleomycin; bleomycin dose by average or bleomycin cycles by range were associate with febrile neutropenia as well as cisplatin dose by range. Similarly, cumulative total etoposide dose or dose by average were associated with leukopenia as well as the cumulative dose of cisplatin. Besides, neutropenia was associated with both, cumulative or cumulative by average dose of cisplatin or etoposide, and cumulative bleomycin. Finally, alopecia was significantly associated only with cumulative and cumulative by average dose of bleomycin.

Table 4. Univariate logistic regression analysis of risk of severe ADRs (III–IV) according to non-genetic factors.
After stepwise forward procedure, using associations with a p < 0.2, multivariate logistic regression analyses for the risk of severe ADRs, including genetic and non-genetic factors, were performed. Table 5 show only statistically significant association models for severe ADRs. We obtained significant models for febrile neutropenia, leukopenia, vomiting, nausea, and alopecia.
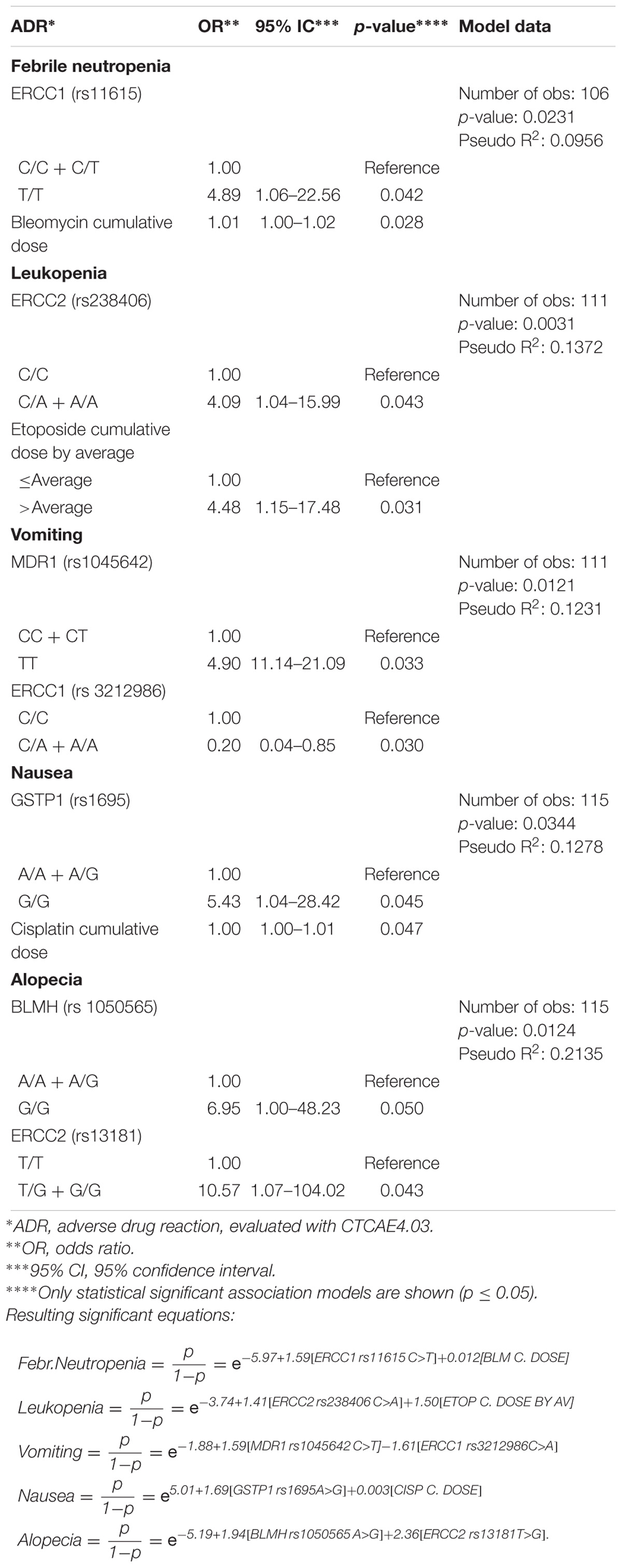
Table 5. Multivariate logistic regression analysis and risk of severe ADRs (grades III–IV), after stepwise forward procedure (cut-off p < 0.2).
