 Xinhui Du1,2,3*†
Xinhui Du1,2,3*† Hua Wei4†
Hua Wei4† Boya Zhang1,2,3Bangmin Wang1,2,3Zhehuang Li1,2,3
Boya Zhang1,2,3Bangmin Wang1,2,3Zhehuang Li1,2,3 Lon Kai Pang5
Lon Kai Pang5 Ruiying Zhao6
Ruiying Zhao6 Weitao Yao1,2,3
Weitao Yao1,2,3- 1Bone Soft Tissue Department, The Affiliated Cancer Hospital of Zhengzhou University and Henan Cancer Hospital, Zhengzhou, China
- 2Key Laboratory for Digital Assessment of Spinal-Pelvic Tumor and Surgical Aid Tools Design (Zhengzhou), Zhengzhou, Henan, China
- 3Key Laboratory for Perioperative Digital Assessment of Bone Tumors (Henan), Zhengzhou, Henan, China
- 4Department of Anesthesiology, Pain and Perioperative Medicine, The First Affiliated Hospital of Zhengzhou University, Zhengzhou, Henan, China
- 5Baylor College of Medicine, Houston, TX, United States
- 6Department of Integrative Biology and Pharmacology, McGovern Medical School, The University of Texas Health Science Center at Houston, Houston, TX, United States
In osteosarcoma patients, metastasis of the primary cancer is the leading cause of death. At present, management options to prevent metastasis are limited and non-curative. In this study, we review the current state of knowledge on the molecular mechanisms of metastasis and discuss promising new therapies to combat osteosarcoma metastasis. Genomic and epigenomic changes, metabolic reprogramming, transcription factors, dysregulation of physiologic pathways, and alterations to the tumor microenvironment are some of the changes reportedly involved in the regulation of osteosarcoma metastasis. Key factors within the tumor microenvironment include infiltrating lymphocytes, macrophages, cancer-associated fibroblasts, platelets, and extracellular components such as vesicles, proteins, and other secreted molecules. We conclude by discussing potential osteosarcoma-limiting agents and their clinical studies.
Introduction
Osteosarcoma is the most common primary malignant bone tumor in children and young adults. Current treatment options for osteosarcoma include neoadjuvant chemotherapy, wide tumor resection, and adjuvant chemotherapy. Unfortunately, these treatment options are limited in efficacy, and management outcomes have not improved in the last 30 years. The 5-year overall survival of osteosarcoma patients with primary localized tumors is 60%–70%, whereas survival drops to approximately 20% in patients with metastasis (1). Distant metastasis is found in approximately 10% of patients at diagnosis, but eventually develops in approximately 50% of patients, commonly contributing to death (2). Hence, one approach to improving overall survival in patients with osteosarcoma is to prevent or delay tumor metastasis. While the mechanisms governing osteosarcoma metastasis remain unclear, developments in molecular technology have enabled us to study osteosarcoma and other cancers more closely. These findings help to pave the way towards novel, effective, and hopefully curative therapies.
In this review, we discuss recent studies that highlight potential factors implicated in osteosarcoma metastasis (Figure 1), and highlight a few emerging anti-cancer agents with potential anti-metastatic activity.
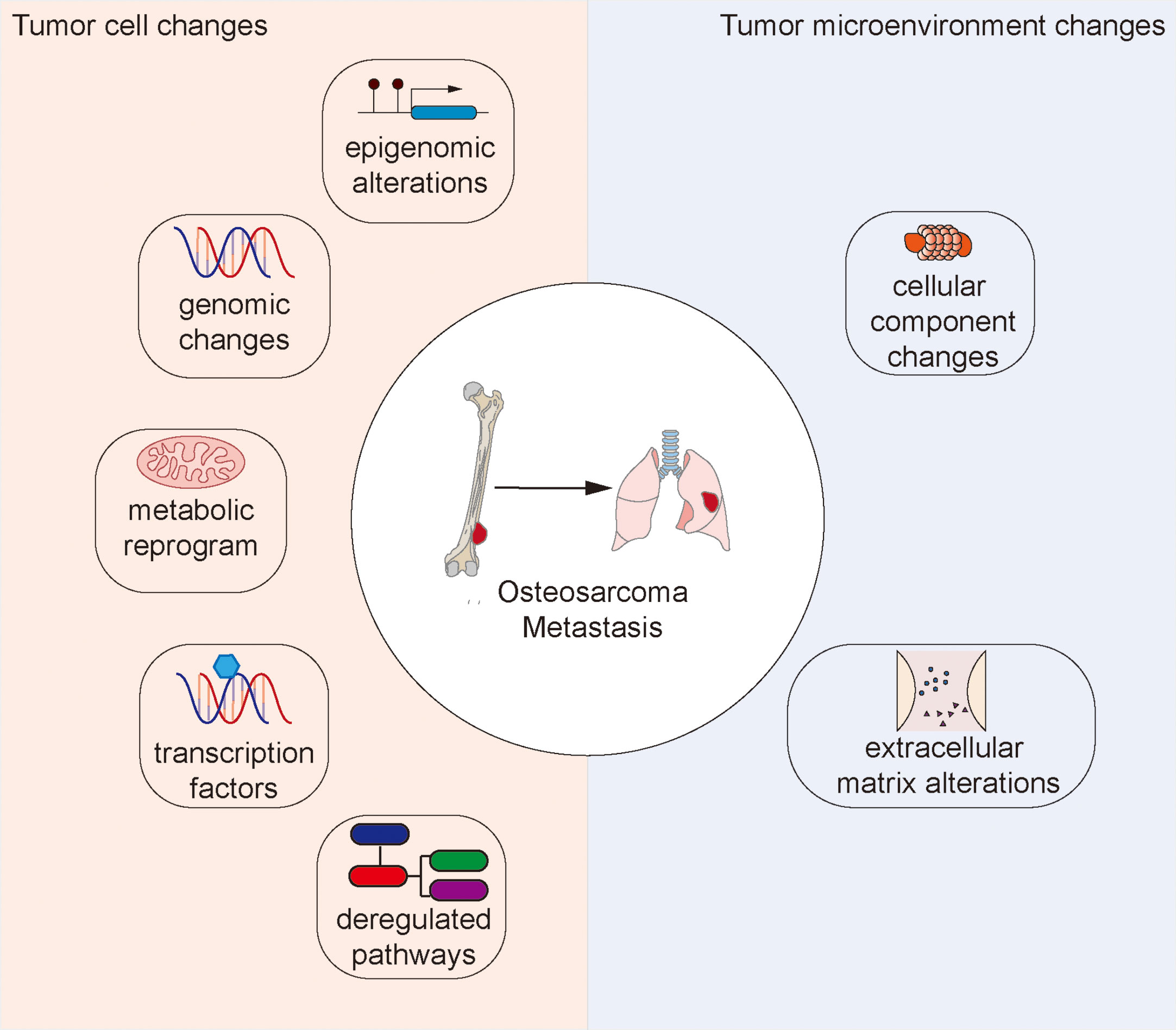
Figure 1 Changes to the tumor cell and tumor microenvironment that facilitate osteosarcoma metastasis.
Tumor cell alterations
Genomic alterations
The genomic profile of osteosarcoma differs greatly from that of other malignant tumors. For example, unlike in breast cancer or melanoma, few targetable recurrent point mutations exist within the protein-coding genes identified in osteosarcoma. In addition, widespread recurrent somatic copy number alterations (SCNAs) and structural rearrangements have been detected and proposed to be responsible for osteosarcoma carcinogenesis and progression. Even among osteosarcoma patients, SCNAs and structural rearrangements are highly heterogeneous (3).
Among osteosarcoma samples, metastatic tumors demonstrate significantly higher mutational burden and genomic instability than primary tumors. Mutated genes are enriched in the PI3K-Akt pathway at both the early and late stages of tumor evolution and in the MAPK pathway at the metastatic stage (4).
Examination of metastatic samples of osteosarcoma revealed alterations in key genes that may play vital roles in metastasis. These alterations include the loss of TP53, RB1, and CDKN2A, or the gain of MYC and MDM2 (4). TP53 is commonly mutated in various cancers including osteosarcoma, and most of the mutations occur in the DNA-binding domain and are characterized as either structural or contact mutations. In addition to inhibitory effects on wild-type TP53 activity, gain-of-function activity promoting tumor progression was also noted. Studies have shown that contact mutations are stronger drivers of osteosarcoma metastasis (5).
RB1 is a well-established tumor suppressor gene reported to be mutated in multiple malignant tumor types including osteosarcoma. RB1 mutation in osteosarcoma is responsible for tumor carcinogenesis and progression. At a molecular level, RB1 loss leads to aberrant spliceosome function due to the upregulation of E2F3a, a mediator of spliceosome gene expression (6).
Amplification of 17p11.2 chromosomal region containing TOP3A led to increased expression of TOP3A, which supported the maintenance of telomeres through the alternative lengthening of telomeres (ALT) mechanism in osteosarcoma (3).
Structural rearrangements in osteosarcoma can also result in novel fusion genes that may participate in tumor progression and metastasis. For example, the fusion gene Rab22a-NeoF1 was detected in osteosarcoma samples. The resultant fusion protein activates RhoA and promotes cell migration, invasion, and lung metastasis after acetylation on K7 (7). When secreted, it also alters the function of adjacent tumor-negative cells and stimulates macrophages toward M2 polarization (8).
Personalized therapy targeting patient-specific genes with copy number alterations and expression changes was tested in patient-derived tumor xenografts and showed a significant decrease in tumor burden (9).
Epigenomic changes
Epigenetic changes are commonly found in osteosarcoma and are involved in multiple aspects of tumor progression including metastasis (10). For example, the methyltransferase DNMT3A inhibits miR-149 expression by DNA methylation to activate the NOTCH1/Hedgehog pathway, thereby promoting the proliferation and metastasis of osteosarcoma (11). The long non-coding RNA (lncRNA) THAP9-AS1 binds to and promotes methylation of the SOCS3 promoter region with DNA methyltransferases (DNMTs) and activates the JAK2/STAT3 signaling pathway to facilitate osteosarcoma growth and metastasis (12). In fact, inhibiting DNMT-1 sensitized osteosarcoma cells to cabozantinib and other targeted agents by repressing the Notch pathway and subsequently upregulating expression of miR-34a (13).
