 Hanxun Yue
Hanxun Yue Zenan Hu2,3†
Zenan Hu2,3† Rui Hu
Rui Hu Ya Zheng
Ya Zheng Yuping Wang
Yuping Wang Yongning Zhou
Yongning Zhou
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Oncol. , 22 June 2022
Sec. Cancer Genetics
Volume 12 - 2022 | https://doi.org/10.3389/fonc.2022.918778
This article is part of the Research Topic Reviews in Cancer Genetics View all 20 articles
Aldehyde dehydrogenases 1 family member A1(ALDH1A1) gene codes a cytoplasmic enzyme and shows vital physiological and pathophysiological functions in many areas. ALDH1A1 plays important roles in various diseases, especially in cancers. We reviewed and summarized representative correlative studies and found that ALDH1A1 could induce cancers via the maintenance of cancer stem cell properties, modification of metabolism, promotion of DNA repair. ALDH1A1 expression is regulated by several epigenetic processes. ALDH1A1 also acted as a tumor suppressor in certain cancers. The detoxification of ALDH1A1 often causes chemotherapy failure. Currently, ALDH1A1-targeted therapy is widely used in cancer treatment, but the mechanism by which ALDH1A1 regulates cancer development is not fully understood. This review will provide insight into the status of ALDH1A1 research and new viewpoint for cancer therapy.
Aldehyde dehydrogenases (ALHDs) are a group of cytoplasmic enzymes that use nicotinamide adenine dinucleotide (NAD) as a coenzyme to oxidize aldehydes into the corresponding carboxylic acids (1, 2). The human genome is thought to contain 19 functional ALDH genes, including ALDH1 family genes. ALDH1A1 is one of the main members of the ALDH1 family. The ALDH1A1 gene is located in subregion 13 of region 21, on the long arm of human chromosome 9. This gene encodes homotetrameric cytoplasmic proteins in various tissues. ALDH1A1 has a greater affinity for the oxidation of both all-trans and 9-cis-retinal molecules than ALDH1A2 and ALDH1A3 (3). ALDH1A has an NAD+ binding pocket (amino acids 8–135 and 159–270), a catalytic site (amino acids 271–470), and an oligomerization domain (amino acids 140–158 and 486–459). ALDH1A1 can exist in monomeric, dimeric, or tetrameric forms. The tetrameric and monomeric forms of ALDH1A1 are the most abundant. ALDH1A1 exists predominantly as a tetramer at high concentrations but the activity of its monomeric form is the highest (4). According to the GenBank database (https://www.ncbi.nlm.nih.gov/genbank/), ALDH1A1 was highly expressed in the liver, duodenum, and other tissues.
In the past decade, researchers have found that ALDH1A1 had vital physiological and pathophysiological functions in many systems, such as the central nervous system, as well as inflammatory and metabolic disorders (5–7). ALDH1A1 overexpression has been found to play an important role in obesity, diabetes, and other diseases (8–13). Because the retinoic acid (RA) signaling pathway is involved in the regulation of gene expression in cancer stem cells (CSCs), researchers have focused on the role of ALDH1A1 in cancers worldwide (14–16). Although several reviews on ALDH1A1 are available, no review has been published that discusses the roles of ALDH1A1 in all cancers (6, 17–19) (Table 1).
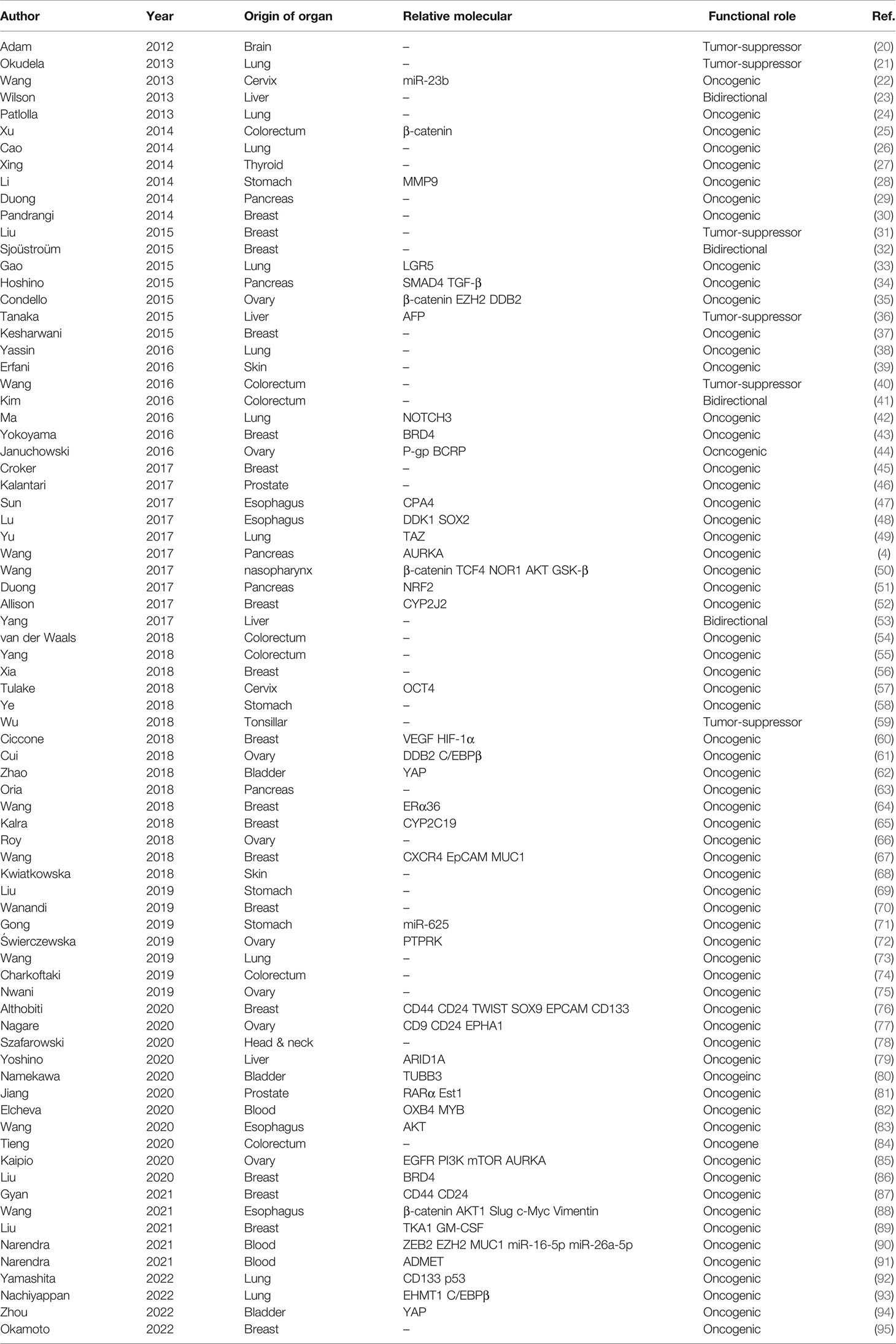
Table 1 Representative Studies of ALDH1A1 in Cancers.
To understand the roles of ALDH1A1 in cancers, we reviewed and summarized representative correlative studies in this article. We summarized the consensus and controversies regarding the functions, regulatory mechanism, diagnostic value, and selective inhibitors of ALDH1A1. Based on the results of our experiments and bioinformatic assay, the potential uses of ALDH1A1 in hepatocellular carcinoma (HCC) will be discussed.
Since 2010, numerous studies have verified the fact that ALDH1A1 could promote tumor initiation and tumor progression. Several years ago, Yassin et al. had found that ALDH1A1 overexpression could reflect the poor historical subtype and advanced tumor grade in lung cancer patients (38). In a comparative study performed by Cao et al., the ALDH1A1 levels were found to be much higher in non-small cell lung cancer (NSCLC) patients at advanced stages than those with early-stage tumors (26). In contrast, triple−negative cases without ALDH1A1, CD133, and mutant p53 expression in lung adenocarcinomas were shown to have a much better prognosis than other cases (92).
The same is true for cancers of the digestive system. Li et al. found that ALDH1A1 overexpression was significantly associated with larger tumor size, deeper invasion, extensive lymph node metastasis, and advanced stage of gastric cancer. ALDH1A1 could represent an independent prognostic factor for both overall survival (OS) and recurrence-free survival (RFS) (28). ALDH1A1 overexpression was also a poor prognostic indicator of survival in patients with gastric neuroendocrine carcinoma (58). Liu et al. reported that ALDH1A1 overexpression was significantly associated with poorly differentiated histology in gastric cancer (69). Xu et al. found that ALDH1A1 overexpression had the same effect in patients with colorectal cancer as observed in gastric cancer (25, 54, 55). An analysis of the Oncomine database showed that ALDH1A1 was significantly upregulated in HCC tissues, compared to non-tumorous liver tissues (53). Peng et al. demonstrated that rs7852860 variants of the ALDH1A1 gene were associated with susceptibility to anti-tuberculosis drug-induced liver injury (96).
