 Yihan Yu
Yihan Yu Kanglu Li1†
Kanglu Li1† Yizhong Peng
Yizhong Peng Wei Wu
Wei Wu Fengxia Chen
Fengxia Chen Zhicai Zhang
Zhicai Zhang
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Oncol., 05 April 2023
Sec. Surgical Oncology
Volume 13 - 2023 | https://doi.org/10.3389/fonc.2023.1165380
This article is part of the Research TopicDiagnosis and Treatment of Bone MetastasesView all 17 articles
Cancer metastasis is a major cause of mortality from several tumors, including those of the breast, prostate, and the thyroid gland. Since bone tissue is one of the most common sites of metastasis, the treatment of bone metastases is crucial for the cure of cancer. Hence, disease models must be developed to understand the process of bone metastasis in order to devise therapies for it. Several translational models of different bone metastatic tumors have been developed, including animal models, cell line injection models, bone implant models, and patient-derived xenograft models. However, a compendium on different bone metastatic cancers is currently not available. Here, we have compiled several animal models derived from current experiments on bone metastasis, mostly involving breast and prostate cancer, to improve the development of preclinical models and promote the treatment of bone metastasis.
Metastasis is a frequent malignant manifestation of cancer in the mid to late stages of tumor progression. Metastasis to the bone, one of the most common sites, occurs when cancer cells migrate from the original site and invade bone tissue. It indicates adverse prognosis, and can cause severe pain, fractures, impaired mobility, and death. The invasion of cancer cells into target sites involves several stages. Initially, they invade the surroundings of the original site, breaching the vasculature and entering the circulation. Then, depending on molecular signals on cell membranes or in their microenvironment, they invade a particular target organ along their path of circulation (1, 2). Although the precise process has not been elucidated yet, the invasion appears to last many months if not years (3). Once a bulk of invasive cancer cells agglomerate into a mass, metastasis begins. Cancer cells modify the surrounding tissues and vasculature to favor their growth. Cancer treatment often involves a combination of radiation, chemotherapy, and medications to reduce the pain and inflammation.
Breast cancer, one of the most prevalent malignant tumors, exhibits a 40% likelihood to eventually develop bone metastases (4, 5). Bone tissue is the most common target site of breast cancer. Bone metastasis reflects potential skeletal-related events and poor clinical results. To improve the current therapies for bone-metastasized breast cancer, animal models that mimic the human tumor microenvironment have been used in preclinical experiments (6). Prostate cancer is the second most frequently occurring cancer in men. It preferentially metastasizes to the bone, and presents a worse prognosis at the metastatic stage. Rarely lethal when restricted to its primary site, the 5-year-survival rate of prostate cancer decreases by 29.8% when it metastasizes to the bone, explaining its rank as the fifth leading cause of tumor-related mortality in males (7). Antimetastatic agents need to be urgently developed and the prognosis following bone metastasis must be improved.
Multiple animal models have been used in clinical research to explore the mechanisms and prognosis of tumor metastasis. Translational models have been used to study the advanced stages of tumor metastases, reveal potential protein targets, and develop metastasis-related treatments. However, fully reproducing human bone metastases in animal models is difficult. Nevertheless, by selecting different cell lines, animal strains, and tumor transplantation methods, animal models can be constructed to answer various questions.
In this review, we have discussed the animal models of bone metastasis most commonly used in preclinical experiments and their underlying mechanisms. No single model can represent all the genetic mechanisms of bone metastasis, which requires whole-body organisms. Here, we have compiled a selection of animal models to assist in future studies (Figure 1).
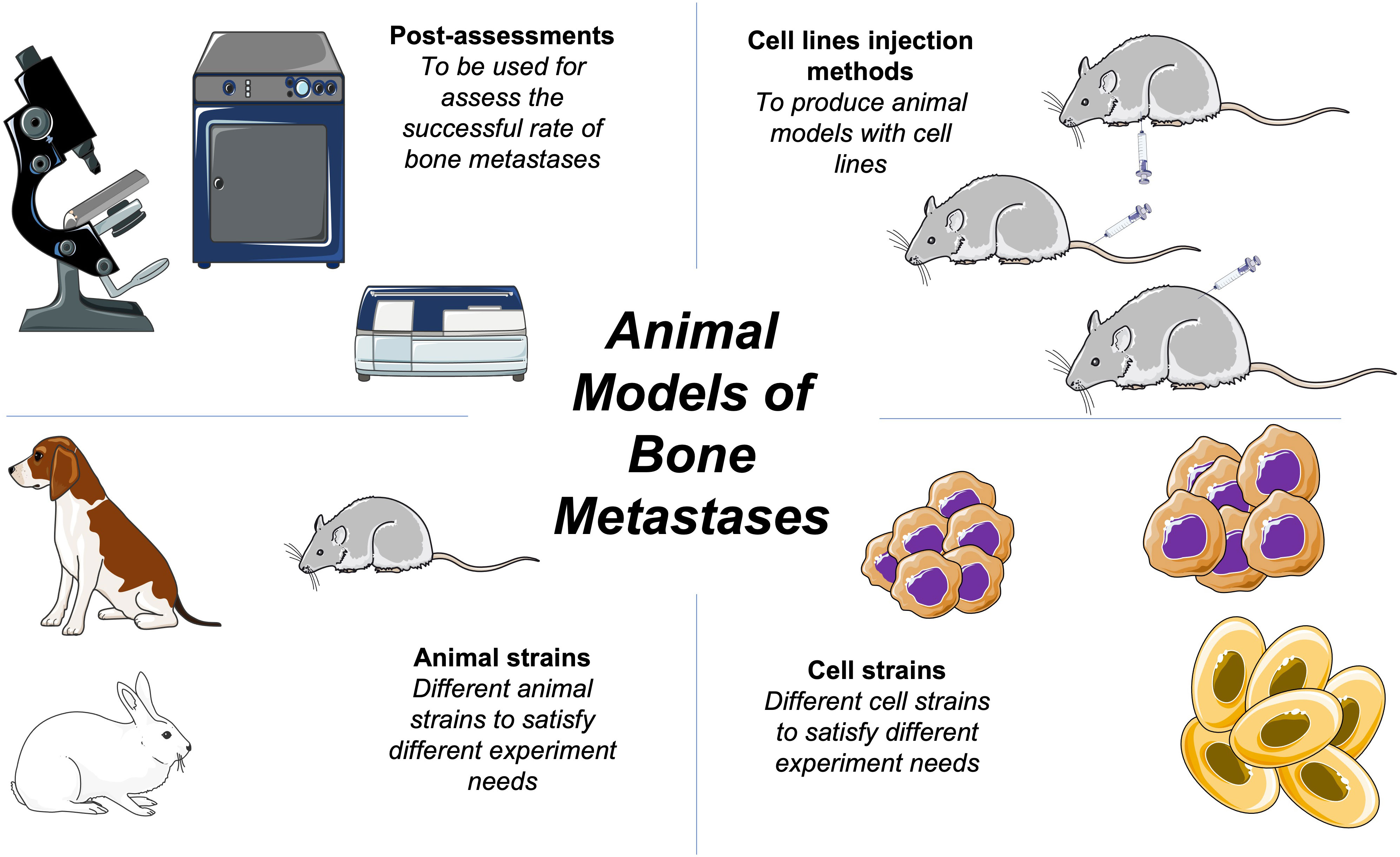
Figure 1 Schematic of basic bone metastases animal models methods.
