 Emily Toscano-Guerra1,2,3,4*
Emily Toscano-Guerra1,2,3,4* Valentina Maggio1
Valentina Maggio1 Javier García1
Javier García1 Maria Eugenia Semidey2,5
Maria Eugenia Semidey2,5 Ana Celma2,6
Ana Celma2,6 Juan Morote2,5Inés de Torres2,5Marina Giralt3Roser Ferrer-Costa2,3
Juan Morote2,5Inés de Torres2,5Marina Giralt3Roser Ferrer-Costa2,3 Rosanna Paciucci1,2,3*
Rosanna Paciucci1,2,3*- 1Cell Signaling and Cancer Progression Laboratory, Vall d’Hebron Institute of Research (VHIR), Barcelona, Spain
- 2Department of Biochemistry and Molecular Biology, Autonomous University of Barcelona, Barcelona, Spain
- 3Clinical Biochemistry Department, Biochemistry Service, Vall d’Hebron Hospital, Barcelona, Spain
- 4Facultad Ciencias e Ingeniería, Universidad Peruana Cayetano Heredia, Lima, Peru
- 5Department of Pathology, Vall d’Hebron Hospital, Barcelona, Spain
- 6Department of Urology, Vall d’Hebron Hospital, Barcelona, Spain
Background: Prostate cancer (PCa) is a leading cause of cancer-related deaths in European men, emphasizing the urgent need for effective risk assessment strategies. The TP53 gene, a tumor suppressor gene frequently mutated in cancer, commonly harbors the rs1042522 single nucleotide polymorphism (SNP), known as the P72R SNP, which may influence PCa susceptibility. This study investigated the prevalence of the P72R SNP in European Caucasian PCa samples and its association with PCa risk.
Methods: Genotyping was conducted on 12 hormone-naïve aggressive PCa cultures (hnPCs) from untreated patients (Gleason ≥8), 11 radical prostatectomies (RP), and 94 serum samples using DNA Sanger sequencing and melting curve analysis. Comparative analysis utilized data from the GnomAD database’s European Caucasian non-cancer population.
Results: Our results demonstrate a significantly higher frequency of the P72R SNP in PCa samples and serums compared to the general European non-cancer population. A robust and statistically significant association (p < 0.0001) between the SNP and prostate cancer risk was identified, with an odds ratio of 7.937 (95% CI 5.37-11.00). Notably, the G allele (R72) showed a pronounced prevalence in high Gleason score (≥8) patients, although statistical significance was not reached. These results highlight a potential association with undifferentiated and malignant PCa lesions.
Conclusion: The compelling association between the P72R SNP and prostate cancer risk underscores the potential utility of this marker for the early identification of patients at risk of aggressive metastatic prostate cancer. This insight could empower further research to intervene at an early stage by offering enhanced opportunities for timely and targeted interventions.
Introduction
Prostate cancer (PCa), which is highly prevalent in developed countries, is the most common cancer in men and the third most common cause of cancer-related deaths in Europe after lung and colorectal cancer (1, 2). In the majority of cases, cancer is efficiently eradicated and cured by surgical prostatectomy, but late-diagnosed or metastatic aggressive cancers have a five-year relative survival rate of less than 30% (3). The tumor suppressor gene TP53 plays a pivotal role in the prevention of tumor development. Not surprisingly, it has also been described as the most commonly mutated gene in human cancers (4). Somatic TP53 mutations occur more frequently as later events and are drivers of aggressive and metastatic cancers, including PCa (5).
Several hereditary genetic variations, found in approximately 5-20% of different cancer types, are associated with pathogenic variants of genes, such as BRCA2 and ATM. In PCa, pathogenic germline variants of the TP53 gene have been described with a relative risk of 4.7-8.6, which is comparable to the frequencies described for those genes (6).
Germline variant alleles, known as single-nucleotide polymorphisms (SNPs), are present in over 1% of the population and occur naturally. Although many of these variants are considered to have minor or not impact on human health, recent significant studies indicate otherwise, particularly in PCa (7, 8). Some common SNPs can indeed affect the structure and function of proteins, thereby contributing to severe health outcomes. In PCa, specific SNPs, such as those in the BTNL2 gene, have been associated with elevated prostate cancer risk, and structural variants have been linked to poor clinical outcomes and disease progression (9, 10).
Despite significant progress in understanding PCa genetics, the understanding of some controversial germline genetic variations is still incomplete. In particular, the P72R single nucleotide polymorphism (rs1042522) has shown conflicting results in its association with prostate cancer risk and aggressiveness in various reports (11–14). These studies suggest that P72R may influence PCa susceptibility, although the exact nature of this influence remains debated within the scientific community.
By investigating the association of the rs1042522 SNP of the TP53 gene in cancer tissues, primary tumor cultures, and serum samples, our study aims to provide additional evidence of the association of this SNP with prostate cancer risk in a European Caucasian population. Here, we describe the identification of the rs1042522 in the majority of hnPCs analyzed. Its presence was confirmed in a cohort of 11 radical prostatectomy (RP) tissues and 94 serum samples from Caucasian patients with prostate cancer. The association between the variant allele and prostate cancer risk was also investigated by comparing with the healthy (non-cancerous) European (non-Finnish) population from GnomAD v2.1.1.
These results highlight the potential value of rs1042522 assessment for identifying patients at risk for aggressive metastatic prostate cancer.
Materials and methods
Prostate primary culture from hormone-naïve patients
Primary cultures from prostate tumor needle biopsies of hormone-naïve patients (i.e. without previous treatment) were established in our laboratory from a cohort of patients with aggressive/metastatic prostate cancer, selected for (i) high levels of serum PSA (≥50 ng/mL), (ii) positive digital rectal examination (DRE), and (iii) Gleason ≥ 8. hnPCs were cultured at 37°C in an atmosphere of 5% CO2 with complete DMEM-F12 medium containing 2 mM L-glutamine, 100 U of penicillin/mL, 100 µg streptomycin/mL, 0.1 mM nonessential amino acids, 1 mM sodium pyruvate and 7% fetal bovine serum, Supplement 1X, human FGF-10 (10 ng/µL), human EGF (20 ng/µL), and vitamin A and E (200 ng/µL). Growth factors and vitamins were added freshly, and supplements were prepared and stored at -20°C until use. The supplement (100X) was prepared in DMEM-F12 containing glucose (6 mg/mL), transferrin (1000 µg/mL), human insulin (2.500 µg/mL), putrescine (97 µg/mL), sodium selenite (30 µM), and hydrocortisone (100 µM).
Radical prostatectomy samples
Eleven radical prostatectomy (RP) samples from patients with localized operable tumors (Gleason score ≤7) were compared with the hnPCs’s. Samples were collected by the Service of Urology of the Hospital Vall d´Hebron (HVH), preserved in optimal cutting temperature (OCT) compound, and stored at -80°C until frozen sectioning.
