
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Oncol. , 27 April 2023
Sec. Breast Cancer
Volume 13 - 2023 | https://doi.org/10.3389/fonc.2023.1146008
This article is part of the Research Topic Overcoming Resistance to Systemic Therapy in Breast Cancer View all 13 articles
 Rafael Vázquez-Romo1†
Rafael Vázquez-Romo1† Oliver Millan-Catalan2†
Oliver Millan-Catalan2† Erika Ruíz-García3
Erika Ruíz-García3 Antonio D. Martínez-Gutiérrez2Alberto Alvarado-Miranda1
Antonio D. Martínez-Gutiérrez2Alberto Alvarado-Miranda1 Alma D. Campos-Parra4
Alma D. Campos-Parra4 César López-Camarillo5
César López-Camarillo5 Nadia Jacobo-Herrera6
Nadia Jacobo-Herrera6 Eduardo López-Urrutia7
Eduardo López-Urrutia7 Mariano Guardado-Estrada8
Mariano Guardado-Estrada8 David Cantú de León4*
David Cantú de León4* Carlos Pérez-Plasencia2,7*
Carlos Pérez-Plasencia2,7*Introduction: Metastatic breast cancer causes the most breast cancer-related deaths around the world, especially in countries where breast cancer is detected late into its development. Genetic testing for cancer susceptibility started with the BRCA 1 and 2 genes. Still, recent research has shown that variations in other members of the DNA damage response (DDR) are also associated with elevated cancer risk, opening new opportunities for enhanced genetic testing strategies.
Methods: We sequenced BRCA1/2 and twelve other DDR genes from a Mexican-mestizo population of 40 metastatic breast cancer patients through semiconductor sequencing.
Results: Overall, we found 22 variants –9 of them reported for the first time– and a strikingly high proportion of variations in ARID1A. The presence of at least one variant in the ARID1A, BRCA1, BRCA2, or FANCA genes was associated with worse progression-free survival and overall survival in our patient cohort.
Discussion: Our results reflected the unique characteristics of the Mexican-mestizo population as the proportion of variants we found differed from that of other global populations. Based on these findings, we suggest routine screening for variants in ARID1A along with BRCA1/2 in breast cancer patients from the Mexican-mestizo population.
As the number of new breast cancer cases and fatalities continues to rise worldwide (1), detection and treatment of this disease is more of a pressing issue for researchers and health professionals. Widely adopted screening programs have proven efficient at detecting stage I-II cases before they develop further into stage III and compromise survival, effectively decreasing the fatal cases. Yet, a significant amount of these deaths occur in stage IV or metastatic patients (2, 3).
An important aspect of the screening programs that has become increasingly widespread with the advancement of technology is genetic testing for cancer susceptibility. The first surveyed genes were BRCA 1 and 2, where germline variants are associated with around 25% of breast cancer cases (4). But recent research has revealed that several other genes that also participate in the DNA damage response (DDR) also confer increased breast cancer risk, such as PALB2, TP53, RAD50, RAD51D, and CHEK2, among others (5, 6). For example, variants in PALB2 (Partner and Localizer of BRCA2) are associated with an estimated cumulative risk of breast cancer of 14% (7). Somatic alterations, on the other hand, are associated with high-grade tumor progression; for instance, TP53 variants correlate with metastasis spread and relapse risk (8, 9). Both germline and somatic alterations drive tumor development cooperatively and influence response to treatment (10, 11).
Variants in the DNA damage response (DDR) genes –a complex machinery encompassing several pathways that maintain the integrity of DNA within the cell (12)– produce malfunctioning proteins that restrict the ability of cells to repair DNA lesions, rendering them susceptible to genetic instability and cancer development (13); such is the case of ARID1A, a recently studied subunit of the SWI/SNF chromatin remodeler complex whose variants have been associated with breast cancer brain metastasis (14).The presence of these variants also influences treatment choice, as patients carrying them can benefit from treatment alternatives that target the DDR to create synthetic lethality by inhibiting the Poly ADP-ribose (PARP) –a polymerase that synthesizes DNA in the final steps of the repair process– with purpose-designed drugs (15, 16).
Our group is interested in studying the distribution of BRCA variants, particularly in the Mexican-mestizo population, where we have found unreported variants (17, 18), confirming that variation distribution can vary significantly from one geographical location to another (19, 20). So, surveying local populations looking for characteristic individual variations or patterns is an important stepping stone toward universal tailored diagnostics and treatments, especially in Latin American countries where breast cancer is mostly diagnosed in later stages (21).
In this work, we sequenced fourteen DDR genes in a Mexican cohort of 40 metastatic breast cancer patients, searching for an association between variants in DDR genes and response to treatment. The genes we sequenced belong mainly to the DNA Interstrand Crosslink Repair, a DDR pathway that has been recently associated which high tumor burden in breast carcinomas (22); the rest were DNA damage sensors (ATM, CHK2) and transcriptional activators (ARID1A, TP53) that had been recently associated with metastatic breast cancer (23–25). We found 19 unique variants, from which 9 had not been reported before, and a correlation between the combination of alterations in the ARID1A, BRCA1, BRCA2, and FANCA genes and progression-free survival. To our knowledge, the association between alterations in DDR genes other than BRCA and treatment response in metastatic breast cancer had not been surveyed yet in the Mexican-mestizo population.
This study included prospectively tumor biopsy and clinical data from forty stage-IV breast cancer patients that attended the Instituto Nacional de Cancerología (INCan, Mexico City, Mexico. The study was approved by INCan’s Review Board and Ethics Committee (016/010/IBI; CEI/1001/16); all patients signed informed consent. After the surgical excision, tumor biopsies were segmented into two pieces, one for pathological confirmation and another for DNA extraction.
A total of 40 patients were enrolled diagnosed with metastatic breast cancer confirmed by positron emission tomography (PET) and computed tomography (CT) scans. All patients were treated according to the National Comprehensive Cancer Network (NCCN) guidelines. Clinical outcome was evaluated by The Response Evaluation Criteria in Solid Tumors (RECIST Version 1.1) at baseline and at 6 months (26). Progression-free survival (PFS) was defined as the time from the commencement of treatment until disease progression or the last visit. Overall survival (OS) was defined as the time from diagnosis until death or the last visit.
Tumor DNA was extracted using the QIAamp DNA Blood Mini kit (Qiagen, cat. no. 51106). following the manufacturer’s recommendations. DNA integrity was verified by agarose electrophoresis and the concentration was determined using RNase P Detection Reagent (FAM) (Applied Biosystems, cat. no. 4316831).
