 Rong Wang1,2†
Rong Wang1,2† Hong-Hu Zhu
Hong-Hu Zhu
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Oncol., 27 September 2022
Sec. Hematologic Malignancies
Volume 12 - 2022 | https://doi.org/10.3389/fonc.2022.972606
This article is part of the Research TopicReviews in Hematologic MalignanciesView all 10 articles
Acute myeloid leukemia (AML) is a heterogeneous disease characterized by malignant proliferation of myeloid hematopoietic stem/progenitor cells. NPM1 represents the most frequently mutated gene in AML and approximately 30% of AML cases carry NPM1 mutations. Mutated NPM1 result in the cytoplasmic localization of NPM1 (NPM1c). NPM1c interacts with other proteins to block myeloid differentiation, promote cell proliferation and impair DNA damage repair. NPM1 is a good prognostic marker, but some patients ultimately relapse or fail to respond to therapy. It is urgent for us to find optimal therapies for NPM1-mutated AML. Efficacy of multiple drugs is under investigation in NPM1-mutated AML, and several clinical trials have been registered. In this review, we summarize the present knowledge of therapy and focus on the possible therapeutic interventions for NPM1-mutated AML.
Nucleophosmin (NPM1) is the most common mutated gene in acute myeloid leukemia (AML). AML with NPM1 mutations accounts for approximately 30% of adult AML, which is characterized by the cytoplasmic localization of NPM1 (NPM1c) (1). NPM1-mutated AML was recognized as a distinct entity in the World Health Organization classification of myeloid neoplasms.
NPM1, shuttling between the nucleus and cytoplasm, is predominantly located in the nucleus (2, 3). NPM1 protein contains three structural domains including N terminal, central and C terminal domain. Nuclear export signals (NESs), located in N terminal domain, promote the translocation of NPM1 from the nucleus to the cytoplasm (3, 4). Nucleolar localization signals (NoLS), formed in highly conserved aromatic region of C terminal domain, is critical for the localization of NPM1 to the nucleus (5). The nuclear export of NPM1 is mediated by the interaction of two NESs and the nuclear exporter exportin 1 (XPO1) (3). NPM1 is a multifunctional protein involved in diverse cellular functions such as ribosome synthesis, genomic stability, cellular growth and stress response (6–9).
NPM1 mutations result in the generation of a new C-terminal NES and the loss of tryptophan residues 288 and 290, which endow mutated-NPM1 stronger nuclear export ability and ultimately lead to the cytoplasmic localization of NPM1 (10, 11). NPM1c mediates cytoplasmic dislocation of promyelocytic leukemia (PML) nuclear bodies (NB) (12). Researchers found that NPM1c interacts and delocalizes PU.1, FBW7γ and APE1, which block myeloid differentiation, promote cell proliferation and impair DNA damage repair, respectively (13–15).
NPM1-mutated AML is a kind of AML with favorable prognosis. The overall survival rate was about 40% and complete remission (CR) rate was about 80% (16). However, approximately 50% of patients will eventually relapse (17). The standard therapy of NPM1-mutated AML patients includes “3+7” induction chemotherapy and consolidation therapy. NPM1 often co-exists with fms-like receptor tyrosine kinase-3 internal tandem duplication (FLT3-ITD), which results in poor survival and high relapse rates. Allogeneic hematopoietic stem cell transplantation (allo-HSCT) and FLT3 inhibitors may be considered as important choices for these high-risk patients. It should be underscored that despite the favorable outcome of NPM1-mutated AML patients, disease-free survival (DFS) and overall survival (OS) of older NPM1-mutated patients remain disappointing and worse than those in younger NPM1-mutated patients (18). This may be partly due to treatment options, disease biology and age-related factors.
It has been about 15 years since NPM1-mutated AML was first discovered. However, there is no consensus over how to treat this type of AML, especially relapsed NPM1-mutated AML. Up to now, several studies targeting NPM1-mutated AML are undergoing, including inhibiting NPM1c functions, interfering with abnormal transport of NPM1c protein, promoting NPM1c degradation and immunotherapy such as monoclonal antibodies. Herein, we summarize available data (Table 1) and ongoing clinical trials (Table 2) and focus on the potential targeted therapy (Figure 1) of NPM1-mutated AML.

Table 1 Summary of venetoclax-based therapies in NPM1-mutated AML.
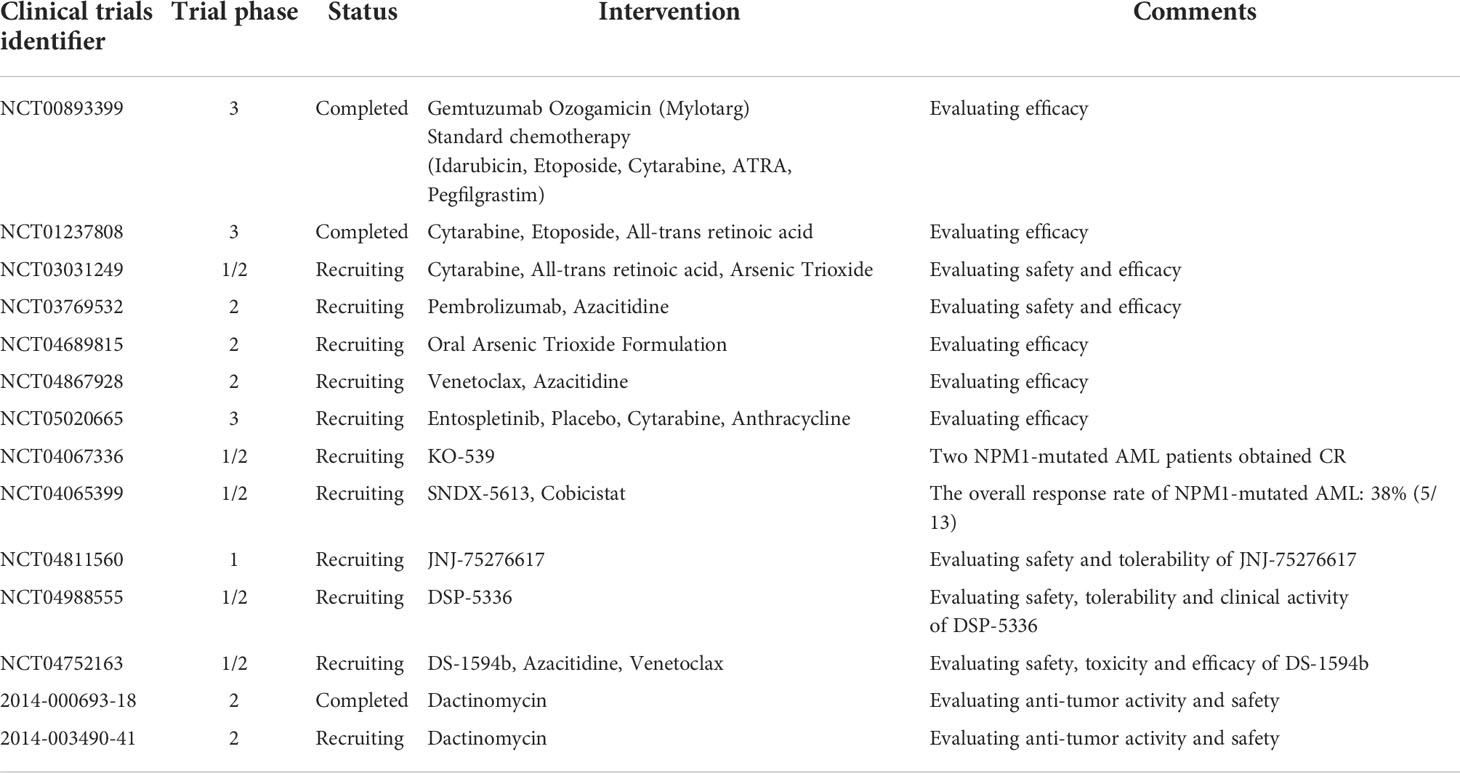
Table 2 Summary of ongoing or completed clinical trials in NPM1-mutated AML.