Extensive and in-depth studies on ALDH1A1 have been performed in breast cancer patients. First, Croker et al. determined the RNA expression of ALDH1A1 in breast cancer cells and found that ALDH1A1 overexpression contributed functionally the proliferation, adhesion, migration, extravasation, and micrometastasis of breast cancer (45). Althobiti et al. found that ALDH1A1 overexpression was associated with poor prognostic features, including an increased tumor grade, poor Nottingham prognostic index, extensive lymph node metastasis, and a greater extent of luminal B and triple-negative subtypes of breast cancer (76). In the African population, Gyan et al. found that ALDH1A1 was expressed at a high level in 90% of breast cancer specimens. This study further confirmed the increased oncogenicity of the CD44+/CD24-/ALDH1A1+ combination phenotype and its association with the increased tumor grade and clinical prognostic stage (87). Xia et al. conducted a population-based study to analyze the relationship between ALDH1A1 polymorphisms, alcohol consumption, and mortality among women diagnosed with breast cancer. They found that after adjusting all the results for multiple comparisons, rs7027604 was significantly associated with all-cause mortality in the rs1424482 CC genotype, and the rs7027604 AA genotype was positively associated with non-breast cancer mortality. Among long-term light drinkers, rs1888202 was associated with decreased all-cause mortality, while the association was not significant among non-drinkers or moderate/heavy drinkers. The increased risk of all-cause mortality associated with rs63319 was limited to women with a low level of native American ancestry (56). Furthermore, Wanandi et al. found that the expression of ALDH1A1 was higher in breast cancer stem cells than in the MCF-7 cell line, but was almost similar to that observed in the more aggressive cell line MDA-MB-231. These results suggested that ALDH1A1 overexpression might be related to the stemness and aggressiveness of breast cancers cells (70).
ALDH1A1 has been reported to promote cancers of the reproductive system. Nagare et al. found that most ALDH1A1-positive high-grade ovarian cancer cells resided in the G1 phase of the cell cycle. They also reported that the ALDH1A1-positive cells co-expressing the combination of CD9, CD24, or EPHA1 were more oncogenic and aggressive than ALDH1A1-negative cells (77). Tulake et al. found that ALDH1A1 and OCT4 were upregulated in both cervical squamous cell carcinoma and cervical intraepithelial neoplasia, as compared to healthy subjects. They found that ALDH1A1 expression levels were also increased in the peripherical blood obtained from cervical cancer patients; thus, ALDH1A1 expression could be regarded as an indicator of cervical cancer (57).
ALDH1A1 is also involved in the development of cancers that originate in other systems. A study involving Iranian prostate cancer patients by Kalantari et al. showed that the level of ALDH1A1 expression was positively correlated with tumor invasiveness (46). Among the most common cancer stem cell markers, only ALDH1A1 overexpression significantly affected the five-year OS of primary head and neck squamous cell carcinoma patients (78). ALDH1A1 levels were also higher in papillary thyroid carcinoma tissues than in normal thyroid tissues. ALDH1A1 overexpression was significantly associated with extrathyroidal extension and reflected a poorer RFS and distant recurrence-free survival (27). ALDH1A1 overexpression was also found to occur in skin cancers, particularly in melanomas (39).
Although ALDH1A1 is seen as an oncogenic factor, it also exhibits different characteristics in some cancers. In a study performed by Adam et al., ALDH1A1 was co-expressed with GFAP and S100 in mature astrocytes and was a better prognostic marker for glioblastoma patients (20). In an experiment conducted by Wang et al., both cytoplasmic and nuclear expression levels were assessed in epithelial cells. Surprisingly, in the tissue microarray and whole-tissue cohorts, univariate analysis indicated that the cytoplasmic expression of ALDH1A1 cannot be considered a prognostic marker for colorectal cancers. Furthermore, nuclear expression levels of ALDH1A1 were significantly associated with longer disease-specific survival and nuclear expression levels in low-grade adenomas, and were predominantly higher than those in high-grade adenomas, primary colorectal cancer, and the corresponding liver metastases (40). Although ALDH1A1 is ubiquitously expressed in the liver, its function in HCC is still ambiguous. Tanaka et al. found that there was no significant difference in the ALDH1A1 level between HCC and non-cancerous liver tissues. In their study, the group with high ALDH1A1 levels was significantly associated with low serum levels of AFP, a small tumor diameter, low levels of lymphovascular invasion, a more differentiated pathology, and a less advanced stage (36). Yang et al. studied the relationship between ALDH1A1 and HCC using the GEO database and found high ALDH1A1 mRNA expression levels were significantly associated with longer 57–month recurrence-free survivals (53). Okudela et al. also found that the level of ALDH1A1 expression was negatively related to carcinogenesis in NSCLC patients. ALDH1A1 was remarkably downregulated in adenocarcinomas and large cell cancers. Among adenocarcinomas, the downregulation of ALDH1A1 tended to be more significant in high-grade, poorly differentiated tumors, and tumors with a stronger proliferating activity. Moreover, the incidence of this reduction was higher in smokers than in non-smokers (21). Liu et al. suggested that the high level of expression of ALDH1A1 mRNA in tumor tissues may be an independent predictor of favorable triple-negative breast cancer, based on an analysis performed using three databases and meta-analyses (31). Wu et al. reported that ALDH1A1-positive cells were a unique component of the crypt cellular microenvironment and were not stem cells. They also found that NGFR-positive and ALDH1A1-positive cells were lost during tumorigenesis with the expression of LGR5 in the tonsillar crypt niche; this may mark the breakdown of the normal microenvironment (59).
However, several groups of researchers believe that the behavior of ALDH1A1 is complex. In a study performed by Kim et al., ALDH1A1 overexpression decreased the proliferation and invasiveness of colorectal cancer cells, while colorectal cancer liver metastasis was more likely to occur in SW480/ALDH1A1-transfected mice (41). Sjoüstroüm et al. hypothesized that in breast cancer cells, ALDH1A1 overexpression was associated with either a better or a worse prognosis, depending on the cut-off. If weakly stained cells were considered to be positively stained, ALDH1A1 overexpression was associated with a better prognosis in two cohorts. If strongly stained cells were considered to be positively stained, ALDH1A1 overexpression was associated with a worse prognosis in one of the cohorts. In addition, stromal ALDH1A1 staining was associated with improved distant disease-free survival, and gene expression analysis showed that there was a relationship between ALDH1A1 overexpression and a favorable prognosis (32).
Chemotherapy plays an important role in cancer treatment. However, many factors, including ALDH1A1, can cause chemotherapy failure. A study by Ma et al. showed that cisplatin induced NOTCH3 expression, and NOTCH3 overexpression was a prognostic factor for shorter progression-free survival and OS in NSCLC patients. They suggested that the chemoresistance of NSCLC patients was attributable to the promotion of ALDH1A1 expression by NOTCH3 and stimulation of autophagy (42). Wang et al. performed a UPLC−MS-based metabolomics analysis and revealed the metabolic dysregulation in lung adenocarcinoma. They found that the metabolic features of lung cancer cells were altered by ALDH1A1 overexpression, and levels of most metabolites, such as glucose-6-phosphate, fructose 1,6-diphosphate, propionyl-CoA, malic acid, phosphatidylcholine, glycerol phosphatidylcholine, GMP, citrulline, and arginine succinic acid were increased. These metabolites were involved in metabolic pathways such as glycolysis, the tricarboxylic acid cycle, glycerophospholipid metabolism, nucleotide metabolism, and the urea cycle. Among these, ALDH1A1 may amplify drug resistance in tumors through nucleotide metabolic pathways (73).
ALDH1A1-induced drug resistance is also common in cancers of the digestive system. Wang et al. found that ALDH1A1 promoted the development of resistance to 5-fluorouracil in esophageal squamous cell carcinoma (83). Oria et al. found that ALDH1A1 reduced the sensitivity of pancreatic cells toward gemcitabine and chemoradiation treatment (63). Fortunately, Duong et al. suggested that a therapeutic strategy involving a combination of dasatinib and gemcitabine might overcome gemcitabine resistance, as it would decrease the level of ALDH1A1 expression in pancreatic cancer. They also found that NRF2 could promote ALDH1A1 expression and the silencing of NRF2 could enhance the anti-proliferative effects of the chemotherapeutic agent 5-fluorouracil in pancreatic cancer cells (29, 51).
ALDH1A1 is highly expressed in breast cancer cells, and the knockdown of ALDH1A1 can significantly sensitize breast cancer cells to chemotherapy and radiotherapy (45). Wang et al. found that tamoxifen could promote ERα36 binding and the activation of estrogen-responsive elements in the ALDH1A1 promoter, to increase the transcription of ALDH1A1, which accounted for the resistance to hormone therapy and metastasis of breast cancer (64). In another study of breast cancer patients on adjuvant therapy, Kalra et al. found that CYP2C19 and ALDH1A1*2 (17 bp deletion), were significantly associated with the disease outcome, including OS, recurrence, and metastasis. Both these genes were involved in the pharmacokinetics of cyclophosphamide. Allison et al. hypothesized that ALDH1A1 was activated in the MDA-MB-468 breast cancer cell line, which stably expressed CYP2J2 and attenuated caspase-3/7 activity and the production of reactive oxygen species induced by cytotoxic agents, such as paclitaxel, doxorubicin, sorafenib, and staurosporin (52).
