 Aminu Mohammed
Aminu Mohammed Md. Shahidul Islam
Md. Shahidul Islam
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Pharmacol. , 22 August 2018
Sec. Ethnopharmacology
Volume 9 - 2018 | https://doi.org/10.3389/fphar.2018.00893
This article is part of the Research Topic Phytochemicals in Preventing and Managing Metabolic Syndrome View all 54 articles
Spices possess tremendous therapeutic potential including hypoglycemic action, attributed to their bioactive ingredients. However, there is no study that critically reviewed the hypoglycemic potency, safety and the bioavailability of the spice-derived bioactive ingredients (SDBI). Therefore, the aim of the study was to comprehensively review all published studies regarding the hypoglycemic action of SDBI with the purpose to assess whether the ingredients are potential hypoglycemic agents or adjuvant. Factors considered were concentration/dosages used, the extent of blood glucose reduction, the IC50 values, and the safety concern of the SDBI. From the results, cinnamaldehyde, curcumin, diosgenin, thymoquinone (TQ), and trigonelline were showed the most promising effects and hold future potential as hypoglycemic agents. Conclusively, future studies should focus on improving the tissue and cellular bioavailability of the promising SDBI to achieve greater potency. Additionally, clinical trials and toxicity studies are with these SDBI are warranted.
Diabetes mellitus (DM) is a chronic metabolic disorder characterized by hyperglycemia resulting from the malfunction in insulin secretion and/or insulin action, both leading to impair metabolism of carbohydrates, lipids, and proteins (ADA, 2015). The prevalence of DM is increasing exponentially to over 425 million people globally, and this figure is likely to rise to 629 million by 2045 (IDF, 2017; Ogurtsova et al., 2017).
At present, the most prominent approach to control DM involves the use of oral synthetic hypogycemic drugs such as sulphonylureas, biguanide, α-glucosidase, and dipeptidyl peptidase-4 (DPP-4) inhibitors. However, these drugs have characteristic profiles of short- and/or long-term side effects, which include hypoglycemia, weight gain, gastrointestinal discomfort and nausea, liver and heart failure (Hung et al., 2012). Additionally, the drugs are costly in the developing countries especially in Asia and African regions. These limitations have prompted the search for potent plant-derived bioactive ingredients as possible alternative therapies for DM. The target is to identify newer compounds that could attenuate hyperglycemia, ameliorate the diabetes associated-complications with fewer adverse effects. These can be standardized and used as the drugs for the treatment of DM.
Spices add flavor, taste, and color in food preparation and most importantly, consumption of spices provide infinite health benefits to humans. Considerable evidence has shown that spices play a vital role in ameliorating DM complications and were documented in several reviews (Khan and Safdar, 2003; Kelble, 2005; Srinivasan, 2005; Mohamed, 2014; Kazeem and Davies, 2016; Bi et al., 2017). However, most of the available reviews focused on the extracts derived from the spices. Although some of the reviews highlighted the hypoglycemic roles of the bioactive ingredients derived from the spices (Upaganlawar et al., 2013; Zhang et al., 2013; Semwal et al., 2015), the critical assessment of their hypoglycemic potency based on the concentration/dose has not yet been well documented. The exaggerations of the data obtained from in vitro and in vivo studies are of concerns. In other words, whether these active ingredients are potential hypoglycemic agents or adjuvants, not clear at all. On the other hand, the lack of bioavailability is the major factor affecting the overall bioactivity of the spice-derived bioactive ingredients (SDBI) (Huang et al., 2010; Yao et al., 2015). Therefore, we intended to comprehensively review all the published studies on the hypoglycemic action of SDBI with critical assessment whether the ingredients are potential hypoglycemic agents or adjuvants. In addition, future prospects, safety and the progress made on the methods used to improve the bioavailability of the promising SDBI were included in this review as well.
In the present study, we considered the SDBI as potential hypoglycemic agents based on multiple citations that showed >50% blood glucose reduction potential at non-toxic dosages. The potent hypoglycemic action using in vitro models (lower IC50 values) and less toxicity associated with the target compounds were also considered in this study. The hypoglycemic roles of the SDBI were categorized and presented based on in vitro (Table 1), in vivo (Supplementary Table 1) or clinical (Table 2) studies. Additionally, proposed hypoglycemic mechanisms depicted by the promising SDBI are presented in Figure 1.
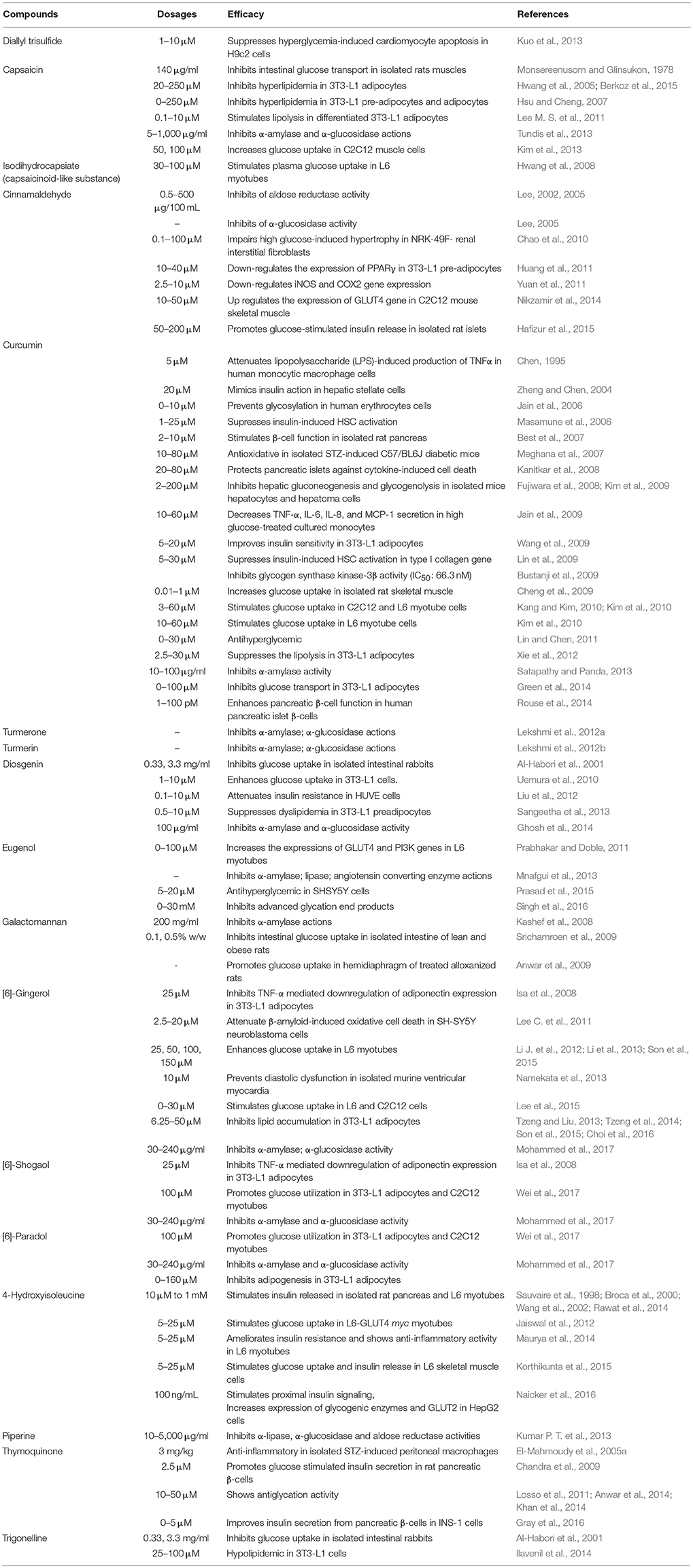
Table 1. In vitro studies of spice-derived ingredients.

Table 2. Clinical trials of spice-derived ingredients.
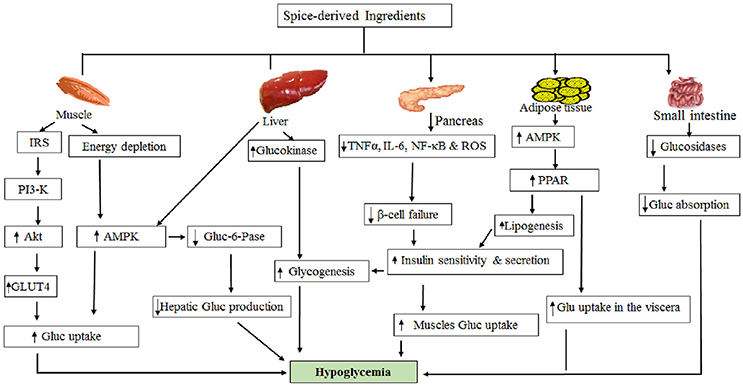
Figure 1. Possible mechanism of hypoglycemic action by spice-derived ingredients.
Diallyl trisulfide (DATS) an organosulfur from garlic (Allium sativum L.) at various concentrations (1–5 μM) suppressed high glucose-induced cardiomyocyte apoptosis via inhibition of NADPH oxidase, reactive oxygen species (ROS) production and downregulated JNK/NF-κB signaling in H9c2 cells (Kuo et al., 2013). This shows the potential of DATS in the management of diabetes-associated inflammation.
Saravanan and colleagues have shown that oral administration of S-Allyl cysteine (SAC) treatment at 150 mg/kg bw for 45 days reduced fasting blood glucose (FBG) by 65%, ameliorated oxidative damages, glycosuria and improved the activities of glucose metabolizing enzymes in STZ-diabetic rats (Saravanan et al., 2009, 2010, 2013; Saravanan and Ponmurugan, 2010, 2011, 2012a,b). Oral supplementation of SAC (0.5–1.0 g/l) for 4 or 10 weeks showed 34% more FBG reduction compared to n-acetyl cysteine, S-ethyl cysteine, S-methyl cysteine, and S-propyl cysteine (<30% FBG reduction) in STZ-induced Balb/cA mice (Hsu et al., 2004; Mong and Yin, 2012). Additionally, SAC was shown to have potent protection against renal inflammation via suppressing NF-κB activity and NF-κB p65 mRNA expression in STZ-induced diabetic rats (Mong and Yin, 2012).
Oral administration of alliin [S-allyl cysteine sulfoxide (SACS)] and S-methyl cysteine sulfoxide (SMCS) at 200 mg/kg bw for 30 days decreased FBG by 44.5 and 38%, respectively in alloxan-induced diabetic rats (Sheela et al., 1995). Furthermore, alliin, a sulfoxided from garlic, decreased serum glycosylated hemoglobin, the activities of phosphatases, lactate dehydrogenase and glucose-6-phosphatase enzymes and increased serum insulin level, liver and intestinal HMG-CoA reductase and hexokinase activities in alloxan-induced diabetic rats (Sheela and Augusti, 1992; Augusti and Sheela, 1996). Conversely, consumption of diallyl diasulfide (DADS) and DATS (40–80 mg/kg bw) for 16 or 3 weeks showed no effects on FBG in STZ-induced diabetic rats (Liu et al., 2005, 2006). Interestingly, treatment of DATS (40 mg/kg bw) for 16 days reduced the expression of phosphorylated JNK and NF-κB, and active caspase 3 in cardiac myocytes of STZ-induced diabetic rats (Kuo et al., 2013). This supports the in vitro data published by Kuo et al. (2013) and further showed DATS ability to ameliorate diabetes-induced elevation of inflammatory mediators such as tumor necrosis factor-alpha (TNFα) in the muscles. Additionally, oral administration of S-allyl-mercapto-captopril (alliin and Captopril conjugate) at 53.5 mg/kg bw for 55 days reduced FBG (42%) and blood pressure in Cohen-Rosenthal diabetic hypertensive rats (Younis et al., 2010). Allicin (derived from hydrolysis of alliin) at 250 mg/kg bw decreased blood glucose levels and improved glucose tolerance after 4 h post-administration period in alloxan-induced diabetic rabbits (Mathew and Augusti, 1973).
Based on the present literature search, studies on the detail toxicities associated with organosulfur compounds under study are scanty. However, Rao and Natarajan (1949) reported the subcutaneous and intraperitoneal LD50 of allicin are 5 and 20 mg/kg bw, respectively.
According to the above-mentioned studies, the spice-derived sulfur containing ingredients showed their hypoglycemic effects not only by decreasing FBG, oxidative stress, inflammatory biomarkers but also by increasing insulin secretion and improving glucose tolerance and glucose metabolism-related enzyme activities. However, based on the levels of hypoglycemic potential of sulfur containing compounds and their derivatives, these compounds (SAC, SMCS, SACS, DADS, DATS, and allicin) cannot be considered as hypoglycemic agents but only as adjuvants.
The hypoglycemic action of capsaicin from Capsicum species seems to be controversial and contradictory. Monsereenusorn and Glinsukon (1978) have reported that capsaicin (140 μg/ml) inhibited intestinal glucose transport (22.6%) mediated by GLUT2, attributed to the Na+-K+-ATPase pump (Monsereenusorn and Glinsukon, 1979). In another study, capsaicin (5–1,000 μg/ml) exhibited α-amylase (IC50: 83 μg/ml) and α-glucosidase (IC50: >500 μg/ml) inhibitory activities (Tundis et al., 2013), implying a possible role in ameliorating post-prandial hyperglycemia. In addition, capsaicin and its derivative (isodihydrocapsiate) at various concentrations (50–100 μM) stimulated glucose uptake, via AMP-activated protein kinase (AMPK) up regulation in C2C12 muscle or L6 myotube cells, respectively (Hwang et al., 2008; Kim et al., 2013). Moreover, capsaicin (0–250 μM) inhibited lipid accumulation in 3T3 L1 pre-adipocytes and adipocytes, implying the role of capsaicin in attenuating insulin resistance (Hsu et al., 2004; Hwang et al., 2005; Lee M. S. et al., 2011).
Intraperitoneal treatment of capsaicin (20–50 mg/kg bw) for 9 weeks attenuated hyperglycemia (44% reduction), improved glucose homeostasis and insulin release in Zucker diabetic fatty (ZDF) rats and partial pancreatectomized diabetic rats (Gram et al., 2007; Kwon et al., 2013). Accordingly, dietary inclusion of capsaicin (0.015%) for 3 weeks decreased hyperglycemia (17%) and ameliorated dyslipidemia, inflammation and insulin resistance in KK-Ay obese/diabetic mice, which linked to its dual action on PPAR-α and TRPV-1 expression/activation (Kang et al., 2011a,b). Similarly, capsaicin (0.0024–0.0042%) inclusion in diet showed maximum FBG reduction of 49% in the same animal model (Okumura et al., 2012).
Notably, administration of capsaicin (10 μg/kg bw) for 20 weeks prevented the onset of type 1 diabetes in a non-obese diabetic mouse model, attributed to the attenuation of antigen-specific T-cells in pancreatic lymph nodes (Nevius et al., 2012). Conversely, animals treated with other dosages (0.1, 1.0, 25.0, and 50.0 μg/kg bw) were hyperglycemic throughout the study period, which is a subject for further studies. More recently, dietary inclusion of capsaicin (0.014–0.1%) for 12 weeks decreased serum and tissue advanced glycation end products (AGEs) and activated the receptor for AGEs (RAGE) in STZ-induced diabetic rats (Hsia et al., 2016). However, the reduction of FBG in the capsaicin-treated groups was not significant compared to the diabetic untreated group (Hsia et al., 2016). This further supports the previous studies that capsaicin administration (0.015%) has no effect on the blood glucose level in the same animal model (Babu and Srinivasan, 1997a; Suresh Babu and Srinivasan, 1998). Furthermore, the dietary inclusion of capsiate (0.025%/7 weeks), a non-pungent capsaicin analog, improved glucose tolerance ability (28%) via improving insulin sensitivity in pancreatectomized diabetic rats (Kwon et al., 2013).
In a randomized, double-blind, placebo-controlled trial, oral administration of capsaicin (5 mg/day) for 4 weeks attenuated insulin resistance and dyslipidemia with no significant effect on FBG in women with gestational diabetes (Yuan et al., 2016). In addition, topical application of capsaicin (0.075%) for 8 weeks ameliorated painful diabetic neuropathy in diabetic patients (Scheffler et al., 1991; Tandan et al., 1992; Biesbroeck et al., 1994; Forst et al., 2002).
The oral LD50 values of capsaicin were within the ranges 90–162 mg/kg bw for mice and rats (Saito and Yamamoto, 1996). However, the intraperitoneal, intravenous, and subcutaneous LD50 values for mice were 7.65, 0.56, and 9 mg/kg bw in mice, indicating possible toxicity (Glinsukon et al., 1980). To further support this, some adverse consequences of capsaicin consumption reported include nausea, vomiting, abdominal pain, burning diarrhea, intense tearing and conjunctivitis (Goldfrank, 2002; Millqvist et al., 2005). Additionally, Marques et al. (2002) have reported that people consuming capsaicin (90–250 mg/day) are more susceptible gastrointestinal cancer compared to the subjects consumed lesser doses of capsaicin (0–29.9 mg/day).
From the above-mentioned studies, although capsaicin showed mild to moderate hypoglycemic activity by inhibiting glucose digesting enzymes activities, improving glucose uptake, decreasing insulin resistance, dyslipidemia, advanced glycation endproducts; the reduction of FBG and hyperglycemia was not promising. Therefore, it may not be a good candidate for DM therapy. Our argument is that none of the studies reported >50% reduction of blood glucose levels despite several weeks of administration. Additionally, capsaicin consumption showed weak antihyperglycemic effect in women with gestational diabetes (Yuan et al., 2016). The toxicities associated with capsaicin consumptions are another great concern. Despite intraperitoneal administration conferred higher action compared to the oral administration, it was more susceptible to adverse consequences and hence should be discouraged. However, capsaicin topical application is encouraged to reduce some complications associated with diabetic neuropathy as this was validated in some clinical trials (Scheffler et al., 1991; Tandan et al., 1992; Biesbroeck et al., 1994; Forst et al., 2002). This justified the use of capsaicin as adjuvant in the management of DM, particularly diabetic neuropathy.
Cinnamaldehyde is an aromatic aldehyde and main bioactive component of cinnamon (Cinnamomum zeylanicum var. cassia Meisn.). Several studies have reported the potential of cinnamaldehyde in the prevention of diabetes related-complications. Lee (2005) reported that cinnamaldehyde (0.005–5 μg/ml) is a potent aldose reductase (IC50: 0.8 μg/ml) and weak α-glucosidase (IC50: 500 μg/ml) inhibitor signifying its potential in attenuating osmotic imbalance in non-insulin dependent tissues and hence, ameliorated diabetic retinopathy.
Cinnamaldehyde (10–50 μM) attenuated lipid accumulations in 3T3 preadipocytes via PPARδ, PPARγ, AMPK, and retinoid X receptor (RXR) expression. and thus, helps to prevent insulin resistance (Huang et al., 2011; Li et al., 2015). Similarly, cinnamaldehyde (2.5–10 μM) prevented STZ-induced pancreatic β-cell damage in RINm5F rat insulinoma cells (Yuan et al., 2011). This effect was linked to the downregulation of iNOS and COX-2 genes expression through blocking the NF-κB and MAPKs activities that ultimately prevented pancreatic ROS elevation and damage. Chao et al. (2010) reported that cinnamaldehyde (0.1–100 μM) reduced high glucose-induced hypertrophy in NRK-49F- renal interstitial fibroblasts through inactivation of the p38 MAPK pathway, linked to diabetic nephropathy. Nikzamir et al. (2014) have demonstrated that cinnamaldehyde (10–50 μM) stimulated glucose transporter 4 (GLUT4) gene expression in C2C12 mouse skeletal muscle. Hafizur et al. (2015) have shown the potential of cinnamaldehyde to induce glucose-stimulated insulin release in isolated islets, which could facilitate glucose transport into the cells and thus reduced hyperglycemia.
Oral administration of cinnamaldehyde (5–20 mg/kg bw) for 45 days reduced FBG (63.3%), lipid accumulation and showed insulinotropic action in STZ-induced diabetic rats (Subash Babu et al., 2007). Interestingly, the same authors have recently reported a more potent FBG reduction (71%) while used the same doses, study period and animal models (Subash Babu et al., 2014). The potent antihyperglycemic action of cinnamaldehyde was linked to the upregulation of GLUT4 protein expression that may facilitate the transport of glucose across the cells (Zhang et al., 2008; Anand et al., 2010; Jawale et al., 2016). Importantly, Zhang et al. (2008) showed a 62% reduction of FBG and improved insulin sensitivity in pancreatic β-cell upon cinnamaldehyde (40 mg/kg bw) consumption in a high-fat diet-fed STZ-induced diabetic rat model. This was supported even at a lower dosage of cinnamaldehyde (143.8 μmol/kg bw) for 4 weeks in high-fat-diet-induced insulin resistant rats (Farrokhfall et al., 2014), and thus, corroborates with the in vitro studies (Huang et al., 2011; Li et al., 2015).
Treatment of cinnamaldehyde (20 mg/kg bw) for 6 weeks curtailed FBG (40%), insulin resistance and diabetes-induced hypertension in STZ-induced diabetic rats, attributed to the restoration of vascular contractility in the treated rats (El-Bassossy et al., 2011). In the same animal model, oral gavage of cinnamaldehyde (20 mg/kg bw) reduced FBG by 21.1 and 69.8% after 4 h and 4 weeks post-treatment period, respectively and ameliorated diabetes-induced alterations (Kumar et al., 2012). Subsequently, administration of cinnamaldehyde (20 mg/kg bw) for 4 weeks was shown to attenuate hyperglycemia, TNF-α mRNA expression and upregulated GLUT-4 mRNA expression in C57BLKS/J db/db mice (Li J. et al., 2012; Guo et al., 2017). In fatty-sucrose diet/streptozotocin (FSD/STZ)-rat model of gestational diabetes, supplementation of cinnamaldehyde (25 mg/kg bw) for 8 weeks reduced FBG (80%) via modulation of PPARγ, proinflammatory cytokines and oxidative stress (Hosni et al., 2017).
Ghrelin a hunger hormone, participate in the regulation of glucose and insulin metabolism. The plasma ghrelin levels are shown to correlate inversely with insulin levels and are associated with insulin resistance and could be a potential target to reduce the progression of type 2 diabetes (Pulkkinen et al., 2010; Tong et al., 2010). Conforming to this, dietary inclusion of cinnamaldehyde (0.2%) for 36 days retarded the endogenous ghrelin release and reduced FBG (10%) in C57BL6 diabetic mice (Camacho et al., 2015).
The low toxicity associated with cinnamaldehyde consumption in rodents via oral route has been well documented (Jenner et al., 1964; Sporn et al., 1965; Zaitsev and Rakhmanina, 1974; Subash Babu et al., 2007). Seemingly, Hooth et al. (2004) reported that the safety of cinnamaldehyde was approved by the Food and Drug Administration (FDA) and has been given Generally Recognized as Safe (GRAS) status in the United States. However, Weibel and Hansen (1989) have reported that cinnamaldehyde elicits some carcinogenic risk by acting as an alkylating agent that could react with cellular macromolecules.
According to the results of the above-mentioned studies, cinnamaldehyde is a potential hypoglycemic agent and adjuvant. Several studies have shown that cinnamaldehyde reduced FBG by >50% at 20 or 40 mg/kg bw in various animal models (Subash Babu et al., 2007, 2014; Zhang et al., 2008; Kumar et al., 2012). Regarding the in vitro studies, this ingredient showed potent hypoglycemic potential at <10 μg/ml or μM and depicted IC50 values of <10 μg/ml as well (Yuan et al., 2011; Kumar et al., 2012). These are of interest in the drug discovery as small amount of the compound stimulated beneficial action in various models. Additionally, the less toxicity associated with cinnamaldehyde intake is of significance in drug design and development. However, the lack of clinical trials with cinnamaldehyde is a major drawback in determining its exact hypoglycemic potential in human subjects. Hence, further studies, particularly clinical studies, are warranted to confirm the hypoglycemic effects of cinnamaldehyde in humans.
Curcumin is the major active principle of turmeric (Curcuma longa L.) and has been reported to possess tremendous potential including hypoglycemic action. Several studies have shown that curcumin (20 μM) stimulated insulinotropic action via PPARγ activation and attenuated oxidative stress in hepatic stellate cells (Zheng and Chen, 2004; Masamune et al., 2006; Lin et al., 2009; Lin and Chen, 2011). Jain et al. (2006, 2009) have reported that curcumin (0–40 μM) prevents glycation, decreased TNF-α, IL-6, IL-8, and MCP-1 secretion in isolated human erythrocytes and high glucose-treated cultured monocytes. Furthermore, curcumin (20–80 μM) protected pancreatic islets against cytokine-induced cell death via scavenging ROS and decreased cytokine induced NF-kB translocation (Kanitkar et al., 2008). These studies have shown that amelioration of oxidative stress could be among the possible mechanism of curcumin hypoglycemic action. Furthermore, curcumin (2–40 μM) improved glucose absorption by activating the volume-regulated anion channel in isolated pancreatic β-cells and C2C12 mouse myoblast cells (Best et al., 2007; Kang and Kim, 2010). Curcumin (2–200 μM) was reported to activate AMPK and suppress gluconeogenic enzymes gene expression in hepatoma cells, which indicates the blood glucose lowering ability of curcumin (Kim et al., 2009, 2010). To support this, curcumin (25 μM) inhibited hepatic gluconeogenesis and glycogenolysis in isolated mice hepatocytes (Fujiwara et al., 2008).