Basing animal models of bone metastasis on general disease models is unreliable. Because the etiology of bone metastasis of human and animal cancers is different, different cancers have different metastatic targets. For example, mouse breast cancer may preferentially metastasize to the lung, while human breast cancer mainly metastasizes to the bone (2). Lung tumors may specifically metastasize to the vertebral column (8, 9). Hence, researchers are required to modify the animal models based on their experiments. The mouse is the most common animal of choice to construct bone metastasis models.
Animal models based on human breast cancer cells are commonly constructed using rodents, such as mice or rats, and used in preclinical experiments (10). Both immunodeficient and immunocompetent animals are used. Nude mice of the Balb/c background are frequently used because they are susceptible to both human and rodent breast cancer cell lines (2). Due to the lack of a thymus, immune responses are hardly generated in most of these mice following the injection of cancer cells, which significantly improves the success rate of model construction. Non-obese diabetic/severe combined immunodeficiency (NOD/SCID) mice are immunodeficient mice commonly used in xenograft experiments. Disabilities in the immune system of NOD/SCID mice affect the growth of lymph cells as well as immune signaling. Yin’s team used NOD/SCID mice paired with the MDA-MB-231 cell line to investigate how runt-related transcription factor 2, an osteogenesis-related factor, promotes breast cancer and bone metastasis (11).
The demand for crossbred or genetically engineered mice has also increased to better meet experimental needs (12–16). Mice that have been crossed and repeatedly backcrossed can offer an in vivo environment better suited to investigate the mechanism of breast cancer bone metastasis (13). In Laura’s experiment, Col1a-Krm2 mice were backcrossed with NOD/SCID/IL2rγnull (NSG) mice for 10 generations to introduce an immunocompromised background (13). They found that cancer metastasis to other organs like the spine may be prevented in rather young animals. By modifying the animal model into adult mice and backcrossing over 10 generations, they could focus on the early stages of human breast cancer metastasis. Devignes’ team also backcrossed Floxed mice bred in previous experiments with FVB/n wild-type mice for 10 generations to achieve genetic reconstitution consistent with their experimental requirements. Based on whether the HIF gene was expressed, mice were divided into two groups to verify whether the HIF signaling pathway in osteoblasts could promote breast cancer cell invasion and bone metastasis (14).
Unlike these experiments, Mercatali’s team used zebrafish as a special model to study bone metastasis (17). Visualizing zebrafish embryos and easy genetic manipulation provide researchers with a new method of studying cancer progression.
The first model of prostate cancer – the Dunning rat – exhibits a spontaneous development of the disease (7). However, this model did not show a tendency for bone metastasis, and R-3327 cells derived from the Dunning rat can only metastasize to the lymph nodes. Dogs are also listed as candidate animal models, but they rarely develop prostate cancer due to the lack of androgen receptors on their cell membranes (7). The internal organization of mice femur includes a high-woven bone structure that is less fibrolamellar in nature, providing conditions amenable for bone metastasis (10, 18).
Transgenic mouse models have the advantage of lacking immune responses to injected cells or xenografts (19). Transgenic adenocarcinoma of the mouse prostate (TRAMP) is one of the most famous transgenic models, exhibiting metastases to the lung and lymph nodes rather than the bone (19, 20). The promoters expressed in neuroendocrine cells, such as the probasin promoter in TRAMP, drive transgenic oncogene expression. NOD/SCID mouse is one of the most used immunodeficient animal models in prostate cancer bone metastasis experiments (21–25). Landgraf created a new model for studying prostate cancer bone metastasis by modifying NSG mice with a humanized tissue-engineered bone construct (hTEBC), which facilitates cancer cell growth (23). Ganguly’s team injected PC3 cells into the tibia of 6-week-old NSG mice to explore whether NOTCH3 induces tumor-specific elevation and secretion via MMP-3 (21).
However, the existing models are still limited to some of the detectable cancer-related factors, and cannot provide a comprehensive or linear picture of bone metastasis.
Both patient-derived cancer tissues and immortalized cancer cell lines are used for transplantation. Patient-derived cancer tissues show genetic concordance between the clinic and the animal models, and help to establish consistent animal models specific to particular cancer cell lines. However, these models may face obstacles in the form of ethics and tissue availability. Cell lines, after several passages, can generate stable primary or secondary cancer sites. Moreover, researchers can genetically edit cell lines by using luciferase genes or knocking out certain genes (26–28).
Immortalized human breast cancer cell lines, such as MDA-MB-231, 4T1, and MCF-7, are more easily available than patient-derived tissues. They possess obvious breast cancer target characteristics, and can also exhibit a tendency for bone metastasis after multiple passages (Table 1) (2, 5, 11, 51). They can help restore human bone metastasis in animal models. The bone-homing capabilities of MDA-MB-231 sub-lines can be enhanced via generation injections, and up to 90% of MDA-MB-231-bone cells can form neoplasms (52–54). Using 5–8-week-old mice is vital to achieve bone metastasis via intracardiac, intra-arterial, or intravenous injections. Farhoodi injected 4T1 cells into the mammary fat pad of Balb/c mice, and then examined their legs for bone metastases. Once its incidence was confirmed, the mice were sacrificed to collect the metastatic tumor cells from the leg bones. These cells were cultivated to purify tumor cells with bone-metastatic tendencies (51). They purified their experimental cells to improve the success rate.
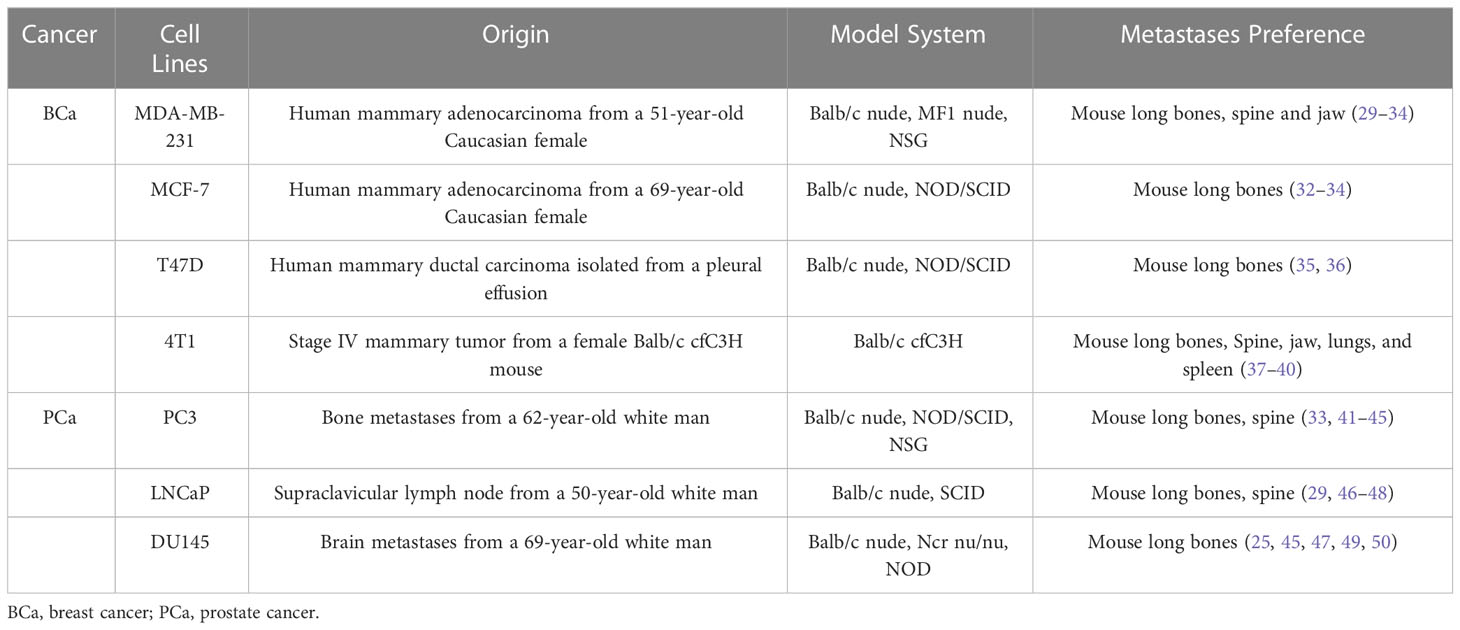
Table 1 Common cancer cell lines in bone metastases.
