
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Oncol. , 26 January 2021
Sec. Neuro-Oncology and Neurosurgical Oncology
Volume 10 - 2020 | https://doi.org/10.3389/fonc.2020.614930
This article is part of the Research Topic Neural Stem Cells of the Subventricular Zone: from Neurogenesis to Glioblastoma Origin View all 14 articles
 Arnaud Lombard1,2†
Arnaud Lombard1,2† Marina Digregorio1†
Marina Digregorio1† Clément Delcamp1
Clément Delcamp1 Bernard Rogister1,3*
Bernard Rogister1,3* Caroline Piette1,4†
Caroline Piette1,4† Natacha Coppieters1†
Natacha Coppieters1†Both in adult and children, high-grade gliomas (WHO grades III and IV) account for a high proportion of death due to cancer. This poor prognosis is a direct consequence of tumor recurrences occurring within few months despite a multimodal therapy consisting of a surgical resection followed by chemotherapy and radiotherapy. There is increasing evidence that glioma stem cells (GSCs) contribute to tumor recurrences. In fact, GSCs can migrate out of the tumor mass and reach the subventricular zone (SVZ), a neurogenic niche persisting after birth. Once nested in the SVZ, GSCs can escape a surgical intervention and resist to treatments. The present review will define GSCs and describe their similarities with neural stem cells, residents of the SVZ. The architectural organization of the SVZ will be described both for humans and rodents. The migratory routes taken by GSCs to reach the SVZ and the signaling pathways involved in their migration will also be described hereafter. In addition, we will debate the advantages of the microenvironment provided by the SVZ for GSCs and how this could contribute to tumor recurrences. Finally, we will discuss the clinical relevance of the SVZ in adult GBM and pediatric HGG and the therapeutic advantages of targeting that neurogenic region in both clinical situations.
Gliomas are the most frequent primary tumors of the central nervous system, both in adults and children. Among them, glioblastoma (GBM), classified as grade IV by the World Health Organization (WHO), is the most frequent in adults, with an overall average annual age-adjusted incidence rate of 3.2 per 100,000 (1, 2). In children, WHO grades III and IV gliomas are generally grouped together as high grade gliomas (HGGs), and are less common, with an annual age-adjusted incidence rate of 0.08 and 0.15 per 100,000, respectively (1). Diffuse intrinsic pontine glioma (DIPG) is the second most common HGG of childhood (3). This tumor has a diffuse growth pattern and is localized in the brainstem. Tumors with identical characteristics are found in other midline structures such as the thalamus and the spinal cord, reason why it has been re-classified as diffuse midline glioma (DMG) in the last WHO 2016 classification (2). In addition, these are phenotypically and molecularly distinct from the other types of HGGs with the characteristic of having histone H3 mutations (2, 3). However, for the interest of this review, DMGs will be included with HGGs in our discussion.
Despite clear molecular and genetic differences between pediatric and adult HGGs, as reviewed by Sturm (4), there seems to be a continuum between these two age groups. For examples, DMG with H3K27M mutations, initially thought to exclusively occur in children, can also be seen in adults (5, 6), while adult GBM characteristics, such as epidermal growth factor receptor (EGFR) amplification, can be seen in adolescents (7). Furthermore, pediatric and adult HGGs share a poor prognosis due to an almost systematic relapse of the tumor despite a multimodal therapy which classically consists of the tumor resection, whenever possible, followed by radiotherapy plus concomitant and adjuvant temozolomide (TMZ) (2, 8). Recurrences most likely emanate from tumor cells which have infiltrated the parenchyma and are practically impossible to successfully eradicate through a complete surgical resection (9). Moreover, some tumor sites are not reachable by surgery which is classically the case for DMG (3).
In addition to cancer cells which have intruded the tumor surroundings, recurrences can be explained by the existence of cancer stem cells (CSCs) or glioma stem cells (GSCs). CSCs-based tumor initiation, growth and maintenance was first proposed over 150 years ago by Virchow, who first suggested that a quiescent sub-population of embryonic stem cells was able to generate tumors (10). Since then, the existence of CSCs in tumors has been demonstrated in various types of cancers (11–13), including in adult GBM (14–17) and pediatric HGG (18–20). Several GSC features supports the hypothesis that these cells contribute to recurrences (16). Indeed, GSCs adapt and survive environmental stresses, present increased resistance to standard therapies and are able to form a novel tumor (21–27). GSCs are mainly present in the tumor mass but have also been detected in the subventricular zone (SVZ), a neurogenic niche persisting after birth and containing resident neural stem cells (NSCs) (19, 21, 23).
The present review will describe the similarities between GSCs and NSCs. The migratory routes of GSCs from the tumor mass to the SVZ and the signaling pathways involved in their migration will also be described hereafter. In addition, we will debate the advantages of the microenvironment provided by the SVZ for GSCs and how this could contribute to tumor recurrences. Finally, we will discuss the clinical relevance of the SVZ in adult GBM and pediatric HGGs and the therapeutic advantages of targeting the SVZ in both clinical situations.
