
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Oncol., 21 September 2022
Sec. Molecular and Cellular Oncology
Volume 12 - 2022 | https://doi.org/10.3389/fonc.2022.1004212
This article is part of the Research TopiclncRNAs in Cancer Metastasis and Therapy Resistance Volume IIView all 19 articles
 Zhengwei Song1†Xiaoguang Wang1†Fei Chen1Qiuli Chen2
Zhengwei Song1†Xiaoguang Wang1†Fei Chen1Qiuli Chen2 Wenjun Liu2Xiaodan Yang1
Wenjun Liu2Xiaodan Yang1 Xun Zhu1Xiaorong Liu1*
Xun Zhu1Xiaorong Liu1* Peter Wang2*
Peter Wang2*Pancreatic cancer is the fourth leading cause of cancer death in the United States. The main methods of treating pancreatic cancer are surgery and chemotherapy, but the treatment efficacy is low with a poor prognosis. Immunotherapy represented by PD-1/PD-L1 has brought a milestone progress in the treatment of pancreatic cancer. However, the unique tumor microenvironment of pancreatic cancer presents challenges for immunotherapy. In addition, m6A is a common RNA modification and a potential molecular target in tumor therapy. The expression pattern of m6A in pancreatic cancer is still unclear. LncRNAs also play an essential role in pancreatic cancer development and treatment. In this study, we found that some m6A regulators were significantly elevated in pancreatic cancer and associated with the expression of PD-1/PD-L1. Moreover, we observed that METTL3 can increase the expression of PD-L1. Notably, METTL3 positively regulates the expression of lncRNA MALAT1 in pancreatic cancer cells. Strikingly, lncRNA MALAT1 increased the expression of PD-L1 in pancreatic cancer cells. This finding indicated that METTL3 regulated the expression of PD-L1 possibly via targeting lncRNA MALAT1 in pancreatic cancer cells. Lastly, MALAT1 governed the viability of pancreatic cancer cells. Taken together, lncRNA MALAT1 is involved in METTL3-mediated promotion of PD-L1 expression in pancreatic cancer.
Pancreatic ductal adenocarcinoma (PDAC) is one of the common malignant tumors of the pancreas and more than 90% of pancreatic cancer are exocrine PDAC (1). According to an epidemiology report, the five-year survival rate of pancreatic cancer after diagnosis is about 10%, and it is estimated that the pancreatic cancer will surpass breast cancer as the third leading cause of death (2). Cancer statistics suggest that there were 32,970 new cases of pancreatic cancer in men and 29,240 new cases of pancreatic cancer in women in the United States (3). The number of people who died from pancreatic cancer was 49,830 for men and 25,970 for women, respectively (3). Surgery and chemotherapy remain the mainstays of treatment for pancreatic adenocarcinoma (4). However, pancreatic cancer is aggressive with no obvious symptoms in the early stage. Most PDAC patients are late to be treated when diagnosed, and less than 20% of the patients are eligible for surgery (5). For unresectable patients, chemotherapy is the main treatment, but its efficacy is not ideal, and the median OS (Overall survival) is basically less than 1 year (6). Recently, immunotherapy, represented by immune checkpoint inhibitors (ICIs), has brought milestone progress to tumor treatment. However, ICIs are almost completely wiped out in pancreatic cancer, and most of them have failed in phase I and II clinical trials (7).
However, studies have shown that pancreatic cancer has a special tumor microenvironment (TME), which brings challenges to immunotherapy and deserves further study (8, 9). Tumor immune microenvironment (TIME) refers to the internal environment in which tumor cells are generated and live, which includes not only tumor cells themselves but also fibroblasts, immune and inflammatory cells, and glue cells that are closely related to tumor cells. However, compared with other cancers, pancreatic cancer has a unique TIME, which presents challenges for immunotherapy, and this may be one of the reasons why programmed death (PD-1)/PD ligand 1 (PD-L1) therapy is not highly sensitive to pancreatic cancer. PD-1, a 288 amino acid type 1 transmembrane protein, is often expressed on the surfaces of several immune cell types, while PD-L1, a 290 amino acid type 1 transmembrane protein, is expressed on hematopoietic cells (10, 11). Pancreatic tumor cells promote the activation of peripheral stromal cells and immunosuppressive cells, including regulatory T cells (Tregs), bone-marrow derived inhibitory cells (MDSCs), and tumor-associated macrophages (TAMs). At the same time, they secrete a series of cytokines and chemokines that cause these cells to flock to the tumor site. On the other hand, activated stromal cells generate a large amount of extracellular matrix that forms a fibrous “barrier” around pancreatic tumor cells, preventing effector cells (T and NK cells) from infiltrating into the tumor which allows tumor cells to evade immune surveillance. Activated immunosuppressive cells secrete immunosuppressive factors and express ligands (e.g., PD-L1 and B7-1/2), forming an immunosuppressive microenvironment. This plays an important role in the occurrence, development, invasion, metastasis and drug resistance of pancreatic cancer (12). The antitumor immune response is a complex, multistep process (13). Therefore, the mechanism of TIME should be further studied to explore new and potentially beneficial targets to improve the efficacy of pancreatic cancer immunotherapy.
Meanwhile, m6A is the most common RNA modification in eukaryotic RNA and plays an important role in cancer progression (14). It has been demonstrated that m6A modification is a dynamic and reversible process, which is composed of methyltransferase complex (Writers), demethylase (Erasers) and function managers (Readers) (15). It is believed that the function of m6A writers is stability of mRNA (2). A recent study show that there are 28 m6A regulators, including METTL3, METTL14, METTL16, WTAP, RBM15, RBM15B, ZC3H13, VIRMA, CBLL1, ZCCHC4, LRPPRC, ELAVL1, YTHDC1, YTHDC2, YTHDF1, YTHDF2, YTHDF3, HNRNPC, HNRNPA2B1, EIF3A, EIF3H, IGF2BP1, IGF2BP2, IGF2BP3, CBLL1, PRRC2A, FTO, ALKBH5 (16). m6A modification plays a role in pre-mRNA splicing, 3’-end processing, nuclear output, translation regulation, mRNA decay and miRNA processing, and its dynamic reversible changes control and determine cell growth and differentiation, suggesting that abnormalities of m6A and modified proteins may also produce pathological effects in the occurrence and progression of tumors (17). Besides, as the most common modification in mRNA, m6A links epigenomics with tumorigenesis and development, and affects the processes of tumor stem cell self-renewal and differentiation, proliferation and apoptosis, invasion and metastasis, drug resistance, and immunosuppression. Therefore, m6A is involved in m6A-modified key proteins that are expected to be potential molecular targets for cancer diagnosis and treatment and drug development. For instance, a study examining DNA and RNA methylation status in circulating tumor cells (CTCs) from lung cancer patients demonstrated for the first time elevated m6A modification levels in CTCs from lung cancer patients (18).
LncRNA, one type of noncoding RNA, participates in tumorigenesis and progression (19–22). LncRNA MALAT1 has been reported to regulate pancreatic oncogenesis. The expression of MALAT1 was highly elevated in PDAC compared with the adjacent normal specimens (23). MALAT1 expression was linked to invasion, tumor stage, poor survival, tumor size and metastasis in PDAC patients (23, 24). Moreover, MALAT1 enhanced invasion, migration and viability of pancreatic cancer cells via reduction of EMT and cancer stem cells as well as induction of apoptosis and cell cycle arrest (25). The role of MALAT1 in PDAC development is not fully elucidated. The expression pattern and pathophysiological role of m6A in pancreatic cancer remain unknown. In addition, the association between m6A methylation modulator and PD-L1 remains unexplored. Therefore, the main purpose of this paper aims to analyze the relationship between m6A RNA methylation regulators, PD-L1 and MALAT1 in pancreatic cancer.
The data was collected from The Cancer Genome Atlas (TCGA) database and the basic information including age, gender, grade, stage, T, M and N of 178 samples was obtained. In this study, we collected 24 m6A-related regulators, including writers, erasers and readers. The writers include RBM27, METTL3, WTAP, RBM15, ZC3H7B, CAPRIN1 and METTL14. The erasers include FTO and ALKBH5. The readers include YTHDF1, YTHDF2, YTHDF3, IGF2BP1/2/3, YTHDC1/2, IGF2BPs, KIAA1429 (VIRMA), EIF3A, EIF3H, HNRNPC, HNRNPA2B1, LRPPRC, ELAVL1 and PRRC2A (26). To determine the interaction of these 20 regulators, they were searched in the Genes/Protein database to gain a preliminary understanding of their biological functions (27).
Consensus clustering is a method of providing quantitative evidence for determining the number and members of possible clusters in a data set, such as microarray gene expression (28). This approach is widely used in cancer genomics. In this study, we adopted this method to explore two PDAC clusters and their association with clinicopathological parameters. Gene set enrichment analysis (GSEA) 3.0 was used to predict the underlying downstream pathways of the two clusters (29). Immune score, stromal score, and tumor purity of each sample were calculated by using the ESTIMATE algorithm (30).
Genes with significant (p<0.01) prognosis were screened from DEGs using univariate COX regression analysis, and gene prognosis models were established by lasso-cox regression. The K-M curve was drawn using the R package survival, and the survival difference between different groups was calculated by the log-rank test to draw the K-M curve. The ROC of the model was calculated using the R package time ROC. Finally, the independent prognostic ability of risk scores was tested by univariate and multivariate Cox regression models. Groups will be divided into two groups including high-risk group and low-risk group by evaluating the distribution of clinical case characteristics using the R package “heatmap”. This study adopted Cox regression models to assess whether the risk score combining with other clinical characteristics, could be an independent prognostic factor.