Different pairs of cell lines can also be combined to test certain concepts. Yin’s team compared MCF-7 and HCC1954 to validate whether KRT13, a protein from the keratin family, promotes stemness, metastasis, and cellular invasiveness (55). Han’s group estimated the metastatic rate of different cell lines (56). They found that the proliferation of MDA-MB-453, UACC-893, and HCC-202 cells increased in the eighth week, while MDA-MB-361, UACC-812, BT-474, and ZR-75-1 cells exhibited moderate proliferation but obvious migration. Using HCC-2218 and HCC1419 cells, tumors did not form, suggesting that both lack the ability to metastasize to the bone. The tumors formed by HCC-202 and MDA-MB-361 cells decreased in size after the sixth week, indicating that these two cell lines may not survive long-term metastasis (56). Eckhardt et al. also tested several cell lines, and NSG mice were used in xenograft studies involving MDA-MB-231 and SUM159 cells (37).
Like other cancer cell lines, those of prostate cancer also originate from both humans and animals (Table 1). R-3327, derived from the Dunning rat, has been used to investigate human prostate cancer due to its spontaneous neoplasm development (57). Other animal-derived cell lines, such as PA-III or AT6-1, naturally form osteolytic and osteoblastic lesions similar to human bone metastases in animal models (57–59). RM1, derived from the mouse prostate, is a highly metastatic cell line, but does not metastasize to the bone (60). Although it can induce consistent bone lesions in mouse models, it is a transformed cell line, not a natural one.
PC3, DU145, and LNCaP are patient-derived cell lines commonly used in prostate cancer animal models. They are easily available and possess the basic prostate cancer cell targets. PC3, derived from the bone metastases of a 62-year-old white man, was selected by isolating highly invasive cells from bone metastatic lesions. Landgraf implanted an hTEBC structure based on the bone-homing properties of PC3 cells, followed by an intracardiac injection of Luc-transfected cancer cells, facilitating the construction of models for transferring the human osteoblast line PC3 to hTEBC and the murine femur (23). Studies on LNCaP, PC3, and DU145 cells, all of which differ in their sensitivity to androgens, showed that prostate cancer-secreted growth differentiation factor 15 modulates the potential for bone remodeling in metastatic bone lesions (49, 61). Lang’s team grouped five common prostate cancer cell lines to verify whether PCAT7, a bone metastasis-related long non-coding RNA, activates the transforming growth factor-β/suppressor of mothers against decapentaplegic signaling pathway by upregulating transforming growth factor-β receptor 1. Its negative correlation with miR-324-5p was also investigated (62). Sohn’s team tried to intracardiacally inject LNCaP cell lines grouped with CD133+. The overexpression of CD133+ in LNCaP cells enhanced their cancer stem cell-like characteristics in terms of colony formation, migration, etc. The CD133+ group exhibited a bone metastasis rate of 80%, compared with 20% in the Vec group. Moreover, the CD133+ group showed a significant violation of the diffuse osteolytic characteristics of the spinal cord and the vertebral bodies (29).
In situ injection of cancer cells best reproduces the process of cancer metastasis in the human body. Injected into mouse mammary fat pads, tumor cells can be seeded through the vasculature towards the target organs – a method that achieves 40–60% of bone metastases in breast cancer animal models (63). To study the function of TIE2, a tyrosine kinase receptor, in osteolytic bone metastasis, Drescher’s team administered both bilateral mammary fat pad injections and left ventricular injections to the grouped mice. The correlation between carcinoma in situ and bone metastasis was evaluated to determine whether TIE2 inhibition stimulates the dormant breast cancer cells and promotes bone metastasis (34). Likewise, Spadazzi’s team injected MCF-7 cells into the left ventricle and mammary fat pads of NSG mice to investigate whether trefoil factor-1 could exert estrogen-induced effects (64).
However, this method suffers from a considerable variation in metastatic tumor growth, besides the comorbidity caused by development of the tumor (Table 2) (73). In addition, it poses the problem of small bone metastases while the primary tumor has grown beyond an ethically reasonable size (5), which seriously compromises the detection of stimulated bone metastases.
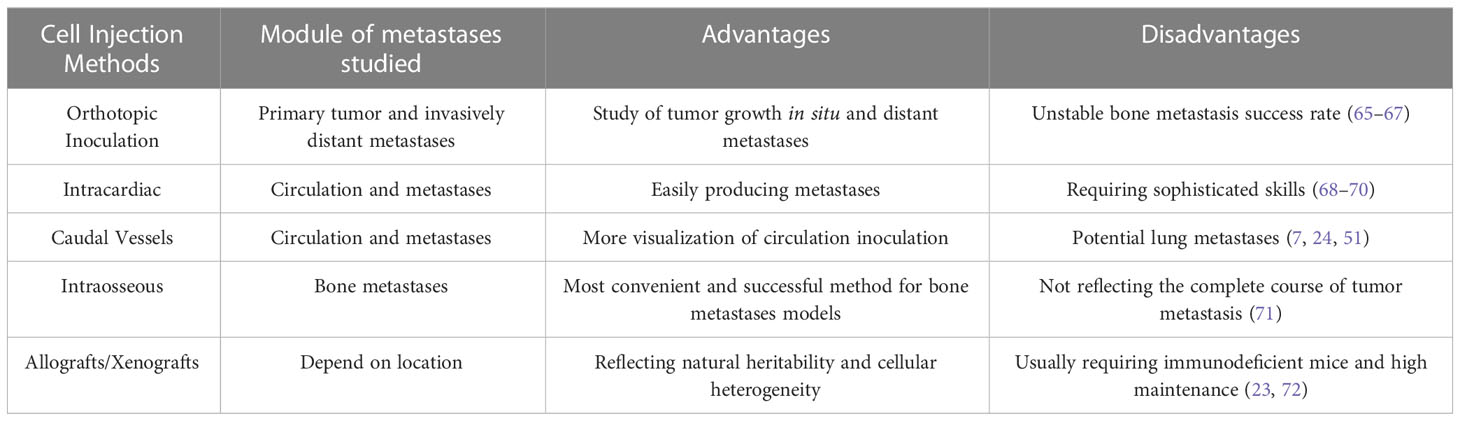
Table 2 Implantation methods for bone metastases models.