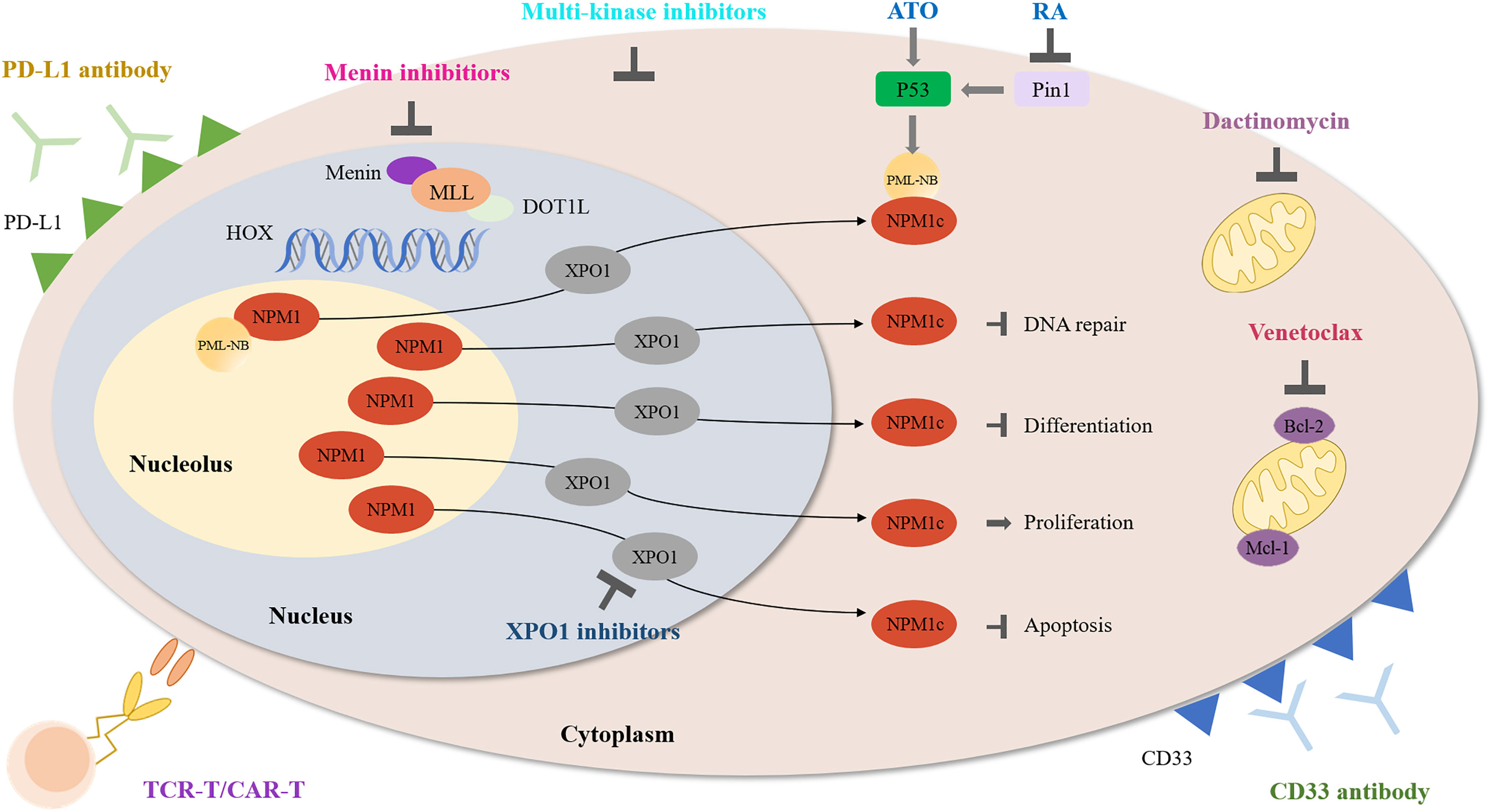
Figure 1 Schematic diagram describing the function of multiple agents in NPM1-mutated AML.
B-cell lymphoma 2 (Bcl-2), an anti-apoptotic protein, is overexpressed in AML. High Bcl-2 expression is associated with survival of AML cells and chemotherapy resistance (19, 20). Venetoclax is a potent and selective small molecule inhibitor of Bcl-2, has shown efficacy in preclinical and clinical practice (21, 22). Recently, safety and efficacy of venetoclax-based therapies, in combination with hypomethylating agents (HMAs) or low-dose cytarabine (LDAC), has been confirmed in several AML clinical studies. To figure out the effect of venetoclax-based therapies on NPM1-mutated AML, we summarize the data from recent prospective clinical studies, real-world reports and the latest ASH abstracts (Table 1).
The phase 1 clinical trial of venetoclax with decitabine or azacitidine (NCT02203773) enrolled 145 AML patients and NPM1-mutated AML accounts for 16% (N = 23) (23). CR and CR with incomplete count recovery (CRi) (CR + CRi) was observed in 21 NPM1-mutated patients. In the phase 3 clinical trial of venetoclax plus azacitidine, 66.7% of NPM1-mutated AML patients achieved CR + CRi (NCT02993523) (24). The phase 1/2, phase 3 clinical trials of venetoclax and LDAC were successively conducted in AML patients (NCT02287233, NCT03069352). Patients with NPM1 mutations represented 11% and 9% of the study cohort and experienced CR + CRi at 89% and 78%, respectively (25, 26). In the clinical trials of venetoclax in combination with intensive chemotherapy (IC), patients with NPM1 mutations also had good responses. NPM1-mutated AML patients attained CR and CRi at 80% and 100% in the venetoclax combined with 5 + 2 (cytarabine + idarubicin) study and venetoclax combined with FLAG + IDA (fludarabine, cytarabine, granulocyte colony-stimulating factor, and idarubicin) study, respectively (ACTRN12616000445471, NCT03214562) (27, 28).
In real-world settings, venetoclax combined with HMA also gained good outcomes in NPM1-mutated AML patients. CR and CRi were achieved at 100%, 71.4% and 66.7% in three reports, respectively (29–31). We also collect venetoclax-based regimens data from the latest ASH abstracts, CR and CRi rates were high in NPM1-mutated AML patients ranging from 76.6% to 100% (32–39).
Furthermore, venetoclax was recently identified as a selective agent for NPM1-mutated AML through clinical drugs screening (40). A retrospective analysis compared outcomes of NPM1-mutated AML patients treated with 3 approaches including venetoclax plus HMA, HMA and intensive chemotherapy (IC). This analysis demonstrated that venetoclax plus HMAs significantly reduced the risk of death and achieved a higher CR rate when compared with standard IC or HMAs (41). Venetoclax plus LDAC showed encouraging activity in eradicating persistent or relapsing mutated NPM1 measurable residual disease (MRD) (42). The multicenter and prospective clinical trials of venetoclax-based regimens are required to confirm its safety and efficacy in NPM1-mutated AML. A phase 2, multicenter trial evaluating the efficacy of venetoclax plus azacitidine in molecular relapse/progression has been registered (NCT04867928).
In addition to the most common combinations between venetoclax and HMA, LDAC or IC, there are several new combinations under investigation, such as the ones with arsenic trioxide (ATO). ATO, as an ancient drug, has exerted its function in several malignancies. Both ATO and venetoclax can downregulate Bcl-2 expression to induce apoptosis (43). Myeloid cell leukemia sequence 1 (MCL-1) is critical for the survival of AML cells and plays an essential role in venetoclax resistance (44, 45). ATO was reported to attenuate MCL-1 upregulation induce by venetoclax (46). The synergistic antileukemic activity of ATO and venetoclax was also confirmed in primary leukemia stem cells from AML patients (46). Therefore, this combination might represent an alternative option for NPM1-mutated AML. ATO and venetoclax synergistically induces the apoptosis of NPM1-mutated OCI-AML3 cells in vitro and showed anti-leukemia activity in two relapsed and/or refractory (R/R) NPM1-mutated AML patients (47).