Fourteen DDR genes were selected for sequencing. Two of them, BRCA1 and BRCA2, were amplified using Ion Ampliseq BRCA 1 and 2 panel (Thermo Fisher Scientific); this panel includes 167 primers pairs in three pools. The remaining genes, ARID1A, ATM, CHK2, FANCA, FANCB, FANCC, FANCD2, PARP1, PALB2, RAD50, RAD51, and TP53 were amplified with the custom panel IAD94476_197_Designed; this panel includes 440 primers in two pools. (Supplementary Table 1). Libraries were prepared from 25 ng DNA, and amplification of each patient’s DDR-genes was identified using a unique Ion Xpress barcode adapter (Thermo Fisher Scientific cat. no. 4471250). For sequencing, we used the Ion PGM Hi-Q Sequencing kit (REFA25589) in the Ion torrent PGM (Personal Genome Machine) instrument (Thermo Fisher Scientific).
The sequences were aligned to the hg19 human reference genome (GRCh37). The.bam files were exported to the Ion Reporter version 5.18 for variation analysis. Pathogenic and probably pathogenic variants were classified according to the American College of Medical Genetics and Genomics guidelines (27)
Kaplan–Meier curves and Cox regressions were calculated using the survival package in R (R version 4.2.2, we used the survival package version 3.4.0.). Variable selection for the Multivariate Cox regressions was performed using a forward stepwise procedure. Statistical significance was defined as p <0.05 (two sided).
The 40 stage-IV screened tumor samples came from patients with a mean age of 53 years, ranging from 27 to 81 (n = 40). Most tumors were ductal (85%) and the remainder, lobular (15%). The molecular type corresponded with previously reported proportions; Luminal A and B tumors were more frequent that HER+ and TNBC. Notably, 16 of the 40 samples came from patients with a family history of cancer (Table 1). Twenty-two of the samples carried at least one variant in the sequenced genes, one had multiple variants in BRCA1, two had multiple variants in TP53, and two had variants in two genes –ARID1A and ATM or FANCA and TP53 (Figure 1, Table 2)
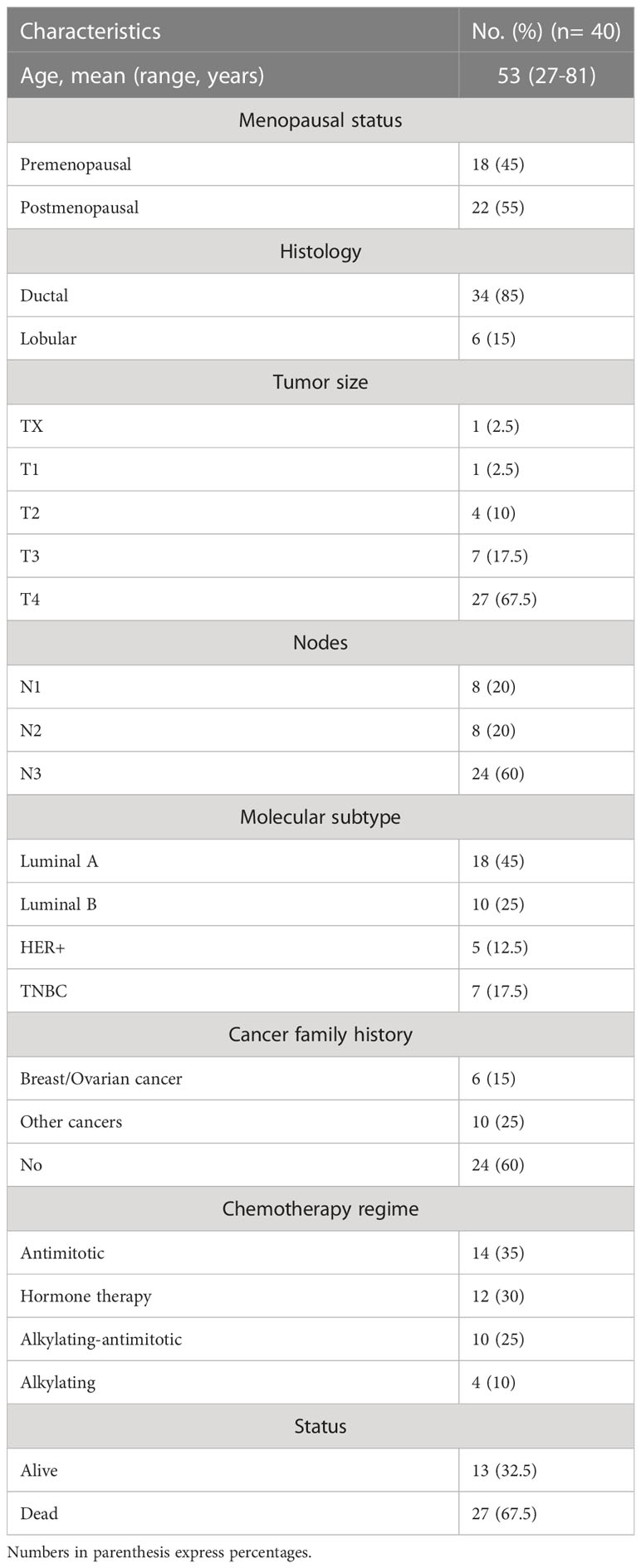
Table 1 Clinical characteristics of 40 unrelated metastatic breast cancer patients.