RNA modifications also play a role in osteosarcoma metastasis. The m6A demethylase FTO mediates mRNA demethylation, promoting the decay of KLF3 mRNA and decreasing its expression, consequently facilitating osteosarcoma proliferation and metastasis (14). Also, the destabilizing effects of FTO on DACT1 mRNA promotes Wnt signaling and consequently osteosarcoma metastasis (15). In addition, ALKBH5-mediated m6A methylation upregulates the expression of USP22 and RNF40, subsequently inhibiting the ubiquitination of histone H2A, promoting osteosarcoma growth and metastasis (16). Upregulation of TRIM7 due to the loss of m6A RNA modifications has also been reported to promote osteosarcoma metastasis and chemoresistance by inducing the ubiquitination of BRMS1 (17).
The prognostic role of epigenetic changes in osteosarcoma have also been extensively studied. Immune-related DNA methylation patterns can be used to predict survival and tumor microenvironment patterns (18). RNA methylation-related signatures of metabolic genes and lncRNAs have also been proposed to be useful tools in the estimation of patient survival and immune landscapes of osteosarcoma (19, 20).
Metabolic reprogramming
Metabolic reprograming is one of the key features of osteosarcoma, and its role in tumor progression, drug resistance, and metastasis is well established (21). Various metabolic gene signatures have been found to predict survival in osteosarcoma patients (19, 22–24). For example, comprehensive metabolic profiling of osteosarcoma based on UHPLC-HRMS unveiled a panel of two metabolites, 5-aminopentanamide and 13(S)-HpOTrE (FA 18:3 + 2O), which was found to be an accurate indicator of lung metastases (25).
Aerobic glycolysis, also known as the Warburg effect, supports biosynthesis and metabolic processes necessary for osteosarcoma growth and metastasis (26). Key enzymes involved in this process, such as PGC1α, PKM2, ALDOA, and LDHA, can directly influence tumor progression and metastasis. For instance, miR-23b-3p downregulates PGC1α and promotes a metabolic shift from oxidative phosphorylation to glycolysis, supporting osteosarcoma progression (27).
PKM2 is another key enzyme regulating glycolysis, which acts on its substrate phosphoenolpyruvate (PEP) to form pyruvate (28). IRF7 was found to downregulate PKM2 via transcriptional suppression, inhibiting aerobic glycolysis in osteosarcoma (29). The SLIT2/ROBO1 axis contributes to the Warburg effect by activating the SRC/ERK/c-MYC/PFKFB2 pathway in osteosarcoma (30). ROCK2 can promote glycolysis and osteosarcoma tumor growth by upregulating HKII via the pPI3K/AKT signaling pathway (31). Aldolase A (ALDOA) stimulation by the lncRNA KCNQ1OT1 sponging miR-34c-5p promotes aerobic glycolysis in osteosarcoma to support metastasis (32).
Lactate dehydrogenase A (LDHA) catalyzes the conversion of pyruvate to lactate. The upregulation of LDHA is involved in cancer cell growth and migration, the development of stem-cell like traits, and chemoresistance (33). KDM6B regulates H3K27me3 demethylation in the promoter region of LDHA, thereby promoting LDHA expression and aerobic glycolysis in osteosarcoma cells, and hence facilitating tumor metastasis (34).
The m6A-reading protein YTH N6-methyladenosine RNA-binding protein 3 (YTHDF3) contributes to osteosarcoma progression by promoting aerobic glycolysis through enhancement of PGK1 mRNA stability in an m6A-dependent manner (35).
IDH1 is an important TCA cycle enzyme that catalyzes the conversion of isocitrate to α-ketoglutarate. High levels of IDH1 have been detected in osteosarcoma and correlated with poor survival. Hsp90-AHA1 was found to upregulate IHD1 and promote growth and metastasis in osteosarcoma (36).
Besides glucose metabolism, changes in lipid and amino acid metabolism have also been reported to participate in osteosarcoma metastasis. Lipid profiles differ in metastatic osteosarcoma cell lines compared to non-metastatic cells. For example, diacylglycerols are overexpressed in metastatic osteosarcoma cells, and the blockage of its synthesis can in fact inhibit cell migration (37). Highly metastatic osteosarcoma cell lines require glutamine for proliferation, and conversely, glutaminase-1 (GLS-1) inhibition limits metastatic progression in osteosarcoma (38).
CD47 is a key factor mediating immune evasion of tumor cells from the innate immune system. Increased uptake of leucine and glutamine in osteosarcoma cells through upregulation of LAT2 activates mTORC1 and subsequent c-Myc-mediated transcription of CD47, enabling evasion of innate immune mechanisms and thereby promoting metastasis (39).
Dysregulated pathways
Dysregulated signaling pathways have also been reported to be involved in osteosarcoma metastasis (Figure 2).
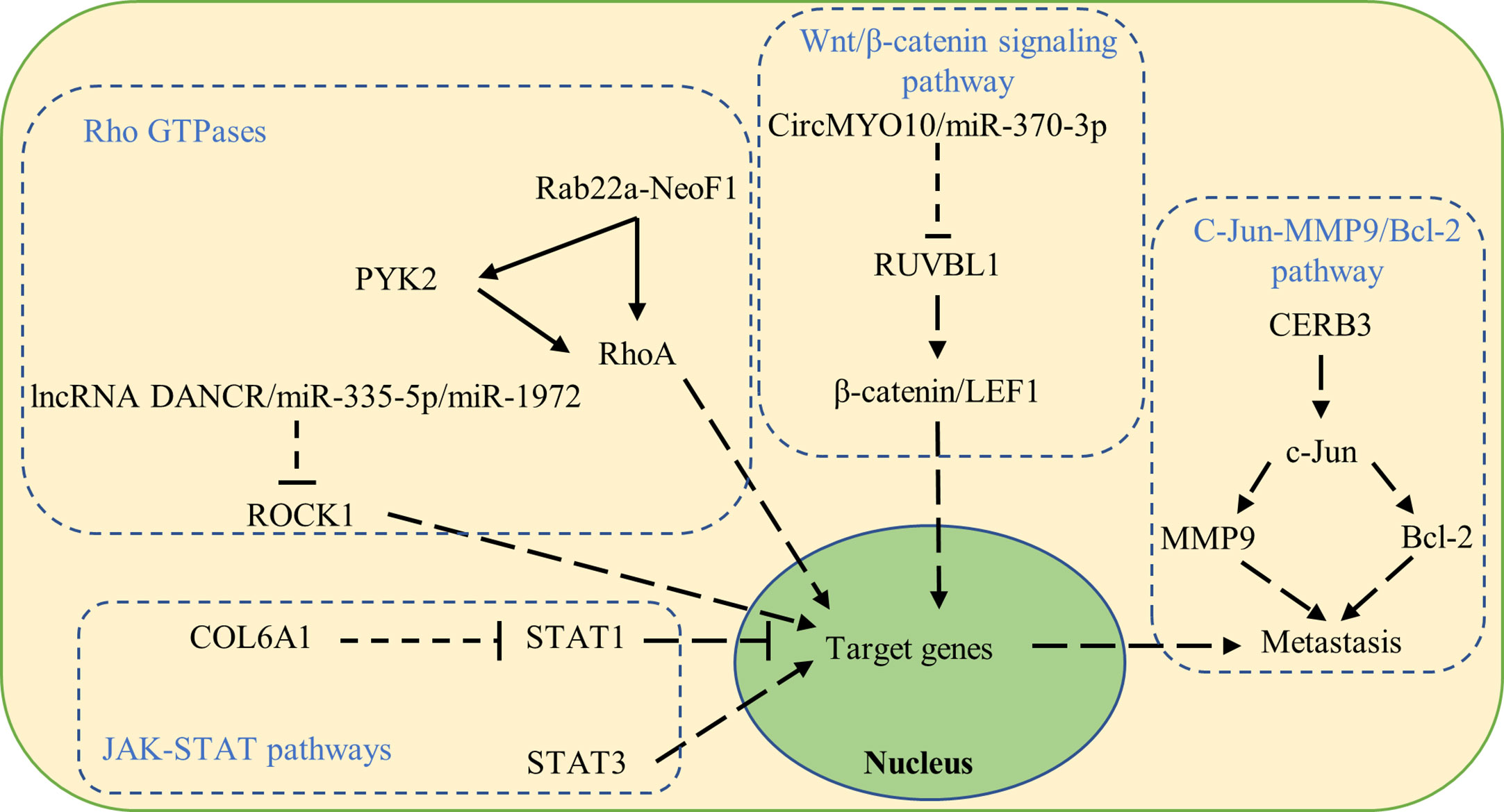
Figure 2 Signaling pathways that contribute to osteosarcoma metastasis when dysregulated.
Wnt/β-catenin signaling pathway
The Wnt/β-catenin signaling pathway is reported to play a crucial role in cell fate determination, proliferation, and migration in cancer. Cytoplasmic β-catenin undergoes ubiquitination and proteasomal degradation mediated by a destruction complex composed of Axin, APC, PP2A, GSK3, and CK1α. On the other hand, nuclear translocated β-catenin acts as a transcriptional coactivator for the TCF/LEF family of transcription factors promoting the expression of Wnt-target genes such as C-myc, RUNX2, and CyclinD1, which subsequently promotes the epithelial–mesenchymal transition and facilitates osteosarcoma metastasis (40, 41).
RUVBL1 can be regulated by CircMYO10/miR-370-3p in osteosarcoma and influences osteosarcoma progression. Molecularly, RUVBL1 enhances the transcriptional activity of the β-catenin/LEF1 complex by mediating chromatin remodeling at the promoter regions of LEF1 target genes, consequently promoting osteosarcoma metastasis (42).
C-Jun-MMP9/Bcl-2 pathway
As upstream signaling agents of MMPs, mitogen-activated protein kinase (MAPK) is a family of serine/threonine kinases that includes extracellular signal-regulated kinase (ERK)1/2, c-Jun N-terminal kinase (JNK) 1/2, and p38. Activation of MAPK is followed by the phosphorylation of various cytosolic substrates that participate in numerous cellular activities such as cell proliferation, differentiation, invasion, migration, and death (43, 44).
Activated by CERB3, c-Jun upregulates MMP9 and Bcl-2 to promote osteosarcoma proliferation and metastasis (45).
Rho GTPases
Rho GTPases belong to the Ras superfamily of GTPases, which are implicated in cell proliferation, cell cycle progression, and migration. Dysregulation of Rho GTPase functions is involved in osteosarcoma progression and metastasis (46).
RhoA activation in tumor cells leads to osteosarcoma metastasis to the lung (47, 48). The fusion protein Rab22a-NeoF1 either directly binds and activates RhoA, or is secreted together with its binding partner PKY2 by tumor-positive cells, taken up by tumor-negative cells, and facilitating RhoA activation via PYK2 (3, 49).
The Rho-associated coiled-coil containing protein kinase 1 (ROCK1) was reported to be a proliferation- and metastasis-related gene in various cancers including osteosarcoma (50). ROCK1 is regulated in osteosarcoma by lncRNA DANCR/miR-335-5p/miR-1972 (51).