The aforesaid results highlight the promising efficacy of venetoclax-based regimens, providing a rationale for further trials in NPM1-mutated AML. Current venetoclax-based regimens are mainly applied in elderly patients who are unfit for chemotherapy or young patients who are ineligible for standard induction therapy. More studies are required to expand the application of this approach, for example, to achieve greater overall survival in young patients. Furthermore, future researches should concentrate on optimizing the venetoclax-based therapies and overcoming venetoclax resistance.
NPM1-mutated AML cells are characterized by high expression of HOXA and HOXB clusters, which are necessary for the maintenance of the leukemic state (48). Histone modifiers MLL1 and DOT1L control HOX and FLT3 expression and differentiation in NPM1-mutated AML (49). Combinatorial inhibition of menin-MLL1 and DOT1L showed synergistic activity against primary AML cells in this study. Another preclinical result also indicated that inhibition of menin-MLL1 reversed leukemic development of NPM1-mutated AML mice models (50). It was reported that menin-MLL1 inhibition combined with venetoclax demonstrated anti-leukemia activity in primary NPM1-mutated AML samples (51). It seems that targeting menin could be a therapeutic strategy in NPM1-mutated AML.
FLT3-ITD often co-exists with mutated NPM1, accounting for approximately 40% of NPM1-mutated AML. Combining menin inhibitors with FLT3 inhibitors induced synergistic inhibition of proliferation and enhanced apoptosis in AML blasts (52). The combination of menin and FLT3 inhibitors significantly reduced leukemia burden and induced the long-term remissions in a PDX model with both NPM1 and FLT3-ITD mutations (53). Since XPO1 inhibition potently downregulate HOX expression in NPM1-mutated AML, the combination of menin and XPO1 inhibitors appeals as a rational therapeutic option in NPM1-mutated AML (48).
Several clinical studies are recruiting to assess the safety and efficacy of menin inhibitors such as SNDX-5613 and KO-539 on leukemia with MLL-rearrangement or NPM1 mutation (NCT04067336, NCT04065399, NCT04811560, NCT04752163, Table 2). Early results demonstrated tolerance and biologic activity of KO-539 (54). This phase 1/2A study evaluated clinical activity in 6 R/R AML patients and KO-539 induced CR in two patients. One patient achieved MRD-negative CR, who had AML with NPM1, DNMT3A, and KMT2D mutations and received KO-539 at 200 mg daily as the eighth line of treatment. SNDX-5613 exhibited safety and promising antileukemic activity in R/R MLL-rearrangement and NPM1-mutated AML in preliminary results (55). As of data cutoff on October 18, 2021, the overall response rate in 13 NPM1-mutated AML patients was 38%. The most common side effects included prolonged QTc, nausea, vomiting and differentiation syndrome.
Exportin 1 (XPO1) is a nuclear exporter implicated in the export of proteins and RNAs (56). NPM1 mutation results in the increased nuclear export ability of mutated NPM1 (10, 11). XPO1 inhibitors can relocate mutated NPM1 to the nucleus. However, XPO1 inhibitors are not NPM1-specific and also inhibit nuclear export of other proteins such as TP53 and P21.
Considering the relationship between XPO1 and NPM1, XPO1 inhibitors might be a promising approach for NPM1-mutated AML. The combination of selinexor and venetoclax showed a synergistic effect on the anti-leukemic activity of AML cells (57). Current studies mainly focus on the effects of XPO1 inhibitors in AML, not specifically in NPM1-mutated AML. To date, the combinations of selinexor and traditional chemotherapy, such as decitabine, cytarabine, mitoxantrone and idarubicin, are under study (58–60). However, systemic toxicities of selinexor, such as nausea and anorexia, limit its clinical usage to twice per week. Eltanexor, a second-generation XPO1 inhibitor, exhibits lower blood-brain penetration, improved tolerability and better anti-leukemic efficacy when compared with selinexor (61, 62). The combination of eltanexor and venetoclax reduce cell viability and induce apoptosis of AML cell lines (63). This combination therapy also enhanced anti-leukemia effect in AML cell-derived and patient-derived xenograft models. Eltanexor seems to be a prospective drug and further investigations are needed to validate the clinical activity in NPM1-mutated AML.
XPO1 is widely expressed in normal cells and interacts with hundreds of proteins, inhibition of XPO1 might generate some side-effects such as hematologic adverse events. Future efforts should focus on combining XPO1 inhibitors with either traditional chemotherapy or novel agents to enhance efficacy and safety.
Arsenic trioxide (ATO) plus all-trans retinoic acid (ATRA) had been proved a successful strategy in acute promyelocytic leukemia (APL), a unique AML subtype characterized by the fusion protein of promyelocytic leukemia (PML)–retinoic acid receptor ɑ (RARɑ). The combination has been proved to induce the degradation of PML-RARɑ fusion protein to cure APL (64).
NPM1-mutated AML cells are more sensitive to ATO because the presence of C-terminal cysteine 288 of NPM1c protein makes cells sensitize to oxidative stress induced by ATO (65). The anti-leukemia ability of ATO and ATRA support the further application in NPM1-mutated AML. Thus, two groups simultaneously demonstrated that the combination of ATO and ATRA induced the degradation of mutated NPM1 protein and apoptosis in both NPM1-mutated AML cell lines and primary cells (66, 67). Furthermore, ATO plus ATRA activated p53 signaling and restored nuclear organization of PML bodies. The combined treatment also significantly reduced bone marrow blasts in 3 NPM1-mutated AML patients and recovered the abnormal localization of both NPM1 and PML (67). ATRA was reported to induce mutated NPM1 degradation through the Pin1/PML/P53 axis, thereby promoting the response of blasts to chemotherapy or ATO (68). It is reported that ATRA improved survival of elderly NPM1-mutated AML patients without FLT3-ITD mutations when added to traditional chemotherapy (69). These findings provide convincing evidence for further clinical application of ATO and ATRA in NPM1-mutated AML. Relevant research are undergoing in NPM1-mutated AML (NCT03031249, NCT04689815, Table 1).
Dactinomycin, a famous antibiotic, exhibits potent antibacterial and anticancer activity by inhibiting topoisomerases and RNA polymerases (70). Investigators found that low dose dactinomycin can efficiently generate stress response in NPM1-mutated cells, illustrating NPM1-mutated AML might be sensitive to nucleolar stress (71). Dactinomycin targets mitochondria particularly primed by mutant NPM1, induces ROS production and restore PML NBs formation. Dactinomycin was initially shown its efficacy on a NPM1-mutated AML patient without FLT3-ITD mutations. The patient achieved morphologic and immunohistochemical CR after two cycles of therapy and showed a molecular CR after the fourth cycle (72). The clinical safety and efficacy of dactinomycin in AML patients with NPM1 mutations was further established (71, 73). Dual targeting of mitochondria with dactinomycin and venetoclax exerts strong anti-leukemic activity in NPM1-mutated AML (74). Dactinomycin seems to be a potential clinical choice for NPM1-mutated AML. Two clinical trials have been registered to evaluate anti-leukemic activity and safety of dactinomycin in NPM1-mutated AML (Table 1).
CD33 is a myeloid differentiation antigen expressed at the very early stages of myeloid cell development and is absent outside the hematopoietic system or on pluripotent hematopoietic stem cells (75). CD33 expression was found in leukemic blasts in almost all AML patients and associated with adverse disease features (76, 77). Gemtuzumab ozogamicin (GO) is CD33-directed immunoconjugate by delivering a DNA-damaging calicheamicin derivative to exert its function.
