 Jian Hu†
Jian Hu† Dongxu Qiu
Dongxu Qiu Anze Yu
Anze Yu Jiao Hu
Jiao Hu Hao Deng
Hao Deng Huihuang Li
Huihuang Li Zhenglin Yi
Zhenglin Yi Jinbo Chen
Jinbo Chen Xiongbing Zu
Xiongbing Zu
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Oncol. , 06 May 2021
Sec. Cancer Molecular Targets and Therapeutics
Volume 11 - 2021 | https://doi.org/10.3389/fonc.2021.607224
This article is part of the Research Topic Targeting the Microenvironment Niche in Solid and Hematologic Malignancies: Basic and Translational Research View all 12 articles
Background: YTH N6-methyladenosine RNA binding protein 1 (YTHDF1) has been indicated proven to participate in the cross-presentation of tumor antigens in dendritic cells and the cross-priming of CD8+ T cells. However, the role of YTHDF1 in prognosis and immunology in human cancers remains largely unknown.
Methods: All original data were downloaded from TCGA and GEO databases and integrated via R 3.2.2. YTHDF1 expression was explored with the Oncomine, TIMER, GEPIA, and BioGPS databases. The effect of YTHDF1 on prognosis was analyzed via GEPIA, Kaplan-Meier plotter, and the PrognoScan database. The TISIDB database was used to determine YTHDF1 expression in different immune and molecular subtypes of human cancers. The correlations between YTHDF1 expression and immune checkpoints (ICP), tumor mutational burden (TMB), microsatellite instability (MSI), and neoantigens in human cancers were analyzed via the SangerBox database. The relationships between YTHDF1 expression and tumor-infiltrated immune cells were analyzed via the TIMER and GEPIA databases. The relationships between YTHDF1 and marker genes of tumor-infiltrated immune cells in urogenital cancers were analyzed for confirmation. The genomic alterations of YTHDF1 were investigated with the c-BioPortal database. The differential expression of YTHDF1 in urogenital cancers with different clinical characteristics was analyzed with the UALCAN database. YTHDF1 coexpression networks were studied by the LinkedOmics database.
Results: In general, YTHDF1 expression was higher in tumors than in paired normal tissue in human cancers. YTHDF1 expression had strong relationships with prognosis, ICP, TMB, MSI, and neoantigens. YTHDF1 plays an essential role in the tumor microenvironment (TME) and participates in immune regulation. Furthermore, significant strong correlations between YTHDF1 expression and tumor immune-infiltrated cells (TILs) existed in human cancers, and marker genes of TILs were significantly related to YTHDF expression in urogenital cancers. TYHDF1 coexpression networks mostly participated in the regulation of immune response and antigen processing and presentation.
Conclusion: YTHDF1 may serve as a potential prognostic and immunological pan-cancer biomarker. Moreover, YTHDF1 could be a novel target for tumor immunotherapy.
N6-methyladenine (m6A), occurring in the N6-position of adenosine, is the most abundant modification of mRNA and noncoding RNAs in mammals (1–3). Moreover, it plays an essential role in RNA nucleation, RNA-protein interaction, mRNA metabolism, mRNA stability and splicing, and mRNA translation (4, 5). Under normal conditions, m6A molecules modify the processing of RNA by serving as “writers”, “erasers”, and “readers” (6, 7). However, abnormal expression of m6A molecules has been proven to promote tumorigenesis by affecting oncogene expression (8–10). Recent evidence reveals that m6A produces a marked effect on cancer development, such as proliferation, migration, and invasion (11), which means that m6A molecules may be promising prognostic biomarkers and therapeutic targets.
YTH N6-methyladenosine RNA binding protein 1 (YTHDF1) belongs to the YTH domain family, and acts as a “reader” of m6A-modified mRNAs. In the cytosol, YTHDF1 plays an essential role in RNA translation initiation by binding to initiation factors (4). Previous studies proved that tumor neoantigens could cause spontaneous antitumor immune responses and predict the response to cancer immunotherapies (12, 13). Recent studies have demonstrated that YTHDF1 plays a crucial role in cancer development and durable neoantigen-specific antitumor immunity (4). For example, YTHDF1 promotes the progression of lung cancer by participating in the m6A demethylase the ALKBH5 pathway (14). In colorectal cancer, YTHDF1 is regarded as an oncogene (15); In breast cancer, overexpression of YTHDF1 predicts worse survival (16). Furthermore, YTHDF1 inhibits the function of antitumor immune cells in the tumor microenvironment (TME) by acting on the infiltrated level of CD8+ T cells and natural killer cells. Blocking the YTHDF1 reactivates the inhibited antitumor immunity and synergistically improves the therapeutic efficacy of anti-PD-L1 inhibitors (17). All of the above studies indicate that YTHDF1 might be a promising prognostic and therapeutic pan-cancer biomarker.
However, the role of YTHDF1 in prognosis and immunology in human cancers has seldomly been analyzed systematically. In this study, the role of YTHDF1 in prognosis and immunology in human cancers was comprehensively analyzed. Furthermore, the potential association between YTHDF1 expression and immune subtypes, molecular subtypes of different cancer types, promising immune biomarkers and tumor-infiltrating lymphocytes (TILs) in the TME was analyzed. In addition, we explored the effect of YTHDF1 expression in urogenital cancers to verify the results in human cancers. This study aimed to explore the potential of YTHDF1 in anticancer immunotherapy in human cancer, thus offering insight to a new antitumor strategy.
All original data were downloaded from The Cancer Genome Atlas (TCGA) (https://cancergenome.nih.gov/) and Gene Expression Omnibus (GEO) (https://www.ncbi.nlm.nih.gov/geo/) databases. Detailed information on human cancers and corresponding individual sample sizes is displayed in Supplementary Table 1. R 3.2.3 was used to integrate the original data and verify the results analyzed by the website database. All applied online web tools were introduced below.
The Oncomine database (https://www.oncomine.org/resource/login.html), TIMER database (https://cistrome.shinyapps.io/timer/), and GEPIA database (http://gepia2.cancer-pku.cn/#analysis) were used to compare YTHDF1 expression between human cancers and paired normal tissue (18–20). The BioGPS database (http://biogps.org) was used to analyze the expression profiles of YTHDF1 in different cancer and paired normal cell lines (21). In the Oncomine database, we set the thresholds to 0.001 for P-value and 1.5 for the fold change.
The GEPIA database (http://gepia2.cancer-pku.cn/#analysis), Kaplan-Meier Plotter database (http://kmplot.com/analysis/), and PrognoScan databases (http://dna00.bio.kyutech.ac.jp/PrognoScan/index.html) were used to explore the prognostic value of YTHDF1 expression in human cancers (20, 22, 23). The GEPIA database is an online website, and its analyzed tumor and normal tissue data were from the TCGA database. We used the GEPIA database to explore the correlation between YTHDF1 expression and overall survival (OS) and disease-free survival (DFS) in 33 cancer types. In the GEPIA database, the median YTHDF1 expression was used as a cutoff value to classify groups. The Kaplan-Meier Plotter database classifies groups by calculating an optimal cutoff value automatically. By using the Kaplan-Meier Plotter database, we identified the association between YTHDF1 expression and OS and relapse-free survival (RFS) in 21 cancer types. Hazard ratios (HRs) with corresponding 95% confidence intervals (CIs) and log-rank P-values were calculated. Data on the PrognoScan website were also collected from the TCGA and GEO database. The prognostic value was considered statistically significant when the P-value was less than 0.05.
The TISIDB database (http://cis.hku.hk/TISIDB/index.php) is an online integrated repository portal collecting abundant human cancer datasets from the TCGA database (24). The correlations between YTHDF1 expression and immune or molecular subtypes of different cancer types were explored via the TISIDB database. Differences with a P-value < 0.05 were considered to be statistically significant.
The relationship between YTHDF1 expression and immune checkpoint (ICP) genes, biomarkers and ESTIMATE score in the TME was explored via the SangerBox website (http://sangerbox.com/Tool), which is a useful online platform for TCGA data analysis (25). Immune checkpoint (ICP) genes were selected according to a previous study (26). Tumor mutational burden (TMB), microsatellite instability (MSI), and neoantigens have been proved to be important biomarkers of the TME (27–29). ESTIMATE (Estimation of Stromal and Immune cells in Malignant Tumor tissues using Expression data) was an algorithm designed by Yoshihara et al. for predicting tumor purity in TME, and this algorithm included stromal score (that captures the presence of stroma 136 in tumor tissue), immune score (that represents the infiltration of immune cells in tumor 137 tissue), and estimate score (that infers tumor purity) (30). Correlations between YTHDF1 expression and the above TME biomarkers were explored via the SangerBox website. Differences with a P-value < 0.05 were considered to be statistically significant.
We first explored the correlation between YTHDF1 expression and six immune cells (B cells, CD4+ T cells, CD8+ T cells, neutrophils, macrophages, and dendritic cells) in the TME of 31 human cancers via the SangerBox website. Then, the correlation between YTHDF1 expression and ten immune cells in the TME was explored in six urogenital cancers by the SangerBox website. These ten immune cells included B cells lineage, CB8+ T cells, cytotoxic lymphocytes, endothelial cells, fibroblasts, monocytic cells, myeloid dendritic cells, neutrophils, natural killer cells, and T cells. We reanalyzed the correlation between YTHDF1 expression and tumor infiltrating lymphocytes (TILs) via the TIMER and GEPIA databases to verify the above results. The TIMER database (https://cistrome.shinyapps.io/timer/), which collected 10897 samples across 32 cancer types from TCGA, was created to analyze the level of tumor-associated immune cell infiltration in the TME (19, 31). Then, we verified the correlation between YTHDF1 expression and 24 tumor-infiltrating lymphocytes (TILs) in six urogenital cancers via the GEPIA database (20). These 24 tumor-infiltrating lymphocytes (TILs) were selected from a previous study (32), and detailed information is displayed in Supplementary Table 2. Finally, the TIMER database was used to explore the relationship between YTHDF1 expression and marker genes of the main immune cells, including B cells, CD8+ T cells, follicular helper T cells (Tfh), T-helper 1 (Th1) cells, T-helper 2 (Th2) cells, T-helper 9 (Th9) cells, T-helper 17 (Th17) cells, T-helper 22 (Th22) cells, Tregs cells, exhausted T cells, M1 macrophages, M2 macrophages, tumor-associated macrophages, monocytes, natural killer (NK) cells, neutrophils and dendritic cells (33–35). The difference at a P-value < 0.05 was considered to be statistically significant.
cBio Cancer Genomics Portal (c-BioPortal) (http://cbioportal.org) collected a multidimensional cancer genomics data set (36). Therefore, the c-BioPortal database was applied to explore YTHDF1 genomic alterations in six urogenital cancers.
The UALCAN database (http://ualcan.path.uab.edu) collected RNA-seq and clinical data of 31 cancer types from TCGA (37), and it offered a useful platform to analyze gene expression in tumor and normal tissues. This database was used to analyze the relationship between individual gene expression and clinicopathological features in human cancers.
The LinkedOmics database (http://www.linkedomics.org/login.php) is a visual platform and is used to explore the gene expression profile (38). We used LinkedOmics to determine the YTHDF1 coexpression genes by using Pearson’s correlation coefficient and showed the results via heat maps, and volcano plots. Then, we explored the Gene Ontology biological process (GO_BP), and KEGG pathways of YTHDF1 and its coexpression genes by using gene set enrichment analysis (GSEA).
