
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Oncol., 05 March 2025
Sec. Gastrointestinal Cancers: Gastric and Esophageal Cancers
Volume 15 - 2025 | https://doi.org/10.3389/fonc.2025.1530430
 Klezzer de Oliveira Carneiro1Taíssa Maíra Thomaz Araújo1
Klezzer de Oliveira Carneiro1Taíssa Maíra Thomaz Araújo1 Ronald Matheus Da Silva Mourão1
Ronald Matheus Da Silva Mourão1 Samir Mansour Moraes Casseb1Samia Demachki1
Samir Mansour Moraes Casseb1Samia Demachki1 Fabiano Cordeiro Moreira1
Fabiano Cordeiro Moreira1 Ândrea Kely Campos Ribeiro Dos Santos2Geraldo Ishak1
Ândrea Kely Campos Ribeiro Dos Santos2Geraldo Ishak1 Daniel de Souza Avelar Da Costa1Leandro Magalhães1
Daniel de Souza Avelar Da Costa1Leandro Magalhães1 Amanda Ferreira Vidal1Rommel Mario Rodriguez Burbano3
Amanda Ferreira Vidal1Rommel Mario Rodriguez Burbano3 Paulo Pimentel de Assumpção1*
Paulo Pimentel de Assumpção1*Introduction: Gastric cancer (GC), which has low survival rates and high mortality, is a major concern, particularly in Asia and South America, with over one million annual cases. Epstein-Barr virus (EBV) is recognized as a carcinogen that may trigger gastric carcinogenesis by infecting the stomach epithelium via reactivated B cells, with growing evidence linking it to GC. This study investigates the transcriptional and microbial profiles of EBV-infected versus EBV-non-infected GC patients.
Methods: Using Illumina NextSeq, cDNA libraries were sequenced, and reads were aligned to the human genome and analyzed with DESeq2. Kegg and differential analyses revealed key genes and pathways. Gene sensitivity and specificity were assessed using ROC curves (p < 0.05, AUC > 0.8). Non-aligned reads were used for microbiome analysis with Kraken2 for bacterial identification. Microbial analysis included LDA score, Alpha and Beta diversity metrics, with significance set at p ≤ 0.05. Spearman’s correlation between differentially expressed genes (DEGs) and bacteria were also examined.
Results: The data revealed a gene expression pattern in EBV-positive gastric cancer, highlighting immune response, inflammation, and cell proliferation genes (e.g., GBP4, ICAM1, IL32, TNFSF10). ROC analysis identified genes with high specificity and sensitivity for discriminating EBV+ gastric cancer, including GBP5, CMKLR1, GM2A and CXCL11 that play pivotal roles in immune response, inflammation, and cancer. Functional enrichment pointed to cytokine-cytokine receptor interactions, antigen processing, and Th17 immune response, emphasizing the role of the tumor microenvironment, shaped by inflammation and immunomodulation, in EBV-associated GC. Microbial analysis revealed changes in the gastric microbiota in EBV+ samples, with a significant reduction in bacterial taxa. The genera Choristoneura and Bartonella were more abundant in EBV+ GC, while more abundant bacteria in EBV- GC included Citrobacter, Acidithiobacillus and Biochmannia. Spearman’s correlation showed a strong link between DE bacterial genera and DEGs involved in processes like cell differentiation, cytokine production, digestion, and cell death.
Conclusion: These findings suggest a complex interaction between the host (EBV+ GC) and the microbiota, possibly influencing cancer progression, and offering potential therapeutic targets such as microbiota modulation or gene regulation. Comparing with EBV- samples further highlights the specific impact of EBV and the microbiota on gastric cancer pathogenesis.
Gastric cancer (GC) is a lethal disease with low overall survival rates worldwide. The majority of new annual GC diagnoses primarily occur in Asian and South American countries (1) and remain a significant public health issue, with over 1 million new cases each year globally. According to data from the International Agency for Research on Cancer in 2022, gastric cancer ranked fifth among the most frequent cancers worldwide in both sexes, representing 4.8% (968,784 cases) (2). Furthermore, gastric adenocarcinoma accounts for over 90% of cases of this neoplasm (3). In 2018, GC incidence rates were nearly twice as high in men as in women (4).
In Brazil, GC was the sixth most frequent cancer (23,021 cases) in 2022 (5). It was estimated that, for each year of the 2020-2022 triennium, 13,360 new cases of stomach cancer occurred among men and 7,870 among women. These values correspond to an estimated risk of 12.81 per 100,000 men and 7.34 per 100,000 women (6).
In this context, the gastric microbiota has emerged as a relevant factor in carcinogenesis. The gastric microbiota, comprising a diverse community of microorganisms, plays a crucial role in stomach health and is closely linked to the development of gastric cancer. Studies suggest that dysbiosis of the gastric microbiota, particularly an increased presence of Helicobacter pylori, contributes to chronic inflammation and damage to the gastric epithelium, promoting conditions conducive to carcinogenesis (7).
Furthermore, the association of gastric cancer with viral infections, such as Epstein-Barr virus (EBV), indicates a significant role for microbial factors in the onset and progression of gastric cancer, with growing evidence linking EBV to gastric carcinogenesis (8). Understanding the gastric microbiota thus opens possibilities for novel therapeutic and preventive approaches (9).
The frequency and distribution of gastric cancer subtypes, as well as the prevalence of H. pylori and EBV, vary across different geographic regions worldwide. The rate of EBV infection in gastric cancer ranges between 4.3% and 18% (10).
An increasing number of reports suggest cooperation between EBV and H. pylori, where the presence of one microorganism may promote the growth or virulence of the other. Although the mechanisms governing this synergistic interaction are not fully understood, evidence suggests that coinfection with H. pylori and EBV significantly increases immune cell recruitment to the infection site, enhancing gastric inflammation and tissue damage. For instance, monochloramine, an oxidant produced in the stomach during H. pylori infection, may trigger the transition of EBV from a latent to a lytic phase (11).
Other pro-inflammatory cytokines emerging from H. pylori-induced gastric inflammation may facilitate EBV proliferation. For example, H. pylori-induced secretion of interferon γ (IFN-γ) promotes an inflammatory environment that exacerbates disease severity (12). Interleukins 6 and 13 (IL-6 and IL-13) promote EBV proliferation, and elevated levels of pro-inflammatory cytokines such as IL-1β, tumor necrosis factor α (TNF-α), and IL-8 have been observed in EBV and H. pylori coinfection associated with severe gastritis (13). Consistent with these findings, plasma IFN-γ levels in gastric cancer patients correlate positively with the extent of EBV reactivation (14).
Additionally, persistent activation of Th17 cells appears to contribute to gastric inflammation associated with H. pylori and EBV coinfection. Th17 cells, a subset of pro-inflammatory CD4+ T helper cells, activate innate immune cells, regulate B cell responses, and are involved in antimicrobial immune responses and wound healing. Th17 cells and their key cytokine, IL-17A, are implicated in the pathogenesis of H. pylori-induced gastritis (15).
Regarding the resident gastric microbiota, recent studies have detected microbiome alterations in gastric cancer compared to non-cancer individuals, indicating dysbiosis, with distinct characteristics that can differentiate GC from other diseases (16). Bacteria found in gastric cardia adenocarcinoma tissues primarily belong to the Firmicutes, Bacteroidetes, and Proteobacteria phyla (17).
At the bacterial genus level, significant increases in the abundance of Achromobacter, Citrobacter, Phyllobacterium, Clostridium, Rhodococcus, and Lactobacillus have been observed in GC compared to chronic gastritis (18). Additionally, a marked difference in microbiota composition exists between non-atrophic gastritis and GC, with bacterial diversity gradually decreasing from non-atrophic gastritis to intestinal metaplasia, and finally to cancer. Notably, H. pylori, Bacteroidetes and Fusobacteria abundance decreases while commensal intestinal bacteria increase (18, 19).
In this context, the present study aimed to investigate the transcriptional and microbial profiles of EBV-infected GC patients, compared to EBV-non-infected GC patients, in order to describe the profile of human transcripts in GC EBV infection, correlate differentially expressed GC genes with EBV positivity and characterize microbiome of EBV+ GC.
Forty-one samples of stomach tumor tissue were collected from patients diagnosed with gastric adenocarcinoma, of which 8 are from patients with GC positive for EBV (GC EBV+) and the remainder were negative for the virus (GC EBV-). None of the EBV-positive samples show co-infection with H. pylori.
The patients included in this study had their medical records and clinical data reviewed for the presence of gastric adenocarcinoma, sex, age, Laurén histological subtype, and H. pylori and EBV infections (Table 1). The individuals were recruited from João de Barros Barreto University Hospital (HUJBB) at the Federal University of Pará (UFPA).
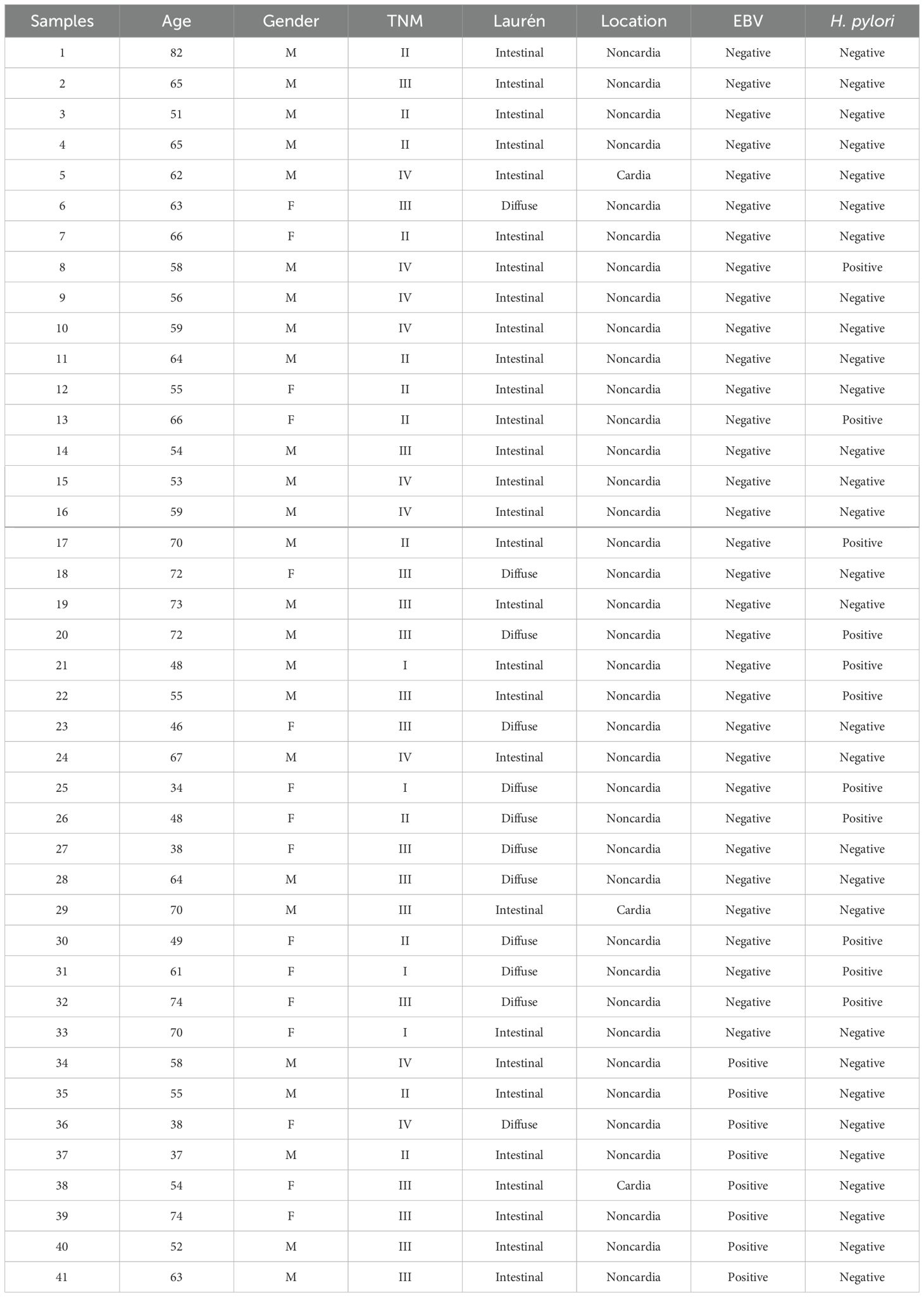
Table 1. Clinicopathologial data of patients.