Curcumin (5–20 μM) was shown to improve insulin sensitivity in 3T3-L1 adipocytes (Wang et al., 2009), which is linked to the suppression of lipolysis and inhibition of glucose transport (Xie et al., 2012; Green et al., 2014). Increased glycogen synthase kinase-3β activity has been implicated in type 2 diabetes insulin resistance, mediated via phosphatidylinositol kinase-3 activation and the inhibition of protein kinase B (Pandey and DeGrado, 2016). Bustanji et al. (2009) have reported that curcumin inhibited glycogen synthase kinase-3β activity (IC50: 66.3 nM). In a more recent study, curcumin (10–100 μg/ml) inhibited α-amylase action (Satapathy and Panda, 2013). Curcumin (0.01–60 μM) was shown to stimulate glucose uptake in isolated rat skeletal muscle and in L6 myotube cells (Cheng et al., 2009). Additionally, curcumin (1 pM−80 μM) was shown to enhances pancreatic β-cell function in isolated human pancreatic islets (Meghana et al., 2007; Rouse et al., 2014).
Lekshmi et al. (2012a) have reported that turmerone from turmeric exhibited potent α-amylase (IC50: 24.5 μg/ml) and α-glucosidase (IC50: 0.28 μg/ml) inhibitory actions. In another study, turmerin, a water-soluble peptide in turmeric rhizomes, was reported to show α-amylase (IC50: 192 μg/ml) and α-glucosidase (IC50: 31 μg/ml) inhibitory actions as well (Lekshmi et al., 2012a). These ingredients have demonstrated the potential in reducing post-prandial hyperglycemia in diabetes.
Several studies have reported the hypoglycemic effect of curcumin using various animal models. Babu and Srinivasan (1997b); Suresh Babu and Srinivasan (1998) have shown that dietary supplementation of curcumin (0.5%) for 8 weeks attenuated hyperlipidemia and renal dysfunction in STZ-induced diabetic rats. Conversely, the authors reported no reduction on the FBG levels in the treated diabetic rats which is consistent with some previous studies (Suryanarayana et al., 2007; Palma et al., 2014). However, the above-mentioned studies have reported potent antioxidant action in the same model which is in line with some previous studies (Sajithlal et al., 1998; Rungseesantivanon et al., 2010; Gupta et al., 2011). These effects of curcumin are in line with the results of in vitro studiesas presented above (Zheng and Chen, 2004; Jain et al., 2006, 2009; Masamune et al., 2006; Kanitkar et al., 2008; Lin et al., 2009; Lin and Chen, 2011).
Oral administration of curcumin (80–100 mg/kg bw) for 3 or 7 weeks reduced FBG (31.4%) and serum glycated hemoglobin (30.6%) in alloxan-induced diabetic rats (Arun and Nalini, 2002). Dietary intervention of curcumin (0.001–0.005% w/v) for 8 weeks delayed the progression of cataract via the downregulation of vascular endothelial growth factor (VEGF) expression in STZ-induced diabetic rats (Suryanarayana et al., 2005; Kowluru and Kanwar, 2007; Mrudula et al., 2007). Supplementation of curcumin (0.5%) for 2 weeks decreased bone resorptive activity via attenuating osteoclastogenesis in STZ-induced diabetic rats (Hie et al., 2009).
Dietary supplementation of curcumin (0.02%) for 6 weeks decreased FBG (22%) in C57BL/KsJ-db/db diabetic mice (Seo et al., 2008). In KKAy diabetic mice, dietary inclusion of curcumin at 0.24% for 5 weeks increased hepatic glycolysis and overall lipids metabolism, which might help in reducing the hyperglycemia (Honda et al., 2006). Kanitkar et al. (2008) have reported that oral treatment of curcumin (7.5 mg/kg bw) for 5 days reduced FBG (69%) and ameliorated pancreatic β-cell damage in STZ-induced diabetic mice. In a series of studies, curcumin (10 or 80 mg/kg bw) treatment for 45 days showed maximum FBG reduction of 57.1%, antihyperlipidemic, insulinotropic and antioxidant activities in type 1 and type 2 diabetic rat models (Murugan and Pari, 2006a,b, 2007; Pari and Murugan, 2007b; Murugan et al., 2008; Hussein and Abu-Zinadah, 2010; Abdel Aziz et al., 2013). Oral administration of photo-irradiated curcumin (10–80 mg/kg bw) for the same period reduced FBG (53.9%) and ameliorated lipid peroxidation in STZ-induced diabetic rats (Mahesh et al., 2004, 2005). This imply that photo-irradiation has no effect on the hypoglycemic action of curcumin, since the reductions of FBG by photo-irradiated (53.9%) or non-photo-irradiated (57.1%) curcumin were not significantly different.
Oral administration of curcumin (15 or 30 mg/kg bw) for 6 weeks reduced FBG (24.4%) and attenuated renal dysfunction at the maximum dosage administered in STZ-induced diabetic rats (Sharma et al., 2006). Similarly, consumption of curcumin (60 mg/kg bw) for 2 weeks to the same animal model improved brain stem function attributed to the regulations of cholinergic, insulin receptor and GLUT-3 in the brain stem (Peeyush et al., 2009; Kumar P. T. et al., 2013). Curcumin treatment for 10 weeks ameliorated hyperglycemia (44.3%), cognitive deficit, cholinergic dysfunction, oxidative stress and inflammation in the same animal model and dosage (Kuhad and Chopra, 2007). Furthermore, Awasthi et al. (2010) have reported that oral administration curcumin (10–50 mg/kg bw) for 3 weeks provented intracerebral STZ-induced impairment in memory and cerebral blood flow. Chiu et al. (2009) showed that curcumin treatment (150 mg/kg bw) for 4 weeks reduced FBG and downregulated the expression of p300 and nuclear factor-κB in STZ-induced diabetic rats. Oral administration of curcumin (200 mg/kg bw) for 2 weeks demonstrated anticholinesterase and antioxidant actions and attenuated diabetes-induced dementia in rats (Agrawal et al., 2010; Chanpoo et al., 2010; Mahfouz, 2011). This is in line with the previous data that curcumin protects pancreatic islets from cytokine-induced cell death via scavenging ROS and decreasing cytokine-induced NF-kB translocation (Kanitkar et al., 2008).
Curcumin treatment (60 mg/kg bw) downregulated β2-adrenoceptor gene expression and upregulated the insulin receptor gene expression in the muscles of STZ-induced diabetic rats, indicating decreased glycogenolysis, gluconeogenesis and increased glycogenesis in the muscles. (Xavier et al., 2012). Dietary inclusion of curcumin (0.5%) for 16 weeks improved the activities of lysosomal enzymes in liver, spleen, heart, lungs, testis and brain of STZ-induced rats (Chougala et al., 2012). El-Bahr (2013) have reported that oral administration of curcumin (15 mg/5 ml/kg bw) for 6 weeks to STZ-induced diabetic rats reduced FGB (43.7%) and improved the in vivo antioxidant status Consumption of curcumin (60 mg/kg bw) for 2 months to alloxan-induced diabetic rats decreased FBG and improved the pancreatic architecture to near normal (Acar et al., 2012; Abdel Aziz et al., 2013; Abdul-Hamid and Moustafa, 2013; Ghosh et al., 2015). Intraperitoneal administration of curcumin (10 mM) for 4 weeks reduced FBG (40%), exhibited pancreatic islet regenerative and antioxidative potential in STZ-induced diabetic rats (El-Azab et al., 2011). In some studies, oral administration of curcumin (100–200 mg/kg bw) for 2 or 8 weeks to STZ-induced diabetic rats ameliorated diabetic nephropathy and cardiomyopathy related symptoms (Soetikno et al., 2012, 2013; Zhao W. C. et al., 2014; Zheng et al., 2014). The proposed mechanism behind this effect was the inhibition of NADPH oxidase-mediating oxidative stress in the spinal cord and downregulation of the sphingosine kinase 1-sphingosine 1-phosphate (SphK1-S1P) signaling pathway (Soetikno et al., 2012; Huang et al., 2013).
Supplementation of curcumin (30–90 mg/kg bw) in yogurt for 31 days to STZ-induced diabetic rats showed antihyperglycemic and antihyperlipidemic actions (Gutierres et al., 2012). Rashid and Sil (2015) have shown that curcumin play a beneficial role against STZ-induced testicular abnormalities in diabetic rats. Consumption of curcumin (100 mg/kg bw) for 8 weeks reduced FBG (56.5%), intracellular Ca2+ level, active caspase cascade and the poly ADP-ribose polymerase (PARP) cleavage. Additionally, theNFκB-mediated inflammation was attenuated when the PI3K/Akt-dependent signaling was activated in the curcumin-treated animals (Rashid and Sil, 2015). This finding has suggested the protective role of curcumin against oxidative and ER stress in testes. Curcumin supplementation (50 or 100 mg/kg bw) for 3 weeks reduced hyperglycemia and the risk of vascular inflammation via attenuation of IL-6, MCP-1, TNF-α, HbA1, and lipid peroxidation in STZ-induced diabetic rats (Jain et al., 2009; Banafshe et al., 2014). In a nut shell, vast amount of data demonstrated curcumin to possess blood glucose and lipid-lowering abilities with subsequent improvement on insulin sensitivity in high fat-fed rats (Naito et al., 2002; Arafa, 2005; Kempaiah and Srinivasan, 2006; Jang et al., 2008; El-Moselhy et al., 2011; Na et al., 2011; Kaur and Meena, 2012; Hussein and El-Maksoud, 2013).
Administration of tetrahydrocurcumin (THC) a curcumin derivative (80 mg/kg bw) for 45 days reduced FBG (55%) and conferred potent antioxidant potential in STZ-induced diabetic rats (Karthikesan et al., 2010a,b). The effect was higher (67%) when co-administered with chlorogenic acid (5 mg/kg bw). This has indicated possible synergy with chlorogenic acid and warrant further study to understand the synergistic mode of interaction of THC and chlorogenic acid. Murugan and Pari have shown that administration of THC at the same dose and study period reduced FBG by 60% compared to 54.4% for curcumin (Pari and Murugan, 2005, 2007,b, 2008; Murugan and Pari, 2006a,b, 2007; Murugan et al., 2008). Additionally, a potent antihyperlipidemic, insulinotropic and antioxidant actions in diabetic rat models were also reported by the authors. This shows that the reduction of the FBG by the THC and curcumin is not significant and, Kanitkar et al. (2008) have reported a 69% reduction by the curcuim alone within short study period.
Chuengsamarn and colleague reported that daily administration of curcumin at 250 mg for 6 and 9 months improved insulin action and lowered atherogenic risks in type 2 diabetic patients (Chuengsamarn et al., 2012, 2014). Previously, Usharani et al. (2008) reported that intake of curcumin capsules (150 mg) twice daily for 8 weeks to type 2 diabetic patients showed improved antioxidative status comparable to that of atorvastatin. Neerati et al. (2014) have recently reported that ingestion of curcumin (475 mg) for 10 day attenuated hyperglycemia and hyperlipidemia in type 2 diabetic patients. These studies compliment the in vitro and in vivo data despite lack of detail hypoglycemic potential in human subjects and signify the greater potential of curcumin in diabetes management.
Considerable amount of data is available, demonstrating curcumin safety and tolerability at the high doses (12 g/day) in several animal models (Lao et al., 2006a,b) and human subjects (Shankar et al., 1980; Chainani-Wu, 2003; Hsu and Cheng, 2007). However, some studies have shown that curcumin and its derivatives may cause hepatotoxicity, skin irritation and stomach ulcers when taken in high doses or for a prolonged period (Babu and Srinivasan, 1997b; Kandarkar et al., 1998; Balaji and Chempakam, 2010). Therefore, it is suggested that curcumin consumption at lower doses has no potential side effects. To further support this daily consumption of curcumin (500 mg) for 2 months was reported not to cause any adverse consequences in humans, except mild nausea and diarrhea (Hsu and Cheng, 2007; Chandran and Goel, 2012).
From the above-mentioned studies, it is evident that curcumin is the most investigated SDBI. Interestingly, numerous studies have reported FBG reduction of >50% with potent amelioration of diabetes-induced damages in various animal models without noticeable toxicity (Mahesh et al., 2004, 2005; Murugan and Pari, 2006a,b, 2007; Pari and Murugan, 2007b; Kanitkar et al., 2008; Murugan et al., 2008; Gutierres et al., 2012). To further support this, several in vitro studies have shown the potent curcumin hypoglycemic potential at concentrations even <10 μM (Best et al., 2007; Jain et al., 2006; Cheng et al., 2009; Wang et al., 2009; Kang and Kim, 2010). The less toxicity of curcumin intake in humans is encouraging and is of pharmacological interest as well.
Diosgenin is a steroidal saponin and dietary ingredient from popularly consumed spice fenugreek (Trigonella foenum-graecum L.). Based on the current literature search, the information regarding the hypoglycemic potential of diosgenin in vitro is scanty. Liu et al. (2012) have reported that diosgenin (0.1–10 μM) attenuated insulin resistance associated endothelial dysfunction via inhibition of IKKβ and IRS-1 pathways in human umbilical vein endothelial cells (HUVECs). However, Fang et al. (2016) have recently linked the inhibition of insulin resistance to increase expression of the phosphorylated estrogen receptor-α (Erα), sarcoma (Src), Akt/protein kinase B and glycogen synthase kinase-3β (GSK-3β). The above data have demonstrated the diosgenin potential in amelioration of diabetes-associated insulin resistance.
In another study, diosgenin (0.5–10 μM) enhanced insulin-dependent glucose uptake and mitigate dyslipidemia via modulation of PPARs in 3T3-L1 preadipocytes (Uemura et al., 2010; Sangeetha et al., 2013). Diosgenin (100 μg/ml) showed uncompetitive mode of inhibition against α-amylase (70.9%) and α-glucosidase (81.7%) actions (Ghosh et al., 2014). Previously, diosgenin (0.33–3.3 mg/ml) was reported to inhibit glucose uptake (IC50: 8 mM) in isolated intestinal rabbits (Al-Habori et al., 2001). The above data suggest the beneficial role of diosgenin in controlling post-prandial hyperglycemia via delaying dietary glucose absorption and facilitating glucose uptake from the circulation.
Dietary inclusion of diosgenin (10 g/kg bw) for 3 weeks reduced FBG (33.4%) and ameliorated dyslipidemia via modulation of Na+-K+-ATPase and increasing Ca2+ ATPase activities in STZ-induced diabetic rats (McAnuff et al., 2002, 2005). The increased action of the ATPases has direct effect on insulin, which plays major role in blood glucose regulation. Interestingly, oral administration of diosgenin (10–60 mg/kg bw) for 2 weeks decreased FBG (58%), elevated plasma insulin levels and tissue hexokinase activity with subsequent attenuation of oxidative stress in STZ-induced diabetic rats (Pari et al., 2012; Sangeetha et al., 2013; Saravanan et al., 2014). In another study, dietary inclusion of diosgenin (0.5 or 2%) for 4 weeks improved glucose tolerance ability as well as insulin sensitivity in high-fat diet-fed KK-Ay/Ta Jcl obese diabetic mice (Uemura et al., 2010).
In coherence with this finding, Naidu et al. (2015) have reported a 62.6% FBG reduction and amelioration of insulin resistance and hyperlipidemia after 30 day administration of diosgenin (60 mg/kg bw) in the same animal model. To further support this, diosgenin (10 mg/kg bw) treatment showed 70% reduction of FBG, improved antioxidant status and insulin levels in STZ-induced diabetic rats (Kalailingam et al., 2014). The higher hypoglycemic action of diosgenin was previously attributed to the reduction of serum levels of cytokines, and adipokines as well as increased PPARγ levels, implying the insulin-sensitizing potential of diosgenin in diabetic condition (Tharaheswari et al., 2014). In another study, oral treatment of diosgenin (40 mg/kg bw) for 7 weeks mitigated vascular dysfunction in STZ-induced diabetic rats (Roghani-Dehkordi et al., 2015). More recently, consumption of diosgenin (40 mg/kg bw) for 45 days decreased FBG (55%) and attenuated hyperlipidemia via inhibition of HMG-CoA reductase activity in STZ-induced diabetic rats (Hao et al., 2015).
Treatment of diosgenin (10–40 mg/kg bw) for 4 or 7 weeks demonstrated antihyperglycemic, antihyperlipidemic, cardioprotective and reno-protective potential in STZ-induced diabetic rats (Golshahi and Roghani-Dehkordi, 2016; Kanchan et al., 2016). However, Sato et al. (2014) have shown a weak reduction of FBG upon diosgenin (3 mg/kg bw) 24 h post-administration in STZ-induced diabetic rats, which may be apparently attributed to the short study period and lower dosage used.
Despite the fact that the detail toxicity studies of diosgenin has not been well documented, the oral LD50 was reported to be >8,000 mg/kg bw in rats (Ryndina et al., 1977). Furthermore, available toxicity studies on some animal models have shown that diosgenin (3.5% w/w) was safe and did not cause any toxicity in the treated animals (Raju and Rao, 2011).
Based on the results of the above-mentioned studies, diosgenin could be regarded as a potential hypoglycemic agent although clinical studies are required to fully confirm its hypoglycemic potential. Regardless of the few data available, diosgenin was observed to reduce FBG by >50% in several diabetic animal models and ameliorated diabetes-associated complications at non-toxic dosages (Pari et al., 2012; Sangeetha et al., 2013; Kalailingam et al., 2014; Saravanan et al., 2014; Hao et al., 2015; Naidu et al., 2015). Additionally, the potent attenuation of insulin resistance and hyperlipidemia at a concentration <10 μM is quite promising (Uemura et al., 2010; Liu et al., 2012; Sangeetha et al., 2013). Furthermore, despite few data regarding the safety issues associated with diosgenin consumption, the less toxic effect reported (LD50: >8,000 mg/kg bw) associated with diosgenin is of a great interest.
Eugenol is an active ingredient of cloves and other spices such as basil (Ocimum basilicum L.) and cinnamon. has diverse pharmacological potential such as hypoglycemic action. Eugenol (2.5–12.5 mM) demonstrated inhibitory actions on α-glucosidase (IC50: 326.1 μM) activity and advanced glycation end products (IC50: 10 μM) formation (Singh et al., 2016). Additionally, Mnafgui et al. (2013) highlighted that eugenol (10–100 μM) inhibited pancreatic α-amylase (IC50: 62.53 mg/ml) and lipase (IC50: 72.34 mg/ml) as well as angiotensin converting enzyme (ACE) activities (IC50: 130.67 mg/ml). However, despite higher IC50 values exhibited by the eugenol, the data signified the eugenol potential in ameliorating post-prandial hyperglycemia and diabetes-related oxidative damage and hypertension. Previously, eugenol (5–20 μM) was reported to prevent hyperglycemia in SHSY5Y cells (Prasad et al., 2015). Furthermore, eugenol (10–100 μM) stimulated muscle glucose uptake via increased GLUT4 and PI3K genes expression in L6 myotubes (Prabhakar and Doble, 2011).
Dietary supplementation of eugenol (200 mg/kg bw) for 2 weeks attenuated nerve and vascular dysfunction with no significant reduction of FBG in STZ-induced diabetic rats (Nangle et al., 2006). However, Mnafgui et al. (2013) have shown 62.5% reduction of FBG with potent antioxidant potential when eugenol (80 mg/kg bw) was administered orally for 30 days in alloxan-induced diabetic rats. Srinivasan et al. (2014) have reported that eugenol (2.5–10 mg/kg bw) treatment for the same study period demonstrated antihyperglycemic and antioxidant potential in STZ-induced diabetic rats. The highest reduction of FBG was about 70.6% with improved activities of key enzymes (hexokinase, pyruvate kinase, glucose-6-phosphatedehydrogenase, glucose-6-phosphatase, fructose-1,6-bisphosphatase) related to carbohydrate metabolism (Srinivasan et al., 2014).
In another study, oral administration of eugenol at 20 and 40 mg/kg bw for 15 weeks reduced FBG by 20 and 28.6%, respectively in high fat-fed C57BL/6J mice (Jeong et al., 2014). Furthermore, oral administration of eugenol (10 mg/kg bw) for 5 days or 6 weeks showed maximum reduction of FBG by 38% and improved the in vivo antioxidant status of STZ-induced diabetic rats (Prasad et al., 2015; Singh et al., 2016). This variation could be linked to the different animal models used. On the other hand, Rauscher et al. (2001) have reported that intraperitoneal treatment of isoeugenol (10 mg/kg bw) for 2 weeks did not show any antihyperglycemic effect in STZ-induced diabetic rats. Additionally, a moderate antioxidant potential was reported in the treated animals, indicating weak hypoglycemic potential (Rauscher et al., 2001).
The LD50 of eugenol administered orally to rats was >1,000 mg/kg (Sober et al., 1950; Taylor et al., 1964; Hagan et al., 1965). However, LaVoie et al. (1986) reported a lower LD50 of 11 mg/kg bw after intratracheal instillation in rats. Similarly, the toxic effects manifested include lung congestion with interstitial hemorrhages, acute emphysema, and acute pulmonary edema. Recently, treatment of eugenol (0.06 μM) showed genotoxicity and cytotoxicity on dental pulp fibroblasts (Escobar-García et al., 2016). Furthermore, eugenol (3 mmol/l) induced oral mucosal fibroblasts within 2 h post-administration period (Jeng et al., 1994).
Based on the above studies the potential of eugenol as hypoglycemic agent is not consistent and thus, need further extensive studies to establish the potency of eugenol hypoglycemic action. However, some studies highlighted >60% FBG reduction at non-toxic dosages (<1,000 mg/kg bw) and attenuation of diabetes-induced complications which are quite encouraging (Mnafgui et al., 2013; Srinivasan et al., 2014; Prasad et al., 2015). Therefore, according to the current literature, the above-mentioned studies have shown the potential of eugenol as adjuvant in the diabetes management.
Galactomannan is a heterogeneous water-soluble polysaccharide from fenugreek with a structural similarity to standard hypoglycemic drug, acarbose. Galactomannan (0.1 and 0.5% w/w) was reported to reduce intestinal glucose uptake in isolated intestine of lean and obese rats and thus improve glycemia (Srichamroen et al., 2009). Furthermore, galactomannan enhanced glucose uptake (51.9%) in isolated hemidiaphragm of treated alloxanized rats (Anwar et al., 2009). Kashef et al. (2008) have shown that galactomannan (200 mg/ml) inhibited the α-amylase activity. This imply that galactomannan could be beneficial in amelioration of post-prandial hyperglycemia in diabetes.
Dietary inclusion of galactomannan (2.5 and 5% w/w) attenuated post-prandial hyperglycemia, hyperlipidemia and abdominal fat deposit in high sucrose-fed rats (Srichamroen et al., 2008). Oral administration of galactomannan to STZ-induced diabetic rats inhibited maltase, lactase and sucrase activities in the small intestine of treated rats (Hamden et al., 2010). These studies support the in vitro data and further confirm the amelioration of post-prandial hyperglycemia by the galactomannan. In another study, oral administration of galactomannan (250–500 mg/kg bw) for 3 weeks reduced FBG (59.4%) and improved serum insulin levels in alloxan induced diabetic rats (Al-Fartosy, 2015). However, a reduction of about 40% on FBG level and improved antioxidant potential were reported upon 2 h post-administration of galactomannan (500 mg/kg bw) in the same animal model (Kamble and Bodhankar, 2013; Kamble et al., 2013). Kandhare et al. (2015) have reported that chronic consumption of galactomannan (60 and 100 mg/kg bw) for 12 weeks ameliorated hyperglycemia (50%) and insulin resistance in C57BL/6 mice.
Galactomannan was reported to be safe up to 8 g/kg with no deleterious effects after 3 days post-administration period (Anwar et al., 2009; Al-Fartosy, 2015). This was similarly reported even after repeated doses for 90 days (Deshpande et al., 2016a). To further support the galactomannan safety, oral administration during gestation induced no significant maternal and embryo-fetal toxicity up to 1,000 mg/kg bw in rats (Deshpande et al., 2016b).
Studies above have shown that little information are available regarding the hypoglycemic potential of galactomannan and thus strenuous to make logical conclusion. However, our observations showed that some studies used galactomannan at high dosages (500 mg/kg bw) or concentrations (200 mg/ml) in addition to being a high molecular weight molecule, signifying weak hypoglycemic action. Therefore, more detail studies are required to fully evaluate the hypoglycemic action of galactomannan both in humans and experimental animal models.
Gingerol ([6]-gingerol) and gingerol-related derivatives (shogaol, paradol and zingerol) are the prominent ingredients of ginger and other members of Zingiberaceae.
Li and co-authors have reported that gingerols (50–150 μM) enhanced glucose uptake in L6 myotubes and muscle C2C12 cells, attributed to an increased surface availability of GLUT4 protein and by activation of AMPK in the cells (Li Y. et al., 2012; Li et al., 2013; Son et al., 2015). Available studies have shown that diabetes leads to an increase accumulation of β-amyloid, a major component of senile plaques, leading to β-cell dysfunction and failure (Maher and Schubert, 2009; Takeda et al., 2011; Luo et al., 2016). Interestingly, [6]-gingerol (2.5–20 μM) attenuated β-amyloid-induced oxidative cell death in SH-SY5Y neuroblastoma cells (Lee C. et al., 2011).