The Oncomine database showed that YTHDF1 mRNA levels were significantly higher in most human cancers, such as bladder, brain and CNS, breast, cervical, colorectal, esophageal, head and neck, kidney, liver, lung, ovarian, pancreatic, prostate cancer, leukemia, melanoma, sarcoma, and other cancer types compared to the corresponding normal tissues. In contrast, YTHDF1 expression was significantly lower only in lymphoma (Figure 1A). The results from the TIMER database showed that YTHDF1 expression was significantly higher in BLCA (bladder urothelial carcinoma), BRCA (breast invasive carcinoma), CHOL(cholangiocarcinoma), COAD (colon adenocarcinoma), ESCA(esophageal carcinoma), HNSC (head and neck cancer), KICH (kidney chromophobe), KIRP (kidney renal papillary carcinoma), LIHC (liver hepatocellular carcinoma), LUAD (lung adenocarcinoma), LUSC (lung squamous cell carcinoma), PRAD (prostate adenocarcinoma), STAD (stomach adenocarcinoma) and UCEC (uterine corpus endometrial carcinoma) than in adjacent normal tissue. However, YTHDF1 mRNA expression was low only in THCA (thyroid carcinoma) (Figure 1B). The results of the GEPIA database analysis are displayed as supplementary results of cancers without paired normal tissues in the TIMER database. Furthermore, the results showed that YTHDF1 mRNA expression was significantly high among most cancer types except LAML (acute myeloid leukemia), which was consistent with the Oncomine database results (Figure 1C). The expression of YTHDF1 in different cancer cell lines and normal tissues was investigated via the BioGPS database, and we found that YTHDF1 had a high expression level in almost all cancer cell lines. Ten cancer cell lines with the highest YTHDF1 expression level are displayed in Figure 1D. In normal cells, the YTHDF1 expression level in immune cells was the highest (Figure 1E). Detailed information is shown in Supplementary Figure 1. The above results suggested that YTHDF1 was overexpressed in cancer tissues and might participate in the process of immune regulation.
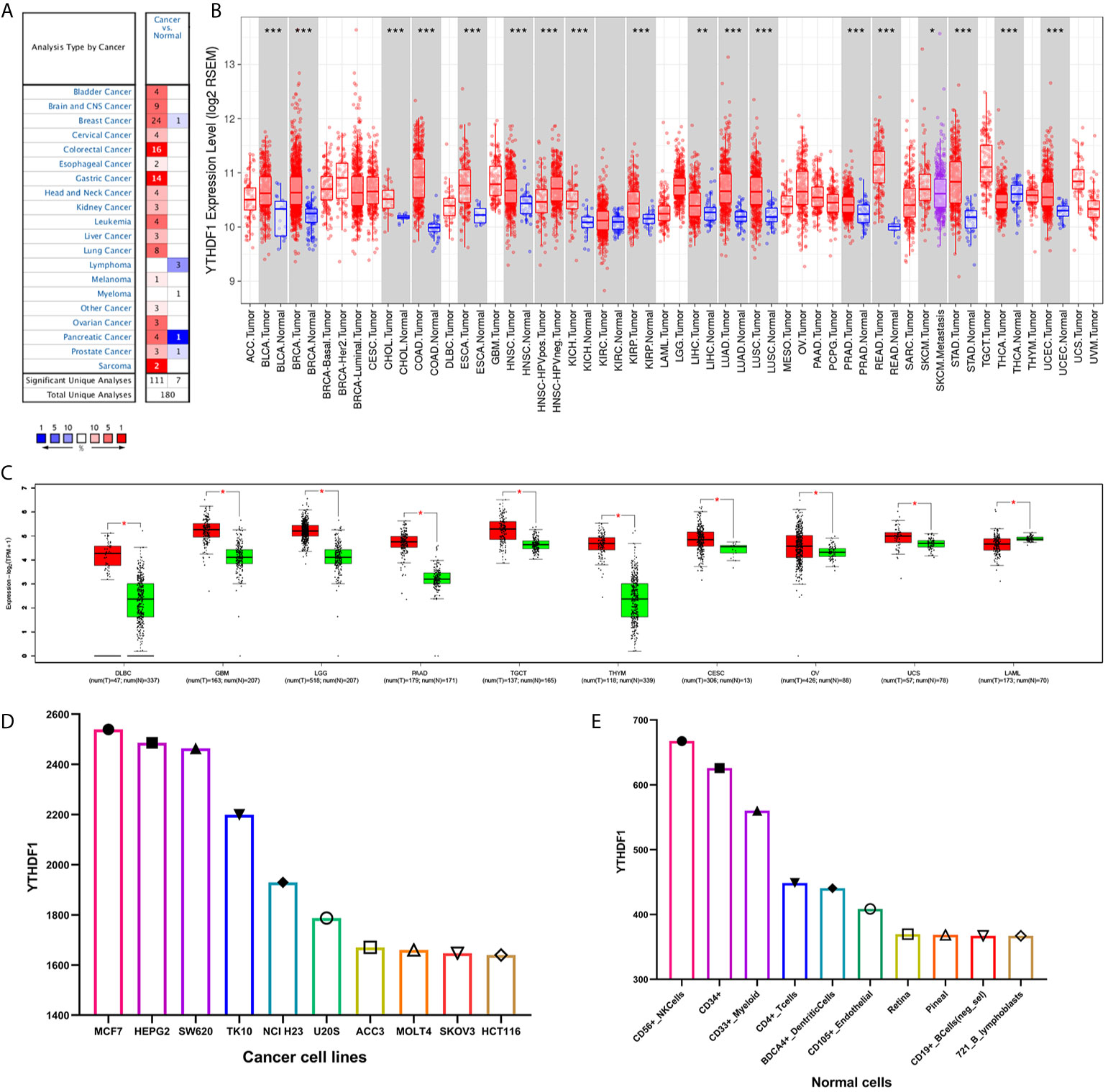
Figure 1 YTHDF1 expression levels in human cancers. (A) YTHDF1 expression in different cancers and paired normal tissue in the Oncomine database. (B) YTHDF1 expression levels in different cancer types from the TCGA database analyzed by the TIMER database. (*P < 0.05, **P < 0.01, ***P < 0.001). (C) YTHDF1 expression in several cancers and paired normal tissue in the GEPIA database. (D) the expression of YTHDF1 in different cancer cell lines analyzed by the BioGPS database. (E) the expression of YTHDF1 in normal tissue analyzed by the BioGPS database.
The prognostic value of YTHDF1 expression in human cancers was analyzed by several databases. In GEPIA, we found that higher YHTDF1 expression was associated with poorer overall survival (OS) and disease-free survival (DFS) in LIHC (n = 364, OS: HR = 1.8, P = 0.0015; n = 364, DFS: HR = 1.6, P = 0.0018; Figures 2A, B), and UVM (uveal melanoma) (n = 78, OS: HR = 3.1, P = 0.027; n = 78, DFS: HR = 2.7, P = 0.034; Figures 2C, D). In addition, patients with higher YTHDF1 expression had poorer OS in mesothelioma (MESO) (n = 82, HR = 1.6, P = 0.049; Figure 2E) and uterine carcinosarcoma (UCS) (n = 740, HR = 2.2, P = 0.00097; Figure 2F), and also had poor DFS in BLCA (n = 400, HR = 1.5, P = 0.025; Figure 2G) and adrenocortical carcinoma (ACC) (n = 76, HR = 2, P = 0.046; Figure 2H). However, higher YTHDF1 expression was related to better OS and DFS in KIRC (kidney renal papillary cell carcinoma) (n = 516, OS: HR = 0.71, P = 0.031; n = 516, DFS: HR = 0.55, P = 0.0015; Figures 2I, J). In the Kaplan-Meier plotter database, higher YTHDF1 expression was associated with poorer RFS in TGCT (testicular germ cell tumors) (n = 105, HR = 5.37, P = 0.011; Figure 2K) and KIRP (n = 183, HR = 3.56, P = 0.027; Figure 2L). More details of the relationship between YTHDF1 and OS or relapse-free survival (RFS) analyzed by the Kaplan-Meier plotter database are shown in Supplementary Figure 2. Furthermore, the correlation between YTHDF1 expression and survival in GEO datasets was evaluated by PrognoScan. Higher expression of YTHDF1 showed worse survival in breast, soft tissue, bladder, lung and brain cancer. Details are shown in Supplementary Table 3. The above results proved that YTHDF1 expression closely related to the prognosis of various cancer types.
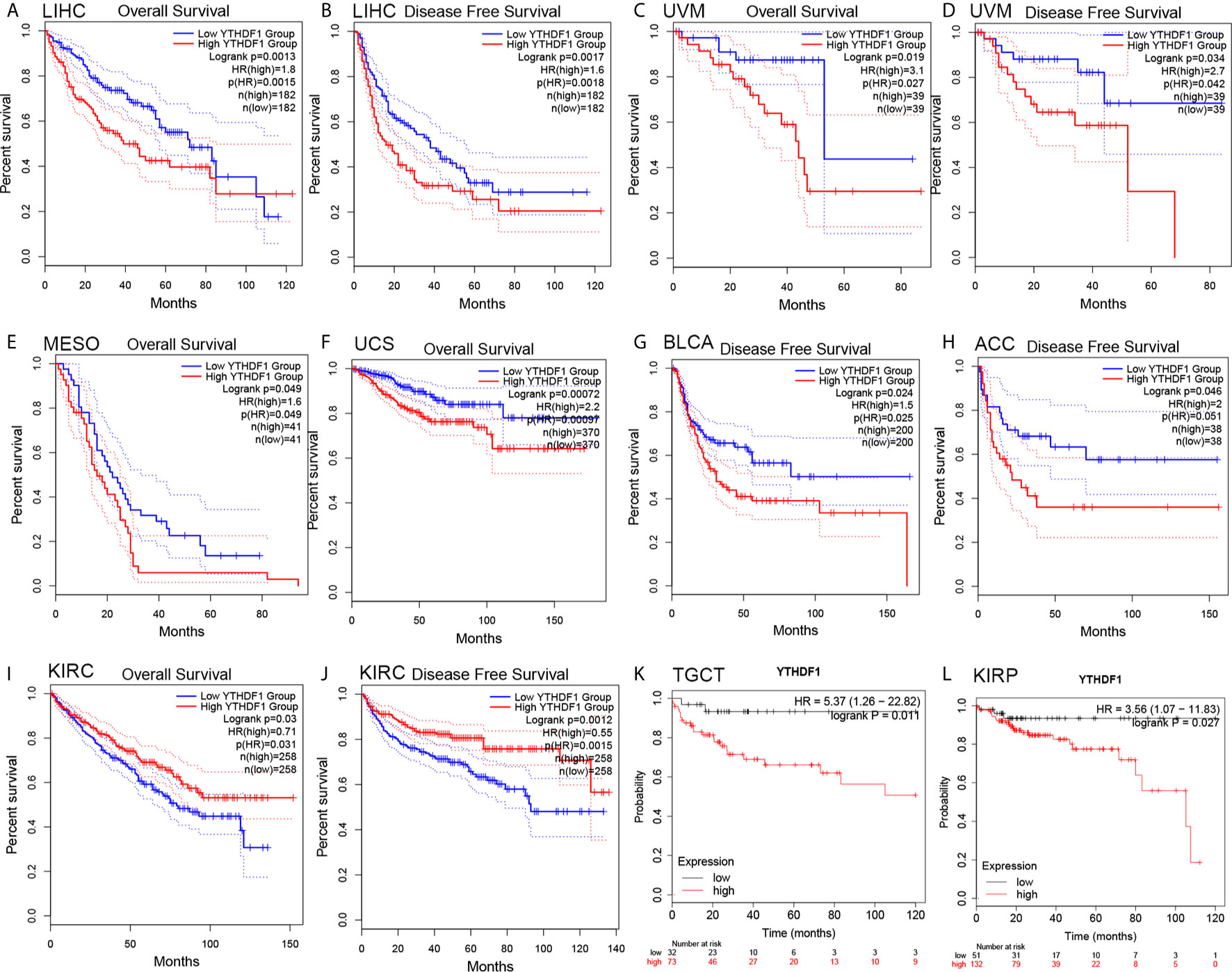
Figure 2 Kaplan-Meier survival curve of human cancers with high and low YTHDF1 expression analyzed by the GEPAI database (A–J) and the Kaplan-Meier plotter database (K, L). (A–D) High YTHDF1 expression was related to worse OS and DFS in LIHC (n = 364) and UVM cohorts (n = 78). (E, F) High YTHDF1 expression was related to worse OS in MESO (n = 82) and UCS cohorts (n = 740). (G, H) High YTHDF1 expression was related to worse DFS in BLCA (n = 400) and ACC cohorts (n = 76). (I, J) High YTHDF1 expression was related to better OS and DFS in KIRC cohorts (n = 316). (K, L) higher YTHDF1 expression was related to poorer RFS in TGCT (n = 105) and KIRP (n = 183). OS, overall survival; DFS, disease free survival; RFS, relapse-free survival.