Some scientists have also suggested subcutaneous allografts to model bone metastasis. Peiffer’s team provided a detailed protocol of resecting subcutaneous prostate cancer allografts from immunocompetent mice (65). Bone metastases, abdominal cavity metastases, and local invasion all occurred in eight mice. This study demonstrated that resection of subcutaneous allografts from mice can lead to the development of metastasis; however, the duration of the experiment was extended by the removal of the prostate gland and precise operations.
Intravascular injection is a way of inoculating cells into the blood circulation. Unlike in orthotopic or ectopic inoculation, tumor cells injected via this method can localize to the target site through the intravascular circulation (Table 2) (66). Intra-arterial injections are usually administered to the left ventricle, limiting the clearance of cells that occurs when they pass through the lung capillaries (10, 53, 67). Tail vein injection, which is the more common intravenous injection today, effectively increases the rate of bone metastasis while also increasing the rate of mortality in mice (51).
Animal models currently rely on intracardiac injections to realize the process of bone metastasis. Tumor cells are injected into the circulation through the left ventricle of mice, after which they go through the processes of adhesion, degradation, and migration to finally cause metastases in different organs, thereby simulating the process of bloodway metastasis of tumors. Using intracardiac injections to probe the role of cancer-associated factors in the regulation of tumor bone metastasis has become the preferred modeling approach (44–46). Zheng et al. used this method to prove that osteoblastic Niche-derived Jagged1 sensitizes bone metastases (15). Wang’s team showed that the bone sialoprotein–αvβ3 integrin axis functioned significantly more efficiently in cancer cell bone metastasis when integrin was overexpressed. For comparison, stained specimens of the brain, lung, tibia, and femur were collected after left ventricular injection in nude mice (52). Although the postoperative mortality is relatively high, the survival rate can still exceed 90% with practice.
Caudal vessel injection can produce a higher rate of metastasis to the leg bone than to other vital organs. This method offers better accuracy than intracardiac injection because the visibility of tail vessels enables researchers to observe the flow of cancer cell fluids within (74). Caudal vascular injections can either be intravenous or arterial. Injecting through the tail artery will reduce the elimination of tumor cells in pulmonary capillaries and improve the success rate of colonization to the bone, while tail vein injection will promote tumor metastasis to the lung (2, 51, 74). In Farhoodi’s experiments, the 4T1 cell model tail artery injection mice showed a significant number of tumor cells localized to the subinguinal fat pad and the leg bone (51). Tumor cells were found in the leg bones of all 32 mice injected through the tail artery, and the rate of bone metastasis following complete tail veil injection was greater than 90% as well. Metastases were also detected in 70% of other target locations 2 weeks post-injection. Hamaidi et al. determined the effect of Lim1 on the adhesion, epithelial–mesenchymal transition, invasion, and metastatic progression of cancer cell surface targets after injection of the renal carcinoma cell line Caki2/786 through the lateral caudal vein of nude mice (75). However, caudal vein injection also resulted in metastatic foci in the lungs of mice.
Multiple factors affect the success of experiments involving vascular injection. Operator skill gaps, standard cell operation procedures, and pressure within the caudal vessels can all influence the growth rate and success of tumor bone metastasis (51). Dilation of the caudal vessels prior to injection or the use of fluorescein to reveal vessel flow can improve the effectiveness of the injection. Non-directed intracardiac injection is still associated with a risk of thrombosis due to the procoagulant activity of tumor cells after accurate completion. The mortality of post-inoculation animal models may be reduced by injecting low-molecular weight heparin into the tail vein 10 minutes before inoculation (76).
Metastatic tumors can bypass the pre-metastatic process if they are directly ectopically implanted into the bone. The growth of tumor cells inside the bone depends on their interaction with bone cells and the bone microenvironment (Table 2) (77, 78). Therefore, while intraosseous injection can help examine local tumor behavior within the bone microenvironment, it cannot be used to study the early stages of bone metastasis (79). Researchers typically inject 50,000–100,000 cancer cells directly into the tibia or femurs of mice, avoiding the possible comorbidity of the animals’ primary tumor (80, 81). Chen et al. observed that Brachyury, one gene affects tail length in mice, was expressed at a low level in the highly metastatic MDA-MB-231 cell line while it was highly expressed in the poorly metastatic T47D cell line when breast cancer cells were injected into the top anterior condylar region of the right tibia of mice. Nude mice showed significant swelling at the injection site 4 weeks post-injection, and X-ray revealed tumor-induced osteolytic lesions (35). After injecting prostate cancer cells into the left tibia of Balb/c nude mice, Thulin’s team performed bone tumor development status assays using peripheral quantitative computed tomography (CT) and microCT to investigate the effect of signal transducer and activator of transcription 3 (STAT3) inhibitors on STAT3-regulated prostate cancer bone metastasis. The STAT3 inhibitor treatment resulted in an intact tibial bone microenvironment with no tumor formation or sclerotic response in mice, whereas the VCaP group showed sclerotic bone tumor response up to 85% (48).
Transplanting allogeneic or xenogeneic tissues into animal models is a common way of modeling bone metastasis (Table 2). Since animals with different genetic backgrounds respond to allogeneic tissues differently, selecting the appropriate tissue source is especially important. In the case of xenografts, patient-derived tumor tissues can better reflect the biological characteristics of tumor bone metastasis in humans (82). Patient-derived xenografts aim to directly transplant human tumor tissue into immunodeficient mice, which represents natural heritability and cellular heterogeneity in human cancer better than simple cell-transplantation models (83). Among animal models, xenografts can only be performed in immunocompromised or immunodeficient animals. Aoki et al. first grew tumor tissue from bone metastases by intraperitoneally injecting it into male thymus-free nu/nu nude mice (42). The tumors were surgically processed to 1-mm3 fragments to be implanted into the proximal left tibia of the nude mice when they reached 10 mm in diameter. They observed tumor growth in all eight mice. Landgraf’s hTEBC model is likewise based on the low immune response of NSG mice to xenografts, while adding humanized components to mimic human tumor bone metastasis as satisfyingly as possible in mice (23).
After injecting cancer cells into mice, bone lesions develop quickly, necessitating researchers to detect physiological conditions, bone changes, and tumor lesions in a timely manner.
Establishing bone metastasis models using luciferase or fluorescent protein-labeled cell lines allows researchers to monitor tumor development in the bones of living animals (15, 39–41). Oliemuller et al. studied the effects of SOX11 on cell invasion and bone metastasis using DCIS-Luc cells, generated by transducing the cells with luciferase 2 lentiviral particles (84). Arriaga’s team bred NPKEYFP mice by crossing NPK mice with the Rosa-CAG-LSL-EYFP-WPRE reporter allele, facilitating in vivo fluorescence visualization and quantification of YFP-positive prostate tumors and metastases (85).
In turn, instrumentation such as the IVIS system can provide more accurate quantitative indicators through fluorescent or bioluminescent readings obtained from tumors (76–78). Typically, tumor growth in the bone is measured once or twice a week. The area of osteolytic lesions and abnormal bone remodeling can be assessed visually by X-ray or in vivo microCT (45–47, 85). Hinz’s team then used the IVIS system. After injecting MDA-MB-231 cells into the left ventricle of NSG mice, they performed IVIS bioluminescence assays weekly to assess osteolytic lesions caused by bone metastasis from triple-negative breast cancer. The inoculation of AKT3-knockout 231-BO cells into NSG mice resulted in enhanced bone metastases (86). Another team validated the effect of intracardiacally injecting MDA-MB-231-derived osteotropic cells into nude mice by examining osteolytic lesions in their hind tibia and femurs by microCT. MicroCT images showed that NKX2-8-silenced cell lines were more likely to produce earlier bone metastases, while its overexpression delayed the appearance of metastases, inhibited osteoclast activity, and reduced bone metastatic lesions (87).