Patient response to chemotherapy has been investigated for long, and ADR after chemotherapy is a substantial clinical problem. For testicular cancers, this is particularly relevant since besides surgery (inguinal orchiectomy) chemotherapy is routinely administered with combination of three cytostatic drugs, bleomycin, etoposide, and cisplatin. Even though chemotherapy is quite successful in the treatment for patients with seminoma and the success rates exceed 90%, adverse reactions are frequently observed in response to one of the drugs or the drug combination. Therefore, in the present study, we have evaluated the role of genetic polymorphisms and other non-genetic factors as potential modifying risk factors for ADRs.
In the univariate analyses (Table 3), we found interesting association between BLMH rs1050565 G/G genotype and severe pain in patients (OR = 16.73, CI = 1.78–157.15, p-value = 0.014). Our observation is in line with the report from White and coworkers who showed an association between acute chest pain and bleomycin infusion (White et al., 1987). Considering that G allele of BLMH leads to the incorporation of 443Val in the enzyme, reducing its biochemical activity, this association supports our finding and high bleomycin plasma levels in patients can be expected.
On the other hand, patients with CYP3A4 rs2740574 A/G genotypes are more likely to develop alopecia (OR = 6.87, CI = 1.02–46.06, p-value = 0.047). This gene encodes for the main enzyme involved in etoposide metabolism2, and metabolizes cisplatin or bleomycin. The presence of G allele leads to reduced transcription of the enzyme, suggesting a relationship between dose and increased plasma levels of etoposide and alopecia.
Cisplatin mainly reacts with N-7 of guanine and adenine to form adducts with the DNA (Kelland, 2007) resulting in the formation of intra and inter strands crosslinks, causing potential errors in DNA repair, resulting in accumulation of damaged DNA, and activation of apoptotic pathway in neoplastic and normal cells. Therefore, it was important to analyze both, the drug-metabolizing enzymes (GSTs) and the DNA damage repair proteins. We observed that GSTT1-null genotype is associated with lymphocytopenia (OR = 17.67, CI = 1.23–252.73, p-value = 0.034) and GSTP1 rs1695 G/G genotype is associated with increased infections (OR = 12.25, CI = 1.05–143.09, p-value = 0.046). Similarly, ERCC2 rs1799793 A/A genotype showed association with anemia (OR = 27.00, CI = 1.68–434.44, p-value = 0.020), ERCC2 rs238406 A/A genotype was associated with leukopenia (OR = 5.5, CI = 1.26–24.10, p-value = 0.024) and ERCC2 rs13181 T/G genotype was linked with alopecia (OR = 10.86, CI = 1.16–101.35, p-value = 0.036), indicating that defects in the metabolism and/or the response to cisplatin could lead to the specific severe ADRs (Table 3). This is in agreement with studies that report that G allele of GSTP1 rs1695 has been associated with an increased risk of myelosuppression, polyneuropathy, and toxicity (Joerger et al., 2012). Conversely, it has been found that the genotype GSTP1 A/A is predicted to show a suboptimal response to chemotherapy with fluorouracil/cisplatin, and a lower survival rate in patients with advanced gastric cancer (Ruzzo et al., 2006). Studies on GSTM1 and GSTT1 have shown that high expression levels of both enzymes result in a low response to chemotherapy, and deletion of these genes shows high degrees of toxicity (Bai et al., 1996; Ambrosone et al., 2001). In ovarian cancer patients, severe emesis grades III–IV were associated with GSTT1-null genotype (Khrunin et al., 2010). In contrast, in patients with GSTM1-null genotype the risk of thrombocytopenia and anemia was lower (Khrunin et al., 2010). For GSTP1, the G/G genotype seems to decrease the susceptibility to grade III neuropathy when compared to that in patients with A/G and/or A/A genotypes (recessive model of inheritance) in ovarian cancer (Khrunin et al., 2010). In this study, however, we did not find an association of these polymorphic enzymes with the above-mentioned adverse reactions. However, the effect of GST in hematological toxicity is reasonable, since GSTs are responsible for etoposide breakdown and elimination2.