All participants were thoroughly informed about the study’s objectives and consulted regarding their participation. Samples were only collected after obtaining informed consent through the Free and Informed Consent Form (TCLE). The use of all samples and the conduct of this study were approved by the Ethics Committee for Research of the João de Barros Barreto University Hospital, under CAAE number 47580121.9.0000.5634.
During tumor sample collection, a 0.5 cm fragment of tissue was excised. These fragments were immediately collected post-gastric resection, stored in RNA later for transport, and subsequently preserved at –80°C.
Initially, approximately 50–100 ng of tissue from each sample were homogenized. Following this, 1 ml of TRIZOL® reagent (Thermo Fisher Scientific) was added to the processed tissue for RNA extraction. TRIZOL® was used to maintain RNA integrity and facilitate cell lysis. After centrifugation at 13,000 rpm for 10 minutes at 4°C, RNA was precipitated with isopropyl alcohol. The resulting total RNA was washed with ethanol, air-dried, and kept at room temperature.
RNA integrity and concentration were analyzed using the Qubit 2.0 Fluorometer (Thermo Fisher Scientific), NanoDrop ND-1000 (Thermo Fisher Scientific), and the 2200 TapeStation System (Agilent). Ideal RNA integrity criteria were values between 1.8 and 2.2 (A260/A280 ratio), >1.8 (A260/A230 ratio), and RNA Integrity Number (RIN) ≥ 5. The total RNA obtained was stored at –80°C until further use.
For library construction, the TruSeq Stranded Total RNA Library Prep Kit with Ribo-Zero Gold (Illumina) was used according to the manufacturer’s instructions. A total of 1 μg of total RNA per sample in a 10 μL volume was used for library preparation. After construction, the libraries’ integrity was assessed again using the 2200 TapeStation System, revealing a final product band of ~260 base pairs.
The cDNA libraries were loaded onto the Illumina NextSeq sequencing system and sequenced using paired-end reads. The NextSeq® 500 ID Output V2 Kit - 150 cycles (Illumina) was employed according to the manufacturer’s guidelines.
Base-calling was performed, and the reads were converted to FASTQ format using the Reporter software, encoding sequence and quality scores in ASCII. Read quality was then visualized in FastQC v0.11.9. Low-quality reads and adapters were removed using Trimmomatic (20).
Filtered reads were aligned to the human transcriptome using hg v38 as a reference index for coding transcripts (18, 19, 21) and quantified using Salmon v1.5.2 (22). Reads were imported from Salmon into R v4.1.0 (R: The R Project for Statistical Computing) with the Tximport v3.14.0 package (23). DESeq2 v3.14 (24) was used to estimate gene-level abundances based on Tximport data. Differentially expressed gene threshold was set to |log2FoldChange| ≥ 1 and adjusted p-value ≤ 0.05. Normalized gene levels was used in subsequent analyses.
To investigate potential pathways and biological functions associated with genes, functional enrichment analysis was conducted. KEGG was used through the ClusterProfiler v4.3.2 package in R (25), with enrichment terms considered significant at an adjusted p-value ≤ 0.05.
Receiver operating characteristic (ROC) curves were plotted for each gene’s expression using the pROC v1.18 package (26), with the area under the curve (AUC) calculated to assess each gene’s sensitivity and specificity for the clinically relevant differential expression.
Reads that were not aligned with the human genome were used for microbiome analysis and Kraken2 tool was used to perform bacterial identification. The differential analysis was carried out using the DESeq2 package from the R software, and the bacterial genus was considered differentially abundant (DA) when |log2FC| > 1 and adjusted p-value ≤ 0.05.
To assess the separation between the EBV-positive and EBV-negative groups based on bacterial abundances, the LDA score was computed using Linear Discriminant Analysis (LDA) from the MASS library on R. Statistical analyses between EBV-positive and EBV-negative LDA scores were performed using the Wilcoxon-Mann-Whitney test.
For the analysis of alpha diversity, Simpson and Shannon indices were used and for beta diversity, PERMANOVA, PCoA and the Bray-Curtis plot were used. All analyzes were carried out using Library Vegan in R, being considered significant when the p value ≤ 0.05.
Finally, Spearman correlation was performed between DE genes and DE bacteria. DE genes correlated (ρ > |0.3|) with at least 10% of DE bacteria were used for the correlation plot and gene ontology analysis. Gene Ontology was performed through the ClusterProfiler v4.3.2 package in R (25), with enrichment terms considered significant at an adjusted p-value ≤ 0.05.
Approximately 6 million reads were obtained per sample, with an average of 18,000 human genes identified. Of these, around 5,000 reads were from bacteria, with an average of 600 bacterial genera per sample (Supplementary Tables 1, 2).
No statistically significant correlations were found between clinicopathological data of patients and EBV status. Additionally, sample size did not allow pairing cases by location, besides there are cases with tumors occupying more than one region, limiting this type of analysis.
The differential expression analysis, carried out by DESeq2 package in R, between EBV-positive GC and EBV-negative GC is presented in Figure 1 (117 upregulated and 202 downregulated, Supplementary Table 3), where majority of human genes are found to be downregulated (light blue on the left), including the MUC6 gene, associated with gastric mucus production, and MYCN, a transcription factor implicated in cell cycle control and proliferation.

Figure 1. Volcano plot of differentially expressed genes in CG EBV+ versus CG EBV- samples.
Conversely, on the right side of the volcano plot, there is an overexpression of genes C1QA, C1QB, C1QC, IL32, ADAMDEC1, CMKLR1, TYROBP, and APOE in EBV-positive gastric cancer samples compared to EBV-negative samples (Figure 1), suggesting an enhanced activation of immune and inflammatory pathways, since C1QA, C1QB and C1QC are genes that encode three polypeptide chains that make up the complement C1q subcomponent and are positively correlated with immune cell markers, such as B cells, CD8+ T cells, CD4+ T cells, M1 macrophages and M2 macrophages, and IL32 encodes a proinflammatory cytokine that is involved in the development of inflammatory diseases and malignancies.
The heatmap shown in Figure 2 illustrates a cluster (highlighted in red) of differentially expressed genes in EBV+ GC samples. Key genes in this cluster include LAPTM5, IRF1, TAP1, GBP4, C1QB, GPNMB, ICAM1, APOE, GM2A, TNFSF10, C1QC, TYMP, GBP5, and IDO1. The white cluster highlights downregulated genes in EBV+ GC samples, with notable genes including FAM3B, CAMK2N1, LIMCH1, SARM1, RAP1GAP, and UBXN10.
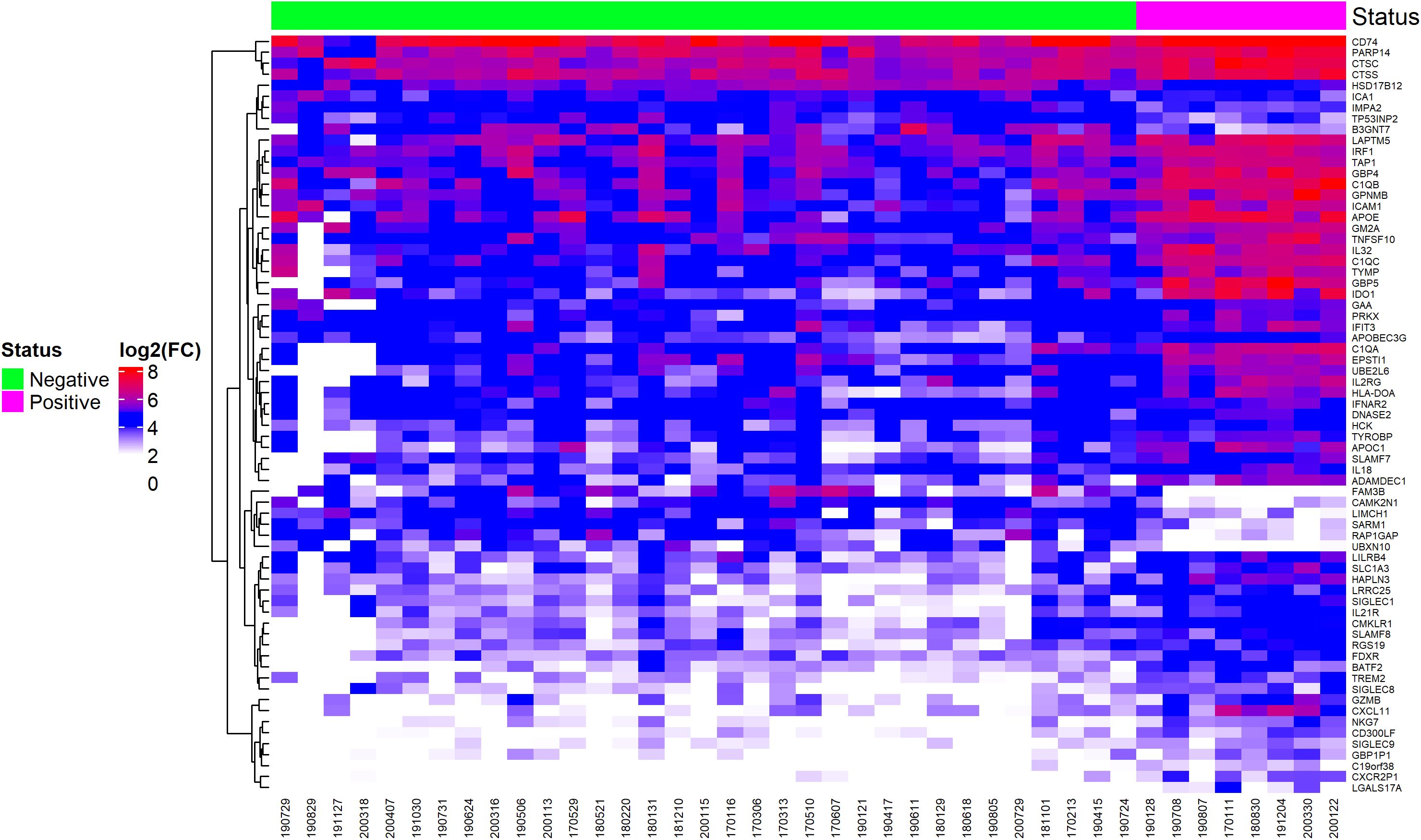
Figure 2. Heatmap of differentially expressed genes, selected by ROC curve analysis, showing EBV-positive and EBV-negative statuses.