JAK-STAT pathways
Signal transducer and activator of transcription (STAT) consists of seven members involved in the regulation of cell proliferation, differentiation, and survival. The activation of STAT1 in osteosarcoma cells suppressed EMT, resulting in increased apoptosis and cell cycle arrest, and decreased colony formation, cell migration, and invasion. Increased expression of COL6A1 promoted STAT1 degradation, which subsequently facilitated osteosarcoma metastasis (52). Furthermore, STAT3 is overexpressed in osteosarcoma and associated with poor survival. Activation of STAT3 upregulates the expression of target oncogenes and facilitates osteosarcoma metastasis (53).
Transcription factors
Dysregulation of transcription factors also contributes to osteosarcoma metastasis. NRF2 regulates intracellular ROS balance, the AMPK/mTOR autophagy signaling pathway, and the Warburg effect. TRIM22 inhibits osteosarcoma progression by binding to and destabilizing NRF2 in a KEAP1-independent manner (54).
RUNX proteins are DNA-binding transcription factors that regulate the expression of multiple genes involved in cellular differentiation and cell-cycle progression. RUNX2 is essential to osteoblast maturation and bone development, and can either suppress or promote carcinogenesis based on the clinical condition (55). Studies of osteosarcoma tumors have revealed that levels of RUNX2 DNA, RNA, and protein are significantly elevated in osteosarcoma tumors. Chromobox homolog4 (CBX4) is overexpressed in osteosarcoma cell lines and tissues, and promotes osteosarcoma metastasis by transcriptionally upregulating RUNX2 via the recruitment of GCN5 to the RUNX2 promoter (56).
Cyclic AMP-responsive element-binding protein 3 (CERB3), also known as LZIP or LUMAN, is a member of the leucine zipper transcription factor family. Its tumor-promoting role in osteosarcoma is regulated by circular RNA circTADA2A-miR-203a-3p. Molecularly, CREB3 can bind directly to the c-Jun promoter and regulate the transcriptional activity of c-Jun in osteosarcoma. MMP9 and Bcl-2 can be regulated by c-Jun and participate in CREB3-c-Jun modulated osteosarcoma progression (45).
The transcription activators YAP/YAZ regulate EMT through AXL in osteosarcoma and influences cell differentiation, cell fate, and metastasis (57).
Tumor microenvironment
The tumor microenvironment includes cellular components, extracellular matrix, vesicles, and secreted molecules that interact with each other to regulate tumor progression, immune evasion, drug resistance, and metastasis (58, 59).
The cellular components of the tumor microenvironment are mainly composed of infiltrating lymphocytes, macrophages, fibroblasts, and platelets. The composition and functions of these cells are dynamically regulated by local tumor cells and can be influenced by therapeutic agents. The recruitment and/or activation of certain cells in the microenvironment play pivotal roles in osteosarcoma metastasis (60).
The prognostic role of tumor-infiltrating lymphocytes in the osteosarcoma tumor microenvironment has been explored. The presence of tumor-infiltrating CD4+ or CD8+ cells was correlated with improved overall survival and progression-free survival in osteosarcoma patients (61).
In addition to tumor-infiltrating lymphocytes, the functional states of macrophages in the tumor microenvironment have also been associated with osteosarcoma progression and metastasis. M1-polarized macrophages are generally regarded as tumor-suppressing, while M2-type macrophages exhibit tumor-promoting roles in osteosarcoma. Molecularly, M2-type macrophages secrete cytokines such as IL-10, TGF-β, and VEGF to promote osteosarcoma EMT and metastasis (62). The M2-polarized macrophages are primarily induced by the activation of Stat3 secondary to stimulation by tumor cell secretions such as exosomes or vesicles. For instance, tumor-derived exosomes have been reported to induce M2 macrophage polarization via Tim-3 to promote osteosarcoma metastasis (62). PYK2 secretion by osteosarcoma cells recruits bone marrow-derived cells (BMDCs) and induces M2 macrophage polarization by activating Stat3 in macrophage cells (8). In the presence of chemotherapy, macrophages secrete IL-18 and enable the upregulation of LAT2 in adjacent osteosarcoma cells, which, in turn, promotes tumor evasion by upregulating CD47 (39).
Cancer-associated fibroblasts (CAFs) comprise a large proportion of cells in the tumor microenvironment. These cells can be identified by the presence of α-smooth muscle actin, fibroblast activation protein, and vimentin. Activated CAFs are thought to promote tumor cell growth, invasion, metastasis, drug resistance, and reprogramming (63). At a molecular level, CAFs build up and remodel the extracellular matrix, enabling tumor cells to invade through the TME. In addition, CAFs modulate cancer cell behavior through the secretion of growth factors, cytokines, and chemokines such as IL-1beta, IL-6, IL-8, TGF-β, and collagen (63).
CAFs can be activated and reprogrammed by various mechanisms, contributing to tumor metastasis. Increased levels of COL6A1 in tumor cells are packaged into exosomes and transported to activated CAFs, which, in turn, promote tumor invasion and metastasis by secreting TGF-β (52). CAFs in the lung can also be reprogrammed to support osteosarcoma metastasis under the influence of TGF-β1 found in osteosarcoma-derived extracellular vesicles (64).
Platelet aggregation and activation can be induced by tumor cells to support tumor metastasis in osteosarcoma. Osteosarcoma cells highly express PDPN, which binds with CLEC-2 on the surface of platelets, leading to platelet activation and subsequent tumor metastasis. At a molecular level, activated platelets secrete various growth factors and cytokines such as PDGF, TGF-β, and LPA, thereby inducing EMT and promoting cell migration and invasion in osteosarcoma. In addition, aggregated platelets form clusters with tumor cells, which are then trapped in the microvasculature of various organs such as the lung, triggering tumor metastasis (65).
The extracellular matrix (ECM) is extensively altered in osteosarcoma, beginning with the collagens and proteoglycans that make it up. Increased expression of several sarcomatous matrix proteins has been associated with poor response to chemotherapy and poor prognosis in clinical studies of osteosarcoma. NELL1 is a secreted osteoinductive protein, which has bone anabolic and anti-osteoclastic effects. NELL1 can promote osteosarcoma metastasis by regulating the expression of key matricellular proteins through the induction of FAK/Src signaling (66).
The procollagen C-proteinase enhancer protein (PCOLCE) is a secreted glycoprotein that enhances procollagen C-proteinase participation in ECM reconstruction. PCOLCE is upregulated by TWIST1 in osteosarcoma and promotes osteosarcoma metastasis to the lung (67).
The extracellular matrix glycoprotein tenascin-C is highly expressed in the tumor microenvironment and promotes the migration, invasion, and metastatic progression of osteosarcoma. Tenascin-C functions by binding with its receptor integrin α9β1, which abolishes actin stress fiber formation and inhibits YAP and its downstream target gene expression (68).
Extracellular vesicles (EVs) are secreted by both tumor cells and their adjacent non-tumor counterparts with diameters ranging from 30 to 150 nm (69). These vesicles are rich in biologically active components such as proteins, lipids, and nucleic acids, and play important roles in the exchange of biomolecules between different cell types (70). Many studies have correlated EVs with carcinogenesis, progression, and metastasis in osteosarcoma (71–73).
Results of the recent clinical trials of advanced or metastatic osteosarcoma
To date, there remains no established effective treatment for metastatic osteosarcoma. Multiple clinical trials have been conducted in recent years to investigate the viability of novel agents or treatment combinations. We compiled key findings from clinical trials in advanced or metastatic osteosarcoma within the last 7 years (summarized in Table 1).
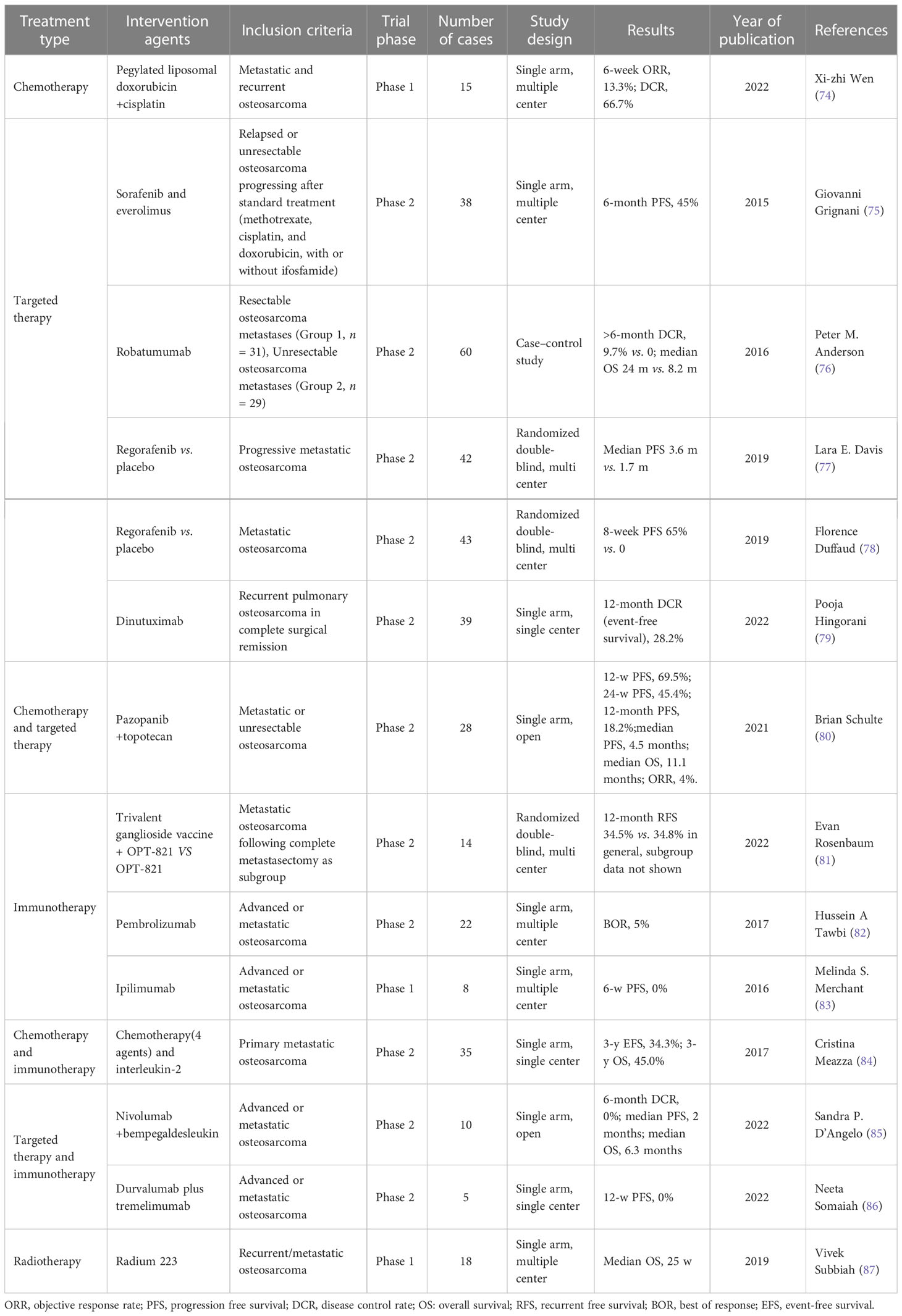
Table 1 Results of recent clinical trials involving patients with advanced or metastatic osteosarcoma.