ALDH1A1-induced drug resistance also seriously hampered ovarian cancer and leukemia treatment. Data from a study performed by Nwani et al. demonstrated that ALDH1A1 was upregulated in ovarian cancer cells that survived exposure to platinum (75). Roy et al. found that ALDH1A1 overexpression was associated with a poor response to platinum-based therapy in patients with high-grade serous ovarian cancer (66). via Kaipio et al. found that ALDH1A1 expression was improved after neoadjuvant chemotherapy in high-grade serous ovarian cancers and that in treatment-naive tumors, ALDH1A1 overexpression was correlated with drug resistance and a reduced duration of survival. Notably, they mentioned that EGFR, PI3K-mTOR, and the AURKA inhibitor were toxic to cancer cells in tests that assessed drug sensitivity and resistance (85). Individuals with chronic myelogenous leukemia acquired resistance to cyclophosphamide, owing to the inactivation of its active metabolite aldophosphamide via the overexpression of ALDH1A1 (91).
Several previous studies have reported that many molecules and signal pathways are involved in the mechanism underlying ALDH1A1 regulation in cancers (28, 33, 47, 60, 72) (Figure 1). Our review described comprehensively the mechanisms of ALDH1A1 in the different processes of cancers and ALDH1A1 expression regulation. ALDH1A1 could induce cancers via the maintenance of CSC properties, modification of metabolism and promotion of DNA repair (43, 80, 81, 86). ALDH1A1 expression is regulated by several epigenetic processes, including, phosphorylation methylation, acetylation, methylation and miRNA modification (34, 71, 82, 90, 91).
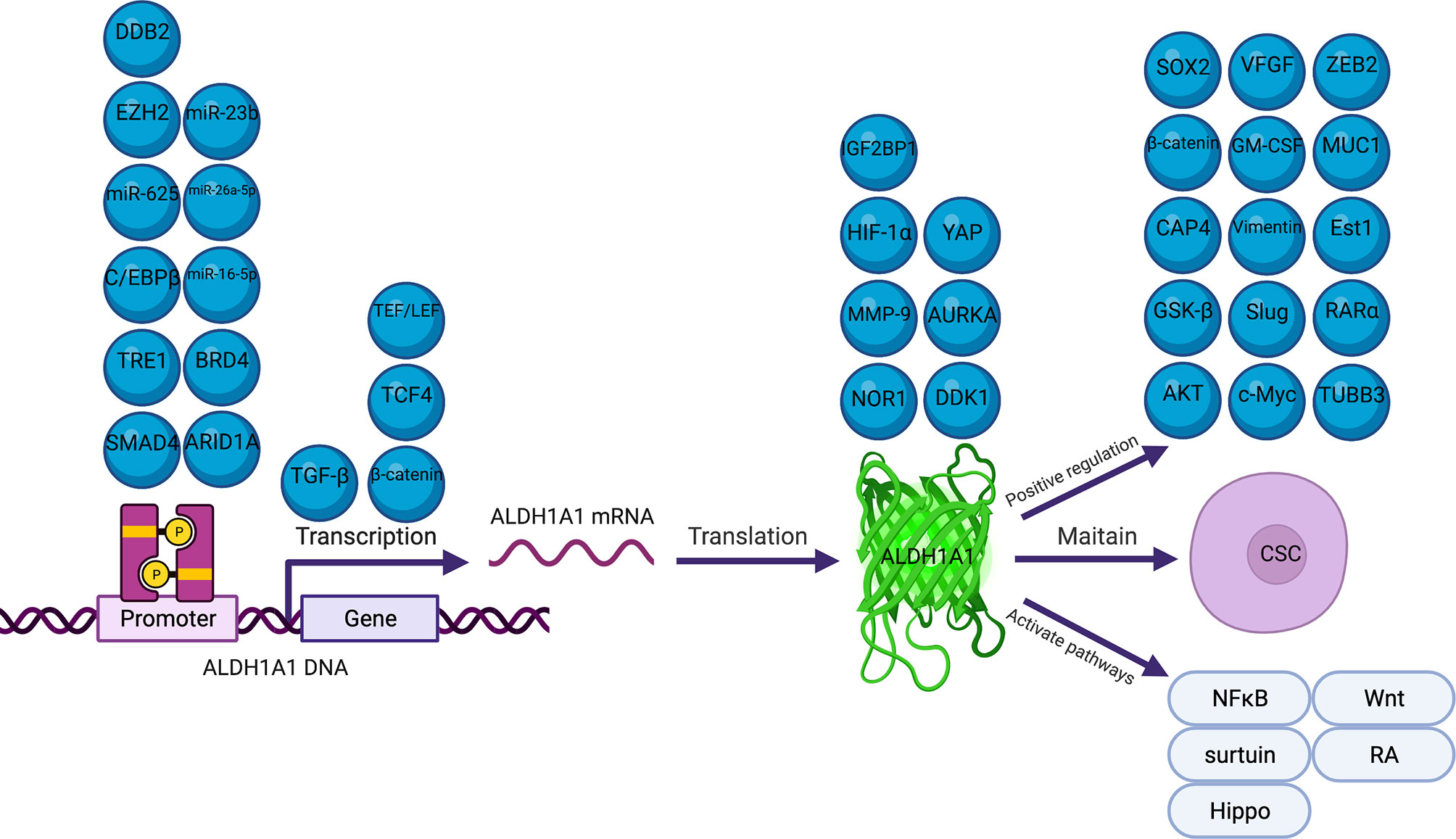
Figure 1 Molecules Involved in the Regulation of ALDH1A1.
CSCs have self-renewal capacity and differentiation potential and contribute to multiple tumor malignancies, such as recurrence, metastasis, heterogeneity, multidrug resistance, and radiation resistance (97). ALDH1A1 is thought to maintain CSC properties in a variety of cancers (49, 88). Yu et al. suggested that ALDH1A1 was a critical gene involved in the mediation of TAZ-induced lung tumorigenesis and CSC phenotypes. They found that TRE1, which was localized in the -256 ~ +52 region of the ALDH1A1 promoter, was majorly responsible for the activation of TAZ (49). Lu et al. suggested that DKK1 maintained the cancer stem-like properties of esophageal cancer cells via the ALDH1A1/SOX2 axis (48). Another group of researchers found that ALDH1A1 could also maintain esophageal CSC properties through promoting the stability of β-catenin and activating the AKT signal pathway (83). Wang et al. reported that the ectopic overexpression of NOR1 suppressed ALDH1A1 and β-catenin expression; β-catenin/TCF4 targeted the regulation of ALDH1A1 transcription; and the silencing of ALDH1A1 reduced AKT and GSK-β expression levels and resulted in the feedback inhibition of β-catenin expression. As a result, NOR1 could suppress the tumorigenic properties of CSCs in nasopharyngeal carcinoma via this signal circuit (50). The most recent study by Nachiyappan et al. showed that EHMT1 could promote tumor progression and maintain the stemness of alveolar rhabdomyosarcoma via the stabilization of C/EBPβ, which could activate the ALDH1A1 promotor (93).
As a metabolic enzyme, the effect of ALDH1A1 on metabolism plays an important role in cancer progression. Charkoftaki et al. tried to identify the role of ALDH1A1 in lipid metabolism in colon cancer cells using an integrated multi-omics approach. They found that the suppression of ALDH1A1 could downregulate oxidative phosphorylation, mitochondrial function, the sirtuin signaling pathway, and the retinol metabolism pathway. This approach provided greater insights into the pathways through which ALDH1A1 drives the development of cancers (74). Liu et al. made a breakthrough in research regarding the mechanism by which ALDH1A1 initiates breast cancer. They found that ALDH1A1 decreased the intracellular pH in breast cancer cells, in order to promote the phosphorylation of TAK1, activate the NFκB signal pathway, and increase the secretion of GM-CSF, and this led to myeloid-derived suppressor cell expansion and immunosuppression (89).
Phosphorylation is one of the most extensively and diligently studied posttranslational modifications which orchestrates a variety of cellular functions like cell growth, differentiation and apoptosis (98). Wang et al. showed that the phosphorylation-dependent regulation of ALDH1A1 was mediated by AURKA. They found AURKA could phosphorylate ALDH1A1 at the locations T267, T422, T439, at which phosphorylation primarily regulated ALDH1A1 activity. AURKA-mediated phosphorylation could rapidly facilitate the dissociation of tetrameric ALDH1A1 into a highly active monomeric species. Surprisingly, ALDH1A1 also reciprocates and prevents the degradation of AURKA, thereby triggering a positive activation loop that drives highly aggressive phenotypes in pancreatic cancer (4).
Acetylation influence a myriad of cellular and physiological processes, including transcription, phase separation, autophagy, mitosis, differentiation and neural function (99). In a study by Yoshino et al., chromatin immunoprecipitation assay showed that the level of H3K27 acetylation was significantly increased in the promoter region of ALDH1A1, while the HDAC1 level was significantly decreased in the ARID1A knocked-out cholangiocarcinoma cell line. Therefore, ARID1A may function as a tumor suppressor in cholangiocarcinoma through the transcriptional downregulation of ALDH1A1, along with a decrease in the levels of histone H3K27 acetylation (79).
