 Rebeca Diez-Alarcia1,2Inés Ibarra-Lecue1
Rebeca Diez-Alarcia1,2Inés Ibarra-Lecue1 Ángela P. Lopez-Cardona3,4Javier Meana1,2
Ángela P. Lopez-Cardona3,4Javier Meana1,2 Alfonso Gutierrez-Adán3
Alfonso Gutierrez-Adán3 Luis F. Callado1,2Ekaitz Agirregoitia5
Luis F. Callado1,2Ekaitz Agirregoitia5 Leyre Urigüen1,2*
Leyre Urigüen1,2*- 1Department of Pharmacology, University of the Basque Country UPV/EHU, Leioa, Spain
- 2Centro de Investigación Biomédica en Red de Salud Mental, Madrid, Spain
- 3Department of Animal Reproduction, Instituto Nacional de Tecnología Agraria y Alimentaria, Madrid, Spain
- 4G.I. Biogénesis, Universidad de Antioquia, Antioquia, Colombia
- 5Department of Physiology, University of the Basque Country, Leioa, Spain
Cannabinoid receptors are able to couple to different families of G proteins when activated by an agonist drug. It has been suggested that different intracellular responses may be activated depending on the ligand. The goal of the present study was to characterize the pattern of G protein subunit stimulation triggered by three different cannabinoid ligands, Δ9-THC, WIN55212-2, and ACEA in mouse brain cortex. Stimulation of the [35S]GTPγS binding coupled to specific immunoprecipitation with antibodies against different subtypes of G proteins (Gαi1, Gαi2, Gαi3, Gαo, Gαz, Gαs, Gαq/11, and Gα12/13), in the presence of Δ9-THC, WIN55212-2 and ACEA (submaximal concentration 10 μM) was determined by scintillation proximity assay (SPA) technique in mouse cortex of wild type, CB1 knock-out, CB2 knock-out and CB1/CB2 double knock-out mice. Results show that, in mouse brain cortex, cannabinoid agonists are able to significantly stimulate not only the classical inhibitory Gαi/o subunits but also other G subunits like Gαz, Gαq/11, and Gα12/13. Moreover, the specific pattern of G protein subunit activation is different depending on the ligand. In conclusion, our results demonstrate that, in mice brain native tissue, different exogenous cannabinoid ligands are able to selectively activate different inhibitory and non-inhibitory Gα protein subtypes, through the activation of CB1 and/or CB2 receptors. Results of the present study may help to understand the specific molecular pathways involved in the pharmacological effects of cannabinoid-derived drugs.
Introduction
During the last decade a wide number of studies have focused on the potential involvement of the endocannabinoid system in a variety of psychiatric and neurological disorders. The putative psychoactive ingredient of Cannabis sativa (marijuana plant), Δ9-tetrahydrocannabinol (Δ9-THC), as well as the endogenous cannabinoids anandamide (arachidonoyl ethanolamide) and 2-arachidonoylglycerol (2-AG) act primarily through cannabinoid CB1 and CB2 receptors. These cannabinoid receptors are GPCRs mostly coupled to Gi/o proteins (Howlett et al., 2002). The CB1 receptor is mainly distributed in the CNS, particularly in cortex, basal ganglia, hippocampus, and cerebellum (Mackie, 2005; De Jesus et al., 2006) and generally acts presinaptically inhibiting the release of neurotransmitters. CB2 receptors are expressed at much lower levels in the CNS compared with CB1 receptors (reviewed in Atwood and Mackie, 2010). As Gi/o coupled GPCRs, CB1 and CB2 receptors inhibit adenylyl cyclase, but moreover, both receptors are able to activate MAPK, inhibit voltage gated Ca2+ channels and activate inwardly rectifying K+ channels (Childers et al., 1993).
The activation of CB1 receptor in the brain leads to the modulation of neuronal excitability, which may be in part responsible of the psychoactive effects of exogenous cannabinoids. In this context, a considerable amount of studies have been performed in order to elucidate the effects of cannabinoids (natural or synthetics) in the development of mental alterations, such as addiction, cognitive deficits, anxiety or psychosis. Importantly, different or opposite behavioral effects have been observed after the administration of Δ9-THC or synthetic cannabinoid ligands (Fattore et al., 2003; Panagis et al., 2014; Rubino and Parolaro, 2016). It has been demonstrated that for most G protein-coupled receptors, distinct agonists can differentially regulate several signaling pathways through the same receptor by a selective activation of different intracellular effectors. This is a mechanism known as functional selectivity or biased agonism. In this way, cannabinoid receptors have been demonstrated to be capable of coupling to different families of G proteins and/or to beta-arrestin when activated by an agonist drug suggesting that different intracellular responses may be activated depending on the ligand (Glass and Northup, 1999; Bosier et al., 2010). For instance, for the CB1 receptor has been reported that, whereas 2-AG and WIN55,212 have little preference for inhibition of cAMP and phosphorylation of ERK1/2, anandamide and CP55940 were biased toward cAMP inhibition (Khajehali et al., 2015). Moreover, in a recent study Dhopeshwarkar and Mackie (2016) demonstrated that CB2 receptor ligands display strong and varied functional selectivity at canonical (inhibition of adenylyl cyclase) and non-canonical (arrestin recruitment) pathways. Moreover, the intracellular signaling activated by a receptor depends on the cellular system where it is expressed, which may vary across different neuronal environments. In this context, it has been demonstrated that opioid and cannabinoid receptors function through the same pool of G proteins when they are co-transfected, whereas in cells endogenously expressing these receptors signaling occurs through distinct pools of G proteins (Shapira et al., 2000). Thus, this fact should be taken into consideration when interpreting results acquired in artificially transfected cells vs. native biological systems.
To our knowledge, no study has compared G protein signaling by different cannabinoid drugs in native brain tissue. Thus, in the current study, we performed [35S]GTPγS scintillation proximity assay (SPAs) coupled with the use of specific antibodies against different Gα protein subunits to evaluate the functional selectivity of different cannabinoid ligands by activating CB1 and/or CB2 cannabinoid receptors in mouse brain cortex.
Materials and Methods
Animal Procedures
Adult C57BL/6J (WT), CB1 knock-out (CB1-/-) (Marsicano et al., 2002), CB2 knock-out (CB2-/-) (Buckley et al., 2000), and CB1/CB2 double knock-out (CB1-/-/CB2-/-) mice were used in this study. Animals (males, aged 7–8 weeks-old) were housed (6–8 animals per cage) in standard cages under controlled conditions of temperature (23 + 1°C) and photoperiod (light/dark cycle 14 h: 10 h) and free access to standard rodent chow and water.
Animal Welfare and Ethical Statements
All experimental procedures using mice were performed in accordance with the European Directive for the Protection of Vertebrate Animals used for experimental and Other Scientific Purposes (European Union Directive #86/606/EEC) and approved by the Ethics Committees for Animal Welfare of the University of the Basque Country (UPV/EHU) permit number CEBA1882011 and by the Institutional Review Board (INIA), permit number CEEA2012/021.
Rationale for Choice of Cannabinoid Ligands
In the present study, we decided to investigate the effects of three different cannabinoid ligands. Δ9-THC was chosen for being the main psychoactive component of marijuana plant and the putative responsible of the development of mental disorders in humans. WIN55212-2, a synthetic cannabinoid structurally different from Δ9-THC, is a potent, non-selective CB1/CB2 receptor agonist that is frequently used in the studies that try to elucidate the effects of Cannabis in the brain. Finally, we wanted to study a ligand structurally similar to endogenous cannabinoids, such as the synthetic anandamide analog arachidonyl-2-chloroethylamide (ACEA). O-2050 was chosen as a neutral cannabinoid antagonist. O-2050 has been proved to be a neutral CB1 receptor antagonist in several studies (Canals and Milligan, 2008; Hudson et al., 2010; Brents et al., 2011; Wiley et al., 2011), with quite similar affinity for CB1 and CB2 receptors. Although, there is some data in the literature suggesting its activity as inverse or even partial agonist at CB1 receptors in various tissues (Makwana et al., 2010; Wiley et al., 2011) in a previous work of our group, we showed that O-2050 has no effect over [35S]GTPγS binding and behaves as an antagonist blocking WIN55212-2-mediated activation (Erdozain et al., 2012) (Figure 1).
FIGURE 1. Chemical structure of the different cannabinoid ligands used. Representative scheme of the chemical structure of the different cannabinoid ligands used in this study (Δ9-tetrahydrocannabinol (Δ9-THC), WIN55212-2, arachidonyl-2-chloroethylamide (ACEA) and O-2050).
Rationale for Choice of G Protein α Subunit Subtypes
In the present study, we decided to investigate the ability of cannabinoid receptors to activate different G proteins subtype. We chose at least one G protein subtype representative of each main G protein family and mainly focusing in the inhibitory G proteins for being the cannabinoid canonical pathway (Figure 2).