The human pancreatic cancer cell lines, BxPC-3 (with epithelial properties) and PANC-1 (with more mesenchymal properties), were cultured in DMEM medium supplemented with and 10% FBS and 1% penicillin/streptomycin solution. The cells were cultured in an incubator with 5% CO2 at 37°C. Human MALAT1 cDNAs were subcloned into pcDNA3 vector. Human METTL3 cDNAs were subcloned into pLenti-C-mGFP vector. Specific small hairpin RNAs (shRNAs) targeting METTL3 (shMETTL3) or MALAT1 (shMALAT1) and the control shRNA (shNC) were obtained (GenePharma, Shanghai, China).
Total RNA was extracted from pancreatic cancer cells using 1ml TRLzol Reagent. Then, RNA was used for reverse transcription and PCR was performed using SYBR Green Kit Data were analyzed by the ΔΔCt approach. GAPDH was used as the control. MALAT1: FW: GGA TCC TAG ACC AGC ATG CC; RV: AAA GGT TAC CAT AAG TAA GTT CCA GAA AA (31). The detailed method for PCR was described previously (32).
The pancreatic cells were lysed by RIPA buffer and a bicinchoninic acid (BCA) assay was used for detection of protein qualification. After proteins were separated onto SDS-PAGE, the proteins were transferred onto a PVDF membrane. The membrane was incubated with 5% non-fat milk and then incubated with METTL3 (1:1000), PD-L1 (1:1,000) or Tubulin (1:1,000) antibody for overnight at 4°C. The membranes were washed by TBST and then incubated with the secondary antibody for 1 h. Then, ECL method was used to examine the protein expression (33).
The treated pancreatic cancer cells were cultured in 96-well plates for 48 and 72 hours. The viability of pancreatic cancer cells was determined by CCK8 assay as described previously (34). Briefly, 10 μL CCK8 reagent was added to each well and incubated for 2.5-3 hours in a cell culture incubator. The OD450 values were obtained by the microplate reader.
In this study, the Pearson correlation coefficient was widely used to measure the degree of correlation between two variables, and its value was between -1 and 1. To assess the impact of M6A-related risk characteristics on the prognosis of PDAC, we compared the prognostic differences between the high-risk and low-risk groups. Kaplan-Meier plotter was used to analyze the relationship between gene expression profile and survival information in PATIENTS with PDAC. In addition, multivariate Cox regression analysis was performed to determine prognostic factors for PDAC patients. Student t test was used to validate significance between two groups. ANOVE was used to validate significance among three or more groups. P <0.05 is considered statistically significant.
Initially, we analyzed the frequency of mutations in 24 expressed m6A regulators and found that 18 regulators all had a mutation frequency of 1%~2%. Mutations occurred in 7.3% of the 178 samples, the most common mutation being missense mutation (Figure 1A). To further understand the expression of m6A RNA methylation regulator in tumor and normal samples, the heatmap was conducted and the results have revealed that compared with normal samples, there were 24 regulators of m6A, whose expression was relatively higher in cancer tissues (Figure 1B). Besides, the expression difference of m6A RNA methylation regulators between tumor and normal samples was significant since the p-value is less than 0.05 (Figure 1C).
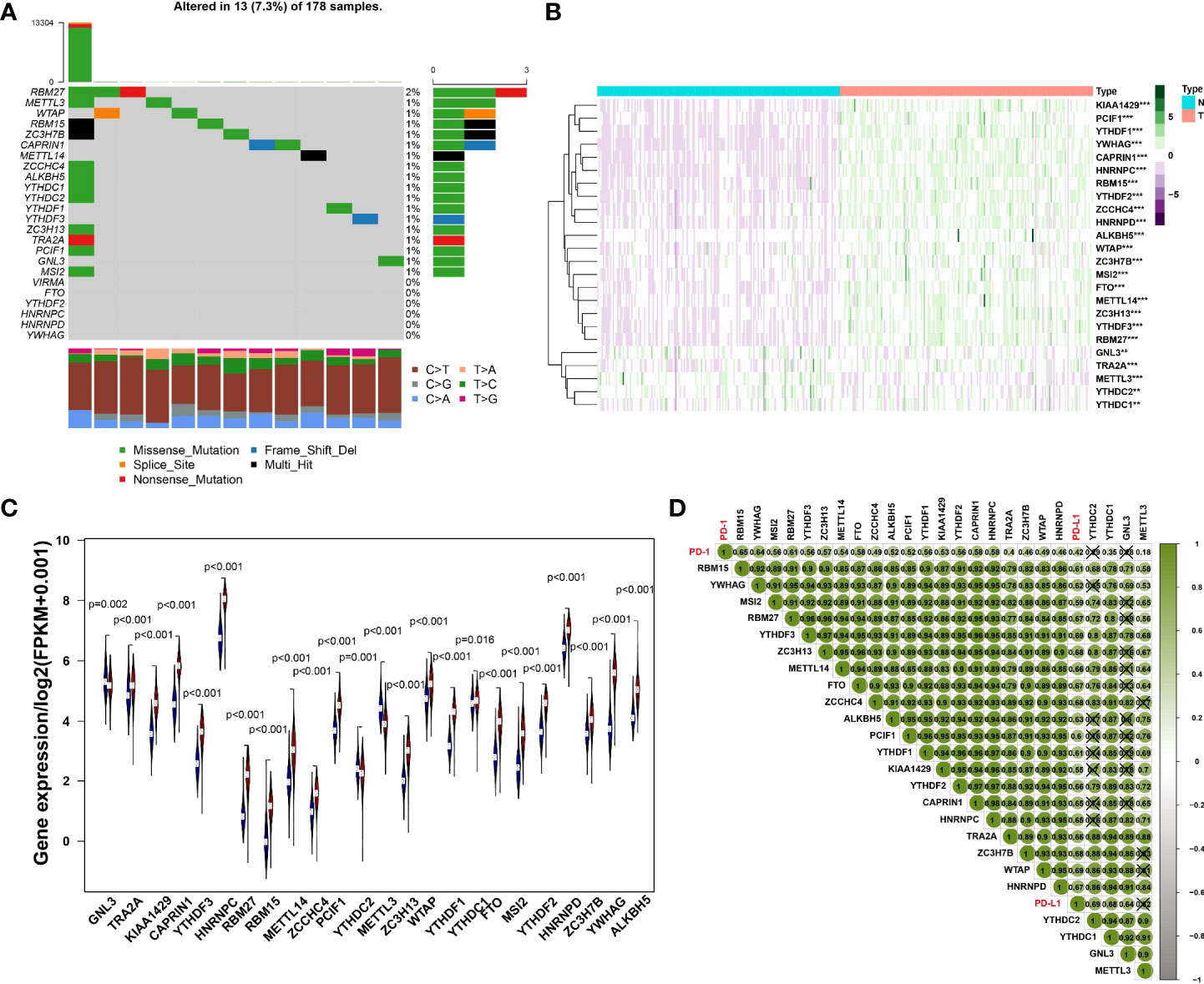
Figure 1 (A) The mutation frequency in each regulator. (B) Heatmap of m6A RNA methylation regulator expression level in each sample. **p<0.01; ***p<0.001. (C) The expression difference of m6A RNA methylation regulator between tumor and normal samples. (D) Correlation among PD-1, PD-L1 and m6A RNA methylation regulators.
The results of Pearson correlation suggested that PD-1 is associated with 20 regulators, including RBM15, YWHAG, MSI2, RBM27, YTHDF3, ZC3H13, METTL14, FTO, ZCCHC4, ALKBH5, PCIF1, YTHDF1, KIAA1429, YTHDF2, CAPRIN1, HNRNPC, TRA2A, ZC3H7B, WTAP, HNRNPD, YTHDC1 and METTL3. The correlation coefficient of PD-1 and RBM15 was the strongest. Similarly, the correlation between PD-L1 and WTAP was the strongest with a correlation coefficient of 0.69, while the relationship between PD-L1 and KIAA1429 was the weakest with a correlation coefficient of 0.55. All regulators of PD-1, PD-L1 and M6A were positively correlated (Figure 1D).
Consensus clustering method was adopted to aggregates data such as transcriptome and proteome profiles. From Figure 2A, it is believed that k = 2 has the optimal clustering stability from k = 2 to 9. Then, the consensus clustering has identified the PDAC cohort of TCGA into two clusters and demonstrated their relationship with clinicopathological parameters (Figure 2A). Subsequently, the heatmap of correlation of m6A RNA methylation regulators with characteristics of PDAC patients and the tracking plot have been concisely displayed (Figures 2B, C). Furthermore, the overall survival (OS) for PDAC patients was analyzed by using Kaplan-Meier curves. It is noteworthy that cluster2 had a significantly higher survival rate than cluster1 (Figure 2D).