Next, the role of YTHDF1 expression on immune and molecular subtypes among human cancers was explored with the TISIDB website. Immune subtypes were classified into six types, including C1 (wound healing), C2 (IFN-gamma dominant), C3 (inflammatory), C4 (lymphocyte depleted), C5 (immunologically quiet) and C6 (TGF-b dominant). The results showed that YTHDF1 expression was related to different immune subtypes in BRCA, COAD, HNSC, KIRC, LGG (brain lower grade glioma), LUAD, LUSC, OV (ovarian serous cystadenocarcinoma), PRAD, SKCM (skin cutaneous melanoma), STAD, and UCEC (Figure 3). Additionally, YTHDF1 expression differed in different immune subtypes of one cancer type. Taking KIRC as an example, YTHDF1 showed high expression in C1 and C5 types and low expression in C2 types. For different molecular subtypes of cancers, a significant connection with YTHDF1 expression existed in BRCA, COAD, ESCA, HNSC, KIRP, LGG, LUSC, OV, PCPG (Pheochromocytoma and Paraganglioma), READ (rectum adenocarcinoma), STAD and UCEC (Figure 4). YTHDF1 expression in different immune and molecular subtypes of other cancers is shown in Supplementary Figures 3, 4. Based on the above results, we concluded that YTHDF1 expression differs in immune subtypes and molecular subtypes of various human cancer types.
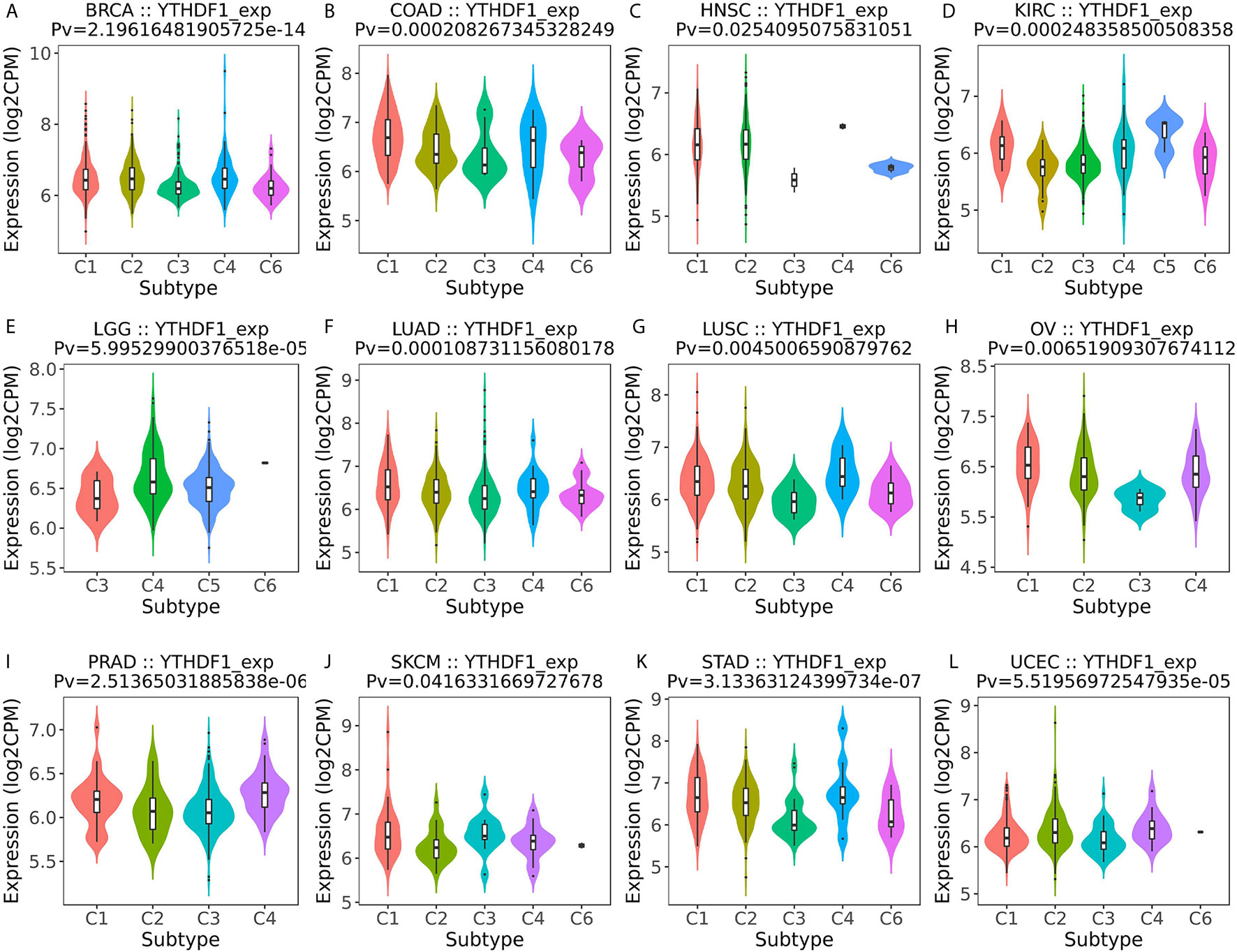
Figure 3 The relationship between YTHDF1 expression and pan-cancer immune subtypes. (A) in BRCA, (B) in COAD, (C) in HNSC, (D) in KIRC, (E) in LGG, (F) in LUAD, (G) in LUSC, (H) in OV, (I) in PRAD, (J) in SKCM, (K) in STAD, (L) in UCEC.
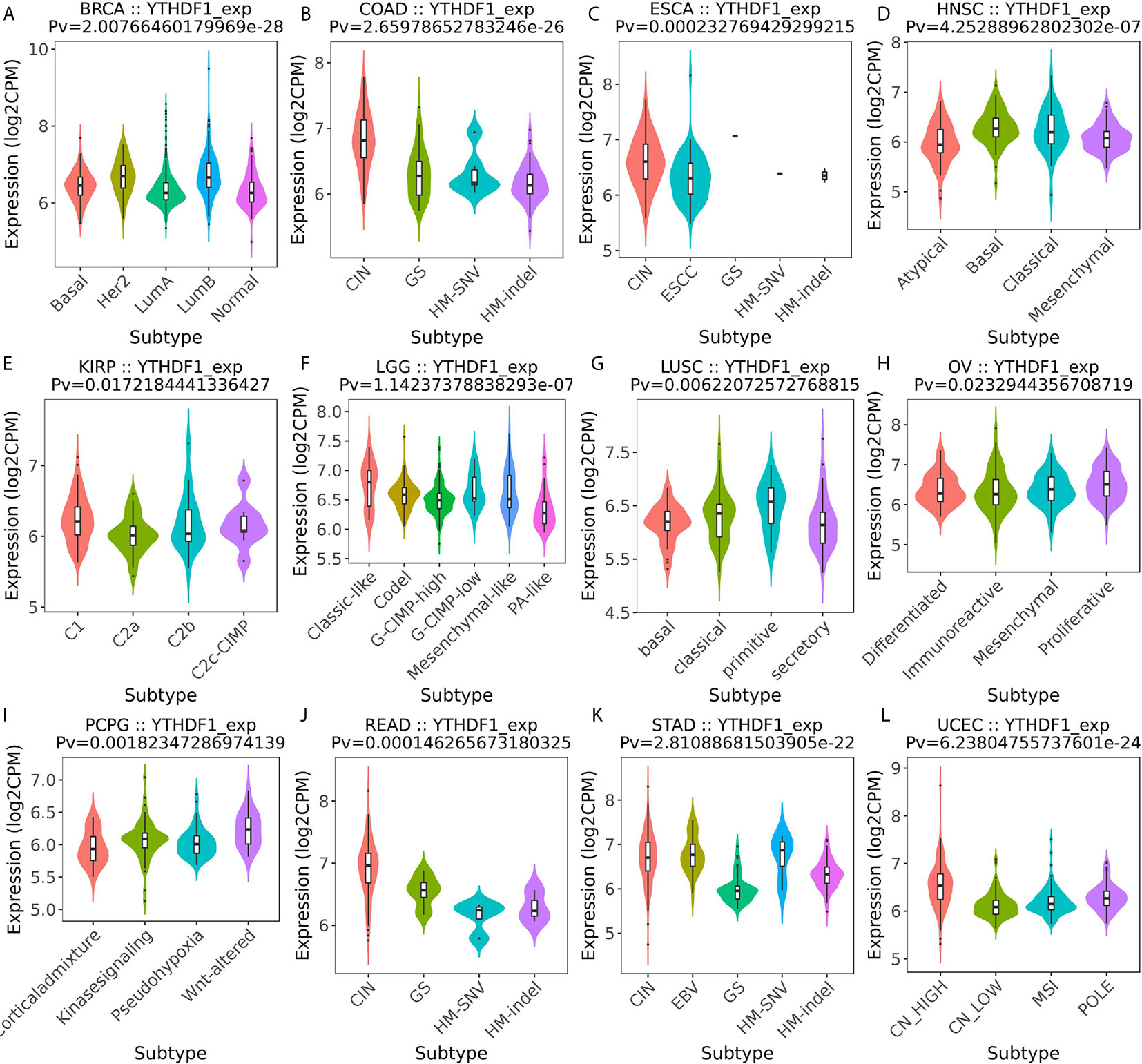
Figure 4 The relationship between YTHDF1 expression and pan-cancer molecular subtypes. (A) in BRCA, (B) in COAD, (C) in ESCA, (D) in HNSC, (E) in KIRP, (F) in LGG, (G) in LUSC, (H) in OV, (I) in PCPG, (J) in READ, (K) in STAD, (L) in UCEC.
Studies have proven that immune checkpoint (ICP) genes have a great influence on immune cell infiltration and immunotherapy (39). Subsequently, we explored the associations between YTHDF1 expression and ICP genes in human cancers to explore the potential of YTHDF1 in immunotherapy. Among 47 ICP genes, strong relationships with YTHDF1 expression were found in many types of cancer, such as GBM (glioblastoma multiforme), OV, LUSC, PRAD, UCEC, TGCT, KIRP, LIHC, BRCA, COAD, KIRC, THCA, LAML, LGG, KICH, and UVM (Figure 5). In OV, PRAD, UCEC, KIRP, LIHC, KIRC, LAML, LGG, KICH, and UVM, YTHDF1 expression was positively related to immune checkpoint genes, especially in LIHC, in which 32 of 47 immune checkpoint genes had connections with YTHDF1 expression. This suggests that YTHDF1 might coordinate the activity of these ICP genes in different signal transduction pathways, and potentially serve as an ideal immunotherapy target. And high YTHDF1 expression might predict good therapeutic efficacy of immunotherapy targeting ICP genes. In GBM, LUSC, TGCT, BRCA, COAD and THCA, YTHDF1 is negatively related with ICP genes, which means that high YTHDF1 expression might predict unsatisfying immunotherapy outcomes when targeting ICP genes. On the other hand, YTHDF1 inhibitors might be potential alternative treatments. Thus, we hypothesized that YTHDF1, working as a potential pan-cancer biomarker or as a new immunotherapy target, may predict the immunotherapy response or achieve a promising therapeutic outcome, respectively.