The analysis of univariate associations among severe ADRs and non-genetic factors (Table 4) showed an association between cumulative dose of bleomycin, bleomycin dose by average and bleomycin cycles by range; and that these parameters are associated with febrile neutropenia, as well as cisplatin dose by range. Similarly, cumulative dose of etoposide (total or by average) is associated with leukopenia as well as the cumulative dose of cisplatin. Besides, neutropenia is associated with both, cumulative or cumulative by average dose of cisplatin or etoposide. These results are consistent with the studies that report the relationship between hematological ADRs and BEP drugs, particularly etoposide and cisplatin56. Moreover, alopecia was significantly associated only with dose of bleomycin (cumulative and by average).
Multivariate analyses to obtain risk association models of severe ADRs, including polymorphisms and non-genetic variables (Table 5) yielded good models to partly explain febrile neutropenia (Pseudo R2: 0.0956), leukopenia (Pseudo R2: 0.1372), vomiting (Pseudo R2: 0.1231), nausea (Pseudo R2: 0.1278), and alopecia (Pseudo R2: 0.2135). Interestingly, in these models, only dosage but not the demographic variables were relevant for severe ADRs.
For better understanding Figure 1 shows a scheme of this research and relevant results. After recruitment, genotyping and data collection from patients, genetic and non-genetic factors were submitted to logistic univariate statistical analyses. Then, logistic multivariate models were adjusted using a stepwise forward procedure with a cut-off p-value ≤ 0.2. The multivariate models are described by pseudo R2 values and equations for the obtained significant ADRs association models (febrile neutropenia, leukopenia, vomiting, nausea, and alopecia).
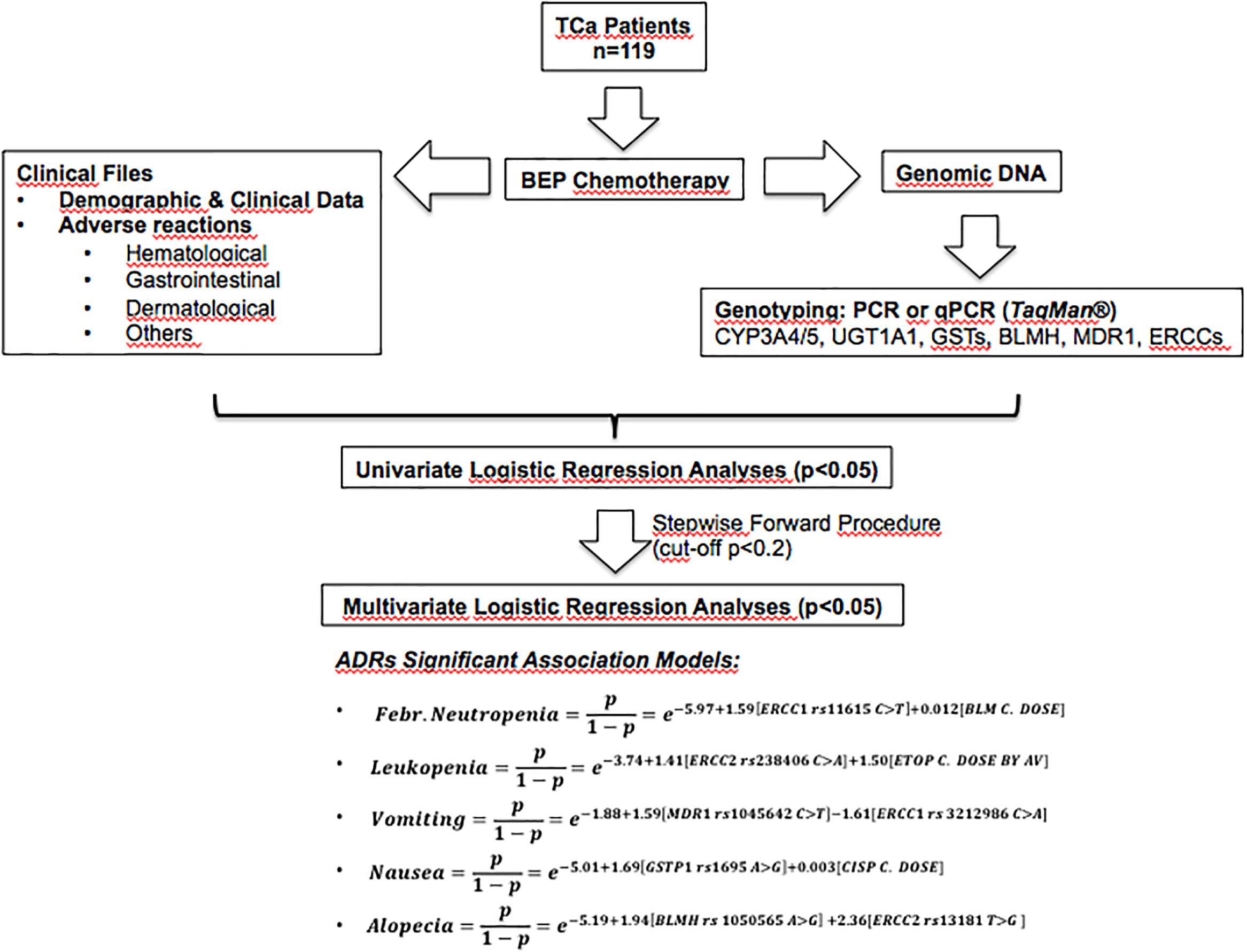
Figure 1. Scheme of this research and relevant results. BEP, bleomycin-etoposide-cisplatin therapy; ADRs, adverse drug reactions; TCa, testicular cancer.