Serum samples from PCa patients
A cohort of 94 surplus serum samples from Southern European Spanish male patients followed up at the HVH Biochemistry Service, diagnosed with PCa by tissue biopsy and elevated PSA, were randomly selected according to biobank availability, sample quality (non-hemolysed) and ethnicity. Blood samples were collected in SST yellow tubes containing separator gels were centrifuged at 3,500 rpm for 15 min at 4°C. The resulting serum was collected in 2 mL tubes, properly labelled and codified in the biobanking system, and subsequently stored at -80°C.
The study was reviewed and approved by the HVH Institutional Review Board (Medical Research Ethics Committee, protocol number PR(AG) 96/2015).
DNA sequencing
The variant rs1042522 of the TP53 gene was first identified in hnPCs and RPs by analysis of complementary DNA (cDNA) using RNA reverse transcription (RT) and Sanger sequencing. Briefly, Total RNA from cells/tissues was isolated using RNeasy mini kit (QIAGEN) according to the manufacturer’s protocol. For tissue samples, a combination of Phenol-based RNA isolation and purification using silica method was used. Good RNA quality was confirmed using a 2100 Bioanalyzer (Agilent), and samples with RIN≥7 were used. cDNA was synthesized from 1.0 µg (cells/tissues) of total RNA using the NZY M-MuLV First Strand cDNA Synthesis Kit (NZY Tech) and 2720 Thermocycler (Applied Biosystems). The resulting amplicons were sequenced using the Sanger method (Macrogen Europe service).
DNA genotyping
Genomic DNA from hnPCs and RPs was extracted using a DNeasy Mini Kit (Qiagen, Spain), following the manufacturer’s guidelines. For genomic DNA extraction from serum samples, the MagNA Pure 24 Instrument and total NA Isolation Kit (Roche) were used, a fully automated system based on magnetic glass particle technology (MGP) that allows the processing of up to 24 samples in approximately 70 min. Genotyping of the P72R SNP of TP53 was conducted for all samples by melting curve analysis with a customized LightSNiP assay (TIB MOLBIOL) on a LightCycler 480 II instrument (Roche). Reactions were carried out in a final volume of 20 μL, comprising 2.0 μL of LightCycler FastStart DNA Master HybProbe mix (Roche), 1.0 μL of LightSNiP mix, 3 mM MgCl2, and 50 ng of DNA. The ThermoCycler was set up under the conditions outlined in Supplementary Table S1. Subsequently, the melting curves were assessed using the Melt Curve Genotyping software (Roche).
Control group
To study the association of the P72R SNP with prostate cancer, the frequencies found in the (non-cancer) European (non-Finnish) population from GnomAD v2.1.1, containing 134,187 samples, were used as a control group. In addition, we studied the Spanish Iberian Population from 1000 genomes Project (www.ensembl.org) containing 214 samples.
Gleason score and grade groups
The Gleason score classification system assesses the aggressiveness of prostate tumors based on the architecture of the cancerous tissues and the degree of cell differentiation, as observed under a microscope. This score, which ranges from 6 to 10, is the sum of two histological patterns (the most common plus the highest grade) within a single tissue sample (Supplementary Table S2). To better understand and describe cancer aggressiveness, the Gleason score was adjusted to a grade group encompassing values ranging from one to five. The correlation between the scores and their respective groups is shown in Supplementary Table 2.
Statistics
Associations between frequencies and proportions were analyzed using contingency tables and Fisher’s test. Values were considered statistically significant at P < 0.05. These analyses were conducted using the GraphPad Prism 9 software. Data and image analyses were performed using Circos software (http://circos.ca/).
Results
The P72R SNP frequency in primary prostate tumour cultures and cancer tissues
More than 20 different SNPs, have been described in the TP53 gene (15) including the rs1042522 or P72R variant, at amino acid 72 of p53 (Supplementary Figure S1A). This variant contains a guanine at position 357 (CGC) of the coding region sequence, which changes the encoded amino acid from proline (CCC) to arginine (R or ARG). (Supplementary Figure S1B). Supplementary Figure S1C shows that arginine induced a decrease in hydrophobicity (turquoise bars) relative to the most abundant proline (P or PRO), which may affect the interaction of the p53 protein with other ligands (CCAR2 or HRMT1L2). To study the potential role of this TP53 variant in the aggressiveness of unresectable prostate tumors, we examined 12 primary tumor cultures from hormone-naïve patients (hnPCs). Sanger sequencing revealed the P72R SNP (ARG variant) at a frequency of 82.0% (9/11), whereas the PRO variant frequency was 18.0% (2/11), and the heterozygote samples predominantly expressed ARG (Table 1, Figure 1A). To confirm these results, the hnPCs and, additionally, 11 prostatectomy tissue samples obtained from different PCa patients, were genotyped using gDNA melting curve analysis (Table 1, Figure 1B). In hnPCs, two homozygotes for cytosine (C, PRO), six homozygotes for guanine (G, ARG), and four heterozygotes were found. In the RP tissues, four homozygotes for cytosine, five homozygotes for guanine, and two heterozygotes predominantly expressed ARG. The total frequencies of C and G in primary tumor cultures were 0.33 and 0.67, respectively, while in RP tissues, the frequency of C was 0.45, and that of G was 0.55, indicating that primary tumor cultures have a greater incidence of the P72R allele than tumor tissues from RP, suggesting that the presence of this SNP may help to distinguish inoperable aggressive cancers from the non-metastatic, less aggressive ones that can be cured by surgery.