The trimethylation of H3K27 was significantly correlated with the expression of ALDH1A1. In a study regarding ovarian cancer, Condello et al. identified that β-catenin could regulate ALDH1A1 via the TEF/LEF transcriptional complex, which was a key element of the Wnt signaling pathway. This mechanism could enhance spheroid formation in ovarian cancer cells. They reported that DDB2 recruited EZH2 to the ALDH1A1 promoter region, thereby facilitating the trimethylation of the local histone H3 at K27, and repressed the transcription of ALDH1A1 (35). Cui et al. also found that DDB2 could bind to the ALDH1A1 gene promoter and facilitate the enrichment of histone H3K27me3, and compete with the transcription factor C/EBPβ for binding to this region, eventually inhibiting the promoter activity of ALDH1A1. This mechanism involved the repression of ovarian cancer cell dedifferentiation (61).
MicroRNAs are small noncoding RNAs that regulate gene expression via recognition of cognate sequences and interference of transcriptional, translational or epigenetic processes (100). Bioinformatics analysis identified miR-16-5p and miR-26a-5p to be hub miRNAs for ALDH1A1 (91). The regulatory mechanism of ALDH1A1 was discussed in a study by Wang et al., who found that miR-23b could directly bind to the 3´-UTR region of ALDH1A1, to cause its reduction (22).
Because ALDH1A1 is an oncogenic factor in many cancers, treatments targeting ALDH1A1 have become a research hotspot. During the diagnosis of NSCLC, the combined application of ALDH1A1 and carcinoembryonic antigen can significantly increase the sensitivity, compared to that observed with the use of carcinoembryonic antigen alone (26). Okamoto et al. developed a new long-acting fluorescence probe that could identify breast cancer stem cells via the targeting of ALDH1A1 (95).
Many researchers have developed specific ALDH1A1-targeting drugs that could be used to treat cancer (Table 2). Some of them believe that the combination of anti-ALDH1A1 therapy and chemotherapy can offset the ALDH1A1-induced drug resistance in cancer patients. Patlolla et al. showed that β-escin could inhibit tobacco carcinogen-induced lung tumor formation by modulating ALDH1A1-positive cells (24). Silybin could efficiently inhibit the proliferation, invasion, and metastasis of prostate cancer cells, by reducing ALDH1A1 expression levels (81). In 2015, Kesharwani et al. reported a new approach for overcoming drug resistance to breast chemotherapy via the targeting of synthetic curcumin analogs against ALDH1A (37). Wang et al. found that quercetin could inhibit the proliferation, clonal expansion, and mammsophere formation of CD44+/CD24- breast cancer stem cells by inhibiting ALDH1A1, CXCR4, EpCAM, and MUC1 (67). Pandrangi et al. found that ellipticine, a plant alkaloid, could inhibit mammosphere formation in ALDH1A1 overexpressed breast cancer stem cells, whereas paclitaxel enhanced mammosphere formation in the same cell lines (30). Condello et al. discovered a novel ALDH1A1 small molecule inhibitor named A37, which could moderately sensitize ovarian cancer cells to cisplatin (35). Liu et al. reported that NCT-501, an ALDH1A1 selective inhibitor, could augment the efficacy of olaparib during ovarian cancer treatment (86). Another group of researchers reported that the ALDH1A1 inhibitor disulfiram and chemotherapeutic agent gemcitabine cooperatively inhibited breast tumor growth and tumorigenesis by purging ALDH+ tumor-initiating cells and activating T-cell immunity (89). Recently, benzimidazole derivatives have been found to act as potent and selective ALDH1A1 inhibitors (8). It has been proven that N42 could also selectively bind to and inhibit ALDH1A1 (9).
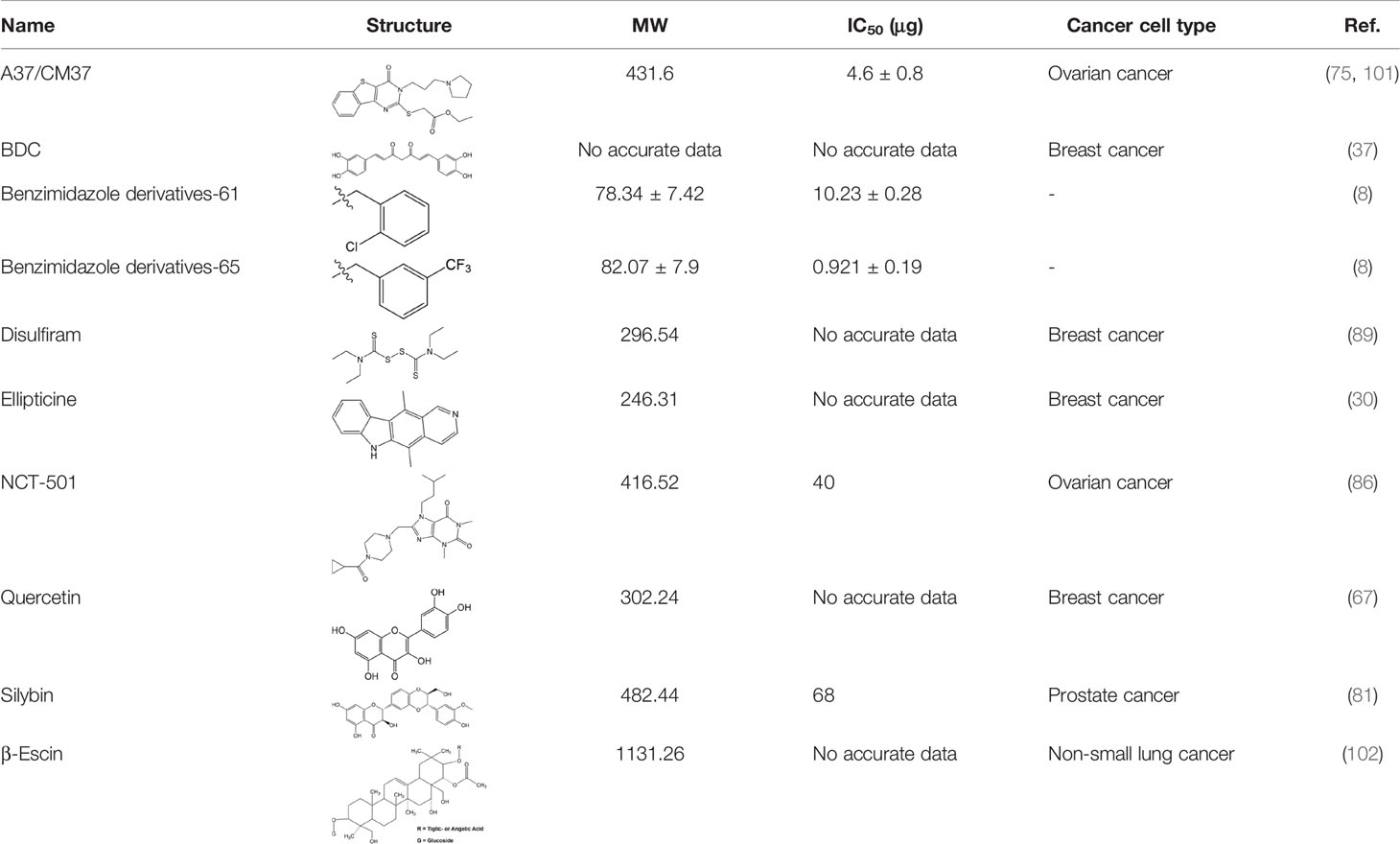
Table 2 Proven Inhibitors of ALDH1A1.
Several ALDH1A1-based cancer prevention and treatment measures have been developed. The oligomer-dependent activity of ALDH1A1 signifies that the targeting of its oligomerization state may be an efficient therapeutic approach that could counteract its protective functions in cancer (4). Januchowski et al. found that ATRA treatment could lead to the downregulation of ALDH1A1, P-gp, and BCRP proteins (44). Yokoyama et al. showed that JQ1, one of the bromodomain and extra-terminal inhibitors, suppressed BRD4-mediated ALDH1A1 expression through a super-enhancer element and its associated enhancer RNA. They also found that the combination of JQ1 and cisplatin could improve the survival of mice with ovarian cancer (43). Moreover, an allogeneic, whole-cell, genetically modified therapeutic melanoma vaccine could generate immune responses to ALDH1A1 and improve long-term survival in advanced melanoma patients (68).