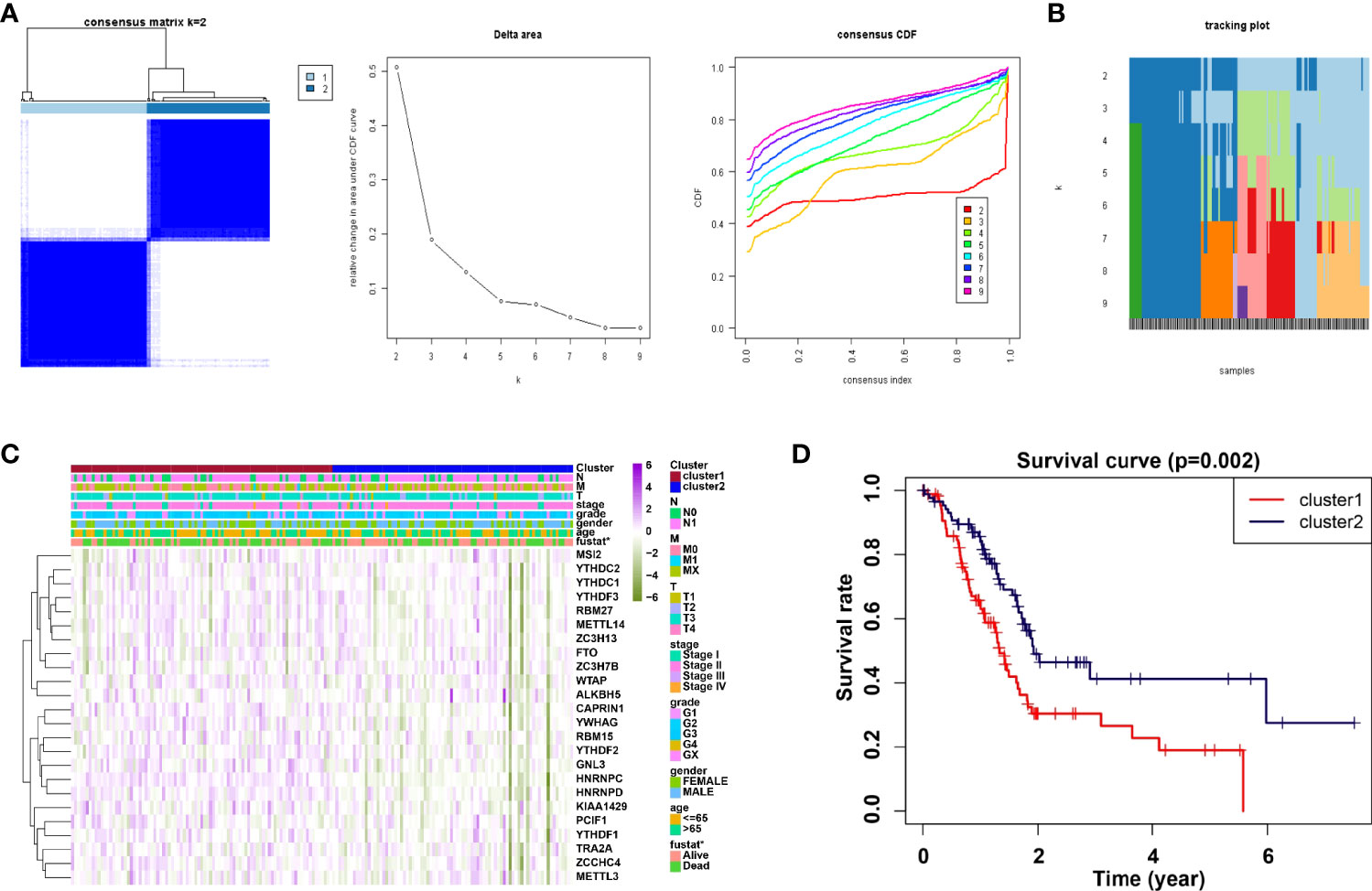
Figure 2 Correlation of consensus clustering for m6A RNA methylation regulators with the characteristics and survival of PDAC patients. (A) Consensus clustering matrix for k=2 (left panel); Consensus clustering cumulative distribution function (CDF) for k=2 to 9 (middle panel); relative change in area under CDF curve for k=2 to 9. (B, C) Heatmap of correlation of m6A RNA methylation regulators with characteristics of PDAC patients. (D) Kaplan-Meier curves of overall survival (OS) for patients.
We analyzed the infiltrating levels of various immune cells in cluster1/2 in PDAC and the results were displayed (Figure 3A). Meanwhile, the graph of estimated proportion of 22 immune cell types in cluster1/2 suggested that the estimated proportion of Macrophages MO of cluster1 was higher than that of Macrophages MO of cluster2, (Figure 3B). In order to further understand the infiltrating levels in cluster1/2 with PDAC, we compared the StromalScore, ImmuneScore and EstimateScore. We found the StromalScore and EstimateScore of cluster2 were higher than that of cluster1, which indicated that cluster2 had a higher degree of immune infiltration than cluster1 (Figures 4A–C). Moreover, cluster1 and cluster2 were involved in the following five signaling pathways: cell cycle, mismatch repair, p53 signaling pathway, RNA degradation and Spliceosome (Figure 4D).
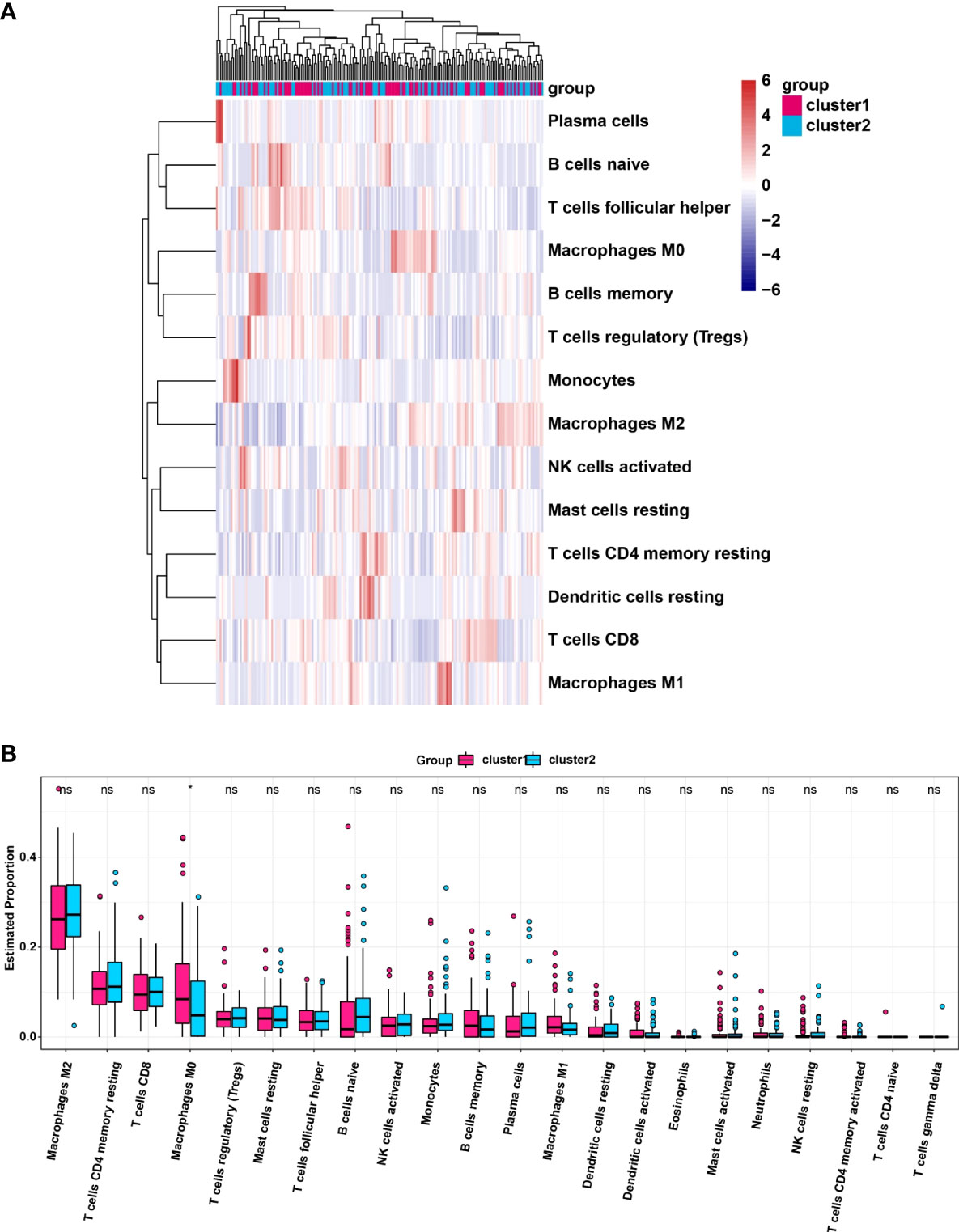
Figure 3 (A) Heatmap of infiltrating levels of various immune cells in cluster1/2 in pancreatic cancer. (B) Estimated proportion of 22 immune cell types in cluster1/2 in pancreatic cancer. *p<0.05; ns, no significance.