Most of these genes are not discussed in literature and their role in EBV infection and cancer is not clear, except for IRF1 and FAM3B, that function as tumor suppressor gene and oncogene, respectively. IRF1 encodes a transcription factor stimulates an immune response against tumor cells and FAM3B encodes a protein called FAM3B/PANDER, which is a hormone that regulates glucose and lipid metabolism and its expression is associated with the progression of multiple types of cancer.
The results of the ROC analysis identified 320 genes with AUC above 0.75 (Supplementary Table 4). The genes LGALS17A (AUC = 1), GBP5 (AUC = 0.99), C1QC (AUC = 0.99), C1QA (AUC = 0.98), C1QB (AUC = 0.97), CMKLR1 (AUC = 0.97), CXCR2P1 (AUC = 0.97), GM2A (AUC = 0.97), CXCL11 (AUC = 0.97), CD300LF (AUC = 0.96), IL32 (AUC = 0.96) (Figure 3) discriminated EBV+ patients with high sensitivity and specificity, suggesting they are potential biomarkers for identifying patients with EBV in gastric cancer.
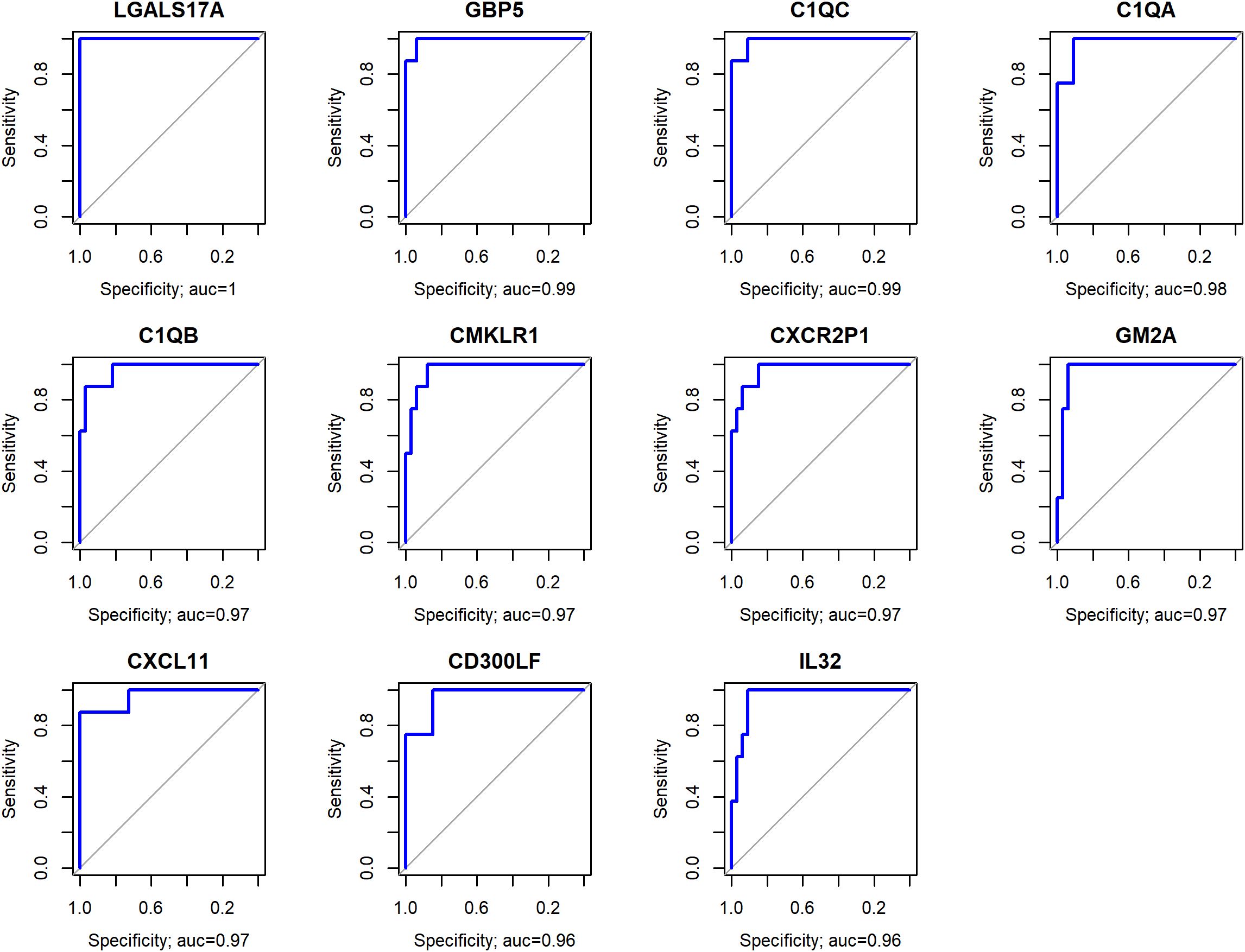
Figure 3. Genes with the highest AUC values, representing the optimal trade-off between sensitivity and specificity in distinguishing EBV+ from EBV- samples.
Among the listed genes, LGALS17A showed the highest AUC value (1), which is related to the very low expression of this gene in EBV- compared to EBV+ patients (p<0.0001), suggesting that this gene has an important role in the pathogenesis of EBV, which needs to be clarified. Importantly, there is no literature available regarding the function of this gene that substantiates its involvement with EBV and/or cancer.
Genes C1QA, C1QB, C1QC, GBP5 CMKLR1, CXCL11 and CXCR2P1 indicates that EBV may be activating inflammatory and immunomodulatory pathways in these tumors. Similarly, the elevated expression of IL32, an inflammatory cytokine, suggests that EBV could be driving a pro-tumoral inflammatory response.
KEGG analysis revealed that many of the differentially expressed genes selected by the ROC curve are involved in immune-related processes, such as cytokine interaction, antigen presentation, Th17 response, and other pathways associated with opportunistic infections (Figure 4).
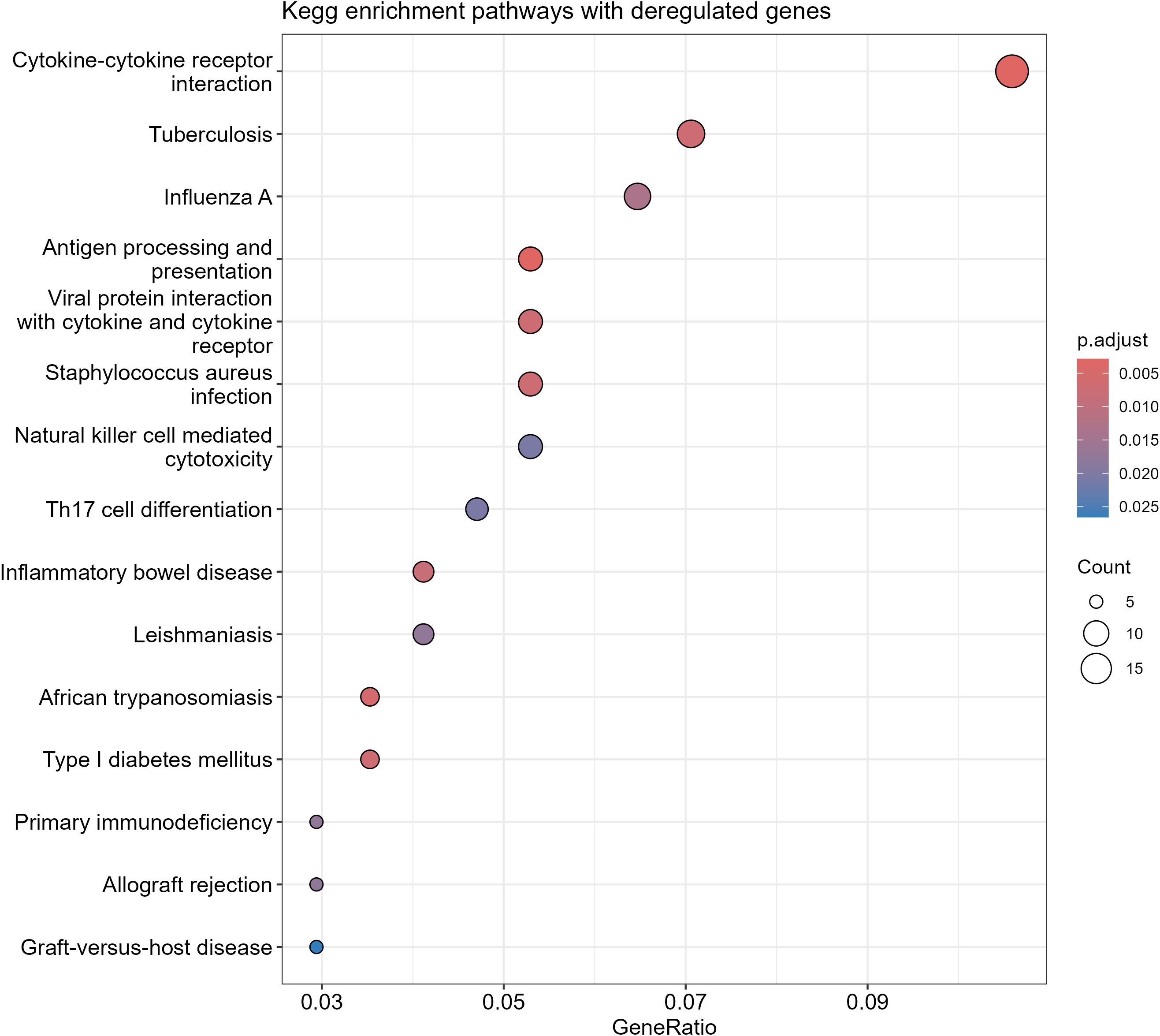
Figure 4. Kegg analysis of differentially expressed genes in EBV+ GC.
When examining the bacterial genera present in the samples, Figure 5 displays differentially abundant bacteria in EBV+ and EBV- GC. The genera Choristoneura and Bartonella were more abundant in EBV+ GC, while differentially abundant bacteria in EBV- GC included Citrobacter, Acidithiobacillus, Biochmannia, Beijerinckia, and Acidaminococcus.