Wen et al. reported a Phase 1 clinical trial investigating the efficacy of the combination therapy of pegylated liposomal doxorubicin and cisplatin in metastatic and recurrent osteosarcoma (74). In 15 cases, the 6-week objective response rate was 13.3% and the disease control rate was 66.7%. Other trials on targeted therapies such as regorafenib (77, 78), dinutuximab (79), robatumumab (76), sorafenib, and everolimus (75) demonstrated limited success with the overall 6-month progression-free survival rate of less than 50%.
The efficacy of combinatorial chemotherapy and targeted therapy treatments has also been tested in metastatic or unresectable osteosarcoma. A single-arm Phase 2 clinical trial involving 28 patients treated with pazopanib and topotecan failed to show any significant improvement in survival (6-month progression-free survival of 45.4%) (80).
Immunotherapy is an emerging treatment modality that has shown promising results in selected cases in melanoma and lung cancers. However, osteosarcoma patients did not seem to respond well to immune checkpoint inhibitors (81–83). The addition of interleukin-2 immunotherapy to a four-agent chemotherapy regimen for treating metastatic osteosarcoma did result in a 3-year event-free survival of 34.3% and 3-year overall survival of 45.0% (84), but a combination of targeted therapy and immunotherapy did not elicit better outcomes (85, 86).
Radiotherapy with radium 223 was also assessed in a clinical trial that involved 18 patients with recurrent or metastatic osteosarcoma (87). This Phase 1 single-arm multi-center trial reported a median overall survival of 25 weeks.
Ongoing clinical trials
There are currently several ongoing clinical trials involving metastatic osteosarcoma registered in ClinicalTrials.gov (Table 2). These include Phase 1, 2, and 3 trials. Interventions being investigated include chemotherapy, immunotherapy, radiotherapy, or targeted therapy alone; and combinatorial chemotherapy and immunotherapy, chemotherapy and targeted therapy, and targeted therapy and immunotherapy. Favorable outcomes from these trials have the potential to transform the landscape of clinical management of metastatic osteosarcoma.
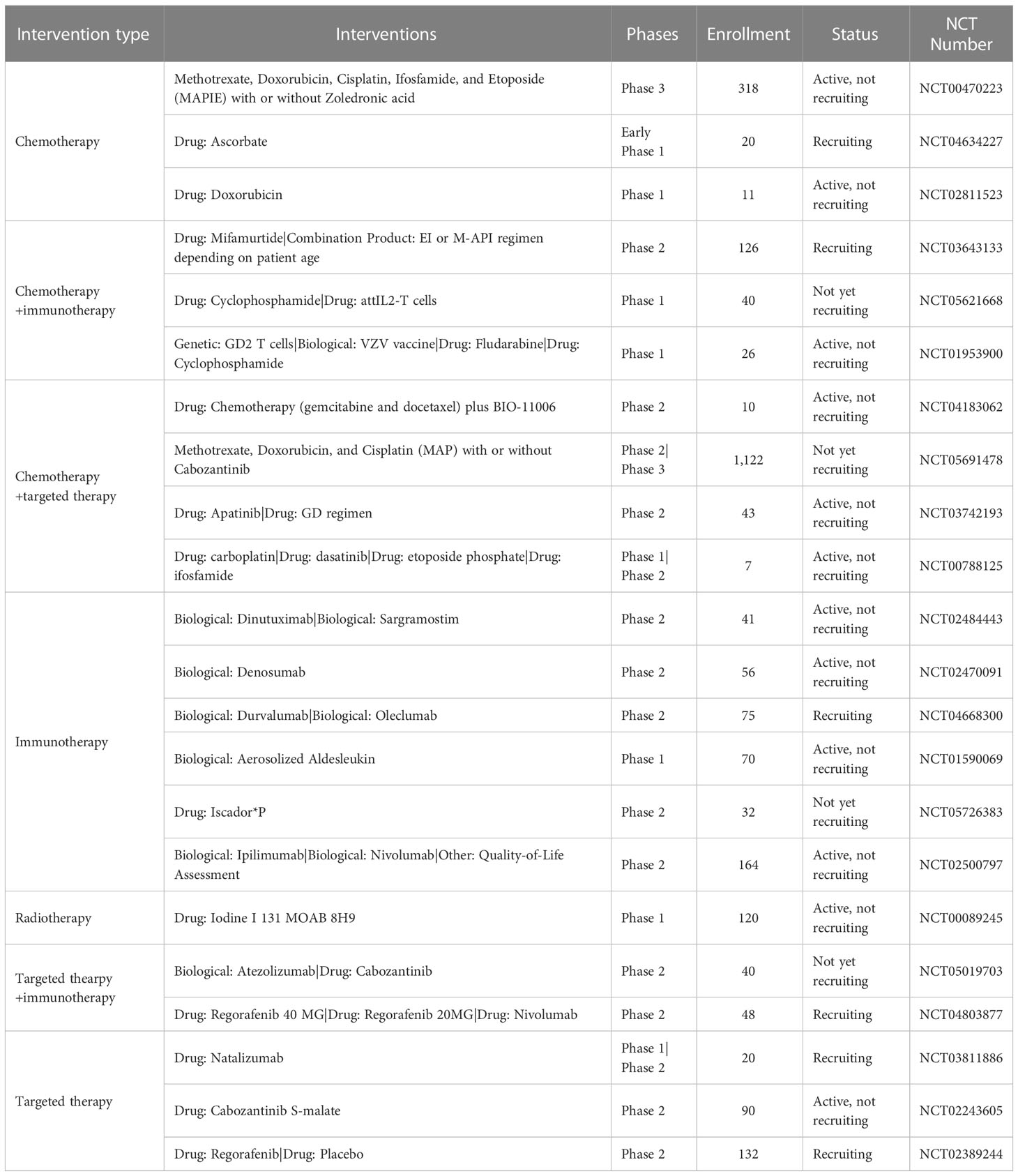
Table 2 Ongoing clinical trials involving patients with metastatic osteosarcoma.
Discussion
Osteosarcoma is the most common primary bone malignancy affecting children and young adults. More than 10% of patients are diagnosed with distant metastasis, and the 5-year overall survival of these patients is approximately 20%. However, current management options to prevent metastasis are limited and ineffective.
Emerging treatment options
Growing research on tumor cell alterations, behavior, and their surrounding microenvironment has informed the investigation of novel treatment options in preclinical settings. These include inhibitors targeting key metastasis-promoting proteins, approved drugs with newly discovered anti-metastatic roles, bioactive nanoparticles, and traditional Chinese medicine agents (Figure 3).
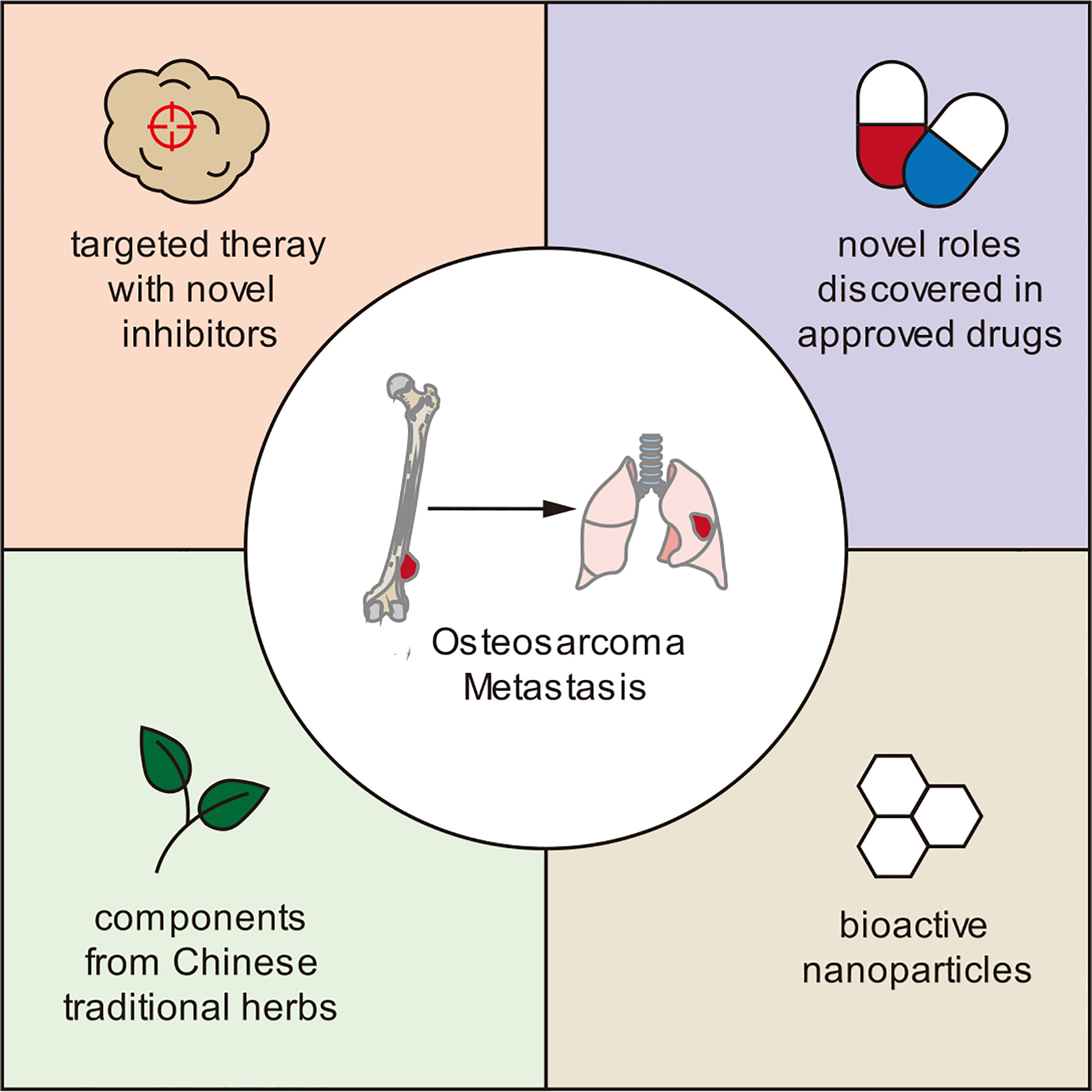
Figure 3 Novel therapies that have demonstrated anti-metastatic effects in osteosarcoma in preclinical studies.
