 Lucas Maciel Vieira1,2*
Lucas Maciel Vieira1,2* Natasha Andressa Nogueira Jorge2
Natasha Andressa Nogueira Jorge2 João Batista de Sousa3
João Batista de Sousa3 João Carlos Setubal4
João Carlos Setubal4 Peter F. Stadler2,5,6,7,8
Peter F. Stadler2,5,6,7,8 Maria Emília Machado Telles Walter1
Maria Emília Machado Telles Walter1- 1Departamento de Ciência da Computação, Instituto de Ciência Exatas, University of Brasília, Brasília, Brazil
- 2Bioinformatics Group, Department of Computer Science, and Interdisciplinary Center for Bioinformatics, Leipzig, Germany
- 3Division of Coloproctology, Department of Surgery, University of Brasília School of Medicine, Brasília, Brazil
- 4Department of Biochemistry, Institute of Chemistry, University of São Paulo, São Paulo, Brazil
- 5Max Planck Institute for Mathematics in the Science, Leipzig, Germany
- 6Institute for Theoretical Chemistry, University of Vienna, Wien, Austria
- 7Facultad de Ciencias, Universidad National de Colombia, Sede Bogotá, Colombia
- 8Santa Fe Institute, Santa Fe, CA, United States
Background: Colorectal cancer (CRC) is a heterogeneous cancer. Its treatment depends on its anatomical site and distinguishes between colon, rectum, and rectosigmoid junction cancer. This study aimed to identify diagnostic and prognostic biomarkers using networks of CRC-associated transcripts that can be built based on competing endogenous RNAs (ceRNA).
Methods: RNA expression and clinical information data of patients with colon, rectum, and rectosigmoid junction cancer were obtained from The Cancer Genome Atlas (TCGA). The RNA expression profiles were assessed through bioinformatics analysis, and a ceRNA was constructed for each CRC site. A functional enrichment analysis was performed to assess the functional roles of the ceRNA networks in the prognosis of colon, rectum, and rectosigmoid junction cancer. Finally, to verify the ceRNA impact on prognosis, an overall survival analysis was performed.
Results: The study identified various CRC site-specific prognosis biomarkers: hsa-miR-1271-5p, NRG1, hsa-miR-130a-3p, SNHG16, and hsa-miR-495-3p in the colon; E2F8 in the rectum and DMD and hsa-miR-130b-3p in the rectosigmoid junction. We also identified different biological pathways that highlight differences in CRC behavior at different anatomical sites, thus reinforcing the importance of correctly identifying the tumor site.
Conclusions: Several potential prognostic markers for colon, rectum, and rectosigmoid junction cancer were found. CeRNA networks could provide better understanding of the differences between, and common factors in, prognosis of colon, rectum, and rectosigmoid junction cancer.
Introduction
Colorectal cancer (CRC) is one of the most common and lethal cancers in the world (1). The most common histopathological type of CRC is adenocarcinoma (2). CRC can be classified according to its three major affected sites: colon, rectum, and rectosigmoid junction. Together these make up the large bowel. The colon corresponds to the largest portion, the rectum is located at the end, and the rectosigmoid junction is the transition between colon sigmoid and rectum. A tumor site is classified as belonging to the rectosigmoid junction when differentiation between rectum and sigmoid is not possible (3). CRC tumor site identification is important, due to different treatment strategies: for the colon, radical resection, depending on the stage, combined with chemotherapy is used; for rectum, only radical surgery or neoadjuvant chemorradiation followed or not by radical resection (4); while for the rectosigmoid junction, the best treatment still remains unknown (5). Because erroneous diagnosis of the CRC site can lead to overtreatment with chemotherapy, the identification of the CRC tumor site is a process that should be carefully analyzed, especially in the rectosigmoid junction (4). Better understanding of the biological characteristics of CRC at each site may provide insight into its development and progression.
Many studies have highlighted the importance of long non-coding RNAs (lncRNAs) in understanding the biological mechanisms of CRC and other types of cancer. LncRNAs are molecules greater than 200 nucleotides that do not encode for proteins and act mainly as transcriptional regulators by interacting with other molecules, such as miRNAs and mRNAs (6). These interaction mechanisms between lncRNAs and other molecules are explained by the competing endogenous RNA (ceRNA) hypothesis, which describes the interactions and their influence on altered protein expression levels (7). CeRNA network analyses have reported differentially expressed lncRNAs involved in breast, gastric, and many other types of cancer (8–10). Specifically for CRC, many studies also analyzed the ceRNAs networks and indicated potential diagnosis and prognosis biomarkers for colon, rectal, and colorectal cancer in general (1, 11–16). Most of these studies investigated the ceRNAs of CRC without differentiating the anatomical sites; however, different CRC sites present unique characteristics and treatment responses. Therefore, the identification of exclusive biomarkers for colon, rectum, or rectosigmoid junction could aid in understanding differences in the disease prognosis and progress. To the best of our knowledge, our study is the first to establish specific ceRNA networks for (i) colon; (ii) rectum; and (iii) rectosigmoid junction, and to associate them with specific biological mechanisms in order to clarify the differences and common factors between these sites.
In this study, we analyzed the functional and prognostic roles of lncRNAs, miRNAs, and mRNAs in colon, rectum, and rectosigmoid junction cancer, based on specific ceRNA networks constructed by using data from The Cancer Genome Atlas (TCGA) rectal adenocarcinoma (READ) and colon adenocarcinoma (COAD) projects.
Materials And Methods
Data
The RNA expression value raw count and clinical information data of patients with colon, rectum, and rectosigmoid junction CRC sites were downloaded from TCGA (17). The selection criteria was: (1) open access to information; (2) sample types from primary tumor or solid normal tissue; (3) patients with adenocarcinomas.
Analysis of Differentially Expressed RNAs
We used the GDCRNATools v1.6 (18) R package and implemented the limma (19) method to obtain differentially expressed (DE) lncRNAs, miRNAs, and protein coding genes (PCGs) for cancer and normal tissue. The expression profiles were normalized by the voom method implemented in the GDCRNATools. The RNAs presenting FDR ≤ 0.05 and |logF C| ≥ 2 were considered statistically significant.
CeRNA Network Construction
The ceRNA networks, for each CRC site, were constructed using the GDCRNATools v1.6 package of R, and the DE lncRNAs, miRNAs, and mRNAs. The network is based on the mRNA-miRNA-lncRNA interactions predicted by the spongeScan (20) algorithm and the starBase v2.0 (21) database. In the ceRNA networks, positively correlated mRNAs and lncRNAs act as sponges by sharing a significant number of miRNA binding sites and suppressing their functioning. The ceRNA networks generated show the possible molecule interactions related to each CRC site.
Functional Analysis
The functional analysis to assess the biological processes in the ceRNAs was done with the enrichment module of GDCRNATools. The gene list used for Gene Ontology (GO) (22), Kyoto Encyclopedia of Genes and Genomes (KEGG) (23), and Disease Ontology (DO) (24) came from the org.Hs.eg.db database v3.11.4, and all the human pathways from KEGG were considered. The adjusted p-value ≤ 0.05 was set as the threshold for statistical significance for GO, KEGG, and DO. Although some pathways present FDR > 0.05, they were still included as they represent good discussion points for the CRC functional analysis.
Survival Analysis
The Cox Proportional-Hazards (CoxPH) model from GDCRNATools was used to calculate the hazard ratio (HR) of the ceRNA molecules. An outlier removal was performed and only molecules with |higherLimit − HR| ≤ 6 and |lowerLimit − HR| ≤ 6 were considered. The survival curve was constructed by using the Kaplan Meier (KM) analysis from GDCRNATools. CoxPH and KM were used p < 0.05 as the threshold for statistical significance.
Results
Differentially Expressed RNAs
We obtained a total RNA expression raw count of 541 cancer and 48 non-tumor samples from 539 patients with CRC from the TCGA-COAD and TCGA-READ projects. A differential expression analysis was performed for each type of CRC. In the colon analysis, we found 140 upregulated and 75 downregulated lncRNAs, 213 upregulated and 136 downregulated miRNAs, and 1,179 upregulated and 1,906 downregulated PCGs (Figure 1A). In the rectum, we found 46 upregulated and 37 downregulated lncRNAs, 119 upregulated and 99 downregulated miRNAs, and 535 upregulated and 1,532 downregulated PCGs (Figure 1B). In the rectosigmoid junction, we found 149 upregulated and 59 downregulated lncRNAs, 181 upregulated and 108 downregulated miRNAs, and 1,005 upregulated and 1,880 downregulated PCGs (Figure 1C).