Furthermore, in a number of previous studies, [6]-gingerol was shown to play a beneficial role in reducing lipid accumulation in 3T3 cells via downregulating PPARγ and decreasing Akt/GSK3β pathway (Isa et al., 2008; Tzeng and Liu, 2013; Tzeng et al., 2014; Choi et al., 2016; Suk et al., 2016). Reducing lipid accumulation may delay the onset and progression of insulin resistance in diabetes. [6]-Gingerol (10 μM) was also reported to prevent diabetes-induced diastolic dysfunction in isolated murine ventricular myocardia (Namekata et al., 2013). In our recent study, [6]-gingerol (30–240 μg/ml) inhibited α-amylase (IC50: 81.8 μM) and α-glucosidase (IC50: 21.6 μM) actions, signifying its potential in ameliorating post-prandial hyperglycemia (Mohammed et al., 2017).
[6]-Shogaol (25 μM) inhibited the TNF-α mediated downregulation of adiponectin expression in 3T3-L1 adipocytes via inhibition of c-Jun-NH2-terminal kinase action (Isa et al., 2008). This prevents increased production of pro-inflammatory mediators and oxidative stress markers. Wei et al. (2017) have shown that 6-shogaol (100 μM) promoted glucose utilization via AMPK phosphorylation in 3T3-L1 adipocytes and C2C12 myotubes. [6]-Shogaol (30–240 μg/ml) showed weak α-amylase (IC50: 443.2 μM) and α-glucosidase (IC50: 326.1 μM) inhibition via non-competitive mode of inhibition (Mohammed et al., 2017).
It has been reported that [6]-paradol (100 μM) stimulated glucose utilization via AMPK phosphorylation in 3T3-L1 adipocytes and C2C12 myotubes, which apparently improved insulin sensitivity of the target tissues (Wei et al., 2017). More recently, [6]-paradol (30–240 μg/ml) exhibited weak inhibitory actions toward α-amylase (IC50: 664.6 μM) and α-glucosidase (IC50: 243.3 μM) actions (Mohammed et al., 2017).
Singh et al. (2009) reported that oral treatment of [6]-gingerol (100 mg/kg bw) for 12 days reduced FBG (57.1%) in db/db mice with potent antihyperlipidemic and antioxidant actions. Oral consumption of [6]-gingerol (75 mg/kg bw) for 3 weeks reduced FBG (42%) via upregulation of GLUT4, IRS-1, IRS-2, PI3K, AKT, PPARα pathways in sodium arsenate hyperglycemic mice (Chakraborty et al., 2012). This supports the previous in vitro studies that showed the [6]-gingerol potential to increase GLUT4 protein availability and activate AMPK (Li Y. et al., 2012; Li et al., 2013; Son et al., 2015). In addition, modulation of enzymes activities involved in gluconeogenesis and glycogenolysis was also proposed as possible mechanism involved in the hypoglycemic effect of [6]-gingerol (Son et al., 2015).
Intraperitoneal treatment of [6]-gingerol (3 or 75 mg/kg bw) for 8 weeks demonstrated antihyperglycemic (10%), cardioprotective potential, improved post-prandial glucose utilization and insulin sensitivity in STZ-induced diabetic rats (Shao et al., 2016). Similarly, Sampath et al. (2016, 2017) have shown maximum FBG (50%) reduction and potent aldose reductase inhibition upon intraperitoneal administration of [6]-gingerol (25 and 75 mg/kg bw) three times per week for 16 weeks to C57BL/6J hyperlipidemic mice. The oral treatment showed higher reduction of FBG relative to the study period compared to the intraperitoneal injection.
Oral administration of [6]-paradol (33.75 mg/kg bw) for 8 weeks decreased FBG (37.6%) in high-fat diet-fed mice (Wei et al., 2017).
Oral administration of zingerone (10 mg/kg bw) for 4 weeks reduced FBG (64.1%), improved the levels of hematological parameters and attenuated dyslipidemia in STZ-induced diabetic rats (Jothi et al., 2016a,b).
Fewer data are available regarding the potential toxicity associated with the intake of [6]-gingerol and its derivatives. Consumption of [6]-gingerol (20–80 μM) induced genotoxicity, lysosomal and mitochondrial damage in human hepatoma G2 (HepG2) cells (Yang et al., 2010). However, consumption of either [6]-gingerol or [6]-shogaol (2,000 mg) for 4 days was reported not to cause any potential toxicity in human subjects and are well tolerated (Zick et al., 2008). On the other hand, the LD50 of zingerone was reported to be 1,000 mg/kg bw (Rao et al., 2009).
From the above-mentioned studies, it is obvious that information regarding the hypoglycemic potential and toxicity of [6]-gingerol and its derivatives are scanty and that makes the overall comment inconclusive. Moreover, according to the data, [6]-gingerol showed higher hypoglycemic potential compared to its derivatives (Singh et al., 2009; Namekata et al., 2013; Sampath et al., 2016; Mohammed et al., 2017). Therefore, more studies are required on [6]-gingerol and notwithstanding, these compounds could be regarded as hypoglycemic adjuvant as reported in the above-mentioned studies.
4-Hydroxyisoleucine (4-OH-Ile) is an active ingredient of fenugreek and most of the hypoglycemic effect of fenugreek are attributed to 4-OH-Ile. Studies available have reported the beneficial effect of 4-OH-Ile in the control of diabetes and its associated complications. Jaiswal et al. (2012) have reported that 4-OH-Ile (5–25 μM) stimulated glucose uptake in L6-GLUT4 myc myotubes, which has been recently confirmed by Korthikunta et al. (2015). In another study, 4-OH-Ile (10 μM) ameliorated insulin resistance in L6 myotubes (Rawat et al., 2014). Moreover, using the same model, 4-OH-Ile (5–25 μM) was shown to ameliorate insulin resistance and demonstrated potent anti-inflammatory action (Maurya et al., 2014). In HepG2 cells, 4-OH-Ile (100 ng/ml) promoted insulin signaling and the expression of glycogenic enzymes and GLUT2 (Naicker et al., 2016). Previously, 4-OH-Ile (100–1,000 μM) stimulated insulin release in isolated rat pancreas (Sauvaire et al., 1998; Broca et al., 2000; Wang et al., 2002). Therefore, it is clear that 4-OH-Ile showed potential to ameliorates insulin resistance. The possible mechanisms involve increased Akt phosphorylation and reduced activation of Jun N-terminal kinase (JNK)1/2, extracellular signal-regulated kinase (ERK)1/2, p38 mitogen-activated protein kinase (MAPK), and nuclear factor (NF)-κB (Avalos-Soriano et al., 2016).
Haeri et al. (2012) have reported that oral administration of 4-OH-Ile (50 mg/kg bw) for 4 weeks to STZ-induced diabetic rats reduced FBG (41%) with potent hypolipidemic and insulinotropic actions. In another study, a reduction of 34% on FBG was reported in STZ-induced diabetic rats upon treatment of 4-OH-Ile (50 mg/kg bw) for 8 weeks (Narender et al., 2006; Haeri et al., 2009). Intraperitoneal administration of 4-OH-Ile (18–50 mg/kg bw) for 15 min or 5 days showed insulinotropic action with no effect on blood glucose levels in Zucker diabetic fa/fa or STZ-induced diabetic rats (Broca et al., 1999, 2004). This supports the insulinotropic potential of 4-OH-Ile and is in line with in vitro data (Sauvaire et al., 1998; Broca et al., 2000; Wang et al., 2002). In C57BL/KsJ-db/db mice, oral treatment of 4-OH-Ile (50 mg/kg bw/ 10 days) lowered FBG by 55.4% (Singh et al., 2010). Consumption of 4-OH-Ile (40 mg/kg bw) for 7 weeks showed antihyperglycemic action and a potent pancreatic β-cell regeneration in alloxan-induced diabetic mice (Shah et al., 2009).
Nuttall et al. (2008) have reported that ingestion of 4-OH-Ile (1 mmol/kg lean body mass) reduced blood glucose levels and improved utilization compared to the untreated non-diabetic subjects after 4 h post-administration period.
Oral LD50 value of 4-OH-Ile was reported to be >5 g/kg bw indicating that consumption of 4-OH-Ile has no potential toxic effect (Shah et al., 2009).
Our candid opinion here is that 4-OH-Ile did not show a significant hypoglycemic action despite long administration period. However, based on the available data, 4-OH-Ile is insulinotropic and could be used in combination with other drugs to attenuate diabetes-induced oxidative damage, and hence regarded as an adjuvant. Most importantly, detail toxicological studies are required to evaluate the safety of 4-OH-Ile both in humans and experimental animals.
Piperine is the major alkaloid responsible for the pungency of black pepper (Piper nigrum L.). Kumar S. et al. (2013) have reported that piperine showed weak inhibition toward α-lipase (IC50: 2,490 μg/ml), α-glucosidase (IC50: 2,550 μg/ml) and aldose reductase (IC50: 2,375 μg/ml) activities. Inhibition of the activities of these enzymes signify its potential in attenuating diabetes-associated complications.
Kharbanda et al. (2016) have reported that piperine (36 mg/kg bw) isolated from black pepper demonstrated antihyperglycemia in STZ-induced diabetic rats by acting as PPAR-γ agonists. Interestingly, Atal et al. (2016) have shown that co-administration of piperine (10 mg/kg bw) with metformin for 4 weeks reduced FBG (40%) compared to metformin alone (19%) in STZ-induced diabetic mice, indicating synergistic effect between the two drugs. Furthermore, weak reduction on FBG was observed upon administration of piperine (10–50 mg/kg bw) for the same study period and model (Rauscher et al., 2000; Kharbanda et al., 2016). However, the levels of serum insulin, lipid profiles and antioxidant enzymes were significantly improved. Oral administration of piperine (20 or 40 mg/kg bw) for 11 weeks ameliorated hyperglycemia (40%) and oxidative damage in STZ-induced diabetic rats (Arcaro et al., 2014). On the other hand, several studies have reported the beneficial effect of piperine in reducing hyperglycemia and attenuating oxidative stress in high-fat diet rats (Vijayakumar et al., 2004; Shah et al., 2010; Bao et al., 2012; BrahmaNaidu et al., 2014).
The safety aspect of piperine has been controversial. Piyachaturawat et al. (1983) have reported that the LD50 values of piperine via different route are in the order intravenous (15.1 mg/kg bw) < intraperitoneal (43 mg/kg bw) < subcutaneous (200 mg/kg bw) < intragastric (330 mg/kg bw) < intramuscular (400 mg/kg bw). The authors further showed that almost all the animals that received a lethal dose (>LD50) died from respiratory complications in <20 min. However, during sub-chronic study, the death occurred within l−3 days after post-administration period. Some of the histopathologic alterations observed include severe hemorrhagic necrosis and edema in GIT, urinary bladder and adrenal glands (Piyachaturawat et al., 1983). Additionally, its toxic effect has been attributed to its structural similarity with some known carcinogens such as safrole, estragole, and methyleugenol (Ames, 1983). On the other hand, consumption of piperine orally (170 mg/kg bw) or intraperitoneally (85 mg/kg bw) did not cause any adverse consequences in rats, with 3% excreted as piperine in the feces (Bhat and Chandrasekhara, 1986).
The information derived from the above-mentioned studies revealed that piperine is a weak hypoglycemic agent despite longer administration period. None of the studies have shown up to 50% reduction on blood glucose levels. The higher IC50 value depicted toward α-glucosidase and aldose reductase inhibitions indicated weak hypoglycemic action as well. Another major concern is the contradiction on the safety issues regarding pure piperine consumption. However, piperine could be regarded as food adjuvant in the management of diabetes based on the potent antioxidant action observed in the above-mentioned studies. Additionally, the use piperine as a naturally-based bio-enhancers to some drugs is receiving much attention and yielding fruitful results (Moorthi and Kathiresan, 2013; Arcaro et al., 2014).
Thymoquinone (TQ) is the main pharmacologically active ingredient of black cumin seeds (Nigella sativa L.), with proven hypoglycemic potential. TQ (2.5 μM) promoted glucose-stimulated insulin secretion and attenuated oxidative damages induced by protease inhibitors in rat pancreatic β-cells Chandra et al. (2009). Previously, TQ (10–50 μM) demonstrated potent antiglycation (IC50: 7.2 μM) potential (Losso et al., 2011; Anwar et al., 2014; Khan et al., 2014). Furthermore, TQ (3 mg/kg) reduced diabetes-induced elevated levels of macrophage-derived inflammatory mediators such as TNF-α, nitrite and IL-1β in isolated STZ-induced diabetic rat model peritoneal macrophages (El-Mahmoudy et al., 2005a). More recently, TQ (0–5 μM) improved insulin secretion from pancreatic β-cells in INS-1 cells (Gray et al., 2016).
Thymoquinone (0.5–6 mg/kg bw) administered intraperitoneally showed hypoglycemic (39.7% reduction) potential in non-diabetic rats (Hawsawi et al., 2001). Oral administration of TQ (50 mg/kg bw) for 12 weeks reduced FBG (37%), stimulated insulin release and improved histopathological changes in sciatic nerves of the STZ-induced diabetic rats (Kanter, 2008, 2009). However, a reduction of 45% on FBG was reported upon oral gavage of TQ (40 mg/kg bw) for 3 weeks in the same animal models (Bashandy et al., 2015). Interestingly, similar reduction of FBG was observed after 2 h post-TQ (60 mg/kg bw) administration period in STZ-nicotinamide-induced diabetic rats (El-Ameen et al., 2015).
In some studies, TQ (50 mg/kg bw) treatment for 3 or 4 weeks reduced FBG by 63% compared to 76% in insulin treated rats in the same animal models (Fararh et al., 2005, 2010). In addition, the activities of hepatic gluconeogenic enzymes were decreased (Fararh et al., 2010). Moreover, oral administration of TQ (20–80 mg/kg bw) for 45 days reduced FBG (61%), improved glucose tolerance, serum insulin levels and antioxidant status in STZ-nicotinamide-induced diabetic rats (Pari and Sankaranarayanan, 2009; Roghani and Baluchnejadmojarad, 2012). Additionally, improved activities of hexokinase, glucose 6-phosphate dehydrogenase, glucose 6-phosphatase and fructose 1, 6-bisphosphatase were observed as well.
To further support this, administration of TQ (20–80 mg/kg bw) for 12 weeks reduced FBG (>70%) STZ-nicotinamide-induced diabetic rats (Fouad and Alwadani, 2015). Oral TQ (80 mg/kg bw) consumption ameliorated diabetes-induced pancreatic oxidative damages with subsequent improvement of antioxidant status in the same animal model (Sankaranarayanan and Pari, 2011a,b). Furthermore, considerable data are available showing TQ (3–50 mg/kg bw) potential in attenuating diabetes-induced oxidative damages via increased expression of antioxidant enzymes in STZ-induced diabetic or hyperlipidemic rats (Abdelmeguid et al., 2010; Mehrdad and Tourandokht, 2012; Ahmad and Beg, 2013; Al Wafai, 2013; Elmansy and Almasry, 2013; Hafez, 2013; Ashour, 2015; Bashandy et al., 2015; Desai et al., 2015; Al-Trad et al., 2016; Saheb et al., 2016).
Salehi et al. (2012) have reported that TQ treatment (2.5 and 5 mg/kg bw) for 5 weeks improved the spatial memory in STZ-induced diabetic rats via attenuation of lipid peroxidation. Furthermore, oral gavage of TQ (10 mg/kg bw) for 2 weeks reduced FBG (66%) and improved the antioxidant status of STZ-induced diabetic rats (Hamdy and Taha, 2009). El-Mahmoudy and colleagues have reported similar reduction of FBG upon oral consumption of TQ (3 mg/kg bw) for 30 days in LETO-STZ-induced diabetic rats (El-Mahmoudy et al., 2005b). Supplementation of TQ (20 mg/kg bw) during pregnancy and lactation periods to STZ-induced gestational diabetic rats induced FBG (20%) and pro-inflammatory cytokines levels (IL-1b, IL-6, IL-2, and TNF-α) in the offspring (Badr et al., 2011, 2013). Surprisingly, TQ administered intraperitoneally at 3 and 5 mg/kg bw for 8 weeks reduced FBG by 68 and 66%, respectively which could be attributed to the longer period of administration (Sangi et al., 2015).
Tremendous efforts were made to assess the toxicological properties of TQ using various in vitro and in vivo models (El-Dakhakhny, 1965; Badary et al., 1998; Mansour et al., 2001; Al-Ali et al., 2008; Khader et al., 2009; Qadri et al., 2009; Abukhader, 2012). The LD50 of TQ in rats via oral and intraperitoneal administration were 794.3 and 57.5 mg/kg, when in mice the values were 870.9 and 104.7 mg/kg through oral and intraperitoneal route, respectively (Al-Ali et al., 2008). Previously, Badary et al. (1998) have shown that the acute LD50 value in mice was 2.4 g/kg bw via oral ingestion of TQ. This indicates the relatively low toxicity of TQ since the LD50 values were >10 and >100 times higher than the therapeutic dosages for TQ via intraperitoneal and oral routes, respectively. However, few signs of the toxicity such as hypoactivity and difficulty in respiration were observed after acute oral administration of TQ in rats (Badary et al., 1998).
Moreover, sub-chronic administration of TQ (35 and 50 mg/kg bw) induced disruption on embryonic development during the second trimester of rat pregnancy (Abukhader, 2012). Conversely, TQ (30–90 mg/kg bw/day) administration for 3 months caused no mortality or sign of toxicity in mice (Badary et al., 1998). Interestingly, Al-Amri and Bamosa (2009) have reported that oral ingestion of TQ for 3 weeks did not show any potential toxicity and was well tolerated up to dose of 2,600 mg/kg bw in human subjects. However, according to the authors TQ administration showed no therapeutic potential up to the maximum dosage used (Al-Amri and Bamosa, 2009).
Based on the above-mentioned studies, TQ possessed blood glucose lowering potential and could be used to attenuate diabetes-induced complications despite lack of relevant clinical trials. Our rationale is that TQ demonstrated hypoglycemic potential at 3–50 mg/kg bw in animal models (Hamdy and Taha, 2009; Pari and Sankaranarayanan, 2009; Roghani and Baluchnejadmojarad, 2012) and depicted IC50 value <10 μM at the concentrations (10–50 μM) in addition to stimulating insulin release at 2.5 μM (Chandra et al., 2009). Moreover, most of the studies have reported more than 50% reduction on blood glucose levels and potent antioxidant actions (Fararh et al., 2005, 2010; Hamdy and Taha, 2009; Pari and Sankaranarayanan, 2009; Roghani and Baluchnejadmojarad, 2012; Fouad and Alwadani, 2015; Sangi et al., 2015). However, lack of detail hypoglycemic and toxicity studies in human subjects are the major concerns.
Trigonelline is a spice-derived alkaloid from fenugreek and possesses tremendous therapeutic potential including hypoglycemic potential. Trigonelline (0.33 and 3.3 mg/ml) inhibited glucose uptake (IC50: 19 mM) in isolated intestinal rabbits (Al-Habori et al., 2001). More recently, Ilavenil et al. (2014) have shown that trigonelline (75 or 100 μM) attenuated adipocyte differentiation and subsequent hyperlipidemia in 3T3-L1 cells.
Oral administration of trigonelline (50–100 mg/kg bw) for 4 weeks showed maximum FBG reduction of 27%, attenuated TNF-α levels and improved insulin levels in neonatal STZ-induced diabetic rats (Ghule et al., 2012). In addition, glomerular filtration rate, activities of antioxidant enzyme and membrane bound enzymes were improved in treated animals. Subsequently, oral consumption at 10 mg/kg bw for 4 weeks demonstrated antihyperglycemia, antihyperlipidemic and antioxidant potential in alloxan-induced diabetic rabbits (Monago and Nwodo, 2010; Al-Khateeb et al., 2012). The highest reduction of FBG was 74.5% compared to 61.1% for Chlorpropamide (Monago and Nwodo, 2010). More recently, supplementation of trigonelline (150 mg/kg bw) for 30 days reduced FBG (50%), hyperlipidemia and diabetes-induced oxidative damages in high-fat diet-fed low-dose STZ-induced diabetic rats model (Subramanian and Prasath, 2014a,b).
Furthermore, Shah and colleagues have reported similar reduction of FBG (>50%) after 24 h treatment in alloxan-induced diabetic rats (Shah et al., 2006). Additionally, repeated oral administration of trigonelline (75 mg/kg bw) for 7 days have shown about 57% reduction on FBG and improved the histology of pancreas of treated rats (Shah et al., 2006). Interestingly, a potent antioxidant potential of trigonelline (10 mg/kg bw) was later documented in alloxan-induced diabetic rats (Hamadi, 2012). Trigonelline (25–100 mg/kg bw) exhibited maximum FBG reduction of 16% after acute (24 h) administration period in nicotinamide STZ-induced diabetic rats (Kamble and Bodhankar, 2014). However, after trigonelline (50 mg/kg bw) treatment for 8 weeks the FBG reduction was lower (48%) as compared to trigonelline treated and sitagliptin (5 mg/kg bw) combination (63%) when same animal models were used (Kamble and Bodhankar, 2013,b).
Dietary inclusion of trigonelline (0.056%) for 43 days demonstrated weak hypoglycemic action (<10% FBG reduction) and potent antioxidant potential in Goto-Kakizaki type 2 diabetes rats (Yoshinari et al., 2009, 2013; Yoshinari and Igarashi, 2010). In another study, supplementation of trigonelline (40 mg/kg bw) for 48 weeks reduced FBG (75%), ameliorated insulin resistance and peripheral diabetic neuropathy in high-fat diet-fed STZ-induced diabetic rats (Zhou and Zhou, 2012). Furthermore, the same authors have reported the FBG reduction of about 70% after 4-week post-administration period at the same dose and in the same animal model (Zhou et al., 2011). However, a weak FBG reduction (38%) of trigonelline was later reported upon treatment for either 2 or 4 weeks at the same dose and animal models, attributed to the short study period (Tharaheswari et al., 2014, 2015). Conversely, Hamden et al. (2013a) have reported 50% FBG reduction after oral administration of trigonelline (100 mg/kg bw) for 30 days in alloxan-induced diabetic rats. Moreover, the authors showed that trigonelline treatment significantly inhibited the activities of dipeptidyl peptidase-IV, α-glucosidase and angiotensin converting enzyme (Hamden et al., 2013a,b). Interestingly, trigonelline (50 mg/kg bw) treatment orally for 4 weeks reduced FBG (81%) in nicotinamide-STZ-induced diabetic rats (Folwarczna et al., 2016). More recently, similar finding was also noticed after 2-week post-administration of trigonelline (50–100 mg/kg bw) in fructose-induced insulin resistance (Ramadan et al., 2016).
Ingestion of trigonelline (500 mg) reduced blood glucose by about 7% and improved glucose tolerance after 15 min post-treatment period in overweight subjects (Van Dijk et al., 2009). The hypoglycemic action was found not to be dependent on the incretin hormones glucagon-like peptide 1 (GLP-1) or glucose-dependent insulinotropic peptide (Olthof et al., 2011).
Aswar et al. (2009) have shown that oral consumption of trigonelline was safe up to 5,000 mg/kg bw with no noticeable abnormal behavior in rats.
As per data from the above studies, trigonelline seems to be among the promising hypoglycemic agents despite few studies which showed weak or no significant hypoglycemic potential. Longer administration period such as 4 weeks at 10–100 mg/kg bw showed potent reduction (>50%) of FBG in diabetic rats (Shah et al., 2006; Monago and Nwodo, 2010; Al-Khateeb et al., 2012; Zhou et al., 2011; Hamden et al., 2013a; Subramanian and Prasath, 2014a,b; Folwarczna et al., 2016). Additionally, a potent amelioration of diabetes-induced complications was observed even in those studies that showed weak hypoglycemic potential. The weak blood glucose lowering potential in humans could be attributed to the shorter study period (Van Dijk et al., 2009). Although few data are available regarding trigonelline toxicity, its consumption did not show any potential toxic effect in rats (Aswar et al., 2009).
Conventionally, poor bioavailability is considered as a major factor linked to the lower therapeutic efficacy of the orally consumed SDBI. Therefore, improving bioavailability of the ingredients is a promising approach in enhancing their disease preventing efficacy in humans. The oral bioavailability of SDBI entails the portion of the ingested ingredient that get in to the blood circulation in its active form. Because, only bioavailable portion will be absorbed and distributed across the tissues and organs that eventually exert its therapeutic effects. Moreover, poor solubility in gastrointestinal fluids and slow absorption rate from the GIT are the crucial factors that thwart SDBI from reaching the systemic circulation in their active forms (Yao et al., 2015).
According to our critical observation, cinnamaldehyde, curcumin, diosgenin, TQ and trigonelline are the promising hypoglycemic SDBI despite their known poor bioavailability in the physiological system. Interestingly, there has been a renewed interest in developing methods that may improve the bioavailability of the SDBI to prevent or treat human diseases such as diabetes. In this regard, we have briefly presented some fact regarding the bioavailability of these promising ingredients and the methods being used to improved their bioavailability.
Cinnamaldehyde is absorbed rapidly from the gut, utilized and excreted via urine, regardless of the dosages, species and sex of the animals used. Oral consumption of cinnamaldehyde was shown to be metabolized into cinnamic acid partially in the stomach and small intestine and then completely metabolized into cinnamic acid in the liver before it enters the circulation (Chen et al., 2009). Previously, Yuan et al. (1993) and Peters and Caldwell (1994) have reported that the intravenous administration of the various dosages of cinnamaldehyde (5–25 mg/kw bw) to F344 rats decreased blood glucose 30 min after the dose administration. The disappearance of cinnamaldehyde is attributed to the rapid oxidation to cinnamic acid in blood (about 37–60%). This is because 1.7 h half-life has been considered for cinnamaldehyde release from the protein adducts (Yuan et al., 1992). Furthermore, the authors highlighted that the blood level of cinnamaldehyde after oral consumption was maintained 1 μg/ml for 24 h (Yuan et al., 1992). More recently, the elimination time of cinnamaldehyde (125–500 mg/kg bw) were 6.7 and 1.7 h for oral and intravenous administration, respectively and the oral bioavailability of about 20% in the blood (Hooth et al., 2004).