Despite our analysis, the study has some shortcomings. Although we had a relatively appropriate sample size for combinatorial analyses, fewer number of patients examined could mask potential associations, especially for low frequency polymorphisms, particularly in the multivariate analyses. Some other potentially candidate genes/polymorphisms were not evaluated in this study (based in level of evidence), which could be still relevant. Besides, the cumulative doses were obtained at the end of the therapy, which could limit our conclusions about this factor in relation to ADRs. We were not able to analyze additional potential toxicities such as hepatotoxicity and nephrotoxicity due to incomplete clinical data. These, and others missing clinical values, could be relevant, giving rise to a possible differential misclassification bias affecting estimated associations between potentially relevant combinations of risk factors and adverse reactions. Finally, we did not adjust p-values for multiple tests (Bonferroni’s adjustment), which could generate direct implications in α and the p-value. However, it has been proposed that the adjustment is not always recommended, especially when high number of comparison are performed and multiple false negatives need to be avoided, which is the case (Goldman, 2008; Kim and Bang, 2016).
Our findings from the univariate analyses suggest that patients with ERCC2 rs13181 T/G and/or CYP3A4 rs2740574 A/G genotypes are more likely to develop grades III–IV alopecia; patients with ERCC2 rs238406 C/C genotype may develop severe leukopenia; and patients with GSTT1-null genotype could develop lymphocytopenia. Patients with ERCC2 rs1799793 A/A genotype were at higher risk of developing anemia. Patients with BLMH rs1050565 G/G genotype experienced severe pain, and patients with GSTP1 G/G genotype were susceptible to severe infections. As expected, severe ADRs associated with non-genetic variables were hematological (neutropenia and leukopenia). The multivariate analyses showed an association between specific ERCC1/2 genotypes and cumulative dose of BEP drugs with the appearance of severe leukopenia and/or febrile neutropenia. Grades III–IV vomiting, nausea and alopecia could also be partly explained by the presence of specific ERCC1/2, MDR1, GSTP1, and BLMH genotypes. Our study provides additional evidence for the use of pharmacogenetics as a useful tool for potential prediction of severe ADRs in testicular patients treated with BEP chemotherapy.
ML, JC, and NV: experimental analyses, analysis of data, and writing the manuscript. AR: experimental analyses and writing the manuscript. CS, LC, JR, RC, SM-M, and CC: experimental analyses. BC and KP: enrolment of patients. EG-M and JA: conception of the research. CA: analysis of data and enrolment of patients. DC: analysis of data. LQ: conception of the research, analysis of data, and writing the manuscript.
This work was supported by grant FONDECYT #1140434.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The authors wish to thank the testicular cancer patients from the “Instituto Nacional del Cáncer” and “San Juan de Dios” hospital for their altruistic collaboration in pursuit of the common welfare. They also thank the Latin American Society of Pharmacogenomics and Personalized Medicine (SOLFAGEM) by sponsoring this article.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fphar.2019.00206/full#supplementary-material
Agundez, J. A. G. (2004). Cytochrome P450 gene polymorphism and cancer. Curr. Drug Metab. 5, 211–224. doi: 10.2174/1389200043335621
Ambrosone, C. B., Sweeney, C., Coles, B. F., Thompson, P. A., McClure, G. Y., Korourian, S., et al. (2001). Polymorphisms in glutathione S-transferases (GSTM1 and GSTT1) and survival after treatment for breast cancer. Cancer Res. 61, 7130–7135.