Inhibitors targeting key metastasis-promoting proteins
Multiple key drivers of osteosarcoma metastasis have been reported, and inhibitors targeting these specific drivers have been developed and assessed. The covalent CDK7 inhibitor THZ2 demonstrated significant suppression of osteosarcoma tumor growth and metastasis by targeting super-enhancer-associated oncogenes (88). Tegavivint, a novel β-catenin/transducing β-like protein 1 (TBL1) inhibitor, exhibits anti-proliferative activity against osteosarcoma cells in vitro and in vivo through downregulation of the Wnt signaling pathway (89). CDK12 has been reported to facilitate genome stability through the regulation of DDR genes; accordingly, the CDK12 inhibitors THZ531 and E9 were found to disrupt osteosarcoma metastasis (90). BMTP-11 targets IL-11R α and inhibits osteosarcoma tumor growth and lung metastasis (91). A quinoline-based DNA methyltransferase inhibitor can induce cell cycle arrest and osteoblastic differentiation in osteosarcoma. It also showed synergistic effects with doxorubicin and cisplatin in treating osteosarcoma (92).
Approved drugs with newly discovered anti-metastatic roles
Drugs previously FDA-approved for other indications have been reported to inhibit osteosarcoma progression and metastasis. The FDA-approved DNA methylation inhibitor decitabine has demonstrated the ability to decrease proliferation, induce osteoblast differentiation, and reduce metastasis to visceral organs. Decitabine exposure in osteosarcoma reduces the protein expression of the metastasis-associated markers VIMENTIN, SLUG, ZEB1, and MMP9, with a concurrent decrease in mRNA expression of the known stem cell markers SOX2, OCT4, and NANOG. Normal osteoblasts express estrogen receptor α (ERα), whereas osteosarcoma cells do not due to promoter DNA methylation. Treatment of 143B osteosarcoma cells with decitabine led to ERα expression and decreased proliferation and induction of osteoblast differentiation (93).
Pramlintide, an FDA-approved drug for type 2 diabetes, was found to inhibit glycolysis and osteosarcoma tumor growth both in vitro and in vivo by inducing apoptosis (94). Melatonin attenuates chemokine CCL24 levels through inhibition of the JNK pathway to hinder human osteosarcoma cell invasion (95). All-trans retinoic acid prevents osteosarcoma metastasis by inhibiting M2 polarization of tumor-associated macrophages (96).
Bioactive nanoparticles
Bioactive nanoparticles (NPs), such as gold NPs, copper oxide NPs, iron oxide NPs, and zinc oxide nanoparticles (ZnO NPs), have been recently discovered to possess significant tumor-suppressing roles (97–99). ZnO NPs can inhibit osteosarcoma metastasis by degrading β-catenin in the HIF-1 α/BNIP3/LC3B-mediated mitophagy pathway (100).
Traditional Chinese medicine agents
The anti-tumor roles of traditional Chinese medicines and herbs have been explored in osteosarcoma. Ailanthone (AIL), a major component of the Chinese medicine Ailanthus altissima, can induce metabolic reprogramming in osteosarcoma, leading to growth inhibition both in vitro and in vivo. Molecularly, AIL induces cell cycle arrest and apoptosis in osteosarcoma cells by downregulating the serine biosynthetic pathway (101). Other natural compounds or herbs such as degalactotigonin (102) and shikonin (103) have also been reported to inhibit osteosarcoma growth and metastasis.
Navigating the challenges of osteosarcoma
Osteosarcoma research is particularly challenging (Figure 4). The low prevalence of osteosarcoma makes the conducting of rigorous clinical trials especially challenging. Heterogeneity within and between patient tumors also limits the generalizability of study findings. Thankfully, advancements in biotechnology and molecular techniques have paved the way for solutions to some of these challenges. For example, patient-derived xenograft models and organoid cultures have emerged as viable cancer models for experimentation, offering increased biomimicry, which should lead to stronger correlations with patient outcomes. Furthermore, detailed molecular characterization of osteosarcoma has allowed for the development of personalized therapies that target specific biomarkers and patient genomic profiles, increasing efficacy of potential treatments.
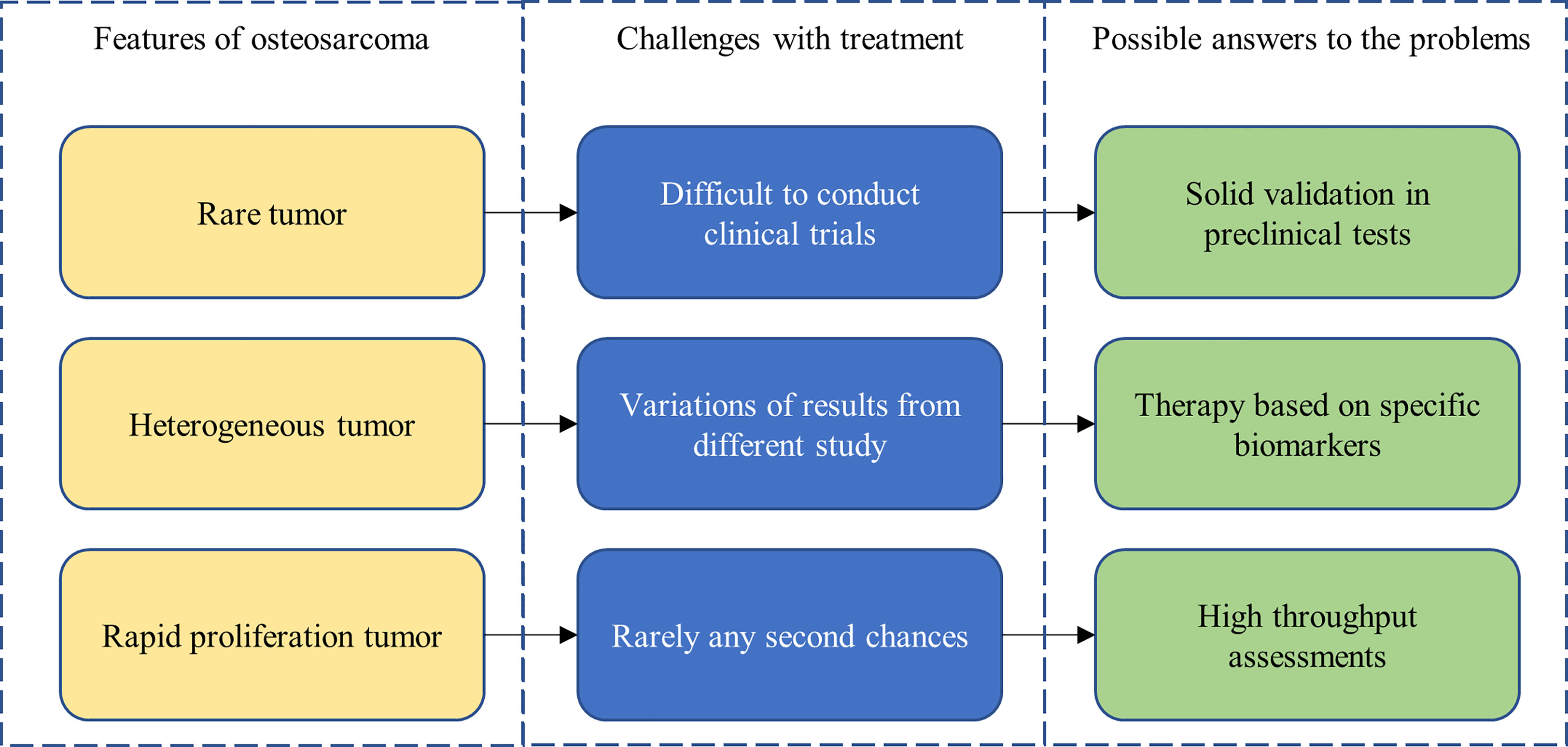
Figure 4 Features of, treatment challenges in, and potential solutions for osteosarcoma.
Conclusion
We reviewed the current literature on contributors to osteosarcoma metastasis, including genomic and epigenomic changes, metabolic reprogramming, transcription factors, dysregulation of physiologic pathways, and alterations to the tumor microenvironment. In addition, we discussed potential emerging therapies to suppress osteosarcoma metastasis. Further research on the molecular mechanisms of osteosarcoma metastasis, combined with growing molecular technologies, can inform the development of novel, personalized, and targeted therapies to ultimately improve outcomes in osteosarcoma patients.
Author contributions
Conceptualization, XD and HW. Writing—original draft preparation, XD. Writing—review and editing, BZ, LP and RZ. Visualization, HW. Supervision, WY. Funding acquisition, XD and HW. All authors contributed to the article and approved the submitted version.
Funding
This study was jointly supported by the grants of the National Natural Science Foundation of China (Grant No. 82102258), the Natural Science Foundation of Henan Province (Grant No. 212300410252), the Henan Provincial Medical Science and Technology Research Project (Grant Nos. LHGJ20190217 and LHGJ20220194), and the Key Technologies R & D Program of Henan Province (Grant No. 212102310617).
Acknowledgments
Special thanks to Sunny and Jessica for their support in preparing the manuscript.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Abbreviations
BMDCs, bone marrow-derived cells; CAF, cancer-associated fibroblast; DCR, disease control rate; DNMTs, DNA methyltransferases; ECM, extracellular matrix; EFS, event-free survival; ERK, extracellular signal-regulated kinase; EVs, extracellular vesicles; GLS-1, glutaminase-1; MAPK, mitogen-activated protein kinase; MMPs, matrix metalloproteinases; NPs, nanoparticles; ORR, objective response rate; OS, overall survival; PFS, progression-free survival; SCNAs, somatic copy number alterations.
