 Bo-Tao Zhang
Bo-Tao Zhang Jia-Ying Xu
Jia-Ying Xu Wei Wang
Wei Wang Yang Zeng2*
Yang Zeng2* Jun Jiang
Jun Jiang
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
MINI REVIEW article
Front. Oncol. , 16 March 2023
Sec. Cancer Epidemiology and Prevention
Volume 13 - 2023 | https://doi.org/10.3389/fonc.2023.1125178
This article is part of the Research Topic Obesity and Cancer: Update on Etiology, Molecular Biomarkers and Biotargets, Clinical Strategies, and Epidemiology View all 10 articles
There is increasing evidence that obesity is associated with the occurrence and development of malignant tumors. When studying the relationship between obesity and malignant tumors, it is very important to choose an appropriate animal model. However, BALB/c nude mice and other animals commonly used to study tumor xenograft (human-derived tumor cell lines) transplantation models are difficult to induce obesity, while C57BL/6 mice and other model animals commonly used for obesity research are not suitable for tumor xenograft transplantation. Therefore, it is difficult to replicate both obesity and malignancy in animal models at the same time. This review summarizes several experimental animal models and protocols that can simultaneously induce obesity and tumor xenografts.
Over the past few decades, obesity has become a growing global health problem. From 1975 to 2016, the global prevalence of obesity nearly tripled, affecting 13% of the world’s adult population (1). A large body of epidemiologic evidence shows that obesity is associated with the incidence and progress of several cancers. According to the World Cancer Research Fund’s Third Expert Report, obesity is an important risk factor for many types of cancer (2). The mechanisms linking obesity and cancer development remain unclear. The impact of obesity on human health may take decades to become apparent. Therefore, the use of experimental animals to study the effects of obesity on cancer is of great importance for the discovery of the phenomenon and the study of the mechanism.
Researchers often use preclinical animal models to study the relationship between obesity and disease. Because gene knockout and transgenic technology cannot fully reflect the pathogenesis and pathogenic factors of obesity, the current modeling method is still based on food inducing. Immunodeficient mice are widely used in cancer research. Because xenografts can be performed, they provide researchers with insight into the growth, invasion, and metastasis of human tumor cells. In addition, researchers have also created several types of genetically engineered mouse models (GEMMs) that can spontaneously develop cancer.
However, replicating both obesity and malignancy in laboratory animals is extremely difficult. Animals commonly used in obesity models cannot engraft heterolytic tumors. On the other hand, it is difficult to induce obesity in animals commonly used in cancer models. This situation leaves researchers with limited options. In this review, we discuss the mouse model and related experimental strategies for obesity and cancer research.
Animal models of obesity are diverse and include both mammalian and nonmammalian species. Non-mammals have certain limitations due to major anatomical and physiological differences from humans (3). Therefore, mammals are usually considered the ideal animal model for obesity research. Among mammals, mice are most used. This is because of their small body size, high reproductive capacity, relatively short life cycle, and relatively easy genome editing (4, 5).
Diet-induced obesity (DIO) is an important model of obesity and results from excessive consumption of a high-fat diet (HFD), which usually contains 45-60% fat (6). DIO can simulate the development of human obesity better than genetic models (7, 8) and commonly use the mouse as the model (9). Consumption of HFD can lead to central obesity and insulin resistance in mice and is a good research alternative to mimic diet-induced obesity in humans.
Among inbred mice, C57BL/6J, BALB/c, KM, and ICR mice are commonly used to reproduce DIO models (10). Other inbred strains, such as SWR/J and A/J mice, are less sensitive to high-fat diets and related complications (11). The C57BL/6J has the advantage of short modeling time and stable metrics, so it is the most widely used. The C57BL/6J is more susceptible to fat accumulation, weight gain, and glucose metabolism disorders when fed a high-fat diet, as manifested by significant changes in abdominal fat weight, Lee’s index, and adipocyte volume.
The weight of C57BL/6J mice gradually increased with age, reaching the peak at approximately 9 months (12). Compared with the younger mice, the older ones (22 months old or older) had less muscle and more fat (13). Male mice are often used in experimental studies to induce obesity because they are more sensitive to high-fat diets and are prone to diet-induced insulin resistance and abnormal glucose tolerance (14, 15). Compared with male mice, female mice gain weight slowly, have a low obesity rate, and are generally resistant to high-fat diet-induced obesity (16, 17). However, because brown adipose tissue is easier to observe in female mice, female C57BL/6J mice are generally used to study the role of brown adipose tissue in energy metabolism (18).
Two types of spontaneously obese mice based on C57BL/6J were identified at the Jackson Laboratory, ob/ob mice in 1950 and db/db mice in 1965. The ob/ob mice lack functional leptin, whereas the db/db mice lack functional leptin receptors. Both types of mice exhibit overeating and are the primary mouse models for studies of monogenic obesity (4, 7). The ob/ob mice have a single base pair mutation in the ob gene, resulting in the absence of functional leptin, increased body weight, hyperphagia, and a low resting metabolic rate. On the other hand, due to a defect in the leptin receptor, leptin signaling is impaired in the db/db mice, resulting in significantly higher serum leptin levels. Therefore, the treatment of reorganization is sufficient to make ob/ob mice normal (19), but it is not effective for db/db mice (20). In addition, the two types of mice are the same in obesity, hypogonadism, and growth hormone (GH) deficiency (4, 6).