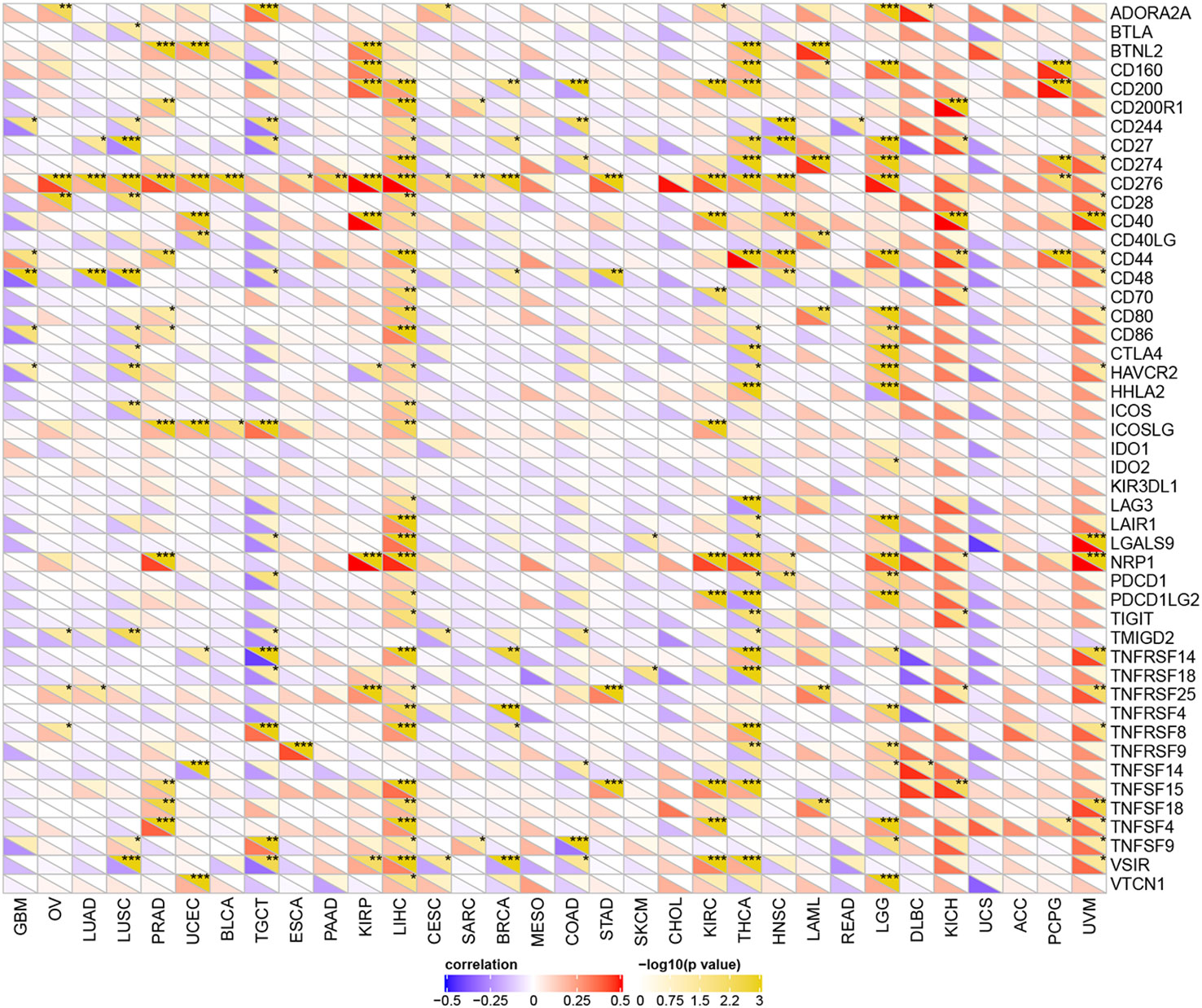
Figure 5 The relationship between YTHDF1 expression and pan-cancer immune checkpoint genes. *P < 0.05; **P < 0.01; ***P < 0.001.
To explore the role of YTHDF1 in the immune mechanism and immune response of the tumor microenvironment (TME), we analyzed the correlations between YTHDF1 expression and TMB, MSI, and neoantigens. TMB, MSI, and neoantigens in tumor microenvironment are related to antitumor immunity and could predict the therapeutic efficacy of tumor immunotherapy (13, 40–42). Our results showed that YTHDF1 expression had significant positive associations with TMB in LUAD, LUSC, BLCA, PAAD (pancreatic adenocarcinoma), SARC (sarcoma), BRCA, STAD, LGG, and ACC and negative relations in COAD and THCA (Figure 6A). For MSI, there were positive correlations with YTHDF1 expression in LUAD, LUSC, UCEC, BLCA, ESCA, and KIRC and negative correlations with in PRAD, COAD, and DLBC (lymphoid neoplasm diffuse large B-cell lymphoma) (Figure 6B). Analyzing the relationships between YTHDF1 expression and neoantigens, we found that YTHDF1 expression was positively related to neoantigens in LUAD, BRCA, HNSC, LIHC, and LGG and negatively related to neoantigens in COAD (Figure 6C). Then, we explored the associations between YTHDF1 expression and three kinds of ESTIMATE (Figure 6D). The results indicated that there were strong positive correlations with YTHDF1 expression in LGG and negative correlations in ACC, PCPG, GBM, OV, LUAD, LUSC, UCEC, BLCA, TGCT, PAAD, KIRP, CESC, BRCA, COAD, STAD, SKCM, KIRC, THCA, HNSC, READ, and UCS. The above results further verified our conjecture that YTHDF1 may influence antitumor immunity by regulating the composition and immune mechanism in the TME.
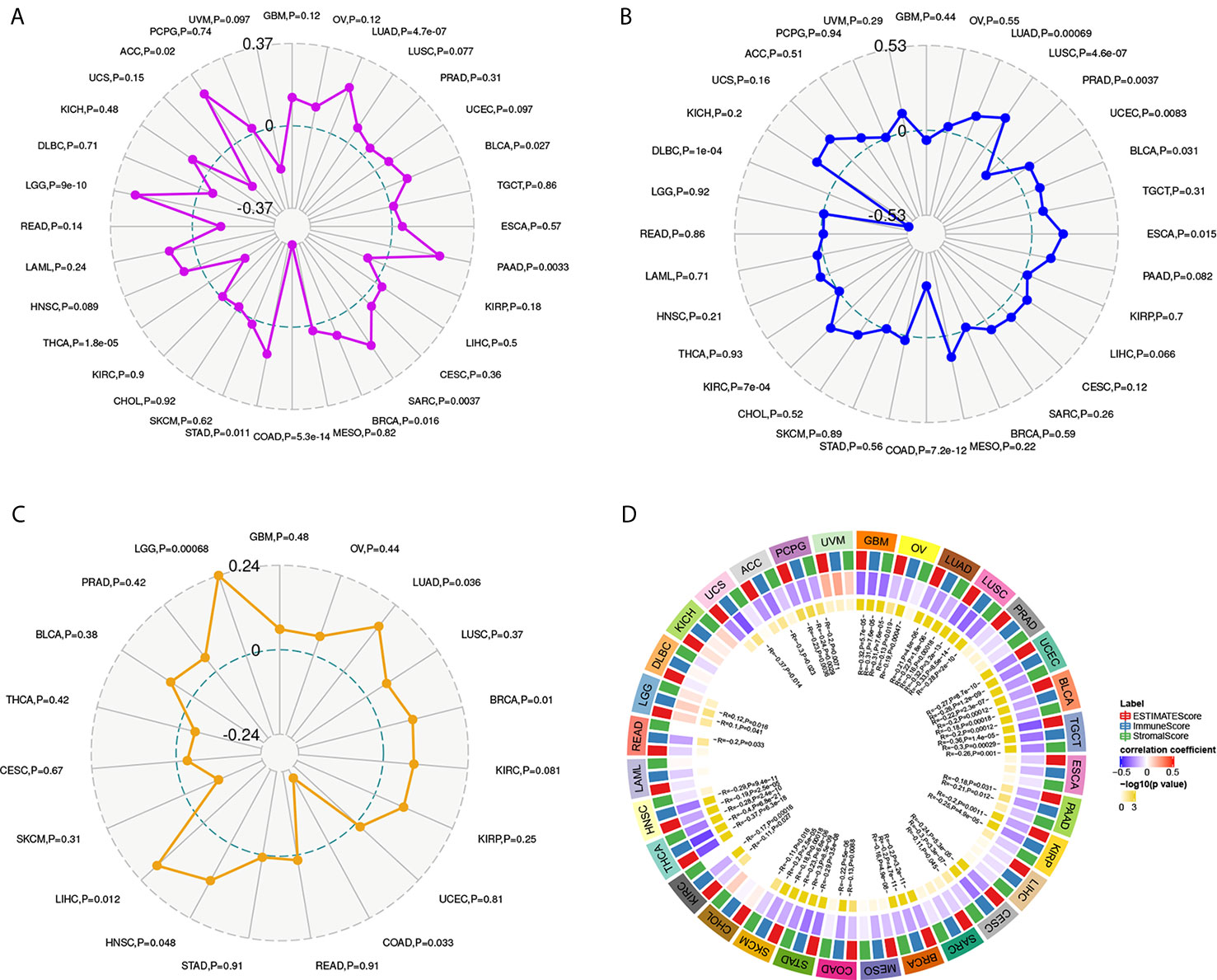
Figure 6 The relationship between YTHDF1 expression and TMB (A), MSI (B), neoantigen (C) and ESTIMATE score (D) in human cancers. TMB, tumor mutational burden; MSI, microsatellite instability; MMR genes, mismatch repair genes; ESTIMATE, Estimation of Stromal and Immune cells in Malignant Tumor tissues using Expression data.
After proving the differential YTHDF1 expression in different immune subtypes, we explored the potential correlation between YTHDF1 expression and immune cell infiltration in human cancers, and the results indicated that there was significant correlation in 31 cancer types. YTHDF1 expression had a strong relationship with dendritic cell in 12 cancer types, macrophage in 9 cancer types, neutrophil in 8 cancer types, CD8+ T cells in 13 cancer types, B cells in 10 cancer types and CD4+ T cells in 6 cancer types (Figure 7A). The TIMER database’s results consisted of the above results, and all details are shown on Supplementary Table 4.
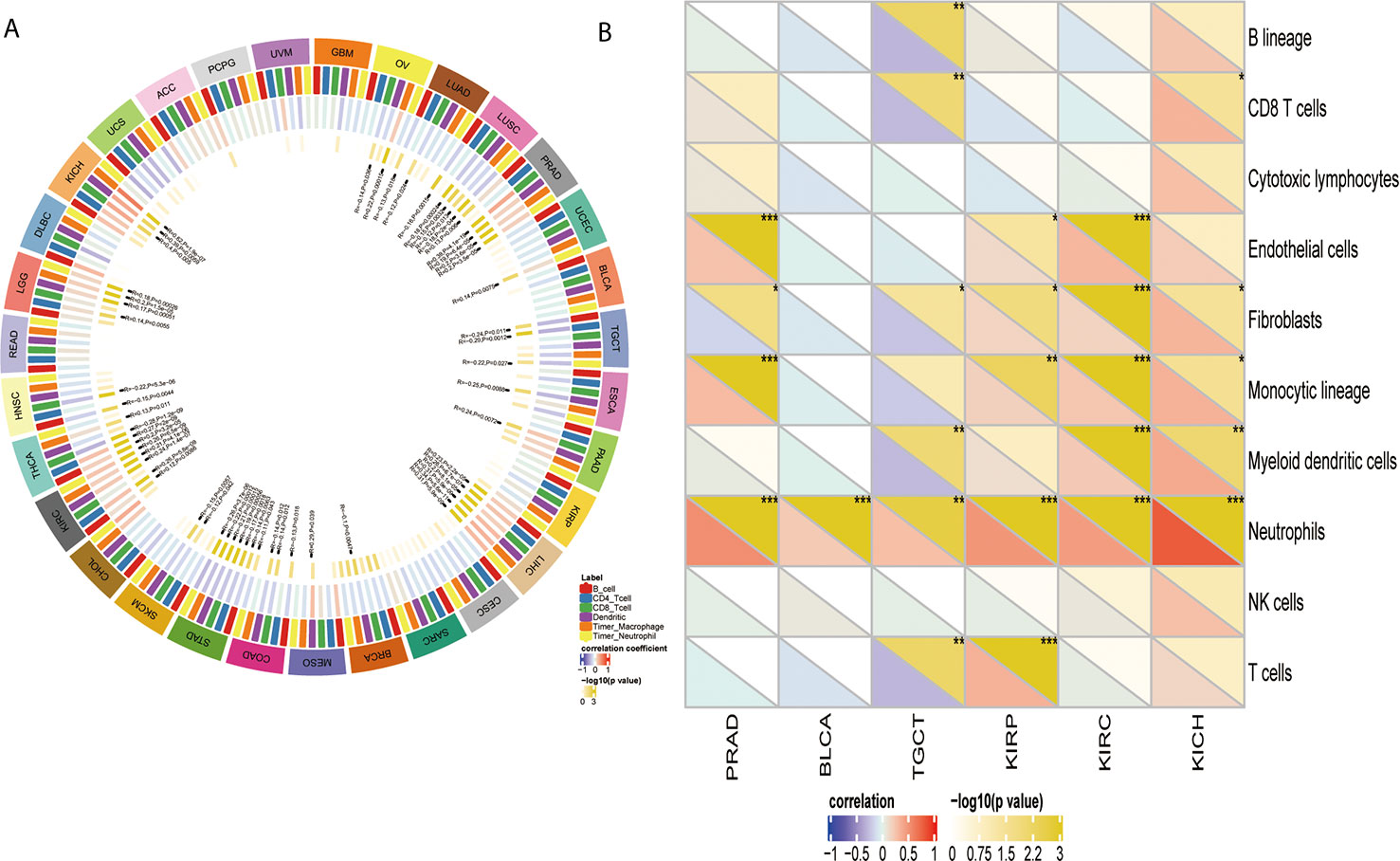
Figure 7 The relationship between YTHDF1 expression and infiltrating immune cells of human cancers and urogenital cancers. (A) the relationship between YTHDF1 expression level and infiltrating levels of B cells, CD4+ T cells, CB8+ T cells, macrophages, neutrophils, dendritic cell in human cancers. (B) the relationship between YTHDF1 expression level and infiltrating levels of B cell lineages, CB8+ T cells, cytotoxic lymphocytes, endothelial cells, fibroblasts, monocytic cell lineages, myeloid dendritic cells, neutrophils, natural killer cells, T cells in six urogenital cancers. *P < 0.05; **P < 0.01; ***P < 0.001.