In this review, we summarize and discuss studies that examined the roles of ALDH1A1 in cancers. Over the past decade, researchers have discovered that ALDH1A1 could induce cancers via the maintenance of cancer stem cell properties, modification of metabolism, promotion of DNA repair. ALDH1A1 expression is regulated by several epigenetic processes. ALDH1A1 also acted as a tumor suppressor in certain cancers. ALDH1A1 is highly expressed in the liver, and this is attributable to its bidirectional functions in HCC. The detoxification of ALDH1A1 often causes chemotherapy failure. Currently, ALDH1A1-targeted therapy is widely used in cancer treatment, but the mechanism by which ALDH1A1 regulates cancer development is not fully understood. In our previous studies, we observed that ALDH1A1 expression was significantly improved in HCC and the knockdown of ALDH1A1 weakened the proliferation and invasion of the Huh-7 cell line. Unlike in other cancers, ALDH1A1 did not maintain the properties of liver CSCs (23). Therefore, we believe that ALDH1A1 can induce HCC by mechanisms other than those involving CSCs. In clinical practice, the detoxification of ALDH1A1 may prolong the survival of HCC patients. This hypothesis needs to be tested in future studies. In the future, we will continue to explore the roles of ALDH1A1 in cancers, especially in HCC.
HY and ZH accomplished the manuscript and equally contributed to this study. YNZ and YW designed this study. YZ, RH and ZG provided guidance for this study. All authors contributed to the article and approved the submitted version.
This work was supported by the Foundation of the First Hospital of Lanzhou University, China (ldyyyn2021-3), the Open Fund of Gansu Provincial Key Laboratory for Biotherapy and Regenerative Medicine (zdsyskfkt-201705).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
We appreciate the linguistic assistance provided by TopEdit (www.topeditsci.com) during the preparation of this manuscript.
1. Marcato P, Dean CA, Giacomantonio CA, Lee PW. Aldehyde Dehydrogenase: Its Role as a Cancer Stem Cell Marker Comes Down to the Specific Isoform. Cell Cycle (2011) 10(9):1378–84. doi: 10.4161/cc.10.9.15486
2. Ma I, Allan AL. The Role of Human Aldehyde Dehydrogenase in Normal and Cancer Stem Cells. Stem Cell Rev Rep (2011) 7(2):292–306. doi: 10.1007/s12015-010-9208-4
3. Tomita H, Tanaka K, Tanaka T, Hara A. Aldehyde Dehydrogenase 1A1 in Stem Cells and Cancer. Oncotarget (2016) 7(10):11018–32. doi: 10.18632/oncotarget.6920
4. Wang J, Nikhil K, Viccaro K, Chang L, White J, Shah K. Phosphorylation-Dependent Regulation of ALDH1A1 by Aurora Kinase A: Insights on Their Synergistic Relationship in Pancreatic Cancer. BMC Biol (2017) 15(1):10. doi: 10.1186/s12915-016-0335-5
5. Calleja LF, Yoval-Sánchez B, Hernández-Esquivel L, Gallardo-Pérez JC, Sosa-Garrocho M, Marín-Hernández Á, et al. Activation of ALDH1A1 by Omeprazole Reduces Cell Oxidative Stress Damage. FEBS J (2021) 288(13):4064–80. doi: 10.1111/febs.15698
6. Carmichael K, Evans RC, Lopez E, Sun L, Kumar M, Ding J, et al. Function and Regulation of ALDH1A1-Positive Nigrostriatal Dopaminergic Neurons in Motor Control and Parkinson’s Disease. Front Neural Circuits (2021) 15:644776. doi: 10.3389/fncir.2021.644776
7. Lerner S, Eilam R, Adler L, Baruteau J, Kreiser T, Tsoory M, et al. ASL Expression in ALDH1A1(+) Neurons in the Substantia Nigra Metabolically Contributes to Neurodegenerative Phenotype. Hum Genet (2021) 140(10):1471–85. doi: 10.1007/s00439-021-02345-5
8. Ma Z, Jiang L, Li B, Liang D, Feng Y, Liu L, et al. Discovery of Benzimidazole Derivatives as Potent and Selective Aldehyde Dehydrogenase 1A1 (ALDH1A1) Inhibitors With Glucose Consumption Improving Activity. Bioorg Med Chem (2021) 46:116352. doi: 10.1016/j.bmc.2021.116352
9. Haenisch M, Nguyen T, Fihn CA, Goldstein AS, Amory JK, Treuting P, et al. Investigation of an ALDH1A1-Specific Inhibitor for Suppression of Weight Gain in a Diet-Induced Mouse Model of Obesity. Int J Obes (Lond) (2021) 45(7):1542–52. doi: 10.1038/s41366-021-00818-1
10. Landrier JF, Kasiri E, Karkeni E, Mihály J, Béke G, Weiss K, et al. Reduced Adiponectin Expression After High-Fat Diet is Associated With Selective Up-Regulation of ALDH1A1 and Further Retinoic Acid Receptor Signaling in Adipose Tissue. FASEB J (2017) 31(1):203–11. doi: 10.1096/fj.201600263RR
11. Nikhil K, Viccaro K, Shah K. Multifaceted Regulation of ALDH1A1 by Cdk5 in Alzheimer’s Disease Pathogenesis. Mol Neurobiol (2019) 56(2):1366–90. doi: 10.1007/s12035-018-1114-9
12. Li X, Chen W, Huang X, Jing W, Zhang T, Yu Q, et al. Synaptic Dysfunction of Aldh1a1 Neurons in the Ventral Tegmental Area Causes Impulsive Behaviors. Mol Neurodegener (2021) 16(1):73. doi: 10.1186/s13024-021-00494-9
13. Yang Y, Wu G, Li Q, Zheng Y, Liu M, Zhou L, et al. Angiogenesis-Related Immune Signatures Correlate With Prognosis, Tumor Microenvironment, and Therapeutic Sensitivity in Hepatocellular Carcinoma. Front Mol Biosci (2021) 8:690206. doi: 10.3389/fmolb.2021.690206
14. Chanda B, Ditadi A, Iscove NN, Keller G. Retinoic Acid Signaling is Essential for Embryonic Hematopoietic Stem Cell Development. Cell (2013) 155(1):215–27. doi: 10.1016/j.cell.2013.08.055
15. Duester G, Mic FA, Molotkov A. Cytosolic Retinoid Dehydrogenases Govern Ubiquitous Metabolism of Retinol to Retinaldehyde Followed by Tissue-Specific Metabolism to Retinoic Acid. Chem Biol Interact (2003) 143-144:(201–10). doi: 10.1016/s0009-2797(02)00204-1
16. Qiu JJ, Zeisig BB, Li S, Liu W, Chu H, Song Y, et al. Critical Role of Retinoid/Rexinoid Signaling in Mediating Transformation and Therapeutic Response of NUP98-RARG Leukemia. Leukemia (2015) 29(5):1153–62. doi: 10.1038/leu.2014.334
17. Liu Y, Lv DL, Duan JJ, Xu SL, Zhang JF, Yang XJ, et al. ALDH1A1 Expression Correlates With Clinicopathologic Features and Poor Prognosis of Breast Cancer Patients: A Systematic Review and Meta-Analysis. BMC Cancer (2014) 14(444). doi: 10.1186/1471-2407-14-444
18. Li B, Yang K, Liang D, Jiang C, Ma Z. Discovery and Development of Selective Aldehyde Dehydrogenase 1A1 (ALDH1A1) Inhibitors. Eur J Med Chem (2021) 209:(112940). doi: 10.1016/j.ejmech.2020.112940
19. Dancik GM, Voutsas IF, Vlahopoulos S. Lower RNA Expression of ALDH1A1 Distinguishes the Favorable Risk Group in Acute Myeloid Leukemia. Mol Biol Rep (2022) 49(4):3321–31. doi: 10.1007/s11033-021-07073-7
20. Adam SA, Schnell O, Poschl J, Eigenbrod S, Kretzschmar HA, Tonn JC, et al. ALDH1A1 is a Marker of Astrocytic Differentiation During Brain Development and Correlates With Better Survival in Glioblastoma Patients. Brain Pathol (2012) 22(6):788–97. doi: 10.1111/j.1750-3639.2012.00592.x
21. Okudela K, Woo T, Mitsui H, Suzuki T, Tajiri M, Sakuma Y, et al. Downregulation of ALDH1A1 Expression in non-Small Cell Lung Carcinomas–its Clinicopathologic and Biological Significance. Int J Clin Exp Pathol (2013) 6(1):1–12.
22. Wang W, Li Y, Liu N, Gao Y, Li L. MiR-23b Controls ALDH1A1 Expression in Cervical Cancer Stem Cells. BMC Cancer (2017) 17(1):292. doi: 10.1186/s12885-017-3192-x
23. Wilson GS, Hu Z, Duan W, Tian A, Wang XM, McLeod D, et al. Efficacy of Using Cancer Stem Cell Markers in Isolating and Characterizing Liver Cancer Stem Cells. Stem Cells Dev (2013) 22(19):2655–64. doi: 10.1089/scd.2012.0703
24. Patlolla JM, Qian L, Biddick L, Zhang Y, Desai D, Amin S, et al. β-Escin Inhibits NNK-Induced Lung Adenocarcinoma and ALDH1A1 and RhoA/Rock Expression in a/J Mice and Growth of H460 Human Lung Cancer Cells. Cancer Prev Res (Phila) (2013) 6(10):1140–9. doi: 10.1158/1940-6207.Capr-13-0216
25. Xu SL, Zeng DZ, Dong WG, Ding YQ, Rao J, Duan JJ, et al. Distinct Patterns of ALDH1A1 Expression Predict Metastasis and Poor Outcome of Colorectal Carcinoma. Int J Clin Exp Pathol (2014) 7(6):2976–86.