Monogenic obesity models have become important research tools in modern drug discovery. The ob/ob mouse is commonly used to evaluate the efficacy of new obesity drugs in overcoming the obesity phenotype caused by overeating (21), and db/db mice are commonly used to study the efficacy of antidiabetic drugs (22). These models require only short-term rather than long-term feeding to induce obesity. However, monogenic models generally do not represent the full pathogenesis of human obesity. Monogenic obesity in humans accounts for only a small proportion of obesity, and a few of human obesity can be explained by mutations in leptin or leptin receptors alone.
Compared to the monogenic model, the polygenic model can better simulate the pathogenesis of human obesity. The C57BL/6J mouse is the most used obese mouse model, which is susceptible to obesity induced by overeating. However, only 60% of C57BL/6J mice gain weight under high-fat diet conditions. The susceptibility of C57BL/6J mice to diet-induced obesity is typically characterized by changes in plasma insulin and leptin levels and insulin sensitivity at 6 weeks of age (23). New Zealand obese (NZO) mice are polygenic inbred mice predisposed to obesity and type 2 diabetes. Unlike C57BL/6J mice, NZO mice can gain weight on a standard diet (24).
Mice have similar biological, physiological and pathological characteristics to humans and exhibit a high degree of genetic similarity, making them an ideal animal model for the study of tumors. Much of the current understanding of human cancer characteristics are based on long-term in vitro culture of tumor cell lines and their inoculation into mice.
Currently, most tumor implantation models used in basic or translational oncology research are based on established cell lines (25). They usually function as allografts of primary mouse tumors or xenografts of human tumors. In both types of models, cancer cells can be injected orthotopically or ectopically (mainly subcutaneously) and subsequently monitored for growth or metastasis (intraperitoneally, intravenously, or intracardially).
Since 1950, allografts have been used primarily as a preclinical model for drug development and cancer therapy (26). For example, researchers established the leukemia model using male DBA/2 mice and found that AZD2014, an mTORC1/2 inhibitor, inhibited the growth and proliferation of L1210 leukemia cells (27). The toxicology of some cytotoxic drugs has also been successfully studied in allograft models. However, allograft tumor models are of limited value for the study of human tumors. Therefore, xenograft tumor models have replaced allograft tumor models as the primary tool for preclinical drug testing since 1990.
The discovery of the thymus-free nude mouse was a major breakthrough in cancer research, allowing human tumors to be replicated in xenogeneic experimental animals. Immunodeficient mice have remarkable xenograft success rates and are able to preserve the original tissue structure and function of human cancers. Representative immunodeficient mice include nude mice, severe combined immunodeficiency (SCID) mice, non-obese diabetic/SCID (NOD/SCID) mice, and NOD-SCID-IL2Rg-/- (NSG) mice (Table 1). SCID mice have been shown to be more suitable for human cancer cell xenografts than nude mice and have advantages for studying the biology of human tumors in vivo and their response to therapy (28).
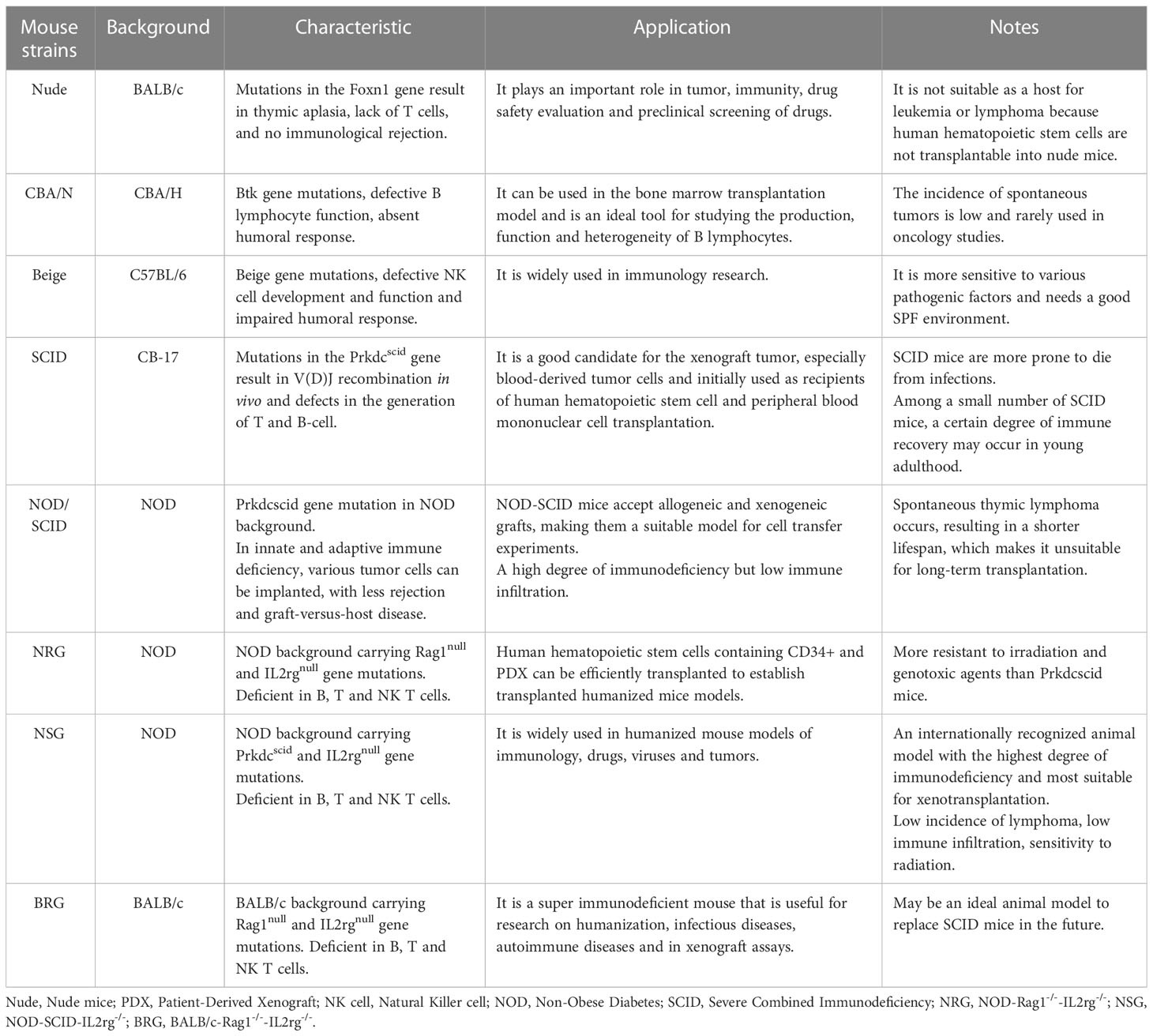
Table 1 Characteristics and application of common immunodeficient mice.