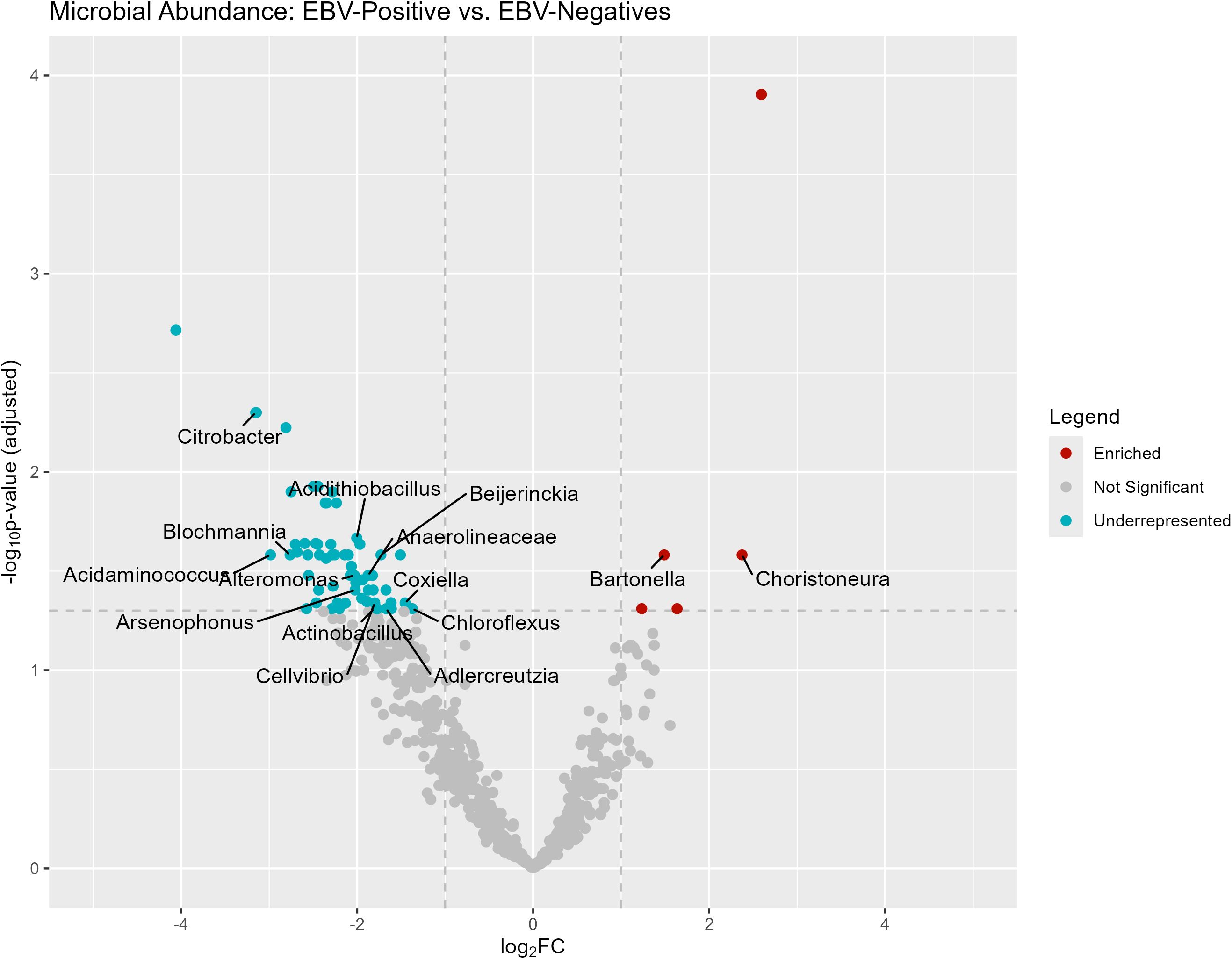
Figure 5. Differentially abundant genera in EBV+ and EBV- GC samples.
Bacterial abundance in EBV-positive and EBV-negative GC samples is represented in a heatmap, showing that EBV+ samples had low abundance for most genera, while a small subset (Shigella, Klebsiella, and Salmonella) remained higher than other bacterial clusters (Figure 6). This same group also maintained high abundance in EBV-negative samples. See Supplementary Table 2 for details.
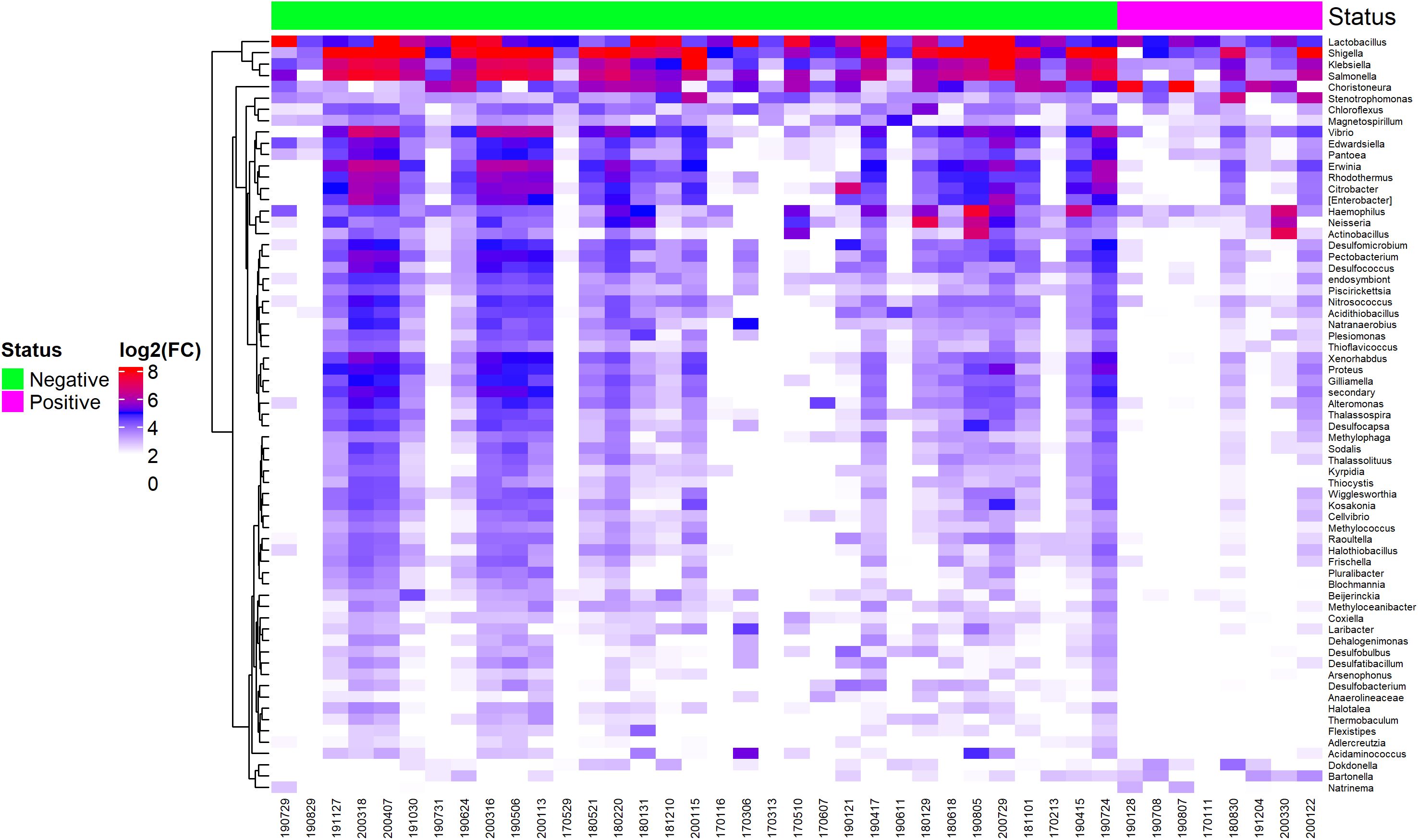
Figure 6. Heatmap of differentially abundant bacterial genera between EBV-positive and EBV-negative statuses.
Additionally, we have conducted a Linear Discriminant Analysis (LDA) to assess the separation between the EBV-positive and EBV-negative groups based on bacterial abundances. The resulting LDA scores are visualized in the Figure 7 (boxplot). As shown, the EBV-positive group exhibits significantly higher LDA scores compared to the EBV-negative group (Wilcoxon; p-value = 0.00001). This clear separation indicates that the bacterial abundance profiles differ between the two groups.
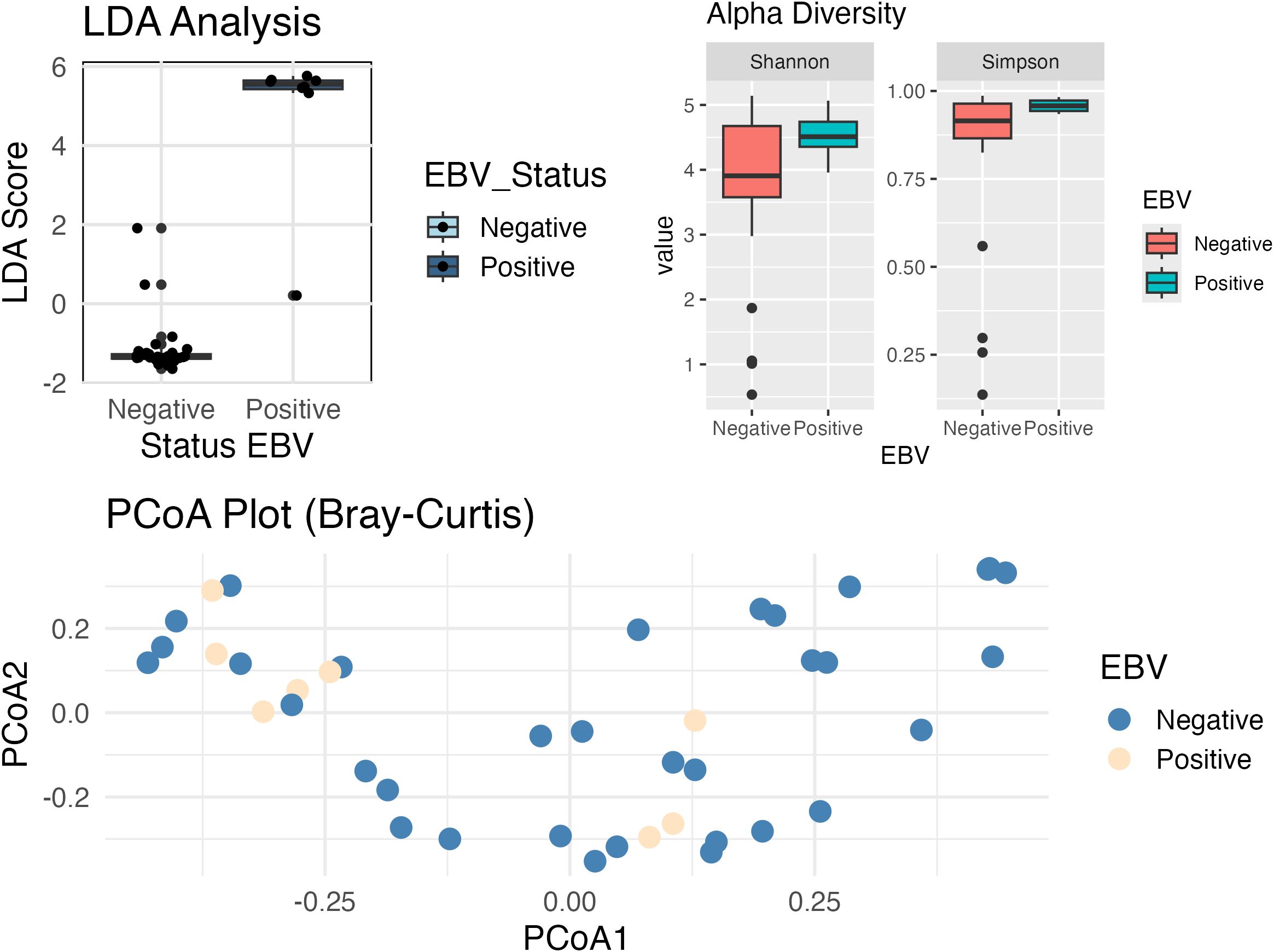
Figure 7. Boxplot comparing Linear Discriminant Analysis (LDA) scores between EBV-negative and EBV-positive samples. EBV-positive samples have higher LDA scores compared to the EBV-negative samples (p. value < 0.001), suggesting a clear separation between the two conditions. Alpha diversity analysis shows significant differences in the diversity and uniformity of bacterial communities between the groups, based on the Shannon (p-value = 0.038) and Simpson (p-value = 0.024) indexes. The Shannon index indicates that EBV+ patients have a more balanced distribution between species, reflecting less dominance of some species, while the Simpson analysis suggests lower bacterial diversity in this group. Beta diversity analysis demonstrate that the overall composition of the bacterial community (PCoA plot; PERMANOVA, p-value = 0.141) did not exhibit statistically significant separation between the groups, suggesting that both share a substantial overlap in taxonomic composition.