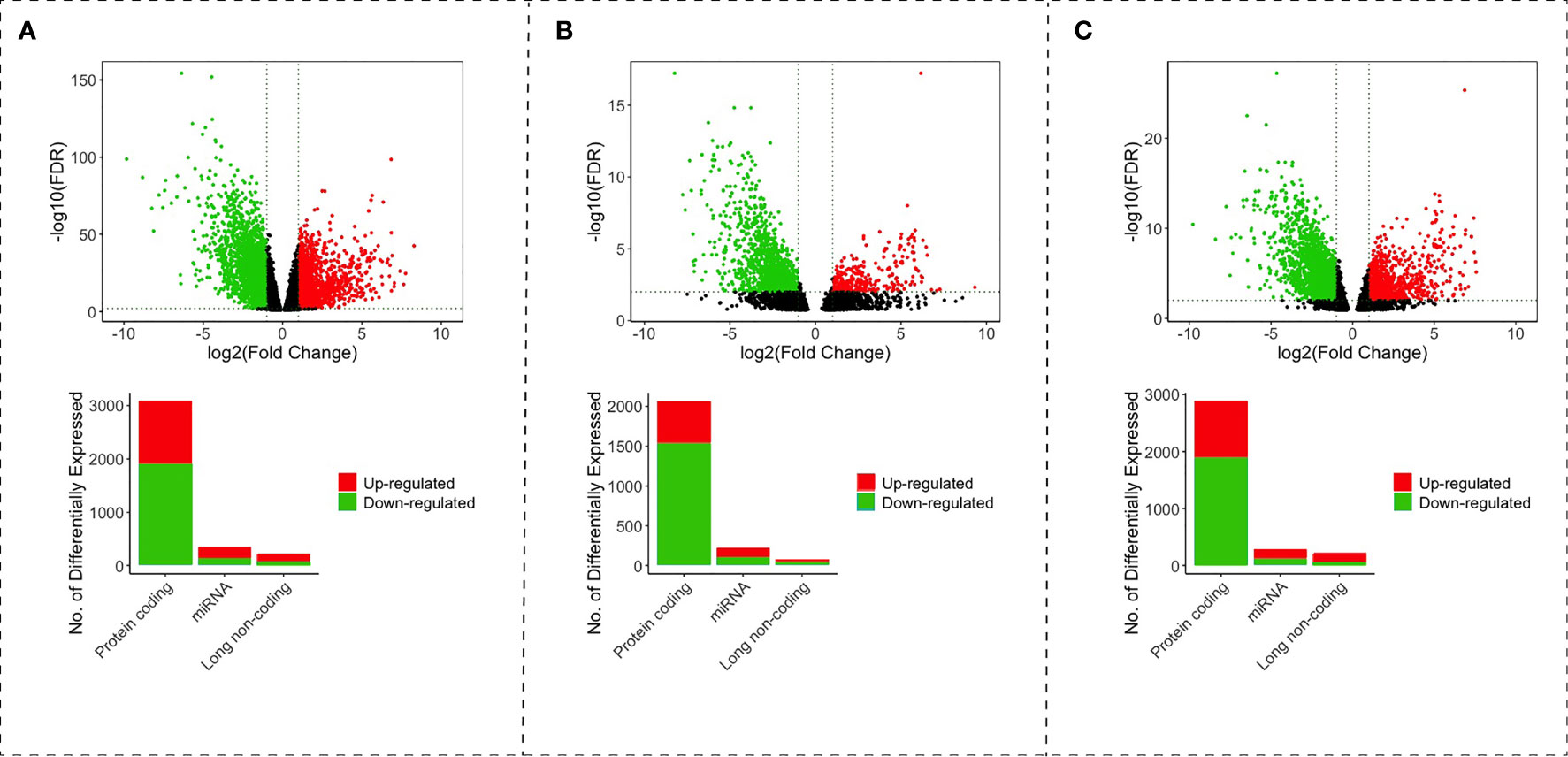
Figure 1 Volcano and bar plot with differentially expressed lncRNAs, miRNAs and mRNAs of colon (A), rectum (B), and rectosigmoid junction (C) sites. The red points and bars represent upregulated RNAs. The green points and bars represent downregulated RNAs.
CeRNA Networks
A ceRNA network, represented by a graph with its vertex (nodes) representing the molecules and the lines connecting them representing the interactions between the molecules, was established for each of the CRC sites. For colon, a ceRNA network consisting of 239 nodes and 506 interactions was established. For rectum, a ceRNA network consisting of 79 nodes and 136 interactions was established. For rectosigmoid junction, a ceRNA network consisting of 131 nodes and 210 interactions was established. We also analyzed the intersection between the ceRNA networks and found that: colon and rectum share 2 nodes and 2 interactions; colon and rectosigmoid junction share 48 nodes and 77 interactions; rectum and rectosigmoid junction share 12 nodes and 23 interactions; and all three sites share 47 nodes and 76 interactions. Furthermore, we performed an analysis to find individual nodes and interactions for each CRC site, finding that: colon has 142 nodes and 351 unique interactions; rectum has 18 nodes and 35 unique interactions; and rectosigmoid junction has 24 nodes and 34 unique interactions. Figure 2 shows all the described networks.
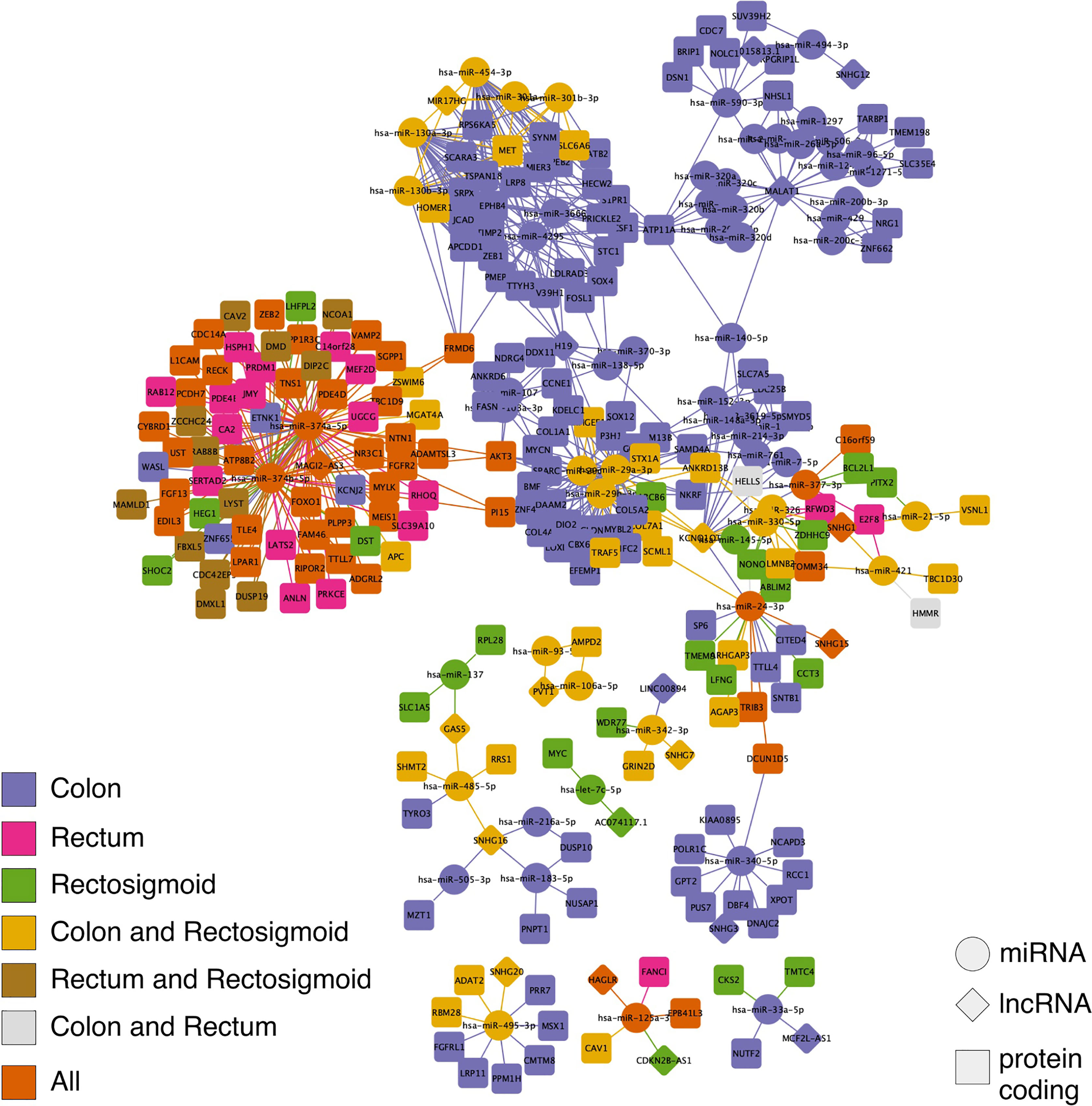
Figure 2 Competing endogenous RNA (ceRNA) network in colon, rectum and rectosigmoid junction sites. The diamonds represent lncRNAs, the circles represent miRNAs, and the squares represent PCGs. The molecules and interactions of each CRC site can be identified by color.