Ectopic transplantation typically inoculates human cancer cell lines or pieces of tumor tissue under the skin in the axilla, back and hind legs of mice. After subcutaneous inoculation, the tumor tissue is surrounded by a thick fibrous capsule and rarely metastasizes to adjacent tissues. Tumor growth can be easily observed and the treatment efficacy can be evaluated.
The microenvironment of in situ implanted tumors is different from that of ectopically implanted tumors, and therefore their growth rates are different. Because growing in an optimal microenvironment, in situ implanted tumors generally exhibit more active proliferation, metastasis, and invasion, which better mimics the growth of tumors in the human body (29). Fu XY et al. orthotopically implanted human colon cancer cells in the colon of nude mice. The transplanted tumors almost exactly replicated the characteristics of the corresponding human cancer, which included local tumor growth, abdominal metastasis with peritoneal seeding, liver metastasis, lymph node metastasis, and intestinal obstruction (30). Carmelo Nucera et al. established an orthotopic model of human thyroid cancer using the anaplastic thyroid carcinoma cell line 8305C and observed tumor growth and metastasis (31). However, because the volume and number of tumors in the visceral organs are not easily measured, there are cases where tumor ectopic transplantation is more appropriate.
The above ectopic and orthotopic transplantation models, also called spontaneous tumor metastasis model. The method of injecting cancer cells directly into the blood to study their spread and metastasis is called experimental tumor metastasis model. The experimental metastasis model is used to study the growth of malignant tumors in distant organs. Intravenous injection can shorten the time of tumor formation in target organs. Inoculation via the tail vein is one of the most used methods in the experimental metastasis model. For example, Nan Huo et al. established a lung metastasis model for thyroid cancer by injecting TPC-1 cells into BALB/c nude mice via the tail vein (32).
In the 1980s, the development of transgenic and gene-targeting technologies in mouse embryonic stem cells facilitated the generation of GEMM. The most common ways to generate GEMM are to activate oncogenes or inactivate tumor-suppressor genes in vivo through the use of transgenic and gene targeting methods, such as knock-outs and knock-ins. Gordon et al. established the first transgenic mice in 1980, harboring randomly integrated oncogenes under the control of a tissue-specific promoter (33). The initial set of genetic engineering tools was set against the background of the emergence of genome-editing technologies such as restriction endonucleases, DNA cloning and sequencing, and then developed lentiviral vector, electrotransfection and microinjection techniques. In 2016, the single-base gene editing technology developed from CRISPR-Cas9 avoided DNA double-strand breaks and further expanded the scope of base editing. The innovation of gene editing technologies has significantly reduced the time needed to establish a GEMM (34). GEMM has been used in the study of colorectal cancer (35), renal cell carcinoma (36) and breast cancer (37). In addition, it can be used in preclinical trials for hormonal and targeted therapies as well as immunotherapy. PD-1 KO and PD-L1 KO mice have been exploited to develop drugs for cancer treatment (38).
In vivo animal models are important research tools to study the underlying mechanisms of the association between obesity and cancers. Among genetic models of obesity, mice deficient in leptin signaling are the most used. When mice were fed standard chow, the genetic model showed early-onset obesity and comorbid diseases such as insulin resistance and hepatic steatosis. Their main disadvantage is the exclusion of the factors other than leptin that may affect cancer cells and tumor microenvironment. For example, obesity accelerates the progress of Kras-driven pancreatic ductal adenocarcinoma, but not lung cancer (39).
The DIO mouse model is believed to mimic human obesity well and to explain the potential biological link between obesity and cancer. The DIO model was established by feeding mice a diet high in sugar, fat or both. While several feeding regimens have been developed, the most commonly used diets contain 30% to 60% kcal from fat, which is fed to the mice for 10 to 12 weeks prior to tumor formation.