CD33 expression was significantly higher in the NPM1-mutated AML cases compared with the NPM1-unmutated cases (78). The results support the therapeutic application of CD33 antibodies in NPM1-mutated AML. A study showed that the addition of GO to standard chemotherapy improves the event-free survival (EFS) and OS in de novo AML patients aged 50–70 years (79). Among this cohort, NPM1-mutated AML patients accounted for 33% of all cases.
One clinical study was registered to evaluate the efficacy of GO in NPM1-mutated AML (NCT00893399). The study failed to show significant benefits on EFS when GO was added to intensive therapy, which might be due to a higher early mortality in the GO arm. However, patients who achieved CR + CRi after induction therapy significantly had fewer relapses in the GO arm than in the standard arm (80). In the following attempts, the combinations of GO and other treatments are required to be optimized in NPM1-mutated AML.
Programmed cell-death protein PD-1 and its ligands PD-L1 are immune checkpoint molecules that are involved in T-cell activation and dampen T-cell anti-tumor response. PD-1/PD-L1 pathway plays an essential role in tumor immune evasion, thus promoting the progression of tumor (81).
NPM1-mutated AML patients have a stronger cytotoxic T-lymphocyte response against mutated NPM1-derived peptides compared with healthy volunteers (82). Immune responses might be a contributing factor for the better prognosis of NPM1-mutated patients (83). High PD-L1 expression was detected in NPM1-mutated AML patients and predicted worse overall survival (84, 85). It should be noted that NPM1 was identified as a transcriptional regulator of PD-L1 and is associated with poor prognosis in triple-negative breast cancer (86). The aforementioned results indicated that PD-L1 might be a potential therapeutic target in NPM1-mutated AML. Unfortunately, current study suggested clinical activity of PD-L1 antibody in AML is limited (87, 88). Thus, more fundamental research and clinical studies are needed to investigate the exact role of PD-L1 in NPM1-mutated AML.
Hypomethylating agents, such as azactidine, have a dual effect against tumor immunity. In addition to enhancing anti-tumor immune response, HMAs can restrain immune response by upregulating PD-1 and PD-L1 expression, which can promote the exhaustion of tumor-specific T cells (89). It seems necessary to combine HMAs with immune checkpoint inhibitors such as PD-1 or PD-L1 antibody (90). Recently, a clinical study to evaluate the safety and efficacy of pembrolizumab when administered in combination with azacitidine in NPM1-mutated AML patients with molecular relapse was recruiting (NCT03769532, Table 1).
The adoptive immunotherapy, such as T cell receptor (TCR) and chimeric antigen receptor (CAR) T cell therapy, is an important milestone in the development of genetically modified cell therapies for leukemia. Due to low antigen expression in healthy tissues, TCR-T and CAR-T targeting tumor-associated antigens could be accompanied by severe toxicity. Neoantigens are derived from tumor-specific gene mutations but most neoantigens are encoded by patient-specific passenger mutations, which can be lost due to immunoediting and ultimately result in immune evasion (91). Nevertheless, neoantigens from driver gene mutations are unlikely to induce immune evasion because leukemic cells need to express the driver gene to maintain their malignant phenotype (92). Therefore, neoantigens derived from driver gene mutations are ideal targets for immunotherapy.
Mutated NPM1 is an essential driver gene and occurs in approximately 30% of AML. Besides the primary genetic lesion, NPM1 mutations also cooperate with other mutations to contribute to leukemogenesis (93). Moreover, NPM1-mutated protein does not exist in normal tissues, so it is an ideal leukemic-specific antigen and a potential target for NPM1-mutated AML. Recently, TCR-T and CAR-T directed against NPM1-mutated peptides obtained preliminary success in NPM1-mutated AML.
Van der Lee et al. transduced CD8+ and CD4+ T cells with the TCR for NPM1-mutated peptide, which demonstrated efficient specificity against NPM1-mutated and HLA-A2-restricted primary leukemic blasts (94). T cells transduced with TCR for NPM1-mutated protein could efficiently kill AML cells and prolonged OS of NSG mice engrafted with HLA-A*02:01-positive NPM1-mutated OCI-AML3 human cells. NPM1-mutated CAR-T cells showed efficient and specific anti-leukemia activity against NPM1c+HLA-A2+ leukemia cells and primary AML blasts (95). CAR-T cells could significantly reduce leukemia burden and prolonged survival of NSG mice engrafted with OCI-AML3 cells. Both TCR-T and CAR-T exhibit strong specificity and cytotoxicity against NPM1-muated AML without evident side effects. Further studies are warranted to investigate in clinical application and overcome potential drawbacks.
NPM1-mutated AML is a clinically heterogeneous group because it almost always exists in the context of other mutations. NPM1 mutations often co-occur with FLT3, DNMT3A or other mutations to contribute to leukemogenesis (96, 97). The latest report classified NPM1-mutated AML into two novel subtypes, primitive and committed subtype, based on a stem cell signature through RNA-seq (98). Interestingly, they found that leukemic cells in the primitive subtype are more sensitive to certain kinase inhibitors. The addition of kinase inhibitors to the treatment might achieve therapeutic benefits in this specific subtype of NPM1-mutated AML. These results may prompt us to make a more accurate risk stratification of NPM1-mutated AML based on multidisciplinary technology, thereby giving a guidance for clinical treatment. Furthermore, some controversial issues in diagnosis and treatments of NPM1-mutated AML still exists. Falini et al. recently summarized how he diagnose and treat NPM1-mutated AML and he constructively proposed that NPM1 mutational status, the timing of HSCT, MRD monitoring and ELN genetic-based risk stratification should be considered during the therapy (99).
NPM1 mutations are ideal targets for MRD monitoring because they are AML-specific, frequent, stable at relapse and do not drive clonal hematopoiesis of indeterminate potential. Investigators found that MRD, as determined by real-time quantitative PCR (RT-qPCR) of NPM1-mutated transcripts, provides important prognostic information for AML (100). Patients with persistence of NPM1-mutated transcripts in blood after the second cycle of chemotherapy was associated with a greater risk of relapse and a lower rate of survival compared with those without such transcripts. In multivariate analysis, the presence of MRD was the only significant prognostic factor for relapse and death. RT-qPCR remains the standard method for MRD monitoring in NPM1-mutated AML, the application of highly sensitive digital droplet PCR and NGS will be expanded in the future.
Considering the above findings, the combination of multiple agents is the dominant trend in the future treatment of NPM1-mutated AML, such as venetoclax-based regimens and XPO1 inhibitors combinations. The pathogenesis of NPM1-mutated AML and diverse drugs combinations need to be further studied. Joint efforts should be made to overcome the limitation of currently promising drugs, such as resistance for venetoclax and toxicity for XPO1 inhibitors. Novel targeted drugs for NPM1-mutated AML are also urgently developed. We are looking forward to acquiring the consensus on treatment of NPM1-mutated AML.
H-HZ and S-ZZ conceived the idea of the paper. RW, PX, and L-LC contributed equally to data collection and wrote the manuscript. All authors contributed to the article and approved the submitted version.