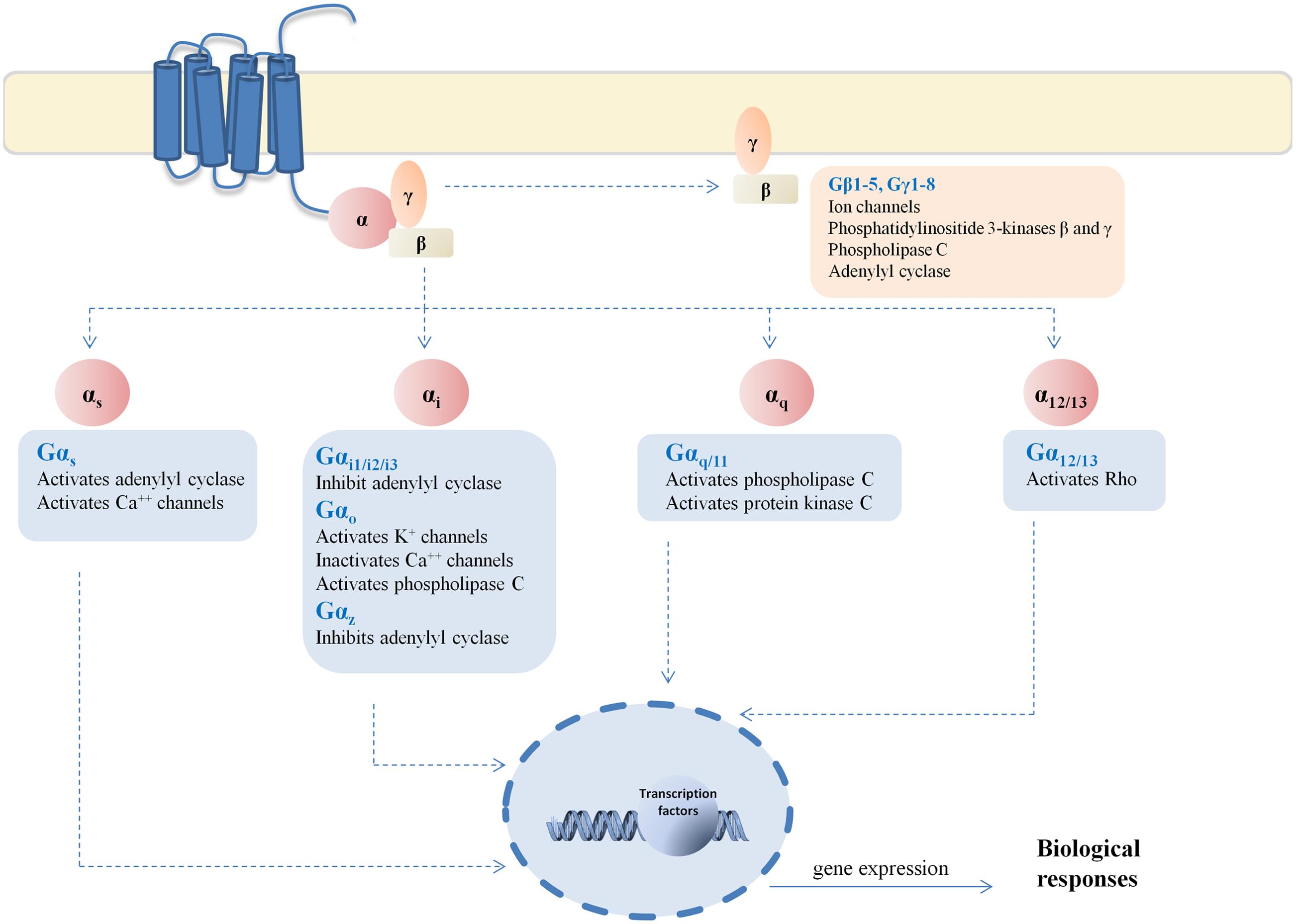
FIGURE 2. G protein signaling pathways. Representative scheme of the signaling pathways linked to each G protein evaluated in this study.
Mouse Brain Cortex Membrane Homogenates Preparation
After sacrifice by cervical dislocation, the brains were rapidly removed, cortices dissected and fresh frozen, and stored immediately at -80°C until use. Preparation of membrane enriched fraction (P2 fraction) was performed as previously described (Gonzalez-Maeso et al., 2000) with minor modifications. Mouse brain cortex samples (approximately 200 mg) from seven mice each time were thawed at 4°C and homogenized with a glass/teflon grinder (IKA labortechnik, Satufen, Germany) (10 strokes at maximum speed) in 30 volumes of homogenization buffer (50 mM Tris-HCl (Invitrogen, Barcelona, Spain), 1 mM EGTA (Sigma-Aldrich, St. Louis, MO, USA), 3 mM MgCl2 (Sigma-Aldrich, St. Louis, MO, USA) and 1 mM DTT (Invitrogen, Barcelona, Spain); pH 7.4; supplemented with 250 mM sucrose (Panreac Química S.A.U, Barcelona, Spain). The homogenates were centrifuged at 1,100 × g for 10 min at 4°C (Sorvall RC-5C centrifuge, SM-24 rotor; FisherScientific, Madrid, Spain). The pellets (P1 fraction) were discarded and the supernatants were then recentrifuged at 40,000 × g for 10 min (4°C). The resultant pellets were resuspended in 20 volumes of fresh cold centrifugation buffer (50 mM Tris-HCl, 1 mM EGTA, 3 mM MgCl2 and 1 mM DTT; pH 7.4) with a glass stick and recentrifuged at 40,000 × g for 10 min (4°C). The obtained pellets were then resuspended in five volumes of centrifugation buffer. Protein content was determined by the method of Bradford with BSA (Sigma-Aldrich®, St. Louis, MO, USA) as standard. Linear regression analysis and extrapolation of the data were carried out with GraphPad Prism 5® software (GraphPad Software, Inc., San Diego, CA, USA). Finally, aliquots of 0.5, 1, and 2 mg were then centrifuged at 21,000 × g (Eppendorf 5810R centrifuge; Eppendorf, Madrid, Spain) during 15 min at 4°C. The supernatant layer was carefully discarded and the pellets stored at -80°C until assay.
Antibody-Capture [35S]GTPγS Scintillation Proximity Assay (SPA)
Specific activation of different subtypes of Gα proteins was determined using a homogeneous protocol of [35S]GTPγS SPA coupled with the use of specific antibodies essentially as previously described (Erdozain et al., 2012). [35S]GTPγS binding was performed in 96-well Isoplates (PerkinElmer Life Sciences, Maanstraat, Germany) and in a final volume of 200 ml containing 1 mM EGTA, 3 mM MgCl2, 100 mM NaCl, 0.2 mM DTT, 50 mM Tris-HCl at pH 7.4, 0.4 nM [35S]GTPγS, 15 μg of protein per well, and different concentrations of GDP depending on the Gα subunit subtype tested. At the end of the 2 h incubation period (30°C), 20 μl of Igepal 1% + SDS 0.1% were added to each well, and plates were incubated at 22°C for 30 min with gentle agitation. Specific antibody for the Gα subunit of interest was then added to each well before an additional 90 min RT incubation period (the antibodies and dilutions employed are described in Table 1). Polyvinyltoluene (PVT) SPA beads coated with protein A (PerkinElmer, S.L., Tres Cantos, Madrid, Spain) were then added (0.75 mg of beads per well), and plates were incubated for 3 h at RT with gentle agitation. Finally, plates were centrifuged (5 min at 1000 × g), and bound radioactivity was detected on a MicroBeta TriLux scintillation counter (PerkinElmer S.L., Tres Cantos, Madrid, Spain). In order to test their effect on the [35S]GTPγS binding to the different Gα subunit subtypes, a single submaximal concentration of the drugs (10 μM) Δ9-THC, WIN55212-2, ACEA and/or O-2050, was used. This submaximal concentration was chosen as previously reported (Erdozain et al., 2012) in our previous experimental assays in which we established the standard conditions for this assays. This concentration is the one which give us binding values around the Emax for any drug and subunit subtype combination studied (Supplementary Figure S1). Non-specific binding was defined as the remaining [35S]GTPγS binding in the presence of 10 μM unlabelled GTPγS.
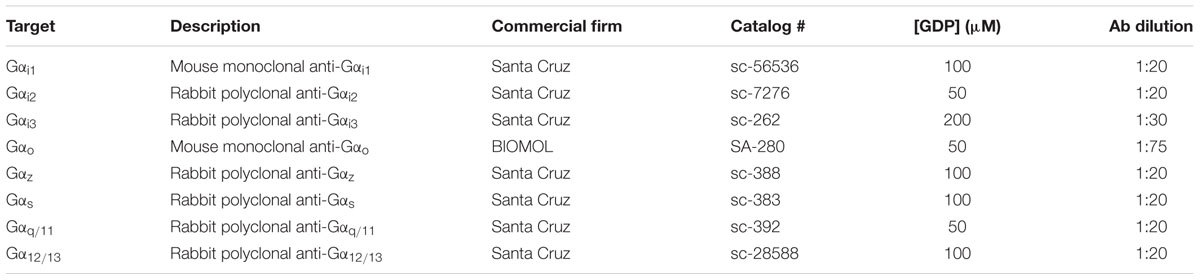
TABLE 1. Antibodies, dilutions, and GDP concentrations employed in the [35S]GTPγS scintillation proximity assays.