Next, we thoroughly analyze the connection between YTHDF1 expression and immune cell infiltration in six urogenital cancers. The results showed that YTHDF1 expression was correlated with the infiltration levels of neutrophils in BLCA, CD8+ T cells, fibroblasts, monocytic cell lineage, myeloid dendritic cells, and neutrophils in KICH; endothelial cells, fibroblasts, monocytic cell lineage, and neutrophils in PRAD; B cell lineage, CB8+ T cells, fibroblasts, myeloid dendritic cells, neutrophils, and T cells in TGCT; endothelial cells, fibroblasts, monocytic cell lineage, neutrophils, and T cells in KIRP and endothelial cells, fibroblasts, monocytic cell lineage, myeloid dendritic cells and neutrophils in KIRC (Figure 7B). Next, we verified the above relationship via GEPIA and TIMER, and most results from those 2 databases were consistent with the above results. YTHDF1 expression was positively related to immune cells in all urogenital cancers except TGCT. Detailed information on the relationship between YTHDF1 expression and TILs in the GEPIA and TIMER databases is shown in Supplementary Figures 5–10. In general, the above results strongly identified that YTHDF1 expression influenced immune cell infiltration in urogenital cancers.
Then, we used the TIMER database to further explore the correlation between YTHDF1 expression and different marker subsets of immune cells in six urogenital cancers. The correlation was adjusted by purity to decrease the bias caused by clinical samples. The results showed that YTHDF1 was strongly connected with marker genes of macrophages, Th1 cells, Th2 cells, Tfh cells, Treg cells, and Th17 cells in urogenital cancers. For example, YTHDF1 expression had a strong association with STAT1 in Th1 cells, STAT6 in Th2 cells, STAT3 in Th17 cells, STAT5B in Treg cells, BCL6 in Tfh cells, and IRF5 in M1 macrophages in most cancer types (Supplementary Table 5). We used BLCA (n = 408) and PRAD (n = 497) as examples to demonstrate the potential immune function of YTHDF1 in urogenital cancers. According to the results of Table 1, YTHDF1 has a close connection with all included marker genes of monocytes, tumor-associated macrophages (TAMs), M2 Macrophages, Th1 cells and regulatory T cells (Tregs) in PRAD. However, YTHDF1 expression in BLCA does not have a strong link since it is only correlated with most marker genes of Th2 cells, and regulatory T cells (Tregs). Different relationships between YTHDF1 expression and gene markers of immune cell infiltration in PRAD and BLCA may explain why there are different survival outcomes in various cancer types. These results again indicate that YTHDF1 expression plays an essential role in the infiltration of immune cells in human cancers, and that YTHDF1 might regulate the function of immune cells by participating in a mechanism similar to that of marker genes.
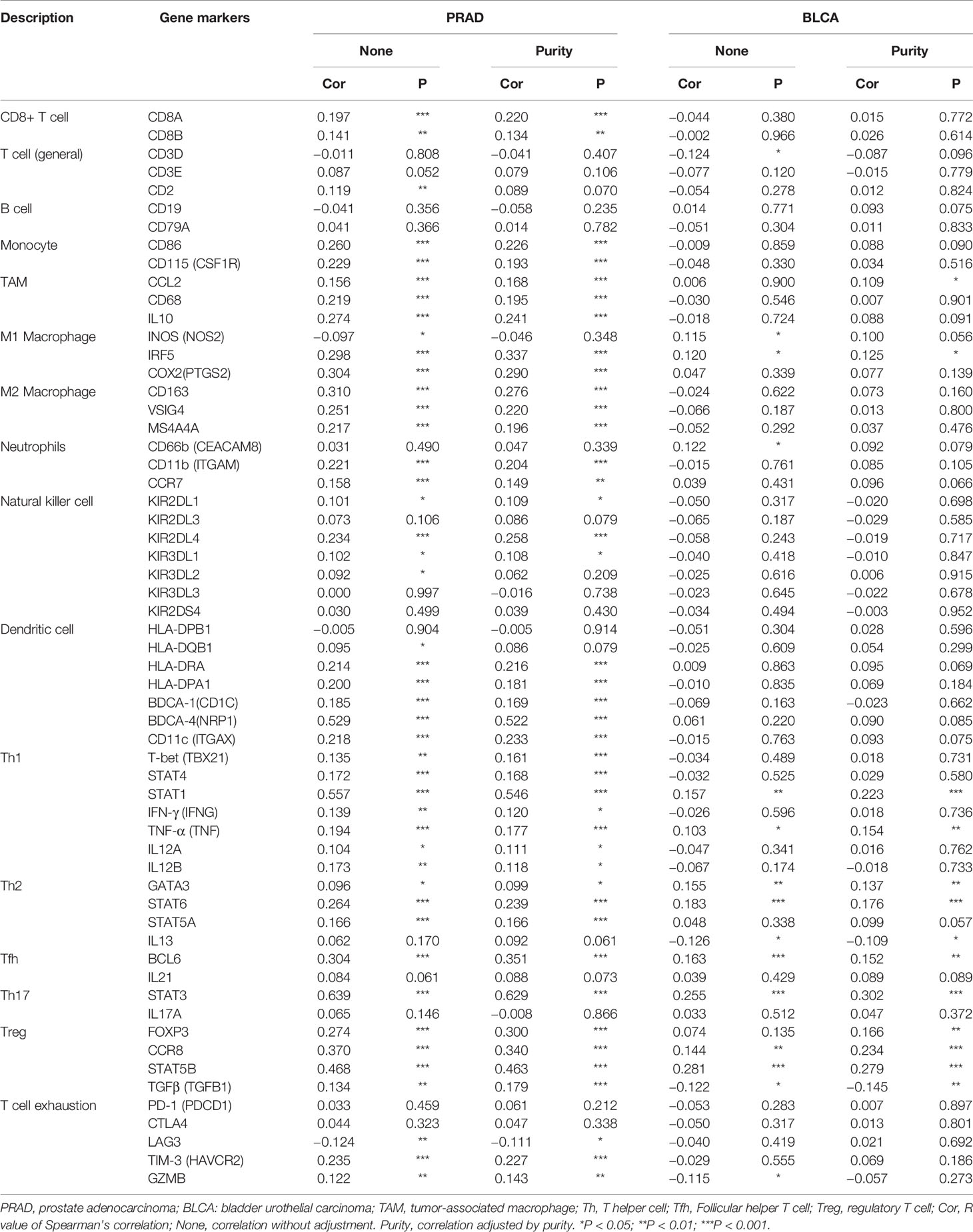
Table 1 Correlation between YTHDF1 and relate genes and markers of immune cells analyzed by TIMER.
Analysis of YTHDF1 genomic alterations in urogenital cancers was explored with the cBioPortal website. The results showed that genomic alterations of YTHDF1 occurred in 0.7% of patients (Figure 8A). The types of YTHDF1 gene alterations were diverse, resulting in changes in gene expression (Figure 8B). Copy number variation (CNV) mainly occurred in PRAD and BLCA, while no CNV occurred in KICH and TGCT (Figure 8C). Next, we used the UALCAN database to explore YTHDF1 expression in urogenital cancers with different clinical characteristics. Taking BLCA as an example, we found that YTHDF1 was significantly differentially expressed existed in different cancer stages, histological subtypes, patient sex, molecular subtypes, nodal metastasis status and TP53 mutation status of BLCA (Figures 8D–I). Additionally, differential YTHDF1 expression in another five urogenital cancers with different clinical characteristics is displayed in Supplementary Figure 11. All the above results suggested that YTHDF1 genomic alteration and differential expression indeed occur in cancer tissue, and might play an essential role in cancer onset and progression.
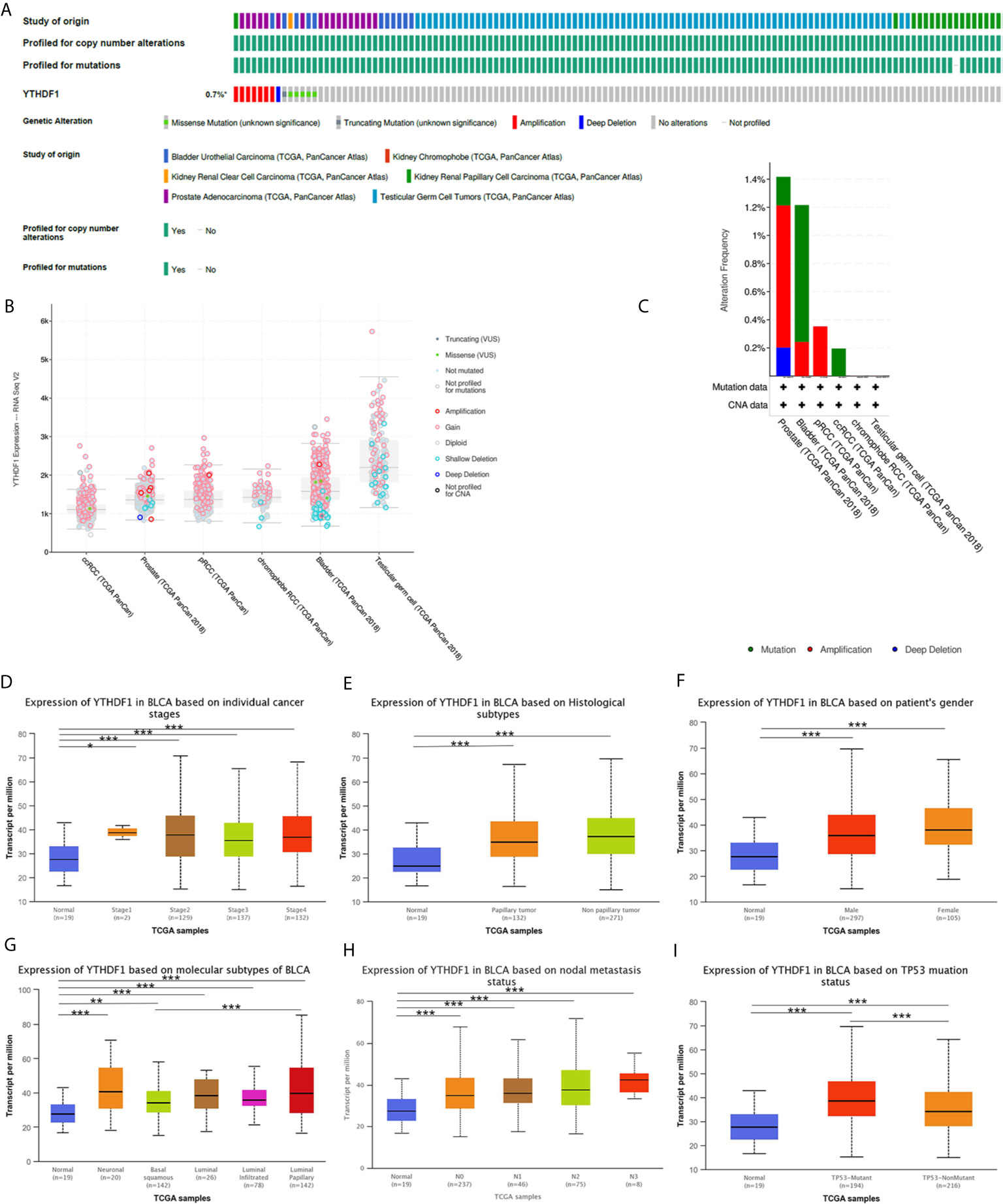
Figure 8 YTHDF1 genomic alterations in six urogenital cancers analyzed by the cBioPortal database (A–C) and YTHDF1 differential expression in bladder cancer with different clinical subgroups (D–I) analyzed by the UALCAN database. (A) OncoPrint of YTHDF1 gene alterations in cancer cohort. (Different colors means different types of genetic alterations and amplification accounts for the largest proportion). (B) main type of YTHDF1 gene alterations in cancer groups. (C) Details of YTHDF1 gene alteration types in cancer cohort; (D–I) YTHDF1 differential expression in bladder cancer with individual cancer stages (n = 419) (D), histological subtypes (n = 422) (E), patient sex (n = 421) (F), molecular subtypes (n = 427) (G), nodal metastasis status (n = 385) (H), TP53 mutation status (n = 429) (I). (*P < 0.05, **P < 0.01, ***P < 0.001).