Leading Innovative and Entrepreneur Team Introduction Programme of Zhejiang (2020R01006 and 2019R01001) and the “Pioneer” R&D program of Zhejiang (2022C03005).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
1. Falini B, Mecucci C, Tiacci E, Alcalay M, Rosati R, Pasqualucci L, et al. Cytoplasmic nucleophosmin in acute myelogenous leukemia with a normal karyotype. N Engl J Med (2005) 352(3):254–66. doi: 10.1056/NEJMoa041974
2. Borer RA, Lehner CF, Eppenberger HM, Nigg EA. Major nucleolar proteins shuttle between nucleus and cytoplasm. Cell (1989) 56(3):379–90. doi: 10.1016/0092-8674(89)90241-9
3. Wang W, Budhu A, Forgues M, Wang XW. Temporal and spatial control of nucleophosmin by the ran-Crm1 complex in centrosome duplication. Nat Cell Biol (2005) 7(8):823–30. doi: 10.1038/ncb1282
4. Yu Y, Maggi LB Jr., Brady SN, Apicelli AJ, Dai MS, Lu H, et al. Nucleophosmin is essential for ribosomal protein L5 nuclear export. Mol Cell Biol (2006) 26(10):3798–809. doi: 10.1128/mcb.26.10.3798-3809.2006
5. Nishimura Y, Ohkubo T, Furuichi Y, Umekawa H. Tryptophans 286 and 288 in the c-terminal region of protein B23.1 are important for its nucleolar localization. Biosci Biotechnol Biochem (2002) 66(10):2239–42. doi: 10.1271/bbb.66.2239
6. Hingorani K, Szebeni A, Olson MO. Mapping the functional domains of nucleolar protein B23. J Biol Chem (2000) 275(32):24451–7. doi: 10.1074/jbc.M003278200
7. Okuda M, Horn HF, Tarapore P, Tokuyama Y, Smulian AG, Chan PK, et al. Nucleophosmin/B23 is a target of Cdk2/Cyclin e in centrosome duplication. Cell (2000) 103(1):127–40. doi: 10.1016/s0092-8674(00)00093-3
8. Bertwistle D, Sugimoto M, Sherr CJ. Physical and functional interactions of the arf tumor suppressor protein with Nucleophosmin/B23. Mol Cell Biol (2004) 24(3):985–96. doi: 10.1128/mcb.24.3.985-996.2004
9. Colombo E, Marine JC, Danovi D, Falini B, Pelicci PG. Nucleophosmin regulates the stability and transcriptional activity of P53. Nat Cell Biol (2002) 4(7):529–33. doi: 10.1038/ncb814
10. Falini B, Bolli N, Shan J, Martelli MP, Liso A, Pucciarini A, et al. Both carboxy-terminus nes motif and mutated Tryptophan(S) are crucial for aberrant nuclear export of nucleophosmin leukemic mutants in npmc+ aml. Blood (2006) 107(11):4514–23. doi: 10.1182/blood-2005-11-4745
11. Bolli N, Nicoletti I, De Marco MF, Bigerna B, Pucciarini A, Mannucci R, et al. Born to be exported: Cooh-terminal nuclear export signals of different strength ensure cytoplasmic accumulation of nucleophosmin leukemic mutants. Cancer Res (2007) 67(13):6230–7. doi: 10.1158/0008-5472.Can-07-0273
12. Zou Q, Tan S, Yang Z, Zhan Q, Jin H, Xian J, et al. Npm1 mutant mediated pml delocalization and stabilization enhances autophagy and cell survival in leukemic cells. Theranostics (2017) 7(8):2289–304. doi: 10.7150/thno.19439
13. Gu X, Ebrahem Q, Mahfouz RZ, Hasipek M, Enane F, Radivoyevitch T, et al. Leukemogenic nucleophosmin mutation disrupts the transcription factor hub that regulates granulomonocytic fates. J Clin Invest (2018) 128(10):4260–79. doi: 10.1172/jci97117
14. Bonetti P, Davoli T, Sironi C, Amati B, Pelicci PG, Colombo E. Nucleophosmin and its aml-associated mutant regulate c-myc turnover through Fbw7 gamma. J Cell Biol (2008) 182(1):19–26. doi: 10.1083/jcb.200711040
15. Vascotto C, Lirussi L, Poletto M, Tiribelli M, Damiani D, Fabbro D, et al. Functional regulation of the Apurinic/Apyrimidinic endonuclease 1 by nucleophosmin: Impact on tumor biology. Oncogene (2014) 33(22):2876–87. doi: 10.1038/onc.2013.251
16. Becker H, Marcucci G, Maharry K, Radmacher MD, Mrózek K, Margeson D, et al. Favorable prognostic impact of Npm1 mutations in older patients with cytogenetically normal De novo acute myeloid leukemia and associated gene- and microrna-expression signatures: A cancer and leukemia group b study. J Clin Oncol Off J Am Soc Clin Oncol (2010) 28(4):596–604. doi: 10.1200/jco.2009.25.1496
17. Hubmann M, Köhnke T, Hoster E, Schneider S, Dufour A, Zellmeier E, et al. Molecular response assessment by quantitative real-time polymerase chain reaction after induction therapy in Npm1-mutated patients identifies those at high risk of relapse. Haematologica (2014) 99(8):1317–25. doi: 10.3324/haematol.2014.104133
18. Gale RE, Green C, Allen C, Mead AJ, Burnett AK, Hills RK, et al. The impact of Flt3 internal tandem duplication mutant level, number, size, and interaction with Npm1 mutations in a Large cohort of young adult patients with acute myeloid leukemia. Blood (2008) 111(5):2776–84. doi: 10.1182/blood-2007-08-109090
19. Vaux DL, Cory S, Adams JM. Bcl-2 gene promotes haemopoietic cell survival and cooperates with c-myc to immortalize pre-b cells. Nature (1988) 335(6189):440–2. doi: 10.1038/335440a0
20. Campos L, Rouault JP, Sabido O, Oriol P, Roubi N, Vasselon C, et al. High expression of bcl-2 protein in acute myeloid leukemia cells is associated with poor response to chemotherapy. Blood (1993) 81(11):3091–6. doi: 10.1182/blood.V81.11.3091.3091
21. Pan R, Hogdal LJ, Benito JM, Bucci D, Han L, Borthakur G, et al. Selective bcl-2 inhibition by abt-199 causes on-target cell death in acute myeloid leukemia. Cancer Discovery (2014) 4(3):362–75. doi: 10.1158/2159-8290.Cd-13-0609
22. Konopleva M, Pollyea DA, Potluri J, Chyla B, Hogdal L, Busman T, et al. Efficacy and biological correlates of response in a phase ii study of venetoclax monotherapy in patients with acute myelogenous leukemia. Cancer Discovery (2016) 6(10):1106–17. doi: 10.1158/2159-8290.Cd-16-0313
23. DiNardo CD, Pratz K, Pullarkat V, Jonas BA, Arellano M, Becker PS, et al. Venetoclax combined with decitabine or azacitidine in treatment-naive, elderly patients with acute myeloid leukemia. Blood (2019) 133(1):7–17. doi: 10.1182/blood-2018-08-868752
24. DiNardo CD, Jonas BA, Pullarkat V, Thirman MJ, Garcia JS, Wei AH, et al. Azacitidine and venetoclax in previously untreated acute myeloid leukemia. N Engl J Med (2020) 383(7):617–29. doi: 10.1056/NEJMoa2012971
25. Wei AH, Strickland SA Jr., Hou JZ, Fiedler W, Lin TL, Walter RB, et al. Venetoclax combined with low-dose cytarabine for previously untreated patients with acute myeloid leukemia: Results from a phase Ib/Ii study. J Clin Oncol (2019) 37(15):1277–84. doi: 10.1200/jco.18.01600
26. Wei AH, Montesinos P, Ivanov V, DiNardo CD, Novak J, Laribi K, et al. Venetoclax plus ldac for newly diagnosed aml ineligible for intensive chemotherapy: A phase 3 randomized placebo-controlled trial. Blood (2020) 135(24):2137–45. doi: 10.1182/blood.2020004856
