
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Oncol. , 31 May 2023
Sec. Cancer Immunity and Immunotherapy
Volume 13 - 2023 | https://doi.org/10.3389/fonc.2023.1170502
 Anastasios D. Giannou1,2,3,4*†
Anastasios D. Giannou1,2,3,4*† Jan Kempski1,2,5†
Jan Kempski1,2,5† Tao Zhang1,2†
Tao Zhang1,2† Jöran Lücke1,2,3†Ahmad Mustafa Shiri1,2
Jöran Lücke1,2,3†Ahmad Mustafa Shiri1,2 Dimitra E. Zazara6,7
Dimitra E. Zazara6,7 Ioannis Belios7Andres Machicote1,2
Ioannis Belios7Andres Machicote1,2 Philipp Seeger3Theodora Agalioti3
Philipp Seeger3Theodora Agalioti3 Joseph Tintelnot5,8Adrian Sagebiel3
Joseph Tintelnot5,8Adrian Sagebiel3 Miriam Tomczak1,2Lennart Bauditz1,2Tanja Bedke1,2
Miriam Tomczak1,2Lennart Bauditz1,2Tanja Bedke1,2 Lorenz Kocheise1,2Baris Mercanoglu3
Lorenz Kocheise1,2Baris Mercanoglu3 Mohammad Fard-Aghaie3
Mohammad Fard-Aghaie3 Emmanouil Giorgakis9,10
Emmanouil Giorgakis9,10 Panagis M. Lykoudis11,12Anastasia Pikouli11Julia-Kristin Grass3Ramez Wahib3Jan Bardenhagen3
Panagis M. Lykoudis11,12Anastasia Pikouli11Julia-Kristin Grass3Ramez Wahib3Jan Bardenhagen3 Benjamin Brunswig3Asmus Heumann3
Benjamin Brunswig3Asmus Heumann3 Tarik Ghadban3Anna Duprée3Michael Tachezy3Nathaniel Melling3
Tarik Ghadban3Anna Duprée3Michael Tachezy3Nathaniel Melling3 Petra C. Arck7
Petra C. Arck7 Pablo Stringa13
Pablo Stringa13 Maria Virginia Gentilini14Gabriel E. Gondolesi13Ryosuke Nakano15
Maria Virginia Gentilini14Gabriel E. Gondolesi13Ryosuke Nakano15 Angus W. Thomson15,16Daniel Perez3Jun Li3Oliver Mann3
Angus W. Thomson15,16Daniel Perez3Jun Li3Oliver Mann3 Jakob R. Izbicki3
Jakob R. Izbicki3 Nicola Gagliani2,3Ioannis C. Maroulis4‡
Nicola Gagliani2,3Ioannis C. Maroulis4‡ Samuel Huber1,2*‡
Samuel Huber1,2*‡Background: The immune system plays a pivotal role in cancer progression. Interleukin 22 binding protein (IL-22BP), a natural antagonist of the cytokine interleukin 22 (IL-22) has been shown to control the progression of colorectal cancer (CRC). However, the role of IL-22BP in the process of metastasis formation remains unknown.
Methods: We used two different murine in vivo metastasis models using the MC38 and LLC cancer cell lines and studied lung and liver metastasis formation after intracaecal or intrasplenic injection of cancer cells. Furthermore, IL22BP expression was measured in a clinical cohort of CRC patients and correlated with metastatic tumor stages.
Results: Our data indicate that low levels of IL-22BP are associated with advanced (metastatic) tumor stages in colorectal cancer. Using two different murine in vivo models we show that IL-22BP indeed controls the progression of liver but not lung metastasis in mice.
Conclusions: We here demonstrate a crucial role of IL-22BP in controlling metastasis progression. Thus, IL-22 might represent a future therapeutic target against the progression of metastatic CRC.
An increasing number of studies highlight the role of the immune system and the tumor microenvironment in cancer progression (1–4). Notably, immune-checkpoint inhibitors, which aim to activate the anti-tumor immune response, are among the most promising new cancer therapies. However, specific immune responses in chronic inflammation can also promote tumor development. Understanding the mechanisms that control the interaction between the immune system and cancer cells is pivotal for the generation of future, more effective immunotherapies.
The pro-tumorigenic effects of the cytokine interleukin 22 (IL-22) have gained increasing attention in recent years. Although IL-22 can have many beneficial functions such as promoting intestinal integrity, supporting the mucosal barrier function and protecting against genotoxic stress (5–9), it can also act on cancer cells directly and fuel cancer progression (7, 10–14). Those dichotomic effects of IL-22 highlight the importance of a tight control of its activity. Interestingly, IL-22 activity is controlled by another protein called IL-22 binding protein (IL-22BP, IL22RA2), which binds IL-22 and prevents it from signaling through its IL-22RA1 – IL-10R2 heterodimeric receptor (15, 16).
The primary cellular sources of IL-22BP can be different depending on the underlying disease. In the case of inflammatory bowel disease (IBD), CD4+ T cells, dendritic cells and eosinophils have been identified as the primary sources of IL-22BP (17, 18). In contrast, dendritic cells (DCs) are the major source of IL-22BP in the healthy intestine and in colorectal cancer (CRC) (11, 19–21). The regulation of IL-22BP expression is still incompletely understood. While TNFα has been shown to regulate IL-22BP in CD4+ T cells in chronic intestinal inflammation, this cytokine does not regulate IL-22BP expression in DCs (18). One study found that IL-22BP production by monocyte-derived DCs is regulated by retinoic acid (19). Recently, our research group as well as others have shown a crucial role of lymphotoxin signaling in the production of IL-22BP by DCs (20, 21).
IL-22BP has been shown to regulate tumorigenesis in the intestine in mice and humans (11, 20). Indeed, high IL-22BP levels in the primary tumors of patients with colorectal cancer (CRC) are associated with a favorable clinical outcome in terms of the overall survival (20). The presence of distant metastasis is a crucial factor influencing the outcome of patients with CRC. However, it is currently unknown how IL-22BP affects CRC-related metastasis formation and progression. Similarly, the cellular sources and the regulation of IL-22BP in CRC metastasis remain unclear.
Here, we describe a down-regulation of IL-22BP expression in the primary tumors of patients with metastatic CRC compared to patients without lymph node involvement or distant metastasis. In parallel, the expression of IL-22BP is increased in liver metastasis compared to normal liver tissue. Using mouse models, we could show that IL-22BP protects mice from the progression of liver but not lung metastasis. Furthermore, we describe DCs as the primary cellular source of IL-22BP in liver metastasis. Collectively, our data point towards a crucial role of IL-22BP not only in the development of the primary tumor but also in the metastatic process.