Based on the data available, improving cinnamaldehyde bioavailability focused on three major processes including use of cinnamaldehyde derivatives or metabolites, micelle, microencapsulation and nanoparticles approaches (Hooth et al., 2004; Raffai et al., 2014; Wani et al., 2014; Jo et al., 2015). However, with the exception of using cinnamaldehyde derivatives, none of the methods was so far employed regarding the hypoglycemic potential of cinnamaldehyde either in vitro, in vivo or in human subjects. These approaches could be used to explore the hypoglycemic potential of cinnamaldehyde and therefore, warrant for further study in this regard.
It is well-established that curcumin is poorly bioavailable and thus its pharmacological effects are compromised. The low plasma and tissue levels of curcumin has been attributed not only to its poor absorption but rapid hepatic metabolism and systemic elimination (Anand et al., 2007; Cui et al., 2009; Bansal et al., 2011). For instance, about 51 ng/ml of curcumin was detected in the serum after 4 h oral consumption of curcumin (12 g) in healthy human subjects (Lao et al., 2006b). However, Marczylo et al. (2009) showed a relatively higher distribution of curcumin (340 mg/kg bw) in plasma (16.1 ng/ml), urine (2.0 ng/ml), intestinal mucosa (1.4 mg/g), liver (3,671.8 ng/g), kidney (206.8 ng/g), and heart (807.6 ng/g) after 2 h post-oral treatment. Previously, oral ingestion of curcumin (400 mg) showed about <20 μg/tissue levels in the kidney or liver, when no curcumin or trace amount was found in the urine in rats after 24 h post-administration period (Ravindranath and Chandrasekhara, 1982).
Furthermore, about 60–67% of curcumin (10–400 mg/kg bw/ 12 days) was absorbed and maintained at relatively constant amount in the circulation independent of the dose administered (Ravindranath and Chandrasekhara, 1982). In another study, administration of curcumin (0.1 g/kg bw) intraperitoneally showed tissue distribution of 177.04, 26.06, 26.90, 7.51, and 0.41 μg/g in the intestines, spleen, liver, kidneys and brain, respectively after 1 h treatment in mice (Pan et al., 1999). Regarding the curcumin metabolism, curcumin undergoes bioreduction to dihydrocurcumin and tetrahydrocucurmin which are then converted to either glucuronide or sulfate conjugates in the body system (Garcea et al., 2004).
In summary, the above-mentioned studies demonstrated the poor bioavailability of curcumin. Interestingly, tremendous efforts are introduced to alternatively increases the bioavailability, prolonged circulation, better permeability, and resistance to metabolic reactions of curcumin. These processes include the use of everted sacs of rat intestines, use of adjuvant that interferes with glucuronidation and the use of liposomal curcumin (Suresh and Srinivasan, 2007; Shaikh et al., 2009). Others are the use of nanoparticles, curcumin phospholipid complex and the structural analogs of curcumin (Suresh and Srinivasan, 2007). For instance, the oral bioavailability of curcumin was reported to improve by 9-fold using nanoparticles approach (Shaikh et al., 2009). Moreover, the bioavailability of THC has been recently shown to be higher compared to the curcumin (Aggarwal et al., 2014).
However, with regard to improving hypoglycemic potential of curcumin, available literatures have shown that the use of curcumin derivatives and nanoparticles approaches were employed in some diabetic models. The hypoglycemic potential of the former has been addressed in the earlier section of this review (Pari and Murugan, 2005, 2007b,?; Murugan and Pari, 2006a,b; Murugan et al., 2008; Karthikesan et al., 2010a,b; Lekshmi et al., 2012a,b). For the later, Grama et al. (2013) have reported that oral administration of nano-curcumin (2 mg/kg bw) for 11 weeks reduced FBG (37%) and delayed cataract formation in STZ-induced diabetic rats. Recently, intranasal delivery of nano-micelle curcumin for 7 days was shown to significantly promote corneal epithelial/nerve healing in STZ-induced diabetic mice (Guo et al., 2016). In a randomized clinical trial, ingestion of nano-curcumin (80 mg) for 3 months reduced FBG and glycated hemoglobin by about 32 and 19%, respectively in type-2 diabetic patients (Rahimi et al., 2016).
Based on the above few studies, it is obvious that nanoparticles approach may be another option to improve the antihyperglycemic as well as hypoglycemic efficacy of curcumin when compared to the use of curcumin metabolites or derivatives as pure compounds. However, the reduction of FBG was less with curcumin nanoparticles compared to that of curcumin alone or its derivatives. Hence, further studies are required to ascertain the efficacy of curcumin nanoparticles or come up with a more improved method.
The therapeutic applications of diosgenin are greatly tempered due to the poor pharmacokinetics. Cayen et al. (1979) have reported that 1 μg/ml of diosgenin was recovered from the serum of human subjects that received diosgenin (3 g/day) for 4 weeks, indicating poor absorption and bioavailability of diosgenin. Furthermore, oral bioavailability of diosgenin was highlighted to be 6% in rats and aqueous solubility was found to be 0.95 μg/ml (Okawara et al., 2010, 2013).
To improve the solubility and intestinal permeability of diosgenin, Kim et al. (2012) have shown that conjugating the hydrophilic unit, tetraethylene glycol to form diosgenin-tetraethylene glycol conjugate improved the hypoglycemic action of diosgenin. Although both the diosgenin and the conjugate treatment (10–20 mg/kg bw) for 9 weeks did not show any significant FBG reduction, the conjugate treated group showed better potential compared to the diosgenin alone (Kim et al., 2012; Okawara et al., 2013). Interestingly, some methods are available to improve the bioavailability of diosgenin, although not directly investigated in any diabetic model. The use of diosgenin and β-cyclodextrin inclusion complex, deglycosylation of diosgenin and diosgenin nanocrystals are receiving much attention in the recent years (Gao et al., 2012; Okawara et al., 2013, 2014; Liu et al., 2016). For instance, the use of diosgenin and β-cyclodextrin inclusion complexes improved the bioavailability of diosgenin by 45% in rats (Okawara et al., 2013).
Poor bioavailability in the systemic circulation has been highlighted as the major limitation for using TQ in clinic trials. Alkharfy et al. (2015) have attributed the poor bioavailability of TQ to the rapid elimination and relatively delay absorption following oral administration. Previously, Pathan et al. (2011) have detected TQ in the plasma (about 58%) for 12 h after oral administration of TQ (20 mg/kg bw) in rats. In another study, TQ was reported to accumulate greatly in the entire nuclei of kidney cells (Effenberger-Neidnicht et al., 2011).
The consequences of TQ hydrophobicity leads to reduce amounts reaching the target which in turn increase toxicity to normal tissues. Taking into consideration many researchers have developed more aqueous-soluble TQ derivatives and encapsulate in a nanoformulations to overcome the poor bioavailability of TQ. The encapsulation increases the bioavailability, protects the TQ from prematured enzyme degradation and limits TQ diffusion to normal tissues (Schneider-Stock et al., 2014). Interestingly, this method has been widely used in various disease condition and improved actions with less toxicity were reported (Ravindran et al., 2010; Singh et al., 2013; Ong et al., 2016; Kalam et al., 2017). Unfortunately, no study is available that reported the potential of TQ-derived nanoparticle in diabetic models and hence further studies in this regard are warranted.
The solubility of trigonelline was reported to be higher compared to other SDBI and thus showed moderate rate of absorption and high elimination rate in the rabbit (Zhao et al., 2003). Similarly, most of the consumed trigonelline is usually absorbed at the small intestine and not degraded by the microflora in germ-free and specific pathogen-free rats (Yuyama, 1999). Consumption of coffee (another rich-source of trigonelline) showed peak plasma concentrations of 5.5–6.5 μM after 2–3 h in human subjects (Lang et al., 2010, 2013). The authors further showed delay clearance and, hence, accumulation of trigonelline in the plasma with an average half-life of about 5 h. In addition, about 50% of the trigonelline consumed was detected in urine 0–8 h post-ingestion (Lang et al., 2010, 2013). Moreover, about 20% of the ingested trigonelline was reported to be excreted in the urine as trigonelline when approximately 9% was excreted as N'-methyl-2-pyridone-5-carboxylic acid (Yuyama and Suzuki, 1991; Yuyama and Kawano, 1996). However, about 100% of trigonelline was recovered unchanged from the urine in the administered rats (Shibata and Taguchi, 1991). Hence, further studies are needed not only to confirm the true metabolism of trigonelline but also to clear the above-mentioned controversies.
According to the literature, no study is available that highlight the method that could improve the trigonelline bioavailability. This could be attributed to the higher solubility of trigonelline compared to the other SDBI.
Data gathered in the present study have shown that SDBI hold promising hypoglycemic potential. Despite many of the ingredients showed weak hypoglycemic effects, cinnamaldehyde, curcumin, diosgenin, TQ and trigonelline have demonstrated promising hypoglycemic potential and need further scientific scrutiny to maximize their use as hypoglycemic therapies and adjuvants. Therefore, future studies on the most promising SDBI should be focused on bringing these ingredients to the forefront for the treatment of diabetes. These can be achieved via extensive clinical trials, improving the tissue bioavailability and distribution as well as detail toxicological studies. Furthermore, more detail studies are required on the combinatory effects of standard hypoglycemic drugs and the ingredients that showed adjuvant property. The idea is to evaluate whether the combined administration could attenuate advance consequences of the synthetic drugs as they showed strong amelioration of diabetes-associated complications.
AM gathered all the previously published articles and drafted the manuscript in its current form under the direct guidance of MI.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fphar.2018.00893/full#supplementary-material
Abdel Aziz, M. T., El-Asmar, M. F., Rezq, A. M., Mahfouz, S. M., Wassef, M. A., Fouad, H. H., et al. (2013). The effect of a novel curcumin derivative on pancreatic islet regeneration in experimental type-1 diabetes in rats (long term study). Diabetol. Metab. Syndr. 5, 1–14. doi: 10.1186/1758-5996-5-75
Abdelmeguid, N. E., Fakhoury, R., Kamal, S. M., and Al Wafai, R. J. (2010). Effects of Nigella sativa and thymoquinone on biochemical and subcellular changes in pancreatic β-cells of streptozotocin-induced diabetic rats. J. Diabetes 2, 256–266. doi: 10.1111/j.1753-0407.2010.00091.x
Abdul-Hamid, M., and Moustafa, N. (2013). Protective effect of curcumin on histopathology and ultrastructure of pancreas in the alloxan treated rats for induction of diabetes. J. Basic Appl. Zool. 66, 169–179. doi: 10.1016/j.jobaz.2013.07.003
Abukhader, M. M. (2012). The effect of route of administration in thymoquinone toxicity in male and female rats. Indian J. Pharm. Sci. 74, 195–200. doi: 10.4103/0250-474X.106060
Acar, A., Akil, E., Alp, H., Evliyaoglu, O., Kibrisli, E., Inal, A., et al. (2012). Oxidative damage is ameliorated by curcumin treatment in brain and sciatic nerve of diabetic rats. Int. J. Neurosci. 122, 367–372. doi: 10.3109/00207454.2012.657380
Aggarwal, B. B., Deb, L., and Prasad, S. (2014). Curcumin differs from tetrahydrocurcumin for molecular targets, signaling pathways and cellular responses. Molecules. 20, 185–205. doi: 10.3390/molecules20010185
Agrawal, R., Mishra, B., Tyagi, E., Nath, C., and Shukla, R. (2010). Effect of curcumin on brain insulin receptors and memory functions in STZ (ICV) induced dementia model of rat. Pharmacol. Res. 61, 247–252. doi: 10.1016/j.phrs.2009.12.008
Ahmad, S., and Beg, Z. H. (2013). Hypolipidemic and antioxidant activities of thymoquinone and limonene in atherogenic suspension fed rats. Food Chem. 138, 1116–1124. doi: 10.1016/j.foodchem.2012.11.109
Al Wafai, R. J. (2013). Nigella sativa and thymoquinone suppress cyclooxygenase-2 and oxidative stress in pancreatic tissue of streptozotocin-induced diabetic rats. Pancreas 42, 841–849. doi: 10.1097/MPA.0b013e318279ac1c
Al-Ali, A., Alkhawajah, A. A., Randhawa, M. A., and Shaikh, N. A. (2008). Oral and intraperitoneal LD50 of thymoquinone, an active principle of Nigella sativa, in mice and rats. J. Ayub. Med. Coll. Abbottabad. 20, 25–27.
Al-Amri, A. M., and Bamosa, A. O. (2009). Phase I safety and clinical activity study of thymoquinone in patients with advanced refractory malignant disease. Med. J. 10, 107–111.
Al-Fartosy, A. J. (2015). Protective effect of galactomannan extracted from Iraqi Lycium Barbarum L. fruits against alloxan-induced diabetes in rats. Am. J. Biochem. Biotechnol. 11, 73–83. doi: 10.3844/ajbbsp.2015.74.83
Al-Habori, M., Raman, A., Lawrence, M. J., and Skett, P. (2001). In vitro effect of fenugreek extracts on intestinal sodium-dependent glucose uptake and hepatic glycogen phosphorylase A. Int. J. Exp. Diabetes Res. 2, 91–99. doi: 10.1155/EDR.2001.91
Alkharfy, K. M., Ahmad, A., Khan, R. M., and Al-Shagha, W. M. (2015). Pharmacokinetic plasma behaviors of intravenous and oral bioavailability of thymoquinone in a rabbit model. Eur. J. Drug Metab. Pharmacokinet. 40, 319–323. doi: 10.1007/s13318-014-0207-8
Al-Khateeb, E., Hamadi, S. A., Al-Hakeemi, A. A., Abu-Taha, M., and Al-Rawi, N. (2012). Hypoglycemic effect of trigonelline isolated from Iraqi fenugreek seeds in normal and alloxan-diabetic rabbits. Eur. Sci. J. 8, 16–24.
Al-Trad, B., Al-Batayneh, K., El-Metwally, S., Alhazimi, A., Ginawi, I., Alaraj, M., et al. (2016). Nigella sativa oil and thymoquinone ameliorate albuminuria and renal extracellular matrix accumulation in the experimental diabetic rats. Eur. Rev. Med. Pharmacol. Sci. 20, 2680–2688.
American Diabetes Association (ADA) (2015). Report of the expert committee on the diagnosis and classification of diabetes mellitus. Diabetes Care 38, S8–S16. doi: 10.2337/dc15-S005
Ames, B. N. (1983). Dietary carcinogens and anticarcinogens: oxygen radials and degenerative diseases. Science 221, 1256–1264. doi: 10.1126/science.6351251
Anand, P., Kunnumakkara, A. B., Newman, R. A., and Aggarwal, B. B. (2007). Bioavailability of curcumin: problems and promises. Mol. Pharm. 4, 807–818. doi: 10.1021/mp700113r
Anand, P., Murali, K. Y., Tandon, V., Murthy, P. S., and Chandra, R. (2010). Insulinotropic effect of cinnamaldehyde on transcriptional regulation of pyruvate kinase, phosphoenolpyruvate carboxykinase, and GLUT4 translocation in experimental diabetic rats. Chem. Biol. Interact. 186, 72–81. doi: 10.1016/j.cbi.2010.03.044
Anwar, S., Desai, S., and Mandlik, R. (2009). Exploring antidiabetic mechanisms of action of galactomannan: a carbohydrate isolated from fenugreek seeds. J. Complement. Integr. Med. 6, 1–11. doi: 10.2202/1553-3840.1218
Anwar, S., Khan, M. A., Sadaf, A., and Younus, H. (2014). A structural study on the protection of glycation of superoxide dismutase by thymoquinone. Int. J. Biol. Macromol. 69, 476–481. doi: 10.1016/j.ijbiomac.2014.06.003
Arafa, H. M. (2005). Curcumin attenuates diet-induced hypercholesterolemia in rats. Med. Sci. Monit. 11, 228–234.
Arcaro, C. A., Gutierres, V. O., Assis, R. P., Moreira, T. F., Costa, P. I., Baviera, A. M., et al. (2014). Piperine, a natural bioenhancer, nullifies the antidiabetic and antioxidant activities of curcumin in streptozotocin-diabetic rats. PLoS ONE 3:e113993. doi: 10.1371/journal.pone.0113993
Arun, N., and Nalini, N. (2002). Efficacy of turmeric on blood sugar and polyol pathway in diabetic albino rats. Plant Food Hum. Nutr. 57, 41–52. doi: 10.1023/A:1013106527829
Ashour, T. H. (2015). Thymoquinone therapy improves hyperglycemia, erythrocyte indices, erythropoietin production and erythrocyte osmotic resistance in rat model of streptozotocin-induced diabetes. Br. J. Med. Med. Res. 5, 350–361. doi: 10.9734/BJMMR/2015/13409
Aswar, U., Mohan, V., and Bodhankar, S. L. (2009). Effect of trigonelline on fertility in female rats. Int. J. Green Pharm. 3, 220–223. doi: 10.22377/ijgp.v3i3.87
Atal, S., Atal, S., Vyas, S., and Phadnis, P. (2016). Bio-enhancing effect of piperine with metformin on lowering blood glucose level in alloxan induced diabetic mice. Phcog. Res. 8, 56–60. doi: 10.4103/0974-8490.171096
Augusti, K. T., and Sheela, C. G. (1996). Antiperoxide effect of S-allyl cysteine sulfoxide, an insulin secretagogue, in diabetic rats. Cell Mol. Life Sci. 15, 115–120. doi: 10.1007/BF01923354
Avalos-Soriano, A., De la Cruz-Cordero, R., Rosado, J. L., and Garcia-Gasca, T. (2016). 4-Hydroxyisoleucine from fenugreek (Trigonella foenum-graecum): effects on insulin resistance associated with obesity. Molecules 21, 1596–1596. doi: 10.3390/molecules21111596
Awasthi, H., Tota, S., Hanif, K., Nath, C., and Shukla, R. (2010). Protective effect of curcumin against intracerebral streptozotocin induced impairment in memory and cerebral blood flow. Life Sci. 86, 87–94. doi: 10.1016/j.lfs.2009.11.007
Babu, P. S., and Srinivasan, K. (1997a). Influence of dietary capsaicin and onion on the metabolic abnormalities associated with streptozotocin induced diabetes mellitus. Mol. Cell Biochem. 175, 49–57.
Babu, P. S., and Srinivasan, K. (1997b). Hypolipidemic action of curcumin, coloring principle of turmeric (Curcuma longa) in streptozotocin induced diabetic rats. Mol. Cell Biochem. 166, 169–175.
Badary, O. A., Al-Shabanah, O. A., Nagi, M. N., Al-Bekairi, A. M., and Elmazar, M. (1998). Acute and subchronic toxicity of thymoquinone in mice. Drug Devel. Res. 44, 56–61.
Badr, G., Alwasel, S., Ebaid, S., Mohany, M., and Alhazza, I. (2011). Perinatal supplementation with thymoquinone improves diabetic complications and T cell immune responses in rat offspring. Cell Immunol. 267, 133–140. doi: 10.1016/j.cellimm.2011.01.002
Badr, G., Mahmoud, M. H., Farhat, K., Waly, H., Al-Abdin, O. Z., and Rabah, D. M. (2013). Maternal supplementation of diabetic mice with thymoquinone protects their offspring from abnormal obesity and diabetes by modulating their lipid profile and free radical production and restoring lymphocyte proliferation via PI3K/AKT signaling. Lipids Health Dis. 12:37. doi: 10.1186/1476-511X-12-37
Balaji, S., and Chempakam, B. (2010). Toxicity prediction of compounds from turmeric (Curcuma longa L). Food Chem. Toxicol. 48, 2951–2959. doi: 10.1016/j.fct.2010.07.032
Banafshe, H. R., Hamidi, G. A., Noureddini, M., Mirhashemi, S. M., Mokhtari, R., and Shoferpour, M. (2014). Effect of curcumin on diabetic peripheral neuropathic pain: possible involvement of opioid system. Eur. J. Pharmacol. 723, 202–206. doi: 10.1016/j.ejphar.2013.11.033
Bansal, S. S., Kausar, H., Aqil, F., Jeyabalan, J., Vadhanam, M. V., Gupta, R. C., et al. (2011). Curcumin implants for continuous systemic delivery: safety and biocompatibility. Drug Deliv. Transl. Res. 1, 332–341. doi: 10.1007/s13346-011-0028-0
Bao, L., Bai, S., and Borijihan, G. (2012). Hypolipidemic effects of a new piperine derivative GB-N from Piper longum in high-fat diet-fed rats. Pharm. Biol. 50, 962–967. doi: 10.3109/13880209.2012.654395
Bashandy, S. A. E., AbdelJaleel, J. A., Abdallah, H. M. I., and Harraz, S. E. S. (2015). Therapeutic implications of thymoquinone in the management of diabetes mellitus and its complications. Am. J. Phytomed. Clin. Ther. 3, 287–301.
Berkoz, M., Yildirim, M., Arvas, G., Turkmen, O., and Allahyerdiyev, O. (2015). Effect of capsaicin on transcription factors in 3T3-L1 cell line. Eastern J Med. 20, 34–45.
Best, L., Elliott, A. C., and Brown, P. D. (2007). Curcumin induces electrical activity in rat pancreatic β-cells by activating the volume-regulated anion channel. Biochem. Pharmacol. 73, 1768–1775. doi: 10.1016/j.bcp.2007.02.006
Bhat, B. G., and Chandrasekhara, N. (1986). Studies on the metabolism of piperine: absorption, tissue distribution and excretion of urinary conjugates in rats. Toxicology 40, 83–92. doi: 10.1016/0300-483X(86)90048-X
Bi, X., Lim, J., and Henry, C. J. (2017). Spices in the management of diabetes mellitus. Food Chem. 217, 281–293. doi: 10.1016/j.foodchem.2016.08.111
Biesbroeck, R., Bril, V., Hollander, P., Kabadi, U., Schwartz, S., Singh, S. P., et al. (1994). A double-blind comparison of topical capsaicin and oral amitriptyline in painful diabetic neuropathy. Adv. Ther. 12, 111–120.
Boddupalli, B. M., Ramani, R., Anisetti, R. N., and Chandragiri, V. L. (2015). Prevention of pioglitazone induced weight gain by co-administration of piperine. Br. J. Pharm. Res. 7, 276–281. doi: 10.9734/BJPR/2015/18416
BrahmaNaidu, P., Nemani, H., Meriga, B., Mehar, S. K., Potana, S., and Ramgopalrao, S. (2014). Mitigating efficacy of piperine in the physiological derangements of high fat diet induced obesity in Sprague Dawley rats. Chem. Biol. Interact. 221, 42–51. doi: 10.1016/j.cbi.2014.07.008
Broca, C., Breil, V., Cruciani-Guglielmacci, C., Manteghetti, M., Rouault, C., and Derouet, M. (2004). Insulinotropic agent ID-1101 (4-hydroxyisoleucine) activates insulin signaling in rat. Am. J. Physiol. Endocrinol. Metab. 287, E463–E471. doi: 10.1152/ajpendo.00163.2003
Broca, C., Gross, R., Petit, P., Sauvaire, Y., Manteghetti, M., and Tournier, M. (1999). 4-Hydroxyisoleucine: experimental evidence of its insulinotropic and antidiabetic properties. Am. J. Physiol. Endocrinol. Metab. 277, E617–E623. doi: 10.1152/ajpendo.1999.277.4.E617
Broca, C., Manteghetti, M., Gross, R., Baissac, Y., Jacob, M., and Petit, P. (2000). 4-Hydroxyisoleucine: effects of synthetic and natural analogues on insulin secretion. Eur. J. Pharmacol. 390, 339–345. doi: 10.1016/S0014-2999(00)00030-3
Bustanji, Y., Taha, M. O., Almasri, I. M., Al-Ghussein, M. A., Mohammad, M. K., and Alkhatib, H. S. (2009). Inhibition of glycogen synthase kinase by curcumin: investigation by simulated molecular docking and subsequent in vitro/in vivo evaluation. J. Enzyme Inhib. Med. Chem. 24, 771–778. doi: 10.1080/14756360802364377
Camacho, S., Michlig, S., de Senarclens-Bezençon, C., Meylan, J., Meystre, J., Pezzoli, M., et al. (2015). Anti-obesity and anti-hyperglycemic effects of cinnamaldehyde via altered ghrelin secretion and functional impact on food intake and gastric emptying. Sci. Rep. 5:7919. doi: 10.1038/srep07919
Cayen, M. N., Ferdinandi, E. S., Greselin, E., and Dvornik, D. (1979). Studies on the disposition of diosgenin in rats, dogs, monkeys and man. Atherosclerosis 33, 71–87. doi: 10.1016/0021-9150(79)90199-0
Chainani-Wu, N. (2003). Safety and anti-inflammatory activity of curcumin: a component of tumeric (Curcuma longa). J. Altern. Complement. Med. 9, 161–168. doi: 10.1089/107555303321223035
Chakraborty, D., Mukherjee, A., Sikdar, S., Paul, A., Ghosh, S., and Khuda-Bukhsh, A. R. (2012). [6]-Gingerol isolated from ginger attenuates sodium arsenite induced oxidative stress and plays a corrective role in improving insulin signaling in mice. Toxicol. Lett. 210, 34–43. doi: 10.1016/j.toxlet.2012.01.002
Chandra, S., Mondal, D., and Agrawal, K. C. (2009). HIV-1 protease inhibitor induced oxidative stress suppresses glucose stimulated insulin release: protection with thymoquinone. Exp. Biol. Med. 234, 442–453. doi: 10.3181/0811-RM-317
Chandran, B., and Goel, A. (2012). A randomized, pilot study to assess the efficacy and safety of curcumin in patients with active rheumatoid arthritis. Phytother. Res. 26, 1719–1725. doi: 10.1002/ptr.4639
Chanpoo, M., Petchpiboonthai, H., Panyarachu, B., and Anupunpisit, V. (2010). Effect of curcumin in the amelioration of pancreatic islets in streptozotocin-induced diabetic mice. J. Med. Assoc. Thai. 93, S152–S159.