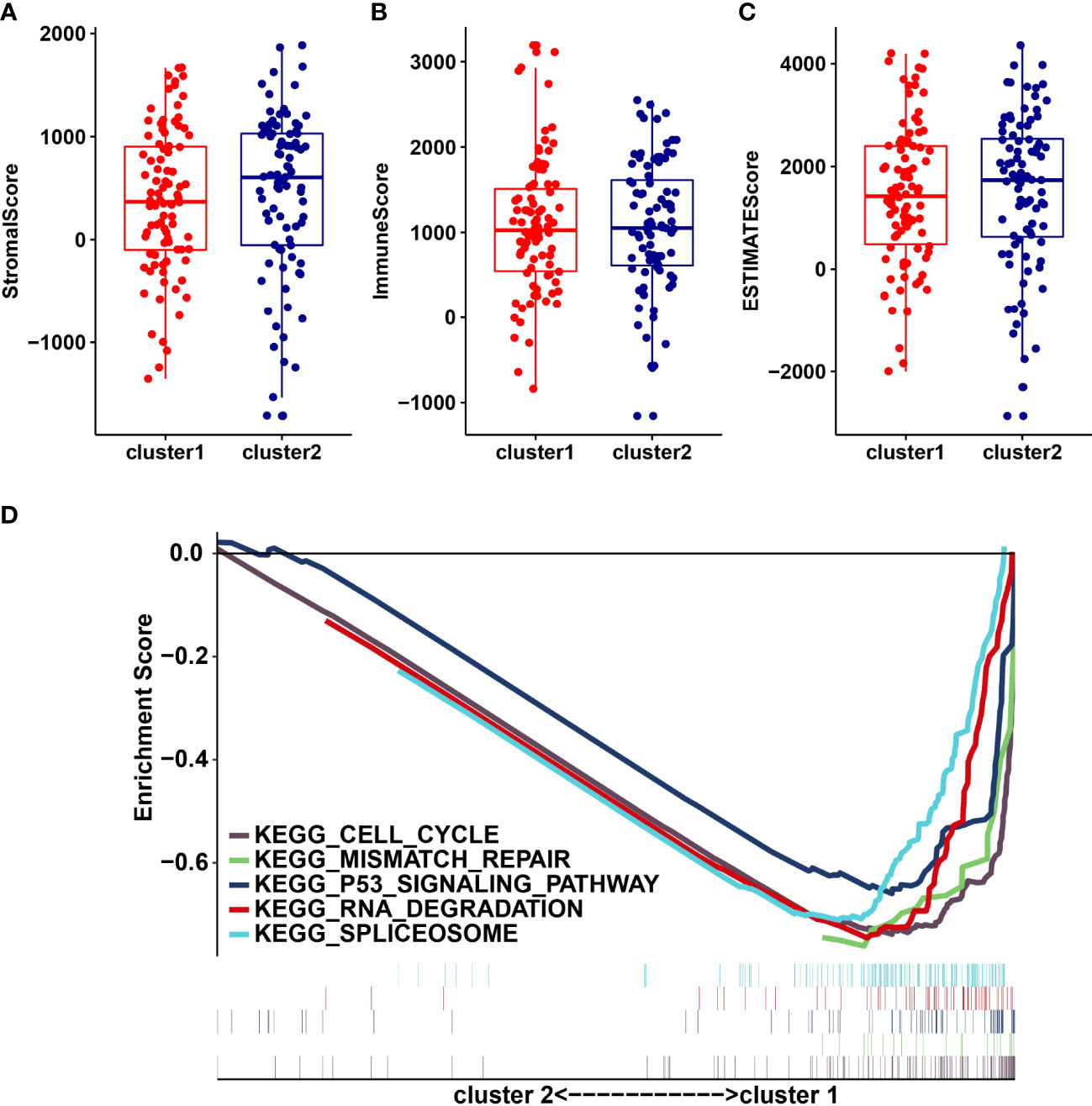
Figure 4 StromalScore (A), ImmunoScore (B), EstinateScore (C) in the cluster1/2 subtypes are illustrated. (D): The signaling pathways are involved in cluster1 and cluster2.
Univariate analysis of 24 m6A RNA methylation regulators was performed to identify genes, which may significantly associate with prognosis. Indeed, the results revealed that GNL3, CAPRIN1, PCIF1, METTL3, YWHAG and ALKBH5 were significantly associated with OS with hazard ratios of 1.807, 2.289, 0.534, 0.647, 1.759 and 0.474, respectively. The 95 percent confidence intervals were (1.128-2.893), (1.315-3.984), (0.325-0.897), (0.443-0.945), (1.068-2.895) and (0.306-0.735), respectively (Figure 5A). Overall, the hazard ratio of GNL3, CAPRIN1 and YWHAG was greater than 1, while the hazard ratio of PCIF1, METTL3 and ALKBH5 was less than 1. Then through the lasso regression algorithm, the coefficient of prognostic genes was identified (Figures 5B, C).
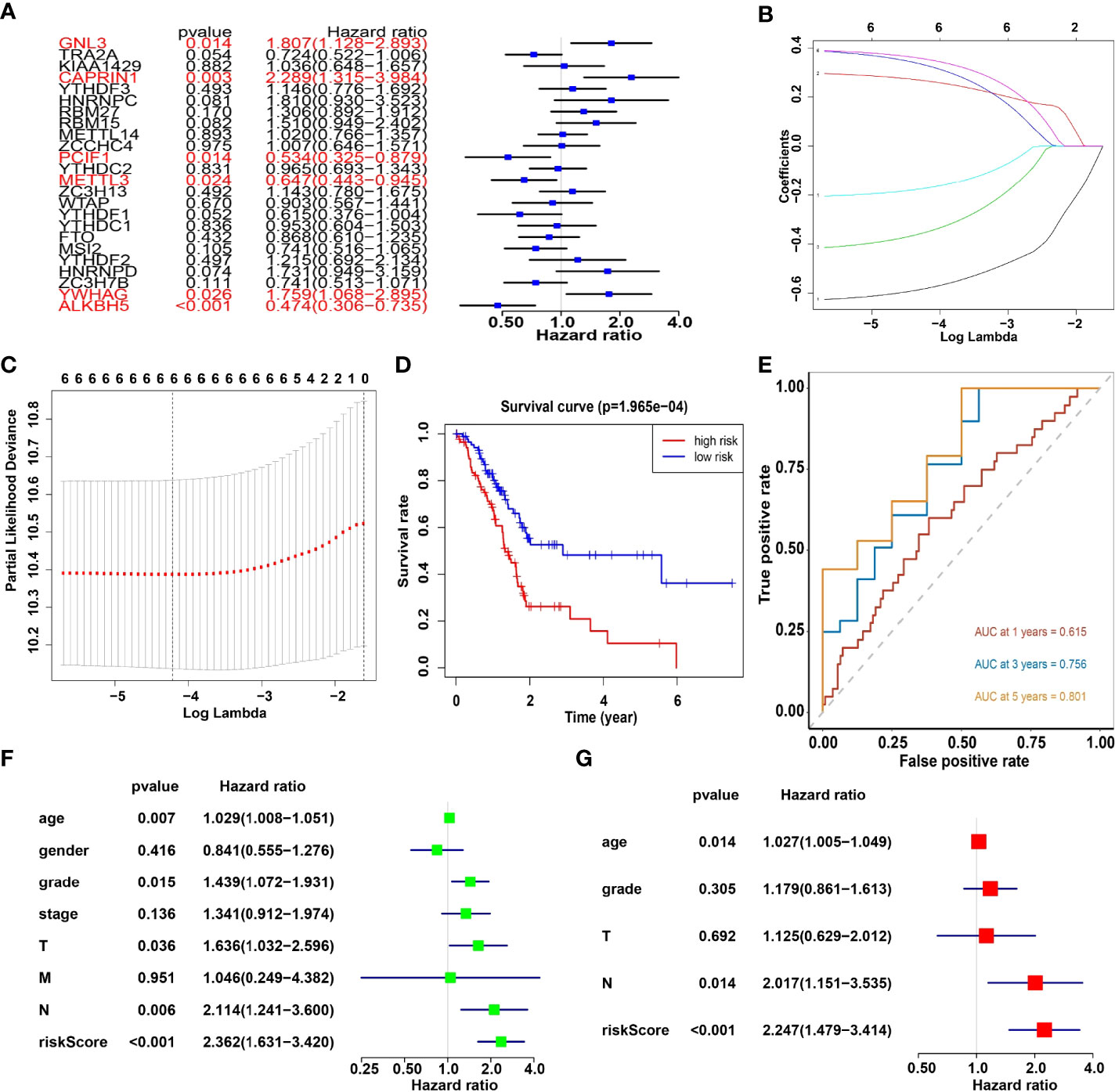
Figure 5 (A) Univariate analysis of 24 regulators. (B, C) LASSO Cox regression algorithm. (D) The Kaplan-Meier curve of high risk and low risk group. (E) Time-dependent ROC curves. (F, G) Univariate and multivariate Cox regression analysis of the risk scores in TCGA.
Moreover, survival analysis showed higher survival rates in the low-risk group than in the high-risk group (Figure 5D). The AUC at 1 years, 3 years and 5 years is 0.615, 0.756 and 0.801 (Figure 5E). Furthermore, to determine whether prognostic marker-based risk scores are independent prognostic indicators for pancreatic cancer patients, univariate and multivariate Cox regression analyses of risk scores were performed. The results proposed that N and risk score were independent prognostic indexes (p-value=0.014, HR=0.017; p-value<0.001, HR=2.247) (Figures 5F, G). The clinical features of PDAC cohort has been displayed in Figure 6A. The riskscore of cluster2 was higher than that of cluster1, and similarly, the riskscore of the low-risk group was higher than that of the high-risk group. Among G1, G2, G3 and G4 groups, G3 group had the highest riskscore, while G4 group had the lowest riskscore (Figure 6B). It is noteworthy that PD-1 and PD-L1 were highly expressed in pancreatic cancer cells compared with normal cells. Compared with cluster2, PD-L1 was highly expressed in cluster1(Figure 6C).