26. Cao YT, Li JH, Wang YT, Fu YW, Xu J. Serum ALDH1A1 is a Tumor Marker for the Diagnosis of non-Small Cell Lung Cancer. Tumori (2014) 100(2):214–8. doi: 10.1700/1491.16419
27. Xing Y, Luo DY, Long MY, Zeng SL, Li HH. High ALDH1A1 Expression Correlates With Poor Survival in Papillary Thyroid Carcinoma. World J Surg Oncol (2014), 12(29). doi: 10.1186/1477-7819-12-29
28. Li XS, Xu Q, Fu XY, Luo WS. ALDH1A1 Overexpression is Associated With the Progression and Prognosis in Gastric Cancer. BMC Cancer (2014), 14(705). doi: 10.1186/1471-2407-14-705
29. Duong HQ, Yi YW, Kang HJ, Bae I, Jang YJ, Kwak SJ, et al. Combination of Dasatinib and Gemcitabine Reduces the ALDH1A1 Expression and the Proliferation of Gemcitabine-Resistant Pancreatic Cancer MIA PaCa-2 Cells. Int J Oncol (2014) 44(6):2132–8. doi: 10.3892/ijo.2014.2357
30. Pandrangi SL, Chikati R, Chauhan PS, Kumar CS, Banarji A, Saxena S. Effects of Ellipticine on ALDH1A1-Expressing Breast Cancer Stem Cells–an In Vitro and in Silico Study. Tumour Biol (2014) 35(1):723–37. doi: 10.1007/s13277-013-1099-y
31. Liu Y, Baglia M, Zheng Y, Blot W, Bao PP, Cai H, et al. ALDH1A1 mRNA Expression in Association With Prognosis of Triple-Negative Breast Cancer. Oncotarget (2015) 6(38):41360–9. doi: 10.18632/oncotarget.6023
32. Sjöström M, Hartman L, Honeth G, Grabau D, Malmström P, Hegardt C, et al. Stem Cell Biomarker ALDH1A1 in Breast Cancer Shows an Association With Prognosis and Clinicopathological Variables That is Highly Cut-Off Dependent. J Clin Pathol (2015) 68(12):1012–9. doi: 10.1136/jclinpath-2015-203092
33. Gao F, Zhou B, Xu JC, Gao X, Li SX, Zhu GC, et al. The Role of LGR5 and ALDH1A1 in non-Small Cell Lung Cancer: Cancer Progression and Prognosis. Biochem Biophys Res Commun (2015) 462(2):91–8. doi: 10.1016/j.bbrc.2015.04.029
34. Hoshino Y, Nishida J, Katsuno Y, Koinuma D, Aoki T, Kokudo N, et al. Smad4 Decreases the Population of Pancreatic Cancer-Initiating Cells Through Transcriptional Repression of ALDH1A1. Am J Pathol (2015) 185(5):1457–70. doi: 10.1016/j.ajpath.2015.01.011
35. Condello S, Morgan CA, Nagdas S, Cao L, Turek J, Hurley TD, et al. β-Catenin-Regulated ALDH1A1 is a Target in Ovarian Cancer Spheroids. Oncogene (2015) 34(18):2297–308. doi: 10.1038/onc.2014.178
36. Tanaka K, Tomita H, Hisamatsu K, Nakashima T, Hatano Y, Sasaki Y, et al. ALDH1A1-Overexpressing Cells are Differentiated Cells But Not Cancer Stem or Progenitor Cells in Human Hepatocellular Carcinoma. Oncotarget (2015) 6(28):24722–32. doi: 10.18632/oncotarget.4406
37. Kesharwani RK, Srivastava V, Singh P, Rizvi SI, Adeppa K, Misra K. A Novel Approach for Overcoming Drug Resistance in Breast Cancer Chemotherapy by Targeting New Synthetic Curcumin Analogues Against Aldehyde Dehydrogenase 1 (ALDH1A1) and Glycogen Synthase Kinase-3 Beta (GSK-3beta). Appl Biochem Biotechnol (2015) 176(7):1996–2017. doi: 10.1007/s12010-015-1696-x
38. Yassin Fel Z. Aldehyde Dehyderogenase (ALDH1A1) Delineating the Normal and Cancer Stem Cells in Spectral Lung Lesions: An Immunohistochemical Appraisal. Pathol Res Pract (2016) 212(5):398–409. doi: 10.1016/j.prp.2016.02.011
39. Erfani E, Roudi R, Rakhshan A, Sabet MN, Shariftabrizi A, Madjd Z. Comparative Expression Analysis of Putative Cancer Stem Cell Markers CD44 and ALDH1A1 in Various Skin Cancer Subtypes. Int J Biol Markers (2016) 31(1):e53-61. doi: 10.5301/jbm.5000165
40. Wang L, Lin D, Fu Y, Lai M. Nuclear Aldehyde Dehydrogenase 1A1 (ALDH1A1) Expression is a Favorable Prognostic Indicator in Colorectal Carcinoma. Pathol Res Pract (2016) 212(9):791–9. doi: 10.1016/j.prp.2016.06.009
41. Kim JC, Ha YJ, Tak KH, Roh SA, Kim CW, Kim TW, et al. Complex Behavior of ALDH1A1 and IGFBP1 in Liver Metastasis From a Colorectal Cancer. PloS One (2016) 11(5):e0155160. doi: 10.1371/journal.pone.0155160
42. Ma Y, Li M, Si J, Xiong Y, Lu F, Zhang J, et al. Blockade of Notch3 Inhibits the Stem-Like Property and is Associated With ALDH1A1 and CD44 via Autophagy in non-Small Lung Cancer. Int J Oncol (2016) 48(6):2349–58. doi: 10.3892/ijo.2016.3464
43. Yokoyama Y, Zhu H, Lee JH, Kossenkov AV, Wu SY, Wickramasinghe JM, et al. BET Inhibitors Suppress ALDH Activity by Targeting ALDH1A1 Super-Enhancer in Ovarian Cancer. Cancer Res (2016) 76(21):6320–30. doi: 10.1158/0008-5472.Can-16-0854
44. Januchowski R, Wojtowicz K, Sterzyska K, Sosiska P, Andrzejewska M, Zawierucha P, et al. Inhibition of ALDH1A1 Activity Decreases Expression of Drug Transporters and Reduces Chemotherapy Resistance in Ovarian Cancer Cell Lines. Int J Biochem Cell Biol (2016) 78:(248–59). doi: 10.1016/j.biocel.2016.07.017
45. Croker AK, Rodriguez-Torres M, Xia Y, Pardhan S, Leong HS, Lewis JD, et al. Differential Functional Roles of ALDH1A1 and ALDH1A3 in Mediating Metastatic Behavior and Therapy Resistance of Human Breast Cancer Cells. Int J Mol Sci (2017) 18(10):2039. doi: 10.3390/ijms18102039
46. Kalantari E, Saadi FH, Asgari M, Shariftabrizi A, Roudi R, Madjd Z. Increased Expression of ALDH1A1 in Prostate Cancer is Correlated With Tumor Aggressiveness: A Tissue Microarray Study of Iranian Patients. Appl Immunohistochem Mol Morphol (2017) 25(8):592–8. doi: 10.1097/pai.0000000000000343
47. Sun L, Cao J, Guo C, Burnett J, Yang Z, Ran Y, et al. Associations of Carboxypeptidase 4 With ALDH1A1 Expression and Their Prognostic Value in Esophageal Squamous Cell Carcinoma. Esophagus (2017) 30(6):1–5. doi: 10.1093/dote/dox011
48. Lu Z, Zhou C, Hu J, Xiong L, Cong Z, Shen Y. DKK1 Maintained Cancer Stem-Like Properties of Esophageal Carcinoma Cells via ALDH1A1/SOX2 Axis. Int J Clin Exp Pathol (2017) 10(9):9489–95.