Most obesity-related complications are due to inflammation (40). Chronic inflammation in adipose tissue, especially white adipose tissue (WAT), stimulates cancer progression through mechanisms such as altered levels of adipokines and inflammatory mediators, and insulin resistance (41–43). Short-term HFD feeding is difficult to obtain an ideal model sufficient to study the relationship between obesity and cancer (44). Therefore, long-term obesity models need to be established to simulate the relationship between human obesity and tumors. The feeding time of the HFD-induced obesity mouse model ranged from 4 weeks to 56 weeks, and 10 weeks to 12 weeks were usually selected (Table 2). DIO mice gain weight, increase fasting blood glucose levels, and develop obesity-related phenotypes such as hyperinsulinemia, insulin resistance, hepatic steatosis, hypertension, and dyslipidemia (59). Whether reversal of the obesity phenotype affects tumor prognosis is a key question in this field. Dietary pattern switching experiments have shown that once DIO is established, a low-fat diet (LFD) for a prolonged period, such as 5 weeks, is sufficient to reverse obesity-induced chronic inflammation and tumor progression (44, 52).
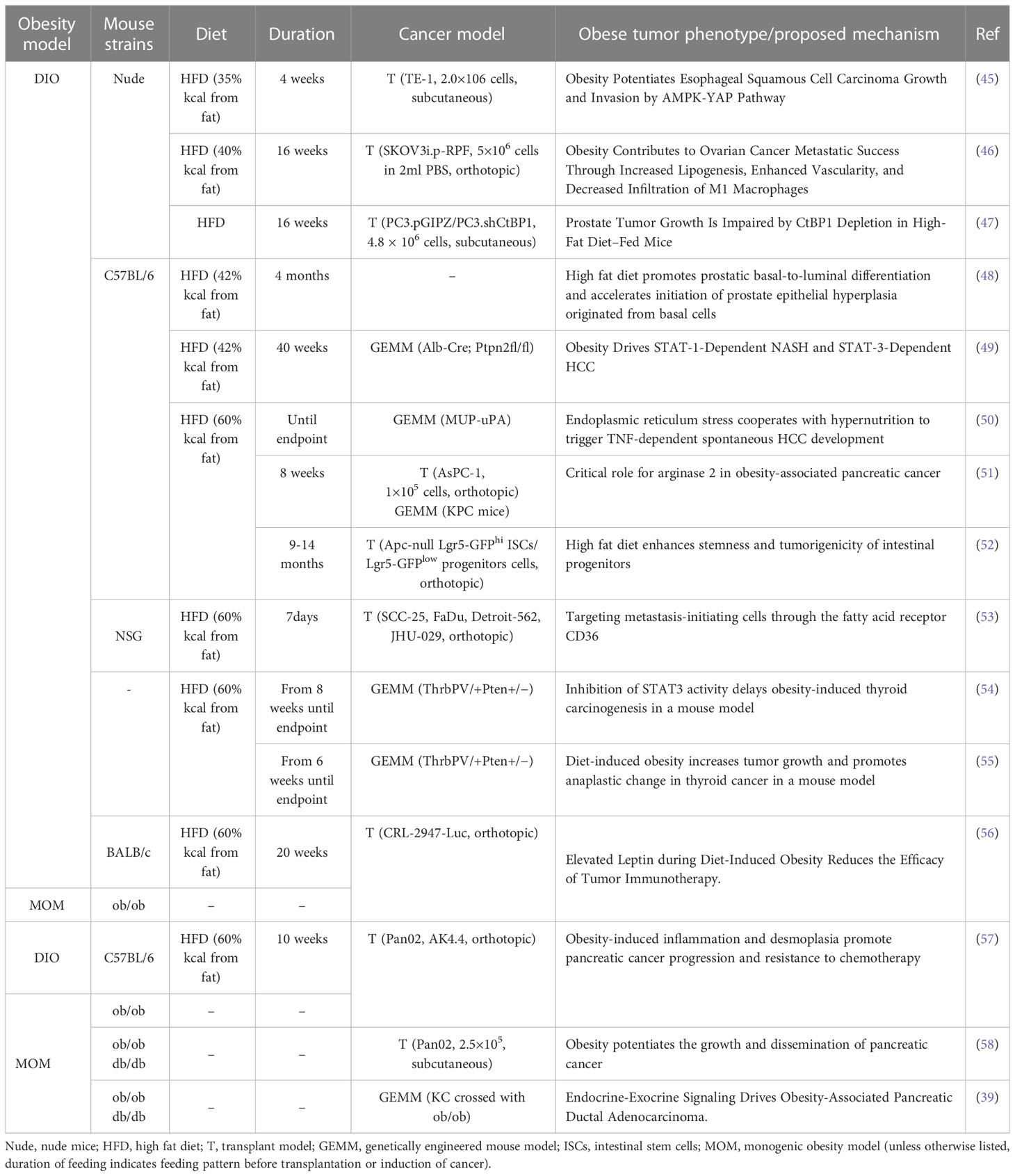
Table 2 Overview of obesity-associated cancer model.
Nude mice used to establish tumor xenograft models, such as BALB/c, are generally difficult to induce obesity. Stemmer K et al. found that Foxn1 nude mice (B6. Cg-Foxn1nu/J) on a C57BL/6 background fed a high-fat diet under thermoneutral (33°C) conditions significantly increased their body weight (60), making them an excellent model for studying obesity and tumors.