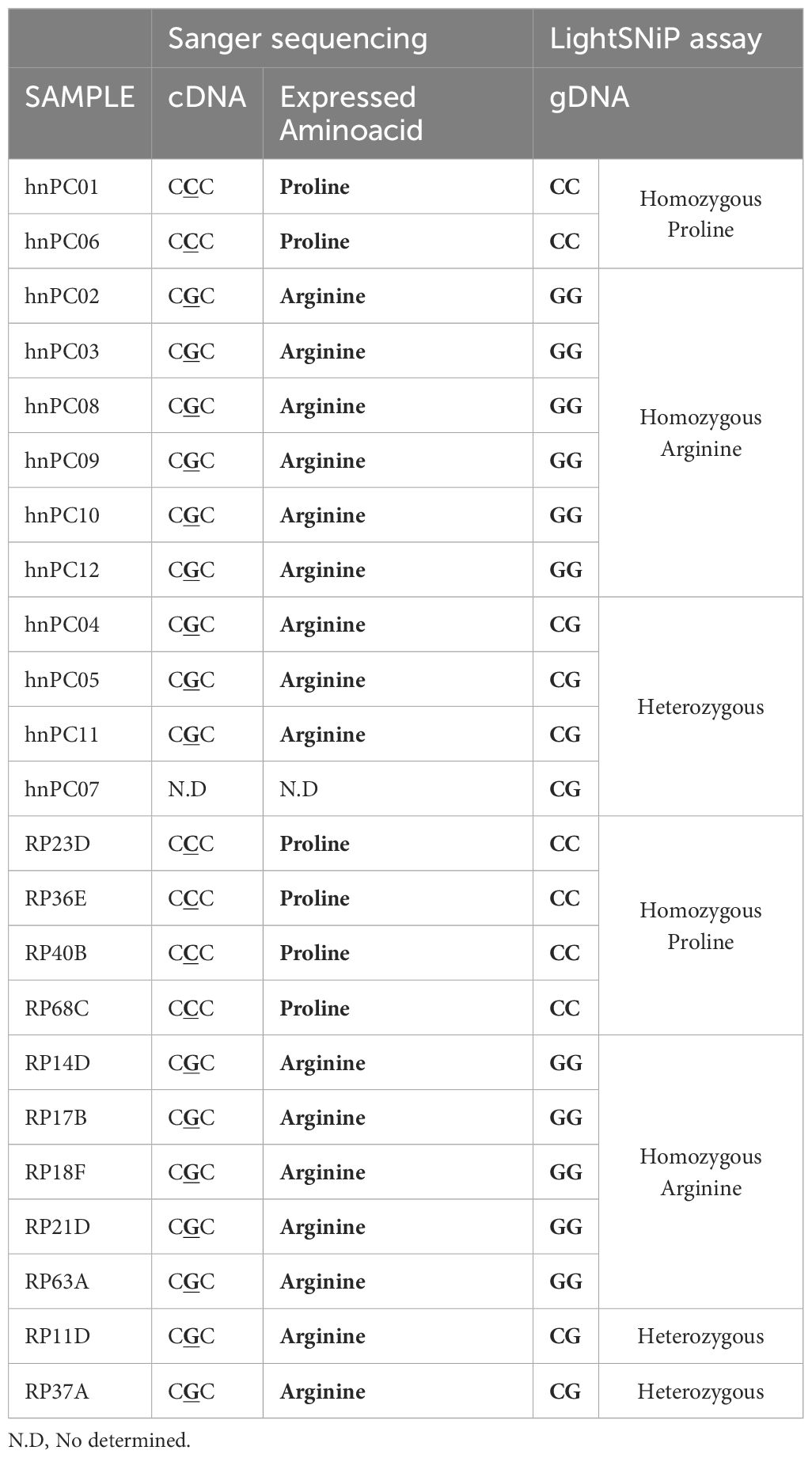
Table 1. P72R genotyping in primary cultures and radical prostatectomy tissues.
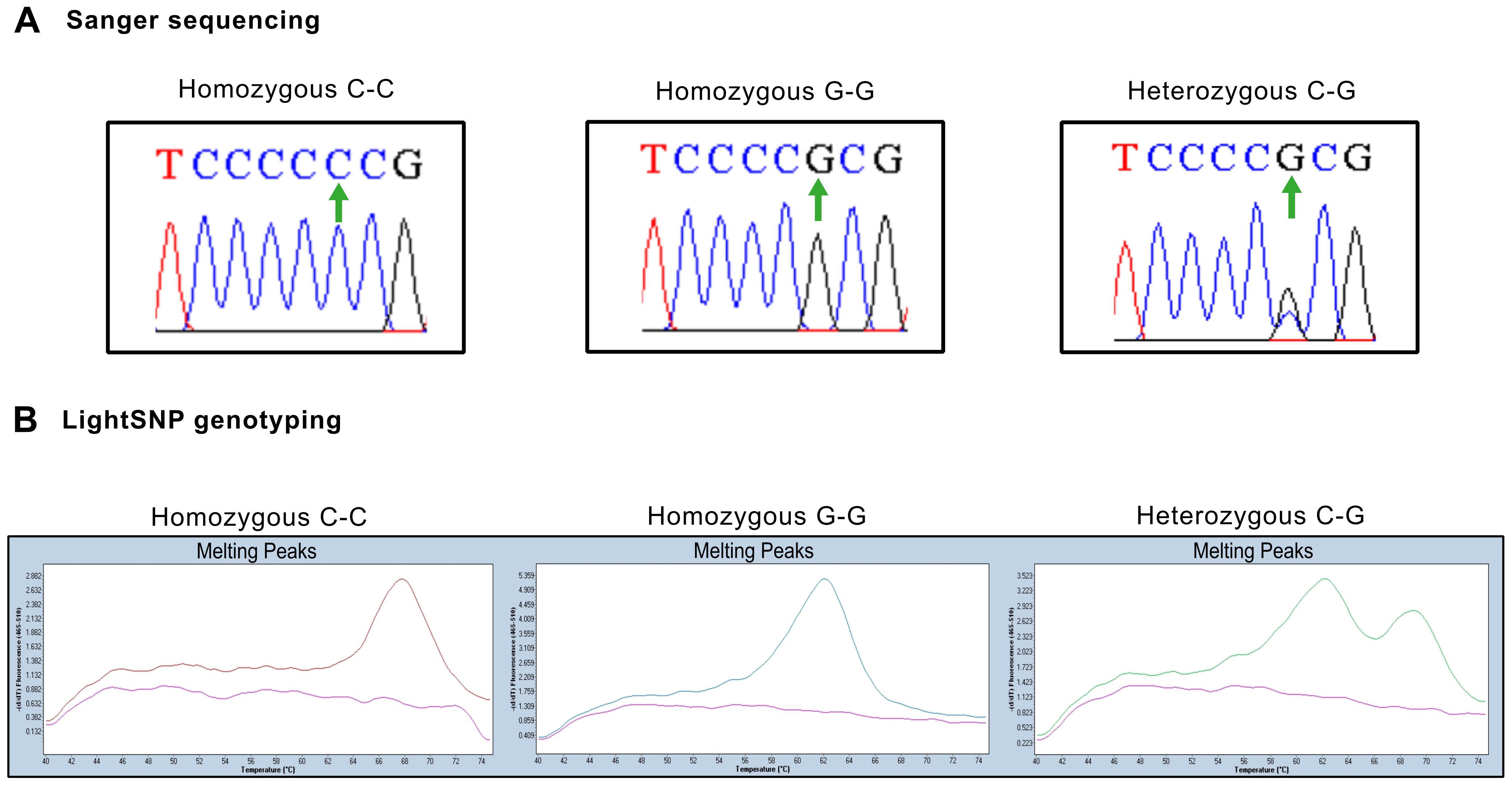
Figure 1. Representative genotyping results for serum and primary culture samples (A) Representative Sanger sequencing chromatograms of homozygous and heterozygous samples. Green arrows indicate position 357 in the coding sequence. (B) Representative melting curves from serum samples illustrating peaks of absorbance at different dissociation temperatures. Left panel: Homozygous samples with cytosine in both strands show a dissociation peak at 68°C. Center panel: Homozygous samples with guanine in both strands show a dissociation peak at 60°C. Right panel: Heterozygous samples with cytosine and guanine residues are identified by both peaks.