Chao, L. K., Chang, W. T., Shih, Y. W., and Huang, J. S. (2010). Cinnamaldehyde impairs high glucose-induced hypertrophy in renal interstitial fibroblasts. Toxicol. Appl. Pharmacol. 244, 174–180. doi: 10.1016/j.taap.2009.12.030
Chen, M. M. Y. (1995). Inhibition of tumor necrosis factor by curcumin, a phytochemical. Biochem. Pharmacol. 49, 1551–1556. doi: 10.1016/0006-2952(95)00171-U
Chen, Y., Yueming, M., and Wei, M. (2009). Pharmacokinetics and bioavailability of cinnamic acid after oral administration of Ramulus Cinnamomi in rats. Eur. J. Drug Met. Pharm. 34, 51–56. doi: 10.1007/BF03191384
Cheng, T. C., Lin, C. S., Hsu, C. C., Chen, L. J., Cheng, K. C., and Cheng, J. T. (2009). Activation of muscarinic M-1 cholinoceptors by curcumin to increase glucose uptake into skeletal muscle isolated from Wistar rats. Neurosci. Lett. 465, 238–241. doi: 10.1016/j.neulet.2009.09.012
Chiu, J., Khan, Z. A., Farhangkhoee, H., and Chakrabarti, S. (2009). Curcumin prevents diabetes-associated abnormalities in the kidneys by inhibiting p300 and nuclear factor-κB. Nutrition 25, 964–972. doi: 10.1016/j.nut.2008.12.007
Choi, J., Kim, K. J., Kim, B. H., Koh, E. J., Seo, M. J., and Lee, B. Y. (2016). 6-Gingerol suppresses adipocyte-derived mediators of inflammation in vitro and in high-fat diet-induced obese zebra fish. Planta Med. 83, 245–253. doi: 10.1055/s-0042-112371
Chougala, M. B., Bhaskar, J. J., Rajan, M. G., and Salimath, P. V. (2012). Effect of curcumin and quercetin on lysosomal enzyme activities in streptozotocin-induced diabetic rats. Clin. Nutr. 31, 749–755. doi: 10.1016/j.clnu.2012.02.003
Chuengsamarn, S., Rattanamongkolgul, S., Luechapudiporn, R., Phisalaphong, C., and Jirawatnotai, S. (2012). Curcumin extract for prevention of type 2 diabetes. Diabetes Care 35, 2121–2127. doi: 10.2337/dc12-0116
Chuengsamarn, S., Rattanamongkolgul, S., Phonrat, B., Tungtrongchitr, R., and Jirawatnotai, S. (2014). Reduction of atherogenic risk in patients with type 2 diabetes by curcuminoid extract: a randomized controlled trial. J. Nutr. Biochem. 25, 144–150. doi: 10.1016/j.jnutbio.2013.09.013
Cui, J., Yu, B., Zhao, Y., Zhu, W., Li, H., Lou, H., et al. (2009). Enhancement of oral absorption of curcumin by self-microemulsifying drug delivery systems. Int. J. Pharm. 371, 148–155. doi: 10.1016/j.ijpharm.2008.12.009
Desai, S. D., Shaik, H. S., Kusal, K. D., and Haseena, S. (2015). Effect of thymoquinone on MDA and SOD levels in sterptozotocine induced diabetic albino rats. Pharm. Sci. Res. 7, 523–526.
Deshpande, P., Mohan, V., and Thakurdesai, P. (2016a). Preclinical safety evaluation of low molecular weight galactomannans based standardized fenugreek seeds extract. EXCLI J. 15, 446–459. doi: 10.17179/excli2016-461
Deshpande, P., Mohan, V., Thakurdesai, P., Pore, M., and Gumaste, S. (2016b). Prenatal developmental toxicity evaluation of low molecular weight galactomannans based standardized fenugreek seed extract during organogenesis period of pregnancy in rats. Int. J. Pharm. Pharm. Sci. 8, 248–253.
Effenberger-Neidnicht, K., Breyer, S., Mahal, K., Diestel, R., Sasse, F., and Schobert, R. (2011). Cellular localisation of antitumoral 6-alkyl thymoquinones revealed by an alkyne-azide click reaction and the streptavidin–biotin system. Chem. Biol. Chem. 12, 1237–1241. doi: 10.1002/cbic.201000762
El-Ameen, N. M. H., Elhassan Taha, M. M., Abdelwahab, S. I., Khalid, A., Elfatih, F., Kamel, M. A., et al. (2015). Anti-diabetic properties of thymoquinone is un associated with glycogen phosphorylase inhibition. Pharmacog J. 7, 406–410. doi: 10.5530/pj.2015.6.16
El-Azab, M. F., Attia, F. M., and El-Mowafy, A. M. (2011). Novel role of curcumin combined with bone marrow transplantation in reversing experimental diabetes: effects on pancreatic islet regeneration, oxidative stress, and inflammatory cytokines. Eur. J. Pharmacol. 658, 41–48. doi: 10.1016/j.ejphar.2011.02.010
El-Bahr, S. M. (2013). Curcumin regulates gene expression of insulin like growth factor, β-cell CLL/lymphoma 2 and antioxidant enzymes in streptozotocin induced diabetic rats. BMC Complement. Altern. Med. 13:368. doi: 10.1186/1472-6882-13-368
El-Bassossy, H. M., Elberry, A. A., Ghareib, S. A., Azhar, A., Banjar, Z. M., and Watson, M. L. (2016). Cardioprotection by 6-gingerol in diabetic rats. Biochem. Biophys. Res. Commun. 477, 908–914. doi: 10.1016/j.bbrc.2016.06.157
El-Bassossy, H. M., Fahmy, A., and Badawy, D. (2011). Cinnamaldehyde protects from the hypertension associated with diabetes. Food Chem. Toxicol. 49, 3007–3012. doi: 10.1016/j.fct.2011.07.060
El-Dakhakhny, M. (1965). Studies on the Egyptian Nigella sativa L. IV. Some pharmacological properties of the seeds' active principle in comparison to its dihydro compound and its polymer. Arzneimittelforschung 15, 1227–1229.
El-Mahmoudy, A., Shimizu, Y., Shiina, T., Matsuyama, H., Nikami, H., and Takewaki, T. (2005a). Macrophage-derived cytokine and nitric oxide profiles in type I and type II diabetes mellitus: effect of thymoquinone. Acta Diabetol. 42, 23–30. doi: 10.1007/s00592-005-0170-6
El-Mahmoudy, A., Shimizu, Y., Shiina, T., Matsuyama, H., Nikami, H., and Takewaki, T. (2005b). Successful abrogation by thymoquinone against induction of diabetes mellitus with streptozotocin via nitric oxide inhibitory mechanism. Int. Immunopharmacol. 5, 195–207. doi: 10.1016/j.intimp.2004.09.001
Elmansy, R. A., and Almasry, S. M. (2013). Morphological and immunohistochemical analysis of the effects of thymoquinone on the neurovascular component of Jejunal submucosa of diabetic rat model. J. Am. Sci. 9, 224–236.
El-Moselhy, M. A., Taye, A., Sharkawi, S. S., El-Sisi, S. F. I., and Ahmed, A. F. (2011). The antihyperglycemic effect of curcumin in high fat diet fed rats. Role of TNF-α and free fatty acids. Food Chem. Toxicol. 49, 1129–1140. doi: 10.1016/j.fct.2011.02.004
Escobar-García, M., Rodríguez-Contreras, K., Ruiz-Rodríguez, S., Pierdant-Pérez, M., Cerda-Cristerna, B., and Pozos-Guillén, A. (2016). Eugenol toxicity in human dental pulp fibroblasts of primary teeth. J. Clin. Pediatr. Dent. 40, 312–318. doi: 10.17796/1053-4628-40.4.312
Fang, K., Dong, H., Jiang, H., Li, F., Wang, D., Yang, D., et al. (2016). Diosgenin and 5-methoxypsoralen ameliorate insulin resistance through ER-α/PI3K/Akt-signaling pathways in HepG2 Cells. Evid. Based Complement. Alternat. Med. 2016, 1–11. doi: 10.1155/2016/7493694
Fararh, K. M., Ibrahim, A. K., and Elsonosy, Y. A. (2010). Thymoquinone enhances the activities of enzymes related to energy metabolism in peripheral leukocytes of diabetic rats. Res. Vet. Sci. 88, 400–404. doi: 10.1016/j.rvsc.2009.10.008
Fararh, K. M., Shimizu, Y., Shiina, T., Nikami, H., Ghanem, M. M., and Takewaki, T. (2005). Thymoquinone reduces hepatic glucose production in diabetic hamsters. Res. Vet. Sci. 79, 219–223. doi: 10.1016/j.rvsc.2005.01.001
Farrokhfall, K., Khoshbaten, A., Zahediasl, S., Mehrani, H., and Karbalaei, N. (2014). Improved islet function is associated with antiinflammatory, antioxidant and hypoglycemic potential of cinnamaldehyde on metabolic syndrome induced by high tail fat in rats. J. Funct. Foods. 10, 397–406. doi: 10.1016/j.jff.2014.07.014
Folwarczna, J., Janas, A., Pytlik, M., Cegieła, U., Sliwinski, L., Krivošíková, Z., et al. (2016). Effects of trigonelline, an alkaloid present in coffee, on diabetes-induced disorders in the rat skeletal system. Nutrients 8:133. doi: 10.3390/nu8030133
Forst, T., Pohlmann, T., Kunt, T., Goitom, K., Schulz, G., Löbig, M., et al. (2002). The influence of local capsaicin treatment on small nerve fibre function and neurovascular control in symptomatic diabetic neuropathy. Acta Diabetol. 39, 1–6. doi: 10.1007/s005920200005
Fouad, A. A., and Alwadani, F. (2015). Ameliorative effects of thymoquinone against eye lens changes in streptozotocin diabetic rats. Environ. Toxicol. Pharmacol. 40, 960–965. doi: 10.1016/j.etap.2015.09.010
Fujiwara, H., Hosokawa, M., Zhou, X., Fujimoto, S., Fukuda, K., Toyoda, K., et al. (2008). Curcumin inhibits glucose production in isolated mice hepatocytes. Diabetes Res. Clin. Pract. 80, 185–191. doi: 10.1016/j.diabres.2007.12.004
Gao, S., Basu, S., Yang, Z., Deb, A., and Hu, M. (2012). Bioavailability challenges associated with development of saponins as therapeutic and chemopreventive agents. Curr. Drug Targets 13, 1885–1899. doi: 10.2174/138945012804545498
Garcea, G., Jones, D. J., Singh, R., Dennison, A. R., Farmer, P. B., Sharma, R. A., et al. (2004). Detection of curcumin and its metabolites in hepatic tissue and portal blood of patients following oral administration. Br. J. Cancer 90, 1011–1015. doi: 10.1038/sj.bjc.6601623
Ghosh, S., Bhattacharyya, S., Rashid, K., and Sil, P. C. (2015). Curcumin protects rat liver from streptozotocin-induced diabetic pathophysiology by counteracting reactive oxygen species and inhibiting the activation of p53 and MAPKs mediated stress response pathways. Toxicol. Rep. 2, 365–376. doi: 10.1016/j.toxrep.2014.12.017
Ghosh, S., More, P., Derle, A., Patil, A. B., Markad, P., Asok, A., et al. (2014). Diosgenin from Dioscorea bulbifera: novel hit for treatment of type II diabetes mellitus with inhibitory activity against α-amylase and α-glucosidase. PLoS ONE 9:e106039. doi: 10.1371/journal.pone.0106039
Ghule, A. E., Jadhav, S. S., and Bodhankar, S. L. (2012). Trigonelline ameliorates diabetic hypertensive nephropathy by suppression of oxidative stress in kidney and reduction in renal cell apoptosis and fibrosis in streptozotocin induced neonatal diabetic (nSTZ) rats. Int. Immunopharmacol. 14, 740–748. doi: 10.1016/j.intimp.2012.10.004
Glinsukon, T., Stitmunnaithum, V., Toskulkao, C., Buranawuti, T., and Tangkrisanavinont, V. (1980). Acute toxicity of capsaicin in several animal species. Toxicon 18, 215–220. doi: 10.1016/0041-0101(80)90076-8
Golshahi, J., and Roghani-Dehkordi, F. (2016). Diosgenin attenuates cardiac oxidative stress in streptozotocin-induced diabetic rat. J. Basic Clin. Pathophysiol. 4, 13–18. doi: 10.22070/jbcp.2016.279
Gram, D. X., Ahrèn, B., Nagy, I., Olsen, U. V., Brand, C. L., Sundler, F., et al. (2007). Capsaicin-sensitive sensory fibers in the islets of Langerhans contribute to defective insulin secretion in Zucker diabetic rat, an animal model for some aspects of human type 2 diabetes. Eur. J. Neurosci. 25, 213–223. doi: 10.1111/j.1460-9568.2006.05261.x
Grama, C. N., Suryanarayana, P., Patil, M. A., Raghu, G., Balakrishna, N., Kumar, M. N., et al. (2013). Efficacy of biodegradable curcumin nanoparticles in delaying cataract in diabetic rat model. PLoS ONE 8:e78217 doi: 10.1371/journal.pone.0078217
Gray, J. P., Zayasbazan, B. D., Yuan, T., Seeram, N., Rebar, R., Follmer, R., et al. (2016). Thymoquinone, a bioactive component of Nigella sativa, normalizes insulin secretion from pancreatic β-cells under glucose overload via regulation of malonyl-CoA. Am. J. Physiol. Endocrinol. Metab. 310, E394–E404. doi: 10.1152/ajpendo.00250.2015
Green, A., Krause, J., and Rumberger, J. M. (2014). Curcumin is a direct inhibitor of glucose transport in adipocytes. Phytomedicine 21, 118–122. doi: 10.1016/j.phymed.2013.08.014
Guo, C., Li, M., Qi, X., Lin, G., Cui, F., Li, F., et al. (2016). Intranasal delivery of nanomicelle curcumin promotes corneal epithelial wound healing in streptozotocin-induced diabetic mice. Sci. Rep. 6:29753. doi: 10.1038/srep29753
Guo, X., Sun, W., Huang, L., Wu, L., Hou, Y., Qin, L., et al. (2017). Effect of cinnamaldehyde on glucose metabolism and vessel function. Med. Sci. Monit. 23, 3844–3853. doi: 10.12659/MSM.906027
Gupta, S. K., Kumar, B., and Nag, T. C. (2011). Curcumin prevents experimental diabetic retinopathy in rats through its hypoglycemic, antioxidant, and anti-inflammatory mechanisms. J. Ocular Pharmacol. Ther. 27, 123–130. doi: 10.1089/jop.2010.0123
Gutierres, V. O., Pinheiro, C. M, Assis, R. P., Vendramini, R. C., Pepato, M. T., and Brunetti, I. L. (2012). Curcumin-supplemented yoghurt improves physiological and biochemical markers of experimental diabetes. Br. J. Nutr. 108, 440–448. doi: 10.1017/S0007114511005769
Haeri, M. R., Izaddoost, M., Ardekani, M. R. S., Nobar, M. R., and White, K. N. (2009). The effect of fenugreek 4-hydroxyisoleucineon liver function biomarkers and glucose in diabetic and fructose-fed rats. Phytother. Res. 23, 61–64. doi: 10.1002/ptr.2557
Haeri, M. R., Limaki, H. K., White, C. J., and White, K. N. (2012). Non-insulin dependent anti-diabetic activity of (2S, 3R, 4S) 4-hydroxyisoleucineof fenugreek (Trigonella foenum graecum) in streptozotocin-induced type I diabetic rats. Phytomedicine. 19, 571–574. doi: 10.1016/j.phymed.2012.01.004
Hafez, D. A. (2013). Effects of Nigella sativa oil and thymoquinone on renal oxidative stress and apoptosis rate instreptozotocin-diabetic rats. J. Am. Sci. 9, 327–333.
Hafizur, R. M., Hameed, A., Shukrana, M., Raza, S. A., Chishti, S., Kabir, N., et al. (2015). Cinnamic acid exerts anti-diabetic activity by improving glucose tolerance in vivo and by stimulating insulin secretion in vitro. Phytomedicine. 22, 297–300. doi: 10.1016/j.phymed.2015.01.003
Hagan, E. C., Jenner, P. M., Jones, W. I., Fitzhugh, O. G., Long, E. L., Brouwer, J. G., et al. (1965). Toxic properties of compounds related to safrole. Toxicol. Appl. Pharmacol. 7, 18–24. doi: 10.1016/0041-008X(65)90069-4
Hamadi, S. A. (2012). Effect of trigonelline and ethanol extract of Iraqi Fenugreek seeds on oxidative stress in alloxan diabetic rabbits. J. Assoc. Arab. Univ. Basic Appl. Sci. 12, 23–26. doi: 10.1016/j.jaubas.2012.02.003
Hamden, K., Bengara, A., Amri, Z., and Elfeki, A. (2013a). Experimental diabetes treated with trigonelline: effect on key enzymes related to diabetes and hypertension, β-cell and liver function. Mol. Cell Biochem. 381, 85–94. doi: 10.1007/s11010-013-1690-y
Hamden, K., Jaouadi, B., Carreau, S., Bejar, S., and Elfeki, A. (2010). Inhibitory effect of fenugreek galactomannan on digestive enzymes related to diabetes, hyperlipidemia, and liver-kidney dysfunctions. Biotechnol. Bioprocess. Eng. 15, 407–413. doi: 10.1007/s12257-009-3037-9
Hamden, K., Mnafgui, K., Amri, Z., Aloulou, A., and Elfeki, A. (2013b). Inhibition of key digestive enzymes related to diabetes and hyperlipidemia and protection of liver-kidney functions by trigonelline in diabetic rats. Sci. Pharm. 81, 233–246. doi: 10.3797/scipharm.1211-14
Hamdy, N. M., and Taha, R. A. (2009). Effects of Nigella sativa oil and thymoquinone on oxidative stress and neuropathy in streptozotocin-induced diabetic rats. Pharmacology. 84, 127–134. doi: 10.1159/000234466
Hao, S., Xu, R., Li, D., Zhu, Z., Wang, T., and Liu, K. (2015). Attenuation of streptozotocin-induced lipid profile anomalies in the heart, brain, and mRNA expression of HMG-CoA reductase by diosgenin in rats. Cell Biochem. Biophys. 72, 741–749. doi: 10.1007/s12013-015-0525-8
Hawsawi, Z. A., Ali, B. A., and Bamosa, A. O. (2001). Effect of Nigella sativa (Black seed) and thymoquinone on blood glucose in albino rats. Ann. Saudi Med. 21, 242–244. doi: 10.5144/0256-4947.2001.242
Hie, M., Yamazaki, M., and Tsukamoto, I. (2009). Curcumin suppresses increased bone resorption by inhibiting osteoclastogenesis in rats with streptozotocin-induced diabetes. Eur. J. Pharmacol. 621, 1–9. doi: 10.1016/j.ejphar.2009.08.025
Honda, S., Aoki, F., Tanaka, H., Kishida, H., Nishiyama, T., Okada, S., et al. (2006). Effects of ingested turmeric oleoresin on glucose and lipid metabolisms in obese diabetic mice: a DNA microarray study. J. Agric. Food Chem. 54, 9055–9062. doi: 10.1021/jf061788t
Hooth, M. J., Sills, R. C., Burka, L. T., Haseman, J. K., Witt, K. L., Orzech, D. P., et al. (2004). Toxicology and carcinogenesis studies of microencapsulated trans-cinnamaldehyde in rats and mice. Food Chem. Toxicol. 42, 1757–1768. doi: 10.1016/j.fct.2004.07.002
Hosni, A. A., Abdel-Moneim, A. A., Abdel-Reheim, E. S., Mohamed, S. M., and Helmy, H. (2017). Cinnamaldehyde potentially attenuates gestational hyperglycemia in rats through modulation of PPARγ, proinflammatory cytokines and oxidative stress. Biomed. Pharmacother. 88, 52–60. doi: 10.1016/j.biopha.2017.01.054
Hsia, S. M., Lee, W. H., Yen, G. C., and Wu, C. H. (2016). Capsaicin, an active ingredient from chilli peppers, attenuates glycative stress and restores RAGE levels in diabetic rats. J. Funct. Foods. 21, 406–417. doi: 10.1016/j.jff.2015.11.043
Hsu, C. C., Yen, H. F., Yin, M. C., Tsai, C. M., and Hsieh, C. H. (2004). Five cysteine-containing compounds delay diabetic deterioration in Balb/cA mice. J. Nutr. 134, 3245–3249. doi: 10.1093/jn/134.12.3245
Hsu, C. H., and Cheng, A. L. (2007). Clinical studies with curcumin. Adv. Exp. Med. Biol. 595, 471–480. doi: 10.1007/978-0-387-46401-5_21
Huang, B., Yuan, H. D., Kim, D. Y., Quan, H. Y., and Chung, S. H. (2011). Cinnamaldehyde prevents adipocyte differentiation and adipogenesis via regulation of peroxisome proliferator-activated receptor-γ (PPARγ) and AMP-activated protein kinase (AMPK) pathways. J. Agric. Food Chem. 59, 3666–3673. doi: 10.1021/jf104814t.
Huang, J., Huang, K., Lan, T., Xie, X., Shen, X., Liu, P., et al. (2013). Curcumin ameliorates diabetic nephropathy by inhibiting the activationof the SphK1-S1P signaling pathway. Mol. Cell Endocrinol. 365, 231–240. doi: 10.1016/j.mce.2012.10.024
Huang, Q., Yu, H., and Ru, Q. (2010). Bioavailability and delivery of nutraceuticals using nanotechnology. J. Food Sci. 75, 50–57. doi: 10.1111/j.1750-3841.2009.01457.x
Hung, H. Y., Qian, K., Morris-Natschke, S. L., Hsu, C. S., and Lee, K. S. (2012). Recent discovery of plant-derived anti-diabetic natural products. Nat. Prod. Rep. 29, 580–606. doi: 10.1039/c2np00074a
Hussein, H. K., and Abu-Zinadah, O. A. (2010). Antioxidant effect of curcumin extracts in induced diabetic wister rats. Int. J. Zool. Res. 6, 266–276. doi: 10.3923/ijzr.2010.266.276
Hussein, M. A., and El-Maksoud, H. A. (2013). Biochemical effects of Resveratrol and Curcumin combination on obese diabetic rats. Mol. Clin. Pharmacol. 4, 1–10.
Hwang, J. T., Park, I. J., Shin, J. I., Lee, Y. K., Lee, S. K., Baik, H. W., et al. (2005). Genistein, EGCG, and capsaicin inhibit adipocyte differentiation process via activating AMP-activated protein kinase. Biochem. Biophys. Res. Comm. 338, 694–699. doi: 10.1016/j.bbrc.2005.09.195
Hwang, S. L., Yang, B. K., Lee, J. Y., Kim, J. H., Kim, B. H., Suh, K. H., et al. (2008). Isodihydrocapsiate stimulates plasma glucose uptake by activation of AMP-activated protein kinase. Biochem. Biophys. Res. Comm. 371, 289–293. doi: 10.1016/j.bbrc.2008.04.061
Ilavenil, S., Arasu, M. V., Lee, J. C., Kim, D. H., Roh, S. G., Park, H. S., et al. (2014). Trigonelline attenuates the adipocyte differentiation and lipid accumulation in 3T3-L1 cells. Phytomedicine 21, 758–765. doi: 10.1016/j.phymed.2013.11.007
International Diabetes Federation (IDF) (2017). Diabetes Atlas, 8th Edn. Available online at http://www.diabetesatlas.org/ (Accessed on 14th February, 2018).