Western Blot
For Western blot experiments, membrane enriched fraction (P2 fraction) pellets from mouse brain tissue (cortex) were resuspended in TBS, reaching a concentration of 4 mg protein/ml. Commercial Laemmli 2x (95%) and β-mercaptoethanol (5%) (Sigma-Aldrich®, St. Louis, MO, USA) were added to each sample, reaching a final protein concentration of 2 mg/ml. Finally, all the samples were heated at 95°C for 5 min in a Thermoblock (Biometra, Goettingen, Germany) and kept at -20°C until assay. Electrophoresis was carried out in SDS polyacrylamide gels, composed of 5% stacking (0.5 M Tris-HCl, pH 6.8, 10% SDS) and 12% resolving (1.5 M Tris-HCl, pH 8.8, 10% SDS) gels, using a miniprotean system (Bio-Rad Laboratories). Equal protein loading in the gel was verified by simultaneous immunodetection of β-actin (mouse monoclonal antibody anti-β-actin, Sigma Biosciences, St. Louis, MO, USA) with the different Gα subunit subtypes. Proteins were then transferred to nitrocellulose membranes (1 h, 0.3 A) using an electrophoretic transfer system (Bio-Rad Laboratories). The non-specific binding sites in the membranes were blocked for 1 h at RT in blocking solution (3% non-fat dry milk, pH = 7.4 in PBS). Membranes were incubated overnight at 4°C in incubation buffer (3% non-fat dry milk + 0.1% Tween-20 in PBS) containing the appropriate dilution of the specific primary anti-Gα subunit antibody. Antibody specificity, as previously described in the literature (Gettys et al., 1994; Valdizan et al., 2010), was confirmed in our experimental conditions by Western blot (data not shown). Membranes were washed with PBS and incubated for 1 h at RT and constant agitation with the fluorescent conjugated secondary antibodies (Alexa Fluor® 680 and/or IRDye 800 conjugated antibodies) suitable diluted in incubation buffer. Finally, membranes were rewashed with PBS and immunoreactivity was detected and quantified using the Odyssey Infrared Imaging System (LI-COR Biosciences, Lincoln, NE, USA) and Odyssey Software. Broad-Range pre-stained SDS-PAGE molecular weight standard (Bio-Rad Laboratories, Hercules, CA, USA) was used.
Data Analysis and Statistical Procedures
Data were analyzed with GraphPad PrismTM 5.01 software. In order to allow better interpretation of the data, specific binding obtained from [35S]GTPγS SPAs were transformed to percentage of basal binding (binding values observed in the absence of any exogenous drug) obtained for each Gα protein subunit studied. The statistical comparison of the SPA results was carried out by a two-tailed one sample Student’s t-test with a significance level of p < 0.05. Immunodensity data obtained from Western blotting assays were transformed to percentage of the control, being the control the mean of immunodensities obtained for WT mice. The statistical comparison of the Western blot results was carried out by a one-way ANOVA test, followed by Dunnet’s post hoc test for multiple comparisons, with a significance level of p < 0.05. All data are expressed as mean ± SEM values.
Materials
[35S]GTPγS (4625 × 1010 Bq/mmol) was purchased from PerkinElmer Life Sciences (Maanstraat, Germany). Tetrahydrocannabinol (Δ9-THC) was purchased from THCPharm GmbH (Frankfurt, Germany); WIN55212-2 and GTPγS were purchased from Sigma-Aldrich (St. Louis, MO, USA); Arachidonyl-2-chloroethylamide ACEA and O-2050 were from Tocris Bioscience (Bristol, UK). All other chemical reagents were of analytical quality and were purchased from Merck (Darmstadt, Germany) or Sigma-Aldrich (St. Louis, MO, USA).
Results
Effects of Δ9-THC, WIN55212-2, and ACEA on G Protein Activation in Mouse Brain Membranes
Cannabinoid receptor ligands were used for the characterization of the functional coupling of cannabinoid receptors to the different G protein α subunit subtypes (Gαi1, Gαi2, Gαi3, Gαo, Gαz, Gαs, Gαq/11, and Gα12/13) in mouse brain tissue. First, we investigated which Gα subunit subtypes were activated by the natural cannabinoid Δ9-THC (10 μM) in mouse brain cortex membrane homogenates (Figure 3A). As expected, we found that Δ9-THC was able to significantly activate several classical AC inhibitory subunits, as Gαi1 (113 ± 3%), Gαo (110 ± 2%), and Gαz (113 ± 5%), while exerted no effect on Gαi2 or Gαi3. Δ9-THC was also able to activate the Gαq/11 subunit (118 ± 5%). However, no changes were observed when we studied AC stimulatory subunit Gαs and the RhoA activator Gα12/13 subunit. To further test if these effects of Δ9-THC were cannabinoid-receptor mediated, the same assays were carried out in the presence of a putative neutral antagonist of the CB1 receptor, O-2050. In all cases, the activation of these G protein subunits was blocked when membranes were co-incubated with the cannabinoid antagonist O-2050. Next, we investigated the effects on G protein subunit activation induced by the synthetic cannabinoid agonist WIN55212-2 (Figure 3B). We found that WIN55212-2 significantly increased the binding of [35S]GTPγS to the all the inhibitory subunits Gαi1 (129 ± 6%), Gαi3 (129 ± 5%), Gαo (120 ± 4%) and Gαz (134 ± 6%), except Gαi2. WIN55212-2 was also able to activate the Gαq/11 subunit (131 ± 7%), but not the AC stimulatory subunit Gαs. Surprisingly, WIN55212-2 was also able to significantly stimulate the RhoA activator Gα12/13 (130 ± 4%). In the same way as previously described for Δ9-THC, the activation of these G protein subunits by WIN55212-2 was always blocked by the co-incubation with the cannabinoid antagonist O-2050, except for the case of Gαz (106 ± 1%). Finally, we investigated the effect of the synthetic anandamide analog ACEA on G protein subunit activation in mouse brain tissue (Figure 3C). When evaluating the classical AC inhibitory subunits, we found that ACEA stimulated Gαi1 (121 ± 4%), Gαi3 (120 ± 5%), and Gαo (116 ± 4%). However, as occurred with Δ9-THC and WIN55212-2, no stimulation was observed in Gαi2, suggesting that none of the cannabinoids evaluated exert their effects through Gαi2 signaling. Moreover, ACEA had no effect on Gαz. As previously observed for the other two cannabinoid ligands evaluated, ACEA also activated Gαq/11 subunit (122 ± 7%) while had no effect on Gαs. Thus, it seems that none of these cannabinoid ligands are able to activate this AC stimulatory subunit either. No changes were observed when we studied the effects of ACEA on the RhoA activator Gα12/13 subunit. Again, the activation of these G protein subunits was blocked when membranes were co-incubated with the cannabinoid antagonist O-2050.
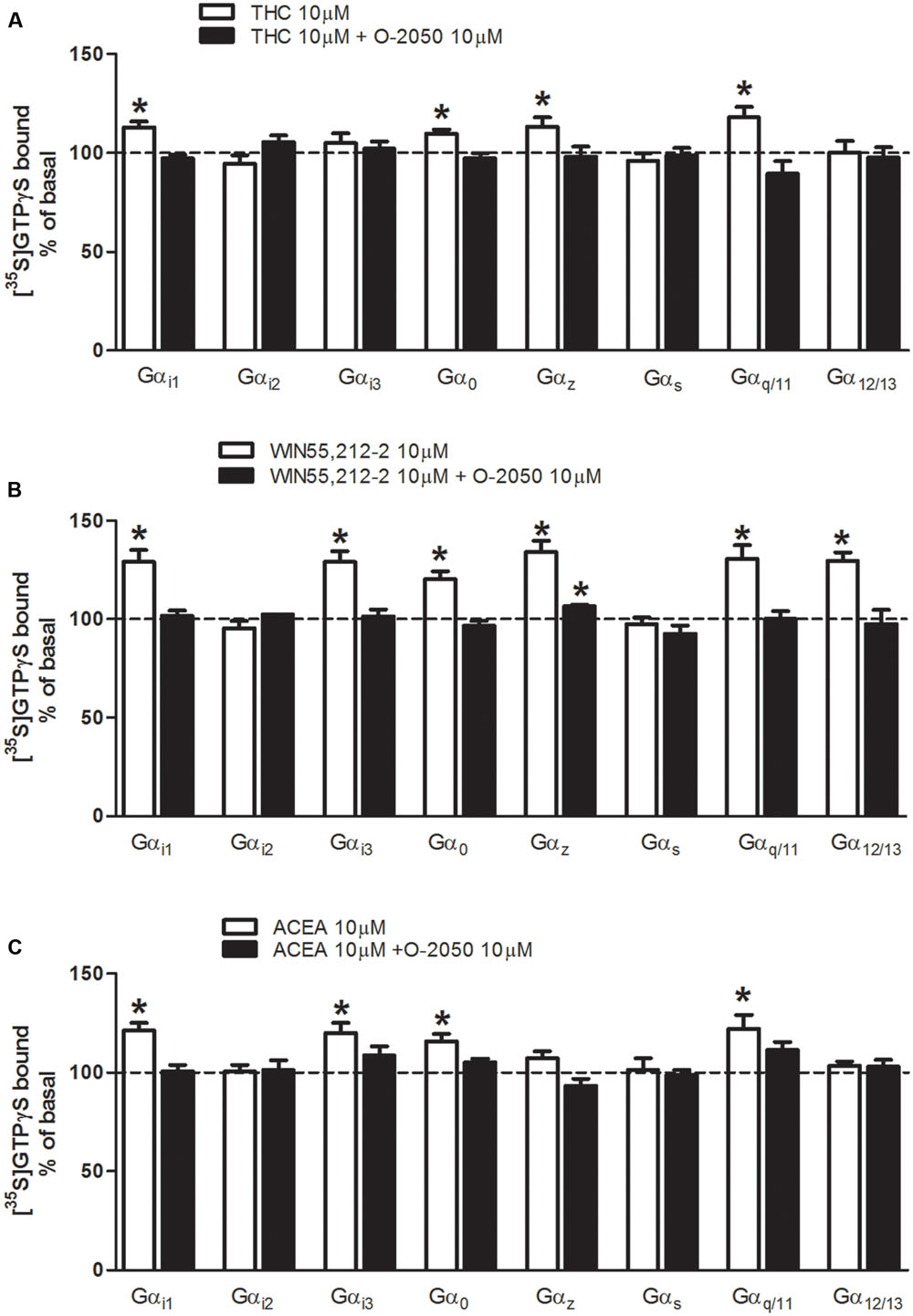
FIGURE 3. Effects of THC, WIN55212-2, and ACEA on G protein activation in mouse brain membranes. [35S]GTPγS scintillation proximity assays coupled to immunoprecipitation with specific antibodies against different Gα subunits (Gαi1, Gαi2, Gαi3, Gαo, Gαz, Gαs, Gαq/11, and Gα12/13) in mouse brain cortical membranes co-incubated with (A) THC (10 μM) (B) WIN55212-2 (10 μM), or (C) ACEA (10 μM) in the presence or absence of the antagonist O-2050 (10 μM). Data are shown as percentage of [35S]GTPγS basal binding values obtained for each specific subunit. Bars represent mean ± SEM of four to six different experiments carried out in triplicate. Asterisks highlight those normalized values of stimulation or inhibition of basal binding which are statistically different from 100% (Student’s t-test; ∗p < 0.05).