We have previously published an association between high IL-22BP levels in the primary tumor and longer survival in patients with colorectal cancer (20). Using the same data set, we again divided the patients based on the expression of IL-22BP into patients with high and low expression of this protein (Figure 1A). We found a statistically significant relationship between IL-22BP levels and the nodal and metastasis staging of tumors (based on the TNM classification, Figures 1B, C). In general, patients with cancers limited to the primary tumor location (i.e., no lymph node involvement or distant metastasis, UICC stage I-II) showed a higher IL-22BP expression than patients in which the tumor had already spread to lymph nodes or distant organs (UICC stage III-IV, Figure 1D). Importantly, we could confirm this association between UICC tumor stage and IL-22BP expression levels using the publicly available The Cancer Genome Atlas (TCGA) COAD (Colon adenocarcinoma) dataset (Figure 1E) (22).
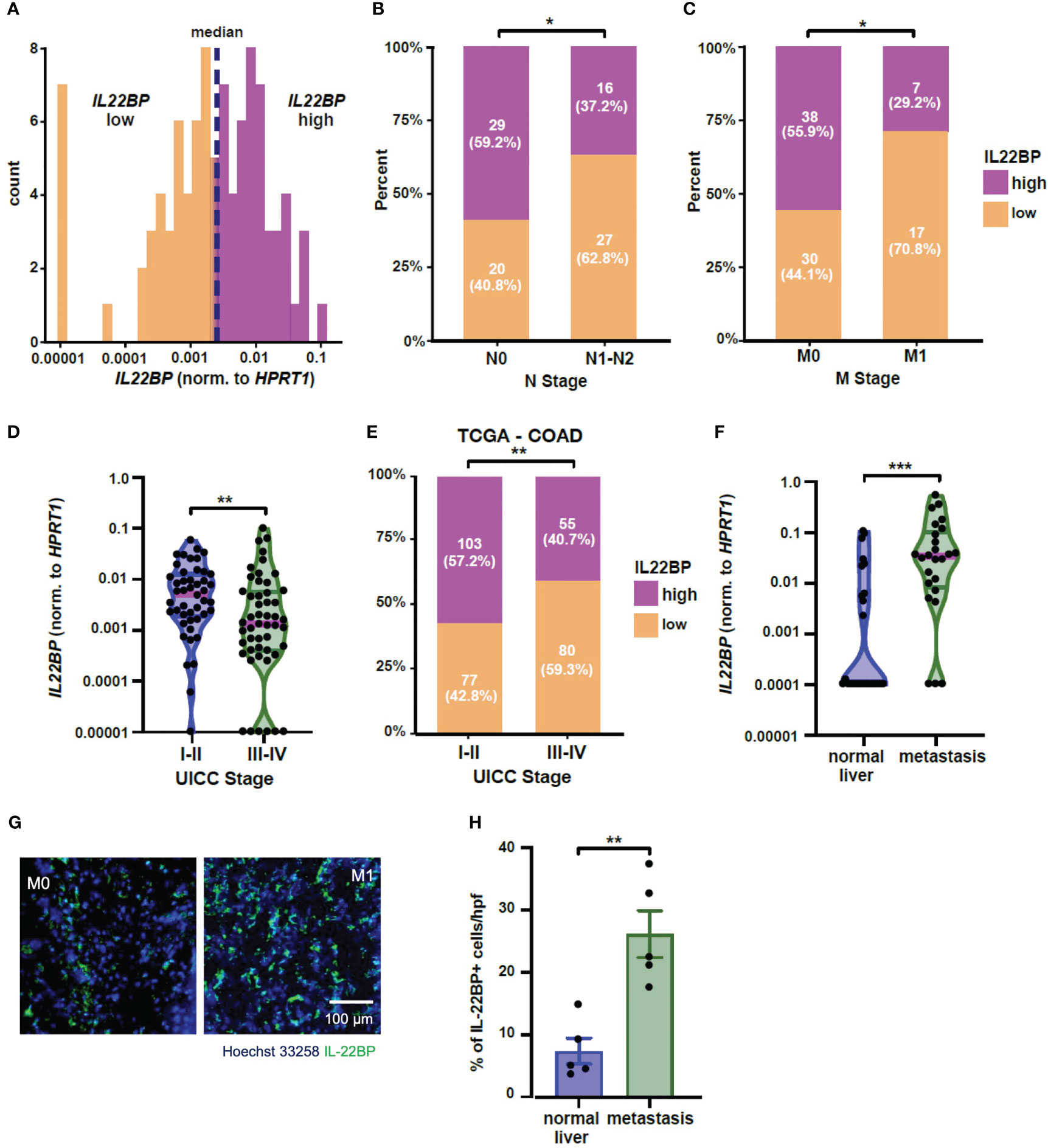
Figure 1 IL-22BP expression is upregulated in liver metastasis. (A) Histogram showing the expression levels of IL22BP in patients with colorectal cancer and the grouping into high and low IL22BP expression using the median value (n=96). (B) Bar graph comparing the distribution of high and low IL22BP expression in the primary tumor of patients with (N1-N2) or without (N0) lymph node metastasis (n=92). (C) Bar graph comparing the distribution of high and low IL22BP expression in the primary tumor of patients with (M1) or without (M0) distant metastasis (n=92). (D) Violin plot of the expression of IL22BP in the primary tumor of patients with early (UICC I-II) or advanced-stage (UICC III-IV) CRC (n=92). (E) Bar graph comparing the distribution between high and low IL22BP expression and UICC Stage in the publicly available TCGA-COAD dataset. (F) Violin plot of the expression of IL22BP in the normal liver tissue (n=27) and liver metastasis (n=25). (G) Representative immunofluorescence picture showing the presence of IL-22 and IL-22BP in normal liver tissue and liver metastasis. (H) The summarizing statistics of the immunofluorescence analysis of IL-22BP+ cells in normal liver tissue and liver metastasis (n = 5). The data presented in (D) and (F) are the median and interquartile ranges. * = P < 0.05; ** = P<0.01; *** = P < 0.001 as assessed by chi-square test (B, C, E) and Mann-Whitney U test (D, F–H).
Next, we analyzed the expression of IL-22BP in another CRC-patient cohort who underwent resection of their liver metastases. Interestingly, our data showed an upregulation of IL-22BP expression in the liver metastasis compared to the expression in healthy liver parenchyma of those patients (Figure 1F). Those findings could be confirmed on a protein level using immunofluorescence (Figures 1G, H).
In summary, our data indicate a down-regulation of IL-22BP in the primary tumors of patients with advanced-stage CRC. However, the expression of this protein is in parallel increased in the liver metastasis compared to the healthy liver. Based on this data, we hypothesized a crucial role of IL-22BP in the process of CRC metastasis formation.