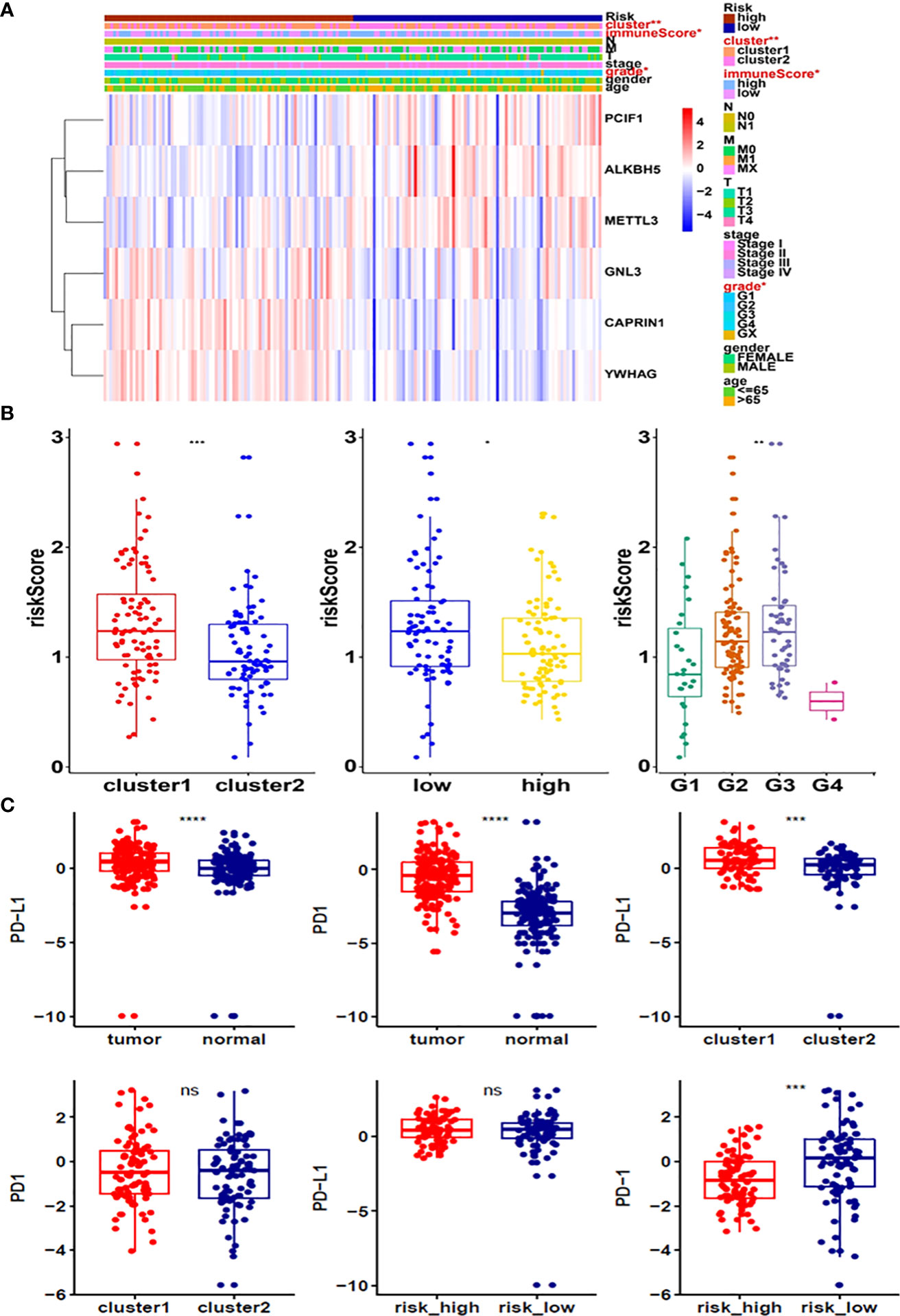
Figure 6 (A) Heatmap of clinicopathological features of pancreatic cancer cohort. (B) Distribution of risk scores stratified by cluster1/2. (C) The expression of PD-1 and PD-L1 in tumors, cluster1/2 and high/low-risk groups. *p<0.05; **p<0.01; ***p<0.001; ****p<0.0001. ns, no significance.
To confirm the association between METTL3 and PD-L1 in pancreatic cancer cells, we transfected METTL3 cDNA and shMETTL3 plasmids to BxPC-3 and PANC-1 cells. We observed that overexpression of METTL3 increased the expression of PD-L1, whereas shMETTL3 infection led to downregulation of PD-L1 in pancreatic cancer cells (Figure 7A). It has been known that METTL3 can regulate the expression of lncRNA MALAT1. Next, we tested whether METTL3 modulation can govern the expression level of lncRNA MALAT1 in pancreatic cancer cells. Indeed, upregulation of METTL3 increased the MALAT1 level, and downregulation of METTL3 reduced the expression of MALAT1 in pancreatic cancer cells (Figure 7B). Moreover, MALAT1 overexpression increased PD-L1 expression level, while reduction of MALAT1 reduced PD-L1 level in pancreatic cancer cells (Figure 7C). Taken together, METTL3 regulates the expression of PD-L1 partly due to regulation of lncRNA MALAT1 in pancreatic cancer.
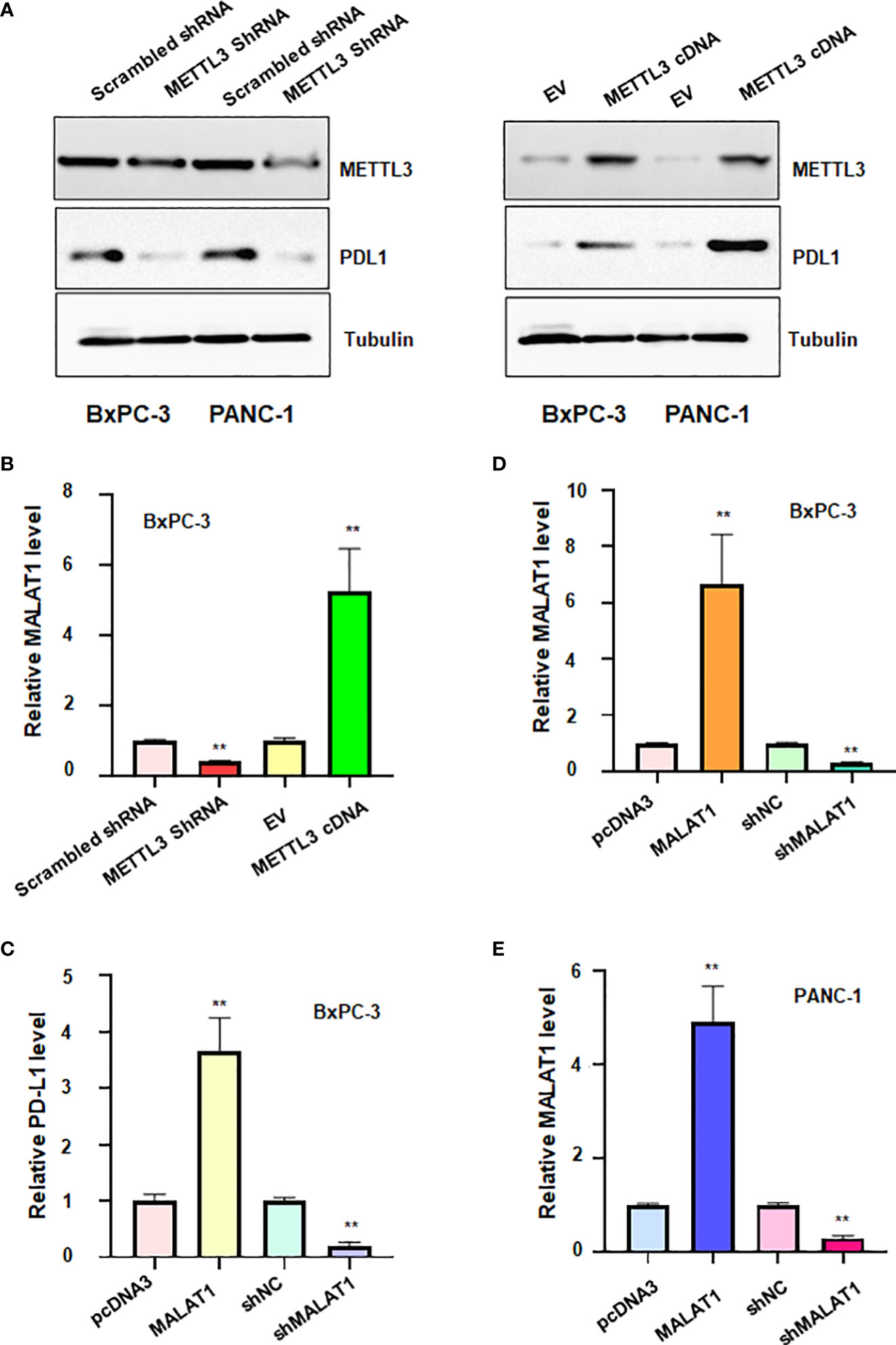
Figure 7 The relationship between METTL3, lncRNA MALAT1 and PD-L1 in PADC cells. (A) Western blotting was used to measure the expression of PD-L1 in BxPC-3 and PANC-1 cells after METTL3 modulation. (B) RT-PCR was used to measure the expression of lncRNA MALAT1 in BxPC-3 cells after METTL3 modulation. (C) RT-PCR was used to test the expression of PD-L1 in BxPC-3 cells after lncRNA MALAT1 changes. D-E: RT-PCR was used to measure the expression of MALAT1 in BxPC-3 (D) and PANC-1 cells (E) after MALAT1 modulation. **p<0.01.