Effects of the Cannabinoid Antagonist O-2050 on G Protein Activation in Mouse Brain Membrane Homogenates
O-2050 was initially synthesized and described as a neutral CB1 receptor antagonist, however, there are some evidences suggesting that is able to act as an inverse agonist or even as a partial agonist (Wiley et al., 2011). For this reason, and in order to validate O-2050 as a useful pharmacological tool to antagonize the effect mediated by cannabinoid receptors, [35S]GTPγS SPAs were performed in mouse cortical membranes in the presence of O-2050 (10 μM) alone. Under these experimental conditions, neither stimulation nor inhibition of [35S]GTPγS basal binding values were observed for any of the Gα subunit subtypes studied, with the exception of Gαz (119 ± 1%) (Figure 4).
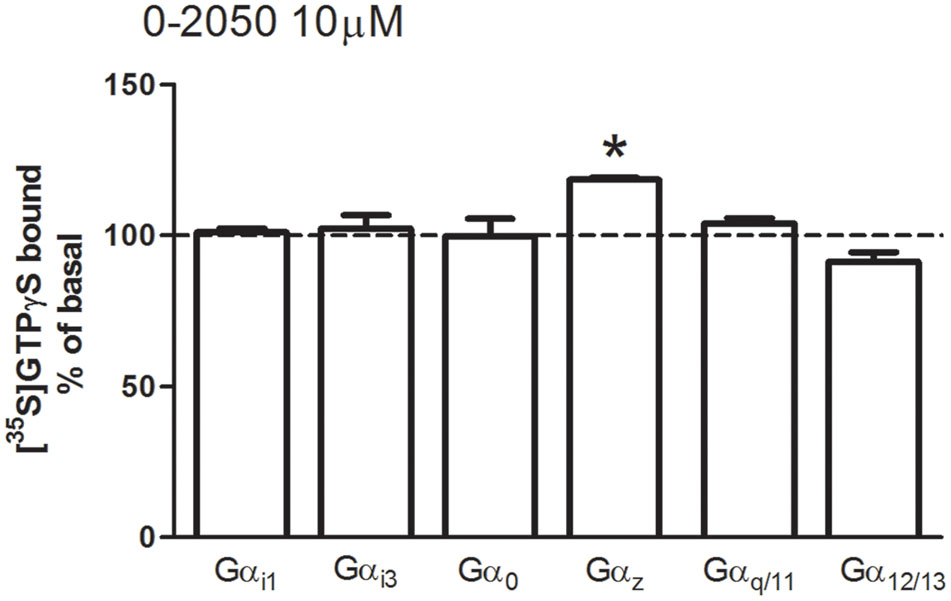
FIGURE 4. Effects of the cannabinoid antagonist O-2050 on G protein activation in mouse brain membranes. [35S]GTPγS scintillation proximity assays coupled to immunoprecipitation with anti-Gαi1, Gαi3, Gαo, Gαz, Gαq/11, and Gα12/13 specific antibodies in mouse brain cortical membranes co-incubated with the cannabinoid antagonist O-2050 (10 μM). Data are shown as percentage of [35S]GTPγS basal binding values obtained for each specific subunit. Bars represent mean ± SEM of four to six different experiments carried out in triplicate. Asterisks highlight those normalized values of stimulation or inhibition of basal binding which are statistically different from 100% (Student’s t-test; ∗p < 0.05).
Effects of Δ9-THC, WIN55212-2, and ACEA on G Protein Activation in Cannabinoid Receptors Knockout Mice
To further elucidate the role of each cannabinoid receptor subtype in the agonist-mediated activation of the different Gα subunit subtypes, [35S]GTPγS SPA was performed in brain tissue of CB1-/-, CB2-/-, and CB1-/-/CB2-/- mice. For that purpose, brain membranes were incubated with the different cannabinoid ligands (THC, WIN55212-2, or ACEA) and with the specific antibodies against that Gα for which an stimulation with these agonists was observed in WT. Figure 5A shows the stimulation of the different Gα subunits when brain membranes of the four genotypes were incubated with Δ9-THC. The significant stimulation of the inhibitory Gαi1 subunit observed in the WT mice was completely absent in the CB1-/- and in the CB1/CB2 double ko mice, but was still present in the CB2-/- (108 ± 2%), which suggests that the Δ9-THC-mediated stimulation of Gαi1 is induced by the activation of CB1 receptor. On the other hand, opposite results were obtained for Gαio and Gαz, the other two inhibitory subunits that were stimulated by Δ9-THC. As previously described, there was a significant Δ9-THC-induced stimulation of Gαo and Gαz subunits in the WT. This stimulation was also observed in the CB1-/- (115 ± 4% for Gαo and 120 ± 4% for Gαz) but not in the CB2-/- or the CB1-/-/CB2-/- mice. These data may indicate that, in mouse brain cortical membranes, Δ9-THC acts through the CB2 receptor to stimulate these inhibitory Gαo and Gαz subunits. Finally, the Δ9-THC-induced activation of the Gαq/11 subunit observed in the WT mice was not found in the CB1-/- and CB1-/-/CB2-/- mice, while remained unchanged in the CB2-/- (116 ± 2%). This result indicates that Δ9-THC stimulates the Gαq/11 subunit acting mainly through the CB1 receptor.
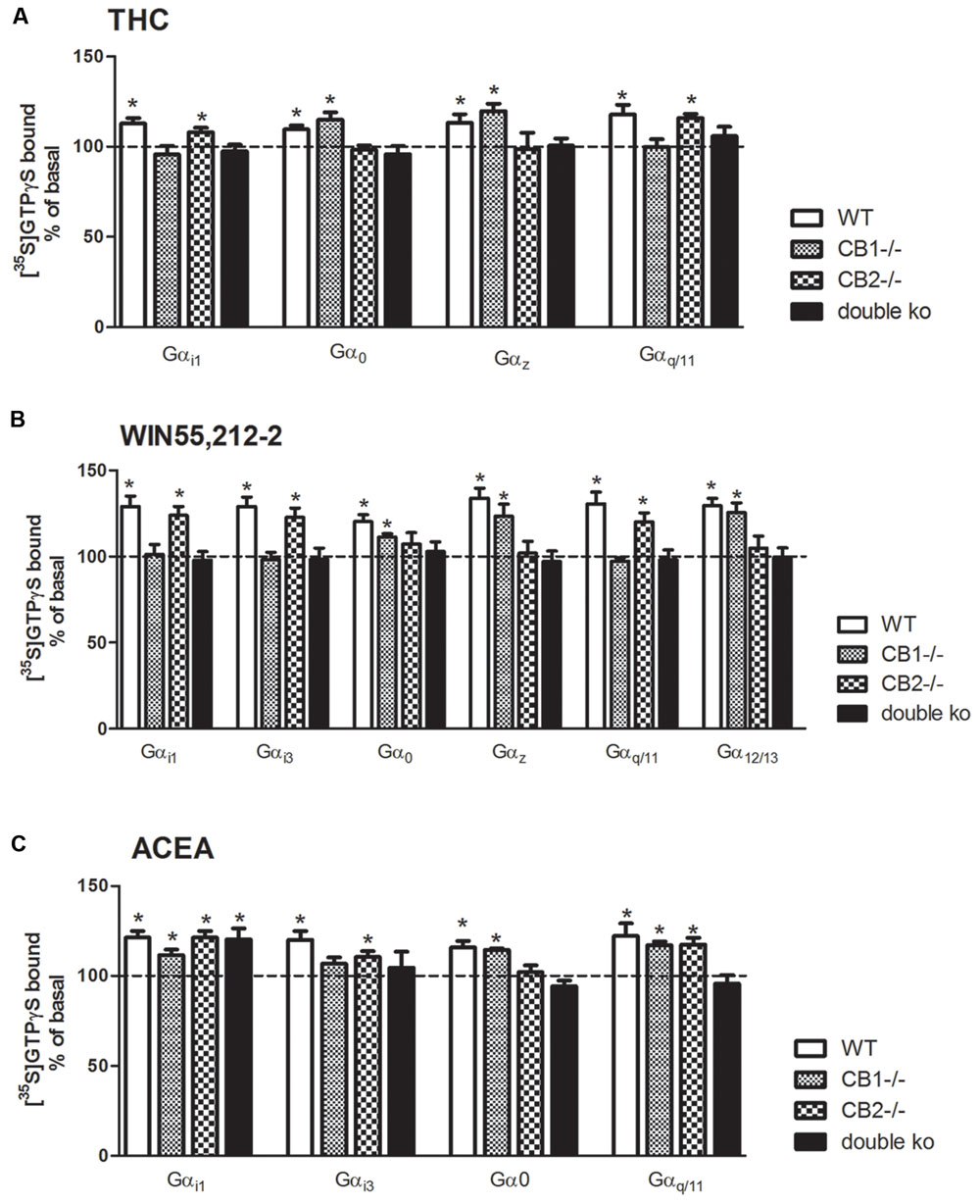
FIGURE 5. Effects of THC, WIN55, 212-2, and ACEA on G protein activation in cannabinoid receptors knockout mice. [35S]GTPγS scintillation proximity assays coupled to immunoprecipitation with specific antibodies against different Gα subunits in mouse brain cortical membranes of WT, CB1 knockout (CB1-/-), CB2 knockout (CB2-/-) and double CB1 and CB2 knockout (CB1-/-/CB2-/-/double KO) co-incubated with (A) THC (10 μM) (B) WIN55212-2 (10 μM), or (C) ACEA (10 μM). Data are shown as percentage of [35S]GTPγS basal binding values obtained for each specific subunit. Bars represent mean ± SEM of four to six different experiments carried out in triplicate. Asterisks highlight those normalized values of stimulation or inhibition of basal binding which are statistically different from 100% (Student’s t-test; ∗p < 0.05).