27. Chua CC, Roberts AW, Reynolds J, Fong CY, Ting SB, Salmon JM, et al. Chemotherapy and venetoclax in elderly acute myeloid leukemia trial (Caveat): A phase ib dose-escalation study of venetoclax combined with modified intensive chemotherapy. J Clin Oncol (2020) 38(30):3506–17. doi: 10.1200/jco.20.00572
28. DiNardo CD, Lachowiez CA, Takahashi K, Loghavi S, Xiao L, Kadia T, et al. Venetoclax combined with flag-Ida induction and consolidation in newly diagnosed and relapsed or refractory acute myeloid leukemia. J Clin Oncol (2021) 39(25):2768–78. doi: 10.1200/jco.20.03736
29. Winters AC, Gutman JA, Purev E, Nakic M, Tobin J, Chase S, et al. Real-world experience of venetoclax with azacitidine for untreated patients with acute myeloid leukemia. Blood Adv (2019) 3(20):2911–9. doi: 10.1182/bloodadvances.2019000243
30. Tong J, Zhao N, Hu X, Yao W, Cheng Y, Zhou L, et al. Efficacy of venetoclax combined with decitabine-based treatment for heavily pre-treated relapsed or refractory aml patients in a real-world setting. Cancer Manag Res (2021) 13:5613–21. doi: 10.2147/cmar.S316561
31. Tenold ME, Moskoff BN, Benjamin DJ, Hoeg RT, Rosenberg AS, Abedi M, et al. Outcomes of adults with Relapsed/Refractory acute myeloid leukemia treated with venetoclax plus hypomethylating agents at a comprehensive cancer center. Front Oncol (2021) 11:649209. doi: 10.3389/fonc.2021.649209
32. Kadia TM, Cortes JE, Konopleva MY, Borthakur GM, Pemmaraju N, Daver NG, et al. Venetoclax combined with cladribine + low dose arac (Ldac) alternating with 5-azacytidine produces high rates of minimal residual disease (Mrd) negative complete remissions (Cr) in older patients with newly diagnosed acute myeloid leukemia (Aml). Blood (2019) 134(Supplement_1):2647. doi: 10.1182/blood-2019-122477%JBlood
33. Law LY, Stevenson R, Ho G, Nair B, Laura A, Sayegh A. Venetoclax-based regimen for treatment of patients with acute myeloid leukemia in community based practices. Blood (2019) 134(Supplement_1):5081. doi: 10.1182/blood-2019-121816%JBlood
34. Madanat YF, Patel H, Patel O, Yan J, Khatib J, Vusirkala M, et al. Factors associated with clinical outcomes after venetoclax-based combination therapy in acute myeloid leukemia and high-grade myeloid neoplasms. Blood (2020) 136(Supplement 1):43. doi: 10.1182/blood-2020-139794%JBlood
35. Arslan S, Zhang J, Dhakal P, Moran JA, Naidoo N, Lombardi Story J, et al. Outcomes of therapy with venetoclax combined with hypomethylating agents in favorable-risk acute myeloid leukemia (Aml). Blood (2020) 136(Supplement 1):41–2. doi: 10.1182/blood-2020-142780%JBlood
36. Hammond D, Loghavi S, Konopleva M, Kadia TM, Daver N, Ohanian M, et al. Response patterns and mrd by flow cytometry and ngs in patients with mutant-idh acute myeloid leukemia treated with venetoclax in combination with hypomethylating agents. Blood (2020) 136(Supplement 1):35–6. doi: 10.1182/blood-2020-141380%JBlood
37. Venugopal S, Issa GC, Konopleva M, DiNardo CD, Borthakur G, Jabbour EJ, et al. Npm1 mutations do not retain a favorable prognostic impact in adults with advanced relapsed or refractory (R/R) acute myeloid leukemia (Aml). Blood (2021) 138(Supplement 1):2287. doi: 10.1182/blood-2021-154059%JBlood
38. Maiti A, DiNardo CD, Rausch CR, Pemmaraju N, Garcia-Manero G, Ohanian M, et al. Phase ii trial of ten-day decitabine with venetoclax (Dec10-ven) in acute myeloid leukemia: Updated outcomes in genomic subgroups. Blood (2021) 138(Supplement 1):694. doi: 10.1182/blood-2021-153227%JBlood
39. Yu W-J, Jia J, Wang J, Tang F-F, Gong L, Liu X, et al. Safety and short-term efficacy of venetoclax combined with azacitidine in acute myeloid leukemia: A single institution experience. Blood (2021) 138(Supplement 1):4413. doi: 10.1182/blood-2021-148259%JBlood
40. Bisaillon R, Moison C, Thiollier C, Krosl J, Bordeleau ME, Lehnertz B, et al. Genetic characterization of abt-199 sensitivity in human aml. Leukemia (2020) 34(1):63–74. doi: 10.1038/s41375-019-0485-x
41. Lachowiez CA, Loghavi S, Kadia TM, Daver N, Borthakur G, Pemmaraju N, et al. Outcomes of older patients with Npm1-mutated aml: Current treatments and the promise of venetoclax-based regimens. Blood Adv (2020) 4(7):1311–20. doi: 10.1182/bloodadvances.2019001267
42. Tiong IS, Dillon R, Ivey A, Teh TC, Nguyen P, Cummings N, et al. Venetoclax induces rapid elimination of Npm1 mutant measurable residual disease in combination with low-intensity chemotherapy in acute myeloid leukaemia. Br J Haematol (2021) 192(6):1026–30. doi: 10.1111/bjh.16722
43. Scholz C, Richter A, Lehmann M, Schulze-Osthoff K, Dörken B, Daniel PT. Arsenic trioxide induces regulated, death receptor-independent cell death through a bcl-2-Controlled pathway. Oncogene (2005) 24(47):7031–42. doi: 10.1038/sj.onc.1208868
44. Glaser SP, Lee EF, Trounson E, Bouillet P, Wei A, Fairlie WD, et al. Anti-apoptotic mcl-1 is essential for the development and sustained growth of acute myeloid leukemia. Genes Dev (2012) 26(2):120–5. doi: 10.1101/gad.182980.111
45. Lin KH, Winter PS, Xie A, Roth C, Martz CA, Stein EM, et al. Targeting mcl-1/Bcl-Xl forestalls the acquisition of resistance to abt-199 in acute myeloid leukemia. Sci Rep (2016) 6:27696. doi: 10.1038/srep27696
46. Cho H, Jang JE, Eom JI, Jeung HK, Chung H, Kim JS, et al. Arsenic trioxide synergistically promotes the antileukaemic activity of venetoclax by downregulating mcl-1 in acute myeloid leukaemia cells. Exp Hematol Oncol (2021) 10(1):28. doi: 10.1186/s40164-021-00221-6
47. Zhu HH, Qian JJ, Sun WJ, You LS, Wang QQ, Naranmandura H, et al. Venetoclax and arsenic showed synergistic anti-leukemia activity in vitro and in vivo for acute myeloid leukemia with the Npm1 mutation. Am J Hematol (2020) 95(3):E55–e7. doi: 10.1002/ajh.25719
48. Brunetti L, Gundry MC, Sorcini D, Guzman AG, Huang YH, Ramabadran R, et al. Mutant Npm1 maintains the leukemic state through hox expression. Cancer Cell (2018) 34(3):499–512.e9. doi: 10.1016/j.ccell.2018.08.005
49. Kühn MW, Song E, Feng Z, Sinha A, Chen CW, Deshpande AJ, et al. Targeting chromatin regulators inhibits leukemogenic gene expression in Npm1 mutant leukemia. Cancer Discovery (2016) 6(10):1166–81. doi: 10.1158/2159-8290.Cd-16-0237
50. Uckelmann HJ, Kim SM, Wong EM, Hatton C, Giovinazzo H, Gadrey JY, et al. Therapeutic targeting of preleukemia cells in a mouse model of Npm1 mutant acute myeloid leukemia. Science (2020) 367(6477):586–90. doi: 10.1126/science.aax5863