Isa, Y., Miyakawa, Y., Yanagisawa, M., Goto, T., Kang, M. S., Kawada, T., et al. (2008). 6-Shogaol and 6-gingerol, the pungent of ginger, inhibit TNF-α mediated downregulation of adiponectin expression via different mechanisms in 3T3-L1 adipocytes. Biochem. Biophys. Res. Comm. 373, 429–434. doi: 10.1016/j.bbrc.2008.06.046
Jain, S. K., Rains, J., Croad, J., Larson, B., and Jones, K. (2009). Curcumin supplementation lowers TNF-α, IL-6, IL-8, and MCP-1 secretion in high glucose-treated culturedmonocytes and blood levels of TNF-α, IL-6, MCP-1, glucose, and glycosylated hemoglobin in diabetic rats. Antioxid. Redox. Signal. 11, 241–249. doi: 10.1089/ars.2008.2140
Jain, S. K., Rains, J., and Jones, K. (2006). Effect of curcumin on protein glycosylation, lipid peroxidation, and oxygen radical generation in human red blood cells exposed to high glucose levels. Free. Radic. Biol. Med. 41, 92–96. doi: 10.1016/j.freeradbiomed.2006.03.008
Jaiswal, N., Maurya, C. K., Venkateswarlu, K., Sukanya, P., Srivastava, A. K., Narender, T., et al. (2012). 4-Hydroxyisoleucine stimulates glucose uptake by increasing surface GLUT4 level in skeletal muscle cells via phosphatidylinositol-3-kinase-dependent pathway. Eur. J. Nutr. 51, 893–898. doi: 10.1007/s00394-012-0374-9
Jang, E. M., Choi, M. S., Jung, U. J., Kim, M. J., Kim, H. J., Jeon, S. M., et al. (2008). Beneficial effects of curcumin on hyperlipidemia and insulin resistance in high-fat-fed hamsters. Metab. Clin. Exp. 57, 1576–1583. doi: 10.1016/j.metabol.2008.06.014
Jawale, A., Datusalia, A. K., Bishno, M., and Sharma, S. S. (2016). Reversal of diabetes-induced behavioral and neurochemical deficits by cinnamaldehyde. Phytomedicine 23, 923–930. doi: 10.1016/j.phymed.2016.04.008
Jeng, J. H., Hahn, L. J., Lu, E. J., Wang, Y. J., and Kuo, M. Y. P. (1994). Eugenol triggers different pathobiological effects on human oral mucosal fibroblasts 1. J. Dental. Res. 73, 1050–1055. doi: 10.1177/00220345940730050601
Jenner, P. M., Hagan, E. C., Taylor, J. M., Cook, E. L., and Fitzhugh, O. G. (1964). Food flavorings and compounds of related structure I. Acute oral toxicity. Food Cosmet. Toxicol. 2, 327–343. doi: 10.1016/S0015-6264(64)80192-9
Jeong, K. J., Kim, D. Y., Quan, H. Y., Jo, H. K., Kim, G. W., and Chung, S. H. (2014). Effects of eugenol on hepatic glucose production and AMPK signaling pathway in hepatocytes and C57BL/6J mice. Fitoterapia 93, 150–162. doi: 10.1016/j.fitote.2013.12.023
Jo, Y. J., Chun, J. Y., Kwon, Y. J., Min, S. G., Hong, G. P., and Choi, M. J. (2015). Physical and antimicrobial properties of trans-cinnamaldehyde nanoemulsions in water melon juice. LWT-Food Sci. Technol. 60, 444–451. doi: 10.1016/j.lwt.2014.09.041
Jothi, M. A., Parameswari, C. S., and Vincent, S. (2016a). Antidiabetic, hypolipidemic, and histopathological analysis of zingerone instreptozotocin-induced diabetic rats. Asian J. Pharm. Clin. Res. 9, 220–224. doi: 10.13040/IJPSR.0975-8232.7(6).2385-93
Jothi, M. A., Parameswari, C. S., and Vincent, S. (2016b). Hematological studies on the effect of zingerone on streptozotocininduced diabetic rats. Int. J. Pharm. Res. Bio. Sci. 5, 21–26.
Kalailingam, P., Kannaian, B., Tamilmani, E., and Kaliaperumal, R. (2014). Efficacy ofnatural diosgenin on cardiovascular risk, insulin secretion, and beta cells instreptozotocin (STZ)-induced diabetic rats. Phytomedicine 21, 1154–1161. doi: 10.1016/j.phymed.2014.04.005
Kalam, M. A., Raish, M., Ahmed, A., Alkharfy, K. M., Mohsin, K., Alshamsan, A., et al. (2017). Oral bioavailability enhancement and hepatoprotective effects of thymoquinone by self-nanoemulsifying drug delivery system. Mater. Sci. Eng. C. 76, 319–329. doi: 10.1016/j.msec.2017.03.088
Kamble, H., Kandhare, A. D., Bodhankar, S., Mohan, V., and Thakurdesai, P. (2013). Effect of low molecular weight galactomannans from fenugreek seeds on animal models of diabetes mellitus. Biomed. Aging Pathol. 3, 145–151. doi: 10.1016/j.biomag.2013.06.002
Kamble, H. V., and Bodhankar, S. L. (2013). Antihyperglycemic activity of trigonelline and sitagliptin in nicotinamide-streptozotocin induced diabetes in Wistar rats. Biomed. Aging Pathol. 3, 125–130. doi: 10.1016/j.biomag.2013.05.006
Kamble, H. V., and Bodhankar, S. L. (2013b). Trigonelline and sitagliptin attenuates nicotinamide-streptozotocin induced diabetic nephropathy in Wistar rats. Int. J. Pharm. Pharm. Sci. 5, 583–589.
Kamble, H. V., and Bodhankar, S. L. (2014). Cardioprotective effect of concomitant administration of trigonelline and sitagliptin on cardiac biomarkers, lipid levels, electrocardiographic and heamodynamic modulation on cardiomyopathy in diabetic Wistar rats. Biomed. Aging Pathol. 4, 335–342. doi: 10.1016/j.biomag.2014.07.009
Kanchan, D. M., Somani, G. S., Peshattiwar, V. V., Kaikini, A. A., and Sathaye, S. (2016). Renoprotective effect of diosgenin in streptozotocin induced diabetic rats. Pharmacol. Rep. 68, 370–377. doi: 10.1016/j.pharep.2015.10.011
Kandarkar, S. V., Sawant, S. S., Ingle, A. D., Deshpande, S. S., and Maru, G. B. (1998). Subchronic oral hepatotoxicity of turmeric in mice-histopathological and ultrastructural studies. Indian J. Exp. Biol. 36, 675–679.
Kandhare, A. D., Bodhankar, S. L., Mohan, V., and Thakurdesai, P. A. (2015). Prophylactic efficacy and possible mechanisms of oligosaccharides based standardized fenugreek seed extract on high-fat diet-induced insulin resistance in C57BL/6 mice. J. Appl. Pharm. Sci. 5, 35–45. doi: 10.7324/JAPS.2015.50307
Kang, C., and Kim, C. (2010). Synergistic effect of curcumin and insulin on muscle cell glucose metabolism. Food Chem. Toxicol. 48, 2366–2373. doi: 10.1016/j.fct.2010.05.073
Kang, J. H., Tsuyoshi, G., Han, I. S., Kawada, T., Kim, Y. M., and Yu, R. (2011a). Dietary capsaicin reduces obesity-induced insulin resistance and hepatic steatosis in obese mice fed a high-fat diet. Obesity. 18, 780–787. doi: 10.1038/oby.2009.301
Kang, J. H., Tsuyoshi, G., Le Ngoc, H., Kim, H. M., Tu, T. H, Noh, H. J., et al. (2011b). Dietary capsaicin attenuates metabolic dysregulationin genetically obese diabetic mice. J. Med. Food. 14, 310–315. doi: 10.1089/jmf.2010.1367
Kanitkar, M., Gokhale, K., Galande, S., and Bhonde, R. R. (2008). Novel role of curcumin in the prevention ofcytokine-induced islet death in vitro anddiabetogenesis in vivo. Br. J. Pharmacol. 155, 702–713. doi: 10.1038/bjp.2008.311
Kanter, M. (2008). Effects of Nigella sativa and its major constituent, thymoquinone on sciatic nerves in experimental diabetic neuropathy. Neurochem. Res. 33, 87–96. doi: 10.1007/s11064-007-9419-5
Kanter, M. (2009). Protective effects of thymoquinone on streptozotocin-induced diabetic nephropathy. J. Mol. Histol. 40, 107–115. doi: 10.1007/s10735-009-9220-7
Karthikesan, K., Pari, L., and Menon, V. P. (2010a). Antihyperlipidemic effect of chlorogenic acid and tetrahydrocurcumin in rats subjected to diabetogenic agents. Chem. Biol. Interact. 188, 643–650. doi: 10.1016/j.cbi.2010.07.026
Karthikesan, K., Pari, L., and Menon, V. P. (2010b). Protective effect of tetrahydrocurcumin and chlorogenic acid against streptozotocin-nicotinamide generated oxidative stress induced diabetes. J. Funct. Food. 2, 134–142. doi: 10.1016/j.jff.2010.04.001
Kashef, R. K. H., Hassan, H. M. M., Afify, A. S., Ghabbour, S. I., and Saleh, N. T. (2008). Effect of soybean galactomannan on the activities of α-amylase, trypsin, lipase and starch digestion. J. Appl. Sci. Res. 4, 1893–1897.
Kaur, G., and Meena, C. (2012). Amelioration of obesity, glucose intolerance, and oxidative stress in high-fat diet and low-dose streptozotocin-induceddiabetic rats by combination consisting of “curcumin withpiperine and quercetin”. ISRN Pharmacol. 2012. doi: 10.5402/2012/957283
Kazeem, M. I., and Davies, T. C. (2016). Anti-diabetic functional foods as sources of insulin secreting, insulin sensitizing and insulin mimetic agents. J. Funct. Food. 20, 122–138. doi: 10.1016/j.jff.2015.10.013
Kelble, A. (2005). Spices and type 2 diabetes. Nutr. Food Sci. 8, 392–413. doi: 10.1108/00346650510585868
Kempaiah, R. K., and Srinivasan, K. (2006). Beneficial influence of dietary curcumin, capsaicin and garlic onerythrocyte integrity in high-fat fed rats. J. Nutr. Biochem. 17, 471–478. doi: 10.1016/j.jnutbio.2005.09.005
Khader, M., Bresgen, N., and Eckl, P. M. (2009). In vitro toxicological properties of thymoquinone. Food Chem. Toxicol. 47, 129–133. doi: 10.1016/j.fct.2008.10.019
Khan, A., and Safdar, M. (2003). Role of diet, nutrients, spices and natural products in diabetes mellitus. Pak. J. Nutr. 2, 1–12. doi: 10.3923/pjn.2003.1.12
Khan, M. A., Anwar, S., Aljarbou, A. N., Al-Orainy, M., Aldebasi, Y. H., Islam, S., et al. (2014). Protective effect of thymoquinone on glucose or methylglyoxal-induced glycation of superoxide dismutase. Int. J. Biol. Macromol. 65, 16–20. doi: 10.1016/j.ijbiomac.2014.01.001
Kharbanda, C., Alam, M. S., Hamid, H., Javed, K., Bano, S., Ali, Y., et al. (2016). Novel piperine derivatives with antidiabetic effect as PPAR-γ agonists. Chem. Biol. Drug Des. 88, 354–362. doi: 10.1111/cbdd.12760
Kim, D. H., Hong, B. N., Le, H. T., Hong, H. N., Lim, C. W., Park, K. H., et al. (2012). Small molecular weight PEGylation of diosgenin in an in vivo animal study for diabetic auditory impairment treatment. Bioorg. Med. Chem. Lett. 22, 4609–4612. doi: 10.1016/j.bmcl.2012.05.094
Kim, J. H., Park, J. M., Kim, E. K., Lee, J. O., Lee, S. K., Jung, J. H., et al. (2010). Curcumin stimulates glucose uptake through AMPK-p38 MAPK pathways in L6 Myotube cells. J. Cell Physiol. 223, 771–778. doi: 10.1002/jcp.22093
Kim, S. H., Hwang, J. T., Park, H. S., Kwon, D. Y., and Kim, M. S. (2013). Capsaicin stimulates glucose uptake in C2C12 muscle cells via the reactive oxygen species (ROS)/AMPK/p38 MAPK pathway. Biochem. Biophys. Res. Commun. 439, 66–70. doi: 10.1016/j.bbrc.2013.08.027
Kim, T., Davis, J., Zhang, A. J., He, X., and Mathews, S. T. (2009). Curcumin activates AMPK and suppresses gluconeogenic gene expression in hepatoma cells. Biochem. Biophys. Res. Commun. 388, 377–382. doi: 10.1016/j.bbrc.2009.08.018
Korthikunta, V., Pandey, J., Singh, R., Srivastava, R., Srivastava, A. K., Tamrakar, A. K., et al. (2015). In vitro anti-hyperglycemic activity of 4-hydroxyisoleucine derivatives. Phytomedicine. 22, 66–70. doi: 10.1016/j.phymed.2014.09.007
Kowluru, R. A., and Kanwar, M. (2007). Effects of curcumin on retinal oxidative stress and inflammation in diabetes. Nutr. Metab. 4:8. doi: 10.1186/1743-7075-4-8
Kuhad, A., and Chopra, K. (2007). Curcumin attenuates diabetic encephalopathy in rats: behavioral and biochemical evidences. Eur. J. Pharmacol. 576, 34–42. doi: 10.1016/j.ejphar.2007.08.001
Kumar, P. T., George, N., Antony, S., and Paulose, C. S. (2013). Curcumin restores diabetes induced neurochemical changes in the brain stem of Wistar rats. Eur. J. Pharmacol. 702, 323–331. doi: 10.1016/j.ejphar.2013.01.012
Kumar, S., Sharma, S., and Vasudeva, N. (2013). Screening of antidiabetic and antihyperlipidemic potential of oilfrom Piper longum and piperine with their possible mechanism. Expert. Opin. Pharmacother. 14, 1723–1736. doi: 10.1517/14656566.2013.815725
Kumar, S., Vasudeva, N., and Sharma, S. (2012). GC-MS analysis and screening of antidiabetic, antioxidant and hypolipidemic potential of Cinnamomum tamala oil in streptozotocin induced diabetes mellitus in rats. Cardiovasc. Diabetol. 11:95. doi: 10.1186/1475-2840-11-95
Kuo, W. W., Wang, W. J., Tsai, C. Y., Way, C. L., Hsu, H. H., and Chen, L. M. (2013). Diallyl trisufide (DATS) suppresses high glucose-induced cardiomyocyte apoptosis byinhibiting JNK/NFκB signaling via attenuating ROS generation. Int. J. Cardiol. 168, 270–280. doi: 10.1016/j.ijcard.2012.09.080
Kwon, D. Y., Kim, Y. S., Ryu, S. Y., Chan, M. R., Yon, G. H., Yang, H. J., et al. (2013). Capsiate improves glucose metabolism by improving insulin sensitivity better thancapsaicin in diabetic rats. J. Nutr. Biochem. 24, 1078–1085. doi: 10.1016/j.jnutbio.2012.08.006
Lang, R., Dieminger, N., Beusch, A., Lee, Y. M., Dunkel, A., Suess, B., et al. (2013). Bioappearance and pharmacokinetics of bioactives upon coffee consumption. Anal. Bioanal. Chem. 405, 8487–8503. doi: 10.1007/s00216-013-7288-0
Lang, R., Wahl, A., Skurk, T., Yagar, E. F., Schmiech, L., Eggers, R., et al. (2010). Development of a hydrophilic liquid interaction chromatography-high-performance liquid chromatography-tandem mass spectrometry based stable isotope dilution analysis and pharmacokinetic studies on bioactive pyridines in human plasma and urine after coffee consumption. Anal. Chem. 82, 1486–1497. doi: 10.1021/ac902616k
Lao, C. D., Demierre, M. F., and Sondak, V. K. (2006a). Targeting events in melanoma carcinogenesis for the prevention of melanoma. Expert Rev. Anticancer Ther. 6, 1559–1568. doi: 10.1586/14737140.6.11.1559
Lao, C. D., Ruffin, M. T., Normolle, D., Heath, D. D., Murray, S. I., Bailey, J. M., et al. (2006b). Dose escalation of a curcuminoid formulation. BMC Complement. Altern. Med. 6:10. doi: 10.1186/1472-6882-6-10
LaVoie, E. J., Adams, J. D., Reinhardt, J., Rivenson, A., and Hoffmann, D. (1986). Toxicity studies on clove cigarette smoke and constituents of clove: determination of the LD50 of eugenol by intratracheal instillation in rats and hamsters. Arch. Toxicol. 59, 78–81. doi: 10.1007/BF00286727
Lee, C., Park, G. H., Kim, C. Y., and Jang, J. H. (2011). [6]-Gingerol attenuates β-amyloid-induced oxidative cell death via fortifying cellular antioxidant defense system. Food Chem. Toxicol. 49, 1261–1269. doi: 10.1016/j.fct.2011.03.005
Lee, H. S. (2002). Inhibitory activity of Cinnamomum cassia bark-derived component against rat lens aldose reductase. J. Pharm. Pharmaceut. Sci. 5, 226–230.
Lee, H. S. (2005). Cuminaldehyde: aldose reductase and α-glucosidase inhibitor derived from Cuminum cyminum L. seeds. J. Agric. Food Chem. 53, 2446–2450. doi: 10.1021/jf048451g
Lee, J. O., Kim, N., Lee, H. J., Moon, J. W., Lee, S. K., Kim, S. J., et al. (2015). [6]-Gingerol affects glucose metabolism by dual regulation via the AMPKα2-mediated AS160–Rab5 pathway and AMPK-mediated insulin sensitizing effects. J. Cell Biochem. 116, 1401–1410. doi: 10.1002/jcb.25100
Lee, M. S., Kim, C. T., Kim, I. H., and Kim, Y. (2011). Effects of capsaicin on lipid catabolism in 3T3-L1 adipocytes. Phytother. Res. 25, 935–939. doi: 10.1002/ptr.3339
Lekshmi, P. C., Arimboor, R., Indulekha, P. S., and Nirmala, M. A. (2012a). Turmeric (Curcuma longa L.) volatile oil inhibits key enzymes linked to type 2 diabetes. Int. J. Food Sci. Nutr. 63, 832–834. doi: 10.3109/09637486.2011.607156
Lekshmi, P. C., Arimboor, R., Raghu, K. G., and Nirmala, M. A. (2012b). Turmerin, the antioxidant protein from turmeric (Curcuma longa) exhibits antihyperglycaemic effects. Nat. Prod. Res. 26, 1654–1658. doi: 10.1080/14786419.2011.589386
Li, J., Liu, T., Wang, L., Guo, X., Xu, T., Wu, L., et al. (2012). Antihyperglycemic and antihyperlipidemic action of cinnamaldehyde in C57blks/j db/db mice. J. Tradit. Chin. Med. 32, 1–2. doi: 10.1016/S0254-6272(13)60053-9
Li, J. E., Futawaka, K., Yamamoto, H., Kasahara, M., Tagami, T., Liu, T. H., et al. (2015). Cinnamaldehyde contributes to insulin sensitivity by activating PPARδ, PPARγ, and RXR. Am. J. Chinese Med. 43, 879–892. doi: 10.1142/S0192415X15500512
Li, Y., Tran, V. H., Duke, C. C., and Roufogalis, B. D. (2012). Gingerols of Zingiber officinale enhance glucose uptake by increasing cell surface GLUT4 in cultured L6 myotubes. Planta Med. 78, 1549–1555. doi: 10.1055/s-0032-1315041
Li, Y., Tran, V. H., Koolaji, H., Duke, C. C., and Roufogalis, B. D. (2013). (S)- [6]-Gingerol enhances glucose uptake in L6 myotubes by activation of AMPK in response to [Ca2+]i. J. Pharm. Pharm. Sci. 16, 304–312. doi: 10.18433/J34G7P
Lin, J., and Chen, A. (2011). Curcumin diminishes the impacts of hyperglycemia on the activation of hepatic stellate cells by suppressing membrane translocation and gene expression of glucose transporter-2. Mol. Cell Endocrinol. 333, 160–171. doi: 10.1016/j.mce.2010.12.028
Lin, J., Zheng, S., and Chen, A. (2009). Curcumin attenuates the effects of insulin on stimulating hepatic stellate cell activation by interrupting insulin signalling and attenuating oxidative stress. Lab. Invest. 89, 1397–1409. doi: 10.1038/labinvest.2009.115
Liu, C. T., Hse, H., Lii, C. K., Chen, P. S., and Sheen, L. Y. (2005). Effects of garlic oil and diallyl trisulfide on glycemic control in diabetic rats. Eur. J. Pharmacol. 516, 165–173. doi: 10.1016/j.ejphar.2005.04.031
Liu, C. T., Wong, P. L., Lii, C. K., Hse, H., and Sheen, L. Y. (2006). Antidiabetic effect of garlic oil but not diallyl disulfide in rats with streptozotocin-induced diabetes. Food Chem. Toxicol. 44, 1377–1384. doi: 10.1016/j.fct.2005.07.013
Liu, C. Z., Chang, J. H., Zhang, L., Xue, H. F., Liu, X. G., Liu, P., et al. (2016). Preparation and evaluation of diosgenin nanocrystals to improve oral bioavailability. AAPS Pharm. Sci. Tech. 18, 2067–2076. doi: 10.1208/s12249-016-0684-y
Liu, K., Zhao, W., Gao, X., Huang, F., Kou, J., and Liu, B. (2012). Diosgenin ameliorates palmitate-induced endothelial dysfunction and insulin resistance via blocking IKKβ and IRS-1 pathways. Atherosclerosis 223, 350–358. doi: 10.1016/j.atherosclerosis.2012.06.012
Losso, J. N., Bawadi, H. A., and Chintalapati, M. (2011). Inhibition of the formation of advanced glycation end products by thymoquinone. Food Chem. 128, 55–61. doi: 10.1016/j.foodchem.2011.02.076
Luo, J., Wärmländer, S. K., Gräslund, A., and Abrahams, J. P. (2016). Reciprocal molecular interactions between the Aβ peptide linked to Alzheimer's disease and insulin linked to diabetes mellitus type II. ACS Chem. Neurosci. 7, 269–274. doi: 10.1021/acschemneuro.5b00325
Maher, P. A., and Schubert, D. R. (2009). Metabolic links between diabetes and Alzheimer's disease. Expert. Rev. Neurother. 9, 617–630. doi: 10.1586/ern.09.18
Mahesh, T., Sri Balasubashini, M. M., and Menon, V. P. (2004). Photo-irradiated curcumin supplementation in streptozotocin-induced diabetic rats: effect on lipid peroxidation. Therapie. 59, 639–644. doi: 10.2515/therapie:2004110
Mahesh, T., Sri Balasubashini, M. M., and Menon, V. P. (2005). Effect of photo-irradiated curcumin treatment against oxidative stress instreptozotocin-induced diabetic rats. J. Med. Food. 8, 251–255. doi: 10.1089/jmf.2005.8.251
Mahfouz, M. K. M. (2011). Curcumin improves insulin sensitivity and ameliorates serum pro-inflammatory cytokines levels indiabetes rat model irrespective of type of diabetes. J. Am. Sci. 7, 794–799.
Mansour, M. A., Ginawi, O. T., El-Hadiyah, T., El-Khatib, A. S., Al-Shabanah, O. A., and Al-Sawaf, H. A. (2001). Effects of volatile oil constituents of Nigella sativa on carbon tetrachloride-induced hepatotoxicity in mice: evidence for antioxidant effects of thymoquinone. Res. Commun. Mol. Pathol. Pharmacol. 110, 239–252.
Marczylo, T. H., Steward, W. P., and Gescher, A. J. (2009). Rapid analysis of curcumin and curcumin metabolites in rat biomatrices using a novel ultra-performance liquid chromatography (UPLC) method. J. Agric. Food Chem. 57, 797–803. doi: 10.1021/jf803038f
Marques, S., Oliveira, N. G., Chaveca, T., and Rueff, J. (2002). Micronuclei and sister chromatid exchanges induced by capsaicin in human lymphocytes. Mutat. Res. 517, 39–46. doi: 10.1016/S1383-5718(02)00040-2
Masamune, A., Suzuki, N., Kikuta, K., Satoh, M., Satoh, K., and Shimosegawa, T. (2006). Curcumin blocks activation of pancreatic stellate cells. J. Cell Biochem. 97, 1080–1093. doi: 10.1002/jcb.20698
Mathew, P. T., and Augusti, K. T. (1973). Studies on the effect of allicin (diallyl disulphide-oxide) on alloxan diabetes. I. Hypoglycaemic action and enhancement of serum insulin effect and glycogen synthesis. Indian J. Biochem. Biophys. 10, 209–121.
Maurya, C. K., Singh, R., Jaiswal, N., Venkateswarlu, K., Narender, T., and Tamrakar, A. K. (2014). 4-Hydroxyisoleucine ameliorates fatty acid-induced insulin resistanceand inflammatory response in skeletal muscle cells. Mol. Cell Endocrinol. 395, 51–60. doi: 10.1016/j.mce.2014.07.018
McAnuff, M. A., Harding, W. W., Omoruyi, F. O., Jacobs, H., Morrison, E. Y. A., and Asemota, H. N. (2005). Hypoglycemic effects of steroidal sapogenins isolated from Jamaican bitter yam, Dioscorea polygonoides. Food Chem. Toxicol. 43, 1667–1672. doi: 10.1016/j.fct.2005.05.008
McAnuff, M. A., Omoruyi, F. O., Morrison, E. Y. A., and Asemota, H. N. (2002). Plasma and liver lipid distributions in streptozotocin induced diabetic rats fed sapogenin extract of the Jamaican bitter yam (Dioscorea polygonoides). Nutr. Res. 22, 1427–1434. doi: 10.1016/S0271-5317(02)00457-8
McAnuff-Harding, M. A., Omoruyi, F. O., and Asemota, H. N. (2006). Intestinal disaccharidases and some renal enzymes in streptozotocin-induceddiabetic rats fed sapogenin extract from bitter yam (Dioscorea polygonoides). Life Sci. 78, 2595–2600. doi: 10.1016/j.lfs.2005.10.046
Meghana, K., Sanjeev, G., and Ramesh, B. (2007). Curcumin prevents streptozotocin-induced islet damage by scavenging free radicals: a prophylactic and protective role. Eur. J. Pharmacol. 577, 183–191. doi: 10.1016/j.ejphar.2007.09.002
Mehrdad, R., and Tourandokht, B. (2012). Dose-dependent effect of thymoquinone on markers of oxidative stress in renal tissue of diabetic rats. Daneshvar. Med. 19, 57–64.