Figure 5B shows the stimulation of different Gα subunits when brain membranes of the four genotypes were incubated with the synthetic cannabinoid agonist WIN55212-2. The significant stimulation of the inhibitory Gαi1 and Gαi3 subunits induced by WIN55212-2 in the WT mice was not found in the CB1-/- nor in the CB1-/-/CB2-/- mice, but was still present in the CB2-/- (124 ± 5% for Gαi1 and 123 ± 5% for Gαi3). On the contrary, the inhibitory subunits Gαo and Gαz, which were significantly stimulated in the WT mice, remained stimulated in the CB1-/- (111 ± 2% for Gαo and 123 ± 7% for Gαz) but not stimulation was found in the CB2-/- nor in the CB1-/-/CB2-/- mice. These results suggest that the inhibitory signaling of WIN55212-2 in the mice brain through Gαi1 and Gαi3 activation seems to be mediated by the CB1 receptor, while the stimulation of Gαo and Gαz would be mediated by the CB2 receptor activation. The significant activation of the Gαq/11 subunit induced by WIN55212-2 in the WT mice was completely absent in the CB1-/- mice, as well as in the CB1/CB2 double ko mice. On the contrary, a significant stimulation of Gαq/11 subunit (120 ± 5%) was observed in the CB2-/- membranes, suggesting that this stimulation is mediated by the CB1 receptor. Strikingly, the observed stimulation of the RhoA activator subunit Gα12/13 by WIN55212-2 in the WT disappeared in the absence of CB2 receptor (both in CB2-/- and CB1-/-/CB2-/- mice) suggesting an important role of this CB2 receptor in the intracellular signaling through Gα12/13 in the brain.
Finally, the same experiments were performed incubating the brain membranes with the synthetic anandamide analog ACEA (Figure 5C). Surprisingly, the significant stimulation of Gαi1 that was observed in the WT mice was still found in all the genotypes evaluated. These results suggest that the inhibitory effect of ACEA mediated by the Gαi1 subunit activation may be independent of cannabinoid receptors. On the other hand, the significant stimulation of Gαi3 subunit induced by ACEA was not observed in brain membranes of CB1-/- mice but was still significant in CB2-/- membranes (111 ± 3%). No stimulation was observed in the CB1-/-/CB2-/- mice. Regarding the Gαo subunit, there was a significant stimulation in the absence of CB1 receptors (115 ± 1%), while this stimulation was not observed in the brain membranes of CB2-/- mice, suggesting the necessary role of this receptor in the activation of Gαo induced by the agonist ACEA. The activation of the Gαq/11 subunit was observed in both CB1-/- (117 ± 2%) and CB2-/- (118 ± 4%) but not in the CB1-/-/CB2-/- mice. Thus, as for Δ9-THC and WIN55212-2, the activation of Gαi3 and Gαo subunits was mediated by their interaction with CB1 and CB2 receptors, respectively. However, the stimulation of [35S]GTPγS binding to Gαq/11 subunit seems to be triggered by the activation of both CB1 and CB2 cannabinoid receptors.
Expression of G Protein Subunits in Knockout Mice for Cannabinoid Receptors
In order to determine if a physiological adaptation of knockout mice to the genetic manipulation to inactivate CB1 and/or CB2 receptors may influence our results by the alteration of the expression level of the different Gα protein subunits, Western blotting assays were carried out in brain cortex membranes of WT, CB1-/-, CB2-/-, and CB1-/-/CB2-/- mice.
In the case of Gαi1 (Figure 6A), no changes were observed for CB1-/- and CB2-/-, but a significant reduction (33 ± 10% from WT) of immunodensity was detected in CB1-/-/CB2-/- mice brain membrane homogenates. No significant differences were found in the expression of Gαo (Figure 6C) or Gα12/13 (Figure 6F) subunits between the WT, CB1-/-, CB2-/-, and CB1-/-/CB2-/- mice brain membranes. However, an increase in the expression of Gαi3 (Figure 6B) in both CB2-/- (139 ± 7%) and CB1-/-/CB2-/- mice (140 ± 7%) was found when compared to the WT and CB1-/- animals. On the contrary, Gαz immunodensity was significantly increased in the CB1-/- mice (129 ± 4%) while no changes were found in the rest of the genotypes when comparing to the WT (Figure 6D). Finally, the expression of Gαq/11 (Figure 6E) was significantly increased in brain membranes of CB2-/- mice (137 ± 7%) but not in WT, CB1-/- and CB1-/-/CB2-/- mice.
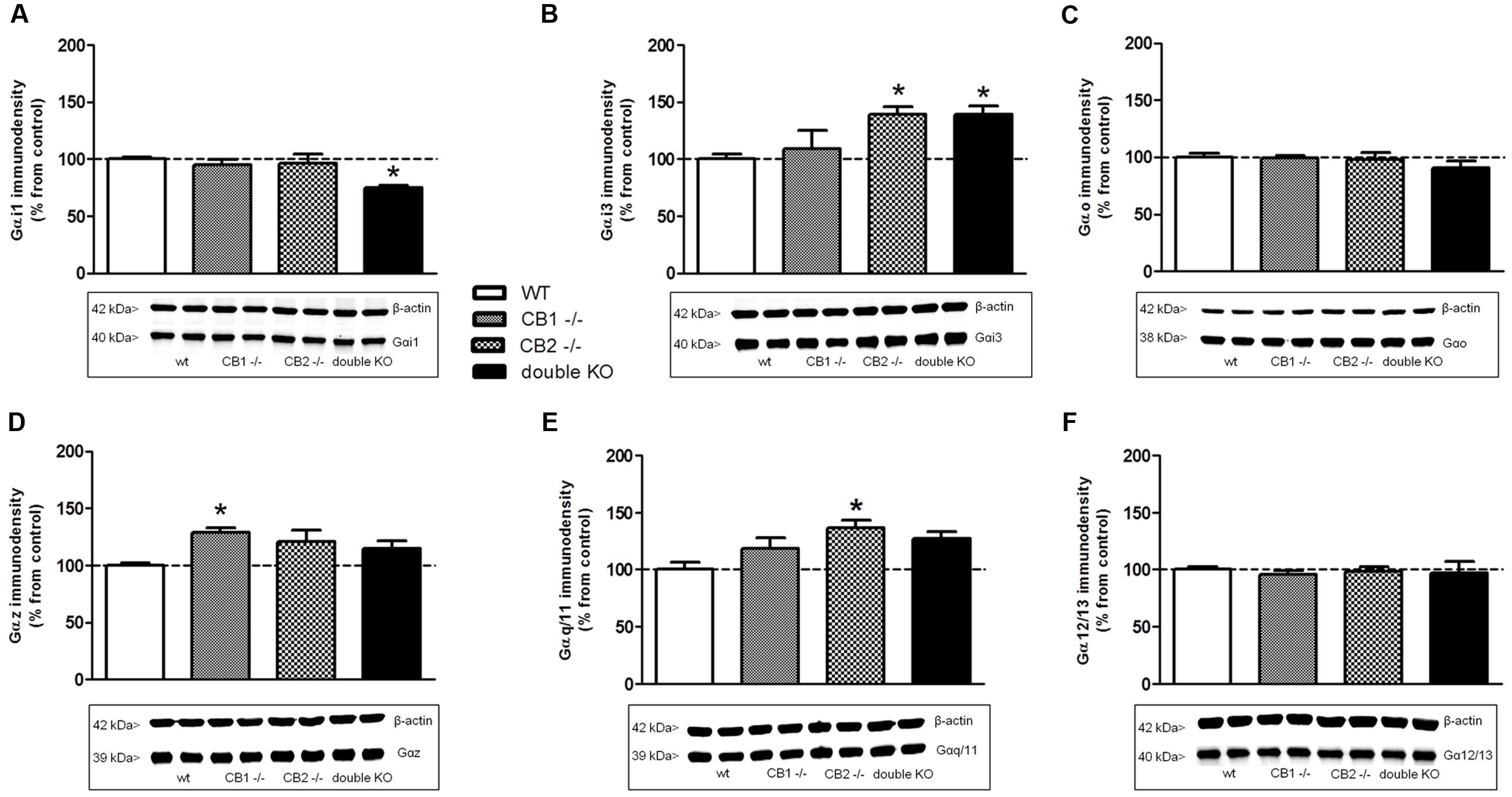
FIGURE 6. Expression of G protein subunits in cannabinoid receptors knockout mice. Immunoreactive signal and representative images obtained by Western blotting with specific antibodies against different Gα subunits: (A) Gαi1, (B) Gαi3, (C) Gαo, (D) Gαz, (E) Gαq/11, and (F) Gα12/13 in mouse brain cortical membranes of WT, CB1 knockout (CB1-/-), CB2 knockout (CB2-/-) and double CB1 and CB2 knockout (CB1-/-/CB2-/-/double KO). Immunoreactivity for β-actin was simultaneously detected on every gel and used as loading control. Normalized values (percentage over controls) of Gα subunits are shown as mean ± SEM of two different experiments carried out in duplicate. Asterisks highlight those values which are statistically different from WT (One-way ANOVA followed by Dunnet’s post hoc test; ∗p < 0.05).