Our human data suggested a role of IL-22BP in the formation of CRC-related liver metastasis. In the next step, we aimed to test, if IL-22BP plays indeed a causative role in the metastatic process. To this end, we performed an intrasplenic injection of the murine colon carcinoma cell line MC38 in wild type (WT, Il22bp+/+) and Il22bp-deficient (Il22bp-/-) mice and compared the generation of liver metastasis 21 days later (Figure 2A), as done previously (23, 24). Interestingly, Il22bp-deficient mice showed an increased number of macroscopic liver metastases and histologically present metastatic foci (Figures 2B-D). Those data indicate a protective effect of IL-22BP on the formation of liver metastasis.
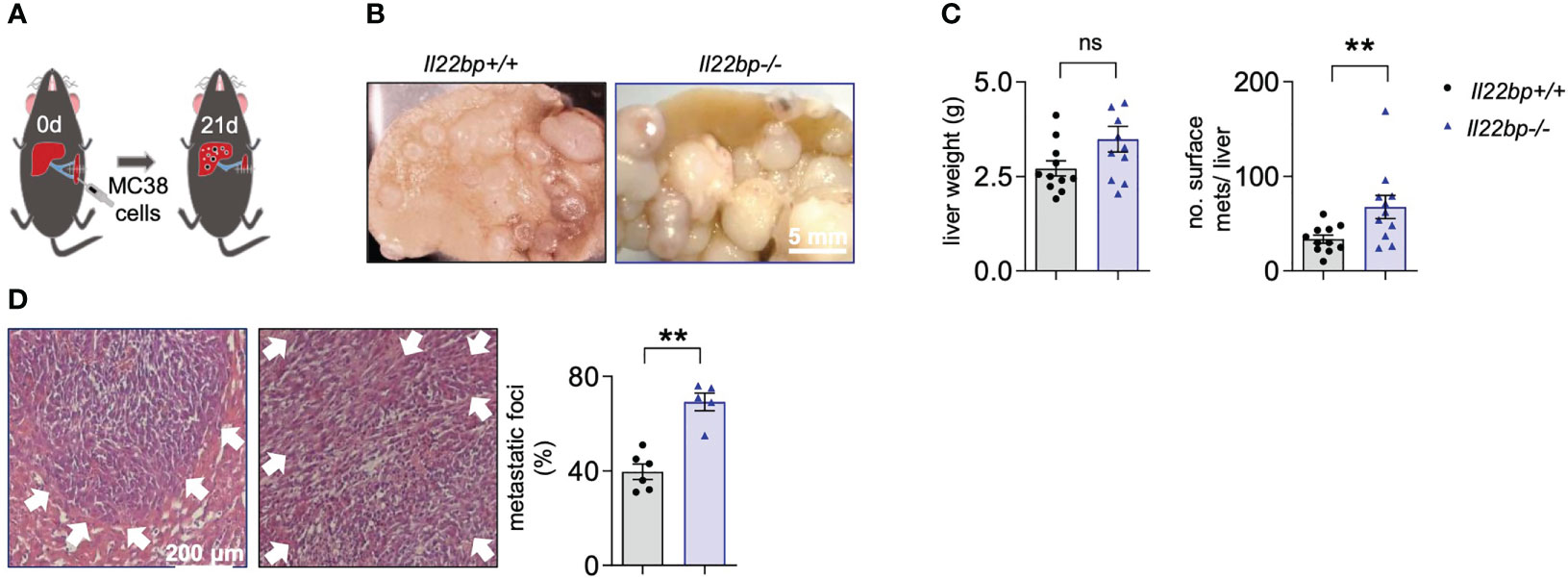
Figure 2 IL-22BP controls the formation of liver metastasis. (A) Schematic overview of intrasplenic injection of MC38 cells for forced liver metastasis induction. (B) Representative macroscopic pictures of liver metastases in Il22bp+/+ and Il22bp-/- mice (C) Number of macroscopic liver metastases and liver weight in Il22bp+/+ and Il22bp-/- mice. (D) Representative microscopic pictures and the corresponding statistics showing the microscopic liver metastases in Il22bp+/+ and Il22bp-/- mice as a percentage of the total liver parenchyma. Data presented as mean ± SEM. ** = P<0.01; as assessed by Mann-Whitney U test. ns =p>0.05. White arrows show the boundaries of microscopic liver metastasis.
In order to verify those results in another setting, we employed the model of intracaecal injection of the Lewis Lung Carcinoma (LLC) cell line and liver metastasis formation was assessed 28 days after the injection (Figure 3A). Indeed, 28 days after the intracaecal injection of the tumor cells, the liver weight and the number of liver metastases was increased in Il22bp-/- mice compared to wild type (Il22bp+/+) mice (Figures 3B-D). Thus, these results confirm that IL-22BP controls the formation of liver metastasis in mice.
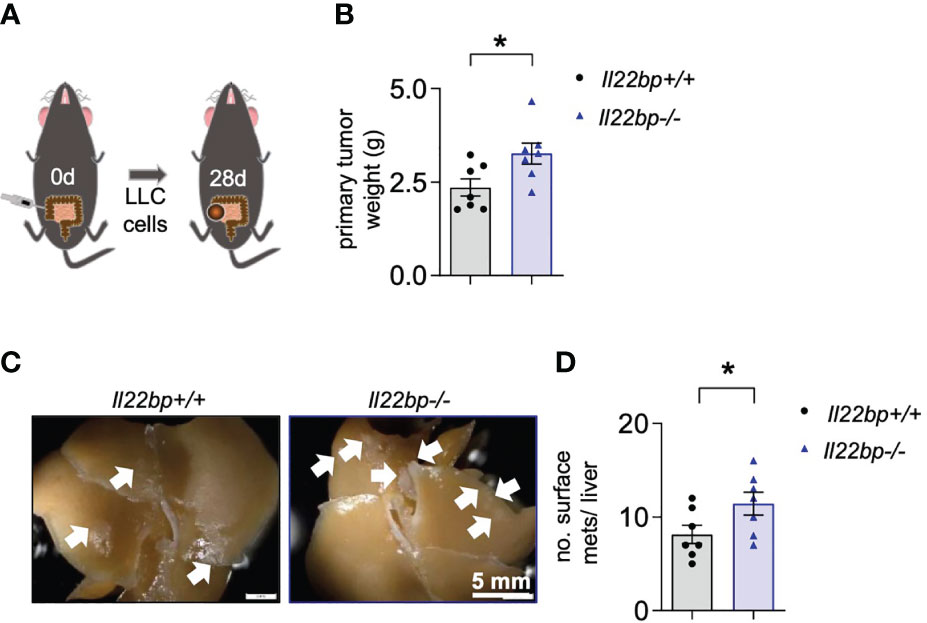
Figure 3 IL-22BP controls the formation of liver metastasis after intracaecal injection of LLC cells. (A) Schematic overview of the intracaecal injection of LLC cells for spontaneous liver metastasis induction. (B) Primary tumor weight of Il22bp+/+ and Il22bp-/- mice 28 days after LLC cell injection (C) Representative macroscopic pictures of liver metastases in Il22bp+/+ and Il22bp-/- mice. (D) Number of liver metastases in Il22bp+/+ and Il22bp-/- mice. Data presented as mean ± SEM. * = p≤0.05 as assessed by Mann-Whitney U test. Arrows show the macroscopic liver metastases.