Millqvist, E., Ternesten-Hasseus, E., Stahl, A., and Bende, M. (2005). Changes in levels of nerve growth factor in nasal secretions after capsaicin inhalation in patients with airway symptoms from scents and chemicals. Environ. Health Perspect. 13, 849–852. doi: 10.1289/ehp.7657
Mnafgui, K., Kaanich, F., Derbali, A., Hamden, K., Derbali, F., Slama, S., et al. (2013). Inhibition of key enzymes related to diabetes and hypertension by Eugenol in vitro and in alloxan-induced diabetic rats. Arch Physiol Biochem. doi: 10.3109/13813455.2013.822521
Mohamed, S. (2014). Functional foods against metabolic syndrome (obesity, diabetes, hypertension and dyslipidemia) and cardiovasular disease. Trend Food Sci. Technol. 35, 114–128. doi: 10.1016/j.tifs.2013.11.001
Mohammed, A., Gbonjubola, V. A., Koorbanally, N. A., and Islam, M. S. (2017). Inhibition of key enzymes linked to type 2 diabetes by compounds isolated from Aframomum melegueta fruit. Pharm. Biol. 55, 1010–1016. doi: 10.1080/13880209.2017.1286358
Monago, C. C., and Nwodo, O. F. C. (2010). Antidiabetic effect of crude trigonelline of Abrus precatorius Linn seed in alloxan diabetic rabbits. J. Pharm. Res. 3, 1916–1919.
Mong, M. C., and Yin, M. C. (2012). Nuclear factor κB-dependent anti-inflammatory effects of S-allylcysteine and S-propyl cysteine in kidney of diabetic mice. J. Agric. Food Chem. 60, 3158–3165. doi: 10.1021/jf3002685
Monsereenusorn, Y., and Glinsukon, T. (1978). Inhibitory effect of capsaicin on intestinal glucose absorption in vitro. Food Cosmetic Toxicol. 16, 469–473. doi: 10.1016/S0015-6264(78)80305-8
Monsereenusorn, Y., and Glinsukon, T. (1979). The inhibitory effect of capsaicin on intestinal glucose absorption in vitro: II. Effect of capsaicin upon intestinal Na+-K+-ATPase activities. Toxicol. Lett. 4, 399–406. doi: 10.1016/0378-4274(79)90052-3
Moorthi, C., and Kathiresan, K. (2013). Curcumin-Piperine/Curcumin-Quercetin/Curcumin-Silibinin dual drug-loaded nanoparticulate combination therapy: a novel approach to target and treat multidrug-resistant cancers. J. Med. Hypoth. Ideas. 7, 15–20. doi: 10.1016/j.jmhi.2012.10.005
Mrudula, T., Suryanarayana, P., Srinivas, P. N., and Reddy, G. B. (2007). Effect of curcumin on hyperglycemia-induced vascular endothelial growth factor expression in streptozotocin-induced diabetic rat retina. Biochem. Biophys. Res. Commun. 361, 528–532. doi: 10.1016/j.bbrc.2007.07.059
Murugan, P., and Pari, L. (2006a). Effect of Tetrahydrocurcumin on lipid peroxidation and lipids in streptozotocin-nicotinamide-induced diabetic rats. Basic Clin. Pharmacol. Toxicol. 99, 122–127. doi: 10.1111/j.1742-7843.2006.pto_447.x
Murugan, P., and Pari, L. (2006b). Antioxidant effect of tetrahydrocurcumin in streptozotocin-nicotinamide induced diabetic rats. Life Sci. 79, 1720–1728. doi: 10.1016/j.lfs.2006.06.001
Murugan, P., and Pari, L. (2007). Influence of tetrahydrocurcumin on hepatic and renal functional markers and protein levels in experimental type 2 diabetic rats. Basic Clin. Pharmacol. Toxicol. 101, 241–245. doi: 10.1111/j.1742-7843.2007.00109.x
Murugan, P., Pari, L., and Rao, C. A. (2008). Effect of tetrahydrocurcumin on insulin receptor status in type 2 diabetic rats: studies on insulin binding to erythrocytes. J. Biosci. 33, 63–72. doi: 10.1007/s12038-008-0022-y
Na, L. X., Zhang, Y. L., Li, Y., Liu, L. Y., Li, R., Kong, T., et al. (2011). Curcumin improves insulin resistance in skeletal muscle of rats. Nutr. Metab. Cardiovasc. Dis. 21, 526–533. doi: 10.1016/j.numecd.2009.11.009
Naicker, N., Nagiah, S., Phulukdaree, A., and Chuturgoon, A. (2016). Trigonella foenum-graecum seed extract, 4-hydroxyisoleucine, and metformin stimulate proximal insulin signaling and increase expression of glycogenic enzymes and GLUT2 in HepG2 cells. Metab. Syndr. Relat. Disord. 14, 114–120. doi: 10.1089/met.2015.0081
Naidu, P. B., Ponmurugan, P., Begum, M. S., Mohan, K., Meriga, B., RavindarNaik, R., et al. (2015). Diosgenin reorganises hyperglycaemia and distorted tissue lipid profile in high-fat diet-streptozotocin-induced diabetic rats. J. Sci. Food Agric. 95, 3177–3182. doi: 10.1002/jsfa.7057
Naito, M., Wu, X., Nomura, H., Kodama, M., Kato, Y., Kato, Y., et al. (2002). The protective effects of tetrahydrocurcumin on oxidative stress in cholesterol-fed rabbits. J. Atherosclerosis Thromb. 9, 243–250. doi: 10.5551/jat.9.243
Namekata, I., Hamaguchi, S., Wakasugi, Y., Ohhara, M., Hirota, Y., and Tanaka, H. (2013). Ellagic acid and gingerol, activators of the sarco-endoplasmic reticulum Ca2+-ATPase, ameliorate diabetes mellitus-induced diastolic dysfunction in isolated murine ventricular myocardia. Eur. J. Pharmacol. 706, 48–55. doi: 10.1016/j.ejphar.2013.02.045
Nangle, M. R., Gibson, T. M., Cotter, M. A., and Cameron, N. E. (2006). Effects of eugenol on nerve and vascular dysfunction in streptozotocin-diabetic rats. Planta Med. 72, 1–7. doi: 10.1055/s-2005-916262
Narender, T., Puri, A., Shweta Khaliq, T., Saxena, R., Bhatia, G., and Chandra, R. (2006). 4-Hydroxyisoleucine an unusual amino acid as antidyslipidemic and antihyperglycemic agent. Bioorg. Med. Chem. Lett. 16, 293–296. doi: 10.1016/j.bmcl.2005.10.003
Neerati, P., Devde, R., and Gangi, A. K. (2014). Evaluation of the effect of curcumin capsules onglyburide therapy in patients with type-2diabetes mellitus. Phytother Res. 28, 1796–1800. doi: 10.1002/ptr.5201
Nevius, E., Srivastava, P. K., and Basu, S. (2012). Oral ingestion of capsaicin, the pungent component of chili pepper, enhances a discreet population of macrophages and confers protection from autoimmune diabetes. Mucosal Immunol. 5, 76–86. doi: 10.1038/mi.2011.50
Nikzamir, A., Palangi, A., Kheirollaha, A., Tabar, H., Malakaskar, A., Shahbazian, H., et al. (2014). Expression of glucose transporter 4 (GLUT4) is increased by cinnamaldehyde in C2C12 mouse muscle cells. Iranian Red Crescent. Med. J. 16:e13426. doi: 10.5812/ircmj.13426
Nuttall, F. Q., Schweim, K., and Gannon, M. C. (2008). Effect of orally administered isoleucine with and without glucose on insulin, glucagon and glucose concentrations in non-diabetic subjects. e-SPEN Eur. J. Clin. Nutr. Metabol. 3, 152–158. doi: 10.1016/j.eclnm.2008.05.001
Ogurtsova, K., da Rocha Fernandes, J. D., Huang, Y., Linnenkamp, U., Guariguata, L., Cho, N. H., et al. (2017). IDF Diabetes Atlas: global estimates for the prevalence of diabetes for 2015 and 2040. Diabetes Res. Clin. Pract. 28, 40–50. doi: 10.1016/j.diabres.2017.03.024
Okawara, M., Hashimoto, F., Todo, H., Sugibayashi, K., and Tokudome, Y. (2014). Effect of liquid crystals with cyclodextrin on the bioavailability of a poorly water-soluble compound, diosgenin, after its oral administration to rats. Int. J. Pharm. 472, 257–261. doi: 10.1016/j.ijpharm.2014.06.032
Okawara, M., Tokudome, Y., Todo, H., Sugibayashi, K., and Hashimoto, F. (2010). Diosgenin disposition in rats after i.v. and p.o. administration. Yakuzaigaku 70, 82–86.
Okawara, M., Tokudome, Y., Todo, H., Sugibayashi, K., and Hashimoto, F. (2013). Enhancement of diosgenin distribution in the skin by cyclodextrin complexation following oral administration. Biol. Pharm. Bull. 36, 36–40. doi: 10.1248/bpb.b12-00467
Okumura, T., Tsukui, T., Hosokawa, M., and Miyashita, K. (2012). Effect of caffeine and capsaicin on the blood glucose levels of obese/diabetic KK-Ay mice. J. Oleo. Sci. 61, 515–523.
Olthof, M. R., van Dijk, A. E., Deacon, C. F., Heine, R. J., and van Dam, R. M. (2011). Acute effects of decaffeinated coffee and the major coffee components chlorogenic acid and trigonelline on glucose tolerance. Nutr. Metabol. 8:10. doi: 10.1186/1743-7075-8-10.
Ong, Y. S., Yazan, L., Ng, W. K., Noordindr, M. M., Sapuan, S., and Foo, J. B (2016). Acute and subacute toxicity profiles of thymoquinone-loaded nanostructured lipid carrier in BalB/c mice. Int. J. Nanomed. 11, 5905–5915. doi: 10.2147/IJN.S114205
Palma, H. E., Wolkmer, P., Gallio, M., Corrèa, M. M. B., Schmatz, R., Thomè, G. R., et al. (2014). Oxidative stress parameters in blood, liver, and kidney of diabetic rats treated with curcumin and/or insulin. Mol. Cell Biochem. 386, 199–210. doi: 10.1007/s11010-013-1858-5
Pan, M. H., Huang, T. M., and Lin, J. K. (1999). Biotransformation of curcumin through reduction and glucuronidation in mice. Drug Metab. Dispos. 27, 486–494.
Pandey, M. K., and DeGrado, T. R. (2016). Glycogen synthase kinase-3 (GSK-3)-targeted therapy and imaging. Theranostics 6, 571–593. doi: 10.7150/thno.14334
Pari, L., Monisha, P., and Mohamed Jalaludeen, A. (2012). Beneficial role of diosgenin on oxidative stress in aorta of streptozotocin induced diabetic rats. Eur. J. Pharmacol. 691, 143–150. doi: 10.1016/j.ejphar.2012.06.038
Pari, L., and Murugan, P. (2005). Effect of tetrahydrocurcumin on blood glucose, plasma insulin and hepatic key enzymes in streptozotocin induced diabetic rats. J. Basic Clin. Physiol. Pharmacol. 16, 257–274. doi: 10.1515/JBCPP.2005.16.4.257
Pari, L., and Murugan, P. (2007). Tetrahydrocurcumin prevents brain lipid peroxidation in streptozotocin-induced diabetic rats. J. Med. Food 10, 323–329. doi: 10.1089/jmf.2006.058
Pari, L., and Murugan, P. (2007b). Influence of tetrahydrocurcumin on tail tendon collagen contents and its properties in rats with streptozotocin-nicotinamide induced type 2 diabetes. Fundam. Clin. Pharmacol. 21, 665–671. doi: 10.1111/j.1472-8206.2007.00542.x
Pari, L., and Sankaranarayanan, C. (2009). Beneficial effects of thymoquinone on hepatic key enzymes in streptozotocin–nicotinamide induced diabetic rats. Life Sci. 85, 830–834. doi: 10.1016/j.lfs.2009.10.021
Pari, P., and Murugan, P. (2008). Antihyperlipidemic effect of curcumin and tetrahydrocurcumin in experimental type 2 diabetic rats. Ren. Fail. 29, 881–889. doi: 10.1080/08860220701540326
Pathan, S. A., Jain, G. K., Zaidi, S. M., Akhter, S., Vohora, D., Chander, P., et al. (2011). Stability-indicating ultra-performance liquid chromatography method for the estimation of thymoquinone and its application in biopharmaceutical studies. Biomed. Chrom. 25, 613–620. doi: 10.1002/bmc.1492
Peeyush, K. T., Gireesh, G., Jobin, M., and Paulose, C. S. (2009). Neuroprotective role of curcumin in the cerebellum of streptozotocin-induced diabetic rats. Life Sci. 85, 704–710. doi: 10.1016/j.lfs.2009.09.012
Peters, M. M., and Caldwell, J. (1994). Studies on trans-cinnamaldehyde. The influence of dose size and sex on its disposition in the rat and mouse. Food Chem. Toxicol. 32, 869–876. doi: 10.1016/0278-6915(94)90084-1
Piyachaturawat, P., Glinsukon, T., and Toskulkao, C. (1983). Acute and subacute toxicity of piperine in mice, rats and hamsters. Toxicol. Lett. 16, 351–359.
Prabhakar, P. K., and Doble, M. (2011). Interaction of phytochemicals with hypoglycemic drugs on glucoseuptake in L6 myotubes. Phytomedicine 18, 285–291. doi: 10.1016/j.phymed.2010.06.016
Prasad, S. N., Bharath, M. M., and Muralidhara, N. (2015). Neurorestorative effects of eugenol, a spice bioactive: evidence in cell model and its efficacy as an intervention molecule to abrogate brain oxidative dysfunctions in the streptozotocin diabetic rat. Neurochem. Int. 95, 24–36. doi: 10.1016/j.neuint.2015.10.012
Pulkkinen, L., Ukkola, O., Kolehmainen, M., and Uusitupa, M. (2010). Ghrelin in diabetes and metabolic syndrome. Int. J. Peptides 2010:248948. doi: 10.1155/2010/248948
Qadri, S. M., Mahmud, H., Föller, M., and Lang, F. (2009). Thymoquinone-induced suicidal erythrocyte death. Food Chem. Toxicol. 47, 1545–1549. doi: 10.1016/j.fct.2009.03.037
Raffai, G., Kim, B., Park, S., Khang, G., Lee, D., and Vanhoutte, P. M. (2014). Cinnamaldehyde and cinnamaldehyde-containing micelles induce relaxation of isolated porcine coronary arteries: role of nitric oxide and calcium. Int. J. Nanomed. 9, 2557–2566. doi: 10.2147/IJN.S56578
Rahimi, H. R., Mohammadpour, A. H., Dastani, M., Jaafari, M. R., Abnous, K., Mobarhan, M. G., et al. (2016). The effect of nano-curcumin on HbA1c, fasting blood glucose, and lipid profile in diabetic subjects: a randomized clinical trial. Avicenna J. Phytomed. 6, 567–577.
Raju, J., and Rao, C. V. (2011). “Diosgenin, a steroid saponin constituent of yams and fenugreek: emerging evidence for applications in medicine,” in Bioactive Compounds in Phytomedicine, ed I. Rasooli Croatia: InTech). 125.
Ramadan, A. A., Afifi, N. A., Erian, E. Y., Saleh, D. O., and Sedik, A. A. (2016). Beneficial effect of trigonelline on the metabolic changes associated with insulin resistance in rats. World J. Pharm. Pharm. Sci. 5, 1238–1250.
Rao, B. N., Rao, B. S., Aithal, B. K., and Kumar, M. R. (2009). Radiomodifying and anticlastogenic effect of zingerone on Swiss albino mice exposed to whole body gamma radiation. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 677, 33–41. doi: 10.1016/j.mrgentox.2009.05.004
Rao, R., and Natarajan, S. (1949). Toxicity of pterygospermin and allicin. P Indian Acad. Sci. A 29, 148–154.
Rashid, K., and Sil, P. C. (2015). Curcumin ameliorates testicular damage in diabetic rats by suppressingcellular stress-mediated mitochondria and endoplasmicreticulum-dependent apoptotic death. Biochimica et. Biophysica. Acta 1852, 70–82. doi: 10.1016/j.bbadis.2014.11.007
Rauscher, F. M., Sanders, R. A., and Watkins, J. B. III. (2000). Effects of piperine on antioxidant pathways in tissuesfrom normal and streptozotocin-induced diabetic rats. J. Biochem. Mol. Toxicol. 14, 329–334. doi: 10.1002/1099-0461(2000)14:6<329::AID-JBT5>3.0.CO;2-G
Rauscher, F. M., Sanders, R. A., and Watkins, J. B. III. (2001). Effects of isoeugenol on oxidative stress pathways innormal and streptozotocin-induced diabetic rats. J. Biochem. Mol. Toxicol. 15, 159–164. doi: 10.1002/jbt.13
Ravindran, J., Nair, H. B., Sung, B., Prasad, S., Tekmal, R. R., and Aggarwal, B. B. (2010). Thymoquinone poly (lactide-co-glycolide) nanoparticles exhibit enhanced anti-proliferative, anti-inflammatory, and chemosensitization potential. Biochem. Pharmacol. 79, 1640–1647. doi: 10.1016/j.bcp.2010.01.023
Ravindranath, V., and Chandrasekhara, N. (1982). Metabolism of curcumin-studies with [3H] curcumin. Toxicology. 22, 337–344. doi: 10.1016/0300-483X(81)90027-5
Rawat, A. K., Korthikunta, V., Gautam, S., Pal, S., Tadigoppula, N., Tamrakar, A. K., et al. (2014). 4-Hydroxyisoleucine improves insulin resistance by promoting mitochondrial biogenesis and act through AMPK and Akt dependent pathway. Fitoterapia 99, 307–317. doi: 10.1016/j.fitote.2014.10.006
Roghani, M., and Baluchnejadmojarad, T. (2012). Dose-dependent effect of thymoquinone on markers of oxidative stress in renal tissue of diabetic rats. Daneshvar Med. 19, 57–64.
Roghani-Dehkordi, F., Roghani, M., and Baluchnejadmojarad, T. (2015). Diosgenin mitigates streptozotocin diabetes-induced vascular dysfunction of the rat aorta: the involved mechanisms. J. Cardiovasc. Pharmacol. 66, 584–592. doi: 10.1097/FJC.0000000000000308
Rouse, M., Younès, A., and Eganm, J. M. (2014). Resveratrol and curcumin enhancepancreatic β-cell function by inhibiting phosphodiesterase activity. J. Endocrinol. 223, 107–117. doi: 10.1530/JOE-14-0335
Rungseesantivanon, S., Thenchaisri, N., Ruangvejvorachai, P., and Patumraj, S. (2010). Curcumin supplementation could improve diabetes-induced endothelial dysfunction associated with decreased vascular superoxide production and PKC inhibition. BMC Complement. Altern. Med. 10:57. doi: 10.1186/1472-6882-10-57
Ryndina, S. E., Shashkina, L. F., and Starkov, M. V. (1977). Toxicity of solasodine and diosgenin. Pharm. Chem. J. 11, 1095–1100. doi: 10.1007/BF00778191
Saheb, S. H., Desai, S. D., Das, K. K., and Haseena, S. (2016). Antioxidant effect of Nigella sativa seed powder and thymoquinone in normal and sterptozotocine induced diabetic albino rats. Int. J. Intg Med Sci. 3, 242–247. doi: 10.16965/ijims.2016.108
Saito, A., and Yamamoto, M. (1996). Acute oral toxicity of capsaicin in mice and rats. J. Toxicol. Sci. 21, 195–200. doi: 10.2131/jts.21.3_195
Sajithlal, G. B., Chithra, P., and Chandrakasan, G. (1998). Effect of curcumin on the advanced glycation and cross-linking of collagen in diabetic rats. Biochem. Pharmacol. 56, 1607–1614. doi: 10.1016/S0006-2952(98)00237-8
Salehi, P., Nasri, S., Roghani, M., Poordahandeh, U., and Baluchnejadmojarad, T. (2012). The effect of thymoquinone on short-term spatial memory, passive avoidance learning and memory of diabetic rats and the involvement of hippocampal oxidative stress. Pajoohandeh J. 17, 219–227.
Sampath, C., Rashid, M. R., Sang, S., and Ahmedna, M. (2017). Specific bioactive compounds in ginger and apple alleviate hyperglycemia in mice with high fat diet-induced obesity via Nrf2 mediated pathway. Food Chem. 226, 79–88. doi: 10.1016/j.foodchem.2017.01.056
Sampath, C., Sang, S., and Ahmedna, M. (2016). In vitro and in vivo inhibition of aldose reductase and advanced glycation end products by phloretin, epigallocatechin 3-gallate and [6]-gingerol. Biomed. Pharmacother. 84, 502–513. doi: 10.1016/j.biopha.2016.09.073
Sangeetha, M. K., ShriShri Mal, N., Atmaja, K., Sali, V. K., and Vasanthi, H. R. (2013). PPAR's and diosgenin a chemico biological insight in NIDDM. Chem. Biol. Interact. 206, 403–410. doi: 10.1016/j.cbi.2013.08.014
Sangi, S. M. A., Sulaiman, M. I., El-wahab, M. F., Ahmedani, E. I., and Ali, S. S. (2015). Antihyperglycemic effect of thymoquinone and oleuropein, on streptozotocin-induced diabetes mellitus in experimental animals. Pharmacogn. Mag. 11, S251–S257. doi: 10.4103/0973-1296.166017
Sankaranarayanan, C., and Pari, L. (2011a). Thymoquinone ameliorates chemical induced oxidative stress and β-cell damage in experimental hyperglycemic rats. Chem. Biol. Interact. 190, 148–154. doi: 10.1016/j.cbi.2011.02.029
Sankaranarayanan, C., and Pari, L. (2011b). Influence of thymoquinone on glycoprotein changes in experimental hyperglycemic rats. Int. J. Nutr. Pharmacol. Neurol. Dis. 1, 15–55. doi: 10.4103/2231-0738.77532
Saravanan, G., and Ponmurugan, P. (2010). Beneficial effect of S-allylcysteine (SAC) on blood glucoseand pancreatic antioxidant system in streptozotocindiabetic rats. Plant Food Hum. Nutr. 65, 374–378. doi: 10.1007/s11130-010-0192-2
Saravanan, G., and Ponmurugan, P. (2011). Ameliorative potential of S-allyl cysteine on oxidative stressin STZ induced diabetic rats. Chem. Biol. Interact. 189, 100–106. doi: 10.1016/j.cbi.2010.10.001
Saravanan, G., and Ponmurugan, P. (2012a). Ameliorative potential of S-allylcysteine: effect on lipid profile and changes intissue fatty acid composition in experimental diabetes. Exp. Toxicol. Pathol. 64, 639–644. doi: 10.1016/j.etp.2010.12.007
Saravanan, G., and Ponmurugan, P. (2012b). Antidiabetic effect of S-allylcysteine: effect on thyroid hormone and circulatoryantioxidant system in experimental diabetic rats. J. Diabetes Complic. 26, 280–285. doi: 10.1016/j.jdiacomp.2012.03.024
Saravanan, G., Ponmurugan, P., and Begum, M. S. (2013). Effect of S-allylcysteine, a sulphur containing amino acid on iron metabolism instreptozotocin induced diabetic rats. J. Trace Element Med. Biol. 27, 143–147. doi: 10.1016/j.jtemb.2012.07.009
Saravanan, G., Ponmurugan, P., Deepa, M. A., and Senthilkumar, B. (2014). Modulatory effects of diosgenin on attenuating the key enzymes activities of carbohydrate metabolism and glycogen content in streptozotocin-induced diabetic rats. Canadian J. Diabetes 38, 409–414. doi: 10.1016/j.jcjd.2014.02.004
Saravanan, G., Ponmurugan, P., Senthil Kumar, G. P., and Rajarajan, T. (2010). Antidiabetic effect of S-allylcysteine: effect on plasma and tissue glycoproteins in experimental diabetes. Phytomedicine 17, 1086–1089. doi: 10.1016/j.phymed.2010.04.008
Saravanan, G., Ponmurugan, P., Senthilkumar, G. P., and Rajarajan, T. (2009). Modulatory effect of S-allylcysteine on glucose metabolismin streptozotocin induced diabetic rats. J. Funct. Food 1, 336–340. doi: 10.1016/j.jff.2009.09.001
Satapathy, T., and Panda, P. K. (2013). Evaluation of in vitro antioxidant, anti-inflammatory and anti-diabetic potential of curcumin. Indo. Am. J. Pharm. Res. 3, 2808–2818.