At the end of the animal test, the mice should be examined simultaneously for extraosseous metastases. All relevant organs and metastases are fixed in 10% formalin for analysis. For histological studies, samples are fixed in paraformaldehyde for 24–48 hours and then decalcified in paraformaldehyde/ethylenediaminetetraacetic acid solution for 2 weeks. The decalcified paraffin-embedded bone should be sectioned for hematoxylin and eosin staining and evaluated using image analysis software. Bone conversion-related growth factors in the serum can also be assayed (88, 89). Metastases from the lung, liver, and brain tissue can likewise be analyzed and studies investigating the correlation between the area and the number of bone metastases can be performed (90).
Bone metastasis is a common manifestation of cancer deterioration in the mid and late stages of the disease. Much research has been done on the invasion of cancer cells, from migration to the bone tissue and beyond; however, much needs to be understood yet. Animal models are vital tools in preclinical metastatic experiments that can help identify the key steps in bone metastasis. Here, we have summarized the experimental animals, cell lines, cell implantation techniques, and evaluation methods used while studying common breast and prostate cancer bone metastases. For preclinical animal testing, immunodeficient animals are used to achieve xenograft growth without eliciting a host immune response. In preclinical studies, many investigators have successfully improved the success of tumor cell colonization to the bone by backcrossing cell lines and transgenic mice. More importantly, most animal tests related to cancer bone metastasis have been performed using cancer cell line injection models. Although the early stages of bone metastasis cannot be studied, these models are effective for studying the interaction between cancer cells and the bone microenvironment.
However, using mice to study human tumor immunity has its limitations. The differences in bone metastasis pathways between humans and animal models can explain why the success of preclinical treatments is not perfectly reproduced in humans. The inability to present a complete and comprehensive picture of the whole process of bone metastasis is also a problem that needs to be addressed while engineering animal models today.
All authors contributed equally to this work. All authors contributed to the article and approved the submitted version.
National Natural Science Foundation of China.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
1. Zhang L, Gaskins K, Yu Z, Xiong Y, Merino MJ, Kebebew E. An in vivo mouse model of metastatic human thyroid cancer. Thyroid Off J Am Thyroid Assoc (2014) 24(4):695–704. doi: 10.1089/thy.2013.0149
2. Tulotta C, Groenewoud A, Snaar-Jagalska BE, Ottewell P. Animal models of breast cancer bone metastasis. Methods Mol Biol Clifton NJ (2019) 1914:309–30. doi: 10.1007/978-1-4939-8997-3_17
3. Gawrzak S, Rinaldi L, Gregorio S, Arenas EJ, Salvador F, Urosevic J, et al. MSK1 regulates luminal cell differentiation and metastatic dormancy in ER+ breast cancer. Nat Cell Biol (2018) 20(2):211–21. doi: 10.1038/s41556-017-0021-z
4. Zhang L, Qu J, Qi Y, Duan Y, Huang YW, Zhou Z, et al. EZH2 engages TGFβ signaling to promote breast cancer bone metastasis via integrin β1-FAK activation. Nat Commun (2022) 13(1):2543. doi: 10.1038/s41467-022-30105-0
5. Horas K, Zheng Y, Zhou H, Seibel MJ. Animal models for breast cancer metastasis to bone: Opportunities and limitations. Cancer Invest. (2015) 33(9):459–68. doi: 10.3109/07357907.2015.1065500
6. Deasy SK, Erez N. A glitch in the matrix: Organ-specific matrisomes in metastatic niches. Trends Cell Biol (2022) 32(2):110–23. doi: 10.1016/j.tcb.2021.08.001
7. Berish RB, Ali AN, Telmer PG, Ronald JA, Leong HS. Translational models of prostate cancer bone metastasis. Nat Rev Urol. (2018) 15(7):403–21. doi: 10.1038/s41585-018-0020-2
8. Liang H, Zhou L, Hu Z, Ge Y, Zhang T, Chen Q, et al. Siglec15 checkpoint blockade for simultaneous immunochemotherapy and osteolysis inhibition in lung adenocarcinoma spinal metastasis via a hollow nanoplatform. Small Weinh Bergstr Ger (2022) 18(29):e2107787. doi: 10.1002/smll.202107787
9. Wang K, Jiang L, Hu A, Sun C, Zhou L, Huang Y, et al. Vertebral-specific activation of the CX3CL1/ICAM-1 signaling network mediates non-small-cell lung cancer spinal metastasis by engaging tumor cell-vertebral bone marrow endothelial cell interactions. Theranostics (2021) 11(10):4770–89. doi: 10.7150/thno.54235
10. Haq M, Goltzman D, Tremblay G, Brodt P. Rat prostate adenocarcinoma cells disseminate to bone and adhere preferentially to bone marrow-derived endothelial cells. Cancer Res (1992) 52(17):4613–9. doi: 10.1038/s41467-022-30105-0
11. Yin X, Teng X, Ma T, Yang T, Zhang J, Huo M, et al. RUNX2 recruits the NuRD(MTA1)/CRL4B complex to promote breast cancer progression and bone metastasis. Cell Death Differ (2022) 29(11):2203–17. doi: 10.1038/s41418-022-01010-2
12. Zuo H, Yang D, Wan Y. Fam20C regulates bone resorption and breast cancer bone metastasis through osteopontin and BMP4. Cancer Res (2021) 81(20):5242–54. doi: 10.1158/0008-5472.CAN-20-3328
13. Brylka L, Jähn-Rickert K, Baranowsky A, Neven M, Horn M, Yorgan T, et al. Spine metastases in immunocompromised mice after intracardiac injection of MDA-MB-231-SCP2 breast cancer cells. Cancers (2022) 14(3):556. doi: 10.3390/cancers14030556
14. Devignes CS, Aslan Y, Brenot A, Devillers A, Schepers K, Fabre S, et al. HIF signaling in osteoblast-lineage cells promotes systemic breast cancer growth and metastasis in mice. Proc Natl Acad Sci USA (2018) 115(5):E992–1001. doi: 10.1073/pnas.1718009115
15. Zheng H, Bae Y, Kasimir-Bauer S, Tang R, Chen J, Ren G, et al. Therapeutic antibody targeting tumor- and osteoblastic niche-derived Jagged1 sensitizes bone metastasis to chemotherapy. Cancer Cell (2017) 32(6):731–747.e6. doi: 10.1016/j.ccell.2017.11.002
16. Ouarné M, Bouvard C, Boneva G, Mallet C, Ribeiro J, Desroches-Castan A, et al. BMP9, but not BMP10, acts as a quiescence factor on tumor growth, vessel normalization and metastasis in a mouse model of breast cancer. J Exp Clin Cancer Res CR. (2018) 37:209. doi: 10.1186/s13046-018-0885-1
17. Mercatali L, La Manna F, Groenewoud A, Casadei R, Recine F, Miserocchi G, et al. Development of a patient-derived xenograft (PDX) of breast cancer bone metastasis in a zebrafish model. Int J Mol Sci (2016) 17(8):1375. doi: 10.3390/ijms17081375
18. Kerschnitzki M, Wagermaier W, Roschger P, Seto J, Shahar R, Duda GN, et al. The organization of the osteocyte network mirrors the extracellular matrix orientation in bone. J Struct Biol (2011) 173(2):303–11. doi: 10.1016/j.jsb.2010.11.014
19. Kang J, La Manna F, Bonollo F, Sampson N, Alberts IL, Mingels C, et al. Tumor microenvironment mechanisms and bone metastatic disease progression of prostate cancer. Cancer Lett (2022) 530:156–69. doi: 10.1016/j.canlet.2022.01.015
20. Gingrich JR, Barrios RJ, Morton RA, Boyce BF, DeMayo FJ, Finegold MJ, et al. Metastatic prostate cancer in a transgenic mouse. Cancer Res (1996) 56(18):4096–102.