The intersection between each of the three site-specific ceRNA networks is regulated by the lncRNAs: MAGI2-AS3, HAGLR-AS3, SNHG1, and SNHG15 (Figure 3). HAGLR-AS3, SNHG1, and SNHG15 compose three different small ceRNA networks by sponging hsa-miR-125a-3p, hsa-miR-377-3p, and hsa-miR-24-3p, respectively. In the MAGI2-AS3 network, we see the ceRNA mechanism affecting a great number of PCGs via hsa-miR-374b-5p and hsa-miR-374a-5p, suggesting that this lncRNA has a great impact on the main CRC mechanism, which occurs at all sites.
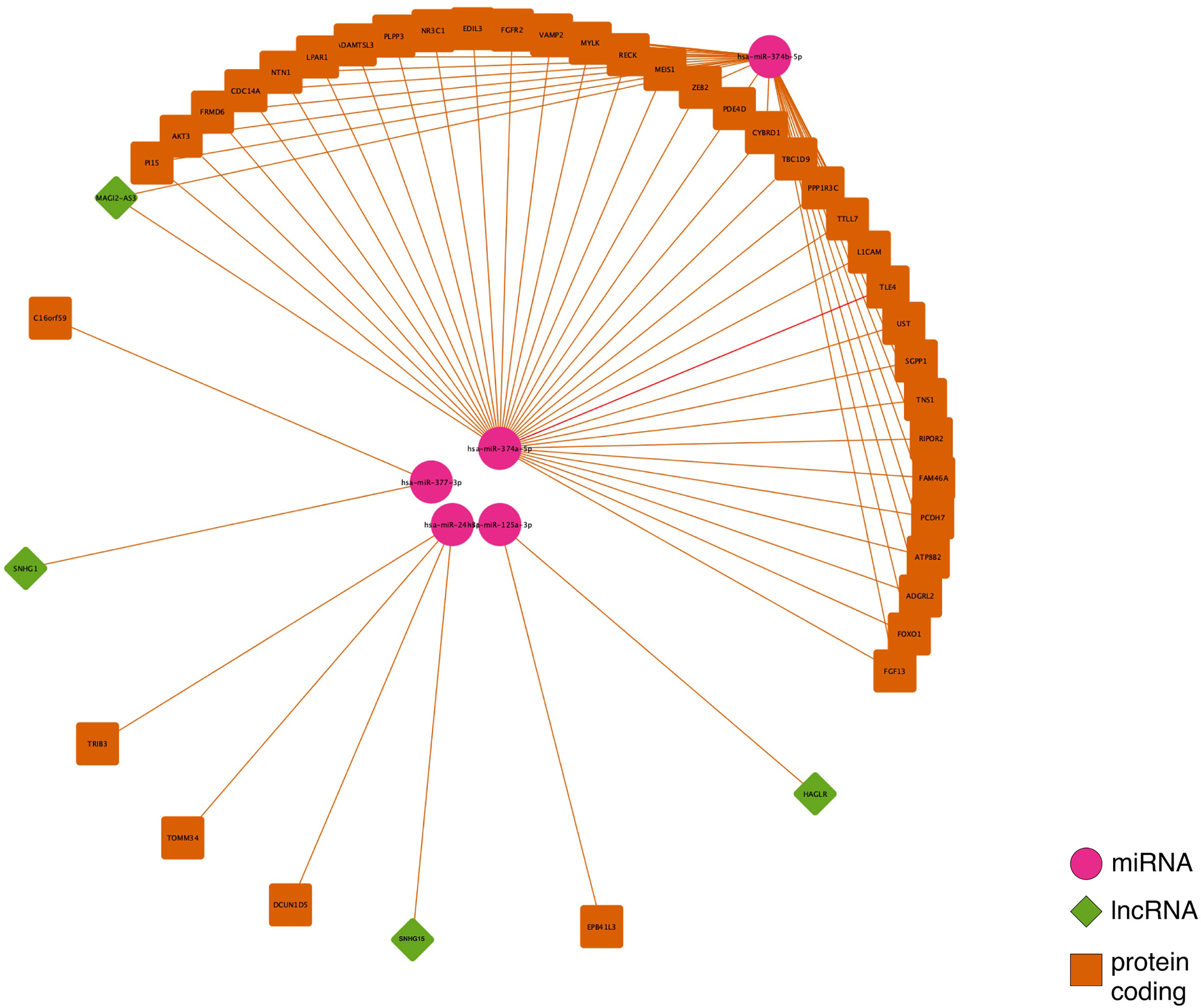
Figure 3 Competing endogenous RNA (ceRNA) network intersection for colon, rectum and rectosigmoid junction sites.
Functional and Survival Analysis
A functional enrichment analysis was performed with GO, KEGG, and DO, to indicate the potential biological roles of the ceRNAs, lncRNAs, and PCGs (Figure 4). Each of the sites presented a different main functional characteristic: colon pathways are mainly related to endothelial differentiation; rectum pathways are mainly related to apoptosis; and rectosigmoid junction pathways are mainly related to signal transduction.
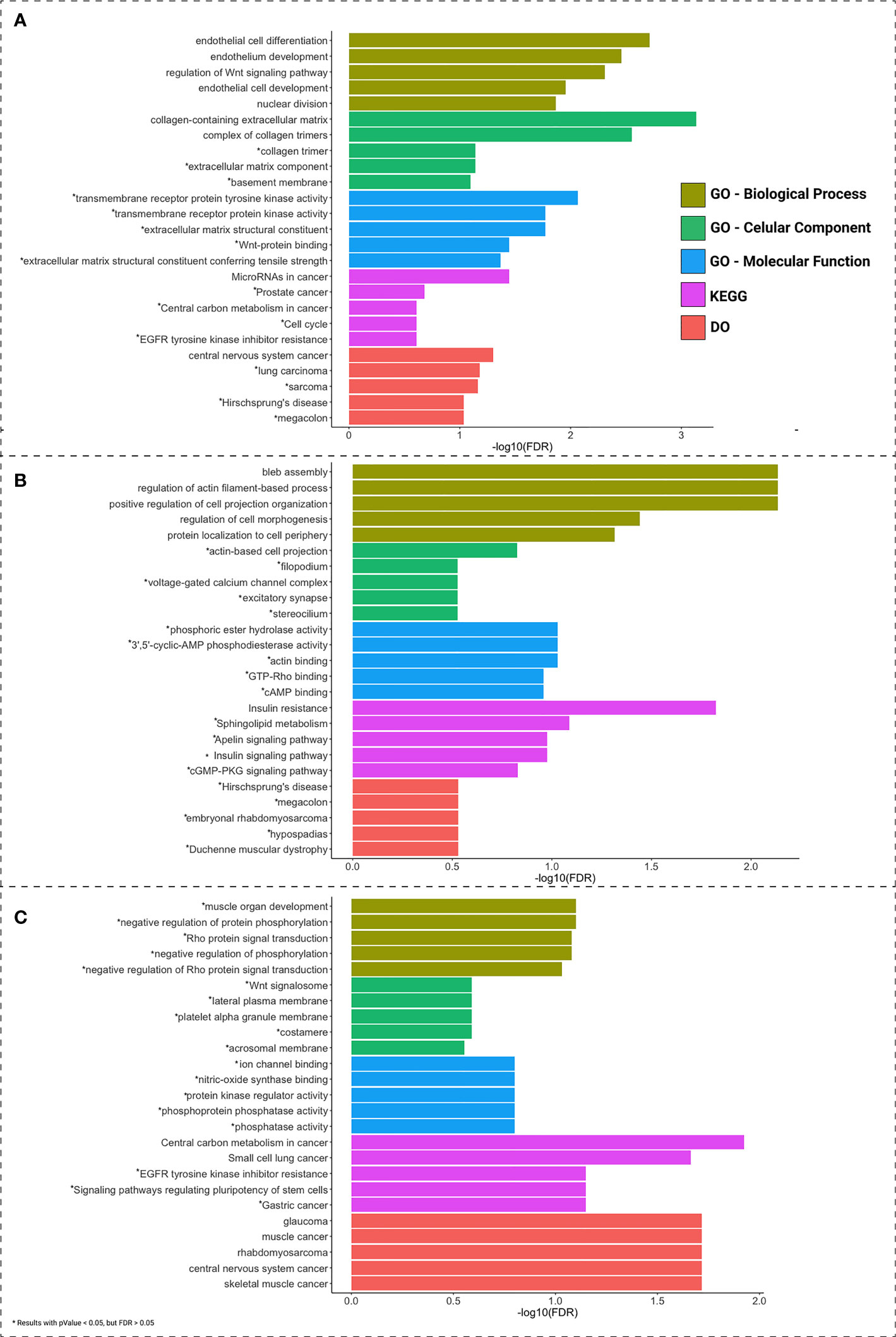
Figure 4 Functional enrichment analysis of PCGs and lncRNAs included in the ceRNA network of colon (A), rectum (B), and rectosigmoid junction (C) sites. The top 5 enrichment results for GO biological processes, cellular component, molecular function, DO and KEGG are shown in different colors. Asterisks (*) indicate pathways presenting FDR > 0.05.