Sato, K., Fujita, S., and Iemitsu, M. (2014). Acute administration of diosgenin or dioscorea improves hyperglycemia with increases muscular steroidogenesis in STZ-induced type 1 diabetic rats. J. Steroid Biochem. Mol. Biol. 143, 152–159. doi: 10.1016/j.jsbmb.2014.02.020
Sauvaire, Y., Petit, P., Broca, C., Manteghetti, M., Baissac, Y., Fernandez-Alvarez, J., et al. (1998). 4-Hydroxyisoleucine a novel amino acid potentiator of insulin secretion. Diabetes 47, 206–210. doi: 10.2337/diab.47.2.206
Scheffler, N. M., Sheitel, P. L., and Lipton, M. N. (1991). Treatment of painful diabetic neuropathy with capsaicin 0.075%. J. Am. Paediatric Med. Assoc. 81, 288–293. doi: 10.7547/87507315-81-6-288
Schneider-Stock, R., Fakhoury, I. H., Zaki, A. M., El-Baba, C. O., and Gali-Muhtasib, H. U. (2014). Thymoquinone: fifty years of success in the battle against cancer models. Drug Dis. Today 19, 18–30. doi: 10.1016/j.drudis.2013.08.021
Semwal, R. B., Semwal, D. K., Combrinck, S., and Viljoen, A. M. (2015). Gingerols and shogaols: important nutraceutical principles from ginger. Phytochemistry 117, 554–568. doi: 10.1016/j.phytochem.2015.07.012
Seo, K. I., Choi, M. S., Jung, U. J., Kim, H. J., Yeo, J., Jeon, S. M., et al. (2008). Effect of curcumin supplementation on blood glucose, plasma insulin, and glucose homeostasis related enzyme activities in diabetic db/db mice. Mol. Nutr. Food Res. 52, 995–1004. doi: 10.1002/mnfr.200700184
Shah, S., Bodhankar, S., Bhonde, R., and Mohan, V. (2009). Regenerative potential of pancreata in alloxan induced diabetic mice by 4-hydroxyisoleucine, comparision with pioglitazone. Int. J. Integrat. Biol. 5, 136–141.
Shah, S., Shah, G., Patel, M., and Singh, S. (2010). Effect of piperine in obesity induced insulin resistance and type-II diabetes mellitus in rats. J. Nat. Remed. 10, 116–122.
Shah, S. N., Bodhankar, S. L., Badole, S. L., Kamble, H. V., and Mohan, V. (2006). Effect of trigonelline: an active compound from Trigonella foenumgraecum Linn. in alloxan induced diabetes in mice. J. Cell Tissue Res. 6, 585–590.
Shaikh, J., Ankola, D. D., Beniwal, V., Singh, D., and Kumar, M. N. (2009). Nanoparticle encapsulation improves oral bioavailability of curcumin by at least 9-fold when compared to curcumin administered with piperine as absorption enhancer. Eur. J. Pharm. Sci. 37, 223–230. doi: 10.1016/j.ejps.2009.02.019
Shankar, T. N., Shantha, N. V., Ramesh, H. P., Murthy, I. A., and Murthy, V. S. (1980). Toxicity studies on turmeric (Curcuma longa): acute toxicity studies in rats, guinea pigs & monkeys. Indian J. Exp. Biol. 18, 73–75.
Shao, Y., Yu, Y., Li, C., Yu, J., Zong, R., and Pei, C. (2016). Synergistic effect of quercetin and 6-gingerol treatment in streptozotocin induced type 2 diabetic rats and poloxamer P-407 induced hyperlipidemia. RSC Adv. 6, 12235–12242. doi: 10.1039/C5RA16493A
Sharma, S., Kulkarni, S. K., and Chopra, K. (2006). Curcumin, the active principle of turmeric (Curcuma longa), ameliorates diabetic nephropathy in rats. Clin. Exp. Pharmacol. Physiol. 33, 940–945. doi: 10.1111/j.1440-1681.2006.04468.x
Sheela, C. G., and Augusti, K. T. (1992). Antidiabetic effects of S-allyl cysteine sulphoxide isolated from garlic Allium sativum Linn. Indian J. Exp. Biol. 30, 523–526.
Sheela, C. G., Kumud, K., and Augusti, K. T. (1995). Anti-diabetic effects of onion and garlic sulfoxide amino acids in rats. Planta Med. 61, 356–357. doi: 10.1055/s-2006-958099
Shibata, K., and Taguchi, H. (1991). Effect of dietary N'-methylnicotinamide or trigonelline on the growth and niacin metabolism in weanling rats. Vitamins 61, 493–499.
Singh, A., Ahmad, I., Akhter, S., Jain, G. K., Iqbal, Z., Talegaonkar, S., et al. (2013). Nanocarrier based formulation of thymoquinone improves oral delivery: stability assessment, in vitro and in vivo studies. Colloids Surf B Biointerfaces 102, 822–832. doi: 10.1016/j.colsurfb.2012.08.038
Singh, A. B., Singh, N., Maurya, R., and Srivastava, A. K. (2009). Anti-hyperglycaemic, lipid lowering and anti-oxidant properties of [6]-gingerol in db/db mice. Int. J. Med. Med. Sci. 1, 536–544.
Singh, A. B., Tamarkar, A. K., Narender, T., and Srivastava, A. K. (2010). Antihyperglycaemic effect of an unusual amino acid (4-hydroxyisoleucine) in C57BL/KsJ-db/db mice. Nat. Prod. Res. 24, 258–265. doi: 10.1080/14786410902836693
Singh, P., Jayaramaiah, R. H., Agawane, S. B., Vannuruswamy, G., Korwar, A. M., Anand, A., et al. (2016). Potential dual role of eugenol in inhibiting advanced glycation end products in diabetes: proteomic and mechanistic insights. Sci. Rep. 6:18798. doi: 10.1038/srep18798
Sober, H. A., Hollander, F., and Sober, E. K. (1950). Toxicity of eugenol determination of LD50 on rats. Exp. Biol. Med. 73, 148–151. doi: 10.3181/00379727-73-17608
Soetikno, V., Sari, F. R., Sukumaran, V., Lakshmanan, A. P., Harima, M., Thandavarayan, R. A., et al. (2012). Curcumin prevents diabetic cardiomyopathy in streptozotocin-induceddiabetic rats: possible involvement of PKC-MAPK signaling pathway. Eur. J. Pharm. Sci. 47, 604–614. doi: 10.1016/j.ejps.2012.04.018
Soetikno, V., Sari, F. R., Sukumaran, V., Lakshmanan, A. P., Mito, S., and Harima, M. (2013). Curcumin decreases renal triglyceride accumulation through AMPK-SREBP signalling pathway in streptozotocin-induced type 1 diabetic rats. J. Nutrion Biochem. 24, 796–802. doi: 10.1016/j.jnutbio.2012.04.013
Son, M. J., Miura, Y., and Yagasaki, K. (2015). Mechanisms for antidiabetic effect of gingerol in cultured cells and obese diabetic model mice. Cytotechnology 67, 641–652. doi: 10.1007/s10616-014-9730-3
Sporn, A., Dinu, I., and Stanciu, V. (1965). Investigation of the toxicity of cinnamic aldehyde. lgiena XIV, 339–345.
Srichamroen, A., Field, C. J., Thomson, A. B., and Basu, T. K. (2008). The modifying effects of galactomannan from Canadian-grown fenugreek (Trigonella foenum-graecum L.) on the glycemic and lipidemic status in rats. J. Clin. Biochem. Nutr. 43, 167–174. doi: 10.3164/jcbn.2008060
Srichamroen, A., Thomson, A. B., Field, C. J., and Basu, T. K. (2009). In vitro intestinal glucose uptake is inhibited by galactomannan from Canadian fenugreek seed (Trigonella foenum graecum L) in genetically lean and obese rats. Nutr. Res. 29, 49–54. doi: 10.1016/j.nutres.2008.11.002
Srinivasan, K. (2005). Plant foods in the management of diabetes mellitus: spices as beneficial antidiabetic food adjuncts. Int. J. Food Sci. Nutr. 56, 399–414. doi: 10.1080/09637480500512872
Srinivasan, S., Sathish, G., Jayanthi, M., Muthukumaran, J., Muruganathan, U., and Ramachandran, V. (2014). Ameliorating effect of eugenol on hyperglycemia by attenuating the key enzymes of glucose metabolism in streptozotocin-induced diabetic rats. Mol. Cell Biochem. 385, 159–168. doi: 10.1007/s11010-013-1824-2
Subash Babu, P., Prabuseenivasan, S., and Ignacimuthu, S. (2007). Cinnamaldehyde-A potential antidiabetic agent. Phytomedicine 14, 15–22. doi: 10.1016/j.phymed.2006.11.005
Subash Babu, P. S., Alshatwi, A. A., and Ignacimuthu, S. (2014). Beneficial antioxidative and antiperoxidative effect of cinnamaldehyde protect streptozotocin-induced pancreatic β-cells damage in Wister rats. Biomol. Ther. 22, 47–54. doi: 10.4062/biomolther.2013.100
Subramanian, S. P., and Prasath, G. S. (2014a). Antidiabetic and antidyslipidemic nature of trigonelline, a major alkaloid of fenugreek seeds studied in high-fat-fed and low-dose streptozotocin-induced experimental diabetic rats. Biomed. Prevt. Nutr. 4, 475–480. doi: 10.1016/j.bionut.2014.07.001
Subramanian, S. P., and Prasath, G. S. (2014b). Trigonelline improves insulin sensitivity and modulates glucose homeostasis in high fat fed-streptozotocin induced type 2 diabetic rats. J. Pharm. Res. 8, 563–569.
Suk, S., Seo, S. G., Yu, J. G., Yang, H., Jeong, E., Jang, Y. J., et al. (2016). A bioactive constituent of ginger, 6-shogaol, prevents adipogenesis and stimulates lipolysis in 3T3-L1 adipocytes. J. Food Biochem. 40, 84–90. doi: 10.1111/jfbc.12191
Suresh Babu, P. S., and Srinivasan, K. (1998). Amelioration of renal lesions associated with diabetes by dietary curcumin in experimental rats. Mol. Cell Biochem. 181, 87–96. doi: 10.1023/A:1006821828706
Suresh, D., and Srinivasan, K. (2007). Studies on the in vitro absorption of spice principles-curcumin, capsaicin and piperine in rat intestines. Food Chem. Toxicol. 45, 1437–1442. doi: 10.1016/j.fct.2007.02.002
Suryanarayana, P., Saraswat, M., Mrudul, T., Krishna, T. P., Krishnaswamy, K., and Reddy, G. B. (2005). Curcumin and turmeric delay streptozotocin-induced diabetic cataract in rats. Invest. Ophthalmol. Vis. Sci. 46, 2092–2099. doi: 10.1167/iovs.04-1304
Suryanarayana, P., Satyanarayana, A., Balakrishna, N., Kumar, P. U., and Reddy, G. B. (2007). Effect of turmeric and curcumin on oxidative stress and antioxidant enzymes in streptozotocin-induced diabetic rat. Med. Sci. Monit. 13, 286–292.
Takeda, S., Sato, N., Rakugi, H., and Morishita, R. (2011). Molecular mechanisms linking diabetes mellitus and Alzheimer disease: beta-amyloid peptide, insulin signaling, and neuronal function. Mol. Biosyst. 7, 1822–1827. doi: 10.1039/c0mb00302f
Tandan, R., Lewis, G. A., Krusinski, P. B., Badger, G. B., and Fries, T. J. (1992). Topical capsaicin in painful diabetic neuropathy. Diabetes Care. 15, 8–14. doi: 10.2337/diacare.15.1.8
Taylor, J. M., Jenner, P. M., and Jones, W. I. (1964). A comparison of the toxicityof some allyl, propenyl, and propyl compounds in the rat. Toxicol. Appl. Pharmacol. 6, 378–387. doi: 10.1016/S0041-008X(64)80002-8
Tharaheswari, M., Jayachandra Reddy, N., Kumar, R., Varshney, K. C., Kannan, M., and Rani, S. S. (2014). Trigonelline and diosgenin attenuate ER stress, oxidative stress mediated damage in pancreas and enhance adipose tissue PPARγ activity in type 2 diabetic rats. Mol. Cell Biochem. 396, 161–174. doi: 10.1007/s11010-014-2152-x
Tharaheswari, M., Nakkala, J. R., Praveen, S., Jeepipall, K., Raja, K., Chandra, V. K., et al. (2015). Fenugreek seed extract and its phytocompounds- trigonelline and diosgenin arbitrate their hepatoprotective effects through attenuation of endoplasmic reticulum stress and oxidative stress in type 2 diabetic rats. Eur. Food Res. Technol. 240, 223–232. doi: 10.1007/s00217-014-2322-9
Tong, J., Prigeon, R. L., Davis, H. W., Bidlingmaier, M., Kahn, S. E., Cummings, D. E., et al. (2010). Ghrelin suppresses glucose-stimulated insulin secretion and deteriorates glucose tolerance in healthy humans. Diabetes 59, 2145–2151. doi: 10.2337/db10-0504
Tundis, R., Menichini, F., Bonesi, M., Conforti, F., Statti, G., Menichini, F., et al. (2013). Antioxidant and hypoglycaemic activities and their relationship to phytochemicals in Capsicum annuum cultivars during fruit development. LWT-Food Sci. Technol. 53, 370–377. doi: 10.1016/j.lwt.2013.02.013
Tzeng, T. F., Chang, C. J., and Liu, I. M. (2014). 6-Gingerol inhibits rosiglitazone-induced adipogenesis in 3T3-L1 adipocytes. Phytother. Res. 28, 187–192. doi: 10.1002/ptr.4976
Tzeng, T. F., and Liu, I. M (2013). 6-Gingerol prevents adipogenesis and the accumulation of cytoplasmic lipid droplets in 3T3-L1 cells. Phytomedicine 20, 481–484. doi: 10.1016/j.phymed.2012.12.006
Uemura, T., Hirai, S., Mizoguchi, N., Goto, T., Lee, J. Y., Taketani, K., et al. (2010). Diosgenin present in fenugreek improves glucose metabolism by promoting adipocyte differentiation and inhibiting inflammation in adipose tissues. Mol. Nutr. Food Res. 54, 1596–1608. doi: 10.1002/mnfr.200900609
Upaganlawar, A. P., Badole, S. L., and Bodhankar, S. L. (2013). “Antidiabetic potential of trigonelline and 4-hydroxyisoleucine in fenugreek,” in Bioactive Food as Dietary Interventions for Diabetes, eds R. R.Watson and V. R. Preedy (San Diego, CA: Academic Press), 59–64.
Usharani, P., Mateen, A. A., Naidu, M. U., Raju, Y. S., and Chandra, N. (2008). Effect of NCB-02, atorvastatin and placebo on endothelial function, oxidative stress and inflammatory markers in patients with type 2 diabetes mellitus: a randomized, parallel-group, placebo-controlled, 8-week study. Drugs Res. Dev. 9, 243–250. doi: 10.2165/00126839-200809040-00004
Van Dijk, A. E., Olthof, M. R., Meeuse, J. C., Seebus, E., Heine, R. J., and van Dam, R. M. (2009). Acute effects of decaffeinated coffee and the major coffee components chlorogenic acid and trigonelline on glucose tolerance. Diabetes Care. 32, 1023–1025. doi: 10.2337/dc09-0207
Vijayakumar, R. S., Surya, D., and Nalini, N. (2004). Antioxidant efficacy of black pepper (Piper nigrum L.) and piperine in rats with high fat diet induced oxidative stress. Redox. Rep. 9, 105–110. doi: 10.1179/135100004225004742
Wang, Q., Ouazzani, J., Andre, S. N., and Potier, P. (2002). A practical synthesis of (2S,3R,4S)-4-hydroxyisoleucine, a potent insulinotropic α-amino acid from fenugreek. Eur. J. Org. Chem. 2002, 834–839. doi: 10.1002/1099-0690(200203)2002:5<834::AID-EJOC834>3.0.CO;2-6
Wang, S. L., Li, Y., Wen, Y., Chen, Y. F., Na, L. X, Li, S. T., et al. (2009). Curcumin, a potential inhibitor of up-regulation of TNF-alpha and IL-6 induced by palmitate in 3T3-L1 adipocytes through NF-kappaB and JNK pathway. Biomed. Environ. Sci. 22, 32–39. doi: 10.1016/S0895-3988(09)60019-2
Wani, K. D., Kadu, B. S., Mansara, P., Gupta, P., and Deore, A. V. (2014). Synthesis, characterization and in vitro study of biocompatible cinnamaldehyde functionalized magnetite nanoparticles (CPGF Nps) for hyperthermia and drug delivery applications in breast cancer. PLoS ONE 9:e107315. doi: 10.1371/journal.pone.0107315
Wei, C. K., Tsai, Y. H., Korinek, M., Hung, P. H., El-Shazly, M., Cheng, Y. B., et al. (2017). 6-Paradol and 6-shogaol, the pungent compounds of ginger, promote glucose utilization in adipocytes and myotubes, and 6-paradol reduces blood glucosein high-fat diet-fed mice. Int. J. Mol. Sci. 18, 1–18. doi: 10.3390/ijms18010168
Weibel, H., and Hansen, J. (1989). Interaction of cinnamaldehyde (a sensitizer in fragrance) with protein. Contact Dermatitis 20, 161–166.
Wongeakin, N., Sridulyakul, P., Jariyapongskul, A., Suksamrarn, A., and Patumraj, S. (2009). Effects of curcumin and tetrahydrocurcumin on diabetes induced endothelial dysfunction. Afr. J. Biochem. Res. 3, 259–265.
Xavier, S., Sadanandan, J., George, N., and Paulose, C. S. (2012). β2-Adrenoceptor and insulin receptor expression in the skeletal muscle of streptozotocin induced diabetic rats: antagonism by vitamin D3 and curcumin. Eur. J. Pharmacol. 687, 14–20. doi: 10.1016/j.ejphar.2012.02.050
Xie, X. Y., Kong, P. R., Wu, J. F., Li, Y., and Li, Y. X. (2012). Curcumin attenuates lipolysis stimulated by tumor necrosis factor-α or isoproterenol in 3T3-L1 adipocytes. Phytomedicine 20, 3–8. doi: 10.1016/j.phymed.2012.09.003
Yang, G., Zhong, L., Jiang, L., Geng, C., Cao, J., Sun, X., et al. (2010). Genotoxic effect of 6-gingerol on human hepatoma G2 cells. Chem. Biol. Interact. 185, 12–17. doi: 10.1016/j.cbi.2010.02.017
Yao, M., McClements, D. J., and Xiao, H. (2015). Improving oral bioavailability of nutraceuticals by engineered nanoparticle-based delivery systems. Curr. Opin. Food Sci. 2, 14–19. doi: 10.1016/j.cofs.2014.12.005
Yoshinari, O., and Igarashi, K. (2010). Anti-diabetic effect of trigonelline and nicotinic acid, on KK-Ay.mice. Curr. Med. Chem. 17, 2196–2202. doi: 10.2174/092986710791299902
Yoshinari, O., Sato, H., and Igarashi, K. (2009). Anti-diabetic effects of pumpkin and its components, trigonelline and nicotinic acid, on Goto-Kakizaki rats. Biosci. Biotechnol. Biochem. 73, 1033–1041. doi: 10.1271/bbb.80805
Yoshinari, O., Takenake, A., and Igarashi, K. (2013). Trigonelline ameliorates oxidative stress in type 2 diabetic Goto-Kakizaki rats. J. Med. Food 16, 34–41. doi: 10.1089/jmf.2012.2311
Younis, F., Mirelman, D., Rabinkov, A., and Rosenthal, T. (2010). S-Allyl-mercapto-captopril: a novel compound in the treatment of Cohen-Rosenthal diabetic hypertensive rats. J. Clin. Hypertens 12, 451–455. doi: 10.1111/j.1751-7176.2010.00270.x
Yuan, H. D., Huang, B., and Chung, S. H. (2011). Protective effect of cinnamaldehyde on streptozotocin-induced damage in rat pancreatic β-cells. Food Sci. Biotechnol. 20, 1271–1276. doi: 10.1007/s10068-011-0175-6
Yuan, J., Dieter, M. P., Bucher, J. R., and Jameson, C. W. (1993). Application of microencapsulation for toxicology studies. III. Bioavailability of microencapsulated cinnamaldehyde. Fundam. Appl. Toxicol. 20, 83–87. doi: 10.1006/faat.1993.1010
Yuan, J. H., Dieter, M. P., Bucher, J. R., and Jameson, C. W. (1992). Toxicokinetics of cinnamaldehyde in F344 rats. Food Chem. Toxicol. 30, 997–1004. doi: 10.1016/0278-6915(92)90109-X
Yuan, L. J., Qin, Y., Wang, L., Zeng, Y., Chang, H., Wang, J., et al. (2016). Capsaicin-containing chili improved postprandial hyperglycemia, hyperinsulinemia, and fasting lipid disorders in women with gestational diabetes mellitus and lowered the incidence of large-for-gestational-age newborns. Clin. Nutr. 35, 388–393. doi: 10.1016/j.clnu.2015.02.011
Yuyama, S. (1999). Absorption of trigonelline from the small intestine of the specific pathogen-free (SPF) and germ-free (GF) rats in vivo. Adv. Exp. Med. Biol. 467, 723–727. doi: 10.1007/978-1-4615-4709-9_94
Yuyama, S., and Kawano, Y. (1996). Urinary excretion of N1-methyl-2-pyridone-5- carboxylic acid and the fate of remaining of trigonelline. Adv. Exp. Med. Biol. 398, 599–603. doi: 10.1007/978-1-4613-0381-7_99
Yuyama, S., and Suzuki, T. (1991). The excretion of N1-methyl-2-pyridone-5-carboxylic acid and related compounds in human subjects after oral administration of nicotinic acid, trigonelline and N1-methyl-2-pyridone-5-carboxylic acid. Adv. Exp. Med. Biol. 294, 475–479. doi: 10.1007/978-1-4684-5952-4_48
Zaitsev, A. N., and Rakhmanina, N. L. (1974). Some data on the toxic properties of phenylethyl and cinnamyl alcohol derivatives. Voplosy. Pitaniia 5, 48–53.
Zhang, D. W., Fu, M., Gao, S. H., and Liu, J. L. (2013). Curcumin and diabetes: a systematic review. Evid. Based Complement. Alternat. Med. 2013:636053. doi: 10.1155/2013/636053
Zhang, W., Xu, Y. C., Guo, F. J., Meng, Y., and Li, M. L. (2008). Anti-diabetic effects of cinnamaldehyde and berberine and their impacts on retinol-binding protein 4 expression in rats with type 2 diabetes mellitus. Chin. Med. J. 121, 2124–2128.
Zhao, H., Xie, Y., Yang, Q., Cao, Y., Tu, H., and Cao, W. (2014). Pharmacokinetic study of cinnamaldehyde in rats by GC-MS after oral and intravenous administration. J. Pharm. Biomed. Anal. 89, 150–157. doi: 10.1016/j.jpba.2013.10.044
Zhao, H. Q., Qu, Y., Wang, X. Y., Lu, X. Y., Zhang, X. H., and Hattori, M. (2003). Determination of trigonelline by HPLC and study on its pharmacokinetics. Yao Xue Xue Bao. 38, 279–282. (in Chinese).
Zhao, W. C., Zhang, B., Liao, M. J., Zhang, W. X., He, W. Y., Wang, H. B., et al. (2014). Curcumin ameliorated diabetic neuropathy partially by inhibition of NADPH oxidase mediating oxidative stress in the spinal cord. Neurosci. Lett. 560, 81–85. doi: 10.1016/j.neulet.2013.12.019
Zheng, C. J., Hu, H., Cao, H., and Li, J. (2014). Effect of JNK/MCP-1 signalling pathway on anti-diabetic neuropathic pain by curcumin in type 2 diabetic rats. Chin. J. Pathophysiol. 30, 1941–1945. doi: 10.3969/j.issn.1000-4718.2014.11.004.
Zheng, S., and Chen, A. (2004). Activation of PPARγ is required for curcumin to induce apoptosis and to inhibit the expression of extracellular matrix genes in hepatic stellate cells in vitro. Biochem. J. 384, 149–157. doi: 10.1042/BJ20040928
Zhou, J. Y., Zhou, S., and Zeng, S. (2011). Experimental diabetes treated with trigonelline: effect on β cell and pancreatic oxidative parameters. Fundam. Clin. Pharmacol. 27, 279–287. doi: 10.1111/j.1472-8206.2011.01022.x
Zhou, J. Y., and Zhou, S. W. (2012). Protection of trigonelline on experimental diabetic peripheral neuropathy. Evid. Based Complement. Altern. Med. 2012:164219. doi: 10.1155/2012/164219
Zick, S. M., Djuric, Z., Ruffin, M. T., Litzinger, A. J., Normolle, D. P., Alrawi, S., et al. (2008). Pharmacokinetics of 6-gingerol, 8-gingerol, 10-gingerol, and 6-shogaol and conjugate metabolites in healthy human subjects. Cancer Epidemiol. Biomark. Prev. 17, 1930–1936. doi: 10.1158/1055-9965.EPI-07-2934
Keywords: adjuvant, diabetes mellitus, hypoglycemic, in vitro, in vivo, spices
Citation: Mohammed A and Islam MS (2018) Spice-Derived Bioactive Ingredients: Potential Agents or Food Adjuvant in the Management of Diabetes Mellitus. Front. Pharmacol. 9:893. doi: 10.3389/fphar.2018.00893
Received: 23 February 2018; Accepted: 23 July 2018;
Published: 22 August 2018.
Edited by:
Adolfo Andrade-Cetto, Universidad Nacional Autónoma de México, MexicoReviewed by:
Marselina Irasonia Tan, Bandung Institute of Technology, IndonesiaCopyright © 2018 Mohammed and Islam. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Md. Shahidul Islam, aXNsYW1kQHVrem4uYWMuemE=; c2lzbGFtMTk3NEB5YWhvby5jb20=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.