21. Ganguly SS, Hostetter G, Tang L, Frank SB, Saboda K, Mehra R, et al. Notch3 promotes prostate cancer-induced bone lesion development via MMP-3. Oncogene (2020) 39(1):204–18. doi: 10.1038/s41388-019-0977-1
22. Polavaram NS, Dutta S, Islam R, Bag AK, Roy S, Poitz D, et al. Tumor- and osteoclast-derived NRP2 in prostate cancer bone metastases. Bone Res (2021) 9(1):24. doi: 10.1038/s41413-021-00136-2
23. Landgraf M, Lahr CA, Sanchez-Herrero A, Meinert C, Shokoohmand A, Pollock PM, et al. Humanized bone facilitates prostate cancer metastasis and recapitulates therapeutic effects of zoledronic acid in vivo. Bone Res (2019) 7:31. doi: 10.1038/s41413-019-0072-9
24. Hu CY, Chen J, Qin XH, You P, Ma J, Zhang J, et al. Long non-coding RNA NORAD promotes the prostate cancer cell extracellular vesicle release via microRNA-541-3p-regulated PKM2 to induce bone metastasis of prostate cancer. J Exp Clin Cancer Res CR. (2021) 40(1):98. doi: 10.1186/s13046-021-01891-0
25. Li L, Ameri AH, Wang S, Jansson KH, Casey OM, Yang Q, et al. EGR1 regulates angiogenic and osteoclastogenic factors in prostate cancer and promotes metastasis. Oncogene (2019) 38(35):6241–55. doi: 10.1038/s41388-019-0873-8
26. Luo A, Xu Y, Li S, Bao J, Lü J, Ding N, et al. Cancer stem cell property and gene signature in bone-metastatic breast cancer cells. Int J Biol Sci (2020) 16(14):2580–94. doi: 10.7150/ijbs.45693
27. Jenkins DE, Hornig YS, Oei Y, Dusich J, Purchio T. Bioluminescent human breast cancer cell lines that permit rapid and sensitive in vivo detection of mammary tumors and multiple metastases in immune deficient mice. Breast Cancer Res BCR (2005) 7(4):R444–454. doi: 10.1186/bcr1026
28. Sawada Y, Kikugawa T, Iio H, Sakakibara I, Yoshida S, Ikedo A, et al. GPRC5A facilitates cell proliferation through cell cycle regulation and correlates with bone metastasis in prostate cancer. Int J Cancer (2020) 146(5):1369–82. doi: 10.1002/ijc.32554
29. Sohn HM, Kim B, Park M, Ko YJ, Moon YH, Sun JM, et al. Effect of CD133 overexpression on bone metastasis in prostate cancer cell line LNCaP. Oncol Lett (2019) 18(2):1189–98. doi: 10.3892/ol.2019.10443
30. Sung NJ, Kim NH, Surh YJ, Park SA. Gremlin-1 promotes metastasis of breast cancer cells by activating STAT3-MMP13 signaling pathway. Int J Mol Sci (2020) 21(23):9227. doi: 10.3390/ijms21239227
31. Marino S, de Ridder D, Bishop RT, Renema N, Ponzetti M, Sophocleous A, et al. Paradoxical effects of JZL184, an inhibitor of monoacylglycerol lipase, on bone remodelling in healthy and cancer-bearing mice. EBioMedicine (2019) 44:452–66. doi: 10.1016/j.ebiom.2019.05.048
32. Zhang Z, Xu Q, Song C, Mi B, Zhang H, Kang H, et al. Serum- and glucocorticoid-inducible kinase 1 is essential for osteoclastogenesis and promotes breast cancer bone metastasis. Mol Cancer Ther (2020) 19(2):650–60. doi: 10.1158/1535-7163.MCT-18-0783
33. Kim B, Kim H, Jung S, Moon A, Noh DY, Lee ZH, et al. A CTGF-RUNX2-RANKL axis in breast and prostate cancer cells promotes tumor progression in bone. J Bone Miner Res (2020) 35(1):155–66. doi: 10.1002/jbmr.3869
34. Drescher F, Juárez P, Arellano DL, Serafín-Higuera N, Olvera-Rodriguez F, Jiménez S, et al. TIE2 induces breast cancer cell dormancy and inhibits the development of osteolytic bone metastases. Cancers (2020) 12(4):868. doi: 10.3390/cancers12040868
35. Chen M, Zou S, He C, Zhou J, Li S, Shen M, et al. Transactivation of SOX5 by brachyury promotes breast cancer bone metastasis. Carcinogenesis (2020) 41(5):551–60. doi: 10.1093/carcin/bgz142
36. Zhao C, Cai X, Wang Y, Wang D, Wang T, Gong H, et al. NAT1 promotes osteolytic metastasis in luminal breast cancer by regulating the bone metastatic niche via NF-κB/IL-1B signaling pathway. Am J Cancer Res (2020) 10(8):2464–79.