To better understand the impact of ceRNA networks, we also performed a functional enrichment analysis for the MAGI2-HAGLR-AS3-SNHG1-SNHG15 intersection ceRNA network (Figure 3), and for each site-specific ceRNA network. The molecules present in the ceRNA network common in all CRC sites are mainly related to cell morphogenesis pathways (Figure 5A), such as regulation of Wnt signaling and cell morphogenesis and the process of insulin resistance. In the colon-specific (Figure 5B) ceRNA network, the most significant pathways are involved in tissue differentiation and development, such as endothelial cell differentiation and connective tissue development, while the rectum-specific (Figure 5C) ceRNA network is related to cell differentiation. No pathways were significant for the rectosigmoid junction site (Figure 5D).
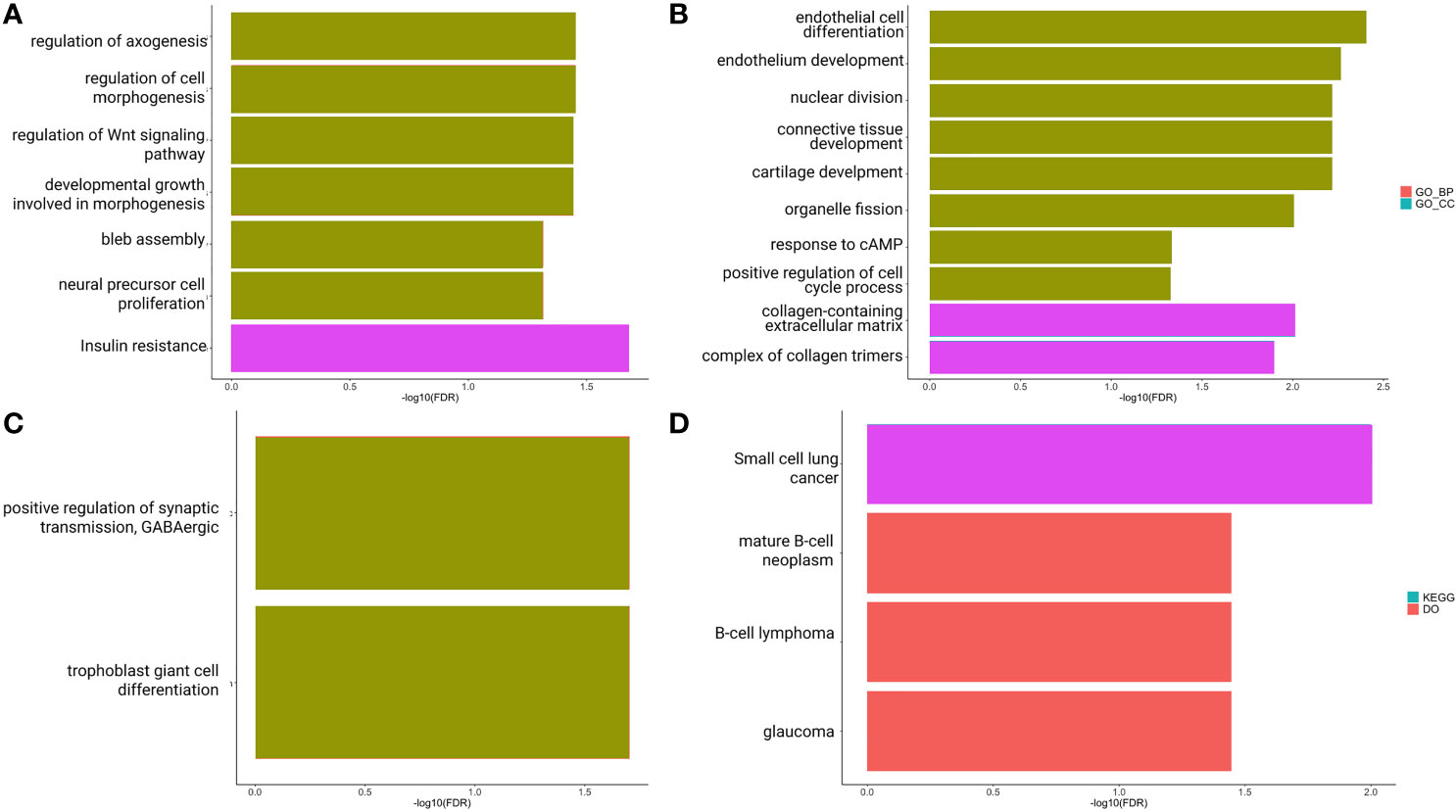
Figure 5 Functional enrichment analysis of PCGs and lncRNAs included in the ceRNA network: common to colon, rectum and rectosigmoid junction (A); specific to colon (B); specific to rectum and (C); specific to rectosigmoid junction (D) sites.
Clinical data from CRC patients was used to obtain the HR and to build the overall survival time curve. For each CRC site, we had data from 391 colon, 84 rectum, and 66 rectosigmoid junction patients. Using CoxPH, we identified 20 molecules from the previous ceRNA networks with relevant HR, being 14 in colon, 3 in rectum, and 4 in rectosigmoid junction molecules (Figure 6). The DMD gene was the only molecule with high HR, present in more than one CRC site.
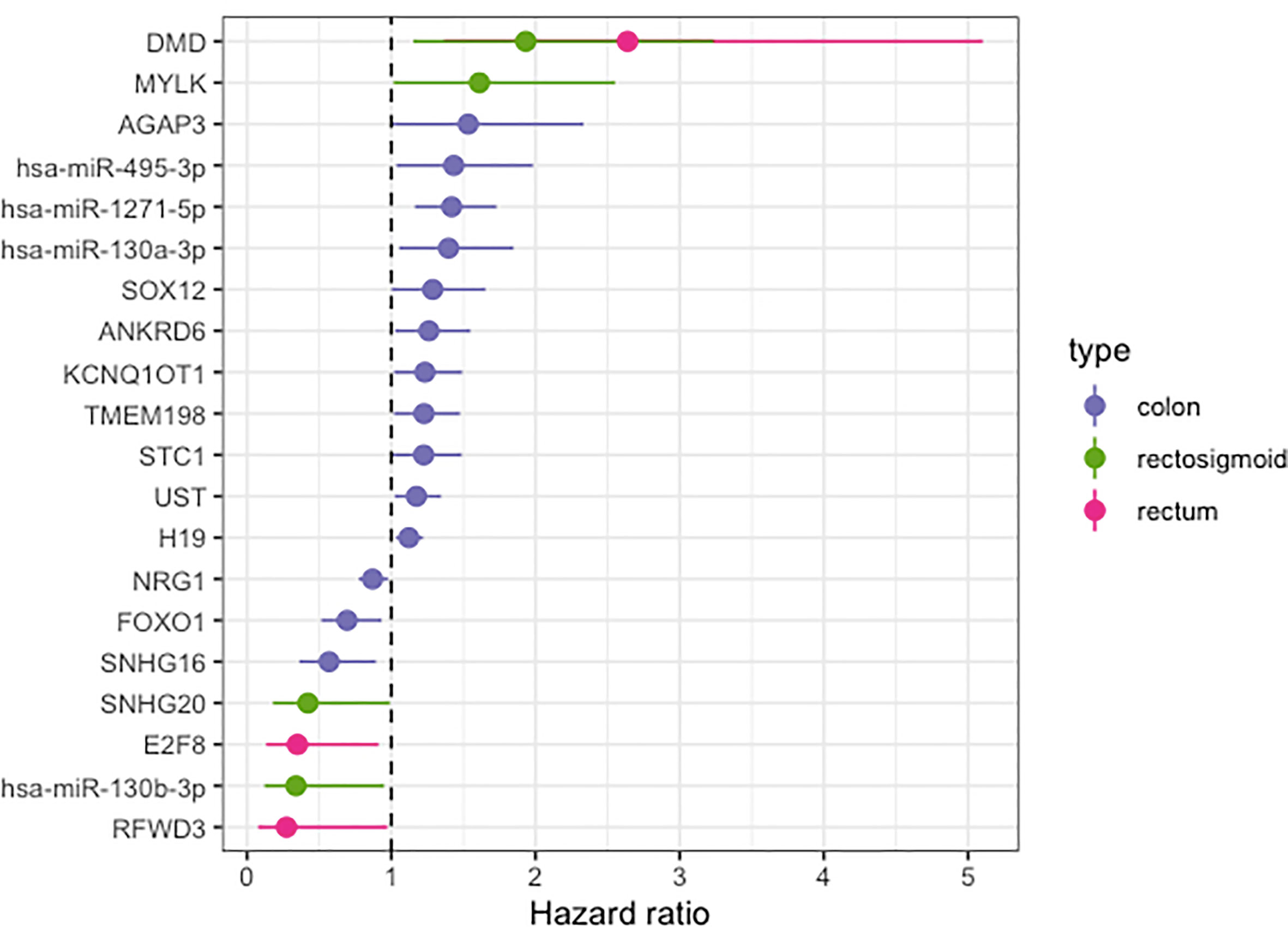
Figure 6 Hazard ratio forest plot of survival associated RNAs in the ceRNA network for colon, rectum and rectosigmoid junction sites. The molecules with hazard ratio < 1 indicate risk factors, and those with hazard ratio > 1 indicate protective factors.