Alpha Diversity analysis (Shannon: p-value = 0.038, Simpson: p-value = 0.024) demonstrated that EBV+ and EBV- patients have significant differences in terms of alpha diversity (diversity within each group).
Results indicate that bacterial communities in EBV+ samples differ in uniformity (Shannon) from EBV-, as it has a more balanced distribution between species, that is, less dominance of one or a few species. Additionally, Simpson’s analysis demonstrates that the EBV+ group has a lower bacterial diversity.
No statistically significant separation was found between groups in the overall composition of the bacterial community (PERMANOVA, p-value = 0.141). This could mean that, although there are significant differences in bacterial uniformity and diversity within groups (alpha diversity), the overall composition does not show a clear separation between groups, as they share many taxa.
This analysis strengthens our findings and highlights the distinct bacterial composition associated with EBV status in GC samples.
One hundred and thirty-five DE genes were found correlated (ρ > |0.3|) with at least 10% of DE bacteria (Figure 8, Supplementary Table 5). The results suggest that alteration in bacterial abundance may be influencing or being influenced by the expression of these genes.
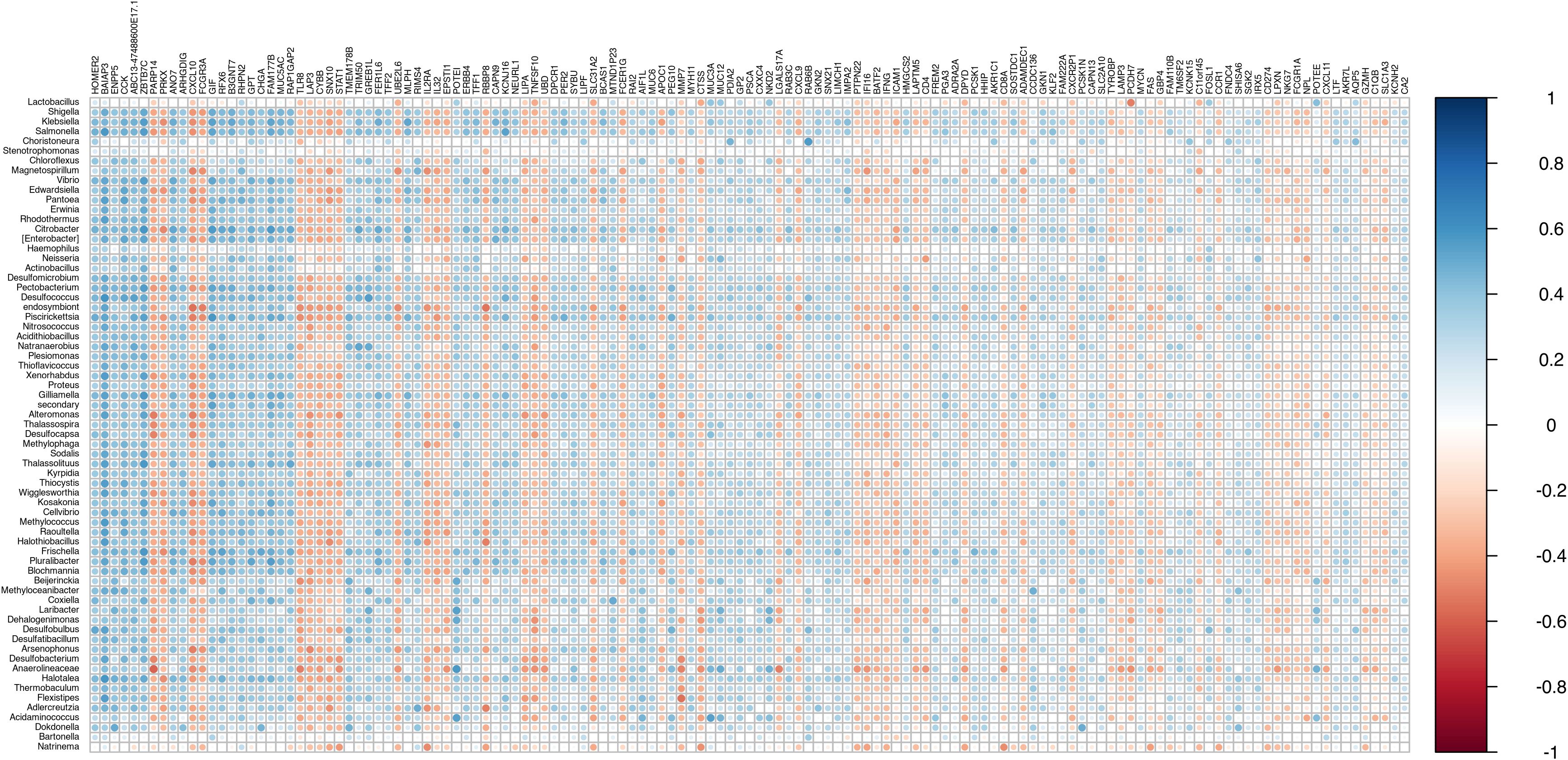
Figure 8. Correlation plot between differentially abundant bacteria and differentially expressed genes. (ρ > |0.3|) with at least 10% of DE bacteria.
A gene ontology analysis was performed using the 135 genes to identify the biological processes involved (Figure 9). The main biological processes described indicate the participation of genes in cell differentiation, regulation of cytokine production, digestion and cell death, suggesting that these pathways can be modulated by the presence of EBV and the bacterial microbiota in the tumor microenvironment.
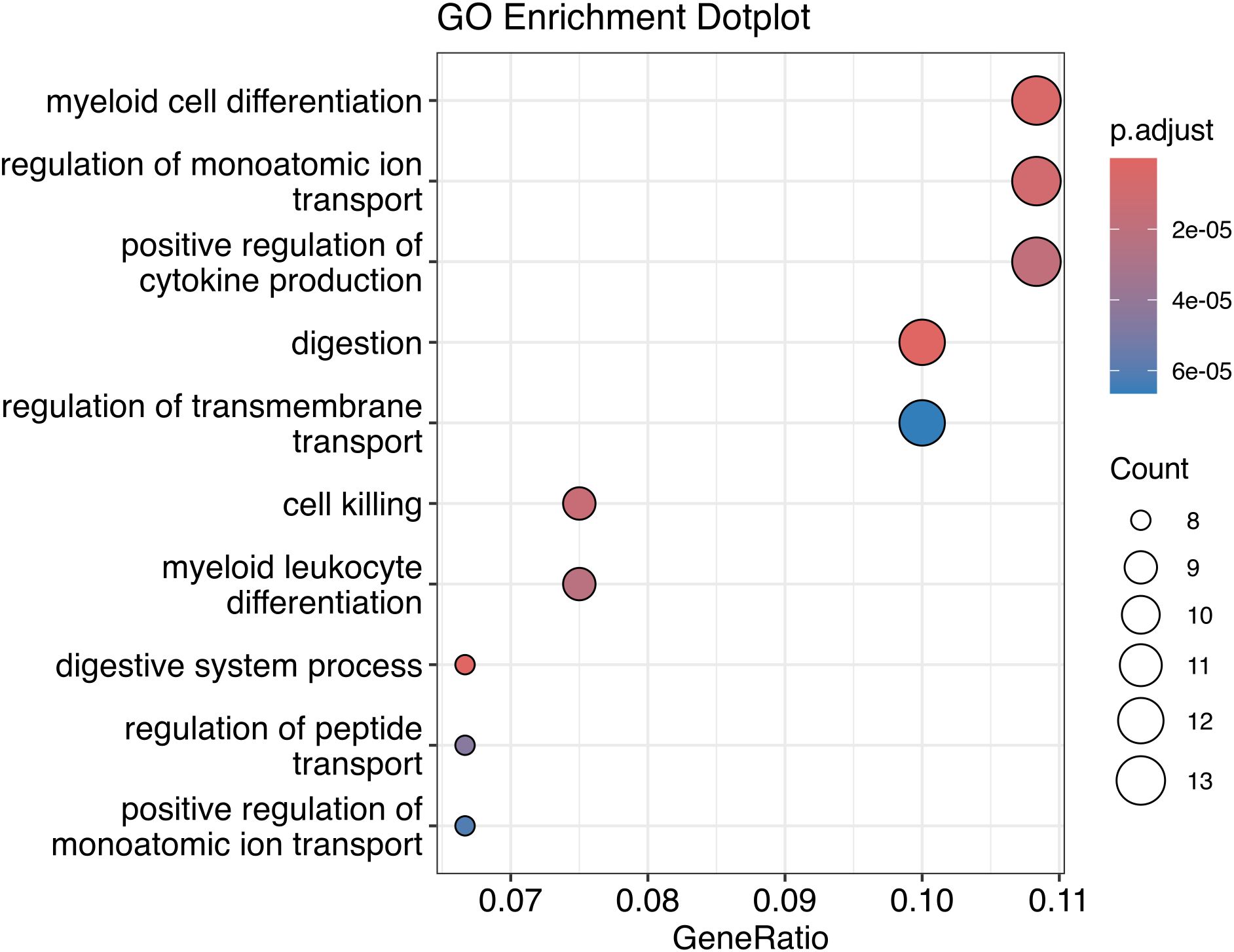
Figure 9. Gene Ontology of DE genes correlated to DE bacteria genera.
The data analysis generated a heatmap indicating a cluster pattern for genes related to immune response in EBV-positive GC patients, including IRF1, C1QB, TNFSF10 and C1QC. The relationship between EBV infection and immune system hyperactivation in the tumor environment is well-established, showing upregulation of various genes positively correlated with immune response and downregulation of genes negatively correlated with immune response (27).
In an immunocompetent host, both innate and adaptive immune responses are triggered by EBV infection, suppressing viral replication. EBV infection increased the proportions of T cells, cytotoxic lymphocytes, CD8+ T cells, NK cells, monocyte lineage cells, and myeloid dendritic cells. In EBV-negative tissues, neutrophils and fibroblasts were present in higher proportions (28, 29).
Importantly, Zhou et al. (29) identified IRF1 (interferon regulatory factor 1) as a key immune-related gene in EBV+ GC, showing increased expression in samples. IRF1 acts as a tumor suppressor and encodes a transcription factor that plays a key role in the body’s defense against infections, cell proliferation, and immune responses. It’s role in various human tumor types has important implications for cancer progression (30).
This gene regulates the expression of multiple genes central to both innate and adaptive immunity, indicating that IRF1 may link the two systems. In addition to its functions in differentiated immune cells, IRF1 has been shown to play roles in the development of various immune cells (dendritic cells, NK cells, CD8+ T cells) that, when activated, counteract the carcinogenic process (31).