Discussion
During the last years, a considerable effort has been made to study the effects of cannabinoids in the brain trying to elucidate the mechanisms by which these compounds may facilitate mental disorders, such as addiction, cognitive deficits, anxiety or psychosis. In this context, these studies have been performed with different cannabinoid ligands (natural or synthetics), in cells or in native tissue and/or in different animal species (mouse, rat, human…).
There is wide evidence supporting the idea that for most of GPCRs, distinct drugs are able to regulate different signaling pathways by the selective activation of different intracellular effectors. The pharmacological relevance of this fact is that the biological responses not only depend on targeting a specific GPCR but also on the particular pathway that this receptor activates. Different studies have focused on the evaluation of the functional selectivity of cannabinoid receptors, but most of them have been performed in transfected cells expressing the CB1 receptor (Glass and Northup, 1999; Bosier et al., 2010). Moreover, much of these studies explore the signaling pathways activated by different agonists by the evaluation of cAMP production or the phoshorylation of intracellular mediators such as ERK or AKT, with no data about the Gα subtype responsible of these downstream effects. These changes on cAMP concentration or ERK/AKT phosphorylation could be the consequence of the activation of different Gα subtypes, Gβγ dimmers, etc. On the other hand, and interestingly, opposite behavioral effects have been observed after the administration of Δ9-THC or synthetic cannabinoid ligands (Fattore et al., 2003; Panagis et al., 2014; Rubino and Parolaro, 2016). For example, when evaluating the cannabinoid effects on brain-stimulation reward, Fattore et al. (2003) showed that the potent non-selective CB1/CB2 receptor agonists WIN55,212-2 and CP 55,940, but not Δ9-THC, effectively restored heroin-seeking behavior. In addition, it has been suggested that the signaling of CB1 receptors may differ between humans and rodents (Straiker et al., 2012).
All these frequently contradictory data highlight the relevance of studying, simultaneously, the effects of different cannabinoid ligands in the same tissue and under the same experimental conditions.
For that reason the goal of the present study was to compare the pattern of G protein subunit stimulation triggered by three structurally different cannabinoids, Δ9-THC, WIN55212-2 and ACEA in mouse brain cortex. To our knowledge, this is the first study evaluating the cannabinoid-induced stimulation of the different Gα subunits in mouse brain tissue.
WIN55212-2, a synthetic cannabinoid structurally different from Δ9-THC, is a potent, non-selective CB1/CB2 receptor agonist that has been used in many studies of cannabinoid receptor function (Pertwee et al., 2010). The synthetic anandamide analog ACEA is a highly selective agonist for the CB1 receptor with a low affinity for CB2 receptors (Hillard et al., 1999).
This study demonstrates that each ligand displays functional selectivity acting as biased agonist for a subset of different G protein subunits. It represents the first characterization of the activation of individual Gα subunits by endogenous cannabinoid receptors in brain cortex. Firstly, we demonstrated that phytocannabinoid Δ9-THC differs from the synthetic agonists WIN55212-2 and ACEA in its ability to stimulate Gαi/o protein subunits in brain cortex.
The Gαi subfamily members Gαi1, Gαi2, and Gαi3 were originally identified by their ability to inhibit AC activity (Plummer et al., 2012; Busnelli et al., 2013; Minetti et al., 2014). Our results show that Δ9-THC, WIN55212-2, and ACEA significantly stimulate Gαi1 subunit. Moreover, data from knockout mice suggest that this effect may be CB1-mediated in the case of Δ9-THC and WIN55 212-2. However, the Gαi1 stimulation is still significant in membranes of all genotypes incubated with ACEA, suggesting that this is a non-CB1 non-CB2 dependant effect and supporting putative actions of ACEA over other receptors (Pertwee et al., 2010). Gαi3 subunit was also stimulated in the presence of WIN55212-2 and ACEA, but not of Δ9-THC. This stimulation seems to be mediated by CB1 receptors as is blocked in the presence of O-2050 and absent in CB1-/- or CB1-/-/CB2-/- mice. In the case of Gαi2, it has been previously described that WIN55212-2 is able to activate this subunit in rat (Prather et al., 2000) and in human brain cortical membranes (Erdozain et al., 2012). However, none of the agonists in the present study stimulated the Gαi2 subunit. This discrepancy may be due to inter-specie and/or regional differences, suggesting that WIN55212-2 may signal through different G protein pools in human and mouse brain cortex.
These three Gαi subunits form the Gαi/o subfamily with the neuronal α-subunit Gαo, which corresponds to the most abundant Gα protein in brain (Sternweis and Robishaw, 1984). In our experimental approach, and in accordance with other studies (Glass and Northup, 1999; Presley et al., 2016), Δ9-THC, WIN55212-2 and ACEA significantly stimulated Gαo. Results obtained in knockout animals show that the stimulation of Gαo in mouse cortex is mediated, at least in part, by CB2 receptors, suggesting a necessary role of this receptor in the cannabinoid-induced activation of Gαo.
The Gαz subtype is the most divergent member of the inhibitory subfamily and is distributed primarily in neuronal and neuroendocrine cells (Hinton et al., 1990). While Δ9-THC and WIN55212-2, similarly, stimulated Gαz, no stimulation of this subunit was observed when membranes were incubated with ACEA, suggesting that ACEA may not signal through this subunit. Additionally, results obtained with knockout mice suggest that the stimulation of Gαz by Δ9-THC and WIN55212-2 may be induced by a CB2-mediated mechanism.
Unlike Gαz, the Gαs family is ubiquitously expressed and couples receptors to AC in a stimulatory fashion (Milligan and Kostenis, 2006). Under the present assay conditions, nor Δ9-THC, WIN55212-2 or ACEA were able to activate this stimulatory subunit. Thus, there is no evidence of Gαs coupling of cannabinoid receptors in the presence of any of these drugs in brain tissue. There are contradictory results about the ability of cannabinoid drugs to activate Gαs proteins. In this way, there are data from both CHO cell lines (Rinaldi-Carmona et al., 1996; Bonhaus et al., 1998) and HEK cells (Presley et al., 2016) expressing CB1 receptor, showing the absence of effect as well as a modest but significant coupling of CB1 to Gαs triggered by different cannabinoids. It has been proposed that ACEA may elevate cAMP through a non-CB1 mechanism, since there is an increase in cAMP in both cells transfected and non-transfected with CB1 and pretreated with pertussis toxin (Presley et al., 2016). It is important to point out that all these studies have been performed in cell lines. Moreover, they use the accumulation of cAMP in the presence of the Gαi/o inhibitor pertussis toxin as an indirect evaluation of potential coupling of CB1 receptors to Gαs. This increase in cAMP production can be mediated by a mechanism different from Gαs activation, as they did not explore directly the activation of this subunit. Therefore, the possible increase in cAMP induced by other actors different from Gαs subunits could not be discarded.
The Gαq/11 proteins, widely expressed through the CNS, mediate PLC activation, leading to the activation of downstream calcium signaling pathways including PKC and MAPKs activation (Sanchez-Fernandez et al., 2014). In this study, a significant stimulation of Gαq/11 was observed in the presence of the three cannabinoids evaluated. It has been previously reported that WIN55212-2 induces the coupling of CB1 to Gαq/11 in different cellular types (Lauckner et al., 2005; McIntosh et al., 2007). Our results show that not only WIN55212-2 but also Δ9-THC and ACEA can activate Gαq/11 subunit in mouse brain. Moreover, the activation of this subunit induced by Δ9-THC and WIN55212-2 seem to be mediated by the CB1 receptor, as demonstrate the data obtained with knockout animals. In the case of ACEA, our data suggest that ACEA modulate Gαq/11 through both CB1 and CB2 cannabinoid receptors.