Clinical data and the expression profile of the HR relevant genes were used for overall survival analysis. These analyses were divided into two sets: the molecules with lowest pValue according to the KM method (Figure 7) and the molecules with lowest pValue according to the KM and COxPh methods (Figure 8).
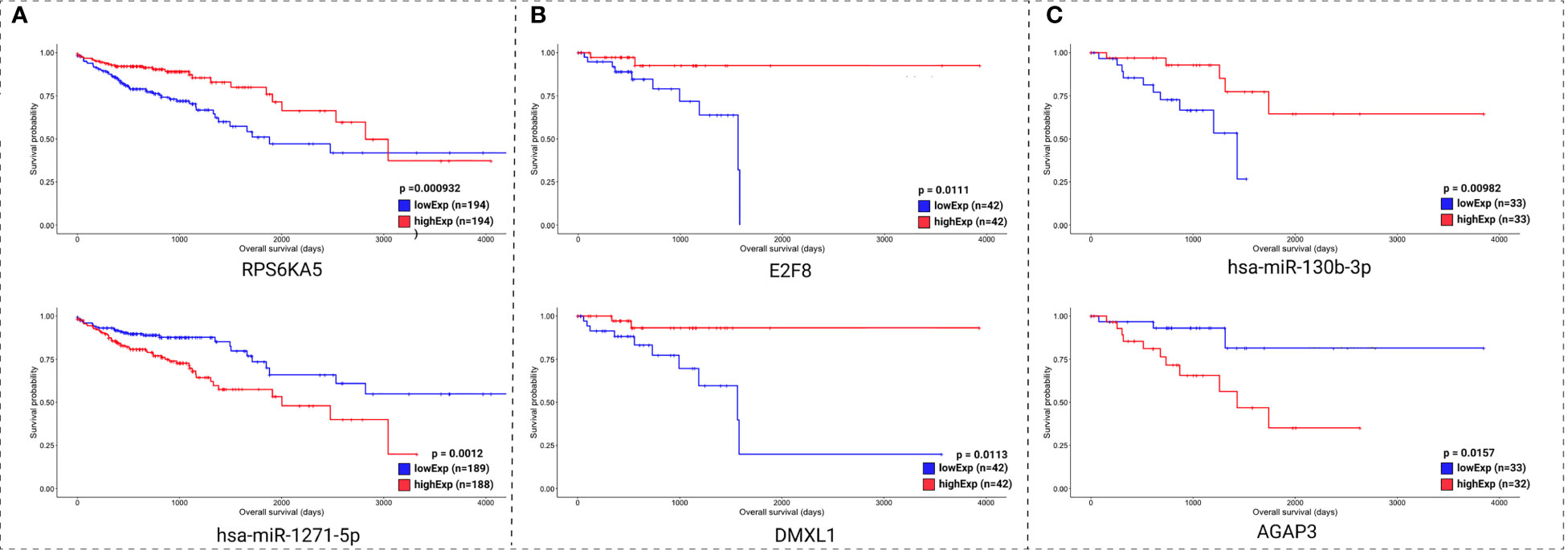
Figure 7 Kaplan-Meier survival curves for the two best scored molecules for colon (A), rectum (B), and rectosigmoid junction (C) sites. Horizontal axis: overall survival time (in days). Vertical axis: survival probability.
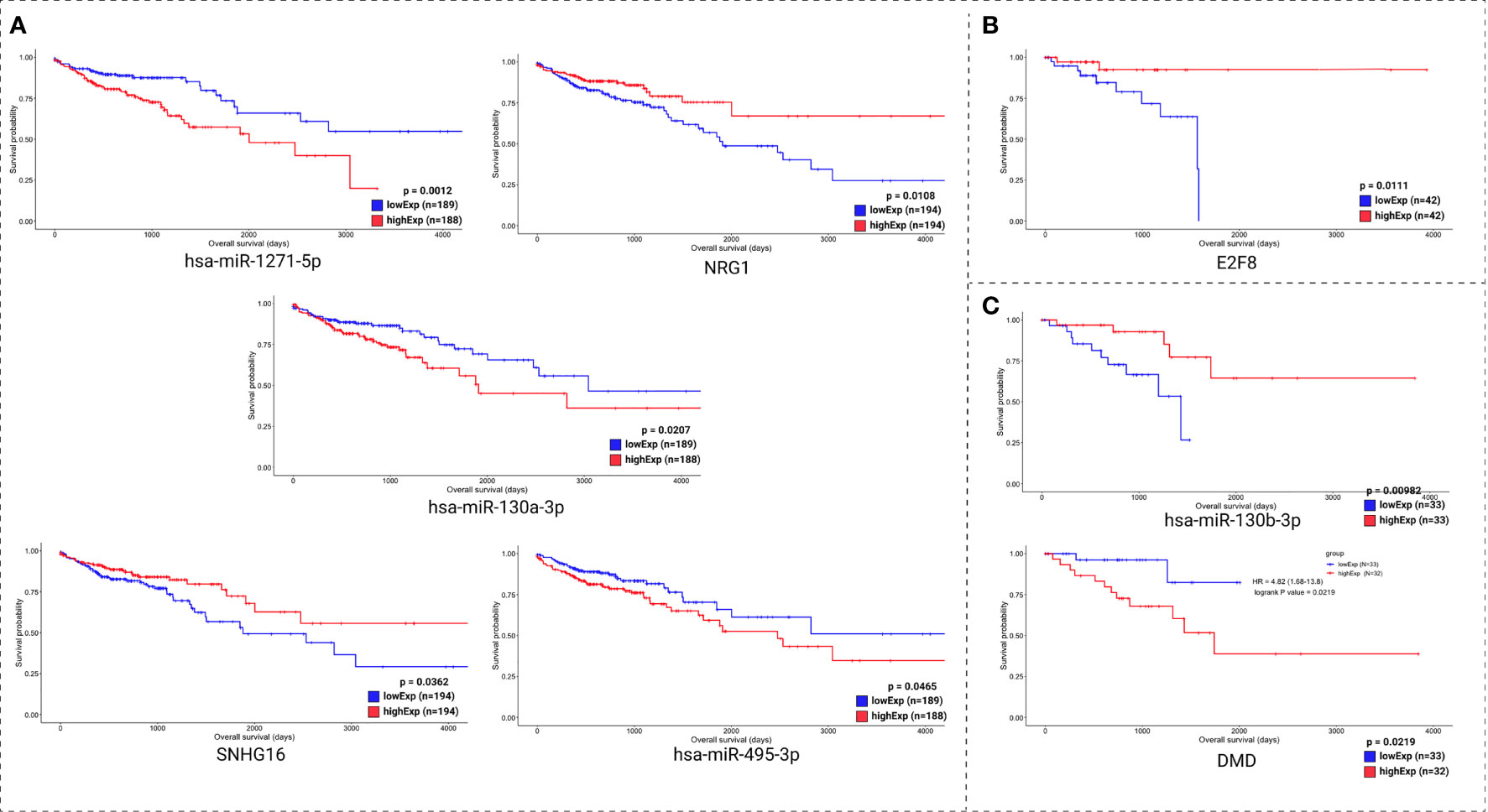
Figure 8 Kaplan-Meier survival curves for the best scored molecules with top HR from CoxPH for colon (A), rectum (B), and rectosigmoid junction (C) sites.