To define the role of lncRNA MALAT1 in regulation of viability of pancreatic cancer cells, we used shMALAT1 or MALAT1 cDNA to modulate the expression of MALAT1 in pancreatic cancer cells. We found that shMALAT1 transfection suppressed the expression of lncRNA MALAT1, while MALAT1 cDNA transfection elevated the expression of MALAT1 in pancreatic cancer cells (Figures 7D, E). Moreover, increased expression of MALAT1 promoted the viability of BxPC-3 and PANC-1 (Figure 8A). Furthermore, depletion of MALAT1 attenuated the cell viability at 48 h and 72 h in pancreatic cancer cells (Figure 8B). Altogether, lncRNA MALAT1 regulates viability of pancreatic cancer cells.
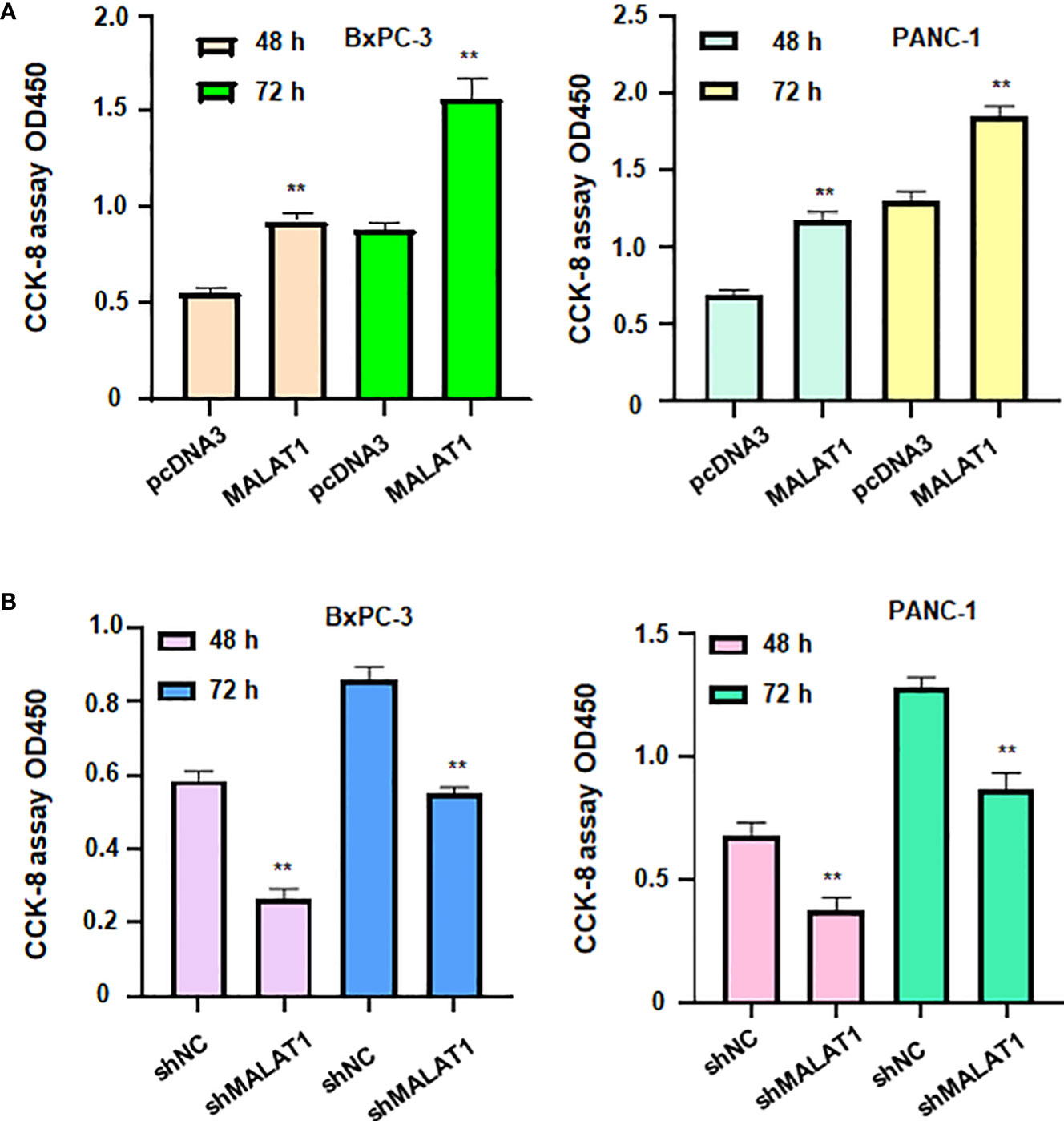
Figure 8 LncRNA MALAT1 regulates viability of pancreatic cancer cells. (A) CCK-8 assay was used to measure the viability of BxPC-3 and PANC-1 cells after MALAT1 overexpression. (B) CCK-8 assay was conducted to measure the viability of BxPC-3 and PANC-1 cells after MALAT1 downregulation. **p<0.01.
Pancreatic cancer is one of the common malignant tumors of the digestive tract and is known as the “king of cancer” in the field of tumor (35). The current treatment methods are mainly chemotherapy and surgery (36). However, the five-year survival rate after diagnosis of pancreatic cancer is about 10%, and it is one of the malignant tumors with poor prognosis (37). Immunotherapy of PD-1/PD-L1 has brought the hope for the treatment of pancreatic cancer (38). However, studies have shown that pancreatic cancer has a special tumor immune microenvironment, which brings challenges to immunotherapy and deserves further study (8). m6A methylation is the most common form of mRNA modification and involves in tumorigenesis (39). However, the role of m6A methylation in pancreatic cancer and the relationship between m6A, PD-1 and the infiltration of TIME in pancreatic cancer were unelucidated.
This study analyzed the relationship between m6A RNA methylation regulators, PD-L1, prognosis and TIME in pancreatic cancer. We found a total of 24 m6A genes that were highly expressed in the tumor samples. PD-1/PD-L1 was significantly associated with 20 m6A regulators. Subsequently, we used consensus clustering to identify two subgroups (cluster 1 and cluster 2) and found that patients in cluster2 displayed better prognosis than cluster1. Furthermore, cluster1 and cluster2 may be associated with cell cycle, p53 pathway, mismatch repair, RN degradation and Spliceosome. Notably, we identified risk signatures, including GNL3, CAPRIN1, METTL3, YWHAG, ALKBH5 and PCIF1.
A recent study found that there is a significantly increasing of METTL3 expression in PDAC cells (40), and this is consistent with our study. Our findings suggested that the expression difference of m6A RNA methylation regulator between tumor and normal samples was significant. Moreover, other evidence suggested that the upregulation of METTL3 can promote proliferation and invasion of pancreatic cancer (41). The chemical resistance in tumor cells will increase because of the rising expression of METTL3 (42). More specifically, a study pointed out that hypomethylation of the METTL3 promoter leads to overexpression of METTL3, which cooperates with NF-κB activating protein (NKAP) to coordinate m6A modification of the primary transcript of miR-25, making it mature miR-25-3p. Moreover, miR-25-3p inhibits PHLPP2 and activates oncogenic AKT-p70S6K signaling, and promotes the occurrence and progression of pancreatic cancer (43). METTL3 expression plays an important role in the TIME of pancreatic cancer (44, 45).
METTL3 has been reported to regulate the expression of PD-L1 in various cancer types. One study showed that METTL3 increased the expression of PD-L1 and intensified the malignant phenotype in oral squamous cell carcinoma (46). Another group identified that METTL3 can upregulate the expression of PD-L1 mRNA in breast cancer cells (47). In line with this report, METTL3 also elevated the PD-L1 mRNA in bladder cancer cells (48). In the present study, we found that METTL3 had a weak association with PD-L1 expression in pancreatic cancer patients. Moreover, METTL3 positively regulates the expression of PD-L1 in pancreatic cancer cells. METTL3 has been known to upregulate the expression of MALAT1 in several cancer types. METTL3 promoted the stability of MALAT1 and enhanced the glioma progression (49). METTL3 targeted MALAT1/miR-26b/HMGA2 axis and caused EMT and promotion of migration and invasion in breast cancer (50). METTL3 regulated MALAT1/E2F1/AGR2 pathway and subsequently controlled Adriamycin resistance in breast cancer (51). We also found that METTL3 controlled the expression of MALAT1 in pancreatic cancer cells.