Obesity is an important risk factor for cancer. Significant attention has been paid to the underlying mechanism between the two diseases. Appropriate animal models replicating both obesity and cancer are highly needed to study their association. A brief review shows that there is currently no single ideal model for this type of research (Table 2). The models listed are good for studying tumor progression and metastasis, but there are also some shortcomings. They cannot determine how diet and obesity contribute to cancer initiation and be used to study cancer survivorship.
The mouse models utilize high-fat diets to achieve obese condition but the typical western diet that is most closely associated with obesity and cancer is composed of a dietary pattern comprised of high protein and fat but most importantly very high in refined sugars (61, 62). This particular dietary pattern is not similar to mouse models and although it would be difficult to replicate in models the shortcomings should be noted (63). Humans who are exposed to high carbohydrate diets will not only lead to weight gain and obesity, but exacerbate glucose/insulin homeostasis which could be an important underlying mechanism associated with the progression of cancer independent of obesity or perhaps in synergy (64). Furthermore, a western dietary pattern has been associated with inflammation (65–67) and this is another important exposure that is missing in most animal models of cancer.
When selecting an appropriate mouse model, factors such as obese phenotype, environmental stimuli, mouse strain and sex should be considered more fully. With the development of different mouse models, the combined application of multiple models makes cancer research more convenient and accurate. Recently, the emergence of a revolutionary CRISPR/Cas9 system has greatly enhanced the efficiency of precise gene editing in various GEMMs. However, the potential risk of off-target effects is a notable concern. An ideal cancer + obesity mouse model should be technically simple, quick in operation, easily reproducible, affordable and short in modeling. Further improvement of obesity-prone mice that can be implanted with human tumor cells will help decipher the mechanism by which obesity affects tumor initiation and progression.
JJ and YZ contributed the central idea and analyzed most of the data. B-TZ and J-YX wrote the initial draft of the paper, WW contributed to refining the ideas and carrying out additional analyses. All authors reviewed the manuscript. All authors contributed to the article and approved the submitted version.
This study was supported by the National Natural Science Foundation of China (82070288), Sichuan Province Science & Technology Program (2022YFS0627), the Health Commission of Sichuan Province (21PJ100), the Office of Science and Technology and Intellectual Property of Luzhou (2022-JYJ-131), and the Affiliated Stomatological Hospital of Southwest Medical University Program (2022Y02).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
1. NCD Risk Factor Collaboration (NCD-RisC). Worldwide trends in body-mass index, underweight, overweight, and obesity from 1975 to 2016: A pooled analysis of 2416 population-based measurement studies in 128.9 million children, adolescents, and adults. Lancet (2017) 390(10113):2627–42. doi: 10.1016/S0140-6736(17)32129-3
2. World Cancer Research Fund/American Institute for Cancer Research. Diet, nutrition, physical activity and cancer: A global perspective. continuous update project expert report 2018 (2018). Available at: https://www.wcrf.org/diet-activity-and-cancer/global-cancer-update-programme/about-the-third-expert-report/.
3. Polakof S, Panserat S, Soengas JL, Moon TW. Glucose metabolism in fish: A review. J Comp Physiol B (2012) 182(8):1015–45. doi: 10.1007/s00360-012-0658-7
4. Kleinert M, Clemmensen C, Hofmann SM, Moore MC, Renner S, Woods SC, et al. Animal models of obesity and diabetes mellitus. Nat Rev Endocrinol (2018) 14(3):140–62. doi: 10.1038/nrendo.2017.161
5. Doulberis M, Papaefthymiou A, Polyzos SA, Katsinelos P, Grigoriadis N, Srivastava DS, et al. Rodent models of obesity. Minerva Endocrinol (2020) 45(3):243–63. doi: 10.23736/S0391-1977.19.03058-X