The P72R SNP is associated with prostate cancer risk
To validate these results, the serum of 94 patients with prostate cancer was genotyped. P72R is a controversial SNP found at different frequencies across populations worldwide, although the majority of subpopulations exhibit lower frequencies (Figure 2A). Among the serum samples, 53.38% (53/94) were homozygous for ARG (hARG), 8.58% (8/94) for PRO (hPRO), and 35.10% (33/94) were heterozygous, whereas the observed frequency of the G allele was 73.9% and the frequency of the C allele was 26.1% (Figure 2B). Regarding controls, in the selected European (non-Finnish) non-cancer population from GnomAD, the frequency of the G allele was 26.3%, whereas the frequency of the C allele was 73.6%. Similar frequencies were also found in the Spanish Iberian population (Figure 2C). The frequencies in our cohort of serum, hnPC, and RP tissues were then compared with those of the control populations (Figure 2C).The results revealed significant differences in the recurrence rate of the P72R SNP in all samples compared with the control population, suggesting an association between the P72R SNP and prostate cancer.
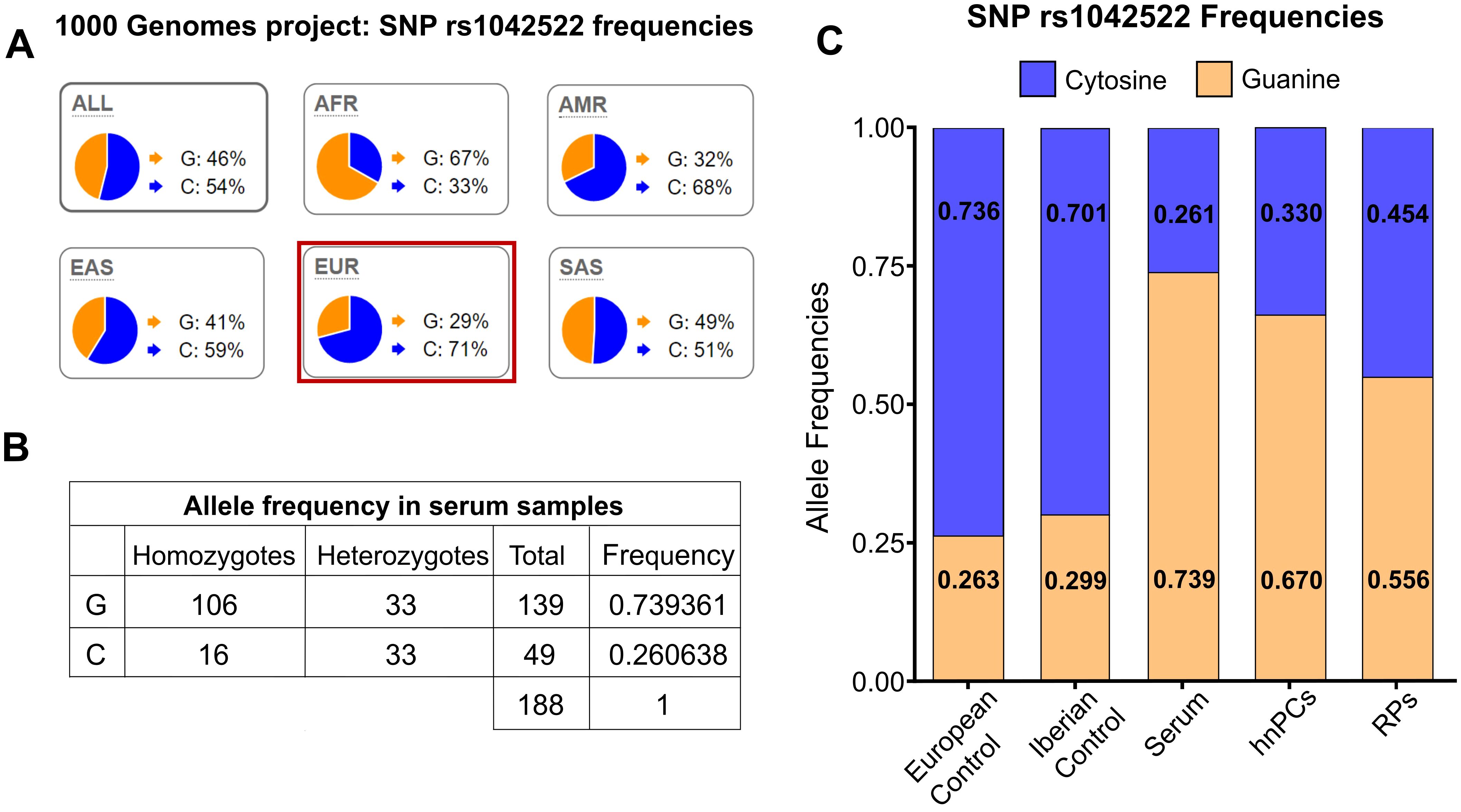
Figure 2. Allele frequencies in the European population and in prostate cancer samples. (A) Global frequency of the rs1042522 SNP. The G allele exhibits a lower frequency in all subpopulations except for the African population. Data and images sourced from Ensemble.org. (B) rs1042522 frequencies in serum samples from patients with prostate cancer. (C) rs1042522 frequencies in primary tumor cultures (hnPCs), serum samples, and radical prostatectomy (RPs) samples compared to those in the control European population and Iberian population.
The significance of this association was assessed by contingency analysis (Fisher´s test), which revealed a significant association between the P72R SNP and prostate cancer in all samples analyzed, with a frequency of 73.93% for the G allele and an odds ratio of 7.937 (95% CI 5.37-11.00) for the serum samples (p<0.0001) (Table 2).