37. Eckhardt BL, Cao Y, Redfern AD, Chi LH, Burrows AD, Roslan S, et al. Activation of canonical BMP4-SMAD7 signaling suppresses breast cancer metastasis. Cancer Res (2020) 80(6):1304–15. doi: 10.1158/0008-5472.CAN-19-0743
38. Zhang D, Iwabuchi S, Baba T, Hashimoto S, Mukaida N, Sasaki S. Involvement of a transcription factor, Nfe2, in breast cancer metastasis to bone. Cancers (2020) 12(10):3003. doi: 10.3390/cancers12103003
39. Sasaki S, Zhang D, Iwabuchi S, Tanabe Y, Hashimoto S, Yamauchi A, et al. Crucial contribution of GPR56/ADGRG1, expressed by breast cancer cells, to bone metastasis formation. Cancer Sci (2021) 112(12):4883–93. doi: 10.1111/cas.15150
40. Sun J, Huang J, Lan J, Zhou K, Gao Y, Song Z, et al. Overexpression of CENPF correlates with poor prognosis and tumor bone metastasis in breast cancer. Cancer Cell Int (2019) 19:264. doi: 10.1186/s12935-019-0986-8
41. Connelly ZM, Jin R, Zhang J, Yang S, Cheng S, Shi M, et al. FOXA2 promotes prostate cancer growth in the bone. Am J Transl Res (2020) 12(9):5619–29. doi: 10.1038/s41413-021-00178-6
42. Aoki Y, Masaki N, Tome Y, Kubota Y, Aoki Y, Bouvet M, et al. Non-invasively imageable tibia-tumor-fragment implantation experimental-bone-metastasis mouse model of GFP-expressing prostate cancer. Vivo Athens Greece (2022) 36(4):1647–50. doi: 10.21873/invivo.12876
43. Shin SH, Kim I, Lee JE, Lee M, Park JW. Loss of EGR3 is an independent risk factor for metastatic progression in prostate cancer. Oncogene (2020) 39(36):5839–54. doi: 10.1038/s41388-020-01418-5
44. Huang S, Wa Q, Pan J, Peng X, Ren D, Li Q, et al. Transcriptional downregulation of miR-133b by REST promotes prostate cancer metastasis to bone via activating TGF-β signaling. Cell Death Dis (2018) 9(7):779. doi: 10.1038/s41419-018-0807-3
45. Meng X, Vander Ark A, Daft P, Woodford E, Wang J, Madaj Z, et al. Loss of TGF-β signaling in osteoblasts increases basic-FGF and promotes prostate cancer bone metastasis. Cancer Lett (2018) 418:109–18. doi: 10.1016/j.canlet.2018.01.018
46. Li Q, Wang M, Hu Y, Zhao E, Li J, Ren L, et al. MYBL2 disrupts the hippo-YAP pathway and confers castration resistance and metastatic potential in prostate cancer. Theranostics (2021) 11(12):5794–812. doi: 10.7150/thno.56604
47. Zhao Z, Li E, Luo L, Zhao S, Liu L, Wang J, et al. A PSCA/PGRN-NF-κB-Integrin-α4 axis promotes prostate cancer cell adhesion to bone marrow endothelium and enhances metastatic potential. Mol Cancer Res MCR (2020) 18(3):501–13. doi: 10.1158/1541-7786.MCR-19-0278
48. Thulin MH, Määttä J, Linder A, Sterbova S, Ohlsson C, Damber JE, et al. Inhibition of STAT3 prevents bone metastatic progression of prostate cancer in vivo. Prostate (2021) 81(8):452–62. doi: 10.1002/pros.24125
49. Siddiqui JA, Seshacharyulu P, Muniyan S, Pothuraju R, Khan P, Vengoji R, et al. GDF15 promotes prostate cancer bone metastasis and colonization through osteoblastic CCL2 and RANKL activation. Bone Res (2022) 10(1):6. doi: 10.1038/s41413-021-00178-6
50. Zhang B, Li Y, Wu Q, Xie L, Barwick B, Fu C, et al. Acetylation of KLF5 maintains EMT and tumorigenicity to cause chemoresistant bone metastasis in prostate cancer. Nat Commun (2021) 12(1):1714. doi: 10.1038/s41467-021-21976-w
51. Farhoodi HP, Segaliny AI, Wagoner ZW, Cheng JL, Liu L, Zhao W. Optimization of a syngeneic murine model of bone metastasis. J Bone Oncol (2020) 23:100298. doi: 10.1016/j.jbo.2020.100298
52. Wang L, Song L, Li J, Wang Y, Yang C, Kou X, et al. Bone sialoprotein-αvβ3 integrin axis promotes breast cancer metastasis to the bone. Cancer Sci (2019) 110(10):3157–72. doi: 10.1111/cas.14172
53. Wetterwald A, van der Pluijm G, Que I, Sijmons B, Buijs J, Karperien M, et al. Optical imaging of cancer metastasis to bone marrow: A mouse model of minimal residual disease. Am J Pathol (2002) 160(3):1143–53. doi: 10.1016/S0002-9440(10)64934-6
54. Nutter F, Holen I, Brown HK, Cross SS, Evans CA, Walker M, et al. Different molecular profiles are associated with breast cancer cell homing compared with colonisation of bone: Evidence using a novel bone-seeking cell line. Endocr Relat Cancer (2014) 21(2):327–41. doi: 10.1530/ERC-13-0158
55. Yin L, Li Q, Mrdenovic S, Chu GCY, Wu BJ, Bu H, et al. KRT13 promotes stemness and drives metastasis in breast cancer through a plakoglobin/c-myc signaling pathway. Breast Cancer Res BCR (2022) 24(1):7. doi: 10.1186/s13058-022-01502-6
56. Han Y, Azuma K, Watanabe S, Semba K, Nakayama J. Metastatic profiling of HER2-positive breast cancer cell lines in xenograft models. Clin Exp Metastasis (2022) 39(3):467–77. doi: 10.1007/s10585-022-10150-1
57. Liepe K, Geidel H, Haase M, Hakenberg OW, Runge R, Kotzerke J. New model for the induction of osteoblastic bone metastases in rat. Anticancer Res (2005) 25(2A):1067–73.
58. Blouin S, Baslé MF, Chappard D. Rat models of bone metastases. Clin Exp Metastasis (2005) 22(8):605–14. doi: 10.1007/s10585-006-9002-5
59. Lamoureux F, Ory B, Battaglia S, Pilet P, Heymann MF, Gouin F, et al. Relevance of a new rat model of osteoblastic metastases from prostate carcinoma for preclinical studies using zoledronic acid. Int J Cancer (2008) 122(4):751–60. doi: 10.1002/ijc.23187
60. Power CA, Pwint H, Chan J, Cho J, Yu Y, Walsh W, et al. A novel model of bone-metastatic prostate cancer in immunocompetent mice. Prostate (2009) 69(15):1613–23. doi: 10.1002/pros.21010
61. Wang W, Yang X, Dai J, Lu Y, Zhang J, Keller ET. Prostate cancer promotes a vicious cycle of bone metastasis progression through inducing osteocytes to secrete GDF15 that stimulates prostate cancer growth and invasion. Oncogene (2019) 38(23):4540–59. doi: 10.1038/s41388-019-0736-3
62. Lang C, Dai Y, Wu Z, Yang Q, He S, Zhang X, et al. SMAD3/SP1 complex-mediated constitutive active loop between lncRNA PCAT7 and TGF-β signaling promotes prostate cancer bone metastasis. Mol Oncol (2020) 14(4):808–28. doi: 10.1002/1878-0261.12634
63. Lee JH, Kim B, Jin WJ, Kim JW, Kim HH, Ha H, et al. Trolox inhibits osteolytic bone metastasis of breast cancer through both PGE2-dependent and independent mechanisms. Biochem Pharmacol (2014) 91(1):51–60. doi: 10.1016/j.bcp.2014.06.005
64. Spadazzi C, Mercatali L, Esposito M, Wei Y, Liverani C, De Vita A, et al. Trefoil factor-1 upregulation in estrogen-receptor positive breast cancer correlates with an increased risk of bone metastasis. Bone (2021) 144:115775. doi: 10.1016/j.bone.2020.115775