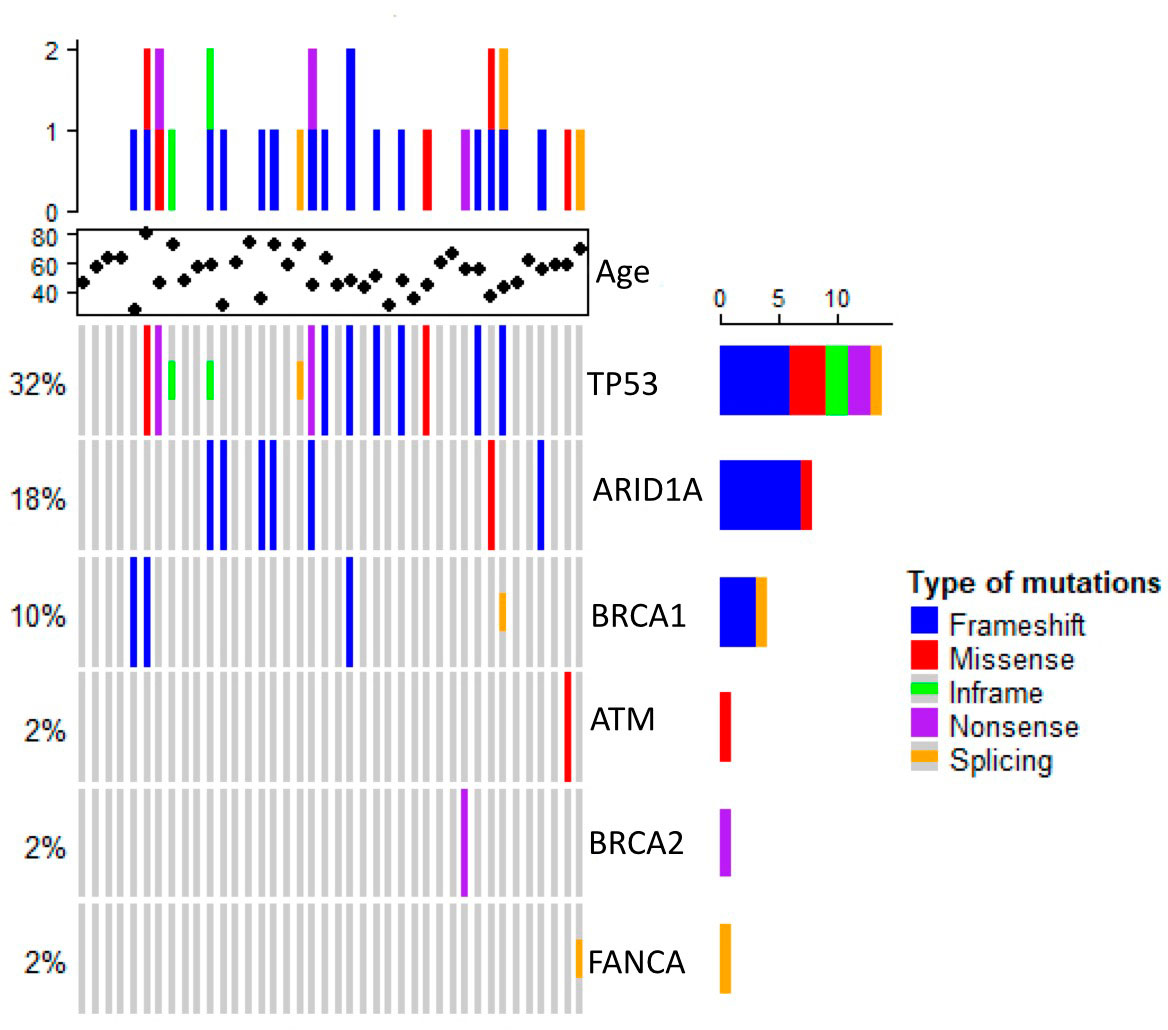
Figure 1 Heat map showing somatic variants profiles of metastatic breast cancer.
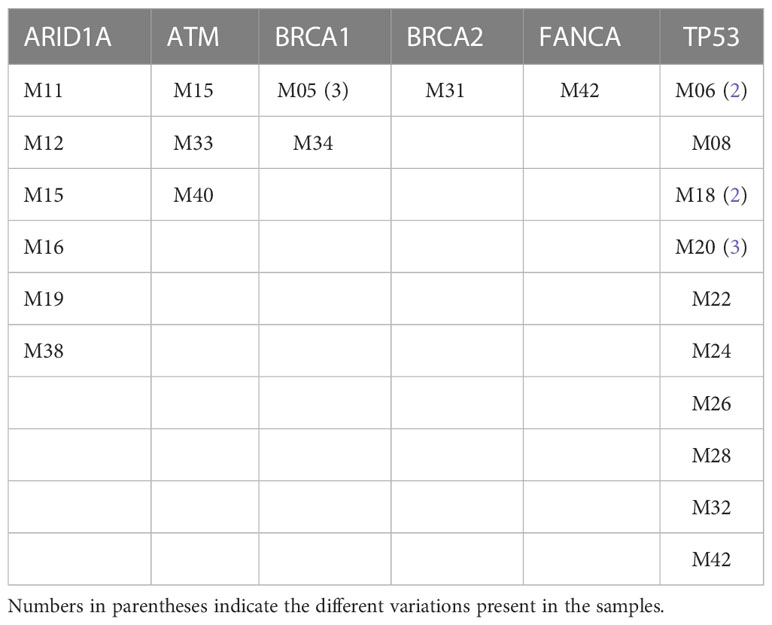
Table 2 Genes with variants in unrelated metastatic breast cancer samples.
Overall, we found 19 unique sequence variants in the 22 samples carrying them, 10 had been already identified and 9 were previously unreported. Ten (52%) of the found variants were in the TP53 gene. The most frequent alterations in this gene –c.742C>T and c.215_216insG– were only the second most frequent, while the first was c.3980_3981insC, in ARID1A. Thirteen variants were found only once in the studied population. Strikingly, no variants in the CHK2, FANCB, FANCC, FANCD2, PALB2, RAD50, or RAD51 genes were present in our cohort (Figure 1, Table 3). Pathogenic or likely pathogenic variants are depicted in Supplementary Table 1.