51. Carter BZ, Tao W, Mak PY, Ostermann LB, Mak D, McGeehan G, et al. Menin inhibition decreases bcl-2 and synergizes with venetoclax in Npm1/Flt3-mutated aml. Blood (2021) 138(17):1637–41. doi: 10.1182/blood.2021011917
52. Dzama MM, Steiner M, Rausch J, Sasca D, Schönfeld J, Kunz K, et al. Synergistic targeting of Flt3 mutations in aml Via combined menin-mll and Flt3 inhibition. Blood (2020) 136(21):2442–56. doi: 10.1182/blood.2020005037
53. Miao H, Kim E, Chen D, Purohit T, Kempinska K, Ropa J, et al. Combinatorial treatment with menin and Flt3 inhibitors induces complete remission in aml models with activating Flt3 mutations. Blood (2020) 136(25):2958–63. doi: 10.1182/blood.2020006575
54. Wang ES, Altman JK, Pettit K, De Botton S, Walter RP, Fenaux P, et al. Preliminary data on a phase 1/2a first in human study of the menin-Kmt2a (Mll) inhibitor ko-539 in patients with relapsed or refractory acute myeloid leukemia. Blood (2020) 136(Supplement 1):7–8. doi: 10.1182/blood-2020-134942%JBlood
55. Stein EM, Aldoss I, DiPersio JF, Stone RM, Arellano ML, Rosen G, et al. Safety and efficacy of menin inhibition in patients (Pts) with mll-rearranged and Npm1 mutant acute leukemia: A phase (Ph) 1, first-in-Human study of sndx-5613 (Augment 101). Blood (2021) 138(Supplement 1):699. doi: 10.1182/blood-2021-146944%JBlood
56. Hutten S, Kehlenbach RH. Crm1-mediated nuclear export: To the pore and beyond. Trends Cell Biol (2007) 17(4):193–201. doi: 10.1016/j.tcb.2007.02.003
57. Luedtke DA, Su Y, Liu S, Edwards H, Wang Y, Lin H, et al. Inhibition of Xpo1 enhances cell death induced by abt-199 in acute myeloid leukaemia Via mcl-1. J Cell Mol Med (2018) 22(12):6099–111. doi: 10.1111/jcmm.13886
58. Ranganathan P, Yu X, Santhanam R, Hofstetter J, Walker A, Walsh K, et al. Decitabine priming enhances the antileukemic effects of exportin 1 (Xpo1) selective inhibitor selinexor in acute myeloid leukemia. Blood (2015) 125(17):2689–92. doi: 10.1182/blood-2014-10-607648
59. Wang AY, Weiner H, Green M, Chang H, Fulton N, Larson RA, et al. A phase I study of selinexor in combination with high-dose cytarabine and mitoxantrone for remission induction in patients with acute myeloid leukemia. J Hematol Oncol (2018) 11(1):4. doi: 10.1186/s13045-017-0550-8
60. Fiedler W, Chromik J, Amberg S, Kebenko M, Thol F, Schlipfenbacher V, et al. A phase ii study of selinexor plus cytarabine and idarubicin in patients with Relapsed/Refractory acute myeloid leukaemia. Br J Haematol (2020) 190(3):e169–e73. doi: 10.1111/bjh.16804
61. Etchin J, Berezovskaya A, Conway AS, Galinsky IA, Stone RM, Baloglu E, et al. Kpt-8602, a second-generation inhibitor of Xpo1-mediated nuclear export, is well tolerated and highly active against aml blasts and leukemia-initiating cells. Leukemia (2017) 31(1):143–50. doi: 10.1038/leu.2016.145
62. Hing ZA, Fung HY, Ranganathan P, Mitchell S, El-Gamal D, Woyach JA, et al. Next-generation Xpo1 inhibitor shows improved efficacy and in vivo tolerability in hematological malignancies. Leukemia (2016) 30(12):2364–72. doi: 10.1038/leu.2016.136
63. Fischer MA, Friedlander SY, Arrate MP, Chang H, Gorska AE, Fuller LD, et al. Venetoclax response is enhanced by selective inhibitor of nuclear export compounds in hematologic malignancies. Blood Adv (2020) 4(3):586–98. doi: 10.1182/bloodadvances.2019000359
64. Zhu J, Koken MH, Quignon F, Chelbi-Alix MK, Degos L, Wang ZY, et al. Arsenic-induced pml targeting onto nuclear bodies: Implications for the treatment of acute promyelocytic leukemia. Proc Natl Acad Sci U.S.A. (1997) 94(8):3978–83. doi: 10.1073/pnas.94.8.3978
65. Huang M, Thomas D, Li MX, Feng W, Chan SM, Majeti R, et al. Role of cysteine 288 in nucleophosmin cytoplasmic mutations: Sensitization to toxicity induced by arsenic trioxide and bortezomib. Leukemia (2013) 27(10):1970–80. doi: 10.1038/leu.2013.222
66. Martelli MP, Gionfriddo I, Mezzasoma F, Milano F, Pierangeli S, Mulas F, et al. Arsenic trioxide and all-trans retinoic acid target Npm1 mutant oncoprotein levels and induce apoptosis in Npm1-mutated aml cells. Blood (2015) 125(22):3455–65. doi: 10.1182/blood-2014-11-611459
67. El Hajj H, Dassouki Z, Berthier C, Raffoux E, Ades L, Legrand O, et al. Retinoic acid and arsenic trioxide trigger degradation of mutated Npm1, resulting in apoptosis of aml cells. Blood (2015) 125(22):3447–54. doi: 10.1182/blood-2014-11-612416
68. Hleihel R, El Hajj H, Wu HC, Berthier C, Zhu HH, Massoud R, et al. A Pin1/Pml/P53 axis activated by retinoic acid in npm-1c-Acute myeloid leukemia. Haematologica (2021) 106(12):3090–9. doi: 10.3324/haematol.2020.274878
69. Schlenk RF, Döhner K, Kneba M, Götze K, Hartmann F, Del Valle F, et al. Gene mutations and response to treatment with all-trans retinoic acid in elderly patients with acute myeloid leukemia. results from the amlsg trial aml Hd98b. Haematologica (2009) 94(1):54–60. doi: 10.3324/haematol.13378
70. Goldberg IH, Rabinowitz M, Reich E. Basis of actinomycin action. i. DNA binding and inhibition of rna-polymerase synthetic reactions by actinomycin. Proc Natl Acad Sci U.S.A. (1962) 48(12):2094–101. doi: 10.1073/pnas.48.12.2094
71. Gionfriddo I, Brunetti L, Mezzasoma F, Milano F, Cardinali V, Ranieri R, et al. Dactinomycin induces complete remission associated with nucleolar stress response in Relapsed/Refractory Npm1-mutated aml. Leukemia (2021) 35(9):2552–62. doi: 10.1038/s41375-021-01192-7
72. Falini B, Brunetti L, Martelli MP. Dactinomycin in Npm1-mutated acute myeloid leukemia. N Engl J Med (2015) 373(12):1180–2. doi: 10.1056/NEJMc1509584
73. Beziat G, Tavitian S, Bertoli S, Huguet F, Largeaud L, Luquet I, et al. Dactinomycin in acute myeloid leukemia with Npm1 mutations. Eur J Haematol (2020) 105(3):302–7. doi: 10.1111/ejh.13438
74. Wu HC, Rérolle D, Berthier C, Hleihel R, Sakamoto T, Quentin S, et al. Actinomycin d targets Npm1c-primed mitochondria to restore pml-driven senescence in aml therapy. Cancer Discovery (2021) 11(12):3198–213. doi: 10.1158/2159-8290.Cd-21-0177
75. Laszlo GS, Estey EH, Walter RB. The past and future of Cd33 as therapeutic target in acute myeloid leukemia. Blood Rev (2014) 28(4):143–53. doi: 10.1016/j.blre.2014.04.001
76. Pollard JA, Alonzo TA, Loken M, Gerbing RB, Ho PA, Bernstein ID, et al. Correlation of Cd33 expression level with disease characteristics and response to gemtuzumab ozogamicin containing chemotherapy in childhood aml. Blood (2012) 119(16):3705–11. doi: 10.1182/blood-2011-12-398370