LncRNA MALAT1 upregulated the expression of PD-L1 via sponging miR-195, leading to promotion of tumorigenesis in diffuse large B cell lymphoma (52). MALAT1 elevated the PD-L1 expression level via inhibition of miR-200a-3p, resulting in non-small lung cancer progression (53). Our data showed that MALAT1 can regulate the expression of PD-L1 in pancreatic cancer cells. LncRNA MALAT1 has been found to maintain the cancer stem cell-like properties in pancreatic cancer cells, including self-renewing ability, chemoresistance and angiogenesis (54). Han et al. reported that MALAT1 interacted with EZH2 and suppressed E-cadherin expression, leading to EZH2-induced invasion and migration in pancreatic cancer (55). Li et al. found that MALAT1 facilitated tumor cell metastasis and proliferation via the promotion of autophagy in pancreatic cancer (56). Zhang et al. revealed that miR-216a triggered apoptosis and G2/M arrest in pancreatic cancer cells via inhibition of MALAT1 expression (57). MALAT1 downregulation retarded pancreatic cancer progression via targeting Hippo-YAP signaling pathway (58). Moreover, MALAT1 was involved in efficacy of gemcitabine treatment in pancreatic cancer patients (59). Recently, MALAT1 was identified to govern pancreatic cancer progression via modulation of miR-129-5p (60). In the current study, we found that depletion of MALAT1 reduced cell viability, whereas overexpression of MALAT1 enhanced viability of pancreatic cancer cells.
In summary, this study is the first time to apply a bioinformatic approach to describe the relationship between m6A and PD-L1 and the TIME in pancreatic cancer. There is a limitation that this work mainly used a bioinformatic strategy to explore the association among m6A, PD-L1 and TME in pancreatic cancer. The role of METTL3 in pancreatic cancer development should be validated in animal study and clinical tissue samples. The mechanism of lncRNA MALAT1-mediated pancreatic oncogenesis should be dissected. How METTL3 regulates PD-L1 expression via regulation of lncRNA MALAT1 is required to be determined in the future.
The original contributions presented in the study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding authors.
ZS, XW, FC, WL, QC, XY and XZ performed the experiments, analyzed the data. ZS, QC and XL wrote the manuscript. PW edited the manuscript and supervised this study. All authors read and approved the final manuscript.
This research was supported by Natural Science Foundation of Zhejiang Province (No.LY21H160046).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
1. Jabbari J, Ahmed A, Walters RW, Silberstein PT. Surgical intervention in early-stage pancreatic adenocarcinoma. J Clin Oncol (2017) 35(4_suppl):443–3. doi: 10.1200/JCO.2017.35.4_suppl.443
2. Zhou H, Yin K, Zhang Y, Tian J and Wang S. The RNA m6A writer METTL14 in cancers: Roles, structures, and applications. Biochim Biophys Acta (BBA) - Rev Cancer (2021) 1876(2):188609. doi: 10.1016/j.bbcan.2021.188609
3. Siegel RL, Miller KD, Fuchs HE and Jemal A. Cancer statistics, 2022. CA: Cancer J Clin (2022) 72(1):7–33. doi: 10.3322/caac.21708
4. Traub B, Link K-H and Kornmann M. Curing pancreatic cancer. Semin Cancer Biol (2021) 76:232–46. doi: 10.1016/j.semcancer.2021.05.030
5. Mizrahi JD, Surana R, Valle JW and Shroff RT. Pancreatic cancer. Lancet (2020) 395(10242):2008–20. doi: 10.1016/S0140-6736(20)30974-0
6. Makohon-Moore A, Iacobuzio-Donahue CA. Pancreatic cancer biology and genetics from an evolutionary perspective. Nat Rev Cancer (2016) 16(9):553–65. doi: 10.1038/nrc.2016.66
7. Leinwand J, Miller G. Regulation and modulation of antitumor immunity in pancreatic cancer. Nat Immunol (2020) 21(10):1152–9. doi: 10.1038/s41590-020-0761-y
8. Guo Y, Wang R, Li J, Song Y, Min J, Zhao T, et al. Comprehensive analysis of m6A RNA methylation regulators and the immune microenvironment to aid immunotherapy in pancreatic cancer. Front Immunol (2021) 12:769425. doi: 10.3389/fimmu.2021.769425
9. Liu J, Chen T, Li S, Liu W, Wang P and Shang G. Targeting matrix metalloproteinases by E3 ubiquitin ligases as a way to regulate the tumor microenvironment for cancer therapy. Semin Cancer Biol (2022) S1044-579X(22):00138–9. doi: 10.1016/j.semcancer.2022.06.004
10. Jiang W, Pan S, Chen X, Wang ZW and Zhu X. The role of lncRNAs and circRNAs in the PD-1/PD-L1 pathway in cancer immunotherapy. Mol Cancer (2021) 20(1):116. doi: 10.1186/s12943-021-01406-7
11. Jiang W, Xia J, Xie S, Zou R, Pan S, Wang ZW, et al. Long non-coding RNAs as a determinant of cancer drug resistance: Towards the overcoming of chemoresistance via modulation of lncRNAs. Drug Resist Update (2020) 50:100683. doi: 10.1016/j.drup.2020.100683
12. Wu J, Cai J. Dilemma and challenge of immunotherapy for pancreatic cancer. Dig Dis Sci (2021) 66(2):359–68. doi: 10.1007/s10620-020-06183-9
13. Schumacher TN, Schreiber RD. Neoantigens in cancer immunotherapy. Science (2015) 348(6230):69–74. doi: 10.1126/science.aaa4971
14. Jiang X, Liu B, Nie Z, Duan L, Xiong Q, Jin Z, et al. The role of m6A modification in the biological functions and diseases. Signal Transduct. Target. Ther (2021) 6(1):74. doi: 10.1038/s41392-020-00450-x
15. He L, Li H, Wu A, Peng Y, Shu G and Yin G. Functions of N6-methyladenosine and its role in cancer. Mol Cancer (2019) 18(1):176. doi: 10.1186/s12943-019-1109-9
16. Hu B-b, Wang X-y, Gu X-Y, Zou C, Gao Z-j, Zhang H, et al. N6-methyladenosine (m6A) RNA modification in gastrointestinal tract cancers: roles, mechanisms, and applications. Mol Cancer (2019) 18(1):178. doi: 10.1186/s12943-019-1099-7
17. Wang X, Zhao BS, Roundtree IA, Lu Z, Han D, Ma H, et al. N(6)-methyladenosine modulates messenger RNA translation efficiency. Cell (2015) 161(6):1388–99. doi: 10.1016/j.cell.2015.05.014
18. Huang W, Qi CB, Lv SW, Xie M, Feng YQ, Huang WH, et al. Determination of DNA and RNA methylation in circulating tumor cells by mass spectrometry. Anal Chem (2016) 88(2):1378–84. doi: 10.1021/acs.analchem.5b03962
19. Liu J, Shang G. The roles of noncoding RNAs in the development of osteosarcoma stem cells and potential therapeutic targets. Front Cell Dev Biol (2022) 10:773038. doi: 10.3389/fcell.2022.773038
20. Chen X, Liu Y, Liu H, Wang ZW and Zhu X. Unraveling diverse roles of noncoding RNAs in various human papillomavirus negative cancers. Pharmacol Ther (2022) 238:108188. doi: 10.1016/j.pharmthera.2022.108188
21. Xie W, Chu M, Song G, Zuo Z, Han Z, Chen C, et al. Emerging roles of long noncoding RNAs in chemoresistance of pancreatic cancer. Semin Cancer Biol (2022) 83:303–18. doi: 10.1016/j.semcancer.2020.11.004
22. Gareev I, Gileva Y, Dzidzaria A, Beylerli O, Pavlov V, Agaverdiev M, et al. Long non-coding RNAs in oncourology. Noncoding RNA Res (2021) 6(3):139–45. doi: 10.1016/j.ncrna.2021.08.001
23. Liu JH, Chen G, Dang YW, Li CJ and Luo DZ. Expression and prognostic significance of lncRNA MALAT1 in pancreatic cancer tissues. Asian Pac J Cancer Prev (2014) 15(7):2971–7. doi: 10.7314/apjcp.2014.15.7.2971
24. Pang EJ, Yang R, Fu XB and Liu YF. Overexpression of long non-coding RNA MALAT1 is correlated with clinical progression and unfavorable prognosis in pancreatic cancer. Tumour Biol (2015) 36(4):2403–7. doi: 10.1007/s13277-014-2850-8
25. Jiao F, Hu H, Yuan C, Wang L, Jiang W, Jin Z, et al. Elevated expression level of long noncoding RNA MALAT-1 facilitates cell growth, migration and invasion in pancreatic cancer. Oncol Rep (2014) 32(6):2485–92. doi: 10.3892/or.2014.3518
26. Zaccara S, Ries RJ and Jaffrey SR. Reading, writing and erasing mRNA methylation. Nat Rev Mol Cell Biol (2019) 20(10):608–24. doi: 10.1038/s41580-019-0168-5
27. Szklarczyk D, Gable AL, Lyon D, Junge A, Wyder S, Huerta-Cepas J, et al. STRING v11: protein-protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res (2019) 47(D1):D607–d613. doi: 10.1093/nar/gky1131
28. Seiler M, Huang CC, Szalma S and Bhanot G. ConsensusCluster: a software tool for unsupervised cluster discovery in numerical data. Omics (2010) 14(1):109–13. doi: 10.1089/omi.2009.0083