Finally, we aimed to test whether this effect is specific for the seeding of cancer cells into the liver or whether it is a broad mechanism which also applies to metastasis formation into other target organs. To this end, we injected LLC cancer cells into the flank of the mice and assessed lung metastasis formation after 35 days (Figure 4A). In contrast to the phenotype observed in the liver, the number of lung metastases was not different between Il22bp+/+ and Il22bp-/- mice (Figures 4B, C). Interestingly, the expression level of Il22bp was higher in lung metastasis compared to liver metastasis (Figure 4D). Taken together, our results indicate that the absence of IL-22BP favors the development of liver but not lung metastasis and that the metastasis-controlling functions of IL-22BP are dependent on the organ-specific microenvironment.

Figure 4 IL-22BP does not influence lung metastasis formation. (A) Schematic overview of flank injection of LLC cells for spontaneous lung metastasis induction. (B) Representative macroscopic pictures of lung metastases in Il22bp+/+ and Il22bp-/- mice. (C) Number of lung metastases in Il22bp+/+ and Il22bp-/- mice. (D) Il22bp mRNA expression in lung and liver metastasis (n=7). Data presented as mean ± SEM. ns =p>0.05; *** =p<0.001 as assessed by Mann-Whitney U test. White arrows show the macroscopic lung metastases.
Different cell types have been reported to produce IL-22BP. Both CD4+ T cells and eosinophils have been described as relevant sources of IL-22BP in the inflamed colon of patients with IBD (17, 18). In contrast, DCs are the major source of IL-22BP in the healthy intestine and in CRC (11, 19, 20). To identify the cellular source of IL-22BP, we intrasplenically injected wild-type mice with MC38 cancer cells (Figure 5A). After 21 days, the mice were sacrificed and we isolated leukocytes from livers with metastasis and from healthy livers of control mice that were not injected with MC38 cancer cells (and thus did not develop liver metastasis). Il22bp expression was analyzed after cellular sorting of T cells, DCs, CD11b+Ly6C+ and CD11b+Ly6G+ cells. Our results demonstrate an increased mRNA expression of Il22bp in DCs in liver metastasis. In contrast, the mRNA expression of Il22bp did not change in the case of CD4+ T cells, CD8+ T cells, CD11b+Ly6C+ and CD11b+Ly6G+ cells. In general, CD11b+Ly6G+, DCs and CD11b+Ly6C cells produced the highest levels of IL-22BP in liver metastasis (Figure 5B). We thus conclude that DCs are not only the major cellular source of IL-22BP in primary CRC tumors, but also are one of the major cellular sources of IL-22BP in CRC-derived liver metastasis.
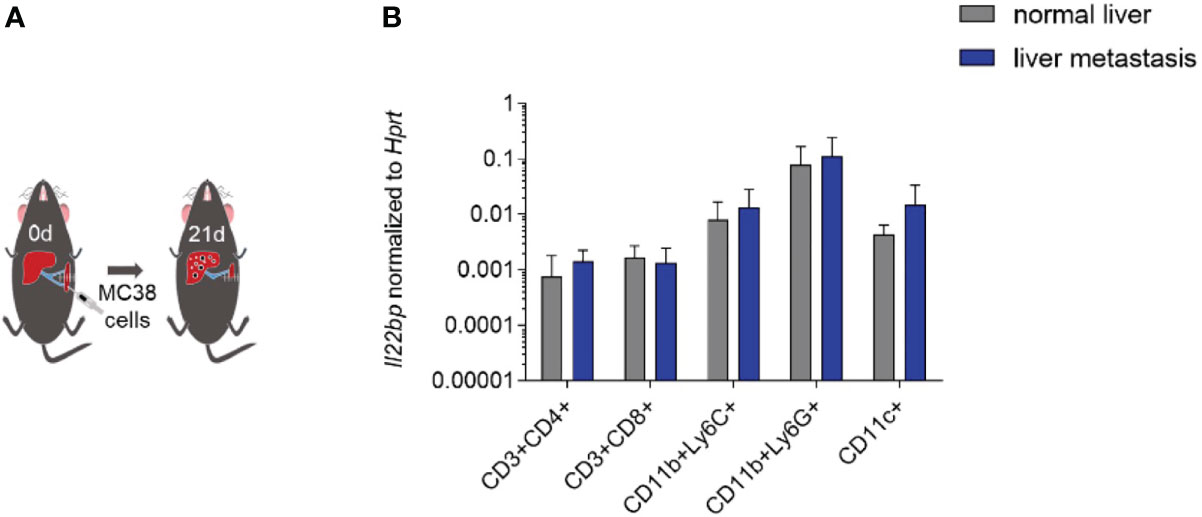
Figure 5 IL-22BP is produced by DCs in liver metastasis. (A) Schematic overview of intraplenic injection of MC38 cells for forced liver metastasis induction. (B) Il22bp expression in CD4+ T cells, CD8+ T cells, CD11c+ cells, CD11b+ Ly6C+ cells and CD11b+Ly6G+ cells. Results of 2 experiments; in each experiment sorted cells were pooled from 5 mice in each group.
Metastasis formation is a multi-step process which culminates in the extravasation of circulating tumor cells into the target organ and the subsequent development of a metastatic niche. The proliferation of the metastatic cells then leads to the formation of established metastases in the target organ. We have recently shown that IL-22 plays a crucial role in the extravasation of cancer cells into the liver parenchyma (25). In order to test if IL-22BP plays also a role in cancer cell extravasation into the liver, we performed an in vivo extravasation experiment. Specifically, we injected GFP-labelled MC38 cells intrasplenically and counted the number of cells extravasating into the liver after 24 hours using flow cytometry (Figure 6A). Notably, we found no difference in the number of extravasating cancer cells between wild type and Il22bp-deficient mice (Figure 6B). This could be explained by the physiologically very low levels of IL-22BP in steady state conditions in the liver. Indeed, our human data have shown that IL-22BP gets upregulated in already established macro-metastases.