Regarding the KM method (Figure 7), we found some PCGs that were not in the group of 20 molecules found with CoxPH (Figure 6), but are relevant for overall patient survival, such as RPS6KA5 for colon, DMXL1 for rectum, and AGAP3 for rectosigmoid junction.
Regarding the CoxPH method (Figure 7), we identified molecules that can be considered potential biomarkers for patient prognosis. The CoxPH methodology also revealed several molecules to be significant for overall survival in each anatomical site (Figure 8), many of which are concurrent with KM method results, such as has-miR-1271-5p, E2F8, and hsa-miR-130-3p.
Most of the relevant molecules for colon cancer patient survival (Figures 7 and 8) are present in distinct regions of the ceRNA network, with the exception of miR-1275-5p and NRG1. These two molecules are connected by MALAT1 ceRNA (Figure 9).
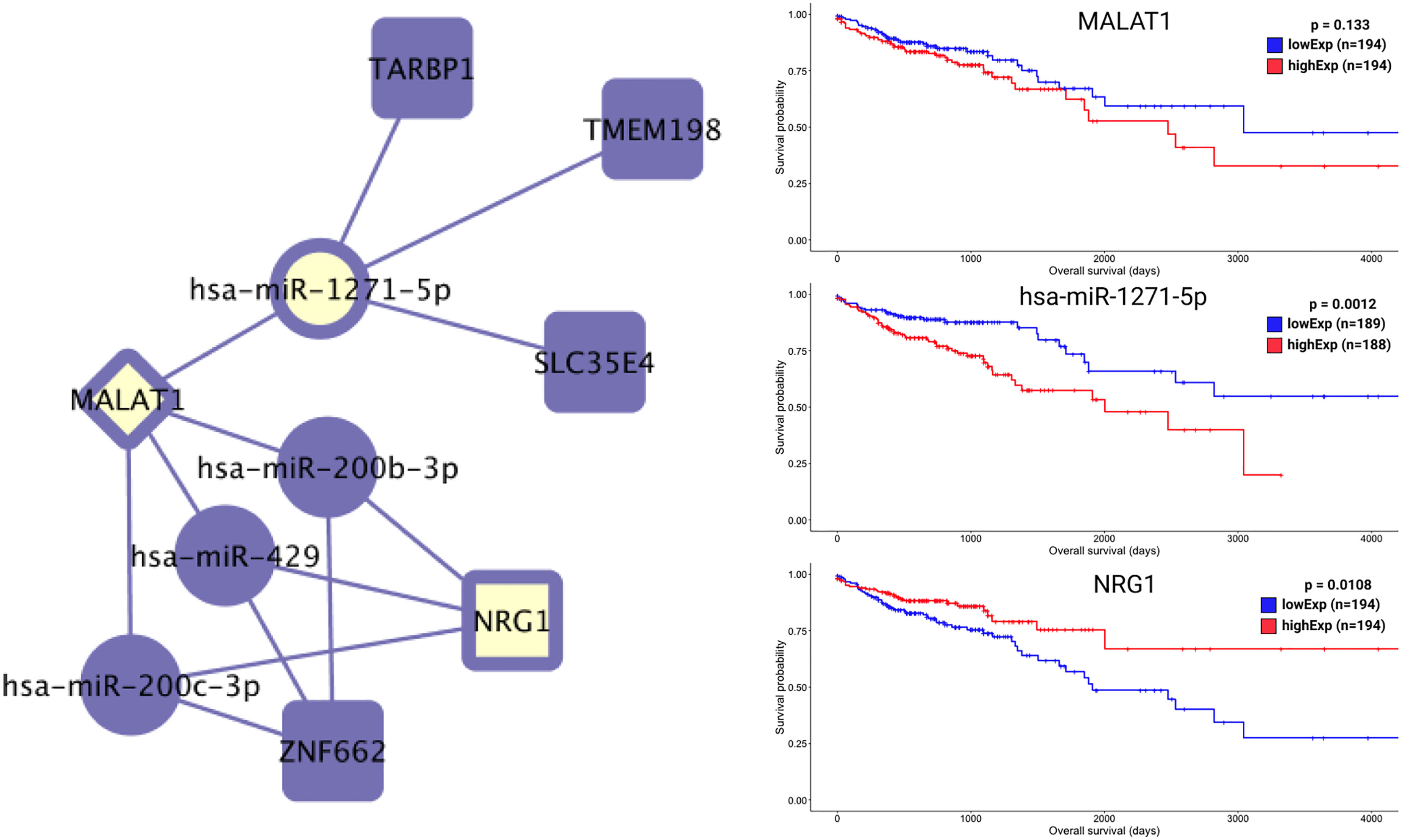
Figure 9 Colon-specific ceRNA network and overall survival for hsa-mir-1271-5p, NRG1 and MALAT1. Here, we can see the opposing putative roles of hsa-mir-1271-5p and NRG1 on overall survival time. MALAT1 is shown as a link that relates hsa-mir-1271-5p and NRG1 regulation, as well as behaving like hsa-mir-1271-5p on patient overall survival time.
Discussion
CRC is one of the most common and lethal cancers worldwide. The treatment therapy is directly related to tumor location. Due to heterogeneous characteristics, tumors located in the rectosigmoid junction are treated as either colon or rectum tumors. Therefore, obtaining molecular markers that could help identify tumor site and molecular characteristics is necessary. In this sense, ceRNA networks allow us to evaluate differentially expressed molecules as well as miRNA-lncRNA-PCG interactions and control mechanisms.
In this study, we used differentially expressed lncRNAs, PCGs, and miRNAs to build specific ceRNA networks for the anatomical sites most affected by CRC and identify different and common factors involved in the progression of CRC. Moreover, we further accessed the relevance of each of these molecules in the ceRNA in regard to their impact on prognosis and their functional implications.
Previous works (11, 15, 16) have reported a protagonist role for the lncRNA H19 as protagonist in regulating various cancer-related mRNAs in colon cancer and in CRC in general. However, in our analysis, H19 is only present in the colon exclusive network and not in that of the rectum, also described by Zhiyuan et al. (14). In this network, H19 presents itself as a risk factor and acts as ceRNA for SOX12, ANKRD6, STC1, and hsa-miR-130a-3p, all of which are present as putative risk factors.
Colon, rectum and rectosigmoid junction presented common ceRNA networks regulated by the lncRNAs MAGI2-AS3, HAGLR-AS3, SNHG1, and SNHG15, suggesting some similarities in CRC development independent of the anatomical site (Figure 3). These common mechanisms are related to the regulation of Wnt signaling, cell morphogenesis, and proliferation (Figure 5A), which are known to be present in cancer. The lncRNAs regulating common ceRNA networks were already correlated with known cancer pathways, such as: MAGI2-AS3 with cell apoptosis and proliferation in CRC (25); HAGLR-AS3 with cell proliferation, invasion, and apoptosis (26); SNHG1 with cell growth and promotion of CRC through the Wnt/β-catenin signaling pathway (27, 28); and SNHG15 with cell proliferation, apoptosis, and activation of the Wnt/β-catenin signal in CRC (29, 30). These studies indicate MAGI2-AS3, HAGLR-AS3, SNHG1, and SNHG15 as potential biomarkers involved in regulation of Wnt signaling, cell morphogenesis, and proliferation. Our study is the first to bring together all of these molecules and related ceRNA networks as common factors in all CRC sites and to indicate their joint use as potential biomarkers for colon, rectum, and rectosigmoid junction cancer common behavior. Within the MAGI2-AS3 network, we found the dystrophin gene (DMD). DMD plays a special role in muscle fiber integrity (31), and it was the only gene identified as a potentially significant risk factor in both rectum and rectosigmoid junction sites. Duchenne muscular dystrophy is a disease known to be associated with DMD, and our functional analysis relates the biological disease’s pathways from DO to the rectum ceRNA (Figure 4B). This gene is part of a network where it is regulated by miRNAs hsa-miR-374a-5p and hsa-miR-374b-5p, and the lncRNA MAGI2-AS3. These three ncRNAs connected to DMD are also ‘sponged’ by the PCG FOXO1, which is critical to tumor suppression and apoptosis (32) and presented a putative protective role in colon CRC tumors. Although Zhong et al. (11) previously reported their interaction, the authors did not mention the DMD and FOXO1 genes, nor did they evaluate their putative role as biomarkers or as survival factors. Therefore, to the best of our knowledge, this is the first time that DMD is reported as a potential biomarker for poor prognosis in CRC. In the case of the rectosigmoid junction, we found DMD and hsa-miR-130b-3p to be relevant to patient prognosis. Some studies have reported the importance of hsa-miR-130b-3p in poor prognosis of CRC (33, 34). It is worth noting that hsa-miR-130b-3p, which is relevant to the rectosigmoid junction is in the same ceRNA network as hsa-miR-130a, which is relevant to colon prognosis. Both molecules are regulated by the lncRNA MIR17HG, which may indicate that this ceRNA network is relevant to both colon and rectosigmoid junction. However, the miRNA responsible for poor patient prognosis is different for each site.