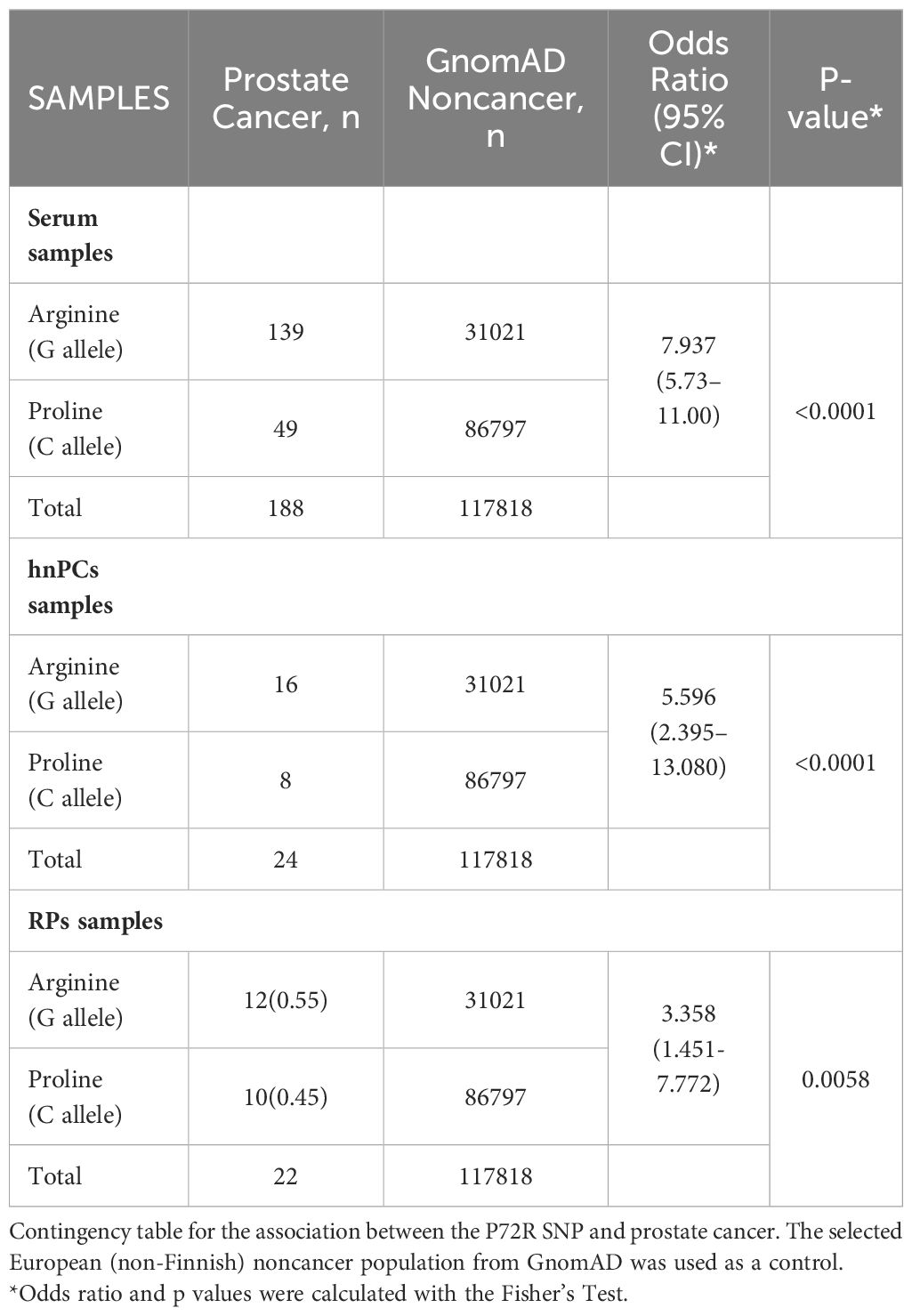
Table 2. Analysis of the association between the rs1042522 variant and prostate cancer.
The P72R SNP was not significantly associated with high Gleason score
The ability of mutant p53 protein to neutralize apoptosis and transform cells in cooperation with EJ-Ras is enhanced when codon 72 encodes ARG (R72) (16). This R72 variant has been reported to bind more efficiently and inactivate PGC-1α (4), a transcriptional target of p53-induced apoptosis in prostate cancer cells, thereby promoting cell invasion and metastasis (4). These observations support our finding that a higher R72 allele frequency is present in more aggressive PCa tumors. To study whether the Gleason Score, or grade (GS) of all genotyped patients was associated with a higher frequency of the R72 allele, we used the Gleason grading group (1 to 5) and divided patients into 2 groups: low-medium aggressive tumors (Gleason grade 1-3), and highly aggressive tumors (Gleason grade 4-5). Patients were genotyped for hARGs, hPROs, or heterozygotes (ARG-PROs). Data from the cohort of 94 serum patients were plotted on a Circos plot according to their Gleason group. Figure 3A shows the distribution of patient proportions for each GS. Although a markedly lower proportion of the P72 allele and a markedly greater frequency of the R72 allele were detected in high Gleason tumors, these differences were not statistically significant (p = 0.2867) (Figures 3A, B).
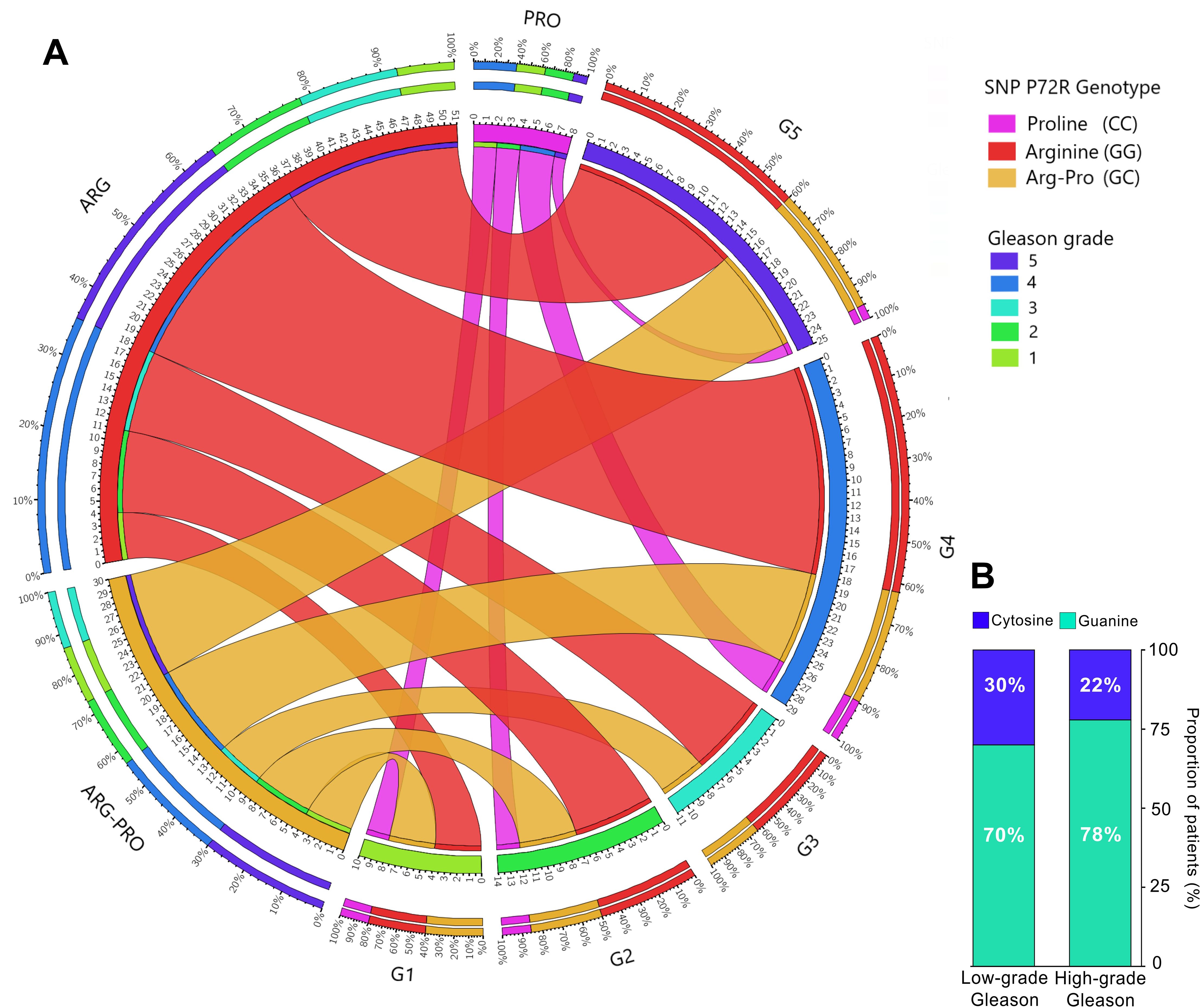
Figure 3. Distribution of the rs1042522 variant in 94 prostate cancer patients according to Gleason grade. (A) Circos plot displaying the distribution of the P72R genotypes according to patient Gleason score. (B) Distribution of the SNPs according to low Gleason grade (1-3) and high Gleason grade (4-5); contingency analysis of these proportions did not yield significant results. Gleason grade 1: Gleason score ≤6; Gleason grade 2-3: Gleason score = 7; Gleason grade 4: Gleason score = 8; Gleason grade 5: Gleason score ≥9.