77. Krupka C, Kufer P, Kischel R, Zugmaier G, Bögeholz J, Köhnke T, et al. Cd33 target validation and sustained depletion of aml blasts in long-term cultures by the bispecific T-Cell-Engaging antibody amg 330. Blood (2014) 123(3):356–65. doi: 10.1182/blood-2013-08-523548
78. De Propris MS, Raponi S, Diverio D, Milani ML, Meloni G, Falini B, et al. High Cd33 expression levels in acute myeloid leukemia cells carrying the nucleophosmin (Npm1) mutation. Haematologica (2011) 96(10):1548–51. doi: 10.3324/haematol.2011.043786
79. Castaigne S, Pautas C, Terré C, Raffoux E, Bordessoule D, Bastie JN, et al. Effect of gemtuzumab ozogamicin on survival of adult patients with de-Novo acute myeloid leukaemia (Alfa-0701): A randomised, open-label, phase 3 study. Lancet (2012) 379(9825):1508–16. doi: 10.1016/s0140-6736(12)60485-1
80. Schlenk RF, Paschka P, Krzykalla J, Weber D, Kapp-Schwoerer S, Gaidzik VI, et al. Gemtuzumab ozogamicin in Npm1-mutated acute myeloid leukemia: Early results from the prospective randomized amlsg 09-09 phase iii study. J Clin Oncol (2020) 38(6):623–32. doi: 10.1200/jco.19.01406
81. Dong H, Strome SE, Salomao DR, Tamura H, Hirano F, Flies DB, et al. Tumor-associated B7-H1 promotes T-cell apoptosis: A potential mechanism of immune evasion. Nat Med (2002) 8(8):793–800. doi: 10.1038/nm730
82. Greiner J, Ono Y, Hofmann S, Schmitt A, Mehring E, Götz M, et al. Mutated regions of nucleophosmin 1 elicit both Cd4+ and Cd8+ T-cell responses in patients with acute myeloid leukemia. Blood (2012) 120(6):1282–9. doi: 10.1182/blood-2011-11-394395%JBlood
83. Greiner J, Schneider V, Schmitt M, Götz M, Döhner K, Wiesneth M, et al. Immune responses against the mutated region of cytoplasmatic Npm1 might contribute to the favorable clinical outcome of aml patients with Npm1 mutations (Npm1mut). Blood (2013) 122(6):1087–8. doi: 10.1182/blood-2013-04-496844
84. Greiner J, Hofmann S, Schmitt M, Götz M, Wiesneth M, Schrezenmeier H, et al. Acute myeloid leukemia with mutated nucleophosmin 1: An immunogenic acute myeloid leukemia subtype and potential candidate for immune checkpoint inhibition. Haematologica (2017) 102(12):e499–501. doi: 10.3324/haematol.2017.176461
85. Brodská B, Otevřelová P, Šálek C, Fuchs O, Gašová Z, Kuželová K. High pd-L1 expression predicts for worse outcome of leukemia patients with concomitant Npm1 and Flt3 mutations. Int J Mol Sci (2019) 20(11):2823. doi: 10.3390/ijms20112823
86. Qin G, Wang X, Ye S, Li Y, Chen M, Wang S, et al. Npm1 upregulates the transcription of pd-L1 and suppresses T cell activity in triple-negative breast cancer. Nat Commun (2020) 11(1):1669. doi: 10.1038/s41467-020-15364-z
87. Saxena K, Herbrich SM, Pemmaraju N, Kadia TM, DiNardo CD, Borthakur G, et al. A phase 1b/2 study of azacitidine with pd-L1 antibody avelumab in Relapsed/Refractory acute myeloid leukemia. Cancer (2021) 127(20):3761–71. doi: 10.1002/cncr.33690
88. Zheng H, Mineishi S, Claxton D, Zhu J, Zhao C, Jia B, et al. A phase I clinical trial of avelumab in combination with decitabine as first line treatment of unfit patients with acute myeloid leukemia. Am J Hematol (2021) 96(2):E46–e50. doi: 10.1002/ajh.26043
89. Daver N, Boddu P, Garcia-Manero G, Yadav SS, Sharma P, Allison J, et al. Hypomethylating agents in combination with immune checkpoint inhibitors in acute myeloid leukemia and myelodysplastic syndromes. Leukemia (2018) 32(5):1094–105. doi: 10.1038/s41375-018-0070-8
90. Chen X, Pan X, Zhang W, Guo H, Cheng S, He Q, et al. Epigenetic strategies synergize with pd-L1/Pd-1 targeted cancer immunotherapies to enhance antitumor responses. Acta Pharm Sin B (2020) 10(5):723–33. doi: 10.1016/j.apsb.2019.09.006
91. Verdegaal EM, de Miranda NF, Visser M, Harryvan T, van Buuren MM, Andersen RS, et al. Neoantigen landscape dynamics during human melanoma-T cell interactions. Nature (2016) 536(7614):91–5. doi: 10.1038/nature18945
92. Blankenstein T, Leisegang M, Uckert W, Schreiber H. Targeting cancer-specific mutations by T cell receptor gene therapy. Curr Opin Immunol (2015) 33:112–9. doi: 10.1016/j.coi.2015.02.005
93. Falini B, Martelli MP, Bolli N, Sportoletti P, Liso A, Tiacci E, et al. Acute myeloid leukemia with mutated nucleophosmin (Npm1): Is it a distinct entity? Blood (2011) 117(4):1109–20. doi: 10.1182/blood-2010-08-299990
94. van der Lee DI, Reijmers RM, Honders MW, Hagedoorn RS, de Jong RC, Kester MG, et al. Mutated nucleophosmin 1 as immunotherapy target in acute myeloid leukemia. J Clin Invest (2019) 129(2):774–85. doi: 10.1172/jci97482
95. Xie G, Ivica NA, Jia B, Li Y, Dong H, Liang Y, et al. Car-T cells targeting a nucleophosmin neoepitope exhibit potent specific activity in mouse models of acute myeloid leukaemia. Nat BioMed Eng (2021) 5(5):399–413. doi: 10.1038/s41551-020-00625-5
96. Ley TJ, Miller C, Ding L, Raphael BJ, Mungall AJ, Robertson A, et al. Genomic and epigenomic landscapes of adult De novo acute myeloid leukemia. N Engl J Med (2013) 368(22):2059–74. doi: 10.1056/NEJMoa1301689
97. Papaemmanuil E, Gerstung M, Bullinger L, Gaidzik VI, Paschka P, Roberts ND, et al. Genomic classification and prognosis in acute myeloid leukemia. N Engl J Med (2016) 374(23):2209–21. doi: 10.1056/NEJMoa1516192
98. Mer AS, Heath EM, Madani Tonekaboni SA, Dogan-Artun N, Nair SK, Murison A, et al. Biological and therapeutic implications of a unique subtype of Npm1 mutated aml. Nat Commun (2021) 12(1):1054. doi: 10.1038/s41467-021-21233-0
99. Falini B, Brunetti L, Martelli MP. How I diagnose and treat Npm1-mutated aml. Blood (2021) 137(5):589–99. doi: 10.1182/blood.2020008211
Keywords: NPM1, AML, targeted therapy, venetoclax, menin inhibitors, XPO1 inhibitors
Citation: Wang R, Xu P, Chang L-L, Zhang S-Z and Zhu H-H (2022) Targeted therapy in NPM1-mutated AML: Knowns and unknowns. Front. Oncol. 12:972606. doi: 10.3389/fonc.2022.972606
Received: 18 June 2022; Accepted: 13 September 2022;
Published: 27 September 2022.
Edited by:
Nicholas Short, University of Texas MD Anderson Cancer Center, United StatesReviewed by:
Branko Cuglievan, University of Texas MD Anderson Cancer Center, United StatesCopyright © 2022 Wang, Xu, Chang, Zhang and Zhu. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Hong-Hu Zhu, emh1aGhkb2NAMTYzLmNvbQ==; Shi-Zhong Zhang, emhhbmdzekBjdGd1LmVkdS5jbg==
†These authors have contributed equally to this work
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.