6. Research Diets. DIO series diets (2020). Available at: https://researchdiets.com/opensource-diets/dio-series-diets.
7. da Silva Xavier G, Hodson DJ. Mouse models of peripheral metabolic disease. Best Pract Res Clin Endocrinol Metab (2018) 32(3):299–315. doi: 10.1016/j.beem.2018.03.009
8. Barrett P, Mercer JG, Morgan PJ. Preclinical models for obesity research. Dis Model Mech (2016) 9(11):1245–55. doi: 10.1242/dmm.026443
9. Picklo MJ Sr, Idso J, Seeger DR, Aukema HM, Murphy EJ. Comparative effects of high oleic acid vs high mixed saturated fatty acid obesogenic diets upon PUFA metabolism in mice. Prostaglandins Leukot Essent Fatty Acids (2017) 119:25–37. doi: 10.1016/j.plefa.2017.03.001
10. Li J, Wu H, Liu Y, Yang L. High fat diet induced obesity model using four strainsof mice: Kunming, C57BL/6, BALB/c and ICR. Exp Anim (2020) 69(3):326–35. doi: 10.1538/expanim.19-0148
11. Surwit RS, Feinglos MN, Rodin J, Sutherland A, Petro AE, Opara EC, et al. Differential effects of fat and sucrose on the development of obesity and diabetes in C57BL/6J and AJ mice. Metabolism (1995) 44(5):645–51. doi: 10.1016/0026-0495(95)90123-x
12. van der Heijden RA, Bijzet J, Meijers WC, Yakala GK, Kleemann R, Nguyen TQ, et al. Obesity-induced chronic inflammation in high fat diet challenged C57BL/6J mice is associated with acceleration of age-dependent renal amyloidosis. Sci Rep (2015) 5:16474. doi: 10.1038/srep16474
13. Houtkooper RH, Argmann C, Houten SM, Cantó C, Jeninga EH, Andreux PA, et al. The metabolic footprint of aging in mice. Sci Rep (2011) 1:134. doi: 10.1038/srep00134
14. Garg N, Thakur S, McMahan CA, Adamo ML. High fat diet induced insulin resistance and glucose intolerance are gender-specific in IGF-1R heterozygous mice. Biochem Biophys Res Commun (2011) 413(3):476–80. doi: 10.1016/j.bbrc.2011.08.123
15. Pettersson US, Waldén TB, Carlsson PO, Jansson L, Phillipson M. Female mice are protected against high-fat diet induced metabolic syndrome and increase the regulatory T cell population in adipose tissue. PloS One (2012) 7(9):e46057. doi: 10.1371/journal.pone.0046057
16. Bruder-Nascimento T, Ekeledo OJ, Anderson R, Le HB, Belin de Chantemèle EJ. Long term high fat diet treatment: An appropriate approach to study the sex-specificity of the autonomic and cardiovascular responses to obesity in mice. Front Physiol (2017) 8:32. doi: 10.3389/fphys.2017.00032
17. Ingvorsen C, Karp NA, Lelliott CJ. The role of sex and body weight on the metabolic effects of high-fat diet in C57BL/6N mice. Nutr Diabetes (2017) 7(4):e261. doi: 10.1038/nutd.2017.6
18. Li J, Zhao WG, Shen ZF, Yuan T, Liu SN, Liu Q, et al. Comparative proteome analysis of brown adipose tissue in obese C57BL/6J mice using iTRAQ-coupled 2D LC-MS/MS. PloS One (2015) 10(3):e0119350. doi: 10.1371/journal.pone.0119350
19. Pelleymounter MA, Cullen MJ, Baker MB, Hecht R, Winters D, Boone T, et al. Effects of the obese gene product on body weight regulation in ob/ob mice. Science (1995) 269(5223):540–3. doi: 10.1126/science.7624776
20. Carroll L, Voisey J, van Daal A. Mouse models of obesity. Clin Dermatol (2004) 22(4):345–9. doi: 10.1016/j.clindermatol.2004.01.004
21. Yashiro H, Hamagami K, Hiyoshi H, Sugama J, Tsuchimori K, Yamaguchi F, et al. SCO-792, an enteropeptidase inhibitor, improves disease status of diabetes and obesity in mice. Diabetes Obes Metab (2019) 21(10):2228–39. doi: 10.1111/dom.13799
22. Ma T, Huo S, Xu B, Li F, Wang P, Liu Y, et al. A novel long-acting oxyntomodulin analogue eliminates diabetes and obesity in mice. Eur J Med Chem (2020) 203:112496. doi: 10.1016/j.ejmech.2020.112496
23. Baumeier C, Saussenthaler S, Kammel A, Jähnert M, Schlüter L, Hesse D, et al. Hepatic DPP4 DNA methylation associates with fatty liver. Diabetes (2017) 66(1):25–35. doi: 10.2337/db15-1716
24. Korovila I, Höhn A, Jung T, Grune T, Ott C. Reduced liver autophagy in high-fat diet induced liver steatosis in new Zealand obese mice. Antioxidants (Basel) (2021) 10(4):501. doi: 10.3390/antiox10040501
25. Gengenbacher N, Singhal M, Augustin HG. Preclinical mouse solid tumour models: status quo, challenges and perspectives. Nat Rev Cancer (2017) 17(12):751–65. doi: 10.1038/nrc.2017.92
26. Suggitt M, Bibby MC. 50 years of preclinical anticancer drug screening: empirical to target-driven approaches. Clin Cancer Res (2005) 11(3):971–81. doi: 10.1158/1078-0432.971.11.3
27. Liao HF, Lin YZ, Yu CC, Tai TS, Hung SK, Yang CC, et al. mTORC1/2 inhibitor served as a more ideal agent against the growth of mouse lymphocytic leukemia both In vitro and in vivo. Anticancer Res (2019) 39(9):4829–35. doi: 10.21873/anticanres.13668
28. Bankert RB, Hess SD, Egilmez NK. SCID mouse models to study human cancer pathogenesis and approaches to therapy: Potential, limitations, and future directions. Front Biosci (2002) 7:c44–62. doi: 10.2741/A758
29. Fidler IJ. Critical factors in the biology of human cancer metastasis: Twenty-eighth G.H.A. clowes memorial award lecture. Cancer Res (1990) 50(19):6130–8.