Discussion
The tumor suppressor gene TP53, reported to be mutated at a high frequency (53.3%) in aggressive metastatic castration resistant prostate cancer, CRPC (4, 17), was the focus of this investigation. The finding that 9 out of 12 primary tumor cultures from patients with aggressive metastatic PCa had a p53 protein with arginine (CGC) at codon 72, but that only 2 cultures had a proline (CCC) was intriguing. An initial explanation for the enrichment of the R72 variant in hnPCs from cancer tissues was the loss of heterozygosity (LOH), particularly in exon 4, which includes codon 72 of the protein. In fact, loss of the proline (C) allele and preferential retention of the arginine (G) allele have been described in primary tumors and metastatic tissues of squamous cell carcinoma of the vulva (18), head and neck (19), colorectal tumors (20), and lung cancer (21). In addition, a recent meta-analysis using the TCGA pan-cancer database and patients selected for P72R heterozygosity reported that 31% (127/409) of heterozygotes had lost the P72 allele in the corresponding tumor tissue (22), indicating that the G allele is preferentially selected for tumor development, possibly due to enhanced p73-induced apoptosis (16) and the modulation of tumor metabolism by regulating PGC-1α (23). Although proper LOH screening was not performed in our cancer cohort, genotyping analysis of serum samples showed a similar frequency of the R72 variant compared to that of hnPCs and cancer tissues from prostatectomy, suggesting that this variant was obtained through germline, instead of LOH.
The results in serum samples, confirmed the high frequency of the G allele, compared to a control European non-cancer population selected from the GnomAD v2.1, indicating a very significant association (p<0.0001) between this SNP and prostate cancer risk, with an odds ratio of 7.937 (95% CI 5.37-11.00). Several reports have described a similarly significant association between the R72 SNP and the risk of developing cutaneous melanoma, breast cancer, and prostate cancer (24–27). A study on PCa compared 187 Iranian patients with 185 control individuals negative for cancer (27). Although the frequency of the G allele in patients with cancer (65.77%) was not as high as the one reported here (73.93%), the difference was statistically significant compared to the control group (p<0.05).
Notably, several reports have shown controversial results (28–30) or failed to demonstrate a significant association between the rs1042522 variant and the risk of any type of cancer (14, 31–34), mostly because of limited sample size or selection bias. For example, in a colorectal cancer study in the Northern European population, no significant association between the presence of the SNP and the risk of cancer was reported (35), but the healthy control samples selected had a high frequency of the ARG allele (61%), in contrast to the estimated 26% frequency of the (non-cancer) European population from GnomAD v2.1.1. Similar to our findings, one study on prostate cancer reported an association between Pro/Pro and a lower risk of prostate cancer (11). However, several other reports have shown conflicting results (11–14, 36–38), suggesting that an appropriate representation of control samples is necessary for accurate results.
To address potential geographical sub-population biases, we further validated our results by analyzing the 1000 Genomes database, filtered by country, and used the Spanish population as a control. In the control population, we observed that the C allele is present at higher frequencies (0.71) compared to the G allele (0.29), similar to the results obtained with the control non-cancer European population from GnomAD (Figure 2C). This indicates that the high frequency of the G allele observed in our serum samples is not attributable to a sub-population effect but it rather reflects its association with aggressive prostate cancer.
These findings underscore the importance of considering population stratification in genetic association studies to avoid potential biases that may skew the results. The use of well-matched control populations is crucial for the accurate interpretation of genetic data. Future studies should continue to account for these variables to ensure the robustness of genetic association findings.
Beyond these conflicting findings regarding ARG frequency at codon 72 of p53, it is crucial to consider the potential impact of this variant on cancer development, although its full understanding is still not clear. Studies in mice have revealed that the presence of the R72 variant is associated with an increased incidence of mammary tumors (39). In addition, this variant is associated with increased phosphorylation of p53 and enhanced transactivation of CDKN1A (p21WAF1) in response to starvation, with consequent augmented growth arrest and reduced apoptosis, a situation that favors survival (40). Furthermore, TP53 mutations tended to preferentially occur in the R72 allele. In the presence of mutant TP53 (e.g., R175H, R273H, and A138V) harboring the R72 SNP, an enhanced capacity for migration, invasion, and metastasis in various cancer cell lines, including PCa, has been reported (23). These observations provide strong evidence that the ARG at codon 72 affects the tumor-suppressive activity of p53.
Although no significant association between the P72R SNP and Gleason score was found in our study, the G allele (R72) was more frequent in patients with a high Gleason score (≥8, group 4-5), suggesting a potential association with more undifferentiated malignant PCa lesions. A few studies have also evaluated the association between this polymorphism and tumor grade and found no significant association with clinical stage or Gleason grade in PCa patients (36, 37, 41, 42). One study revealed a link between the modest influence of this SNP and shorter biochemical recurrence (BCR) after radical prostatectomy (38). Another study in the Japanese population (12) observed a particularly high frequency of the ARG allele in patients with metastases and in those with a high Gleason score (≥8). However, the latter study analyzed the association using the arginine allele as a reference, and the authors calculated the significance level between the Pro/Pro and Pro/Arg genotypes and arginine. As expected, they found no significant differences, as proline did not differ significantly from that in the control group.
To validate and extend our findings, further investigations with a larger sample size and consideration of family history of prostate cancer are needed. Nevertheless, our results suggest that a simple but powerful test (gDNA assessment for the P72R SNP) may be useful for the early identification of patients at risk for aggressive metastatic prostate cancer. Future studies should focus on developing detailed guidelines on how this SNP assessment could be integrated into current prostate cancer screening allowing clinicians to intervene at an early stage.
Data availability statement
Information about the dataset used and/or analyzed during the current study is available in Supplementary Datasheet 1. Additional information is available from the corresponding author upon reasonable request.
Ethics statement
This study was approved by the Medical Research Ethics Committee of Vall d’Hebron Hospital (protocol number: PR(AG) 96/2015). The studies were conducted in accordance with the local legislation and institutional requirements. The participants provided their written informed consent to participate in this study.
Author contributions
ET-G: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Visualization, Writing – original draft. VM: Data curation, Investigation, Methodology, Writing – review & editing. JG: Investigation, Methodology, Writing – review & editing. MS: Methodology, Writing – review & editing, Data curation. AC: Data curation, Writing – review & editing. JM: Resources, Writing – review & editing. IT: Data curation, Writing – review & editing. MG: Data curation, Writing – review & editing. RF: Resources, Writing – review & editing. RP: Conceptualization, Funding acquisition, Supervision, Validation, Writing – original draft, Writing – review & editing.