References
1. Hu Z, Wen S, Huo Z, Wang Q, Zhao J, Wang Z, et al. Current status and prospects of targeted therapy for osteosarcoma. Cells Switzerland; (2022) 11. doi: 10.3390/cells11213507
2. Meltzer PS, Helman LJ. New horizons in the treatment of osteosarcoma. N Engl J Med United States; (2021) 385:2066–76. doi: 10.1056/NEJMra2103423
3. de Nonneville A, Salas S, Bertucci F, Sobinoff AP, Adélaïde J, Guille A, et al. TOP3A amplification and ATRX inactivation are mutually exclusive events in pediatric osteosarcomas using ALT. EMBO Mol Med England; (2022) 14:e15859. doi: 10.15252/emmm.202215859
4. Wang D, Niu X, Wang Z, Song C-L, Huang Z, Chen K-N, et al. Multiregion sequencing reveals the genetic heterogeneity and evolutionary history of osteosarcoma and matched pulmonary metastases. Cancer Res (2019) 79:7–20. doi: 10.1158/0008-5472.CAN-18-1086
5. Xiong S, Chachad D, Zhang Y, Gencel-Augusto J, Sirito M, Pant V, et al. Differential gain-of-Function activity of three p53 hotspot mutants in vivo. Cancer Res (2022) 82:1926–36. doi: 10.1158/0008-5472.CAN-21-3376
6. Tu J, Huo Z, Yu Y, Zhu D, Xu A, Huang M-F, et al. Hereditary retinoblastoma iPSC model reveals aberrant spliceosome function driving bone malignancies. Proc Natl Acad Sci U S A United States; (2022) 119:e2117857119. doi: 10.1073/pnas.2117857119
7. Liang X, Wang X, He Y, Wu Y, Zhong L, Liu W, et al. Acetylation dependent functions of Rab22a-NeoF1 fusion protein in osteosarcoma. Theranostics. (2020) 10:7747–57. doi: 10.7150/thno.46082
8. Zhong L, Liao D, Li J, Liu W, Wang J, Zeng C, et al. Rab22a-NeoF1 fusion protein promotes osteosarcoma lung metastasis through its secretion into exosomes. Signal Transduct Target Ther (2021) 6:59. doi: 10.1038/s41392-020-00414-1
9. Sayles LC, Breese MR, Koehne AL, Leung SG, Lee AG, Liu H-Y, et al. Genome-informed targeted therapy for osteosarcoma. Cancer Discovery (2019) 9:46–63. doi: 10.1158/2159-8290.CD-17-1152
10. Namløs HM, Skårn M, Ahmed D, Grad I, Andresen K, Kresse SH, et al. miR-486-5p expression is regulated by DNA methylation in osteosarcoma. BMC Genomics (2022) 23:142. doi: 10.1186/s12864-022-08346-6
11. Cheng S, Wang W. DNMT3A regulates miR-149 DNA methylation to activate NOTCH1/Hedgehog pathway to promote the development of junctional osteosarcoma. BioMed Res Int (2022) 2022:3261213. doi: 10.1155/2022/3261213
12. Yang S, Wang B, Liu C, Wang Q, Wang R, Su W, et al. THAP9-AS1 promotes tumorigenesis and reduces ROS generation through the JAK2/STAT3 signaling pathway by increasing SOCS3 promoter methylation in osteosarcoma. Oxid Med Cell Longev (2021) 2021:5620475. doi: 10.1155/2021/5620475
13. Wang J-H, Zeng Z, Sun J, Chen Y, Gao X. A novel small-molecule antagonist enhances the sensitivity of osteosarcoma to cabozantinib in vitro and in vivo by targeting DNMT-1 correlated with disease severity in human patients. Pharmacol Res Netherlands; (2021) 173:105869. doi: 10.1016/j.phrs.2021.105869
14. Shan H-J, Gu W-X, Duan G, Chen H-L. Fat mass and obesity associated (FTO)-mediated N6-methyladenosine modification of krüppel-like factor 3 (KLF3) promotes osteosarcoma progression. Bioengineered. (2022) 13:8038–50. doi: 10.1080/21655979.2022.2051785
15. Lv D, Ding S, Zhong L, Tu J, Li H, Yao H, et al. M(6)A demethylase FTO-mediated downregulation of DACT1 mRNA stability promotes wnt signaling to facilitate osteosarcoma progression. Oncogene England; (2022) 41:1727–41. doi: 10.1038/s41388-022-02214-z
16. Yadav P, Subbarayalu P, Medina D, Nirzhor S, Timilsina S, Rajamanickam S, et al. M6A RNA methylation regulates histone ubiquitination to support cancer growth and progression. Cancer Res (2022) 82:1872–89. doi: 10.1158/0008-5472.CAN-21-2106
17. Zhou C, Zhang Z, Zhu X, Qian G, Zhou Y, Sun Y, et al. N6-methyladenosine modification of the TRIM7 positively regulates tumorigenesis and chemoresistance in osteosarcoma through ubiquitination of BRMS1. EBioMedicine. (2020) 59:102955. doi: 10.1016/j.ebiom.2020.102955
18. Shi D, Mu S, Pu F, Liu J, Zhong B, Hu B, et al. Integrative analysis of immune-related multi-omics profiles identifies distinct prognosis and tumor microenvironment patterns in osteosarcoma. Mol Oncol (2022) 16:2174–94. doi: 10.1002/1878-0261.13160
19. Wang G, Wang H, Cheng S, Zhang X, Feng W, Zhang P, et al. N1-methyladenosine methylation-related metabolic genes signature and subtypes for predicting prognosis and immune microenvironment in osteosarcoma. Front Genet (2022) 13:993594. doi: 10.3389/fgene.2022.993594
20. Wu Z-Y, Shi Z-Y. The prognostic value and immune landscapes of m1A/m5C/m6A-associated lncRNA signature in osteosarcoma. Eur Rev Med Pharmacol Sci Italy; (2022) 26:5868–83.
21. He Q, Hao P, He G, Mai H, Liu W, Zhang W, et al. IGF2BP1-regulated expression of ERRα is involved in metabolic reprogramming of chemotherapy resistant osteosarcoma cells. J Transl Med England; (2022) 20:348. doi: 10.1186/s12967-022-03549-7
22. Li L, Li Z, He X, Wang Y, Lu M, Gong T, et al. A nutritional metabolism related prognostic scoring system for patients with newly diagnosed osteosarcoma. Front Nutr Switzerland; (2022) 9:883308. doi: 10.3389/fnut.2022.883308
23. Wan L, Zhang W, Liu Z, Yang Z, Tu C, Li Z. A novel glutamine metabolism-related gene signature in prognostic prediction of osteosarcoma. Int J Gen Med New Zealand; (2022) 15:997–1011. doi: 10.2147/IJGM.S352859
24. Li L-Q, Zhang L-H, Yuan Y-B, Lu X-C, Zhang Y, Liu Y-K, et al. Signature based on metabolic-related gene pairs can predict overall survival of osteosarcoma patients. Cancer Med United States; (2021) 10:4493–509. doi: 10.1002/cam4.3984
25. Lv D, Zou Y, Zeng Z, Yao H, Ding S, Bian Y, et al. Comprehensive metabolomic profiling of osteosarcoma based on UHPLC-HRMS. Metabolomics United States; (2020) 16:120. doi: 10.1007/s11306-020-01745-4
26. Feng Z, Ou Y, Hao L. The roles of glycolysis in osteosarcoma. Front Pharmacol Switzerland; (2022) 13:950886. doi: 10.3389/fphar.2022.950886
27. Zhu R, Li X, Ma Y. miR-23b-3p suppressing PGC1α promotes proliferation through reprogramming metabolism in osteosarcoma. Cell Death Dis England; (2019) 10:381. doi: 10.1038/s41419-019-1614-1
28. Zhang Z, Deng X, Liu Y, Liu Y, Sun L, Chen F. PKM2, function and expression and regulation. Cell Biosci England; (2019) 9:52. doi: 10.1186/s13578-019-0317-8
29. Li Z, Geng M, Ye X, Ji Y, Li Y, Zhang X, et al. IRF7 inhibits the warburg effect via transcriptional suppression of PKM2 in osteosarcoma. Int J Biol Sci Australia; (2022) 18:30–42. doi: 10.7150/ijbs.65255
30. Zhao S-J, Shen Y-F, Li Q, He Y-J, Zhang Y-K, Hu L-P, et al. SLIT2/ROBO1 axis contributes to the warburg effect in osteosarcoma through activation of SRC/ERK/c-MYC/PFKFB2 pathway. Cell Death Dis England; (2018) 9:390. doi: 10.1038/s41419-018-0419-y
31. Deng B, Deng J, Yi X, Zou Y, Li C. ROCK2 promotes osteosarcoma growth and glycolysis by up-regulating HKII via phospho-PI3K/AKT signalling. Cancer Manag Res New Zealand; (2021) 13:449–62. doi: 10.2147/CMAR.S279496
32. Shen Y, Xu J, Pan X, Zhang Y, Weng Y, Zhou D, et al. LncRNA KCNQ1OT1 sponges miR-34c-5p to promote osteosarcoma growth via ALDOA enhanced aerobic glycolysis. Cell Death Dis England; (2020) 11:278. doi: 10.1038/s41419-020-2485-1
33. Sharma D, Singh M, Rani R. Role of LDH in tumor glycolysis: regulation of LDHA by small molecules for cancer therapeutics. Semin Cancer Biol Engl (2022). doi: 10.1016/j.semcancer.2022.11.007
34. Jiang Y, Li F, Gao B, Ma M, Chen M, Wu Y, et al. KDM6B-mediated histone demethylation of LDHA promotes lung metastasis of osteosarcoma. Theranostics. (2021) 11:3868–81. doi: 10.7150/thno.53347
35. Liu D, Li Z, Zhang K, Lu D, Zhou D, Meng Y. N(6)-methyladenosine reader YTHDF3 contributes to the aerobic glycolysis of osteosarcoma through stabilizing PGK1 stability. J Cancer Res Clin Oncol Germany (2022). doi: 10.1007/s00432-022-04337-y
36. Zheng D, Liu W, Xie W, Huang G, Jiang Q, Yang Y, et al. AHA1 upregulates IDH1 and metabolic activity to promote growth and metastasis and predicts prognosis in osteosarcoma. Signal Transduct Target Ther (2021) 6:25. doi: 10.1038/s41392-020-00387-1
37. Roy J, Dibaeinia P, Fan TM, Sinha S, Das A. Global analysis of osteosarcoma lipidomes reveal altered lipid profiles in metastatic versus nonmetastatic cells. J Lipid Res United States; (2019) 60:375–87. doi: 10.1194/jlr.M088559
38. Ren L, Ruiz-Rodado V, Dowdy T, Huang S, Issaq SH, Beck J, et al. Glutaminase-1 (GLS1) inhibition limits metastatic progression in osteosarcoma. Cancer Metab (2020) 8:4. doi: 10.1186/s40170-020-0209-8
39. Wang Z, Li B, Li S, Lin W, Wang Z, Wang S, et al. Metabolic control of CD47 expression through LAT2-mediated amino acid uptake promotes tumor immune evasion. Nat Commun England; (2022) 13:6308. doi: 10.1038/s41467-022-34064-4
40. Vega OA, Lucero CMJ, Araya HF, Jerez S, Tapia JC, Antonelli M, et al. Wnt/β-catenin signaling activates expression of the bone-related transcription factor RUNX2 in select human osteosarcoma cell types. J Cell Biochem United States; (2017) 118:3662–74. doi: 10.1002/jcb.26011
41. Martin JW, Zielenska M, Stein GS, van Wijnen AJ, Squire JA. The role of RUNX2 in osteosarcoma oncogenesis. Sarcoma. (2011) 2011:282745. doi: 10.1155/2011/282745
42. Chen J, Liu G, Wu Y, Ma J, Wu H, Xie Z, et al. CircMYO10 promotes osteosarcoma progression by regulating miR-370-3p/RUVBL1 axis to enhance the transcriptional activity of β-catenin/LEF1 complex via effects on chromatin remodeling. Mol Cancer (2019) 18:150. doi: 10.1186/s12943-019-1076-1
43. Dana PM, Sadoughi F, Asemi Z, Yousefi B. Molecular signaling pathways as potential therapeutic targets in osteosarcoma. Curr Med Chem United Arab Emirates; (2022) 29:4436–44. doi: 10.2174/0929867329666220209110009
44. Su C, Cai X, Xu T, Wu Y, Wang L, Chen P, et al. LIMS2 is downregulated in osteosarcoma and inhibits cell growth and migration. J Oncol Egypt; (2022) 2022:4811260. doi: 10.1155/2022/4811260
45. Wu Y, Xie Z, Chen J, Chen J, Ni W, Ma Y, et al. Circular RNA circTADA2A promotes osteosarcoma progression and metastasis by sponging miR-203a-3p and regulating CREB3 expression. Mol Cancer (2019) 18:73. doi: 10.1186/s12943-019-1007-1
46. Wang J, Yuan L, Xu X, Zhang Z, Ma Y, Hong L, et al. Rho-GEF trio regulates osteosarcoma progression and osteogenic differentiation through Rac1 and RhoA. Cell Death Dis England; (2021) 12:1148. doi: 10.1038/s41419-021-04448-3
47. Du X, Ou Y, Zhang M, Li K, Huang W, Jiang D. The mevalonate pathway promotes the metastasis of osteosarcoma by regulating YAP1 activity via RhoA. Genes Dis China; (2022) 9:741–52. doi: 10.1016/j.gendis.2020.11.009
48. Zhan F, Deng Q, Chen Z, Xie C, Xiang S, Qiu S, et al. SAR1A regulates the RhoA/YAP and autophagy signaling pathways to influence osteosarcoma invasion and metastasis. Cancer Sci Engl (2022).