The discussed observations demonstrate that tumor surveillance by the immune system is compromised by IRF1 loss. Therefore, IRF1 may function as a “systemic guardian”, protecting the host against exogenous mutagens that can lead to carcinogenesis (30). These findings support literature evidence and clinical data, including TCGA analyses, which indicate a strong correlation between EBV positivity and a more favorable prognosis (32, 33).
Moreover, the volcano plot illustrates differential expression of several genes in EBV-positive patients, notably IL32 and C1QA, both associated with immune activation and also observed as upregulated in the heatmap.
EBV has the potential to immortalize B cells and infect epithelial cells. During immortalization, several EBV products induce cytokines or chemokines necessary for proliferation in infected cells. IL-32 is upregulated after EBV infection, with EBV’s latent membrane protein 1 (LMP1) responsible for inducing its expression (34).
Interleukin (IL)-32 is a recently discovered cytokine with potent pro-inflammatory activity. It is expressed in natural killer cells, monocytes, T lymphocytes, peripheral blood mononuclear cells, epithelial and endothelial cells, and fibroblasts, with various isoforms exhibiting different biological activities. IL-32 is involved in establishing an inflammatory loop that, in turn, induces the synthesis of IL-1β, TNFα, IL-6, IL-8, and macrophage inflammatory protein (MIP)-2 (35). Consistently, IL-32 involvement has been documented in infectious diseases, chronic inflammatory conditions, including gastritis, inflammatory bowel disease, and cancer (36–38).
IL-32’s biological activity depends on cell type and context, with isoform-specific variations, allowing it to either promote or inhibit cancer development. Interestingly, IL-32 and the mechanisms regulating its endogenous isoforms are considered potential targets for anti-neoplastic strategy (35, 36, 38, 39).
The complement system is a critical part of the immune system, protecting the body from invading bacteria and deleterious immune complexes via its enzymatic cascade, receptors, and proteins. Of complement constituents, C1q is a key molecule activating the classical pathway, leading to opsonization and phagocytosis. C1q is encoded by a cluster of three genes (C1QA, C1QB, and C1QC), with C1QA encoding the A-chain polypeptide of the serum complement subcomponent C1q, which associates with C1r and C1s to form the first component of the serum complement system (40).
Corroborating our finds, Deng et al. (41) identified C1QA as a differentially expressed gene when comparing EBV-positive GC patients and EBV-negative GC patients. An integrated bioinformatics analysis revealed that EBV-positive GC expressed more immune-related genes, including common immune checkpoints and human leukocyte antigen (HLA) genes, than EBV-negative GC. Immune scores were higher in EBV+ GC, indicating greater immune cell infiltration and identifying C1QA as an EBV+ GC biomarker.
In line with heatmap results, certain genes, including FAM3B, showed low expression in EBV-positive samples.
Liang et al. (42) studied the EBV+ GC mechanism, identifying 216 downregulated genes by EBV-induced hypermethylation, significantly increasing methylation of ACSS1, FAM3B, IHH, and TRABD in EBV-positive tumors.
DNA methylation is arguably the most significant mechanism in EBV-positive GC. Studies that promoters of 886 genes involved in cancer-related pathways were abnormally hypermethylated in EBV-positive GC (AGS) cells, including FAM3B (43–46).
Maciel-Silva et al. (47) reported that FAM3B could inhibit TNF-α-mediated programmed cell death by upregulating anti-apoptotic Bcl-2 family members and reducing caspase-3 proteolytic activity. These results suggest that FAM3B influences prostate tumorigenesis by modulating the expression of the cell survival genes Bcl-2 and Bcl-XL.
In their study, increased expression of FAM3B was found in AGS/CDDP cells, where its overexpression induced cisplatin resistance. Conversely, FAM3B knockdown increased cisplatin sensitivity in AGS/CDDP cells. Furthermore, FAM3B promoted epithelial-mesenchymal transition (EMT) in gastric cancer cells by upregulating snail protein. Snail inhibition reversed the EMT and cisplatin resistance induced by FAM3B (47).
FAM3B plays a pivotal role in cancer initiation and progression. When upregulated, it can promote invasion and metastasis in human colon cancer, esophageal squamous cell carcinoma, and prostate cancer cells and is associated with poor patient prognosis (47). Similarly, elevated FAM3C is strongly linked to poor prognosis in various cancers, including liver, colorectal, gastric, breast, esophageal squamous cell carcinoma, and oral squamous cell carcinoma (48).
These findings reinforce the oncogenic role of FAM3B. However, its downregulation in EBV-positive samples aligns with clinical and scientific evidence suggesting that EBV infection in gastric cancer is associated with a favorable prognosis.
ROC analysis identified several genes with high specificity and sensitivity for discriminating EBV+ samples in GC. Among these, the LGALS17A gene stood out with the highest AUC value, suggesting that it is a reliable biomarker for identifying patients with these conditions. However, despite its potential significance, LGALS17A is a pseudogene that has not yet been studied in the literature.
The GBP5, C1QC, C1QA, C1QB, CMKLR1, CXCR2P1, GM2A, CXCL11, and IL32 genes play crucial roles in various physiological and pathological processes, including immune response, inflammation, and cancer (34, 35, 40, 41, 49–51). These genes also presented high AUC values (above 0.96), indicating that, together, they form a set of potential biomarkers, providing a more comprehensive understanding of the pathological characteristics of EBV-associated gastric cancer, that could contribute to enhanced diagnosis, prognosis, and the development of personalized therapies.
The CXCL11 gene presents robust data in the literature, and recent studies have found that this gene is involved in the activation of multiple oncogenic signaling pathways and is closely related to tumorigenesis, progression, chemotherapy tolerance, immunotherapy efficacy, and poor prognosis (51–53). Notably, Zhang et al. (54) identified the CXCL11 gene as a key factor in the upregulation of PD-L1 expression in gastric cancer (GC) cells, mediated through the activation of the STAT and PI3K–Akt signaling pathways.
Providing a deeper understanding of the gene expression landscape associated with the immune system, the functional enrichment analysis of differentially expressed genes highlighted cytokine-cytokine receptor interaction, antigen processing and presentation, as well as the Th17 immune response. These findings reinforce the role of the tumor microenvironment, shaped by inflammation and immunomodulation, in the pathogenesis of EBV-associated GC.
The cytokine-cytokine receptor interaction was the most enriched process, suggesting that cytokine-mediated signaling plays a central role in EBV-GC. Studies have demonstrated that EBV can induce the secretion of pro-inflammatory cytokines, such as IL-6 and TNF-α, which promote cell survival, angiogenesis, and tumor invasiveness. This pathway also contributes to local immunosuppression, facilitating tumor immune evasion (55).
The enrichment of antigen processing and presentation reflects EBV’s ability to subvert immune mechanisms. This virus can alter the expression of major histocompatibility complex (MHC) molecules, suppressing antigen presentation and, consequently, the activation of cytotoxic T cells (56). This immune evasion is crucial for viral persistence and tumor progression.
Furthermore, the activation of NK cell-mediated cytotoxicity suggests an initial innate immune response against EBV-infected cells. However, the virus has developed strategies to escape this surveillance, such as the expression of viral proteins that inhibit NK cell function (57).
Th17 differentiation has also emerged as an important aspect of the pathogenesis of EBV-associated GC, as well as other cancers like non-Hodgkin lymphoma (58). Th17 cells are known to mediate chronic inflammation, which is closely associated with the development and progression of gastric tumors (59). Studies indicate that EBV can induce an inflammatory phenotype that favors the expansion of Th17 cells, exacerbating the inflammatory tumor microenvironment (41).
Regarding the study of bacterial diversity present in patients with and without EBV, it was observed that the genera Choristoneura and Bartonella were significantly more abundant in EBV + patients.
These genera have not been previously associated with carcinogenesis. However, alterations in the microbiota are known to significantly impact gastric carcinogenesis. According to the literature, the normal gastric microbiota can harbor 128 phyla, though it is primarily dominated by five phyla: Proteobacteria, Firmicutes, Actinobacteria, Bacteroidetes, and Fusobacteria. Helicobacter pylori is the dominant bacterium in normal gastric mucosa and non-atrophic gastritis. Microbiome diversity tends to decrease when H. pylori predominates and increases as H. pylori diminishes, as seen in premalignant lesions and possibly in GC (60).
Generally, during gastric epithelial progression from normal to GC, H. pylori abundance decreases, while oral and intestinal microbiota frequencies and microbial diversity increase. At the phylum level, Firmicutes are significantly higher in GC patients (61). At the genus level, certain bacteria are enriched in GC patients, while others decrease, such as Bacteroides, Verrucomicrobia, Deferribacteres, and the Lachnospiraceae. Conversely, Clostridium, Fusobacterium, Lactobacillus, Citrobacter, Achromobacter, Rhodococcus, Gemella, Pseudomonas, and Acidovorax are elevated in GC patients.
Dysbiosis leads to microecological changes and activates inflammatory factors in the gastrointestinal mucosa, such as oxidative stress activation, nitric oxide (NO) release, and the production and secretion of pro-inflammatory cytokines and cyclooxygenase-2 (COX-2) (62).
Microbial data from this study reveal a significant decrease in bacterial abundance and diversity in EBV-positive GC samples, except for Shigella, Salmonella, and Klebsiella. These findings suggest a potential interaction between EBV infection and the gastric microbiota, contributing to the dysbiotic state observed in gastric cancer. Reduced bacterial diversity and abundance in EBV-positive samples may reflect the virus’s role in altering the gastric microenvironment, favoring immune evasion and tumor progression. EBV has been shown to modulate the immune system through cytokine signaling and inflammation, creating unfavorable conditions for commensal bacterial colonization (55). This dysbiosis may further exacerbate tumor-promoting processes by altering metabolic pathways and immune responses within the gastric niche.
Interestingly, the enrichment of Shigella, Salmonella, and Klebsiella in EBV-positive samples is noteworthy, as these genera are known for their pathogenic potential and association with inflammation. These bacteria have been implicated in gastrointestinal infections, and their persistence in EBV-positive GC samples may indicate a synergistic interaction between viral and bacterial factors, promoting a pro-inflammatory tumor microenvironment (63). For instance, Shigella and Salmonella can induce inflammatory responses via NF-κB activation (64) and other pathways, which may ultimately complement EBV-driven oncogenic processes.
These results align with emerging evidence linking viral infections to microbiota alterations in gastric cancer. EBV’s ability to suppress bacterial diversity while promoting specific pathogenic taxa underscores the complex interaction between microbial and viral factors in shaping the tumor microenvironment (65). Future studies should explore the mechanistic basis of these interactions, focusing on the role of microbial metabolites, immune modulation, and their contributions to gastric tumorigenesis (66).
In summary, gastric dysbiosis (encompassing bacteria, viruses, acid suppressants, antibiotics, and surgery) can lead to gastric immune dysfunction or result in a decrease in dominant bacteria and an increase in the number and virulence of pathogenic microorganisms, which, in turn, promotes GC development.
Finally, Spearman’s correlation demonstrated a strong correlation between DE bacterial genera and DEGs involved in the biological processes of cell differentiation, regulation of cytokine production, digestion and cell death.
Cellular differentiation and regulation of cytokine production show that these genes can influence the local immune response, which is relevant for understanding how gastric cancer can interact with chronic inflammation mediated by EBV and bacteria present in the tumor environment.