30. Fu XY, Besterman JM, Monosov A, Hoffman RM. Models of human metastatic colon cancer in nude mice orthotopically constructed by using histologically intact patient specimens. Proc Natl Acad Sci USA (1991) 88(20):9345–9. doi: 10.1073/pnas.88.20.9345
31. Nucera C, Nehs MA, Mekel M, Zhang X, Hodin R, Lawler J, et al. A novel orthotopic mouse model of human anaplastic thyroid carcinoma. Thyroid (2009) 19(10):1077–84. doi: 10.1089/thy.2009.0055
32. Huo N, Cong R, Sun ZJ, Li WC, Zhu X, Xue CY, et al. STAT3/LINC00671 axis regulates papillary thyroid tumor growth and metastasis via LDHA-mediated glycolysis. Cell Death Dis (2021) 12(9):799. doi: 10.1038/s41419-021-04081-0
33. Gordon JW, Scangos GA, Plotkin DJ, Barbosa JA, Ruddle FH. Genetic transformation of mouse embryos by microinjection of purified DNA. Proc Natl Acad Sci USA (1980) 77(12):7380–4. doi: 10.1073/pnas.77.12.7380
34. Mou H, Kennedy Z, Anderson DG, Yin H, Xue W. Precision cancer mouse models through genome editing with CRISPR-Cas9. Genome Med (2015) 7(1):53. doi: 10.1186/s13073-015-0178-7
35. Sui H, Zhang L, Gu K, Chai N, Ji Q, Zhou L, et al. YYFZBJS ameliorates colorectal cancer progression in ApcMin/+ mice by remodeling gut microbiota and inhibiting regulatory T-cell generation. Cell Commun Signal (2020) 18(1):113. doi: 10.1186/s12964-020-00596-9
36. Lee JC, Yotis DM, Lee JY, Sarabusky MA, Shrum B, Champagne A, et al. Kidney injury molecule-1 inhibits metastasis of renal cell carcinoma. Sci Rep (2021) 11(1):11840. doi: 10.1038/s41598-021-90919-8
37. Mori H, Chen JQ, Cardiff RD, Pénzváltó Z, Hubbard NE, Schuetter L, et al. Pathobiology of the 129:Stat1 mouse model of human age-related ER-positive breast cancer with an immune infiltrate-excluded phenotype. Breast Cancer Res (2017) 19(1):102. doi: 10.1186/s13058-017-0892-8
38. Cao S, Li J, Lu J, Zhong R, Zhong H. Mycobacterium tuberculosis antigens repress Th1 immune response suppression and promotes lung cancer metastasis through PD-1/PDl-1 signaling pathway. Cell Death Dis (2019) 10(2):44. doi: 10.1038/s41419-018-1237-y
39. Chung KM, Singh J, Lawres L, Dorans KJ, Garcia C, Burkhardt DB, et al. Endocrine-exocrine signaling drives obesity-associated pancreatic ductal adenocarcinoma. Cell (2020) 181(4):832–847.e18. doi: 10.1016/j.cell.2020.03.062
40. Ellulu MS, Patimah I, Khaza’ai H, Rahmat A, Abed Y. Obesity and inflammation: the linking mechanism and the complications. Arch Med Sci (2017) 13(4):851–63. doi: 10.5114/aoms.2016.58928
41. Iyengar NM, Hudis CA, Dannenberg AJ. Obesity and cancer: Local and systemic mechanisms. Annu Rev Med (2015) 66:297–309. doi: 10.1146/annurev-med-050913-022228
42. Iyengar NM, Gucalp A, Dannenberg AJ, Hudis CA. Obesity and cancer mechanisms: Tumor microenvironment and inflammation. J Clin Oncol (2016) 34(35):4270–6. doi: 10.1200/JCO.2016.67.4283
43. Pham DV, Park PH. Tumor metabolic reprogramming by adipokines as a critical driver of obesity-associated cancer progression. Int J Mol Sci (2021) 22(3):1444. doi: 10.3390/ijms22031444
44. Tiwari P, Blank A, Cui C, Schoenfelt KQ, Zhou G, Xu Y, et al. Metabolically activated adipose tissue macrophages link obesity to triple-negative breast cancer. J Exp Med (2019) 216(6):1345–58. doi: 10.1084/jem.20181616
45. Liu JH, Wu QF, Fu JK, Che XM, Li HJ. Obesity potentiates esophageal squamous cell carcinoma growth and invasion by AMPK-YAP pathway. J Immunol Res (2020) 2020:6765474. doi: 10.1155/2020/6765474
46. Liu Y, Metzinger MN, Lewellen KA, Cripps SN, Carey KD, Harper EI, et al. Obesity contributes to ovarian cancer metastatic success through increased lipogenesis, enhanced vascularity, and decreased infiltration of M1 macrophages. Cancer Res (2015) 75(23):5046–57. doi: 10.1158/0008-5472.CAN-15-0706
47. Moiola CP, De Luca P, Zalazar F, Cotignola J, Rodríguez-Seguí SA, Gardner K, et al. Prostate tumor growth is impaired by CtBP1 depletion in high-fat diet-fed mice. Clin Cancer Res (2014) 20(15):4086–95. doi: 10.1158/1078-0432.CCR-14-0322
48. Kwon OJ, Zhang B, Zhang L, Xin L. High fat diet promotes prostatic basal-to-luminal differentiation and accelerates initiation of prostate epithelial hyperplasia originated from basal cells. Stem Cell Res (2016) 16(3):682–91. doi: 10.1016/j.scr.2016.04.009
49. Grohmann M, Wiede F, Dodd GT, Gurzov EN, Ooi GJ, Butt T, et al. Obesity drives STAT-1-Dependent NASH and STAT-3-Dependent HCC. Cell (2018) 175(5):1289–1306.e20. doi: 10.1016/j.cell.2018.09.053