The specific networks for colon and rectum present distinct enriched biological pathways, with more specific endothelial development in the colon and cell morphology in the rectum. Due to the low number of samples for the rectosigmoid junction, we were unable to find a statistically significant pathway for this network. However, the pathways found are related to phosphorylation and signal transduction. These different biological pathways highlight differences in CRC behavior between distinct anatomical sites, thus reinforcing the importance of correctly identifying the tumor site.
E2F8 and RFWD3 presented putative protective roles for rectum CRC tumors. E2F8 encodes transcription factors that regulate development by the cell cycle (35), and RFWD3 is known to be essential in the process of repairing DNA interstrand cross-links (36). Both genes are connected with the lncRNA SNHG1 but are regulated by different miRNA. The SNHG1 ceRNA network is common for all CRC sites, but only interacts with RFWD3 and E2F8 in the rectum, indicating a potential role for this network in rectum cancer. The E2F8 gene has been reported as relevant to CRC as well as in regulating cancer progression (35, 37) and our survival analysis indicates better survival for high E2F8 expression levels. Previous studies (35, 37) have identified E2F8 as a biomarker for colon cancer, but they did not evaluate the potential role of SNHG1-RFWD3-E2F8 ceRNA network interaction in rectum cancer.
The RPS6KA5 gene encodes for a tyrosine kinase and has been indicated as a biomarker for colon cancer (38) through interaction with hsa-miR-130a (39). In our colon-specific network, the lncRNA MIR17HG sponges hsa-miR-130a and interacts with RPS6KA5. Hsa-miR-1271-5p, hsa-miR-130a, SOX12, ANKRD6, TMEM198, STC1, H19, and NRG1 all presented potential risk factors for colon cancer. Most of these molecules are present in distinct regions of the ceRNA network, with the exception of miR-1275-5p and NRG1. Both of these molecules are connected to the lncRNA MALAT1 and present opposing putative roles (Figure 9). Some studies (11, 15, 16) have previously reported the effects of H19 ceRNA on CRC, but both our network and survival analyses suggest its influence only in the case of tumors located in the colon. No enrichment pathway of the rectosigmoid junction presented an exclusive HR relevant molecule.
In further consideration of the overall survival evidence, we reaffirm the potential role as prognosis biomarkers for hsa-miR-1271-5p, NRG1, hsa-miR-130a-3p, SNHG16, and hsa-miR-495-3p, in the colon; E2F8, in the rectum; and of DMD and hsa-miR-130b-3p, in the rectosigmoid junction.
This study had some limitations. Firstly, although several novel lncRNAs, PCGs, and miRNAs with clinical significance for CRC were found, the study was performed with TCGA data and no further experimental validation was carried out. Secondly, less information was analyzed for rectum and rectosigmoid junction tissue than for that of colon, which could influence site-specific results. Research on ceRNAs in CRC is still in development and requires further experimental studies and greater amount of data from colon, rectum, and rectosigmoid cancer in order to improve our understanding of the biomarkers found.
In conclusion, this study constructed a ceRNA network for colon, rectum, and rectosigmoid that provides clinical significance and functional implications for cancer at each of these sites. The results indicate several potential prognostic markers for colon, rectum, and rectosigmoid cancer, and also suggest that the ceRNAs found can help explain the differences between, and common factors on, prognosis for these CRC sites.
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Author Contributions
LV contributed to conception and design of the study, wrote the manuscript, and performed the analysis. NJ collaborated and reviewed the study on the bioinformatics methods and biology assumptions. JCS, MW, and PS reviewed and collaborated to key points at the discussion and methodology on a bioinformatics perspective. JBS reviewed and collaborated to key points at the discussion and methodology on a medical perspective. All authors contributed to the article and approved the submitted version.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The results shown here are in whole or part based upon data generated by the TCGA Research Network: https://www.cancer.gov/tcgahttps://www.cancer.gov/tcga. This study was financed in part by the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior – Brasil (CAPES) – Finance Code 001 and the Free State of Saxony (EraPerMed proj. no. 100371715). We acknowledge support from Leipzig University for Open Access Publishing.
References
1. Qiaowei F, Bingrong L. Comprehensive Analysis of a Long Noncoding RNA-Associated Competing Endogenous RNA Network in Colorectal Cancer. OncoTargets Ther (2018) 11:2453. doi: 10.2147/OTT.S158309
2. Ulrich N, Anina Z, Christoph S, Tara M, Matthias M, Tibor S, et al. Mucinous and Signet-Ring Cell Colorectal Cancers Differ From Classical Adenocarcinomas in Tumor Biology and Prognosis. Ann Surg (2013) 258:775. doi: 10.1097/SLA.0b013e3182a69f7e
3. Christian W, James B, Anne L, Elisabeth E. TNM Supplement: A Commentary on Uniform Use. Hoboken, New Jersey: John Wiley & Sons (2019).
4. Ruud L, Marcel F, Gerwin F, Frank B, Aart B. The Localisation of Cancer in the Sigmoid, Rectum or Rectosigmoid Junction Using Endoscopy or Radiology—What Is the Most Accurate Method? J Gastrointest Oncol (2014) 5:469. doi: 10.3978/j.issn.2078-6891.2014.087
5. Xu G, Zheng J, Tianyi M, Zheng L, Hanqing H, Zhixun Z, et al. Radiotherapy Dose Led to a Substantial Prolongation of Survival in Patients With Locally Advanced Rectosigmoid Junction Cancer: A Large Population Based Study. Oncotarget (2016) 7:28408. doi: 10.18632/oncotarget.8630
6. Pieter-Jan V, Kenneth V, Gerben M, Klaas V, Lennart M, Jo V, et al. An Update on Lncipedia: A Database for Annotated Human lncRNA Sequences. Nucleic Acids Res (2015) 43:D174–80. doi: 10.1093/nar/gku1060
7. Leonardo S, Laura P, Yvonne T, Lev K, Paolo PP. A Cerna Hypothesis: The Rosetta Stone of a Hidden RNA Language? Cell (2011) 146:353–8. doi: 10.1016/j.cell.2011.07.014
8. Tayier T, Linlin L, Zumureti A, Shumei F. Comprehensive Analysis of the Aberrantly Expressed Lncrna-Associated Cerna Network in Breast Cancer. Mol Med Rep (2019) 19:4697–710. doi: 10.3892/mmr.2019.10165
9. Gang Z, Shuwei L, Jiafei L, Yuqiu G, Qiaoyan W, Gaoxiang M, et al. Lncrna MT1JP Functions as a Cerna in Regulating FBXW7 Through Competitively Binding to Mir-92a-3p in Gastric Cancer. Mol Cancer (2018) 17:1–11. doi: 10.1186/s12943-018-0829-6
10. Yunpeng Z, Yanjun X, Li F, Feng L, Zeguo S, Tan W, et al. Comprehensive Characterization of Lncrna-Mrna Related Cerna Network Across 12 Major Cancers. Oncotarget (2019) 7:64148. doi: 10.18632/oncotarget.11637