Figure 6 IL-22BP controls the proliferation of metastatic cells but not the extravasation process (A) Schematic overview of intrasplenic injection of GFP-labelled MC38 cells for forced liver metastasis induction. (B) FACS plots and the corresponding statistics showing the number of GFP+ extravasated MC38 cells in the liver parenchyma of Il22bp+/+ and Il22bp-/- mice. (C) Representative microscopic pictures and the corresponding statistics of immunohistochemical analysis of Ki-67+ cells in liver metastasis of Il22bp+/+ and Il22bp-/- mice. Data presented as mean ± SEM. Data presented as mean ± SEM. ns>0.05; * = p≤0.05 as assessed by Mann-Whitney U test. White arrows point to areas of proliferation rich in Ki-67+ cells.
To test if IL-22BP controls the proliferation of metastatic cells that have already extravasated into the liver, we performed a Ki-67 staining of livers isolated from in the injected wild type and Il22bp-deficient mice. Interestingly, we found an increased percentage of proliferating tumor cells in Il22bp-deficient mice compared to wild type mice (Figure 6C). Overall, our data indicate that IL-22BP does not control the early process of cancer cell extravasation into the liver but rather controls the proliferation of tumor cells in already established liver metastasis.
Tumor development is not only influenced by intrinsic genetic changes in the cancer cells but also by the immunological tumor microenvironment (1, 2, 26–28). One factor that has been identified to play a crucial role in the progression of colorectal cancer is IL-22. IL-22 expression is upregulated in CRC and is associated with chemotherapy resistance (10, 11, 13, 29). Notably, the activity of IL-22 is regulated by IL-22BP, a high-affinity, soluble IL-22 receptor that prevents IL-22 from binding to its membrane-bound IL-22 receptor (IL-22RA1) (15, 16). We have recently shown a crucial role of IL-22BP in the development of primary tumors in the intestine in mice and humans (20). In the current study, we examined whether IL-22BP regulates the metastatic cascade in CRC since the prognosis of patients with colorectal cancer is largely dependent on the presence of metastasis. However, due to limited understanding of the mechanisms underlying the metastatic cascade, there are no therapies available that specifically inhibit metastasis formation or progression.
Interestingly, we found that IL-22BP is downregulated in the primary tumors of advanced-stage CRC. Of note, our study uses a relatively small sample size of only 96 patients with CRC for cytokine expression analysis, which may not be representative of the larger population. We have thus extended our study by analyzing the COAD Dataset of The Cancer Genome Atlas (TCGA) (22). Specifically, we compared whether patients with high and low IL-22BP Expression are differentially distributed between patients with UICC Stage I-II (i.e. tumor growth limited to the primary tumor side and UICC Stage III-IV (i.e. dissemination into locoregional lymph nodes or distant organs). Indeed, we could confirm that lymph node involvement and distant metastasis are associated with low IL-22BP expression. Importantly, the findings presented here are limited to patients with CRC and liver metastasis and cannot me generalized to other populations (such as CRC patients with distant metastasis affecting other organs). Finally, it is worth noting that despite the age and gender being equally distributed in both UICC Stage I-II and UICC Stage III-IV patients in our CRC cohort (Table 1), other cofounding factors not analyzed in the current study (such as comorbidities) could also potentially affect the expression levels of cytokines.
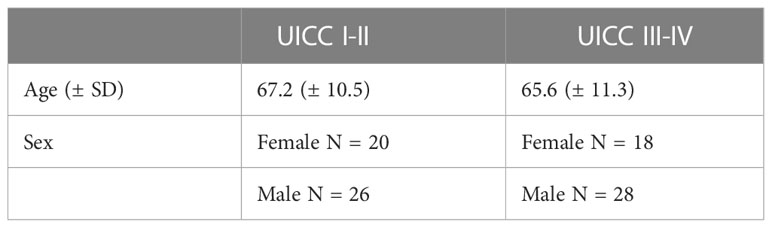
Table 1 Characteristics of Patients with UICC Stage I-II and UICC Stage III-IV CRC.
In contrast to the primary tumor, we found that the expression of IL-22BP is upregulated in liver metastasis compared to healthy liver tissue. Using IL-22BP-/- mice we could show that IL-22BP protects mice from liver metastasis, while it has no effect on the development of lung metastasis. Interestingly, the expression levels of IL-22BP are higher in lung metastasis compared to liver metastasis. Overall, those results suggest that the organ-specific microenvironment might influence the significance and function of IL-22BP in controlling the metastatic cascade.
Metastasis formation is a multi-step process. In the last and crucial steps, circulating tumor cells derived from the primary tumor need to extravasate into the target organ and establish a metastatic niche that promotes the progression of early metastasis. We have recently shown that IL-22 plays a crucial role in mediating the extravasation of cancer cells into the liver (25). IL-22BP does not seem to have such a role, since we here demonstrate that it does not affect cancer cell extravasation into the liver. This may be explained by the fact that the IL-22BP levels in the liver are very low under physiological conditions. However, IL-22BP is strongly upregulated in established liver metastasis. In line with this data, a genetic knock-out of IL-22BP resulted in faster growth of liver metastasis by increasing the proliferation of cancer cells. Thus, we conclude that IL-22BP is a crucial anti-tumorigenic factor that slows down the progression of liver metastasis. Interestingly, these effects are liver-specific since the knock-out of IL-22BP did not influence the formation of lung metastasis.
Dendritic cells, eosinophils and CD4+ T cells have been found to produce IL-22BP. However, the primary source of IL-22BP can differ depending on the disease-specific micro-milieu. Here, we show that CD11b+Ly6G+, CD11c+ and CD11b+Ly6C+ cells are the major source of IL-22BP in liver metastasis. However, of those cells only CD11c+ cells appear to upregulate IL-22BP in the setting of liver metastasis. Thus, dendritic cells are not only the major cellular source in the primary tumor in CRC, but they also represent one of the major cellular sources in CRC liver metastasis. Further studies are needed to decipher the mechanism underlying the upregulation of IL-22BP during the formation of liver metastasis. Moreover, the fact that IL-22BP was measured only on mRNA level in our animal experiments represents a limitation of our study. Currently, an antibody-based staining procedure of murine IL-22BP is complex and has not been successfully reported in a consistent way. Overcoming that limitation in future studies could provide further evidence regarding the differential levels of IL-22BP in tumor infiltrating lymphocytes compared to immune cells in the peritumoral tissue. Moreover, the spatial distribution of those cells in the tumor microenvironment could potentially offer explanations for the fact that IL-22BP controls the development of liver metastasis but not lung metastasis.