49. Xie K, Zhang X, Tao Y. Rab22a-NeoF1: a promising target for osteosarcoma patients with lung metastasis. Signal Transduct Target Ther England; (2020) 5:161. doi: 10.1038/s41392-020-00273-w
50. de Sousa GR, Vieira GM, das Chagas PF, Pezuk JA, Brassesco MS. Should we keep rocking? portraits from targeting rho kinases in cancer. Pharmacol Res Netherlands; (2020) 160:105093. doi: 10.1016/j.phrs.2020.105093
51. Wang Y, Zeng X, Wang N, Zhao W, Zhang X, Teng S, et al. Long noncoding RNA DANCR, working as a competitive endogenous RNA, promotes ROCK1-mediated proliferation and metastasis via decoying of miR-335-5p and miR-1972 in osteosarcoma. Mol Cancer (2018) 17:89. doi: 10.1186/s12943-018-0837-6
52. Zhang Y, Liu Z, Yang X, Lu W, Chen Y, Lin Y, et al. H3K27 acetylation activated-COL6A1 promotes osteosarcoma lung metastasis by repressing STAT1 and activating pulmonary cancer-associated fibroblasts. Theranostics. (2021) 11:1473–92. doi: 10.7150/thno.51245
53. Liu Y, Liao S, Bennett S, Tang H, Song D, Wood D, et al. STAT3 and its targeting inhibitors in osteosarcoma. Cell Prolif England; (2021) 54:e12974. doi: 10.1111/cpr.12974
54. Liu W, Zhao Y, Wang G, Feng S, Ge X, Ye W, et al. TRIM22 inhibits osteosarcoma progression through destabilizing NRF2 and thus activation of ROS/AMPK/mTOR/autophagy signaling. Redox Biol (2022) 53:102344. doi: 10.1016/j.redox.2022.102344
55. Lee YM. RUNX family in hypoxic microenvironment and angiogenesis in cancers. Cells Switzerland; (2022) 11. doi: 10.3390/cells11193098
56. Wang X, Qin G, Liang X, Wang W, Wang Z, Liao D, et al. Targeting the CK1α/CBX4 axis for metastasis in osteosarcoma. Nat Commun (2020) 11:1141. doi: 10.1038/s41467-020-14870-4
57. Lamhamedi-Cherradi S-E, Mohiuddin S, Mishra DK, Krishnan S, Velasco AR, Vetter AM, et al. Transcriptional activators YAP/TAZ and AXL orchestrate dedifferentiation, cell fate, and metastasis in human osteosarcoma. Cancer Gene Ther (2021) 28:1325–38. doi: 10.1038/s41417-020-00281-6
58. Kirchhammer N, Trefny MP, Auf der Maur P, Läubli H, Zippelius A. Combination cancer immunotherapies: emerging treatment strategies adapted to the tumor microenvironment. Sci Transl Med United States (2022) 14:eabo3605. doi: 10.1126/scitranslmed.abo3605
59. Xu M, Zhang T, Xia R, Wei Y, Wei X. Targeting the tumor stroma for cancer therapy. Mol Cancer England; (2022) 21:208. doi: 10.1186/s12943-022-01670-1
60. Zhu T, Han J, Yang L, Cai Z, Sun W, Hua Y, et al. Immune microenvironment in osteosarcoma: components, therapeutic strategies and clinical applications. Front Immunol Switzerland; (2022) 13:907550. doi: 10.3389/fimmu.2022.907550
61. Casanova JM, Almeida J-S, Reith JD, Sousa LM, Fonseca R, Freitas-Tavares P, et al. Tumor-infiltrating lymphocytes and cancer markers in osteosarcoma: influence on patient survival. Cancers (Basel) Switzerland; (2021) 13. doi: 10.3390/cancers13236075
62. Cheng Z, Wang L, Wu C, Huang L, Ruan Y, Xue W. Tumor-derived exosomes induced M2 macrophage polarization and promoted the metastasis of osteosarcoma cells through Tim-3. Arch Med Res United States; (2021) 52:200–10. doi: 10.1016/j.arcmed.2020.10.018
63. Wang J-W, Wu X-F, Gu X-J, Jiang X-H. Exosomal miR-1228 from cancer-associated fibroblasts promotes cell migration and invasion of osteosarcoma by directly targeting SCAI. Oncol Res United States; (2019) 27:979–86. doi: 10.3727/096504018X15336368805108
64. Mazumdar A, Urdinez J, Boro A, Migliavacca J, Arlt MJE, Muff R, et al. Osteosarcoma-derived extracellular vesicles induce lung fibroblast reprogramming. Int J Mol Sci Switzerland; (2020) 21. doi: 10.3390/ijms21155451
65. Takemoto A, Takagi S, Ukaji T, Gyobu N, Kakino M, Takami M, et al. Targeting podoplanin for the treatment of osteosarcoma. Clin Cancer Res United States; (2022) 28:2633–45. doi: 10.1158/1078-0432.CCR-21-4509
66. Qin Q, Gomez-Salazar M, Tower RJ, Chang L, Morris CD, McCarthy EF, et al. NELL1 regulates the matrisome to promote osteosarcoma progression. Cancer Res (2022) 82:2734–47. doi: 10.1158/0008-5472.CAN-22-0732
67. Wang S, Zhong L, Li Y, Xiao D, Zhang R, Liao D, et al. Up-regulation of PCOLCE by TWIST1 promotes metastasis in osteosarcoma. Theranostics. (2019) 9:4342–53. doi: 10.7150/thno.34090
68. Sun Z, Schwenzer A, Rupp T, Murdamoothoo D, Vegliante R, Lefebvre O, et al. Tenascin-c promotes tumor cell migration and metastasis through integrin α9β1-mediated YAP inhibition. Cancer Res (2018) 78:950–61. doi: 10.1158/0008-5472.CAN-17-1597
69. Lak NSM, van der Kooi EJ, Enciso-Martinez A, Lozano-Andrés E, Otto C, Wauben MHM, et al. Extracellular vesicles: a new source of biomarkers in pediatric solid tumors? a systematic review. Front Oncol Switzerland; (2022) p:887210. doi: 10.3389/fonc.2022.887210
70. Gao X, Gao B, Li S. Extracellular vesicles: a new diagnostic biomarker and targeted drug in osteosarcoma. Front Immunol Switzerland; (2022) 13:1002742. doi: 10.3389/fimmu.2022.1002742
71. Sun N, Tran BV, Peng Z, Wang J, Zhang C, Yang P, et al. Coupling lipid labeling and click chemistry enables isolation of extracellular vesicles for noninvasive detection of oncogenic gene alterations. Adv Sci (Weinheim Baden-Wurttemberg Ger Germany; (2022) 9:e2105853. doi: 10.1002/advs.202105853
72. Wu SC, Kim A, Gu Y, Martinez DI, Zocchi L, Chen CC, et al. UHRF1 overexpression promotes osteosarcoma metastasis through altered exosome production and AMPK/SEMA3E suppression. Oncogenesis United States; (2022) 11:51. doi: 10.1038/s41389-022-00430-6
73. Han Z, Peng X, Yang Y, Yi J, Zhao D, Bao Q, et al. Integrated microfluidic-SERS for exosome biomarker profiling and osteosarcoma diagnosis. Biosens Bioelectron England; (2022) 217:114709. doi: 10.1016/j.bios.2022.114709
74. Wen X-Z, Pan Q-Z, Xu B-S, Xiao W, Weng D-S, Zhao J-J, et al. Phase I study of pegylated liposomal doxorubicin and cisplatin in patients with advanced osteosarcoma. Cancer Chemother Pharmacol Germany; (2022) 89:209–15. doi: 10.1007/s00280-021-04371-6
75. Grignani G, Palmerini E, Ferraresi V, D’Ambrosio L, Bertulli R, Asaftei SD, et al. Sorafenib and everolimus for patients with unresectable high-grade osteosarcoma progressing after standard treatment: a non-randomised phase 2 clinical trial. Lancet Oncol England; (2015) 16:98–107. doi: 10.1016/S1470-2045(14)71136-2
76. Anderson PM, Bielack SS, Gorlick RG, Skubitz K, Daw NC, Herzog CE, et al. A phase II study of clinical activity of SCH 717454 (robatumumab) in patients with relapsed osteosarcoma and Ewing sarcoma. Pediatr Blood Cancer United States; (2016) 63:1761–70. doi: 10.1002/pbc.26087
77. Duffaud F, Mir O, Boudou-Rouquette P, Piperno-Neumann S, Penel N, Bompas E, et al. Efficacy and safety of regorafenib in adult patients with metastatic osteosarcoma: a non-comparative, randomised, double-blind, placebo-controlled, phase 2 study. Lancet Oncol England; (2019) 20:120–33. doi: 10.1016/S1470-2045(18)30742-3
78. Davis LE, Bolejack V, Ryan CW, Ganjoo KN, Loggers ET, Chawla S, et al. Randomized double-blind phase II study of regorafenib in patients with metastatic osteosarcoma. J Clin Oncol United States; (2019) 37:1424–31. doi: 10.1200/JCO.18.02374