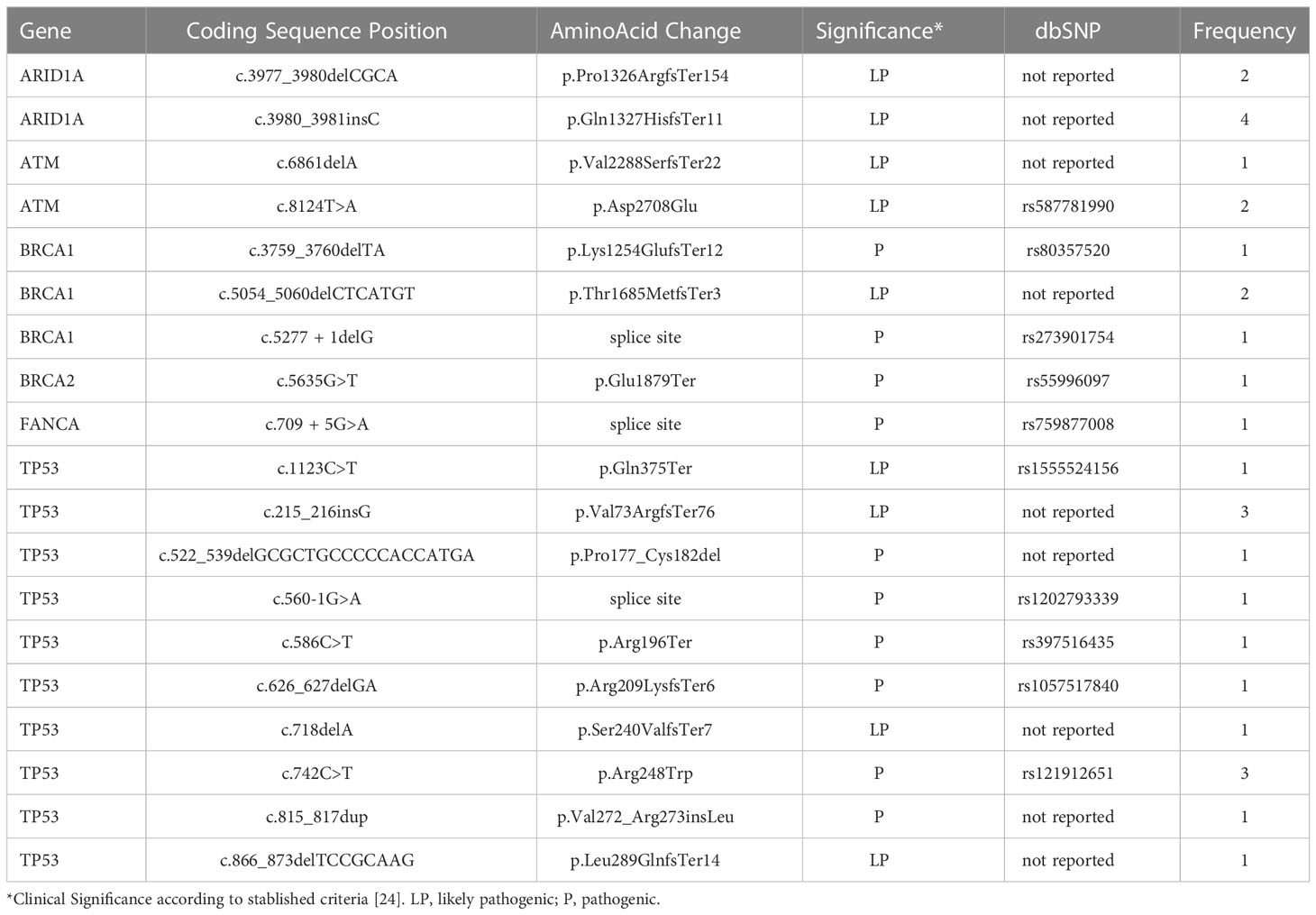
Table 3 Variants found in 40 unrelated metastatic breast cancer samples.
An overall survival analysis was performed considering the patients included with a median follow-up of 5 years (1-13 years). The median PFS of all patients was 6 months (0-45) and median OS was 34 months (1-161). An univariate and multivariate Cox analysis were performed examining the association of the individual variants, with the clinical outcomes (PFS and OS), nevertheless, no significant association was observed (data not shown). So, we considered the genes with highest number of variants observed at the multivariate analysis combined (ARID1A, BRCA1, BRCA2 and FANC genes) and regrouped the variation frequencies of these genes into a single binary variant where at least one variation needed to be met. To assess the association between the combination of these variants and clinical outcome, we compared PFS and OS between patient’s with or without variants in those genes. Median PFS was 8 months in patients without sequence variants in selected genes, versus 4 months in patients with at least one variant of the genes selected (p-value=0.0025) (Figure 2). Besides, the median OS in patients without variants was 51 months versus 31 months for patients with variants of the genes selected (p-value= 0.014) (Figure 3).
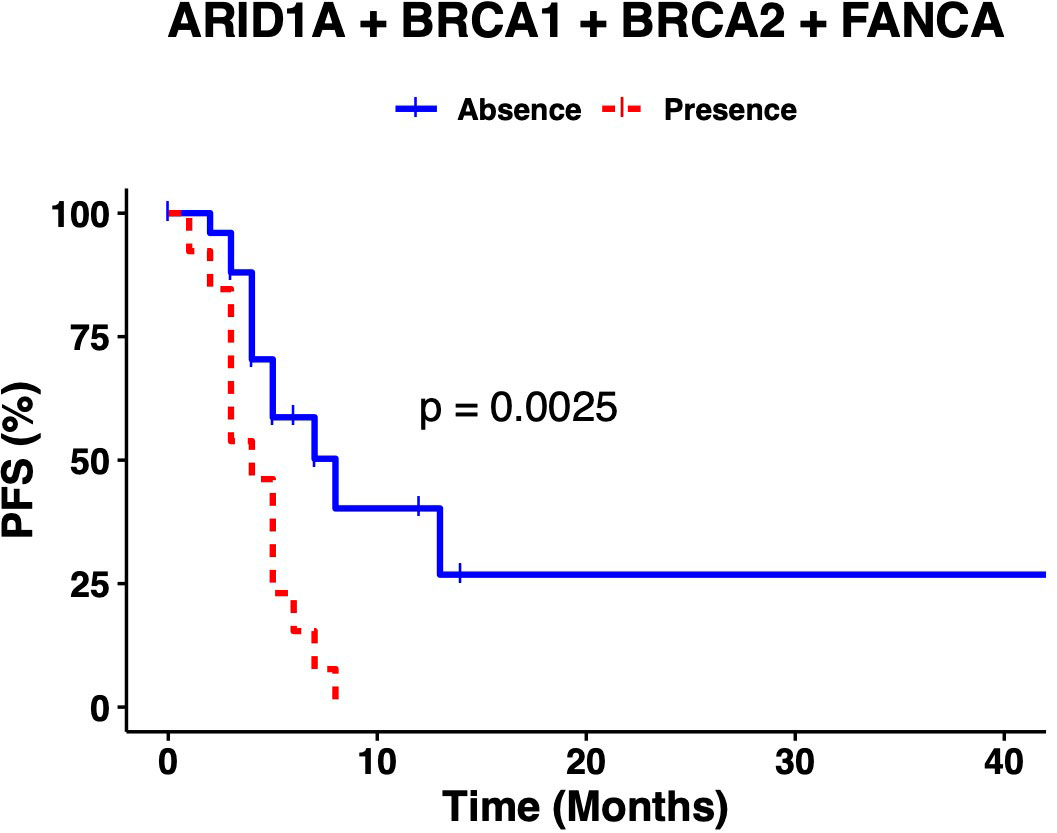
Figure 2 The presence of at least one variant in ARID1A, BRCA1, BRCA2 or FANCA genes correlates with lower PFS.