49. Yu J, Alharbi A, Shan H, Hao Y, Snetsinger B, Rauh MJ, et al. TAZ Induces Lung Cancer Stem Cell Properties and Tumorigenesis by Up-Regulating ALDH1A1. Oncotarget (2017) 8(24):38426–43. doi: 10.18632/oncotarget.16430
50. Wang W, Yi M, Chen S, Li J, Zhang H, Xiong W, et al. NOR1 Suppresses Cancer Stem-Like Cells Properties of Tumor Cells via the Inhibition of the AKT-GSK-3beta-Wnt/beta-Catenin-ALDH1A1 Signal Circuit. J Cell Physiol (2017) 232(10):2829–40. doi: 10.1002/jcp.25706
51. Duong HQ, You KS, Oh S, Kwak SJ, Seong YS. Silencing of NRF2 Reduces the Expression of ALDH1A1 and ALDH3A1 and Sensitizes to 5-FU in Pancreatic Cancer Cells. Antioxidants (Basel) (2017) 6(3):52. doi: 10.3390/antiox6030052
52. Allison SE, Chen Y, Petrovic N, Zhang J, Bourget K, Mackenzie PI, et al. Activation of ALDH1A1 in MDA-MB-468 Breast Cancer Cells That Over-Express CYP2J2 Protects Against Paclitaxel-Dependent Cell Death Mediated by Reactive Oxygen Species. Biochem Pharmacol (2017) 143:(79–89). doi: 10.1016/j.bcp.2017.07.020
53. Yang CK, Wang XK, Liao XW, Han CY, Yu TD, Qin W, et al. Aldehyde Dehydrogenase 1 (ALDH1) Isoform Expression and Potential Clinical Implications in Hepatocellular Carcinoma. PloS One (2017) 12(8):e0182208. doi: 10.1371/journal.pone.0182208
54. van der Waals LM, Borel Rinkes IHM, Kranenburg O. ALDH1A1 Expression is Associated With Poor Differentiation, ‘Right-Sidedness’ and Poor Survival in Human Colorectal Cancer. PloS One (2018) 13(10):e0205536. doi: 10.1371/journal.pone.0205536
55. Yang W, Wang Y, Wang W, Chen Z, Bai G. Expression of Aldehyde Dehydrogenase 1a1 (ALDH1A1) as a Prognostic Biomarker in Colorectal Cancer Using Immunohistochemistry. Med Sci Monit (2018) 24:(2864–72). doi: 10.12659/msm.910109
56. Xia Z, Baumgartner KB, Baumgartner RN, Boone SD, Hines LM, John EM, et al. Associations Between ALDH1A1 Polymorphisms, Alcohol Consumption, and Mortality Among Hispanic and non-Hispanic White Women Diagnosed With Breast Cancer: The Breast Cancer Health Disparities Study. Breast Cancer Res Treat (2018) 168(2):443–55. doi: 10.1007/s10549-017-4600-2
57. Tulake W, Yuemaier R, Sheng L, Ru M, Lidifu D, Abudula A. Upregulation of Stem Cell Markers ALDH1A1 and OCT4 as Potential Biomarkers for the Early Detection of Cervical Carcinoma. Oncol Lett (2018) 16(5):5525–34. doi: 10.3892/ol.2018.9381
58. Ye Y, Zhang S, Chen Y, Wang X, Wang P. High ALDH1A1 Expression Indicates a Poor Prognosis in Gastric Neuroendocrine Carcinoma. Pathol Res Pract (2018) 214(2):268–72. doi: 10.1016/j.prp.2017.10.015
59. Wu V, Auchman M, Mollica PA, Sachs PC, Bruno RD. ALDH1A1 Positive Cells are a Unique Component of the Tonsillar Crypt Niche and are Lost Along With NGFR Positive Stem Cells During Tumourigenesis. Pathology (2018) 50(5):524–9. doi: 10.1016/j.pathol.2018.03.002
60. Ciccone V, Terzuoli E, Donnini S, Giachetti A, Morbidelli L, Ziche M. Stemness Marker ALDH1A1 Promotes Tumor Angiogenesis via Retinoic Acid/HIF-1alpha/VEGF Signalling in MCF-7 Breast Cancer Cells. J Exp Clin Cancer Res (2018) 37(1):311. doi: 10.1186/s13046-018-0975-0
61. Cui T, Srivastava AK, Han C, Wu D, Wani N, Liu L, et al. DDB2 Represses Ovarian Cancer Cell Dedifferentiation by Suppressing ALDH1A1. Cell Death Dis (2018) 9(5):561. doi: 10.1038/s41419-018-0585-y
62. Zhao AY, Dai YJ, Lian JF, Huang Y, Lin JG, Dai YB, et al. YAP Regulates ALDH1A1 Expression and Stem Cell Property of Bladder Cancer Cells. Onco Targets Ther (2018) 11:(6657–63). doi: 10.2147/ott.S170858
63. Oria VO, Bronsert P, Thomsen AR, Foll MC, Zamboglou C, Hannibal L, et al. Proteome Profiling of Primary Pancreatic Ductal Adenocarcinomas Undergoing Additive Chemoradiation Link ALDH1A1 to Early Local Recurrence and Chemoradiation Resistance. Transl Oncol (2018) 11(6):1307–22. doi: 10.1016/j.tranon.2018.08.001
64. Wang Q, Jiang J, Ying G, Xie XQ, Zhang X, Xu W, et al. Tamoxifen Enhances Stemness and Promotes Metastasis of ERalpha36(+) Breast Cancer by Upregulating ALDH1A1 in Cancer Cells. Cell Res (2018) 28(3):336–58. doi: 10.1038/cr.2018.15
65. Kalra S, Kaur RP, Ludhiadch A, Shafi G, Vashista R, Kumar R, et al. Association of CYP2C19*2 and ALDH1A1*1/*2 Variants With Disease Outcome in Breast Cancer Patients: Results of a Global Screening Array. Eur J Clin Pharmacol (2018) 74(10):1291–8. doi: 10.1007/s00228-018-2505-6
66. Roy M, Connor J, Al-Niaimi A, Rose SL, Mahajan A. Aldehyde Dehydrogenase 1A1 (ALDH1A1) Expression by Immunohistochemistry is Associated With Chemo-Refractoriness in Patients With High-Grade Ovarian Serous Carcinoma. Hum Pathol (2018) 73:(1–6). doi: 10.1016/j.humpath.2017.06.025
67. Wang R, Yang L, Li S, Ye D, Yang L, Liu Q, et al. Quercetin Inhibits Breast Cancer Stem Cells via Downregulation of Aldehyde Dehydrogenase 1a1 (ALDH1A1), Chemokine Receptor Type 4 (CXCR4), Mucin 1 (MUC1), and Epithelial Cell Adhesion Molecule (EpCAM). Med Sci Monit (2018) 24:(412–20). doi: 10.12659/msm.908022
68. Kwiatkowska-Borowczyk E, Czerwińska P, Mackiewicz J, Gryska K, Kazimierczak U, Tomela K, et al. Whole Cell Melanoma Vaccine Genetically Modified to Stem Cells Like Phenotype Generates Specific Immune Responses to ALDH1A1 and Long-Term Survival in Advanced Melanoma Patients. Oncoimmunology (2018) 7(11):e1509821. doi: 10.1080/2162402x.2018.1509821
69. Liu WT, Liu WB, Gao M, Zhang YY, Gu KS. Expression of ALDH1A1 and CD133 is Associated With the Prognosis and Effect of Different Chemotherapeutic Regimens in Gastric Cancer. Oncol Lett (2019) 18(5):4573–82. doi: 10.3892/ol.2019.10798
70. Wanandi SI, Syahrani RA, Arumsari S, Wideani G, Hardiany NS. Profiling of Gene Expression Associated With Stemness and Aggressiveness of ALDH1A1-Expressing Human Breast Cancer Cells. Malays J Med Sci (2019) 26(5):38–52. doi: 10.21315/mjms2019.26.5.4
71. Gong X, Xu B, Zi L, Chen X. miR-625 Reverses Multidrug Resistance in Gastric Cancer Cells by Directly Targeting ALDH1A1. Cancer Manag Res (2019) 11:(6615–24). doi: 10.2147/cmar.S208708
72. Swierczewska M, Sterzynska K, Wojtowicz K, Kazmierczak D, Izycki D, Nowicki M, et al. PTPRK Expression Is Downregulated in Drug Resistant Ovarian Cancer Cell Lines, and Especially in ALDH1A1 Positive CSCs-Like Populations. Int J Mol Sci (2019) 20(8):2053. doi: 10.3390/ijms20082053
73. Wang Y, Wang CH, Zhang YF, Zhu L, Lei HM, Tang YB. UPLC-MS-Based Metabolomics Reveals Metabolic Dysregulation in ALDH1A1-Overexpressed Lung Adenocarcinoma Cells. Metabolomics (2019) 15(4):52. doi: 10.1007/s11306-019-1514-5
74. Charkoftaki G, Thompson DC, Golla JP, Garcia-Milian R, Lam TT, Engel J, et al. Integrated Multi-Omics Approach Reveals a Role of ALDH1A1 in Lipid Metabolism in Human Colon Cancer Cells. Chem Biol Interact (2019) 304:(88–96). doi: 10.1016/j.cbi.2019.02.030
75. Nwani NG, Condello S, Wang Y, Swetzig WM, Barber E, Hurley T, et al. A Novel ALDH1A1 Inhibitor Targets Cells With Stem Cell Characteristics in Ovarian Cancer. Cancers (Basel) (2019) 11(4):502. doi: 10.3390/cancers11040502
76. Althobiti M, El Ansari R, Aleskandarany M, Joseph C, Toss MS, Green AR, et al. The Prognostic Significance of ALDH1A1 Expression in Early Invasive Breast Cancer. Histopathology (2020) 77(3):437–48. doi: 10.1111/his.14129