The Gα12/13 proteins regulate important signaling events by the activation of the small GTPase protein RhoA, involved in the regulation of the actin cytoskeleton and cell motility (Kozasa et al., 2011; Yu and Brown, 2015). Under our experimental conditions, a significant stimulation of Gα12/13 subunit was observed when membranes were incubated with WIN55212-2 but not with Δ9-THC or ACEA. To our knowledge, this is the first study reporting that WIN55212-2 signals through Gα12/13 in brain cortex. These data are concordant with other studies suggesting that cannabinoids induce the stimulation of this RhoA-activator (Dalton et al., 2013; Roland et al., 2014). Moreover, our results from knockout mice show that the WIN55212-2-induced signaling through Gα12/13 in the brain seems to be mediated, mainly, by the CB2 receptor.
Although O-2050 had been described as a CB1 antagonist, it displays a complex pharmacological profile. In this context, its good affinity for CB2 receptors complicates its use as a tool to evaluate the unique contribution of CB1 receptor (Wiley et al., 2011). We observed that, when alone, O-2050 activated the Gαz subunit. Therefore, in co-incubations, O-2050 behaved always as an antagonist of the effects of Δ9-THC, WIN55212-2 and ACEA over all the studied Gα subunit subtypes, except for the Gαz subunit. When WIN55212-2 and O-2050 were co-incubated, the stimulation of Gαz was lower but still significant. In this way, the blockade exerted by O-2050 pharmacologically confirmed the involvement of cannabinoid receptors in the observed stimulations.
Studies in knockout mice provide very valuable data in basic research but in addition to the absence of the targeted protein, we cannot discard the appearance of putative neurodevelopmental compensatory mechanisms. In this work, we have used CB1 and/or CB2 receptor knockout mice to elucidate the role of each receptor in the observed effects of different ligands on the stimulation of Gα subunits. Moreover, Western blotting assays were carried out in order to unmask the role of a possible adaptation of cannabinoid receptors knockout mice affecting the expression level of the different Gα subtypes on the different genotypes. The observed stimulations in knockout mice may not be influenced by putative neurodevelopmental compensatory mechanisms involving G proteins density. In this way, although expression of some Gα subunits in knockout mice is different from the WT, these changes do not explain the absence of stimulation in CB1 or CB2 knockout mice. The convergence of our pharmacological and genetic data demonstrate that the results obtained herein with the cannabinoid receptors knockout mice are likely due to the absence of the CB1 and/or CB2 receptors and not to non-specific changes due to neurodevelopmental adaptations.
Conclusion
Our results demonstrate that, in mouse brain native tissue and under our experimental conditions different exogenous cannabinoids are able to selectively activate different inhibitory and non-inhibitory Gα protein subtypes, through the activation of CB1 and/or CB2 receptors. However, it is important to be aware of potential limitations. It has been suggested that the signaling of CB1 receptors is significantly diminished in humans compared to that of rodents, a finding that may have implications for the use of rodent models for studies of CB1 receptor function related to human disease and therapy (Straiker et al., 2012).
Results of the present study may help to dissect the specific signaling pathways involved in the different pharmacological actions of cannabinoids. Moreover, the knowledge of the specific molecular target responsible of these different physiological effects will help in the design of new biased cannabinoid drugs with more specific therapeutic effect and a reduced range of adverse effects.
Author Contributions
RD-A, II-L, and AL-C performed the experiments, LC, EA, AG-A, JM, and LU designed the study, JM, LC, RD-A, AG-A, and LU analyzed and interpreted the results, RD-A and LU drafted the manuscript. All the contributors revised critically and gave their approval to the final version of the manuscript.
Funding
This work was supported by Plan Estatal de I+D+i 2013–2016 and Instituto de Salud Carlos III-Subdirección General de Evaluación y Fomento de la Investigación, Spanish Ministry of Economy, and FEDER (grant number PI13/01529 to LU) and the Basque Government (grant number IT616/13). II-L is a recipient of a Predoctoral Fellowship from the Basque Government. AL-C acknowledges the financial support given by Universidad de Antioquia, COLCIENCIAS through the Francisco José de Caldas fellowship 512/2010.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The authors would like to thank Dr. Guillermo Velasco and Dr. Ismael Galve-Roperh (Universidad Complutense de Madrid) for kindly donating CB1-/- and CB2-/- mice. These lines were used to set up a permanent colony and establish the CB1-/-/CB2-/- double knockout mouse model in our lab.
Supplementary Material
The Supplementary Material for this article can be found online at: http://journal.frontiersin.org/article/10.3389/fphar.2016.00415/full#supplementary-material
Abbreviations
2-AG, 2- arachidonoylglycerol; ACEA, arachidonyl-2-chloroethylamide; CB1-/-, CB1 knock-out mice; CB2-/-, CB2 knock-out mice; CB1-/-/CB2-/-, CB1 and CB2 double knock-out mice; P1, pellet fraction; P2, membrane enriched fraction; PVT, polyvinyltoluene; RT, room temperature; SDS, sodium dodecyl sulfate; SPA, scintillation proximity assay; TBS, tris-buffered saline; WT, wild type; Δ9-THC, Δ9-tetrahydrocannabinol.
References
Atwood, B. K., and Mackie, K. (2010). CB2: a cannabinoid receptor with an identity crisis. Br. J. Pharmacol. 160, 467–479. doi: 10.1111/j.1476-5381.2010.00729.x
Bonhaus, D. W., Chang, L. K., Kwan, J., and Martin, G. R. (1998). Dual activation and inhibition of adenylyl cyclase by cannabinoid receptor agonists: evidence for agonist-specific trafficking of intracellular responses. J. Pharmacol. Exp. Ther. 287, 884–888.
Bosier, B., Muccioli, G. G., Hermans, E., and Lambert, D. M. (2010). Functionally selective cannabinoid receptor signalling: therapeutic implications and opportunities. Biochem. Pharmacol. 80, 1–12. doi: 10.1016/j.bcp.2010.02.013
Brents, L. K., Reichard, E. E., Zimmerman, S. M., Moran, J. H., Fantegrossi, W. E., and Prather, P. L. (2011). Phase I hydroxylated metabolites of the K2 synthetic cannabinoid JWH-018 retain in vitro and in vivo cannabinoid 1 receptor affinity and activity. PLoS ONE 6:e21917. doi: 10.1371/journal.pone.0021917
Buckley, N. E., McCoy, K. L., Mezey, E., Bonner, T., Zimmer, A., Felder, C. C., et al. (2000). Immunomodulation by cannabinoids is absent in mice deficient for the cannabinoid CB(2) receptor. Eur. J. Pharmacol. 396, 141–149. doi: 10.1016/S0014-2999(00)00211-9
Busnelli, M., Peverelli, E., Mantovani, G., Spada, A., and Chini, B. (2013). Deciphering the specific role of G(alphai/o) isoforms: functional selective oxytocin ligands and somatostatin SST5 receptor mutants. Biochem. Soc. Trans. 41, 166–171. doi: 10.1042/BST20120306
Canals, M., and Milligan, G. (2008). Constitutive activity of the cannabinoid CB1 receptor regulates the function of co-expressed Mu opioid receptors. J. Biol. Chem. 283, 11424–11434. doi: 10.1074/jbc.M710300200
Childers, S. R., Pacheco, M. A., Bennett, B. A., Edwards, T. A., Hampson, R. E., Mu, J., et al. (1993). Cannabinoid receptors: G-protein-mediated signal transduction mechanisms. Biochem. Soc. Symp. 59, 27–50.
Dalton, G. D., Peterson, L. J., and Howlett, A. C. (2013). CB(1) cannabinoid receptors promote maximal FAK catalytic activity by stimulating cooperative signalling between receptor tyrosine kinases and integrins in neuronal cells. Cell. Signal. 25, 1665–1677. doi: 10.1016/j.cellsig.2013.03.020
De Jesus, M. L., Salles, J., Meana, J. J., and Callado, L. F. (2006). Characterization of CB1 cannabinoid receptor immunoreactivity in postmortem human brain homogenates. Neuroscience 140, 635–643. doi: 10.1016/j.neuroscience.2006.02.024
Dhopeshwarkar, A., and Mackie, K. (2016). Functional selectivity of CB2 cannabinoid receptor ligands at a canonical and noncanonical pathway. J. Pharmacol. Exp. Ther. 358, 342–351. doi: 10.1124/jpet.116.232561
Erdozain, A. M., Diez-Alarcia, R., Meana, J. J., and Callado, L. F. (2012). The inverse agonist effect of rimonabant on G protein activation is not mediated by the cannabinoid CB1 receptor: evidence from postmortem human brain. Biochem. Pharmacol. 83, 260–268. doi: 10.1016/j.bcp.2011.10.018
Fattore, L., Spano, M. S., Cossu, G., Deiana, S., and Fratta, W. (2003). Cannabinoid mechanism in reinstatement of heroin-seeking after a long period of abstinence in rats. Eur. J. Neurosci. 17, 1723–1726. doi: 10.1046/j.1460-9568.2003.02607.x
Gettys, T. W., Sheriff-Carter, K., Moomaw, J., Taylor, I. L., and Raymond, J. R. (1994). Characterization and use of crude alpha-subunit preparations for quantitative immunoblotting of G proteins. Anal. Biochem. 220, 82–91. doi: 10.1006/abio.1994.1302
Glass, M., and Northup, J. K. (1999). Agonist selective regulation of G proteins by cannabinoid CB(1) and CB(2) receptors. Mol. Pharmacol. 56, 1362–1369.