Bai, F., Nakanishi, Y., Kawasaki, M., Takayama, K., Yatsunami, J., Pei, X. H., et al. (1996). Immunohistochemical expression of glutathione S-transferase-Pi can predict chemotherapy response in patients with nonsmall cell lung carcinoma. Cancer 78, 416–421. doi: 10.1002/(SICI)1097-0142(19960801)78:3<416::AID-CNCR6>3.0.CO;2-H
Barahmani, N., Carpentieri, S., Li, X. N., Wang, T., Cao, Y., Howe, L., et al. (2009). Glutathione S-transferase M1 and T1 polymorphisms may predict adverse effects after therapy in children with medulloblastoma. Neuro Oncol. 11, 292–300. doi: 10.1215/15228517-2008-089
Benhamou, S., and Sarasin, A. (2002). ERCC2/XPD gene polymorphisms and cancer risk. Mutagenesis 17, 463–469. doi: 10.1093/mutage/17.6.463
Božina, N., Bradamante, V., and Lovrić, M. (2009). Genetic polymorphism of metabolic enzymes P450 (CYP) as a susceptibility factor for drug response, toxicity, and cancer risk. Arh. Hig. Rada Toksikol. 60, 217–242. doi: 10.2478/10004-1254-60-2009-1885
Chen, Y., Zhang, H., and He, Q. (2017). Involvement of bleomycin hydrolase and poly(ADP-ribose) polymerase-1 in Ubc9-mediated resistance to chemotherapy agents. Int. J. Oncol. 50, 223–231. doi: 10.3892/ijo.2016.3777
Chiu, T. J., Chen, C. H., Chien, C. Y., Li, S. H., Tsai, H. T., and Chen, Y. J. (2011). High ERCC1 expression predicts cisplatin-based chemotherapy resistance and poor outcome in unresectable squamous cell carcinoma of head and neck in a betel-chewing area. J. Transl. Med. 9:31. doi: 10.1186/1479-5876-9-31
Cho, H. J., Eom, H. S., Kim, H. J., Kim, I. S., Lee, G. W., and Kong, S. Y. (2010). Glutathione-S-transferase genotypes influence the risk of chemotherapy-related toxicities and prognosis in Korean patients with diffuse large B-cell lymphoma. Cancer Genet. Cytogenet. 198, 40–46. doi: 10.1016/j.cancergencyto.2009.12.004
Cizmarikova, M., Wagnerova, M., Schonova, L., Habalova, V., Kohut, A., Linkova, A., et al. (2010). MDR1 (C3435T) polymorphism: relation to the risk of breast cancer and therapeutic outcome. Pharmacogenomics J. 10, 62–69. doi: 10.1038/tpj.2009.41
Clarkson, S. G., and Wood, R. D. (2005). Polymorphisms in the human XPD (ERCC2) gene, DNA repair capacity and cancer susceptibility: an appraisal. DNA Repair 4, 1068–1074. doi: 10.1016/j.dnarep.2005.07.001
Common Terminology Criteria for Adverse Events [CTCAE] (2017). Cancer Therapy Evaluation Program, Common Terminology Criteria for Adverse Events, Version 3.0, DCTD, NCI, NIH, DHHS March 31, 2003. Available at: http://ctep.cancer.gov
de Haas, E. C., Zwart, N., Meijer, C., Nuver, J., Boezen, H. M., Suurmeijer, A. J., et al. (2008). Variation in bleomycin hydrolase gene is associated with reduced survival after chemotherapy for testicular germ cell cancer. J. Clin. Oncol. 26, 1817–1823. doi: 10.1200/JCO.2007.14.1606
Deenen, M. J., Cats, A., Beijnen, J. H., and Schellens, J. H. (2011). Part 2: pharmacogenetic variability in drug transport and phase I anticancer drug metabolism. Oncologist 16, 820–834. doi: 10.1634/theoncologist.2010-0259
Domenichini, A., Adamska, A., and Falasca, M. (2019). ABC transporters as cancer drivers: potential functions in cancer development. Biochim. Biophys. Acta. Gen. Subj. 1863, 52–60. doi: 10.1016/j.bbagen.2018.09.019