79. Hingorani P, Krailo M, Buxton A, Hutson P, Sondel PM, Diccianni M, et al. Phase 2 study of anti-disialoganglioside antibody, dinutuximab, in combination with GM-CSF in patients with recurrent osteosarcoma: a report from the children’s oncology group. Eur J Cancer England; (2022) 172:264–75. doi: 10.1016/j.ejca.2022.05.035
80. Schulte B, Mohindra N, Milhem M, Attia S, Robinson S, Monga V, et al. Phase II study of pazopanib with oral topotecan in patients with metastatic and non-resectable soft tissue and bone sarcomas. Br J Cancer England; (2021) 125:528–33. doi: 10.1038/s41416-021-01448-0
81. Rosenbaum E, Chugh R, Ryan CW, Agulnik M, Milhem MM, George S, et al. A randomised phase II trial of a trivalent ganglioside vaccine targeting GM2, GD2 and GD3 combined with immunological adjuvant OPT-821 versus OPT-821 alone in metastatic sarcoma patients rendered disease-free by surgery. Eur J Cancer England; (2022) 176:155–63. doi: 10.1016/j.ejca.2022.09.003
82. Tawbi HA, Burgess M, Bolejack V, Van Tine BA, Schuetze SM, Hu J, et al. Pembrolizumab in advanced soft-tissue sarcoma and bone sarcoma (SARC028): a multicentre, two-cohort, single-arm, open-label, phase 2 trial. Lancet Oncol England; (2017) 18:1493–501. doi: 10.1016/S1470-2045(17)30624-1
83. Merchant MS, Wright M, Baird K, Wexler LH, Rodriguez-Galindo C, Bernstein D, et al. Phase I clinical trial of ipilimumab in pediatric patients with advanced solid tumors. Clin Cancer Res (2016) 22:1364–70. doi: 10.1158/1078-0432.CCR-15-0491
84. Meazza C, Cefalo G, Massimino M, Daolio P, Pastorino U, Scanagatta P, et al. Primary metastatic osteosarcoma: results of a prospective study in children given chemotherapy and interleukin-2. Med Oncol United States; (2017) 34:191. doi: 10.1007/s12032-017-1052-9
85. D’Angelo SP, Richards AL, Conley AP, Woo HJ, Dickson MA, Gounder M, et al. Pilot study of bempegaldesleukin in combination with nivolumab in patients with metastatic sarcoma. Nat Commun England; (2022) 13:3477. doi: 10.1038/s41467-022-30874-8
86. Somaiah N, Conley AP, Parra ER, Lin H, Amini B, Solis Soto L, et al. Durvalumab plus tremelimumab in advanced or metastatic soft tissue and bone sarcomas: a single-centre phase 2 trial. Lancet Oncol England; (2022) 23:1156–66. doi: 10.1016/S1470-2045(22)00392-8
87. Subbiah V, Anderson PM, Kairemo K, Hess K, Huh WW, Ravi V, et al. Alpha particle radium 223 dichloride in high-risk osteosarcoma: a phase I dose escalation trial. Clin Cancer Res United States; (2019) 25:3802–10. doi: 10.1158/1078-0432.CCR-18-3964
88. Zhang J, Liu W, Zou C, Zhao Z, Lai Y, Shi Z, et al. Targeting super-Enhancer-Associated oncogenes in osteosarcoma with THZ2, a covalent CDK7 inhibitor. Clin Cancer Res United States; (2020) 26:2681–92. doi: 10.1158/1078-0432.CCR-19-1418
89. Nomura M, Rainusso N, Lee Y-C, Dawson B, Coarfa C, Han R, et al. Tegavivint and the β-Catenin/ALDH axis in chemotherapy-resistant and metastatic osteosarcoma. J Natl Cancer Inst (2019) 111:1216–27. doi: 10.1093/jnci/djz026
90. Bayles I, Krajewska M, Pontius WD, Saiakhova A, Morrow JJ, Bartels C, et al. Ex vivo screen identifies CDK12 as a metastatic vulnerability in osteosarcoma. J Clin Invest (2019) 129:4377–92. doi: 10.1172/JCI127718
91. Lewis VO, Devarajan E, Cardó-Vila M, Thomas DG, Kleinerman ES, Marchiò S, et al. BMTP-11 is active in preclinical models of human osteosarcoma and a candidate targeted drug for clinical translation. Proc Natl Acad Sci U S A (2017) 114:8065–70. doi: 10.1073/pnas.1704173114
92. Manara MC, Valente S, Cristalli C, Nicoletti G, Landuzzi L, Zwergel C, et al. A quinoline-based DNA methyltransferase inhibitor as a possible adjuvant in osteosarcoma therapy. Mol Cancer Ther United States; (2018) 17:1881–92. doi: 10.1158/1535-7163.MCT-17-0818
93. Lillo Osuna MA, Garcia-Lopez J, El Ayachi I, Fatima I, Khalid AB, Kumpati J, et al. Activation of estrogen receptor alpha by decitabine inhibits osteosarcoma growth and metastasis. Cancer Res (2019) 79:1054–68. doi: 10.1158/0008-5472.CAN-18-1255
94. Yang Y, Peng Z, Flores ER, Kleinerman ES. Pramlintide: a novel therapeutic approach for osteosarcoma through metabolic reprogramming. Cancers (Basel) Switzerland; (2022) 14. doi: 10.3390/cancers14174310
95. Lu K-H, Su S-C, Lin C-W, Hsieh Y-H, Lin Y-C, Chien M-H, et al. Melatonin attenuates osteosarcoma cell invasion by suppression of c-c motif chemokine ligand 24 through inhibition of the c-jun n-terminal kinase pathway. J Pineal Res England; (2018) 65:e12507. doi: 10.1111/jpi.12507
96. Zhou Q, Xian M, Xiang S, Xiang D, Shao X, Wang J, et al. All-trans retinoic acid prevents osteosarcoma metastasis by inhibiting M2 polarization of tumor-associated macrophages. Cancer Immunol Res United States; (2017) 5:547–59. doi: 10.1158/2326-6066.CIR-16-0259
97. Wang J, Liu N, Su Q, Lv Y, Yang C, Zhan H. Green synthesis of gold nanoparticles and study of their inhibitory effect on bulk cancer cells and cancer stem cells in breast carcinoma. Nanomater (Basel Switzerland) Switzerland; (2022) 12. doi: 10.3390/nano12193324
98. Soto KM, Luzardo-Ocampo I, López-Romero JM, Mendoza S, Loarca-Piña G, Rivera-Muñoz EM, et al. Gold nanoparticles synthesized with common mullein (Verbascum thapsus) and castor bean (Ricinus communis) ethanolic extracts displayed antiproliferative effects and induced caspase 3 activity in human HT29 and SW480 cancer cells. Pharmaceutics Switzerland; (2022) 14. doi: 10.3390/pharmaceutics14102069
99. Alshawwa SZ, Mohammed EJ, Hashim N, Sharaf M, Selim S, Alhuthali HM, et al. In Situ Biosynthesis of reduced alpha hematite (α-Fe(2)O(3)) nanoparticles by stevia rebaudiana l. leaf extract: insights into antioxidant, antimicrobial, and anticancer properties. Antibiot (Basel Switzerland) Switzerland; (2022) 11.
100. He G, Nie J-J, Liu X, Ding Z, Luo P, Liu Y, et al. Zinc oxide nanoparticles inhibit osteosarcoma metastasis by downregulating β-catenin via HIF-1α/BNIP3/LC3B-mediated mitophagy pathway. Bioact Mater (2023) 19:690–702. doi: 10.1016/j.bioactmat.2022.05.006
101. Zhang Y, Gong R, Liu Y, Sun X, Liang J, Zhou Y, et al. Ailanthone inhibits proliferation, migration and invasion of osteosarcoma cells by downregulating the serine biosynthetic pathway. Front Oncol Switzerland; (2022) 12:842406. doi: 10.3389/fonc.2022.842406
102. Zhao Z, Jia Q, Wu M-S, Xie X, Wang Y, Song G, et al. Degalactotigonin, a natural compound from solanum nigrum l., inhibits growth and metastasis of osteosarcoma through GSK3β inactivation-mediated repression of the Hedgehog/Gli1 pathway. Clin Cancer Res United States; (2018) 24:130–44. doi: 10.1158/1078-0432.CCR-17-0692
Keywords: osteosarcoma, metastasis, reprogramming, tumor microenvironment, mechanism
Citation: Du X, Wei H, Zhang B, Wang B, Li Z, Pang LK, Zhao R and Yao W (2023) Molecular mechanisms of osteosarcoma metastasis and possible treatment opportunities. Front. Oncol. 13:1117867. doi: 10.3389/fonc.2023.1117867
Received: 07 December 2022; Accepted: 18 April 2023;
Published: 01 May 2023.
Edited by:
Vasiliki Gkretsi, European University Cyprus, CyprusReviewed by:
Ranjit Kumar Mehta, University of Michigan, United StatesDeng Zhouming, Wuhan University, China
Copyright © 2023 Du, Wei, Zhang, Wang, Li, Pang, Zhao and Yao. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Xinhui Du, emx5eWR1eGluaHVpMjc4NUB6enUuZWR1LmNu
†These authors have contributed equally to this work