Funding
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This study was supported by the Ministerio de Ciencia e Innovation (RTI2018-096055-B-I00) and the SGR grant from the Catalan Government (2021 SGR 01173).
Acknowledgments
The authors extend their gratitude to the whole Biochemistry Service of the Hospital Vall d’Hebron, Barcelona, Spain, for their technical support.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fonc.2024.1398411/full#supplementary-material
Abbreviations
SNP, single nucleotide polymorphism; hnPCs, hormone-naïve primary cultures; PSA, prostate-specific antigen; PCa, prostate cancer; DRE, positive digital rectal examination; BCR, biochemical recurrence; PRO, proline; ARG, arginine; RP, radical prostatectomy; LOH, loss of heterozygosity.
References
1. Ferlay J, Ervik M, Colombet M, Mery L, Piñeros M, Znaor A, et al. Global cancer obeservatory: cancer today. (2020). Available online at: http://gco.iarc.fr/today/home.
2. Wang L, Lu B, He M, Wang Y, Wang Z, Du L. Prostate cancer incidence and mortality: global status and temporal trends in 89 countries from 2000 to 2019. Front Public Health. (2022) 10:811044. doi: 10.3389/fpubh.2022.811044
3. The American Cancer Society. Survival rates for Prostate cancer, cancer.org. Atlanta: American cancer society (2020). Available at: http://www.cancer.org/cancer/types/prostate-cancer/detection-diagnosis-staging/survival-rates.html.
4. McCann JJ, Vasilevskaya IA, McNair C, Gallagher P, Neupane NP, de Leeuw R, et al. Mutant p53 elicits context-dependent pro-tumorigenic phenotypes. Oncogene. (2022) 41:444–58. doi: 10.1038/s41388-021-01903-5
5. Arora K, Barbieri CE. Molecular subtypes of prostate cancer. Curr Oncol Rep. (2018) 20:58. doi: 10.1007/s11912-018-0707-9
6. Maxwell KN, Cheng HH, Powers J, Gulati R, Ledet EM, Morrison C, et al. Inherited TP53 variants and risk of prostate cancer. Eur Urology. (2022) 81:243–50. doi: 10.1016/j.eururo.2021.10.036
7. Conti DV, Darst BF, Moss LC, Saunders EJ, Sheng X, Chou A, et al. Trans-ancestry genome-wide association meta-analysis of prostate cancer identifies new susceptibility loci and informs genetic risk prediction. Nat Genet. (2021) 53:65–75. doi: 10.1038/s41588-020-00748-0
8. Wang A, Shen J, Rodriguez AA, Saunders EJ, Chen F, Janivara R, et al. Characterizing prostate cancer risk through multi-ancestry genome-wide discovery of 187 novel risk variants. Nat Genet. (2023) 55:2065–74. doi: 10.1038/s41588-023-01534-4
9. FitzGerald LM, Kumar A, Boyle EA, Zhang Y, McIntosh LM, Kolb S, et al. Germline missense variants in the BTNL2 gene are associated with prostate cancer susceptibility. Cancer Epidemiology Biomarkers Prev. (2013) 22:1520–8. doi: 10.1158/1055-9965.EPI-13-0345
10. Wang NK, Rouette A, Houlahan KE, Yamaguchi TN, Livingstone J, Jung CH, et al. Abstract 4305: Germline structural variants shape prostate cancer clinical and molecular evolution. Cancer Res. (2023) 83:4305. doi: 10.1158/1538-7445.AM2023-4305
11. Henner WD, Evans AJ, Hough KM, Harris EL, Lowe BA, Beer TM. Association of codon 72 polymorphism of p53 with lower prostate cancer risk. Prostate. (2001) 49:263–6. doi: 10.1002/pros.10021
12. Suzuki K, Matsui H, Ohtake N, Nakata S, Takei T, Nakazato H, et al. A p53 codon 72 polymorphism associated with prostate cancer development and progression in Japanese. J BioMed Sci. (2003) 10:430–5. doi: 10.1007/BF02256434
13. Li MS, Liu JL, Wu Y, Wang P, Teng H. Meta-analysis demonstrates no association between p53 codon 72 polymorphism and prostate cancer risk. Genet Mol Res. (2011) 10:2924–33. doi: 10.4238/2011.November.29.3
14. Zhang L, Shao N, Yu Q, Hua L, Mi Y, Feng N. Association between p53 Pro72Arg polymorphism and prostate cancer risk: a meta-analysis. J BioMed Res. (2011) 25:25–32. doi: 10.1016/S1674-8301(11)60003-1
15. Barnoud T, Parris JLD, Murphy ME. Common genetic variants in the TP53 pathway and their impact on cancer. J Mol Cell Biol. (2019) 11:578–85. doi: 10.1093/jmcb/mjz052
16. Marin MC, Jost CA, Brooks LA, Irwin MS, O’Nions J, Tidy JA, et al. A common polymorphism acts as an intragenic modifier of mutant p53 behaviour. Nat Genet. (2000) 25:47–54. doi: 10.1038/75586
17. Robinson D, Van Allen EM, Wu YM, Schultz N, Lonigro RJ, Mosquera JM, et al. Integrative clinical genomics of advanced prostate cancer. Cell. (2015) 161:1215–28. doi: 10.1016/j.cell.2015.05.001
18. Brooks LA, Tidy JA, Gusterson B, Hiller L, O’Nions J, Gasco M, et al. Preferential retention of codon 72 arginine p53 in squamous cell carcinomas of the vulva occurs in cancers positive and negative for human papillomavirus. Cancer Res. (2000) 60:6875–7.