Felix, C., Walker, A., Lange, B., Williams, T., Winicki, N., Cheung, N., et al. (1998). Association of CYP3A4 genotype with treatment-related leukemia. Proc. Natl. Acad. Sci. 95, 13176–13181. doi: 10.1073/pnas.95.22.13176
Frosina, G. (2009). DNA repair in normal and cancer stem cells, with special reference to the central nervous system. Curr. Med. Chem. 16, 854–866. doi: 10.2174/092986709787549253
Giovannetti, E., Pacetti, P., Reni, M., Leon, L. G., Mambrini, A., Vasile, E., et al. (2011). Association between DNA-repair polymorphisms and survival in pancreatic cancer patients treated with combination chemotherapy. Pharmacogenomics 12, 1641–1652. doi: 10.2217/pgs.11.109
Goldman, M. (2008). Why is Multiple Testing a Problem? Available at: https://www.stat.berkeley.edu/~mgoldman/Section0402.pdf
Gossage, L., and Madhusudan, S. (2007). Cancer pharmacogenomics: role of DNA repair genetic polymorphisms in individualizing cancer therapy. Mol. Diagn. Ther. 11, 361–380. doi: 10.1007/BF03256260
Ha, V. H., Jupp, J., and Tsang, R. Y. (2017). Oncology drug dosing in gilbert syndrome associated with UGT1A1: a summary of the literature. Pharmacotherapy 37, 956–972. doi: 10.1002/phar.1946
Huang, R. S., and Ratain, M. J. (2009). Pharmacogenetics and pharmacogenomics of anticancer agents. Cancer J. Clin. 59, 42–55. doi: 10.3322/caac.20002
Ingelman-Sundberg, M., and Sim, S. C. (2010). Pharmacogenetic biomarkers as tools for improved drug therapy; emphasis on the cytochrome P450 system. Biochem. Biophys. Res. Commun. 396, 90–94. doi: 10.1016/j.bbrc.2010.02.162
Jain, A. K., Jain, S., and Rana, A. C. (2007). Metabolic enzyme considerations in cancer therapy. Malays J. Med. Sci. 14, 10–17.
Joerger, M., Burgers, J. A., Baas, P., Doodeman, V. D., Smits, P. H., Jansen, R. S., et al. (2012). Gene polymorphisms, pharmacokinetics, and hematological toxicity in advanced non-small-cell lung cancer patients receiving cisplatin/gemcitabine. Cancer Chemother. Pharmacol. 69, 25–33. doi: 10.1007/s00280-011-1670-4
Jóna, Á, Miltényi, Z., Póliska, S., Bálint, B. L., and Illés, Á (2016). Effect of Bleomycin Hydrolase gene polymorphism on late pulmonary complications of treatment for Hodgkin Lymphoma. PLoS One 11:e0157651. doi: 10.1371/journal.pone.0157651
Kelland, L. (2007). The resurgence of platinum-based cancer chemotherapy. Nat. Rev. Cancer 7, 573–584. doi: 10.1038/nrc2167
Keshava, C. H., McCanlies, E., and Weston, A. (2004). CYP3A4 polymorphism-potential risk factor for breast and prostate cancer: a HuGE review. Am. J. Epidemiol. 160, 825–841. doi: 10.1093/aje/kwh294
Khrunin, A. V., Moisseev, A., Gorbunova, V., and Limborska, S. (2010). Genetic polymorphisms and the efficacy and toxicity of cisplatin-based chemotherapy in ovarian cancer patients. Pharmacogenomics J. 10, 54–61. doi: 10.1038/tpj.2009.45
Kim, J., and Bang, H. (2016). Three common misuses of P values. Dent. Hypotheses 7, 73–80. doi: 10.4103/2155-8213.190481
Kivistö, K. T., Kroemer, H. K., and Eichelbaum, M. (1995). The role of human cytochrome P450 enzymes in the metabolism of anticancer agents: implications for drug interactions. Br. J. Clin. Pharmacol. 40, 523–530. doi: 10.1111/j.1365-2125.1995.tb05796.x
Lehmann, A. R. (2001). The Xeroderma pigmentosum group D (XPD) gene: one gene, two functions, three diseases. Genes Dev. 15, 15–23. doi: 10.1101/gad.859501