In summary, our data indicate a crucial role of IL-22BP in the progression of liver metastasis. Since IL-22BP is an endogenous antagonist of IL-22 signaling, our data indicate that pharmacological targeting of IL-22 or via boosting of IL-22BP might represent promising treatment options for CRC patients with liver metastasis. However, further studies are needed to assess how current pharmacological therapies are affecting the IL-22 - IL-22BP axis. Indeed, immunotherapies increasingly effective in various malignancies and are also used now in CRC. Similarly, other chemotherapies such as capecitabine can also affect the immunological response (30). Thus, further research is needed to understand possible synergistic effects of available therapeutic strategies and a pharmacological IL-22 blockade.
IL-22BP-/- mice have been described elsewhere (11). Age- and sex-matched knock-out mice and co-housed in-house bred C57/BL6 WT mice 8 to 18 weeks of age were used for all experiments. All animals were cared for in accordance with the institutional review board ‘Behörde für Soziales, Familie, Gesundheit und Verbraucherschutz’ Hamburg, Germany.
Human tissues were obtained freshly after surgical removal of tumors from patients diagnosed with CRC. Both the healthy colonic mucosa and parts of the tumor were collected and frozen until the RNA isolation procedure. In this study, we analyzed the expression levels of cytokines in 96 patients with CRC who were operated between 2010 and 2015 and who did not receive oncological therapy prior to surgical resection. The pathological tumor staging was available for 92 of those patients, the remaining four patients have thus been excluded from the analysis of clinical data. All human studies were approved by the local ethical committee (Ethik-Kommission der Ärztekammer Hamburg).
Human samples obtained from patients with suspicion of Non-alcoholic fatty liver disease, who showed normal histological findings and liver metastasis from CRC patients.
In brief, human liver or liver metastasis slides of tissue were fixed for 10 min in 4% PFA at RT. Tissues were washed with PBS and incubated in PBS-Triton 0.3% for 5 min. After washing they were incubated for 60 min in blocking buffer. Samples were stained overnight with α-IL-22BP antibody (primary mouse anti-mouse antibody, 1:350, antibody from R&D Systems, MAB 10871; Minneapolis, MN). After washing primary antibody (Alexa Fluor 488 rabbit anti-mouse IgG, Invitrogen) staining was performed (1 hour, RT) followed by 5 min staining with Hoechst 33258 (1:5000). For isotype control, the primary antibody was omitted.
For induction of forced liver metastasis, mice received 250 μl PBS containing 3x105 MC38 cells i.s. The injection was performed in the hemi-spleen which was removed 3 min after cancer cell injection. The mice were sacrificed after 3 weeks. Liver macroscopic metastases were counted by using a stereoscope (Olympus Corporation, Germany) (23, 24).
For induction of solid tumors and subsequent spontaneous lung metastasis formation, mice were anesthetized using isoflurane inhalation and received a 100 μl PBS containing 5x105 LLC cells subcutaneously (s.c.) (24). The tumors were resected after 2 weeks. The mice were sacrificed 4 weeks after tumor resection and lung macroscopic metastases were counted by using a stereoscope.
The caecum of anesthetized mice was exteriorized through an abdominal laparotomy. 1×106 LLC cells were injected into the caecal wall between the mucosa and the muscularis externa layers using a 30-gauge needle. A proper implantation into the caecum was confirmed at day 0 by a localized bubble in the cecal wall. The mice were sacrificed after 4 weeks and liver macroscopic metastases were counted by using a stereoscope (31).
Liver specimens were fixed in 4% buffered formalin, and embedded in paraffin or OCT (Sakura, Tokyo, Japan) and stored at -80°C. Tissue sections (4 μm) were prepared and stained with H&E. Metastatic lesion areas were quantified by Image J (ImageJ, U.S. National Institutes of Health, Bethesda, MD).
Paraffin liver sections from Il22bp+/+ and Il22bp-/- mice were stained with hematoxylin and eosin. Images were obtained at a 20x magnification and analysis was conducted using NIH Image J software. For estimation of metastatic site surface (as percentage of liver surface), a point counting grid (7 horizontal and 12 vertical lines/84 points) was superimposed on the images of liver parenchyma fields. The volume fraction of the metastatic foci was then equal to Pi/Pt, where Pi indicates the number of points on the structure of interest, namely on metastatic sites, and Pt is the total number of points on the reference field, which in this case is the liver parenchyma. Therefore, the volume fraction of metastatic sites was expressed as a percentage of the whole liver parenchyma.
Fc-γ receptors were blocked using a mAb (clone 2.4G2). The cells were stained with fluorochrome-conjugated antibodies as described elsewhere (25). BD LSRFortessa and FACSAria (BD Biosciences, San Jose, CA) were used for cell analysis and cell sorting, respectively. Data were analyzed using FlowJo v.6.1 (TreeStar, Ashland, OR).
Total RNA was extracted from colon tissue and cells from colon, lymph nodes, liver and spleen using TRIzol® Reagent (Invitrogen). The High capacity cDNA synthesis Kit (Applied Biosystems) was used for cDNA synthesis. Primers and probes were purchased from Applied Biosystems. For mouse Il22bp (Il22ra2) expression the Mm01192969_m1 primers (Applied Biosystems) were used; for human IL-22BP (IL22RA2) expression the Hs00364814_m1 primers (Applied Biosystems) were used. Real-time PCR was performed using the Kapa Probe Fast qPCR Master Mix (Kapa Biosystems) on the StepOne Plus system (Applied Biosystems). For both human and mouse, relative expression was normalized to HPRT and calculated using the 2-ΔΔCt method.
For the in vivo extravasation assay, mice received 250 μL PBS containing 3x105 MC38 GFP-labelled cells i.s. The mice were sacrificed after 24h and their livers were weighed. To isolate the MC38 GFP-labelled cells, the murine livers were cut in small pieces and minced using a scalpel. The tissues were incubated for 30 min at 37°C on a shaking incubator in HBSS (with Ca2+ and Mg2+) with Collagenase (1 mg/ml) and DNase I (10 U/ml) and supernatants were collected. Then, the supernatants were centrifuged for 4 min at 40g. We repeated this step twice more. Subsequently, the hepatocytes were removed and the cells were diluted in PBS 1x. One fifth of the cells were mixed with beads in 1:10 dilution and were analyzed by flow cytometry. In every extravasation assay, we used a mouse injected with not labelled cells in order to set up the gate of GFP+ cells.