Modulation of digestion may suggest that microbial factors, together with altered gene expression, are implicated in altering gastric tissue homeostasis. Furthermore, cell death may indicate that these genetic and bacterial changes are involved in the survival of tumor cells, perhaps by affecting mechanisms of apoptosis or self-sufficiency in tumor growth.
Therefore, these findings suggest a complex biological process of interaction between the host (in this case, EBV+ gastric cancer) and the microbiota, possibly with a direct impact on cancer progression. These processes may also offer potential therapeutic targets, such as modulation of the microbiota or interventions in the regulation of the genes involved. Comparison with EBV- samples also helps to highlight those related to EBV-associated gastric cancer, providing additional insights into the specific influence of the virus and microbiota on the pathogenesis of gastric cancer.
From the transcriptomic analysis provided by this study, it was possible to investigate the transcriptional landscape of EBV-associated gastric cancer, where high expression of genes involved in immune processes (GBP4, C1QA, ICAM1, CXCL11) and pro-inflammatory processes (IRF, IL32, TNF) was observed in EBV-positive samples. This indicates that the presence of EBV may increase susceptibility to gastric cancer due to alterations in the immune response profile, initiating inflammatory processes as a contributing factor in the development of gastric carcinogenesis, particularly given the significant correlation between these genes and the EBV+ GC status.
Kegg analysis revealed the multifaceted role of EBV in reshaping the gastric tumor microenvironment, promoting pro-tumorigenic biological processes. Understanding these pathways provides valuable insights for the development of targeted therapies, such as cytokine inhibitors and immunotherapy strategies.
Additionally, microbiome analysis suggests that EBV infection in gastric cancer is associated with significant alterations in the gastric microbiota. While most bacterial taxa show reduced abundance in EBV-positive samples, specific pathogens like Shigella, Salmonella, and Klebsiella are enriched, potentially contributing to a pro-inflammatory and tumor-promoting microenvironment. This underscores the complex interaction between viral infection and microbial dynamics in gastric tumorigenesis.
Our findings indicate that both viral and bacterial factors play interdependent roles in modulating the tumor microenvironment, affecting immune response and gastric homeostasis. These results open possibilities for the development of innovative therapeutic strategies, such as microbiome modulation or interventions targeting key genes involved in this complex biological process, aiming to improve the diagnosis and treatment of EBV-associated gastric cancer.
All relevant data is contained within the article: The original contributions presented in the study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding author.
The studies involving humans were approved by Ethics Committee for Research of the João de Barros Barreto University Hospital (CAEE - 47580121.9.0000.5634). The studies were conducted in accordance with the local legislation and institutional requirements. The participants provided their written informed consent to participate in this study. Written informed consent was obtained from the individual(s) for the publication of any potentially identifiable images or data included in this article.
KC: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Resources, Software, Validation, Visualization, Writing – original draft. TA: Conceptualization, Investigation, Resources, Validation, Visualization, Writing – original draft, Writing – review & editing. RD: Conceptualization, Data curation, Formal analysis, Methodology, Resources, Visualization, Writing – review & editing. SC: Writing – review & editing, Data curation, Methodology, Project administration, Validation. SD: Methodology, Validation, Writing – review & editing. FM: Data curation, Formal analysis, Methodology, Writing – review & editing. ÂD: Methodology, Writing – review & editing. GI: Methodology, Writing – review & editing. DD: Conceptualization, Data curation, Formal analysis, Methodology, Software, Writing – review & editing. LM: Data curation, Formal analysis, Methodology, Visualization, Writing – review & editing. AV: Conceptualization, Data curation, Visualization, Writing – review & editing. RB: Conceptualization, Methodology, Validation, Writing – review & editing. Pd: Conceptualization, Funding acquisition, Methodology, Project administration, Resources, Supervision, Validation, Writing – review & editing.
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This work received funding from the Fundação Amazônia de Amparo a Estudos e Pesquisas -FAPESPA (004/21), Conselho Nacional de Desenvolvimento Científico e Tecnológico-CNPq (313303/2021-5) and Ministério Público do Trabalho (11/12/2020 -Ids 372cfc4 and b7c1637).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declare that no Generative AI was used in the creation of this manuscript.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fonc.2025.1530430/full#supplementary-material
1. Rawla P, Barsouk A. Epidemiology of gastric cancer: global trends, risk factors and prevention. pg. (2019) 14:26–38. doi: 10.5114/pg.2018.80001
2. Global cancer observatory: cancer today - world . Available online at: https://gco.iarc.who.int/media/globocan/factsheets/populations/900-world-fact-sheet.pdf (Accessed November 17, 2024).
3. Ilic M, Ilic I. Epidemiology of stomach cancer. WJG. (2022) 28:1187–203. doi: 10.3748/wjg.v28.i12.1187
4. Bray F, Ferlay J, Soerjomataram I, Siegel RL, Torre LA, Jemal A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA A Cancer J Clin. (2018) 68:394–424. doi: 10.3322/caac.21492
5. Ferlay J, Ervik M, Lam F, Laversanne M, Colombet M, Mery L, et al. Global Cancer Observatory: Cancer Today - Brazil. International Agency for Research on Cancer. Available online at: https://gco.iarc.who.int/today/ (Accessed November 17, 2024).
6. INCA. Estimativa 2020: incidência de câncer no Brasil. Inca (2018). Available online at: https://www.inca.gov.br/sites/ufu.sti.inca.local/files/media/document/estimativa-2020-incidencia-de-cancer-no-brasil.pdf (Accessed April 2, 2024).
7. Koga Y. Microbiota in the stomach and application of probiotics to gastroduodenal diseases. World J Gastroenterol. (2022) 28:6702–15. doi: 10.3748/wjg.v28.i47.6702
8. Matsuoka T, Yashiro M. Biomarkers of gastric cancer: Current topics and future perspective. WJG. (2018) 24:2818–32. doi: 10.3748/wjg.v24.i26.2818
9. Dhillon J. Normal components and contaminants. In: Centeno BA, Dhillon J, editors. Monographs in Clinical Cytology. S. Karger AG (2020). p. 34–41. doi: 10.1159/000455733
10. Crowe SE. Helicobacter pylori infection. N Engl J Med. (2019) 380:1158–65. doi: 10.1056/NEJMcp1710945
11. Minoura-Etoh J, Gotoh K, Sato R, Ogata M, Kaku N, Fujioka T, et al. Helicobacter pylori-associated oxidant monochloramine induces reactivation of Epstein-Barr virus (EBV) in gastric epithelial cells latently infected with EBV. J Med Microbiol. (2006) 55:905–11. doi: 10.1099/jmm.0.46580-0
12. Allison CC, Ferrand J, McLeod L, Hassan M, Kaparakis-Liaskos M, Grubman A, et al. Nucleotide oligomerization domain 1 enhances IFN-γ signaling in gastric epithelial cells during Helicobacter pylori infection and exacerbates disease severity. J Immunol. (2013) 190:3706–15. doi: 10.4049/jimmunol.1200591
13. Cárdenas-Mondragón MG, Carreón-Talavera R, Camorlinga-Ponce M, Gomez-Delgado A, Torres J, Fuentes-Pananá EM. Epstein Barr virus and Helicobacter pylori co-infection are positively associated with severe gastritis in pediatric patients. PLoS One. (2013) 8:e62850. doi: 10.1371/journal.pone.0062850
14. Cárdenas-Mondragón MG, Torres J, Sánchez-Zauco N, Gómez-Delgado A, Camorlinga-Ponce M, Maldonado-Bernal C, et al. Elevated levels of interferon-γ Are associated with high levels of Epstein-Barr virus reactivation in patients with the intestinal type of gastric cancer. J Immunol Res. (2017) 2017:7069242. doi: 10.1155/2017/7069242
15. Dávila-Collado R, Jarquín-Durán O, Dong T, Espinoza JL. Epstein-Barr virus and helicobacter pylori co-infection in non-malignant gastroduodenal disorders. Pathogens. (2020) 9:104. doi: 10.3390/pathogens9020104
16. Zhang Y, Zhou L, Xia J, Dong C, Luo X. Human microbiome and its medical applications. Front Mol Biosci. (2022) 8:703585. doi: 10.3389/fmolb.2021.703585
17. Shao D, Vogtmann E, Liu A, Qin J, Chen W, Abnet CC, et al. Microbial characterization of esophageal squamous cell carcinoma and gastric cardia adenocarcinoma from a high-risk region of China. Cancer. (2019) 125:3993–4002. doi: 10.1002/cncr.32403
18. Aviles-Jimenez F, Vazquez-Jimenez F, Medrano-Guzman R, Mantilla A, Torres J. Stomach microbiota composition varies between patients with non-atrophic gastritis and patients with intestinal type of gastric cancer. Sci Rep. (2014) 4:4202. doi: 10.1038/srep04202
19. Ferreira RM, Pereira-Marques J, Pinto-Ribeiro I, Costa JL, Carneiro F, MaChado JC, et al. Gastric microbial community profiling reveals a dysbiotic cancer-associated microbiota. Gut. (2018) 67:226–36. doi: 10.1136/gutjnl-2017-314205
20. Bolger AM, Lohse M, Usadel B. Trimmomatic: a flexible trimmer for Illumina sequence data. Bioinformatics. (2014) 30:2114–20. doi: 10.1093/bioinformatics/btu170
21. Coker OO, Dai Z, Nie Y, Zhao G, Cao L, Nakatsu G, et al. Mucosal microbiome dysbiosis in gastric carcinogenesis. Gut. (2018) 67:1024–32. doi: 10.1136/gutjnl-2017-314281
22. Patro R, Duggal G, Love MI, Irizarry RA, Kingsford C. Salmon provides fast and bias-aware quantification of transcript expression. Nat Methods. (2017) 14:417–9. doi: 10.1038/nmeth.4197
23. Soneson C, Love MI, Robinson MD. Differential analyses for RNA-seq: Transcript-level estimates improve gene-level inferences. F1000Research. (2015) 4:1521. doi: 10.12688/f1000research.7563.2
24. Love MI, Huber W, Anders S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. (2014) 15:550. doi: 10.1186/s13059-014-0550-8