Lyer, L., Das, S., Janisch, L., Wen, M., Ramirez, J., Karrison, T., et al. (2002). UGT1A1∗28 polymorphism as a determinant of irinotecan disposition and toxicity. Pharmacogenomics J. 2, 43–47. doi: 10.1038/sj.tpj.6500072
Mancinelli, L., Cronin, M., and Sadée, W. (2000). Pharmacogenomics: the promise of personalized medicine. AAPS PharmSci. 2:E4. doi: 10.1208/ps020104
Marchewka, Z., Piwowar, A., Ruzik, S., and Długosz, A. (2017). Glutathione S - transferases class Pi and Mi and their significance in oncology. Postepy Hig. Med. Dosw. 71, 541–550. doi: 10.5604/01.3001.0010.3835
Mazerska, Z., Mróz, A., Pawłowska, M., and Augustin, E. (2016). The role of glucuronidation in drug resistance. Pharmacol. Ther. 159, 35–55. doi: 10.1016/j.pharmthera.2016.01.009
Mezvrishvili, Z., and Managadze, L. (2006). Three cycles of etoposide and cisplatin chemotherapy in clinical stage IS nonseminomatous testicular cancer. Int. Urol. Nephrol. 38, 621–624. doi: 10.1007/s11255-006-0038-x
Namipashaki, A., Razaghi-Moghadam, Z., and Ansari-Pour, N. (2015). The essentiality of reporting hardy-weinberg equilibrium calculations in population-based genetic association studies. Cell J. 17, 187–192. doi: 10.22074/cellj.2016.3711
Negoro, Y., Yano, R., Yoshimura, M., Suehiro, Y., Yamashita, S., Kodawara, T., et al. (2018). Influence of UGT1A1 polymorphism on etoposide plus platinum-induced neutropenia in Japanese patients with small-cell lung cancer. Int. J. Clin. Oncol. doi: 10.1007/s10147-018-1358-4 [Epub ahead of print].
Nissar, S., Syed-Sameer, A., Rasool, R., Chowdri, N. A., and Rashid, F. (2017). Glutathione S-transferase: biochemistry, polymorphism and role in colorectal carcinogenesis. J. Carcinog. Mutagen. 8, 1–9. doi: 10.4172/2157-2518.1000287
Park, C. H., Bessho, T., Matsunaga, T., and Sancar, A. (1995). Purification, and characterization of the XPF-ERCC1 complex of human DNA repair excision nuclease. J. Biol. Chem. 270, 22657–22660. doi: 10.1074/jbc.270.39.22657
Phuthong, S., Settheetham-Ishida, W., Natphopsuk, S., Settheetham, D., and Ishida, T. (2017). Haplotype analysis of MDR1 and risk for cervical cancer in Northeasterns Thailand. Asian Pac. J. Cancer. Prev. 18, 1815–1819. doi: 10.22034/APJCP.2017.18.7.1815
Pirmohamed, M., James, S., Meakin, S., Green, C., Scott, A. K., Walley, T. J., et al. (2004). Adverse drug reactions as cause of admission to hospital: prospective analysis of 18 820 patients. BMJ 329, 15–19. doi: 10.1136/bmj.329.7456.15
Pizzocaro, G., Piva, L., Salvioni, R., Zanoni, F., and Milani, A. (1985). Cisplatin, etoposide, bleomycin first-line therapy and early resection of residual tumor in far-advanced germinal testis cancer. Cancer 56, 2411–2415. doi: 10.1002/1097-0142(19851115)56:10<2411::AID-CNCR2820561012>3.0.CO;2-H
Quiñones, L., Berthou, F., Varela, N., Simon, B., Gil, L., and Lucas, D. (1999). Ethnic susceptibility to lung cancer: differences in CYP2E1, CYP1A1 and GSTM1 genetic polymorphisms between french caucasian and chilean populations. Cancer Lett. 141, 167–171. doi: 10.1016/S0304-3835(99)00099-3
Quiñones, L., Rosero, M., Roco, A., Moreno, I., Varela, F., Caceres, D., et al. (2008). Role of Cytochrome P450 enzymes in the metabolism of antineoplasic drugs: present situation and future perspectives. Rev. Med. Chil. 136, 1327–1335.