77. Nagare RP, Sneha S, Krishnapriya S, Ramachandran B, Murhekar K, Vasudevan S, et al. ALDH1A1+ Ovarian Cancer Stem Cells Co-Expressing Surface Markers CD24, EPHA1 and CD9 Form Tumours In Vivo. Exp Cell Res (2020) 392(1):112009. doi: 10.1016/j.yexcr.2020.112009
78. Szafarowski T, Sierdzinski J, Ludwig N, Gluszko A, Filipowska A, Szczepanski MJ. Assessment of Cancer Stem Cell Marker Expression in Primary Head and Neck Squamous Cell Carcinoma Shows Prognostic Value for Aldehyde Dehydrogenase (ALDH1A1). Eur J Pharmacol (2020) 867:(172837). doi: 10.1016/j.ejphar.2019.172837
79. Yoshino J, Akiyama Y, Shimada S, Ogura T, Ogawa K, Ono H, et al. Loss of ARID1A Induces a Stemness Gene ALDH1A1 Expression With Histone Acetylation in the Malignant Subtype of Cholangiocarcinoma. Carcinogenesis (2020) 41(6):734–42. doi: 10.1093/carcin/bgz179
80. Namekawa T, Ikeda K, Horie-Inoue K, Suzuki T, Okamoto K, Ichikawa T, et al. ALDH1A1 in Patient-Derived Bladder Cancer Spheroids Activates Retinoic Acid Signaling Leading to TUBB3 Overexpression and Tumor Progression. Int J Cancer (2020) 146(4):1099–113. doi: 10.1002/ijc.32505
81. Jiang Y, Song H, Jiang L, Qiao Y, Yang D, Wang D, et al. Silybin Prevents Prostate Cancer by Inhibited the ALDH1A1 Expression in the Retinol Metabolism Pathway. Front Cell Dev Biol (2020) 8:574394. doi: 10.3389/fcell.2020.574394
82. Elcheva IA, Wood T, Chiarolanzio K, Chim B, Wong M, Singh V, et al. RNA-Binding Protein IGF2BP1 Maintains Leukemia Stem Cell Properties by Regulating HOXB4, MYB, and ALDH1A1. Leukemia (2020) 34(5):1354–63. doi: 10.1038/s41375-019-0656-9
83. Wang W, He S, Zhang R, Peng J, Guo D, Zhang J, et al. ALDH1A1 Maintains the Cancer Stem-Like Cells Properties of Esophageal Squamous Cell Carcinoma by Activating the AKT Signal Pathway and Interacting With Beta-Catenin. BioMed Pharmacother (2020) 125:(109940). doi: 10.1016/j.biopha.2020.109940
84. Tieng FYF, Abu N, Sukor S, Mohd Azman ZA, Mahamad Nadzir N, Lee LH, et al. L1CAM, CA9, KLK6, HPN, and ALDH1A1 as Potential Serum Markers in Primary and Metastatic Colorectal Cancer Screening. Diagnostics (Basel) (2020) 10(7):444. doi: 10.3390/diagnostics10070444
85. Kaipio K, Chen P, Roering P, Huhtinen K, Mikkonen P, Ostling P, et al. ALDH1A1-Related Stemness in High-Grade Serous Ovarian Cancer is a Negative Prognostic Indicator But Potentially Targetable by EGFR/mTOR-PI3K/aurora Kinase Inhibitors. J Pathol (2020) 250(2):159–69. doi: 10.1002/path.5356
86. Liu L, Cai S, Han C, Banerjee A, Wu D, Cui T, et al. ALDH1A1 Contributes to PARP Inhibitor Resistance via Enhancing DNA Repair in BRCA2(-/-) Ovarian Cancer Cells. Mol Cancer Ther (2020) 19(1):199–210. doi: 10.1158/1535-7163.Mct-19-0242
87. Gyan E, Green A, Ahenkorah-Fondjo L, Jackson A, Toss MS, Akakpo PK, et al. The Role of ALDH1A1 in Contributing to Breast Tumour Aggressiveness: A Study Conducted in an African Population. Ann Diagn Pathol (2021) 51:(151696). doi: 10.1016/j.anndiagpath.2020.151696
88. Wang W, He S, Zhang R, Peng J, Guo D, Zhang J, et al. ALDH1A1 Maintains the Cancer Stem-Like Cells Properties of Esophageal Squamous Cell Carcinoma by Activating the AKT Signal Pathway and Interacting With β-Catenin. BioMed Pharmacother (2020) 125:(109940). doi: 10.1016/j.biopha.2020.109940
89. Liu C, Qiang J, Deng Q, Xia J, Deng L, Zhou L, et al. ALDH1A1 Activity in Tumor-Initiating Cells Remodels Myeloid-Derived Suppressor Cells to Promote Breast Cancer Progression. Cancer Res (2021) 81(23):5919–34. doi: 10.1158/0008-5472.Can-21-1337
90. Narendra G, Raju B, Verma H, Silakari O. Identification of Potential Genes Associated With ALDH1A1 Overexpression and Cyclophosphamide Resistance in Chronic Myelogenous Leukemia Using Network Analysis. Med Oncol (2021) 38(10):123. doi: 10.1007/s12032-021-01569-9
91. Narendra G, Raju B, Verma H, Sapra B, Silakari O. Multiple Machine Learning Models Combined With Virtual Screening and Molecular Docking to Identify Selective Human ALDH1A1 Inhibitors. J Mol Graph Model (2021) 107:(107950). doi: 10.1016/j.jmgm.2021.107950
92. Yamashita N, So T, Miyata T, Yoshimatsu T, Nakano R, Oyama T, et al. Triple-Negative Expression (ALDH1A1-/CD133-/Mutant P53-) Cases in Lung Adenocarcinoma had a Good Prognosis. Sci Rep (2022) 12(1):1473. doi: 10.1038/s41598-022-05176-0
93. Nachiyappan A, Soon JLJ, Lim HJ, Lee VK, Taneja R. EHMT1 Promotes Tumor Progression and Maintains Stemness by Regulating ALDH1A1 Expression in Alveolar Rhabdomyosarcoma. J Pathol (2022) 256(3):349–62. doi: 10.1002/path.5848
94. Zhou J, Sun C, Yang L, Wang J, Jn-Simon N, Zhou C, et al. Liver Regeneration and Ethanol Detoxification: A New Link in YAP Regulation of ALDH1A1 During Alcohol-Related Hepatocyte Damage. FASEB J (2022) 36(4):e22224. doi: 10.1096/fj.20211686R
95. Okamoto A, Funakoshi Y, Oe M, Takai R, Suto H, Nagatani Y, et al. Identification of Breast Cancer Stem Cells Using a Newly Developed Long-Acting Fluorescence Probe, C5S-A, Targeting ALDH1A1. Anticancer Res (2022) 42(3):1199–205. doi: 10.21873/anticanres.15586
96. Peng W, Zhao ZZ, Jiao L, Wu T, Chen H, Zhang CY, et al. Prospective Study of ALDH1A1 Gene Polymorphisms Associated With Antituberculosis Drug-Induced Liver Injury in Western Chinese Han Population. Microbiol Immunol (2021) 65(4):143–53. doi: 10.1111/1348-0421.12877
97. Yang L, Shi P, Zhao G, Xu J, Peng W, Zhang J, et al. Targeting Cancer Stem Cell Pathways for Cancer Therapy. Signal Transduct Target Ther (2020) 5(1):8. doi: 10.1038/s41392-020-0110-5
98. Singh V, Ram M, Kumar R, Prasad R, Roy BK, Singh KK. Phosphorylation: Implications in Cancer. Protein J (2017) 36(1):1–6. doi: 10.1007/s10930-017-9696-z
99. Shvedunova M, Akhtar A. Modulation of Cellular Processes by Histone and non-Histone Protein Acetylation. Nat Rev Mol Cell Biol (2022) 23(5):329–49. doi: 10.1038/s41580-021-00441-y
100. Chen L, Heikkinen L, Wang C, Yang Y, Sun H, Wong G. Trends in the Development of miRNA Bioinformatics Tools. Brief Bioinform (2019) 20(5):1836–52. doi: 10.1093/bib/bby054
101. Condello S, Morgan CA, Nagdas S, Cao L, Turek J, Hurley TD, et al. Beta-Catenin-Regulated ALDH1A1 is a Target in Ovarian Cancer Spheroids. Oncogene (2015) 34(18):2297–308. doi: 10.1038/onc.2014.178
Keywords: aldehyde dehydrogenases, cancer stem cell, carcinogenesis, drug resistance, hepatocellular carcinoma
Citation: Yue H, Hu Z, Hu R, Guo Z, Zheng Y, Wang Y and Zhou Y (2022) ALDH1A1 in Cancers: Bidirectional Function, Drug Resistance, and Regulatory Mechanism. Front. Oncol. 12:918778. doi: 10.3389/fonc.2022.918778
Received: 12 April 2022; Accepted: 17 May 2022;
Published: 22 June 2022.
Edited by:
Ira Ida Skvortsova, Innsbruck Medical University, AustriaReviewed by:
Junlong Zhuang, Nanjing Drum Tower Hospital, ChinaCopyright © 2022 Yue, Hu, Hu, Guo, Zheng, Wang and Zhou. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Yongning Zhou, emhvdXluQGx6dS5lZHUuY24=; Yuping Wang, d2FuZ3l1cGluZ0BsenUuZWR1LmNu
†These authors have contributed equally to this work
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.