Sample size was calculated using G*power (http://www.gpower.hhu.de/) assuming α = 0.05, β = 0.8, and ρ = 0.3. Statistical analysis was performed with GraphPad Prism® Software (GraphPad Software, San Diego, CA, USA). For comparison of groups, the non-parametric two- sided Mann–Whitney test or Fisher’s exact test was used. The significance level a was set to 0.05.
The original contributions presented in the study are included in the article/supplementary material. Further inquiries can be directed to the corresponding authors.
The studies involving human participants were reviewed and approved by Ärztekammer Hamburg. The patients/participants provided their written informed consent to participate in this study. The animal study was reviewed and approved by Behörde für Justiz und Verbraucherschutz, Lebensmittelsicherheit und Veterinärwesen (Hamburg, Germany).
AG and JK collaboratively conceived, designed and carried out most of the experiments, analyzed the data, provided critical intellectual input, and wrote the paper draft. TZ, JLu, AMS, DZ, IB, AM, PSe, TA, JT, AS, MTo, LB, TB, LK, BM, MF-A, AP, J-KG, RW, JB, BB, AH, TG, AD, MTa, and NM performed experiments. EG, PL, PA, PSt, MG, GG, RN, AT, DP, JLi, OM, JI, and NG provided critical intellectual input. IM and SH collaboratively conceived and designed most experiments, supervised the study, and provided critical intellectual input for the final paper draft. All authors contributed to the article and approved the submitted version.
This work was supported in part by the Deutsche Forschungsgemeinschaft (grant SFB841 to SH, NG, and grant SFB1328 to SH, NG), the European Research Council (CoG865466 to SH), Ernst Jung-Stiftung Hamburg (to SH), Stiftung Experimentelle Biomedizin (to SH), European Respiratory Society/short term fellowship (to AG), Else Kröner Memorial Stipendium (to AG), Werner Otto Stiftung (to AG), Erich und Gertrud Roggenbuck Stiftung (to AG), Hamburger Krebsgesellschaft Stiftung (to AG), Mildred Scheel Cancer Career Center Hamburg HaTriCS4 short term fellowship funded by Deutsche Krebshilfe (to JK, JT) and China Scholarship Council (to TZ). SH has an endowed Heisenberg - Professorship awarded by the Deutsche Forschungsgemeinschaft.
The Authors thank the In Vivo Optical Imaging Core Facility and the FACS Core Sorting Unit at the University Medical Center Hamburg-Eppendorf for their technical assistance.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
1. Bindea G, Mlecnik B, Tosolini M, Kirilovsky A, Waldner M, Obenauf AC, et al. Spatiotemporal dynamics of intratumoral immune cells reveal the immune landscape in human cancer. Immunity (2013) 39(4):782–95. doi: 10.1016/j.immuni.2013.10.003
2. Broussard EK, Disis ML. TNM staging in colorectal cancer: T is for T cell and m is for memory. J Clin Oncol (2011) 29(6):601–3. doi: 10.1200/JCO.2010.32.9078
3. Mlecnik B, Tosolini M, Kirilovsky A, Berger A, Bindea G, Meatchi T, et al. Histopathologic-based prognostic factors of colorectal cancers are associated with the state of the local immune reaction. J Clin Oncol (2011) 29(6):610–8. doi: 10.1200/JCO.2010.30.5425
4. Pagès F, Mlecnik B, Marliot F, Bindea G, Ou FS, Bifulco C, et al. International validation of the consensus immunoscore for the classification of colon cancer: a prognostic and accuracy study. Lancet (2018) 391(10135):2128–39. doi: 10.1016/S0140-6736(18)30789-X
5. Gronke K, Hernández PP, Zimmermann J, Klose CSN, Kofoed-Branzk M, Guendel F, et al. Interleukin-22 protects intestinal stem cells against genotoxic stress. Nature (2019) 566(7743):249–53. doi: 10.1038/s41586-019-0899-7
6. Kempski J, Brockmann L, Gagliani N, Huber S. TH17 cell and epithelial cell crosstalk during inflammatory bowel disease and carcinogenesis. Front Immunol (2017) 8:1373. doi: 10.3389/fimmu.2017.01373
7. Lindemans CA, Calafiore M, Mertelsmann AM, O’Connor MH, Dudakov JA, Jenq RR, et al. Interleukin-22 promotes intestinal-stem-cell-mediated epithelial regeneration. Nature (2015) 528(7583):560–4. doi: 10.1038/nature16460
8. Pickert G, Neufert C, Leppkes M, Zheng Y, Wittkopf N, Warntjen M, et al. STAT3 links IL-22 signaling in intestinal epithelial cells to mucosal wound healing. J Exp Med (2009) 206(7):1465–72. doi: 10.1084/jem.20082683
9. Sugimoto K, Ogawa A, Mizoguchi E, Shimomura Y, Andoh A, Bhan AK, et al. IL-22 ameliorates intestinal inflammation in a mouse model of ulcerative colitis. J Clin Invest (2008) 118(2):534–44. doi: 10.1172/JCI33194
10. Kryczek I, Lin Y, Nagarsheth N, Peng D, Zhao L, Zhao E, et al. IL-22+CD4+ T cells promote colorectal cancer stemness via STAT3 transcription factor activation and induction of the methyltransferase DOT1L. Immunity (2014) 40(5):772–84. doi: 10.1016/j.immuni.2014.03.010
11. Huber S, Gagliani N, Zenewicz LA, Huber FJ, Bosurgi L, Hu B, et al. IL-22BP is regulated by the inflammasome and modulates tumorigenesis in the intestine. Nature (2012) 491(7423):259–63. doi: 10.1038/nature11535
12. Perez LG, Kempski J, McGee HM, Pelzcar P, Agalioti T, Giannou A, et al. TGF-β signaling in Th17 cells promotes IL-22 production and colitis-associated colon cancer. Nat Commun (2020) 11(1):2608. doi: 10.1038/s41467-020-16363-w
13. Kirchberger S, Royston DJ, Boulard O, Thornton E, Franchini F, Szabady RL, et al. Innate lymphoid cells sustain colon cancer through production of interleukin-22 in a mouse model. J Exp Med (2013) 210(5):917–31. doi: 10.1084/jem.20122308