The above results identified that YTHDF1 had a significant association with the prognosis and immunity of cancers. Next, we explored YTHDF1 coexpression networks using the LinkedOmics database to verify the potential function of YTHDF1 in tumor tissue, and used BLCA as an example to illustrate the potential effect. In BLCA, 1396 genes (dark red dots) were significantly positively related to YTHDF1, and 1143 genes (dark green dots) were negatively related (false discovery rate (FDR) < 0.01) (Figure 9A). A heat map was used to display the top 50 genes positively and negatively correlated with YTHDF1 (Figures 9B, C). Moreover, all coexpressed genes were displayed in Supplementary Table 6. C20orf11, CSTF1, and LSM14B (r = 0.631) had the strongest association with YTHDF1 expression (r = 0.696, 0.647, 0.631 and P = 1.90E-60, 9.72E-46, 6.43E-46, respectively). Next, gene set enrichment analysis (GSEA) was used to determine the main gene ontology (GO) terms of YTHDF1 coexpression genes. We further analyzed the biological process categories of GO and found that YTHDF1 and its co-expression genes primarily participated in the regulation of immune response, leukocyte activation and inflammatory response (Figure 9D). Then, we carried out Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway analysis, and the results showed that coexpressed genes were enriched in staphylococcus aureus infection, antigen processing and presentation, and cytokine-cytokine receptor interaction (Figure 9E). These results suggested that YTHDF1 expression might play an essential role in human cancers by regulating the immune response of the TME.
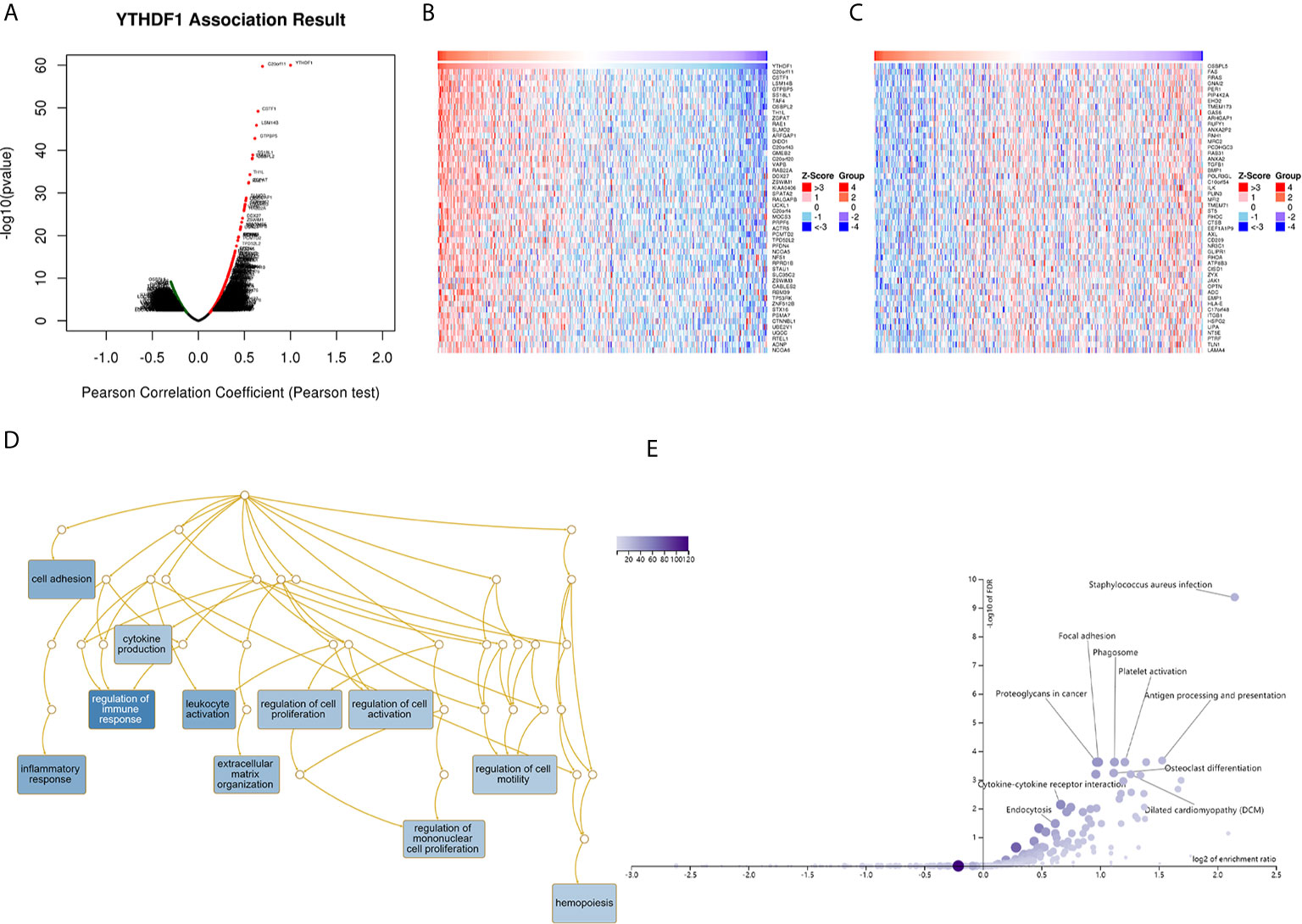
Figure 9 YTHDF1 coexpression genes in BLCA analyzed by the LinkedOmics database. (A) Highly correlated genes of YTHDF1 tested by Pearson test in BLCA cohort. (B, C) Top 50 positive coexpression genes (B) and negative coexpression genes (C) of YTHDF1 in heat map in BLCA; (D) Directed acyclic graph of YTHDF1 GO analysis (biological process) in BLCA cohort. (E) Volcano plot of YTHDF1 KEGG pathways in BLCA cohort.
YTHDF1, a “reader” of m6A, impairs the activation of DCs and CD8+ T cells by increasing the translation of lysosomal cathepsins and thus degrading tumor neoantigens (4, 17). Moreover, in mice with low YTHDF1 expression, there was a higher level of CD8+ T cells and NK cells in the TME, which means enhanced immunosurveillance in the absence of YTHDF1 (17). Moreover, YTHDF1 also plays an important role in oncogenesis and tumor progression in various cancers. For example, YTHDF1 promotes the growth and progression of bladder cancer via the ITGA6-METTL3 pathway (43). In colorectal cancer, YTHDF1 plays a vital oncogenic role in cell self-renewal and differentiation via the Wnt/β-catenin pathway (44). In addition, YTHDF1 also produces a marked effect on lung cancer growth and metastasis by mediating YAP expression and activity (14). Although anti-programmed death 1 (PD-1)/programmed death ligand-1(PD-L1) drugs have achieved considerable clinical efficacy and durable response with low toxicity (45, 46), they are not effective for all cancer types or do not achieve ideal efficacy in all patients (47). A new study identified that YTHDF1 deficiency in classical dendritic cells enhanced the cross-presentation of tumor antigens and the cross-priming of CD8+ T cells, and the therapeutic efficacy of the anti-PD-L1 drug was improved after blocking YTHDF1 (17). The YTHDF1 inhibitor GB1107 was also proven to inhibit the growth and metastasis of lung adenocarcinoma in vivo (48). Consistent with our findings and hypothesis, GB1107 increased tumor M1 macrophage polarization and CD8+ T cell infiltration and potentiated the effects of a PD-L1 immune checkpoint inhibitor (48). Therefore, YTHDF1 can be used as a new anticancer immunotherapy drug target or in combination with known immune checkpoint inhibitors to enhance immune infiltration and responses in cancers. These studies indicated that YTHDF1 might be a promising target in antitumor immunotherapy. Furthermore, combining anti-PD-L1 drugs with YTHDF1 depletion could be used as a new antitumor strategy.
In the first step of our study, we used the Oncomine, TIMER, GEPIA, and BioGPS databases to determine the expression level of YTHDF1 in cancers and normal tissues. The results showed that YTHDF1 had significantly higher expression in most cancer types except lymphoma and THCA, which was consistent with previous studies in lung and colorectal cancer (14, 44). These results indicated that YTHDF1 indeed promotes oncogenesis and tumor progression in human cancers.
Next, the relationship between YTHDF1 expression and prognosis was explored. Similar to previous studies, high YTHDF1 expression patients had a worse prognosis in LIHC and TCGT (49, 50). In addition, in many other cancer types such as ACC, BLCA, MESO, UVM, LIHC, and UCS, high YTHDF1 expression meant worse prognosis, which proved that YTHDF1 was a potential prognostic pan-cancer biomarker. Then, we explored YTHDF1 expression in different immune subtypes and molecular subtypes of human cancers to determine its potential mechanism of action. The results showed that YTHDF1 expression was significantly different in different immune subtypes and molecular subtypes in most cancer types, which might prove that YTHDF1 is a promising diagnostic pan-cancer biomarker and participates in immune regulation. Moreover, we demonstrated that there were significant differences in YTHDF1 expression in different clinical subgroups of urogenital cancers, consistent with a previous study showing that YTHDF1 is expressed differently in KIRC at different stages (50). In fact, YTHDF1 differential YTHDF1 expression existed in all urogenital cancers with diverse clinical characteristics, which indicated that YTHDF1 might play a role in the growth and progression of cancers.
Tumor-infiltrating lymphocytes (TILs) in the TME have been proven to be an independent predictor of cancer patient prognosis and immunotherapeutic efficacy (51, 52). Our study demonstrated that YTHDF1 had a strong association with TILs and played an essential role in the TME. For example, YTHDF1 expression was strongly significantly related to CD8+T cells, macrophages and dendritic cells. As a universal component of the TME, macrophages have been proven to contribute to immune evasion and suppression (53). The TME with low YTHDF1 expression contained higher levels of CD8+ T cells and natural killer (NK) cells in a mouse model, and enhanced the ability to DCs in cross-present of tumor antigens (17). In particular, in PRAD, we found that TYHDF1 expression has a close association with TILs in the TME. For example, YTHDF1 expression has a close association with IFN-gamma in PRAD, which is regarded as a hallmark of antitumor immunity (54). Moreover, YTHDF1 expression is related to all of the examined marker genes of CD8+ T cells, monocytes, tumor-associated macrophages (TAMs), M2 macrophages, Th1 cells, and other known immune stimulatory and immune suppressive cytokines, which proves the potential immune function of YTHDF1 in PRAD.