25. Yu G, Wang L-G, Han Y, He Q-Y. clusterProfiler: an R package for comparing biological themes among gene clusters. OMICS. (2012) 16:284–7. doi: 10.1089/omi.2011.0118
26. Robin X, Turck N, Hainard A, Tiberti N, Lisacek F, Sanchez J-C, et al. pROC: an open-source package for R and S+ to analyze and compare ROC curves. BMC Bioinf. (2011) 12:77. doi: 10.1186/1471-2105-12-77
27. Corallo S, Lasagna A, Filippi B, Alaimo D, Tortorella A, Serra F, et al. Unlocking the potential: Epstein-Barr virus (EBV) in gastric cancer and future treatment prospects, a literature review. Pathogens. (2024) 13:728. doi: 10.3390/pathogens13090728
28. Tangye SG, Palendira U, Edwards ESJ. Human immunity against EBV-lessons from the clinic. J Exp Med. (2017) 214:269–83. doi: 10.1084/jem.20161846
29. Zhou H, Jing S, Liu Y, Wang X, Duan X, Xiong W, et al. Identifying the key genes of Epstein–Barr virus-regulated tumour immune microenvironment of gastric carcinomas. Cell Prolif. (2022) 56:e13373. doi: 10.1111/cpr.13373
30. Kano S, Sato K, Morishita Y, Vollstedt S, Kim S, Bishop K, et al. The contribution of transcription factor IRF1 to the interferon-gamma-interleukin 12 signaling axis and TH1 versus TH-17 differentiation of CD4+ T cells. Nat Immunol. (2008) 9:34–41. doi: 10.1038/ni1538
31. Perevalova AM, Gulyaeva LF, Pustylnyak VO. Roles of interferon regulatory factor 1 in tumor progression and regression: two sides of a coin. Int J Mol Sci. (2024) 25:2153. doi: 10.3390/ijms25042153
32. Nakayama A, Abe H, Kunita A, Saito R, Kanda T, Yamashita H, et al. Viral loads correlate with upregulation of PD-L1 and worse patient prognosis in Epstein-Barr Virus-associated gastric carcinoma. PloS One. (2019) 14:e0211358. doi: 10.1371/journal.pone.0211358
33. The Cancer Genome Atlas Research Network. Comprehensive molecular characterization of gastric adenocarcinoma. Nature. (2014) 513:202–9. doi: 10.1038/nature13480
34. Lai K-Y, Chou Y-C, Lin J-H, Liu Y, Lin K-M, Doong S-L, et al. Maintenance of Epstein-Barr virus latent status by a novel mechanism, latent membrane protein 1-induced interleukin-32, via the protein kinase Cδ Pathway. J Virol. (2015) 89:5968. doi: 10.1128/JVI.00168-15
35. Khawar MB, Abbasi MH, Sheikh N. IL-32: A novel pluripotent inflammatory interleukin, towards gastric inflammation, gastric cancer, and chronic rhino sinusitis. Mediators Inflammation. (2016) 2016:8413768. doi: 10.1155/2016/8413768
36. Han H, Jain AD, Truica MI, Izquierdo-Ferrer J, Anker JF, Lysy B, et al. Small-molecule MYC inhibitors suppress tumor growth and enhance immunotherapy. Cancer Cell. (2019) 36:483–497.e15. doi: 10.1016/j.ccell.2019.10.001
37. Sloot S, Chen YA, Zhao X, Weber JL, Benedict JJ, Mulé JJ, et al. Improved survival of patients with melanoma brain metastases in the era of targeted BRAF and immune checkpoint therapies. Cancer. (2018) 124:297–305. doi: 10.1002/cncr.30946
38. Yan Y, Kumar AB, Finnes H, Markovic SN, Park S, Dronca RS, et al. Combining immune checkpoint inhibitors with conventional cancer therapy. Front Immunol. (2018) 9:1739. doi: 10.3389/fimmu.2018.01739
39. Re VD, Brisotto G, Repetto O, Zorzi MD, Caggiari L, Zanussi S, et al. Overview of Epstein–Barr-virus-associated gastric cancer correlated with prognostic classification and development of therapeutic options. Int J Mol Sci. (2020) 21:9400. doi: 10.3390/ijms21249400
40. Bruiners N, Schurz H, Daya M, Salie M, van Helden PD, Kinnear CJ, et al. A regulatory variant in the C1Q gene cluster is associated with tuberculosis susceptibility and C1qA plasma levels in a South African population. Immunogenetics. (2020) 72:305–14. doi: 10.1007/s00251-020-01167-5
41. Deng S-Z, Wang X-X, Zhao X-Y, Bai Y-M. amp]]amp; Zhang, H.-M. Exploration of the Tumor Immune Landscape and Identification of Two Novel Immunotherapy-Related Genes for Epstein-Barr virus-associated Gastric Carcinoma via Integrated Bioinformatics Analysis. Front Surg. (2022) 9:898733. doi: 10.3389/fsurg.2022.898733
42. Liang Q, Yao X, Tang S, Zhang J, Yau TO, Li X, et al. Integrative identification of Epstein-Barr virus-associated mutations and epigenetic alterations in gastric cancer. Gastroenterology. (2014) 147:1350–62.e4. doi: 10.1053/j.gastro.2014.08.036
43. Zhao J, Liang Q, Cheung K-F, Kang W, Lung RWM, Tong JHM, et al. Genome-wide identification of Epstein-Barr virus-driven promoter methylation profiles of human genes in gastric cancer cells. Cancer. (2013) 119:304–12. doi: 10.1002/cncr.27724
44. Farrell PJ. Epstein-Barr virus and cancer. Annu Rev Pathol. (2019) 14:29–53. doi: 10.1146/annurev-pathmechdis-012418-013023
45. Wang J, Liu W, Zhang X, Zhang Y, Xiao H, Luo B. LMP2A induces DNA methylation and expression repression of AQP3 in EBV-associated gastric carcinoma. Virology. (2019) 534:87–95. doi: 10.1016/j.virol.2019.06.006
46. Zhao Z, Liu W, Liu J, Wang J, Luo B. The effect of EBV on WIF1, NLK, and APC gene methylation and expression in gastric carcinoma and nasopharyngeal cancer. J Med Virol. (2017) 89:1844–51. doi: 10.1002/jmv.24863
47. Maciel-Silva P, Caldeira I, Santos I de A, Carreira ACO, Siqueira FR, Antonioli E, et al. FAM3B/PANDER inhibits cell death and increases prostate tumor growth by modulating the expression of Bcl-2 and Bcl-XL cell survival genes. BMC Cancer. (2018) 18:90. doi: 10.1186/s12885-017-3950-9
48. Zhu Y, Pu Z, Wang G, Li Y, Wang Y, Li N, et al. FAM3C: an emerging biomarker and potential therapeutic target for cancer. biomark Med. (2021) 15:373–84. doi: 10.2217/bmm-2020-0179
49. Shin J, Kim G, Lee JW, Lee JiE, Kim YS, Yu J-H, et al. Identification of ganglioside GM2 activator playing a role in cancer cell migration through proteomic analysis of breast cancer secretomes. Cancer Sci. (2016) 107:828–35. doi: 10.1111/cas.12935
50. Cao F-Y, Wang C-H, Li X, Ma M-Z, Tao G-C, Yang C, et al. Guanylate binding protein 5 accelerates gastric cancer progression via the JAK1-STAT1/GBP5/CXCL8 positive feedback loop. Am J Cancer Res. (2023) 13:1310–28.
51. Wang J, Ouyang X, Zhu W, Yi Q, Zhong J. The Role of CXCL11 and its Receptors in Cancer: Prospective but Challenging Clinical Targets. Cancer Control. (2024) 31:10732748241241162. doi: 10.1177/10732748241241162
52. Li Y, Han S, Wu B, Zhong C, Shi Yu, Lv C, et al. CXCL11 correlates with immune infiltration and impacts patient immunotherapy efficacy: A pan-cancer analysis. Front Immunol. (2022) 13:951247. doi: 10.3389/fimmu.2022.951247
53. Puchert M, Obst J, Koch C, Zieger K, Engele Jürgen. CXCL11 promotes tumor progression by the biased use of the chemokine receptors CXCR3 and CXCR7. Cytokine. (2020) 125:154809. doi: 10.1016/j.cyto.2019.154809
54. Zhang C, Li Z, Xu L, Che X, Wen Ti, Fan Y, et al. CXCL9/10/11, a regulator of PD-L1 expression in gastric cancer. BMC Cancer. (2018) 18:462. doi: 10.1186/s12885-018-4384-8
55. Shinozaki-Ushiku A, Kunita A, Isogai M, Hibiya T, Ushiku T, Takada K, et al. Profiling of virus-encoded microRNAs in Epstein-Barr virus-associated gastric carcinoma and their roles in gastric carcinogenesis. J Virol. (2015) 89:5581–91. doi: 10.1128/JVI.03639-14
56. Yin Y, Manoury B, Fåhraeus R. Self-inhibition of synthesis and antigen presentation by Epstein-Barr virus-encoded EBNA1. Sci (New York N.Y.). (2003) 301:1371–4. doi: 10.1126/science.1088902
57. Png YT, Yang AZY, Lee MY, Chua MJM, Lim CM. The role of NK cells in EBV infection and EBV-associated NPC. Viruses. (2021) 13:300. doi: 10.3390/v13020300
58. Duffield AS, Ascierto ML, Anders RA, Taube JM, Meeker AK, Chen S, et al. Th17 immune microenvironment in Epstein-Barr virus–negative Hodgkin lymphoma: Implications for immunotherapy. Blood Adv. (2017) 1:1324. doi: 10.1182/bloodadvances.2017007260
59. Rezalotfi A, Ahmadian E, Aazami H, Solgi G, Ebrahimi M. Gastric cancer stem cells effect on Th17/Treg balance; A bench to beside perspective. Front Oncol. (2019) 9:226. doi: 10.3389/fonc.2019.00226
60. Fakharian F, Asgari B, Nabavi-Rad A, Sadeghi A, Soleimani N, Yadegar A, et al. The interplay between Helicobacter pylori and the gut microbiota: An emerging driver influencing the immune system homeostasis and gastric carcinogenesis. Front Cell Infection Microbiol. (2022) 12:953718. doi: 10.3389/fcimb.2022.953718
61. Yang Y, Ji R, Zhao X, Cao X, Wang Q, Jiang Q, et al. Alterations in gastric mucosal microbiota in gastric carcinogenesis: A systematic review and meta-analysis. Front Med (Lausanne). (2021) 8:754959. doi: 10.3389/fmed.2021.754959
62. Li L, Peng P, Ding N, Jia W, Huang C, Tang Y. Oxidative stress, inflammation, gut dysbiosis: what can polyphenols do in inflammatory bowel disease? Antioxidants. (2023) 12:967. doi: 10.3390/antiox12040967
63. Ryan JL, Shen Y-J, Morgan DR, Thorne LB, Kenney SC, Dominguez RL, et al. Epstein-Barr virus infection is common in inflamed gastrointestinal mucosa. Digestive Dis Sci. (2012) 57:1887. doi: 10.1007/s10620-012-2116-5
64. Zhao L, Cho WC, Nicolls MR. Colorectal cancer-associated microbiome patterns and signatures. Front Genet. (2021) 12:787176. doi: 10.3389/fgene.2021.787176
65. Kato I, Zhang J, Sun J. Bacterial-viral interactions in human orodigestive and female genital tract cancers: A summary of epidemiologic and laboratory evidence. Cancers. (2022) 14:425. doi: 10.3390/cancers14020425
Keywords: gastric cancer, EBV, metatranscriptomics, gastric microbiome, carcinogenesis
Citation: Carneiro KdO, Araújo TMT, Da Silva Mourão RM, Casseb SMM, Demachki S, Moreira FC, Dos Santos ÂKCR, Ishak G, Da Costa DdSA, Magalhães L, Vidal AF, Burbano RMR and de Assumpção PP (2025) Transcriptional and microbial profile of gastric cancer patients infected with Epstein-Barr virus. Front. Oncol. 15:1530430. doi: 10.3389/fonc.2025.1530430
Received: 18 November 2024; Accepted: 17 February 2025;
Published: 05 March 2025.
Edited by:
Dunfa Peng, University of Miami, United StatesCopyright © 2025 Carneiro, Araújo, Da Silva Mourão, Casseb, Demachki, Moreira, Dos Santos, Ishak, Da Costa, Magalhães, Vidal, Burbano and de Assumpção. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Paulo Pimentel de Assumpção, YXNzdW1wY2FvcHBAZ21haWwuY29t
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.