29. Murohashi M, Hinohara K, Kuroda M, Isagawa T, Tsuji S, Kobayashi S, et al. Gene set enrichment analysis provides insight into novel signalling pathways in breast cancer stem cells. Br J Cancer (2010) 102(1):206–12. doi: 10.1038/sj.bjc.6605468
30. Vegesna R. Inferring tumour purity and stromal and immune cell admixture from expression data. Nat Commun (2013) 4:2612. doi: 10.1038/ncomms3612
31. Castellani G, Buccarelli M, Lulli V, Ilari R, De Luca G, Pedini F, et al. MiR-378a-3p acts as a tumor suppressor in colorectal cancer stem-like cells and affects the expression of MALAT1 and NEAT1 lncRNAs. Front Oncol (2022) 12:867886. doi: 10.3389/fonc.2022.867886
32. Chen T, Liu J, Zhang H, Li J and Shang G. Long intergenic noncoding RNA00265 enhances cell viability and metastasis via targeting miR-485-5p/USP22 axis in osteosarcoma. Front Oncol (2022) 12:907472. doi: 10.3389/fonc.2022.907472
33. Wu L, Yu K, Chen K, Zhu X, Yang Z, Wang Q, et al. Fbxo45 facilitates pancreatic carcinoma progression by targeting USP49 for ubiquitination and degradation. Cell Death Dis (2022) 13(3):231. doi: 10.1038/s41419-022-04675-2
34. Xu Y, Chen X, Pan S, Wang ZW and Zhu X. TM7SF2 regulates cell proliferation and apoptosis by activation of c-Raf/ERK pathway in cervical cancer. Cell Death Discovery (2021) 7(1):299. doi: 10.1038/s41420-021-00689-5
35. Force UPST. Screening for pancreatic cancer: US preventive services task force reaffirmation recommendation statement. JAMA (2019) 322(5):438–44. doi: 10.1001/jama.2019.10232
36. Kamisawa T, Wood LD, Itoi T and Takaori K. Pancreatic cancer. Lancet (2016) 388(10039):73–85. doi: 10.1016/S0140-6736(16)00141-0
37. Semaan A, Maitra A. Rebooting pancreatic cancer knowledge and treatment options. Nat Rev Gastroenterol Hepatol (2018) 15(2):76–8. doi: 10.1038/nrgastro.2017.182
38. Morrison AH, Byrne KT and Vonderheide RH. Immunotherapy and prevention of pancreatic cancer. Trends Cancer (2018) 4(6):418–28. doi: 10.1016/j.trecan.2018.04.001
39. He PC, He C. m6A RNA methylation: from mechanisms to therapeutic potential. EMBO J (2021) 40(3):e105977. doi: 10.15252/embj.2020105977
40. Xia T, Wu X, Cao M, Zhang P, Shi G, Zhang J, et al. The RNA m6A methyltransferase METTL3 promotes pancreatic cancer cell proliferation and invasion. Pathol.-Res. Pract (2019) 215(11):152666. doi: 10.1016/j.prp.2019.152666
41. Li J, Wang F, Liu Y, Wang H and Ni B. N(6)-methyladenosine (m(6)A) in pancreatic cancer: Regulatory mechanisms and future direction. Int J Biol Sci (2021) 17(9):2323–35. doi: 10.7150/ijbs.60115
42. Taketo K, Konno M, Asai A, Koseki J, Toratani M, Satoh T, et al. The epitranscriptome m6A writer METTL3 promotes chemo-and radioresistance in pancreatic cancer cells. Int J Oncol (2018) 52(2):621–9. doi: 10.3892/ijo.2017.4219s
43. Zhang J, Bai R, Li M, Ye H, Wu C, Wang C, et al. Excessive miR-25-3p maturation via N6-methyladenosine stimulated by cigarette smoke promotes pancreatic cancer progression. Nat Commun (2019) 10(1):1858. doi: 10.1038/s41467-019-09712-x
44. Xu F, Zhang Z, Yuan M, Zhao Y, Zhou Y, Pei H, et al. M6A regulatory genes play an important role in the prognosis, progression and immune microenvironment of pancreatic adenocarcinoma. Cancer Invest (2021) 39(1):39–54. doi: 10.1080/07357907.2020.1834576
45. Wang L, Zhang S, Li H, Xu Y, Wu Q, Shen J, et al. Quantification of m6A RNA methylation modulators pattern was a potential biomarker for prognosis and associated with tumor immune microenvironment of pancreatic adenocarcinoma. BMC Cancer (2021) 21(1):876. doi: 10.1186/s12885-021-08550-9
46. Ai Y, Liu S, Luo H, Wu S, Wei H, Tang Z, et al. METTL3 intensifies the progress of oral squamous cell carcinoma via modulating the m6A amount of PRMT5 and PD-L1. J Immunol Res (2021) 2021:6149558. doi: 10.1155/2021/6149558
47. Wan W, Ao X, Chen Q, Yu Y, Ao L, Xing W, et al. METTL3/IGF2BP3 axis inhibits tumor immune surveillance by upregulating N(6)-methyladenosine modification of PD-L1 mRNA in breast cancer. Mol Cancer (2022) 21(1):60. doi: 10.1186/s12943-021-01447-y
48. Ni Z, Sun P, Zheng J, Wu M, Yang C, Cheng M, et al. JNK signaling promotes bladder cancer immune escape by regulating METTL3-mediated m6A modification of PD-L1 mRNA. Cancer Res (2022) 82(9):1789–802. doi: 10.1158/0008-5472.CAN-21-1323
49. Chang YZ, Chai RC, Pang B, Chang X, An SY, Zhang KN, et al. METTL3 enhances the stability of MALAT1 with the assistance of HuR via m6A modification and activates NF-kappaB to promote the malignant progression of IDH-wildtype glioma. Cancer Lett (2021) 511:36–46. doi: 10.1016/j.canlet.2021.04.020
50. Zhao C, Ling X, Xia Y, Yan B and Guan Q. The m6A methyltransferase METTL3 controls epithelial-mesenchymal transition, migration and invasion of breast cancer through the MALAT1/miR-26b/HMGA2 axis. Cancer Cell Int (2021) 21(1):441. doi: 10.1186/s12935-021-02113-5
51. Li S, Jiang F, Chen F, Deng Y and Pan X. Effect of m6A methyltransferase METTL3 -mediated MALAT1/E2F1/AGR2 axis on adriamycin resistance in breast cancer. J Biochem Mol Toxicol (2022) 36(1):e22922. doi: 10.1002/jbt.22922
52. Wang QM, Lian GY, Song Y, Huang YF and Gong Y. LncRNA MALAT1 promotes tumorigenesis and immune escape of diffuse large b cell lymphoma by sponging miR-195. Life Sci (2019) 231:116335. doi: 10.1016/j.lfs.2019.03.040
53. Wei S, Wang K, Huang X, Zhao Z and Zhao Z. LncRNA MALAT1 contributes to non-small cell lung cancer progression via modulating miR-200a-3p/programmed death-ligand 1 axis. Int J Immunopathol Pharmacol (2019) 33:2058738419859699. doi: 10.1177/2058738419859699
54. Jiao F, Hu H, Han T, Yuan C, Wang L, Jin Z, et al. Long noncoding RNA MALAT-1 enhances stem cell-like phenotypes in pancreatic cancer cells. Int J Mol Sci (2015) 16(4):6677–93. doi: 10.3390/ijms16046677
55. Han T, Jiao F, Hu H, Yuan C, Wang L, Jin ZL, et al. EZH2 promotes cell migration and invasion but not alters cell proliferation by suppressing e-cadherin, partly through association with MALAT-1 in pancreatic cancer. Oncotarget (2016) 7(10):11194–207. doi: 10.18632/oncotarget.7156
56. Li L, Chen H, Gao Y, Wang YW, Zhang GQ, Pan SH, et al. Long noncoding RNA MALAT1 promotes aggressive pancreatic cancer proliferation and metastasis via the stimulation of autophagy. Mol Cancer Ther (2016) 15(9):2232–43. doi: 10.1158/1535-7163.MCT-16-0008
57. Zhang Y, Tang X, Shi M, Wen C and Shen B. MiR-216a decreases MALAT1 expression, induces G2/M arrest and apoptosis in pancreatic cancer cells. Biochem Biophys Res Commun (2017) 483(2):816–22. doi: 10.1016/j.bbrc.2016.12.167
58. Zhou Y, Shan T, Ding W, Hua Z, Shen Y, Lu Z, et al. Study on mechanism about long noncoding RNA MALAT1 affecting pancreatic cancer by regulating hippo-YAP signaling. J Cell Physiol (2018) 233(8):5805–14. doi: 10.1002/jcp.26357
59. Wang CJ, Shi SB, Tian J, Xu J and Niu ZX. lncRNA MALAT1, HOTTIP and PVT1 as predictors for predicting the efficacy of GEM based chemotherapy in first-line treatment of pancreatic cancer patients. Oncotarget (2017) 8(56):95108–15. doi: 10.18632/oncotarget.19345
Keywords: lncRNA, MALAT1, PD-L1, METTL3, TME, pancreatic cancer
Citation: Song Z, Wang X, Chen F, Chen Q, Liu W, Yang X, Zhu X, Liu X and Wang P (2022) LncRNA MALAT1 regulates METTL3-mediated PD-L1 expression and immune infiltrates in pancreatic cancer. Front. Oncol. 12:1004212. doi: 10.3389/fonc.2022.1004212
Received: 27 July 2022; Accepted: 15 August 2022;
Published: 21 September 2022.
Edited by:
Aamir Ahmad, University of Alabama at Birmingham, United StatesReviewed by:
Wentao Cho, California State University, United StatesCopyright © 2022 Song, Wang, Chen, Chen, Liu, Yang, Zhu, Liu and Wang. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Xiaorong Liu, YWluaWd1MTk4NzI5QHNpbmEuY24=; Peter Wang, d2FuZ3BldGVyMkBob3RtYWlsLmNvbQ==
†These authors have contributed equally to this work
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.