11. Min-Er Z, Yanyu C, Guannan Z. Lncrna H19 Regulates PI3K–Akt Signal Pathway by Functioning as a Cerna and Predicts Poor Prognosis in Colorectal Cancer: Integrative Analysis of Dysregulated ncRNA-Associated Cerna Network. Springer (2019) 19:148. doi: 10.1186/s12935-019-0866-2
12. Jun-Hao L, Shun L, Hui Z, Liang-Hu Q, Jian-Hua Y. Lncrna CACS15 Contributes to Oxaliplatin Resistance in Colorectal Cancer by Positively Regulating ABCC1 Through Sponging Mir-145. Arch Biochem Biophys (2019) 663:183–91. doi: 10.1016/j.abb.2019.01.005
13. Qian-Rong H, Pan XB. Prognostic lncRNAs, miRNAs, and mRNAs Form a Competing Endogenous RNA Network in Colon Cancer. Frontiers (2019) 9:712. doi: 10.3389/fonc.2019.00712
14. Zhiyuan Z, Sen W, Dongjian J, Wenwei Q, Qingyuan W, Jie L, et al. Construction of a Cerna Network Reveals Potential lncRNA Biomarkers in Rectal Adenocarcinoma. Oncol Rep (2018) 39:2101–13. doi: 10.3892/or.2018.6296
15. Hongda P, Jingxin P, Shibo S, Lei J, Hong L, Zhangru Y. Identification and Development of Long non-Coding RNA-Associated Regulatory Network in Colorectal Cancer. J Cell Mol Med (2019) 23:5200–10. doi: 10.1111/jcmm.14395
16. Hui Z, Zhuo W, Jianzhong W, Rong M, Jifeng F. Long Noncoding Rnas Predict the Survival of Patients With Colorectal Cancer as Revealed by Constructing an Endogenous RNA Network Using Bioinformation Analysis. Cancer Med (2019) 8:863–73. doi: 10.1002/cam4.1813
17. Katarzyna T, Patrycja C, Maciej W. The Cancer Genome Atlas (TCGA): An Immeasurable Source of Knowledge. Contemp Oncol (2015) 19:A68. doi: 10.5114/wo.2014.47136
18. Ruidong L, Han Q, Shibo W, Julong W. Gdcrnatools: An R/Bioconductor Package for Integrative Analysis of lncRNA, Mirna and Mrna Data in GDC. Bioinformatics (2018) 34:2515–7. doi: 10.1093/bioinformatics/bty124
19. Matthew R, Belinda P DIW, Yifang H, Charity L, Wei S, et al. Limma Powers Differential Expression Analyses for Rna-Sequencing and Microarray Studies. Nucleic Acids Res (2015) 43:e47–7. doi: 10.1093/nar/gkv007
20. Pedro F, Sonia T, Toni G, Anton E, Ana C. Spongescan: A Web for Detecting Microrna Binding Elements in lncRNA Sequences. Nucleic Acids Res (2016) 44:W176–80. doi: 10.1093/nar/gkw443
21. Jun-Hao L, Shun L, Hui Z, Liang-Hu Q, Jian-Hua Y. Starbase V2. 0: Decoding Mirna-Cerna, Mirna-Ncrna and Protein–RNA Interaction Networks From Large-Scale CLIP-Seq Data. Nucleic Acids Res (2014) 42:D92–7. doi: 10.1093/nar/gkt1248
22. Michael A, Catherine B, Judith B, David B, Heather B, Michael CJ, et al. Gene Ontology: Tool for the Unification of Biology. Nat Genet (2000) 25:25–9. doi: 10.1038/75556
23. Minoru K, Susumu G. KEGG: Kyoto Encyclopedia of Genes and Genomes. Nucleic Acids Res (2000) 28:27–30. doi: 10.1093/nar/28.1.27
24. Lynn S, Elvira M, James M, Becky T, Mike S, Lance N, et al. Human Disease Ontology 2018 Update: Classification, Content and Workflow Expansion. Nucleic Acids Res (2019) 47:D955–62. doi: 10.1093/nar/gky1032
25. Yuping Z, Bo L, Zhuo L, Lai J, Gang W, Min L, et al. Long Noncoding MAGI2-AS3 Promotes Colorectal Cancer Progression Through Regulating Mir-3163/Tmem106b Axis. J Cell Physiol (2019) 235:4824–33. doi: 10.1002/jcp.29360
26. Weixuan S, Wenting N, Zhaoyi W, Haolong Z, Yezhou L, Xuedong F. Lnc HAGLR Promotes Colon Cancer Progression Through Sponging Mir-185-5p and Activating CDK4 and CDK6 In Vitro and In Vivo. OncoTargets Ther (2020) 13:5913. doi: 10.2147/OTT.S246092
27. Mu X, Xiaoxiang C, Kang L, Kaixuan Z, Xiangxiang L, Bei P, et al. The Long Noncoding RNA SNHG1 Regulates Colorectal Cancer Cell Growth Through Interactions With EZH2 and Mir-154-5p. Mol Cancer (2019) 17:1–16. doi: 10.1186/s12943-018-0894-x
28. Yuping Z, Bo L, Zhuo L, Lai J, Gang W, Min L, et al. Up-Regulation of lncRNA Snhg1 Indicates Poor Prognosis and Promotes Cell Proliferation and Metastasis of Colorectal Cancer by Activation of the Wnt/β-Catenin Signaling Pathway. Oncotarget (2017) 8:111715. doi: 10.18632/oncotarget.22903
29. Min L, Zehua B, Guoying J, Jia Z, Surui Y, Yuyang F, et al. Lnc RNA-SNHG 15 Enhances Cell Proliferation in Colorectal Cancer by Inhibiting Mir-338-3p. Cancer Med (2019) 8:2404–13. doi: 10.1002/cam4.2105
30. Min L, Zehua B, Guoying J, Jia Z, Surui Y, Yuyang F, et al. Long Noncoding RNA SNHG15 Enhances the Development of Colorectal Carcinoma Via Functioning as a Cerna Through Mir-141/Sirt1/Wnt/β-Catenin Axis. Artif Cells Nanomed Biotechnol (2019) 47:2536–44. doi: 10.1080/21691401.2019.1621328
31. Sylvie TG, Julie M, Michel K, Mireille C. Normal and Altered Pre-Mrna Processing in the DMD Gene. Hum Genet (2017) 136:1155–72. doi: 10.1007/s00439-017-1820-9
32. Yun-Cheol C, Ji-Young K, Woo PJ, Kee-Beom K, Hyein O, Kyung-Hwa L, et al. Foxo1 Degradation Via G9a-Mediated Methylation Promotes Cell Proliferation in Colon Cancer. Nucleic Acids Res (2019) 47:1692–705. doi: 10.1093/nar/gky1230
33. Tommaso C, Alessandra F, Carolina V, Lina S, Massimo P, Carmelo L, et al. Microrna-130b Promotes Tumor Development and is Associated With Poor Prognosis in Colorectal Cancer. Neoplasia (2013) 15:1086–99. doi: 10.1593/neo.13998
34. Yanyang Z, Gang M, Yao L, Tomoya I, Jianguo G, Jian L, et al. Microrna 130b Suppresses Migration and Invasion of Colorectal Cancer Cells Through Downregulation of Integrin β1. PloS One (2014) 9:e87938. doi: 10.1371/journal.pone.0087938
35. Zhiyuan Z, Jie L, Huang Y, Wen P, Wenwei Q, Jiou G, et al. Upregulated Mir-1258 Regulates Cell Cycle and Inhibits Cell Proliferation by Directly Targeting E2F8 in CRC. Cell Prolif (2018) 51:e12505. doi: 10.1111/cpr.12505
36. Yo-Chuen L, Yating W, Rosaline H, Sumanprava G, Susan W, K AM, et al. PCNA-Mediated Stabilization of E3 Ligase RFWD3 At the Replication Fork is Essential for Dna Replication. Proc Natl Acad Sci (2018) 115:13282–7. doi: 10.1073/pnas.1814521115
37. Peng-yu Y, Xin-an Z. Knockdown of E2F8 Suppresses Cell Proliferation in Colon Cancer Cells by Modulating the Nf-κb Pathway. Ann Clin Lab Sci (2019) 49:474–80.
38. Peng S, Jianping W, Yao T, Chunxia X, Xingguo Z. Gene Pair Based Prognostic Signature for Colorectal Colon Cancer. Medicine (2018) 97:2–3. doi: 10.1097/MD.0000000000012788
Keywords: colorectal cancer, competing endogenous RNA, TCGA, long non-coding RNA, miRNA, mRNA
Citation: Vieira LM, Jorge NAN, de Sousa JB, Setubal JC, Stadler PF and Walter MEMT (2021) Competing Endogenous RNA in Colorectal Cancer: An Analysis for Colon, Rectum, and Rectosigmoid Junction. Front. Oncol. 11:681579. doi: 10.3389/fonc.2021.681579
Received: 16 March 2021; Accepted: 22 April 2021;
Published: 10 June 2021.
Edited by:
Peti Thuwajit, Mahidol University, ThailandReviewed by:
Nicolò Musso, University of Catania, ItalyAli M. Ardekani, Avicenna Research Institute (ARI), Iran
Copyright © 2021 Vieira, Jorge, de Sousa, Setubal, Stadler and Walter. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Lucas Maciel Vieira, bHVjYXMubWFjaWVsLnZpZWlyYUBnbWFpbC5jb20=