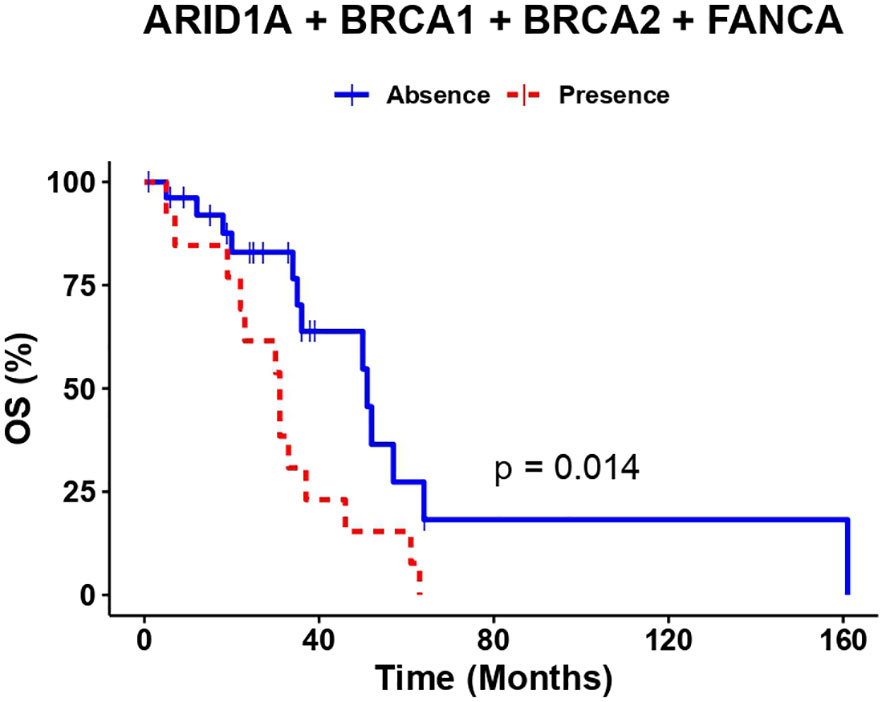
Figure 3 The presence of at least one variant in ARID1A, BRCA1, BRCA2 or FANCA genes correlates with lower OS.
A univariate and multivariate analysis was performed comparing the presence/absence of pathogenic variants in at least one selected gene and clinical outcome (PFS and OS). At PFS Cox models, the presence of at least one pathogenic variant, demonstrated to be predictors of PFS (univariate, HR: 3.3 (95% CI 1.5-7.5), p-value=0.004) (multivariate, HR: 3.74 (95% CI 1.44-9.74), p-value=0.006). Additionally, at OS Cox models, the presence of at least one pathogenic variant, also demonstrated to be predictors of OS (univariate, HR: 2.7 (95% CI 1.2-6), p-value=0.017) (multivariate HR: 2.87 (95% CI 1.07-7.67), p-value=0.035) (Table 4).
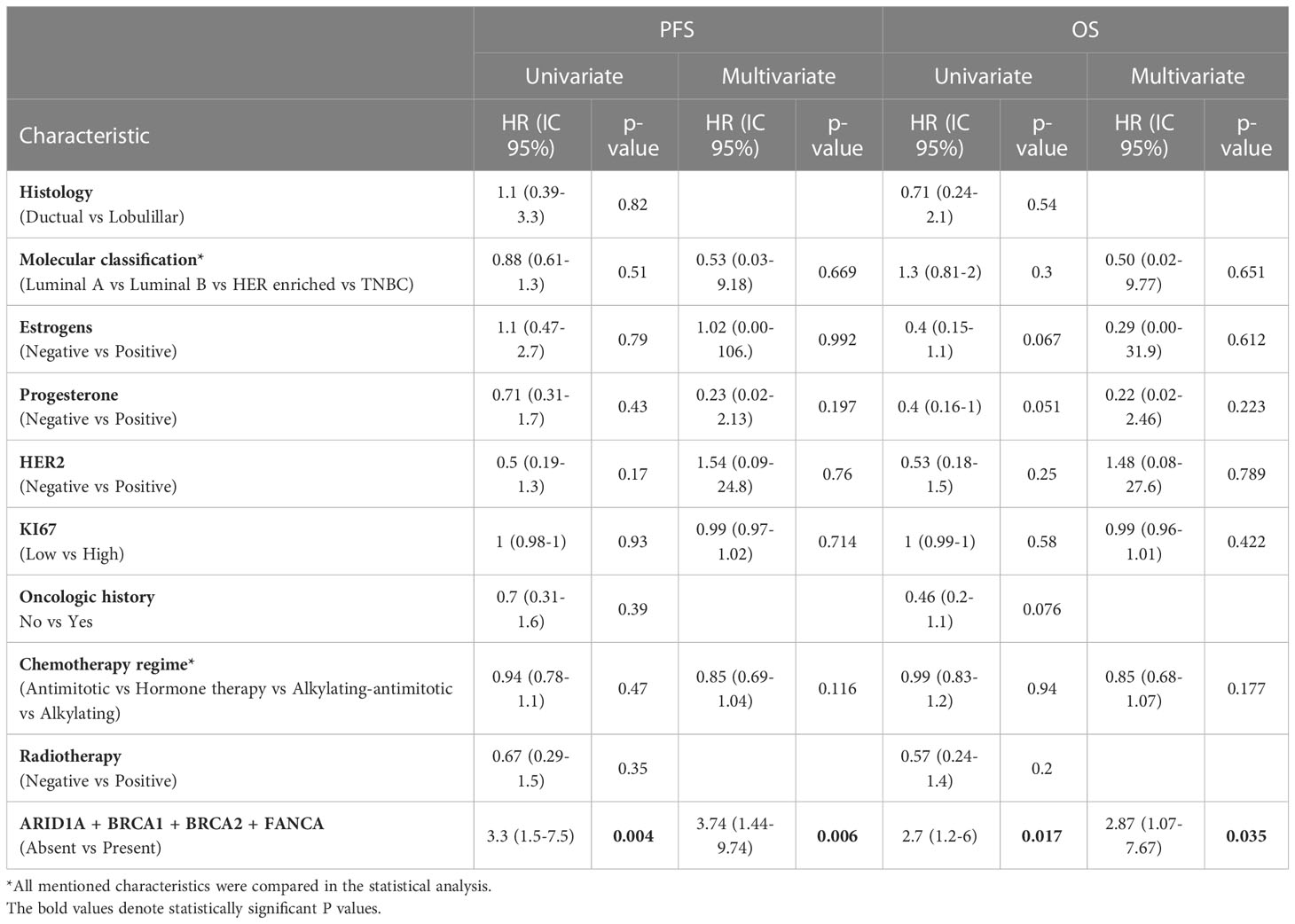
Table 4 Univariate and Multivariate analysis of clinical characteristics and variants in grouped genes (ARID1A + BRCA1 + BRCA2 + FANCA) in the analyzed population.