Gonzalez-Maeso, J., Rodriguez-Puertas, R., Gabilondo, A. M., and Meana, J. J. (2000). Characterization of receptor-mediated [35S]GTPgammaS binding to cortical membranes from postmortem human brain. Eur. J. Pharmacol. 390, 25–36. doi: 10.1016/S0014-2999(99)00827-4
Hillard, C. J., Manna, S., Greenberg, M. J., DiCamelli, R., Ross, R. A., Stevenson, L. A., et al. (1999). Synthesis and characterization of potent and selective agonists of the neuronal cannabinoid receptor (CB1). J. Pharmacol. Exp. Ther. 289, 1427–1433.
Hinton, D. R., Blanks, J. C., Fong, H. K., Casey, P. J., Hildebrandt, E., and Simons, M. I. (1990). Novel localization of a G protein, Gz-alpha, in neurons of brain and retina. J. Neurosci. 10, 2763–2770.
Howlett, A. C., Barth, F., Bonner, T. I., Cabral, G., Casellas, P., Devane, W. A., et al. (2002). International union of pharmacology. XXVII. Classification of cannabinoid receptors. Pharmacol. Rev. 54, 161–202. doi: 10.1124/pr.54.2.161
Hudson, B. D., Hebert, T. E., and Kelly, M. E. (2010). Physical and functional interaction between CB1 cannabinoid receptors and beta2-adrenoceptors. Br. J. Pharmacol. 160, 627–642. doi: 10.1111/j.1476-5381.2010.00681.x
Khajehali, E., Malone, D. T., Glass, M., Sexton, P. M., Christopoulos, A., and Leach, K. (2015). Biased agonism and biased allosteric modulation at the CB1 cannabinoid receptor. Mol. Pharmacol. 88, 368–379. doi: 10.1124/mol.115.099192
Kozasa, T., Hajicek, N., Chow, C. R., and Suzuki, N. (2011). Signalling mechanisms of RhoGTPase regulation by the heterotrimeric G proteins G12 and G13. J. Biochem. 150, 357–369. doi: 10.1093/jb/mvr105
Lauckner, J. E., Hille, B., and Mackie, K. (2005). The cannabinoid agonist WIN55,212-2 increases intracellular calcium via CB1 receptor coupling to Gq/11 G proteins. Proc. Natl. Acad. Sci. U.S.A. 102, 19144–19149. doi: 10.1073/pnas.0509588102
Mackie, K. (2005). Distribution of cannabinoid receptors in the central and peripheral nervous system. Handb. Exp. Pharmacol. 168, 299–325. doi: 10.1007/3-540-26573-2_10
Makwana, R., Molleman, A., and Parsons, M. E. (2010). Evidence for both inverse agonism at the cannabinoid CB1 receptor and the lack of an endogenous cannabinoid tone in the rat and guinea-pig isolated ileum myenteric plexus-longitudinal muscle preparation. Br. J. Pharmacol. 160, 615–626. doi: 10.1111/j.1476-5381.2010.00717.x
Marsicano, G., Wotjak, C. T., Azad, S. C., Bisogno, T., Rammes, G., Cascio, M. G., et al. (2002). The endogenous cannabinoid system controls extinction of aversive memories. Nature 418, 530–534. doi: 10.1038/nature00839
McIntosh, B. T., Hudson, B., Yegorova, S., Jollimore, C. A., and Kelly, M. E. (2007). Agonist-dependent cannabinoid receptor signalling in human trabecular meshwork cells. Br. J. Pharmacol. 152, 1111–1120. doi: 10.1038/sj.bjp.0707495
Milligan, G., and Kostenis, E. (2006). Heterotrimeric G-proteins: a short history. Br. J. Pharmacol. 147(Suppl. 1), S46–S55. doi: 10.1038/sj.bjp.0706405
Minetti, G. C., Feige, J. N., Bombard, F., Heier, A., Morvan, F., Nurnberg, B., et al. (2014). Galphai2 signalling is required for skeletal muscle growth, regeneration, and satellite cell proliferation and differentiation. Mol. Cell. Biol. 34, 619–630. doi: 10.1128/MCB.00957-13
Panagis, G., Mackey, B., and Vlachou, S. (2014). Cannabinoid regulation of brain reward processing with an emphasis on the role of CB1 receptors: a step back into the future. Front. Psychiatry 5:92. doi: 10.3389/fpsyt.2014.00092
Pertwee, R. G., Howlett, A. C., Abood, M. E., Alexander, S. P., Di Marzo, V., Elphick, M. R., et al. (2010). International union of basic and clinical pharmacology. LXXIX. Cannabinoid receptors and their ligands: beyond CB(1) and CB(2). Pharmacol. Rev. 62, 588–631. doi: 10.1124/pr.110.003004
Plummer, N. W., Spicher, K., Malphurs, J., Akiyama, H., Abramowitz, J., Nurnberg, B., et al. (2012). Development of the mammalian axial skeleton requires signalling through the Galpha(i) subfamily of heterotrimeric G proteins. Proc. Natl. Acad. Sci. U.S.A. 109, 21366–21371. doi: 10.1073/pnas.1219810110
Prather, P. L., Martin, N. A., Breivogel, C. S., and Childers, S. R. (2000). Activation of cannabinoid receptors in rat brain by WIN 55212-2 produces coupling to multiple G protein alpha-subunits with different potencies. Mol. Pharmacol. 57, 1000–1010.
Presley, C. S., Abidi, A. H., and Moore, B. M. II. (2016). Cannabinoid receptor 1 ligands revisited: Pharmacological assessment in the ACTOne system. Anal. Biochem. 498, 8–28. doi: 10.1016/j.ab.2015.12.019
Rinaldi-Carmona, M., Calandra, B., Shire, D., Bouaboula, M., Oustric, D., Barth, F., et al. (1996). Characterization of two cloned human CB1 cannabinoid receptor isoforms. J. Pharmacol. Exp. Ther. 278, 871–878.
Roland, A. B., Ricobaraza, A., Carrel, D., Jordan, B. M., Rico, F., Simon, A., et al. (2014). Cannabinoid-induced actomyosin contractility shapes neuronal morphology and growth. Elife 3: e03159. doi: 10.7554/eLife.03159
Rubino, T., and Parolaro, D. (2016). The impact of exposure to cannabinoids in adolescence: insights from animal models. Biol. Psychiatry 79, 578–585. doi: 10.1016/j.biopsych.2015.07.024
Sanchez-Fernandez, G., Cabezudo, S., Garcia-Hoz, C., Beninca, C., Aragay, A. M., Mayor, F., et al. (2014). Galphaq signalling: the new and the old. Cell. Signal. 26, 833–848. doi: 10.1016/j.cellsig.2014.01.010
Shapira, M., Vogel, Z., and Sarne, Y. (2000). Opioid and cannabinoid receptors share a common pool of GTP-binding proteins in cotransfected cells, but not in cells which endogenously coexpress the receptors. Cell. Mol. Neurobiol. 20, 291–304. doi: 10.1023/A:1007058008477
Sternweis, P. C., and Robishaw, J. D. (1984). Isolation of two proteins with high affinity for guanine nucleotides from membranes of bovine brain. J. Biol. Chem. 259, 13806–13813.
Straiker, A., Wager-Miller, J., Hutchens, J., and Mackie, K. (2012). Differential signalling in human cannabinoid CB1 receptors and their splice variants in autaptic hippocampal neurones. Br. J. Pharmacol. 165, 2660–2671. doi: 10.1111/j.1476-5381.2011.01744.x
Valdizan, E. M., Castro, E., and Pazos, A. (2010). Agonist-dependent modulation of G-protein coupling and transduction of 5-HT1A receptors in rat dorsal raphe nucleus. Int. J. Neuropsychopharmacol. 13, 835–843. doi: 10.1017/S1461145709990940
Wiley, J. L., Breivogel, C. S., Mahadevan, A., Pertwee, R. G., Cascio, M. G., Bolognini, D., et al. (2011). Structural and pharmacological analysis of O-2050, a putative neutral cannabinoid CB(1) receptor antagonist. Eur. J. Pharmacol. 651, 96–105. doi: 10.1016/j.ejphar.2010.10.085
Keywords: CB1 receptor, CB2 receptor, functional selectivity, scintillation proximity assay (SPA), G proteins, brain
Citation: Diez-Alarcia R, Ibarra-Lecue I, Lopez-Cardona ÁP, Meana J, Gutierrez-Adán A, Callado LF, Agirregoitia E and Urigüen L (2016) Biased Agonism of Three Different Cannabinoid Receptor Agonists in Mouse Brain Cortex. Front. Pharmacol. 7:415. doi: 10.3389/fphar.2016.00415
Received: 11 August 2016; Accepted: 19 October 2016;
Published: 04 November 2016.
Edited by:
Francisco Ciruela, University of Barcelona, SpainReviewed by:
Luca Ferraro, University of Ferrara, ItalyFernando Rodriguez de Fonseca, Instituto de Investigación Biomédica de Málaga, Spain
Copyright © 2016 Diez-Alarcia, Ibarra-Lecue, Lopez-Cardona, Meana, Gutierrez-Adán, Callado, Agirregoitia and Urigüen. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Leyre Urigüen, bGV5cmUudXJpZ3VlbkBlaHUuZXVz