Quiñones, L. A., Roco, A. M., Cayún, J. P., Escalante, P., Miranda, C., Varela, N., et al. (2017). Clinical applications of pharmacogenomics. Rev. Med. Chil. 145, 483–500. doi: 10.4067/S0034-98872017000400009
Roco, A., Cayún, J. P., Contreras, S., Stojanova, J., and Quiñones, L. (2014). Can pharmacogenetics explain efficacy and safety of cisplatin pharmacotherapy? Rev. Front. Genet. 5:391. doi: 10.3389/fgene.2014.00391
Roco, A., Quiñones, L., Agúndez, J., García-Martín, E., Squicciarini, V., Miranda, C., et al. (2012). Allele frequencies of 23 functionally significant variant alleles related with metabolism of antineoplastic drugs in the Chilean population: comparison with Caucasian and Asian populations. Front. Genet. 3:229. doi: 10.3389/fgene.2012.00229
Ross, C. J., Visscher, H., Rassekh, S. R., Castro-Pastrana, L. I., Shereck, E., Carleton, B., et al. (2011). Pharmacogenomics of serious adverse drug reactions in pediatric oncology. J. Popul. Ther. Clin. Pharmacol. 18, e134–e151.
Ruzzo, A., Graziano, F., Kawakami, K., Watanabe, G., Santini, D., Catalano, V., et al. (2006). Pharmacogenetic profiling and clinical outcome of patients with advanced gastric cancer treated with palliative chemotherapy. J. Clin. Oncol. 24, 1883–1891. doi: 10.1200/JCO.2005.04.8322
Schwartz, D. R., Homanics, G. E., Hoyt, D. G., Klein, E., Abernethy, J., and Lazo, J. S. (1999). The neutral cysteine protease bleomycin hydrolase is essential for epidermal integrity and bleomycin resistance. Proc. Natl. Acad. Sci. U.S.A. 96, 4680–4685. doi: 10.1073/pnas.96.8.4680
Tsai, M. S., Weng, S. H., Kuo, Y. H., Chiu, Y. F., and Lin, Y. W. (2011). Synergistic effect of curcumin and cisplatin via down-regulation of thymidine phosphorylase and excision repair cross-complementary 1 (ERCC1). Mol. Pharmacol. 80, 136–146. doi: 10.1124/mol.111.071316
White, D. A., Schwartzberg, L. S., Kris, M. G., and Bosl, G. J. (1987). Acute chest pain syndrome during bleomycin infusions. Cancer 59, 1582–1585. doi: 10.1002/1097-0142(19870501)59:9<1582::AID-CNCR2820590909>3.0.CO;2-#
Zamble, D. B., and Lippard, S. J. (1995). Cisplatin and DNA repair in cancer chemotherapy. Trends Biochem. Sci. 20, 435–439. doi: 10.1016/S0968-0004(00)89095-7
Zhang, Y., Chang, D., and Zhang, J. (2017). Research advances in resistance to platinum-based chemotherapy in lung cancer. Zhongguo Yi Xue Ke Xue Yuan Xue Bao 39, 150–155. doi: 10.3881/j.issn.1000-503X.2017.01.025
Keywords: pharmacogenetics, polymorphisms, toxicity, testicular cancer, ADRs
Citation: Lavanderos MA, Cayún JP, Roco Á, Sandoval C, Cerpa L, Rubilar JC, Cerro R, Molina-Mellico S, Celedón C, Cerda B, García-Martín E, Agúndez JAG, Acevedo C, Peña K, Cáceres DD, Varela NM and Quiñones LA (2019) Association Study Among Candidate Genetic Polymorphisms and Chemotherapy-Related Severe Toxicity in Testicular Cancer Patients. Front. Pharmacol. 10:206. doi: 10.3389/fphar.2019.00206
Received: 28 December 2018; Accepted: 19 February 2019;
Published: 08 March 2019.
Edited by:
Jorge Fuentealba, Universidad de Concepción, ChileReviewed by:
Sujit Nair, SVKM’s Narsee Monjee Institute of Management Studies, IndiaCopyright © 2019 Lavanderos, Cayún, Roco, Sandoval, Cerpa, Rubilar, Cerro, Molina-Mellico, Celedón, Cerda, García-Martín, Agúndez, Acevedo, Peña, Cáceres, Varela and Quiñones. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Luis A. Quiñones, bHF1aW5vbmVAbWVkLnVjaGlsZS5jbA== Nelson M. Varela, bnZhcmVsYUBtZWQudWNoaWxlLmNs
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.