Variants in DNA damage response (DDR) genes in cancer are important biomarkers for treatment selection and are also functionally important, since malfunctioning DDR can potentially increase genomic instability, eventually leading to treatment resistance or relapse (Reviewed in 28). Here we analyzed a 40-patient cohort of metastatic breast cancer patients in search for variations in DDR genes and found 13 previously identified variants and 9 that had not been reported before. These findings contribute to the understanding of the genomic landscape of metastatic breast cancer in the Mexican-mestizo population which, due to its diverse ancestry (29), is likely to differ from the mostly Caucasian populations of North America (30) and Europe (31) where the genomic characterizations of metastatic breast cancer has been reported.
Sequence variants in TP53 accounted for roughly half of our findings. A similar proportion was previously reported in metastatic tumors (31) but in breast tumors in general, TP53 variants accounted for less than 10% (32), highlighting the high risk of metastasis associated with TP53 variants. Half (5/10) of the variants that we report here had not been reported before, suggesting they are exclusive or more frequent in the Mexican-mestizo population and underlining the importance of studying local populations.
Variants in PALB2 are usually reported in frequencies around 1%, second to the BRCA genes at 3-5% (6, 33, 34). The absence of is PALB2 variants in our cohort is similar to the low incidence observed in a separate study where only two of 115 patients carried PALB2 variants (35), suggesting that PALB2 variants are scarce in the Mexican-mestizo population.
We found a significant association between the presence of at least one pathogenic variant and worse PFS and OS. Three of these genes are related to the DNA Interstrand Crosslink Repair: BRCA1 and BRCA2 are the quintessential breast cancer susceptibility genes (36). FANCA variations are the most frequent in Fanconi anemia (37) and, according to recent reports, it might be the only FANC gene involved in hereditary cancer (38). These findings bolster previous reports on the association between variations in individual genes of the DNA Interstrand Crosslink Repair pathway and breast cancer susceptibility in Iranian (39) and Chinese populations (40). Interestingly, while there is evidence that variations in the DDR pathways –particularly HR– sensitize several cancer types to chemotherapy (15), secondary variants in these genes can generate resistance to alkylating agent therapy (41, 42). These findings suggest that the variants in BRCA1, BRCA2, and FANCA, associated with worse prognosis in our sample set, might have contributed to chemotherapy resistance. The mechanisms underlying this phenomenon will undoubtedly motivate further analysis. Sequence variants in the fourth gene, ARID1A, have been associated with breast cancer brain metastasis, though the specific variants that we found had not been reported before (14).
In Latin America, breast cancer is detected late in its development (21) a trend that can only be reverted through optimized screening strategies. Since tumors with altered ARID1A are sensible to PARP inhibitors (43) and its variants are frequent in cohorts as small as the 40 patients that we report here, we suggest screening tumors for variants ARID1A in the Mexican-mestizo population. Such screening might broaden the treatment options for breast cancer patients, as these variants have been associated with enhanced effects of treatments such as ATR inhibitors and Gemcitabine in ovarian cancer (44). Further studies will confirm whether the high prevalence of ARID1A variants in tumor samples is valid for other Latin American populations and whether there is a functional relationship between ARID1A and the DNA Interstrand Crosslink Repair genes. Additionally, whether the ARID1A variants we observed were acquired during tumor development or already present in the germline and thus related to cancer susceptibility besides response to treatment remains to be determines. If these variants are germline, ARID1A might be a better indicator of cancer risk than PALB2 in the Mexican-mestizo populations.
Our study was limited by the number of genes sequenced and the relatively low number of samples; additionally, the sequencing was performed only from tumor. A larger sample would provide a more comprehensive perspective of the variations in these and other genes; however, our sample included only metastatic breast cancer patients, which represent less than 20% of the total breast cancer cases (45, 46).
Our findings contribute to the description of the sequence variation landscape of metastatic breast cancer in the Mexican-mestizo population. We found the expected high frequency variants in TP53 and BRCA 1 and 2; conversely, PALB2 variants seem scarce compared to other reported populations. The presence of at least one pathogenic variant in the ARID1A, BRCA1, BRCA2, or FANCA genes remains predictor of worse progression-free survival and overall survival.
All data generated or analyzed during this study are included in this published article and are available from the corresponding author on reasonable request.
The studies involving human participants were reviewed and approved by National Cancer Institute of México, Ethics and Scientific Committee. The patients/participants provided their written informed consent to participate in this study.
Conceptualization, CP-P, DC and RV-R; Data curation, AM-G; Formal analysis, OM-C, AC-P, RV-R and AA-M; Investigation, DC; Methodology, ER-G, OM-C, MG-E and NJ-H; Resources, CL-C; Supervision, DC and CP-P; Writing – original draft, CP-P; Writing – review and editing, EL-U and CP-P. All authors contributed to the article and approved the submitted version.
This research was funded by Instituto Nacional de Cancerología Research funds.