Our study also found that YTHDF1 was negatively related to the immune, stromal and ESTIMATE scores of the TME in most human cancer types, but YTHDF1 expression was not always negatively related to TILs in human cancers, which indicated that YTHDF1 plays a different immune regulation role in various cancer types. The correlation between YTHDF1 and tumor mutational burden (TMB), microsatellite instability (MSI), and neoantigens also proved that YTHDF1 closely related with the TME in human cancers. However, further experimental research to prove its function.
In our analysis of YTHDF1 coexpression networks, we found that YTHDF1 and its coexpression genes indeed took part in regulating the immune response and antigen processing and presentation. Furthermore, a previous study proved that anti-PD-L1 drugs achieved better therapeutic efficacy in YTHDF1 low expression conditions (17). A strong relationship between YTHDF1 and ICP genes was found, which provides a theoretical basis for combined molecular targeting immunotherapy for the future. In summary, the above results strongly indicated the potential of YTHDF1 as a target in anticancer immunotherapy.
However, even though we performed a comprehensive and systematic analysis on YTHDF1 and used different databases and R 3.2.2 for cross-verification, there are some limitations to this study. First, the microarray and sequencing data from different databases exhibited differences and lacked granularity and specificity, which might cause systematic bias. Second, in vivo/in vitro experiments are needed to prove our results on the potential function of YTHDF1, which could increase the credibility of our results. Third, even though we concluded that YTHDF1 expression was strongly related to immune cell infiltration and prognosis of human cancers, we lack direct evidence on YTHDF1 influencing prognosis by taking part in immune infiltration, Therefore, the mechanisms by which YTHDF1 participate in immune regulation remain unknown, and the exact pathway needs further study. Fourth, no anti-YTHDF1 therapeutic monoclonal antibodies have been evaluated in clinical trials. Therefore, we have no specific and complete cases with data to identify the benefit of anti-YTHDF1 targeting drugs in the survival of cancer models or inhibiting tumor growth. In the future, a prospective study of YTHDF1 expression and its role in human cancers’ immune infiltration is needed, along with successful development and testing of a new antitumor immunotherapy drug targeting YTHDF1.
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found in the article/Supplementary Material.
JianH, DQ, AY and JiaoH were responsible for experimental design, experimental analysis and thesis writing. HD, HL, and ZY were responsible for data screening, collection and writing. JC and XZ were responsible for the guidance and review of the thesis. All authors contributed to the article and approved the submitted version.
We thank the National Natural Science Foundation of China (81902592), Hunan Province Key R&D Program (2019SK2202) and Xiangya Hospital Youth Fund (2018Q09) for the financial support. This article has not received sponsorship for publication.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
We thank everyone who provided support for this study. Thank You-e He and Belaydi Othmane for the helpful suggestions and language polishing on this manuscript.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fonc.2021.607224/full#supplementary-material
Supplementary Figure 1 | YTHDF1 expression in different cancer cell lines (A) and normal tissue (B) investigated by the BioGPS database.
Supplementary Figure 2 | Kaplan-Meier survival curve of human cancers with high and low YTHDF1 expression analyzed by the Kaplan-Meier plotter database. (A–G) Overall survival of ESCA, HNSC, PPAD, PCPG, READ, THCA, and UCEC with high and low YTHDF1 expression (n = 80, 499, 177, 178, 165, and 502 respectively); (H–N) Progression-free survival of ESSC, HNSC, LUAD, PAAD, READ, STAD, and UCEC with high and low YTHDF1 expression (n = 54, 124, 300, 69, 47, 215, and 422, respectively).
Supplementary Figure 3 | Relationship between YTHDF1 expression and pan-cancer immune subtypes.
Supplementary Figure 4 | Relationship between YTHDF1 expression and pan-cancer molecular subtypes.
Supplementary Figure 5 | Correlation between YTHDF1 expression and 24 tumor infiltrating lymphocytes (TILs) in KICH analyzed by the GEPIA database.
Supplementary Figure 6 | Correlation between YTHDF1 expression and 24 tumor infiltrating lymphocytes (TILs) in PRAD analyzed by the GEPIA database.
Supplementary Figure 7 | Correlation between YTHDF1 expression and 24 tumor infiltrating lymphocytes (TILs) in KIRC analyzed by the GEPIA database.
Supplementary Figure 8 | Correlation between YTHDF1 expression and 24 tumor infiltrating lymphocytes (TILs) in KIRP analyzed by the GEPIA database.
Supplementary Figure 9 | Correlation between YTHDF1 expression and 24 tumor infiltrating lymphocytes (TILs) in BLCA analyzed by the GEPIA database.
Supplementary Figure 10 | Correlation between YTHDF1 expression and 24 tumor infiltrating lymphocytes (TILs) in TGCT analyzed by the GEPIA database.
Supplementary Figure 11 | YTHDF1 differential expression in five urogenital cancers with different clinical subgroups analyzed by the UALCAN database.
Supplementary Table 1 | Sample Numbers of the TCGA Cancer types and corresponding immune and molecular subtypes.
Supplementary Table 2 | Correlation between YTHDF1 expression and survival in the GEO datasets analyzed by the PrognoScan database.
Supplementary Table 3 | Detailed relationship between YTHDF1 expression and pan-cancer immune cell infiltration in the tumor microenvironment.
Supplementary Table 4 | Relationship between YTHDF1 expression and related immune marker sets of urogenital cancers analyzed by the TIMER database.
Supplementary Table 5 | 24 tumors associated infiltrating lymphocytes (TILs) and their gene markers.
Supplementary Table 6 | YTHDF1 coexpression genes in urogenital cancers analyzed by the LinkedOmics database.
YTHDF1, YTH N6-methyladenosine RNA binding protein 1; ICP, immune checkpoints; TMB, tumor mutational burden; MSI: microsatellite instability; TILs, tumor immune-infiltrated cells; m6A, N6-methyladenine; TCGA, the Cancer Genome Atlas; ESTIMATE, Estimation of Stromal and Immune cells in Malignant Tumor tissues using Expression data; OS, overall survival; DFS, disease-free survival; RFS, relapse-free survival; ACC, adrenocortical carcinoma; BLCA, bladder urothelial carcinoma; BRCA, breast invasive carcinoma; CESC, cervical squamous cell carcinoma and endocervical adenocarcinoma; COAD, colon adenocarcinoma; READ, colon adenocarcinoma; ESCA, esophageal carcinoma; GBM, glioblastoma multiforme; HNSC, head and neck squamous cell carcinoma; KICH, kidney chromophobe; KIRC, kidney renal clear cell carcinoma; KIRP, kidney renal papillary cell carcinoma; LAML, acute myeloid leukemia; LGG, brain lower grade glioma; LIHC, liver hepatocellular carcinoma; LUAD, lung adenocarcinoma; LUSC, lung squamous cell carcinoma; MESO, mesothelioma; OV, ovarian serous cystadenocarcinoma; PAAD, pancreatic adenocarcinoma; PCPG, pheochromocytoma and paraganglioma; PRAD, prostate adenocarcinoma; READ, rectum adenocarcinoma; STAD, stomach adenocarcinoma; TGCT, testicular germ cell tumors; THCA, thyroid carcinoma; UCEC, uterine corpus endometrial carcinoma; UCS, uterine carcinosarcoma; TME, tumor microenvironment.
1. He L, Li H, Wu A, Peng Y, Shu G, Yin G. Functions of N6-methyladenosine and its Role in Cancer. Mol Cancer (2019) 18(1):176. doi: 10.1186/s12943-019-1109-9
2. Dominissini D, Moshitch-Moshkovitz S, Schwartz S, Salmon-Divon M, Ungar L, Osenberg S, et al. Topology of the Human and Mouse M6a RNA Methylomes Revealed by M6a-Seq. Nature (2012) 485(7397):201–6. doi: 10.1038/nature11112
3. Jia G, Fu Y, Zhao X, Dai Q, Zheng G, Yang Y, et al. N6-Methyladenosine in Nuclear RNA is a Major Substrate of the Obesity-Associated FTO. Nat Chem Biol (2011) 7(12):885–7. doi: 10.1038/nchembio.687
4. Wang X, Zhao BS, Roundtree IA, Lu Z, Han D, Ma H, et al. N(6)-Methyladenosine Modulates Messenger RNA Translation Efficiency. Cell (2015) 161(6):1388–99. doi: 10.1016/j.cell.2015.05.014
5. Wang X, Lu Z, Gomez A, Hon GC, Yue Y, Han D, et al. N6-Methyladenosine-Dependent Regulation of Messenger RNA Stability. Nature (2014) 505(7481):117–20. doi: 10.1038/nature12730
6. Dang W, Xie Y, Cao P, Xin S, Wang J, Li S, et al. N(6)-Methyladenosine and Viral Infection. Front Microbiol (2019) 10:417. doi: 10.3389/fmicb.2019.00417
7. Roignant JY, Soller M. M(6)a in Mrna: An Ancient Mechanism for Fine-Tuning Gene Expression. Trends Genet TIG (2017) 33(6):380–90. doi: 10.1016/j.tig.2017.04.003
8. Liu J, Eckert MA, Harada BT, Liu SM, Lu Z, Yu K, et al. M(6)a mRNA Methylation Regulates AKT Activity to Promote the Proliferation and Tumorigenicity of Endometrial Cancer. Nat Cell Biol (2018) 20(9):1074–83. doi: 10.1038/s41556-018-0174-4
9. Vu LP, Pickering BF, Cheng Y, Zaccara S, Nguyen D, Minuesa G, et al. The N(6)-methyladenosine (M(6)a)-Forming Enzyme METTL3 Controls Myeloid Differentiation of Normal Hematopoietic and Leukemia Cells. Nat Med (2017) 23(11):1369–76. doi: 10.1038/nm.4416
10. Li Z, Weng H, Su R, Weng X, Zuo Z, Li C, et al. Fto Plays an Oncogenic Role in Acute Myeloid Leukemia as a N(6)-Methyladenosine Rna Demethylase. Cancer Cell (2017) 31(1):127–41. doi: 10.1016/j.ccell.2016.11.017
11. Liu ZX, Li LM, Sun HL, Liu SM. Link Between M6a Modification and Cancers. Front Bioeng Biotechnol (2018) 6:89. doi: 10.3389/fbioe.2018.00089
12. Ott PA, Hu Z, Keskin DB, Shukla SA, Sun J, Bozym DJ, et al. An Immunogenic Personal Neoantigen Vaccine for Patients With Melanoma. Nature (2017) 547(7662):217–21. doi: 10.1038/nature22991