65. Peiffer LB, Hicks J, Sosa RY, De Marzo AM, Sfanos KS, Maynard JP. Modeling human prostate cancer metastasis in mice via resection of subcutaneous allografts. Front Oncol (2022) 12:877536. doi: 10.3389/fonc.2022.877536
66. Yip RKH, Rimes JS, Capaldo BD, Vaillant F, Mouchemore KA, Pal B, et al. Mammary tumour cells remodel the bone marrow vascular microenvironment to support metastasis. Nat Commun (2021) 12(1):6920. doi: 10.1038/s41467-021-26556-6
67. Chang J, Sun X, Ma X, Zhao P, Shi B, Wang Y, et al. Intra-cardiac injection of human prostate cancer cells to create a bone metastasis xenograft mouse model. J Vis Exp JoVE (2022) 189). doi: 10.3791/64589
68. Chen M, Wu C, Fu Z, Liu S. ICAM1 promotes bone metastasis via integrin-mediated TGF-β/EMT signaling in triple-negative breast cancer. Cancer Sci (2022) 113(11):3751–65. doi: 10.1111/cas.15532
69. Labanca E, Yang J, Shepherd PDA, Wan X, Starbuck MW, Guerra LD, et al. Fibroblast growth factor receptor 1 drives the metastatic progression of prostate cancer. Eur Urol Oncol (2022) 5(2):164–75. doi: 10.1016/j.euo.2021.10.001
70. Capietto AH, Lee S, Clever D, Eul E, Ellis H, Ma CX, et al. Effective treatment of established bone metastases can be achieved by combinatorial osteoclast blockade and depletion of granulocytic subsets. Cancer Immunol Res (2021) 9(12):1400–12. doi: 10.1158/2326-6066.CIR-21-0232
71. Engelmann J, Zarrer J, Gensch V, Riecken K, Berenbrok N, Luu TV, et al. Regulation of bone homeostasis by MERTK and TYRO3. Nat Commun (2022) 13(1):7689. doi: 10.1038/s41467-022-33938-x
72. Boudreau MW, Duraki D, Wang L, Mao C, Kim JE, Henn MA, et al. A small-molecule activator of the unfolded protein response eradicates human breast tumors in mice. Sci Transl Med (2021) 13(603):eabf1383. doi: 10.1126/scitranslmed.abf1383
73. Bibby MC. Orthotopic models of cancer for preclinical drug evaluation: Advantages and disadvantages. Eur J Cancer Oxf Engl 1990 (2004) 40(6):852–7. doi: 10.1186/s13046-018-0813-4
74. Kuchimaru T, Kataoka N, Nakagawa K, Isozaki T, Miyabara H, Minegishi M, et al. A reliable murine model of bone metastasis by injecting cancer cells through caudal arteries. Nat Commun (2018) 9:2981. doi: 10.1038/s41467-018-05366-3
75. Hamaidi I, Coquard C, Danilin S, Dormoy V, Béraud C, Rothhut S, et al. The Lim1 oncogene as a new therapeutic target for metastatic human renal cell carcinoma. Oncogene (2019) 38(1):60–72. doi: 10.1038/s41388-018-0413-y
76. Stocking KL, Jones JC, Everds NE, Buetow BS, Roudier MP, Miller RE. Use of low-molecular-weight heparin to decrease mortality in mice after intracardiac injection of tumor cells. Comp Med (2009) 59(1):37–45.
77. Robinson BD, Sica GL, Liu YF, Rohan TE, Gertler FB, Condeelis JS, et al. Tumor microenvironment of metastasis in human breast carcinoma: A potential prognostic marker linked to hematogenous dissemination. Clin Cancer Res Off J Am Assoc Cancer Res (2009) 15(7):2433–41. doi: 10.1158/1078-0432.CCR-08-2179
78. Coleman RE, Gregory W, Marshall H, Wilson C, Holen I. The metastatic microenvironment of breast cancer: Clinical implications. Breast Edinb Scotl (2013) 22 Suppl 2:S50–56. doi: 10.1016/j.breast.2013.07.010
79. Halpern J, Lynch CC, Fleming J, Hamming D, Martin MD, Schwartz HS, et al. The application of a murine bone bioreactor as a model of tumor: bone interaction. Clin Exp Metastasis (2006) 23(7–8):345–56. doi: 10.1007/s10585-006-9044-8
80. Zhu W, Sheng D, Shao Y, Zhang Q, Peng Y. Neuronal calcitonin gene-related peptide promotes prostate tumor growth in the bone microenvironment. Peptides (2021) 135:170423. doi: 10.1016/j.peptides.2020.170423
81. Zheng Y, Seibel MJ, Zhou H. Methods in bone biology: Cancer and bone. In: Duque G, Watanabe K, editors. Osteoporosis Res: Animal models. London: Springer London (2011). p. 83–91. doi: 10.1007/978-0-85729-293-3_7
82. Holzapfel BM, Thibaudeau L, Hesami P, Taubenberger A, Holzapfel NP, Mayer-Wagner S, et al. Humanised xenograft models of bone metastasis revisited: Novel insights into species-specific mechanisms of cancer cell osteotropism. Cancer Metastasis Rev (2013) 32(1–2):129–45. doi: 10.1007/s10555-013-9437-5
83. Lin D, Xue H, Wang Y, Wu R, Watahiki A, Dong X, et al. Next generation patient-derived prostate cancer xenograft models. Asian J Androl. (2014) 16(3):407–12. doi: 10.4103/1008-682X.125394
84. Oliemuller E, Newman R, Tsang SM, Foo S, Muirhead G, Noor F, et al. SOX11 promotes epithelial/mesenchymal hybrid state and alters tropism of invasive breast cancer cells. eLife (2020) 9. doi: 10.7554/eLife.58374
85. Arriaga JM, Panja S, Alshalalfa M, Zhao J, Zou M, Giacobbe A, et al. A MYC and RAS co-activation signature in localized prostate cancer drives bone metastasis and castration resistance. Nat Cancer (2020) 11):1082–96. doi: 10.1038/s43018-020-00125-0
86. Hinz N, Baranowsky A, Horn M, Kriegs M, Sibbertsen F, Smit DJ, et al. Knockdown of AKT3 activates HER2 and DDR kinases in bone-seeking breast cancer cells, promotes metastasis In vivo and attenuates the TGFβ/CTGF axis. Cells (2021) 10(2):430. doi: 10.3390/cells10020430
87. Abudourousuli A, Chen S, Hu Y, Qian W, Liao X, Xu Y, et al. NKX2-8/PTHrP axis-mediated osteoclastogenesis and bone metastasis in breast cancer. Front Oncol (2022) 12:907000. doi: 10.3389/fonc.2022.907000
88. Maimon A, Levi-Yahid V, Ben-Meir K, Halpern A, Talmi Z, Priya S, et al. Myeloid cell-derived PROS1 inhibits tumor metastasis by regulating inflammatory and immune responses via IL-10. J Clin Invest (2021) 131(10):e126089. doi: 10.1172/JCI126089
89. Cai WL, Huang WD, Li B, Chen TR, Li ZX, Zhao CL, et al. microRNA-124 inhibits bone metastasis of breast cancer by repressing interleukin-11. Mol Cancer (2018) 17(1):9. doi: 10.1186/s12943-017-0746-0
Keywords: bone metastases, animal models, breast cancer, prostate cancer, cell lines
Citation: Yu Y, Li K, Peng Y, Wu W, Chen F, Shao Z and Zhang Z (2023) Animal models of cancer metastasis to the bone. Front. Oncol. 13:1165380. doi: 10.3389/fonc.2023.1165380
Received: 14 February 2023; Accepted: 10 March 2023;
Published: 05 April 2023.
Edited by:
Wenwen Zhang, Nanjing Medical University, ChinaReviewed by:
Qiuyu Liu, Henan Provincial People’s Hospital, ChinaCopyright © 2023 Yu, Li, Peng, Wu, Chen, Shao and Zhang. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Fengxia Chen, ZmVuZ3hpYWNoZW5Ad2h1LmVkdS5jbg==; Zengwu Shao, MTk4NVhIMDUzNkBodXN0LmVkdS5jbg==; Zhicai Zhang, emhpY2FpemhhbmdAMTI2LmNvbQ==
†These authors have contributed equally to this work
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.