OM-C is a doctoral student from Programa de Doctorado en Ciencias Biológicas, Universidad Nacional Autónoma de México (UNAM) and was supported by CONACYT (451334). We want to acknowledge administrative support from the Institutional Research Deputy.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fonc.2023.1146008/full#supplementary-material
1. Sung H, Ferlay J, Siegel RL, Laversanne M, Soerjomataram I, Jemal A, et al. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin (2021) 71:209–49. doi: 10.3322/caac.21660
2. Dillekås H, Rogers MS, Straume O. Are 90% of deaths from cancer caused by metastases? Cancer Med-us (2019) 8:5574–6. doi: 10.1002/cam4.2474
3. Daily K, Douglas E, Romitti PA, Thomas A. Epidemiology of De Novo metastatic breast cancer. Clin Breast Cancer (2021) 21:302–8. doi: 10.1016/j.clbc.2021.01.017
4. Mahdavi M, Nassiri M, Kooshyar MM, Vakili-Azghandi M, Avan A, Sandry R, et al. Hereditary breast cancer; genetic penetrance and current status with BRCA. J Cell Physiol (2019) 234:5741–50. doi: 10.1002/jcp.27464
5. Lang G-T, Shi J-X, Huang L, Cao A-Y, Zhang C-H, Song C-G, et al. Multiple cancer susceptible genes sequencing in BRCA-negative breast cancer with high hereditary risk. Ann Transl Med (2020) 8:1417–7. doi: 10.21037/atm-20-2999
6. Kuusisto KM, Bebel A, Vihinen M, Schleutker J, Sallinen S-L. Screening for BRCA1, BRCA2, CHEK2, PALB2, BRIP1, RAD50, and CDH1 mutations in high-risk Finnish BRCA1/2-founder mutation-negative breast and/or ovarian cancer individuals. Breast Cancer Res (2011) 13:R20. doi: 10.1186/bcr2832
7. Antoniou AC, Casadei S, Heikkinen T, Barrowdale D, Pylkäs K, Roberts J, et al. Breast-cancer risk in families with mutations in PALB2. N Engl J Med (2014) 371:497–506. doi: 10.1056/nejmoa1400382
8. Guerra E, Cimadamore A, Simeone P, Vacca G, Lattanzio R, Botti G, et al. p53, cathepsin d, bcl-2 are joint prognostic indicators of breast cancer metastatic spreading. BMC Cancer (2016) 16:649. doi: 10.1186/s12885-016-2713-3
9. Zhang P, Kitchen-Smith I, Xiong L, Stracquadanio G, Brown K, Richter PH, et al. Germline and somatic genetic variants in the p53 pathway interact to affect cancer risk, progression, and drug response. Cancer Res (2021) 81:1667–80. doi: 10.1158/0008-5472.can-20-0177
10. Grünewald TGP, Delattre O. Cooperation between somatic mutations and germline susceptibility variants in tumorigenesis – a dangerous liaison. Mol Cell Oncol (2016) 3:e1086853. doi: 10.1080/23723556.2015.1086853
11. Pennington KP, Walsh T, Harrell MI, Lee MK, Pennil CC, Rendi MH, et al. Germline and somatic mutations in homologous recombination genes predict platinum response and survival in ovarian, fallopian tube, and peritoneal carcinomas. Clin Cancer Res (2014) 20:764–75. doi: 10.1158/1078-0432.ccr-13-2287
12. Kciuk M, Bukowski K, Marciniak B, Kontek R. Advances in DNA repair–emerging players in the arena of eukaryotic DNA repair. Int J Mol Sci (2020) 21:3934. doi: 10.3390/ijms21113934
13. Suh KJ, Ryu HS, Lee K-H, Kim H, Min A, Kim T-Y, et al. Prognostic effects of abnormal DNA damage response protein expression in breast cancer. Breast Cancer Res Tr (2019) 175:117–27. doi: 10.1007/s10549-019-05128-9
14. Morgan AJ, Giannoudis A, Palmieri C. The genomic landscape of breast cancer brain metastases: a systematic review. Lancet Oncol (2021) 22:e7–e17. doi: 10.1016/s1470-2045(20)30556-8
15. Patel PS, Algouneh A, Hakem R. Exploiting synthetic lethality to target BRCA1/2-deficient tumors: where we stand. Oncogene (2021) 40:3001–14. doi: 10.1038/s41388-021-01744-2
16. Rose M, Burgess JT, O’Byrne K, Richard DJ, Bolderson E. PARP inhibitors: clinical relevance, mechanisms of action and tumor resistance. Front Cell Dev Biol (2020) 8:564601. doi: 10.3389/fcell.2020.564601
17. Vaca-Paniagua F, Alvarez-Gomez RM, Fragoso-Ontiveros V, Vidal-Millan S, Herrera LA, Cantú D, et al. Full-exon pyrosequencing screening of BRCA germline mutations in Mexican women with inherited breast and ovarian cancer. PloS One (2012) 7:e37432. doi: 10.1371/journal.pone.0037432
18. Millan-Catalan O, Campos-Parra AD, Vázquez-Romo R, de León DC, Jacobo-Herrera N, Morales-González F, et al. A multi-center study of BRCA1 and BRCA2 germline mutations in Mexican-mestizo breast cancer families reveals mutations unreported in Latin American population. Cancers (2019) 11:1246. doi: 10.3390/cancers11091246
19. Rebbeck TR, Friebel TM, Friedman E, Hamann U, Huo D, Kwong A, et al. Mutational spectrum in a worldwide study of 29,700 families with BRCA1 or BRCA2 mutations. Hum Mutat (2018) 39:593–620. doi: 10.1002/humu.23406
20. Macchini M, Centonze F, Peretti U, Orsi G, Militello AM, Valente MM, et al. Epidemiology and geographic distribution of BRCA1-2 and DNA damage response genes pathogenic variants in pancreatic ductal adenocarcinoma patients. Cancer Treat Rev (2022) 104:102357. doi: 10.1016/j.ctrv.2022.102357
21. Pinto JA, Pinillos L, Villarreal-Garza C, Morante Z, Villarán MV, Mejía G, et al. Barriers in Latin America for the management of locally advanced breast cancer. Ecancermedicalscience (2019) 13:897. doi: 10.3332/ecancer.2019.897
22. Mei P, Freitag CE, Wei L, Zhang Y, Parwani AV, Li Z. High tumor mutation burden is associated with DNA damage repair gene mutation in breast carcinomas. Diagn Pathol (2020) 15:50. doi: 10.1186/s13000-020-00971-7
23. Cheng X, Zhao J-X, Dong F, Cao X-C. ARID1A mutation in metastatic breast cancer: a potential therapeutic target. Front Oncol (2021) 11:759577. doi: 10.3389/fonc.2021.759577
24. Bai H, Yu J, Jia S, Liu X, Liang X, Li H. Prognostic value of the TP53 mutation location in metastatic breast cancer as detected by next-generation sequencing. Cancer Manage Res (2021) 13:3303–16. doi: 10.2147/cmar.s298729