13. Schumacher TN, Schreiber RD. Neoantigens in Cancer Immunotherapy. Sci (New York NY) (2015) 348(6230):69–74. doi: 10.1126/science.aaa4971
14. Jin D, Guo J, Wu Y, Yang L, Wang X, Du J, et al. M(6)a Demethylase ALKBH5 Inhibits Tumor Growth and Metastasis by Reducing YTHDFs-mediated YAP Expression and Inhibiting Mir-107/LATS2-Mediated YAP Activity in NSCLC. Mol Cancer (2020) 19(1):40. doi: 10.1186/s12943-020-01161-1
15. Nishizawa Y, Konno M, Asai A, Koseki J, Kawamoto K, Miyoshi N, et al. Oncogene c-Myc Promotes Epitranscriptome M(6)a Reader YTHDF1 Expression in Colorectal Cancer. Oncotarget (2018) 9(7):7476–86. doi: 10.18632/oncotarget.23554
16. Liu L, Liu X, Dong Z, Li J, Yu Y, Chen X, et al. N6-Methyladenosine-Related Genomic Targets are Altered in Breast Cancer Tissue and Associated With Poor Survival. J Cancer (2019) 10(22):5447–59. doi: 10.7150/jca.35053
17. Han D, Liu J, Chen C, Dong L, Liu Y, Chang R, et al. Anti-Tumour Immunity Controlled Through mRNA M(6)a Methylation and YTHDF1 in Dendritic Cells. Nature (2019) 566(7743):270–4. doi: 10.1038/s41586-019-0916-x
18. Rhodes DR, Kalyana-Sundaram S, Mahavisno V, Varambally R, Yu J, Briggs BB, et al. Oncomine 3.0: Genes, Pathways, and Networks in a Collection of 18,000 Cancer Gene Expression Profiles. Neoplasia (New York NY) (2007) 9(2):166–80. doi: 10.1593/neo.07112
19. Li T, Fan J, Wang B, Traugh N, Chen Q, Liu JS, et al. Timer: A Web Server for Comprehensive Analysis of Tumor-Infiltrating Immune Cells. Cancer Res (2017) 77(21):e108–e10. doi: 10.1158/0008-5472.Can-17-0307
20. Tang Z, Kang B, Li C, Chen T, Zhang Z. GEPIA2: An Enhanced Web Server for Large-Scale Expression Profiling and Interactive Analysis. Nucleic Acids Res (2019) 47(W1):W556–w60. doi: 10.1093/nar/gkz430
21. Wu C, Jin X, Tsueng G, Afrasiabi C, Su AI. BioGPS: Building Your Own Mash-Up of Gene Annotations and Expression Profiles. Nucleic Acids Res (2016) 44(D1):D313–6. doi: 10.1093/nar/gkv1104
22. Mizuno H, Kitada K, Nakai K, Sarai A. PrognoScan: A New Database for Meta-Analysis of the Prognostic Value of Genes. BMC Med Genomics (2009) 2:18. doi: 10.1186/1755-8794-2-18
23. Nagy A, Lanczky A, Menyhart O, Gyorffy B. Validation of miRNA Prognostic Power in Hepatocellular Carcinoma Using Expression Data of Independent Datasets. Sci Rep (2018) 8(1):9227. doi: 10.1038/s41598-018-27521-y
24. Ru B, Wong CN, Tong Y, Zhong JY, Zhong SSW, Wu WC, et al. TISIDB: An Integrated Repository Portal for Tumor-Immune System Interactions. Bioinf (Oxford England) (2019) 35(20):4200–2. doi: 10.1093/bioinformatics/btz210
25. Wang D, Wang Y, Zou X, Shi Y, Liu Q, Huyan T, et al. FOXO1 Inhibition Prevents Renal Ischemia-Reperfusion Injury Via cAMP-response Element Binding Protein/PPAR-γ Coactivator-1α-Mediated Mitochondrial Biogenesis. Br J Pharmacol (2020) 177(2):432–48. doi: 10.1111/bph.14878
26. Danilova L, Ho WJ, Zhu Q, Vithayathil T, De Jesus-Acosta A, Azad NS, et al. Programmed Cell Death Ligand-1 (Pd-L1) and CD8 Expression Profiling Identify an Immunologic Subtype of Pancreatic Ductal Adenocarcinomas With Favorable Survival. Cancer Immunol Res (2019) 7(6):886–95. doi: 10.1158/2326-6066.Cir-18-0822
27. Bonneville R, Krook MA, Kautto EA, Miya J, Wing MR, Chen H-Z, et al. Landscape of Microsatellite Instability Across 39 Cancer Types. JCO Precis Oncol (2017) 1:PO.17.00073. doi: 10.1200/PO.17.00073
28. Chan TA, Yarchoan M, Jaffee E, Swanton C, Quezada SA, Stenzinger A, et al. Development of Tumor Mutation Burden as an Immunotherapy Biomarker: Utility for the Oncology Clinic. Ann Oncol Off J Eur Soc Med Oncol (2019) 30(1):44–56. doi: 10.1093/annonc/mdy495
29. Rooney MS, Shukla SA, Wu CJ, Getz G, Hacohen N. Molecular and Genetic Properties of Tumors Associated With Local Immune Cytolytic Activity. Cell (2015) 160(1-2):48–61. doi: 10.1016/j.cell.2014.12.033
30. Yoshihara K, Shahmoradgoli M, Martínez E, Vegesna R, Kim H, Torres-Garcia W, et al. Inferring Tumour Purity and Stromal and Immune Cell Admixture From Expression Data. Nat Commun (2013) 4:2612. doi: 10.1038/ncomms3612
31. Li B, Severson E, Pignon JC, Zhao H, Li T, Novak J, et al. Comprehensive Analyses of Tumor Immunity: Implications for Cancer Immunotherapy. Genome Biol (2016) 17(1):174. doi: 10.1186/s13059-016-1028-7
32. Miao YR, Zhang Q, Lei Q, Luo M, Xie GY, Wang H, et al. Immucellai: A Unique Method for Comprehensive T-Cell Subsets Abundance Prediction and its Application in Cancer Immunotherapy. Advanced Sci (Weinheim Baden Wurttemberg Germany) (2020) 7(7):1902880. doi: 10.1002/advs.201902880
33. Sousa S, Maatta J. The Role of Tumour-Associated Macrophages in Bone Metastasis. J Bone Oncol (2016) 5(3):135–8. doi: 10.1016/j.jbo.2016.03.004
34. Danaher P, Warren S, Dennis L, D’Amico L, White A, Disis ML, et al. Gene Expression Markers of Tumor Infiltrating Leukocytes. J Immunother Cancer (2017) 5:18. doi: 10.1186/s40425-017-0215-8
35. Siemers NO, Holloway JL, Chang H, Chasalow SD, Ross-MacDonald PB, Voliva CF, et al. Genome-Wide Association Analysis Identifies Genetic Correlates of Immune Infiltrates in Solid Tumors. PloS One (2017) 12(7):e0179726. doi: 10.1371/journal.pone.0179726
36. Gao J, Aksoy BA, Dogrusoz U, Dresdner G, Gross B, Sumer SO, et al. Integrative Analysis of Complex Cancer Genomics and Clinical Profiles Using the Cbioportal. Sci Signaling (2013) 6(269):pl1. doi: 10.1126/scisignal.2004088
37. Chandrashekar DS, Bashel B, Balasubramanya SAH, Creighton CJ, Ponce-Rodriguez I, Chakravarthi B, et al. Ualcan: A Portal for Facilitating Tumor Subgroup Gene Expression and Survival Analyses. Neoplasia (New York NY) (2017) 19(8):649–58. doi: 10.1016/j.neo.2017.05.002
38. Zhou G, Soufan O, Ewald J, Hancock REW, Basu N, Xia J. NetworkAnalyst 3.0: A Visual Analytics Platform for Comprehensive Gene Expression Profiling and Meta-Analysis. Nucleic Acids Res (2019) 47(W1):W234–41. doi: 10.1093/nar/gkz240
39. Topalian SL, Drake CG, Pardoll DM. Immune Checkpoint Blockade: A Common Denominator Approach to Cancer Therapy. Cancer Cell (2015) 27(4):450–61. doi: 10.1016/j.ccell.2015.03.001
40. Lee V, Murphy A, Le DT, Diaz LA Jr. Mismatch Repair Deficiency and Response to Immune Checkpoint Blockade. Oncologist (2016) 21(10):1200–11. doi: 10.1634/theoncologist.2016-0046
41. Peng M, Mo Y, Wang Y, Wu P, Zhang Y, Xiong F, et al. Neoantigen Vaccine: An Emerging Tumor Immunotherapy. Mol Cancer (2019) 18(1):128. doi: 10.1186/s12943-019-1055-6
42. Yarchoan M, Hopkins A, Jaffee EM. Tumor Mutational Burden and Response Rate to PD-1 Inhibition. New Engl J Med (2017) 377(25):2500–1. doi: 10.1056/NEJMc1713444
43. Jin H, Ying X, Que B, Wang X, Chao Y, Zhang H, et al. N(6)-Methyladenosine Modification of ITGA6 mRNA Promotes the Development and Progression of Bladder Cancer. EBioMedicine (2019) 47:195–207. doi: 10.1016/j.ebiom.2019.07.068
44. Bai Y, Yang C, Wu R, Huang L, Song S, Li W, et al. Ythdf1 Regulates Tumorigenicity and Cancer Stem Cell-Like Activity in Human Colorectal Carcinoma. Front Oncol (2019) 9:332. doi: 10.3389/fonc.2019.00332
45. Ma LJ, Feng FL, Dong LQ, Zhang Z, Duan M, Liu LZ, et al. Clinical Significance of PD-1/PD-Ls Gene Amplification and Overexpression in Patients With Hepatocellular Carcinoma. Theranostics (2018) 8(20):5690–702. doi: 10.7150/thno.28742
46. Meng X, Huang Z, Teng F, Xing L, Yu J. Predictive Biomarkers in PD-1/PD-L1 Checkpoint Blockade Immunotherapy. Cancer Treat Rev (2015) 41(10):868–76. doi: 10.1016/j.ctrv.2015.11.001
47. Topalian SL, Hodi FS, Brahmer JR, Gettinger SN, Smith DC, McDermott DF, et al. Safety, Activity, and Immune Correlates of anti-PD-1 Antibody in Cancer. New Engl J Med (2012) 366(26):2443–54. doi: 10.1056/NEJMoa1200690
48. Vuong L, Kouverianou E, Rooney CM, McHugh BJ, Howie SEM, Gregory CD, et al. An Orally Active Galectin-3 Antagonist Inhibits Lung Adenocarcinoma Growth and Augments Response to PD-L1 Blockade. Cancer Res (2019) 79(7):1480–92. doi: 10.1158/0008-5472.Can-18-2244
49. Zhao X, Chen Y, Mao Q, Jiang X, Jiang W, Chen J, et al. Overexpression of YTHDF1 is Associated With Poor Prognosis in Patients With Hepatocellular Carcinoma. Cancer Biomarkers Section A Dis Markers (2018) 21(4):859–68. doi: 10.3233/cbm-170791
50. Lobo J, Barros-Silva D, Henrique R, Jerónimo C. The Emerging Role of Epitranscriptomics in Cancer: Focus on Urological Tumors. Genes (2018) 9(11):552–73. doi: 10.3390/genes9110552
51. Azimi F, Scolyer RA, Rumcheva P, Moncrieff M, Murali R, McCarthy SW, et al. Tumor-Infiltrating Lymphocyte Grade is an Independent Predictor of Sentinel Lymph Node Status and Survival in Patients With Cutaneous Melanoma. J Clin Oncol Off J Am Soc Clin Oncol (2012) 30(21):2678–83. doi: 10.1200/jco.2011.37.8539
52. Ohtani H. Focus on TILs: Prognostic Significance of Tumor Infiltrating Lymphocytes in Human Colorectal Cancer. Cancer Immun (2007) 7:4.
53. Gajewski TF, Schreiber H, Fu YX. Innate and Adaptive Immune Cells in the Tumor Microenvironment. Nat Immunol (2013) 14(10):1014–22. doi: 10.1038/ni.2703
Keywords: YTHDF1, prognosis, tumor microenvironment, immune infiltration, immunotherapy, human cancer
Citation: Hu J, Qiu D, Yu A, Hu J, Deng H, Li H, Yi Z, Chen J and Zu X (2021) YTHDF1 Is a Potential Pan-Cancer Biomarker for Prognosis and Immunotherapy. Front. Oncol. 11:607224. doi: 10.3389/fonc.2021.607224
Received: 16 September 2020; Accepted: 06 April 2021;
Published: 06 May 2021.
Edited by:
Marco Rossi, University of Catanzaro, ItalyReviewed by:
Shanmugarajan Krishnan, Massachusetts General Hospital, United StatesCopyright © 2021 Hu, Qiu, Yu, Hu, Deng, Li, Yi, Chen and Zu. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Xiongbing Zu, enV4Ynh5eXlAMTI2LmNvbQ==; Jinbo Chen, Y2hlbmppbmJvMTk4OUB5YWhvby5jb20=
†These authors have contributed equally to this work
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.