19. Schneider-Stock R, Mawrin C, Motsch C, Boltze C, Peters B, Hartig R, et al. Retention of the arginine allele in codon 72 of the p53 gene correlates with poor apoptosis in head and neck cancer. Am J Pathol. (2004) 164:1233–41. doi: 10.1016/S0002-9440(10)63211-7
20. Schneider-Stock R, Boltze C, Peters B, Szibor R, Landt O, Meyer F, et al. Selective loss of codon 72 proline p53 and frequent mutational inactivation of the retained arginine allele in colorectal cancer. Neoplasia. (2004) 6:529–35. doi: 10.1593/neo.04178
21. Papadakis ED, Soulitzis N, Spandidos DA. Association of p53 codon 72 polymorphism with advanced lung cancer: the Arg allele is preferentially retained in tumours arising in Arg/Pro germline heterozygotes. Br J Cancer. (2002) 87:1013–8. doi: 10.1038/sj.bjc.6600595
22. De Souza C, Madden J, Koestler DC, Minn D, Montoya DJ, Minn K, et al. Effect of the p53 P72R polymorphism on mutant TP53 allele selection in human cancer. J Natl Cancer Inst. (2021) 113:1246–57. doi: 10.1093/jnci/djab019
23. Basu S, Gnanapradeepan K, Barnoud T, Kung CP, Tavecchio M, Scott J, et al. Mutant p53 controls tumor metabolism and metastasis by regulating PGC-1a. Genes Dev. (2018) 32:230. doi: 10.1101/gad.309062.117
24. Shen H, Liu Z, Strom SS, Spitz MR, Lee JE, Gershenwald JE, et al. p53 codon 72 arg homozygotes are associated with an increased risk of cutaneous melanoma. J Invest Dermatol. (2003) 121:1510–4. doi: 10.1046/j.1523-1747.2003.12648.x
25. Volodko N, Salla M, Eksteen B, Fedorak RN, Huynh HQ, Baksh S. TP53 codon 72 Arg/Arg polymorphism is associated with a higher risk for inflammatory bowel disease development. World J Gastroenterol. (2015) 21:10358–66. doi: 10.3748/wjg.v21.i36.10358
26. Dia Y, Diop DJP, Ndiaye R, Dem A, Diouf D, Dieng MM, et al. Arg allele of P53 gene codon72 is a risk factor for breast cancer in Senegalese women. J Mol Genet Med. (2017) 11:300–4. doi: 10.4172/1747-0862.1000300
27. Doosti A, Dehkordi PG. The p53 codon 72 polymorphism and association to prostate cancer in Iranian patients. Afr J Biotechnol. (2011) 10:12821–5. doi: 10.5897/AJB
28. Ezzikouri S, El feydi AE, Chafik A, Benazzouz M, El kihal L, Afifi R, et al. The Pro variant of the p53 codon 72 polymorphism is associated with hepatocellular carcinoma in Moroccan population. Hepatol Res. (2007) 37:748–54. doi: 10.1111/j.1872-034X.2007.00126.x
29. Mohana Devi S, Balachandar V, Arun M, Suresh Kumar S, Balamurali Krishnan B, Sasikala K. Analysis of genetic damage and gene polymorphism in hepatocellular carcinoma (HCC) patients in a South Indian population. Dig Dis Sci. (2013) 58:759–67. doi: 10.1007/s10620-012-2409-8
30. Son MS, Jang MJ, Jeon YJ, Kim WH, Kwon CI, Ko KH, et al. Promoter polymorphisms of pri-miR-34b/c are associated with hepatocellular carcinoma. Gene. (2013) 524:156–60. doi: 10.1016/j.gene.2013.04.042
31. Tang NP, Wu YM, Wang B, Ma J. Systematic review and meta-analysis of the association between P53 codon 72 polymorphism and colorectal cancer. Eur J Surg Oncol (EJSO). (2010) 36:431–8. doi: 10.1016/j.ejso.2010.03.010
32. Weng Y, Lu L, Yuan G, Guo J, Zhang Z, Xie X, et al. p53 codon 72 polymorphism and Hematological Cancer Risk: An Update Meta-Analysis. PloS One. (2012) 7:e45820. doi: 10.1371/journal.pone.0045820
33. Ye J, Li XF, Wang YD, Yuan Y. Arg72Pro polymorphism of TP53 gene and the risk of skin cancer: a meta-analysis. PloS One. (2013) 8:e79983. doi: 10.1371/journal.pone.0079983
34. Xu T, Xu ZC, Zou Q, Yu B, Huang XE. P53 Arg72Pro polymorphism and bladder cancer risk–meta-analysis evidence for a link in Asians but not Caucasians. Asian Pac J Cancer Prev. (2012) 13:2349–54. doi: 10.7314/APJCP.2012.13.5.2349
35. Krüger S, Bier A, Engel C, Mangold E, Pagenstecher C, Doeberitz M von K, et al. The p53 codon 72 variation is associated with the age of onset of hereditary non-polyposis colorectal cancer (HNPCC). J Med Genet. (2005) 42:769–73. doi: 10.1136/jmg.2004.028506
36. Lu Y, Liu Y, Zeng J, He Y, Peng Q, Deng Y, et al. Association of p53 codon 72 polymorphism with prostate cancer: an update meta-analysis. Tumor Biol. (2014) 35:3997–4005. doi: 10.1007/s13277-014-1657-y
37. Sivoňová MK, Vilčková M, Kliment J, Mahmood S, Jurečeková J, Dušenková S, et al. Association of p53 and p21 polymorphisms with prostate cancer. Biomed Rep. (2015) 3:707–14. doi: 10.3892/br.2015.496
38. Duncan A, Nousome D, Ricks R, Kuo HC, Ravindranath L, Dobi A, et al. Association of TP53 single nucleotide polymorphisms with prostate cancer in a racially diverse cohort of men. Biomedicines. (2023) 11:1404. doi: 10.3390/biomedicines11051404
39. Gunaratna RT, Santos A, Luo L, Nagi C, Lambertz I, Spier M, et al. Dynamic role of the codon 72 p53 single-nucleotide polymorphism in mammary tumorigenesis in a humanized mouse model. Oncogene. (2019) 38:3535–50. doi: 10.1038/s41388-018-0630-4
40. Kung CP, Liu Q, Murphy ME. The codon 72 polymorphism of p53 influences cell fate following nutrient deprivation. Cancer Biol Ther. (2017) 18:484–91. doi: 10.1080/15384047.2017.1323595
41. Babaei F, Ahmadi SA, Abiri R, Rezaei F, Naseri M, Mahmoudi M, et al. The TP53 codon 72 polymorphism and risk of sporadic prostate cancer among Iranian patients. Iran J Public Health. (2014) 43:453–9.
42. Aydin M, Bozkurt A, Cikman A, Gulhan B, Karabakan M, Gokce A, et al. Lack of evidence of HPV etiology of prostate cancer following radical surgery and higher frequency of the Arg/Pro genotype in Turkish men with prostate cancer. Int Braz J Urol. (2017) 43:36–46. doi: 10.1590/s1677-5538.ibju.2015.0429
Keywords: TP53, SNP, rs1042522, prostate cancer, European population
Citation: Toscano-Guerra E, Maggio V, García J, Semidey ME, Celma A, Morote J, de Torres I, Giralt M, Ferrer-Costa R and Paciucci R (2024) Association of the rs1042522 SNP with prostate cancer risk: a study of cancer tissues, primary tumor cultures, and serum samples from a Spanish Caucasian population. Front. Oncol. 14:1398411. doi: 10.3389/fonc.2024.1398411
Received: 09 March 2024; Accepted: 22 July 2024;
Published: 13 August 2024.
Edited by:
Hung-Ming Lam, University of Washington, United StatesReviewed by:
Wensheng Zhang, Xavier University of Louisiana, United StatesYupeng Wu, First Affiliated Hospital of Fujian Medical University, China
Copyright © 2024 Toscano-Guerra, Maggio, García, Semidey, Celma, Morote, de Torres, Giralt, Ferrer-Costa and Paciucci. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Rosanna Paciucci, cm9zcGFjYmFyQGdtYWlsLmNvbQ==; Emily Toscano-Guerra, ZW1hLnRnMjdAZ21haWwuY29t