25. Stucci LS, Internò V, Tucci M, Perrone M, Mannavola F, Palmirotta R, et al. The ATM gene in breast cancer: its relevance in clinical practice. Genes-basel (2021) 12:727. doi: 10.3390/genes12050727
26. Eisenhauer EA, Therasse P, Bogaerts J, Schwartz LH, Sargent D, Ford R, et al. New response evaluation criteria in solid tumours: revised RECIST guideline (version 1.1). Eur J Cancer (2009) 45:228–47. doi: 10.1016/j.ejca.2008.10.026
27. Hampel H, Bennett RL, Buchanan A, Pearlman R, Wiesner GL, Committee GDG American College of Medical Genetics and Genomics Professional Practice and Guidelines Committee and National Society of Genetic Counselors Practice Guidelines. A practice guideline from the American college of medical genetics and genomics and the national society of genetic counselors: referral indications for cancer predisposition assessment. Genet Med (2015) 17:70–87. doi: 10.1038/gim.2014.147
28. Huang R, Zhou P-K. DNA Damage repair: historical perspectives, mechanistic pathways and clinical translation for targeted cancer therapy. Signal Transduct Target Ther (2021) 6:254. doi: 10.1038/s41392-021-00648-7
29. Moreno-Estrada A, Gignoux CR, Fernández-López JC, Zakharia F, Sikora M, Contreras AV, et al. The genetics of Mexico recapitulates native American substructure and affects biomedical traits. Science (2014) 344:1280–5. doi: 10.1126/science.1251688
30. Paul MR, Pan T, Pant DK, Shih NNC, Chen Y, Harvey KL, et al. Genomic landscape of metastatic breast cancer identifies preferentially dysregulated pathways and targets. J Clin Invest (2020) 130:4252–65. doi: 10.1172/jci129941
31. Bertucci F, Ng CKY, Patsouris A, Droin N, Piscuoglio S, Carbuccia N, et al. Genomic characterization of metastatic breast cancers. Nature (2019) 569:560–4. doi: 10.1038/s41586-019-1056-z
32. Schon K, Tischkowitz M. Clinical implications of germline mutations in breast cancer: TP53. Breast Cancer Res Tr (2018) 167:417–23. doi: 10.1007/s10549-017-4531-y
33. Shimelis H, LaDuca H, Hu C, Hart SN, Na J, Thomas A, et al. Triple-negative breast cancer risk genes identified by multigene hereditary cancer panel testing. Jnci J Natl Cancer Inst (2018) 110:855–62. doi: 10.1093/jnci/djy106
34. Cragun D, Weidner A, Tezak A, Clouse K, Pal T. Cancer risk management among female BRCA1/2, PALB2, CHEK2, and ATM carriers. Breast Cancer Res Tr (2020) 182:421–8. doi: 10.1007/s10549-020-05699-y
35. Gómez-Flores-Ramos L, Barraza-Arellano AL, Mohar A, Trujillo-Martínez M, Grimaldo L, Ortiz-Lopez R, et al. Germline variants in cancer genes from young breast cancer Mexican patients. Cancers (2022) 14:1647. doi: 10.3390/cancers14071647
36. Armstrong N, Ryder S, Forbes C, Ross J, Quek RG. A systematic review of the international prevalence of BRCA mutation in breast cancer. Clin Epidemiol (2019) 11:543–61. doi: 10.2147/clep.s206949
37. Solomon PJ, Margaret P, Rajendran R, Ramalingam R, Menezes GA, Shirley AS, et al. A case report and literature review of fanconi anemia (FA) diagnosed by genetic testing. Ital J Pediatr (2015) 41:38. doi: 10.1186/s13052-015-0142-6
38. del Valle J, Rofes P, Moreno-Cabrera JM, López-Dóriga A, Belhadj S, Vargas-Parra G, et al. Exploring the role of mutations in fanconi anemia genes in hereditary cancer patients. Cancers (2020) 12:829. doi: 10.3390/cancers12040829
39. Abbasi S, Rasouli M. A rare FANCA gene variation as a breast cancer susceptibility allele in an Iranian population. Mol Med Rep (2017) 15:3983–8. doi: 10.3892/mmr.2017.6489
40. Pan Z-W, Wang X-J, Chen T, Ding X-W, Jiang X, Gao Y, et al. Deleterious mutations in DNA repair gene FANCC exist in BRCA1/2-negative Chinese familial breast and/or ovarian cancer patients. Front Oncol (2019) 9:169. doi: 10.3389/fonc.2019.00169
41. Edwards SL, Brough R, Lord CJ, Natrajan R, Vatcheva R, Levine DA, et al. Resistance to therapy caused by intragenic deletion in BRCA2. Nature (2008) 451:1111–5. doi: 10.1038/nature06548
42. Imyanitov E, Sokolenko A. Mechanisms of acquired resistance of BRCA1/2-driven tumors to platinum compounds and PARP inhibitors. World J Clin Oncol (2021) 12:544–56. doi: 10.5306/wjco.v12.i7.544
43. Shen J, Peng Y, Wei L, Zhang W, Yang L, Lan L, et al. ARID1A deficiency impairs the DNA damage checkpoint and sensitizes cells to PARP inhibitors. Cancer Discovery (2015) 5:752–67. doi: 10.1158/2159-8290.cd-14-0849
44. Mullen J, Kato S, Sicklick JK, Kurzrock R. Targeting ARID1A mutations in cancer. Cancer Treat Rev (2021) 100:102287. doi: 10.1016/j.ctrv.2021.102287
45. Siegel RL, Miller KD, Fuchs HE, Jemal A. Cancer statistics, 2022. CA Cancer J Clin (2022) 72:7–33. doi: 10.3322/caac.21708
Keywords: metastatic breast cancer, DNA damage response, Latin American population, Mexican-mestizo population, Progression free survival (PFS)
Citation: Vázquez-Romo R, Millan-Catalan O, Ruíz-García E, Martínez-Gutiérrez AD, Alvarado-Miranda A, Campos-Parra AD, López-Camarillo C, Jacobo-Herrera N, López-Urrutia E, Guardado-Estrada M, Cantú de León D and Pérez-Plasencia C (2023) Pathogenic variant profile in DNA damage response genes correlates with metastatic breast cancer progression-free survival in a Mexican-mestizo population. Front. Oncol. 13:1146008. doi: 10.3389/fonc.2023.1146008
Received: 16 January 2023; Accepted: 14 April 2023;
Published: 27 April 2023.
Edited by:
Chunyan Dong, Tongji University, ChinaReviewed by:
Gianluca Tedaldi, Laboratory of Biosciences (IRCCS), ItalyCopyright © 2023 Vázquez-Romo, Millan-Catalan, Ruíz-García, Martínez-Gutiérrez, Alvarado-Miranda, Campos-Parra, López-Camarillo, Jacobo-Herrera, López-Urrutia, Guardado-Estrada, Cantú de León and Pérez-Plasencia. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: David Cantú de León, ZGZjYW50dUBnbWFpbC5jb20=; Carlos Pérez-Plasencia, Y2FybG9zLnBwbGFzQGdtYWlsLmNvbQ==
†These authors have contributed equally to this work
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.