50. Nakagawa H, Umemura A, Taniguchi K, Font-Burgada J, Dhar D, Ogata H, et al. ER stress cooperates with hypernutrition to trigger TNF-dependent spontaneous HCC development. Cancer Cell (2014) 26(3):331–43. doi: 10.1016/j.ccr.2014.07.001
51. Zaytouni T, Tsai PY, Hitchcock DS, DuBois CD, Freinkman E, Lin L, et al. Critical role for arginase 2 in obesity-associated pancreatic cancer. Nat Commun (2017) 8(1):242. doi: 10.1038/s41467-017-00331-y
52. Beyaz S, Mana MD, Roper J, Kedrin D, Saadatpour A, Hong SJ, et al. High-fat diet enhances stemness and tumorigenicity of intestinal progenitors [published correction appears in nature. Nature (2016) 531(7592):53–8. doi: 10.1038/nature17173
53. Pascual G, Avgustinova A, Mejetta S, Martín M, Castellanos A, Attolini CS, et al. Targeting metastasis-initiating cells through the fatty acid receptor CD36. Nature (2017) 541(7635):41–5. doi: 10.1038/nature20791
54. Park JW, Han CR, Zhao L, Willingham MC, Cheng SY. Inhibition of STAT3 activity delays obesity-induced thyroid carcinogenesis in a mouse model. Endocr Relat Cancer (2016) 23(1):53–63. doi: 10.1530/ERC-15-0417
55. Kim WG, Park JW, Willingham MC, Cheng SY. Diet-induced obesity increases tumor growth and promotes anaplastic change in thyroid cancer in a mouse model. Endocrinology (2013) 154(8):2936–47. doi: 10.1210/en.2013-1128
56. Murphy KA, James BR, Sjaastad FV, Kucaba TA, Kim H, Brincks EL, et al. Cutting edge: Elevated leptin during diet-induced obesity reduces the efficacy of tumor immunotherapy. J Immunol (2018) 201(7):1837–41. doi: 10.4049/jimmunol.1701738
57. Incio J, Liu H, Suboj P, Chin SM, Chen IX, Pinter M, et al. Obesity-induced inflammation and desmoplasia promote pancreatic cancer progression and resistance to chemotherapy. Cancer Discov (2016) 6(8):852–69. doi: 10.1158/2159-8290.CD-15-1177
58. Zyromski NJ, Mathur A, Pitt HA, Wade TE, Wang S, Nakshatri P, et al. Obesity potentiates the growth and dissemination of pancreatic cancer. Surgery (2009) 146(2):258–63. doi: 10.1016/j.surg.2009.02.024
59. Della Vedova MC, Muñoz MD, Santillan LD, Plateo-Pignatari MG, Germanó MJ, Rinaldi Tosi ME, et al. A mouse model of diet-induced obesity resembling most features of human metabolic syndrome. Nutr Metab Insights (2016) 9:93–102. doi: 10.4137/NMI.S32907
60. Stemmer K, Kotzbeck P, Zani F, Bauer M, Neff C, Müller TD, et al. Thermoneutral housing is a critical factor for immune function and diet-induced obesity in C57BL/6 nude mice. Int J Obes (Lond) (2015) 39(5):791–7. doi: 10.1038/ijo.2014.187
61. Matsushita M, Fujita K, Nonomura N. Influence of diet and nutrition on prostate cancer. Int J Mol Sci (2020) 21(4):1447. doi: 10.3390/ijms21041447
62. Preguiça I, Alves A, Nunes S, Fernandes R, Gomes P, Viana SD, et al. Diet-induced rodent models of obesity-related metabolic disorders-a guide to a translational perspective. Obes Rev (2020) 21(12):e13081. doi: 10.1111/obr.13081
63. Masi LN, Martins AR, Crisma AR, do Amaral CL, Davanso MR, Serdan TDA, et al. Combination of a high-fat diet with sweetened condensed milk exacerbates inflammation and insulin resistance induced by each separately in mice. Sci Rep (2017) 7(1):3937. doi: 10.1038/s41598-017-04308-1
64. García-Montero C, Fraile-Martínez O, Gómez-Lahoz AM, Pekarek L, Castellanos AJ, Noguerales-Fraguas F, et al. Nutritional components in Western diet versus Mediterranean diet at the gut microbiota-immune system interplay. implications for health and disease. Nutrients (2021) 13(2):699. doi: 10.3390/nu13020699
65. Gentile CL, Weir TL. The gut microbiota at the intersection of diet and human health. Science (2018) 362(6416):776–80. doi: 10.1126/science.aau5812
66. Singh RK, Chang HW, Yan D, Lee KM, Ucmak D, Wong K, et al. Influence of diet on the gut microbiome and implications for human health. J Transl Med (2017) 15(1):73. doi: 10.1186/s12967-017-1175-y
Keywords: obesity, cancer, diet-induced obesity, mouse model, preclinical disease model
Citation: Zhang B-T, Xu J-Y, Wang W, Zeng Y and Jiang J (2023) Obesity and cancer: Mouse models used in studies. Front. Oncol. 13:1125178. doi: 10.3389/fonc.2023.1125178
Received: 04 January 2023; Accepted: 07 March 2023;
Published: 16 March 2023.
Edited by:
Eva Surmacz, Allysta Pharmaceuticals, Inc., United StatesReviewed by:
Guilherme Zweig Rocha, State University of Campinas, BrazilCopyright © 2023 Zhang, Xu, Wang, Zeng and Jiang. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jun Jiang, amlhbmdqdW5Ac3dtdS5lZHUuY24=; Yang Zeng, WmVuZ3lhbmc4MjZAc3dtdS5lZHUuY24=
†These authors have contributed equally to this work
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.