14. Brand S, Beigel F, Olszak T, Zitzmann K, Eichhorst ST, Otte JM, et al. IL-22 is increased in active crohn’s disease and promotes proinflammatory gene expression and intestinal epithelial cell migration. Am J Physiol-Gastrointest Liver Physiol (2006) 290(4):G827–38. doi: 10.1152/ajpgi.00513.2005
15. Kotenko SV, Izotova LS, Mirochnitchenko OV, Esterova E, Dickensheets H, Donnelly RP, et al. Identification, cloning, and characterization of a novel soluble receptor that binds IL-22 and neutralizes its activity. J Immunol (2001) 166(12):7096–103. doi: 10.4049/jimmunol.166.12.7096
16. Xu W, Presnell SR, Parrish-Novak J, Kindsvogel W, Jaspers S, Chen Z, et al. A soluble class II cytokine receptor, IL-22RA2, is a naturally occurring IL-22 antagonist. Proc Natl Acad Sci USA (2001) 98(17):9511–6. doi: 10.1073/pnas.171303198
17. Martin JC, Bériou G, Heslan M, Bossard C, Jarry A, Abidi A, et al. IL-22BP is produced by eosinophils in human gut and blocks IL-22 protective actions during colitis. Mucosal Immunol (2015) 9(2):539–49. doi: 10.1038/mi.2015.83
18. Pelczar P, Witkowski M, Perez LG, Kempski J, Hammel AG, Brockmann L, et al. A pathogenic role for T cell–derived IL-22BP in inflammatory bowel disease. Sci (1979) (2016) 354(6310):358–62. doi: 10.1126/science.aah5903
19. Martin JC, Bériou G, Heslan M, Chauvin C, Utriainen L, Aumeunier A, et al. Interleukin-22 binding protein (IL-22BP) is constitutively expressed by a subset of conventional dendritic cells and is strongly induced by retinoic acid. Mucosal Immunol (2014) 7(1):101–13. doi: 10.1038/mi.2013.28
20. Kempski J, Giannou AD, Riecken K, Zhao L, Steglich B, Lücke J, et al. IL-22BP mediates the antitumor effects of lymphotoxin against colorectal tumors in mice and humans. Gastroenterology (2020) 159(4):1417–30.e3. doi: 10.1053/j.gastro.2020.06.033
21. Guendel F, Kofoed-Branzk M, Gronke K, Tizian C, Witkowski M, Cheng HW, et al. Group 3 innate lymphoid cells program a distinct subset of IL-22BP-Producing dendritic cells demarcating solitary intestinal lymphoid tissues. Immunity (2020) 53(5):1015–1032.e8. doi: 10.1016/j.immuni.2020.10.012
22. Cancer Genome Atlas Research Network, Weinstein JN, Collisson EA, Mills GB, Shaw KRM, Ozenberger BA, et al. The cancer genome atlas pan-cancer analysis project. Nat Genet (2013) 45(10):1113–20. doi: 10.1038/ng.2764
23. Soares KC, Foley K, Olino K, Leubner A, Mayo SC, Jain A, et al. A preclinical murine model of hepatic metastases. J Visualized Experiments (2014) 91:51677. doi: 10.3791/51677
24. Giannou AD, Marazioti A, Kanellakis NI, Giopanou I, Lilis I, Zazara DE, et al. NRAS destines tumor cells to the lungs. EMBO Mol Med (2017) 9(5):672–86. doi: 10.15252/emmm.201606978
25. Giannou AD, Kempski J, Shiri AM, Lücke J, Zhang T, Zhao L, et al. Tissue resident iNKT17 cells facilitate cancer cell extravasation in liver metastasis via interleukin-22. Immunity (2023) 56(1):125–142.e12. doi: 10.1016/j.immuni.2022.12.014
26. Pagès F, Berger A, Camus M, Sanchez-Cabo F, Costes A, Molidor R, et al. Effector memory T cells, early metastasis, and survival in colorectal cancer. N Engl J Med (2005) 353(25):2654–66. doi: 10.1056/NEJMoa051424
27. Mlecnik B, Bindea G, Pagès F, Galon J. Tumor immunosurveillance in human cancers. Cancer Metastasis Rev (2011) 30(1):5–12. doi: 10.1007/s10555-011-9270-7
28. Fridman WH, Pagès F, Sautès-Fridman C, Galon J. The immune contexture in human tumours: impact on clinical outcome. Nat Rev Cancer (2012) 12(4):298–306. doi: 10.1038/nrc3245
29. Wu T, Cui L, Liang Z, Liu C, Liu Y, Li J. Elevated serum IL-22 levels correlate with chemoresistant condition of colorectal cancer. Clin Immunol (2013) 147(1):38–9. doi: 10.1016/j.clim.2013.02.007
30. Derakhshani A, Hashemzadeh S, Asadzadeh Z, Shadbad MA, Rasibonab F, Safarpour H, et al. Cytotoxic T-lymphocyte antigen-4 in colorectal cancer: another therapeutic side of capecitabine. Cancers (Basel) (2021) 13(10):2414. doi: 10.3390/cancers13102414
Keywords: metastasis, tumor immunology, IL-22BP, colorectal cancer, cancer therapy
Citation: Giannou AD, Kempski J, Zhang T, Lücke J, Shiri AM, Zazara DE, Belios I, Machicote A, Seeger P, Agalioti T, Tintelnot J, Sagebiel A, Tomczak M, Bauditz L, Bedke T, Kocheise L, Mercanoglu B, Fard-Aghaie M, Giorgakis E, Lykoudis PM, Pikouli A, Grass J-K, Wahib R, Bardenhagen J, Brunswig B, Heumann A, Ghadban T, Duprée A, Tachezy M, Melling N, Arck PC, Stringa P, Gentilini MV, Gondolesi GE, Nakano R, Thomson AW, Perez D, Li J, Mann O, Izbicki JR, Gagliani N, Maroulis IC and Huber S (2023) IL-22BP controls the progression of liver metastasis in colorectal cancer. Front. Oncol. 13:1170502. doi: 10.3389/fonc.2023.1170502
Received: 21 February 2023; Accepted: 16 May 2023;
Published: 31 May 2023.
Edited by:
Antonella Argentiero, National Cancer Institute Foundation (IRCCS), ItalyReviewed by:
Imke Atreya, University Hospital Erlangen, GermanyCopyright © 2023 Giannou, Kempski, Zhang, Lücke, Shiri, Zazara, Belios, Machicote, Seeger, Agalioti, Tintelnot, Sagebiel, Tomczak, Bauditz, Bedke, Kocheise, Mercanoglu, Fard-Aghaie, Giorgakis, Lykoudis, Pikouli, Grass, Wahib, Bardenhagen, Brunswig, Heumann, Ghadban, Duprée, Tachezy, Melling, Arck, Stringa, Gentilini, Gondolesi, Nakano, Thomson, Perez, Li, Mann, Izbicki, Gagliani, Maroulis and Huber. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Anastasios D. Giannou, YS5naWFubm91QHVrZS5kZQ==; Samuel Huber, cy5odWJlckB1a2UuZGU=
†These authors have contributed equally to this work and share first authorship
‡These authors jointly supervised this work
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.