Gimple and colleagues recently suggested the following definition for GSCs; “GSCs are defined by tumor-initiating capacity following serial transplantation, self-renewal, and the ability to recapitulate tumor heterogeneity” (16). These functional characteristics are currently the only tools available for the identification of GSCs as none of the stem marker expression shows sufficient sensitivity and specificity. Indeed, GSCs markers widely overlap with NSC specific ones, thus rendering the identification of GSCs within a heterogeneous tumor rather difficult. Thus, so far, GSCs are still solely recognized based on their functional properties (11, 15).
Multiple signaling pathways involved in normal NSC biology also play a role in GSCs. For example, the Notch pathway is implicated in the maintenance of NSCs in an undifferentiated state via the repression of proneural gene expression and is frequently upregulated in GSCs (28, 29). The Bone Morphogenetic Protein pathway inhibits neurogenesis and promotes gliogenesis in NSCs, while it stimulates astrocyte-like differentiation and reduces proliferation in GSCs (30, 31). The Wnt pathway regulates NSC and GSC proliferation via the accumulation of β-catenin (32, 33). The Sonic Hedgehog pathway is involved in self-renewal of NSCs and GSCs via Gli1 (34, 35). STAT3 is needed for NSC and GSC proliferation and the maintenance of multipotency (36, 37). Finally, EGFR, classically expressed by NSCs and promoting their proliferation, is often overexpressed in GBM and has been associated with tumor initiation, tumor growth, cell invasion, angiogenesis, and resistance to chemo- and radiotherapy (38).
In the same way, multiple transcription factors are common between NSCs and GSCs. Bmi1, a component of the Polycomb Repressive Complex 1, is classically found in undifferentiated NSCs and is involved in the maintenance of their multipotency. It also contributes to glioma aggressiveness via NF-κB and matrix metalloproteinase-9 (39). In addition, the inhibition of c-Myc, a transcription factor involved in the regulation of stem cell renewal and proliferation, triggers GSC apoptosis and reduces neurospheres formation (40). Sox2 which protects NSCs from apoptosis via survivin overexpression (41), is essential for the stemness maintenance of GSCs, together with Oct4 and Nanog (18). Finally, Olig2, a key transcriptional factor normally required for neural progenitor cell (NPC) proliferation (42), is able to reduce the suppressive action of p53 which regulates the proliferation in GSCs (43).
The identification of markers permitting the detection of GSCs is important since it has been estimated that approximately only one GSC every 1,000 tumor cells is present in a GBM tumor (44). Despite this low number of GSCs in the tumor mass, there is now evidence that these cells might contribute to tumor recurrences. Indeed, GSCs can leave the tumor mass, invade the parenchyma and migrate to further locations, including the neurogenic zones, where they escape a surgical intervention (23, 45–47). These GSCs can then remain quiescent until a still unknown mechanism triggers the development of a new tumor (48).
In the adult human brain, there are two well-described neurogenic zones: the SVZ, situated in the walls of the lateral ventricles (LV), and the subgranular zone (SGZ) of the hippocampal dentate gyrus. Although controverted, some evidence suggest that the presence of NSCs is not limited to these two well-known neurogenic niches but could extend to other parts of the human central nervous system (49, 50). In addition, multiple other niches of NSCs have been reported in other mammalians (51). As the SVZ is the largest neurogenic region of the adult brain and because a link between the SGZ and brain tumors is less clear (52), this review will focus on the role of the SVZ in HGG recurrences.
The human SVZ is composed of four distinct layers going from I to IV from the innermost toward the outermost layer. Layer I runs alongside the ventricular cavity and is a monolayer of ependymal cells responsible for the production and secretion of cerebrospinal fluid. Layer II is known as the hypocellular space as it contains cellular processes with only very few cell bodies. Layer III is a cellular ribbon mainly comprising cells expressing glial fibrillary acidic protein (GFAP). Finally, layer IV, the outermost layer adjacent to the parenchyma, is a transition zone mainly composed of myelinated axons and oligodendrocytes (Figure 1) (53). Describing the exact localization of NSCs within the different layers of the SVZ is difficult as it depends on which criteria have been used to define or identify these cells. Recent single-cell RNA sequencing studies helped classifying NSCs into four main populations: quiescent NSCs, activated NSCs, NPCs and neuroblasts (54). Most NSC seem to be quiescent and positive for GFAP and CD133 (55). Mammalian NSCs resembling glial cells and sharing common characteristics and markers including GFAP, are mainly detected in layer III of the SVZ (56). The transcription factor Sox2 has been validated for the detection of NSCs in the human fetal brain (57). Sox2 is expressed by quiescent and activated NSCs, but not by NPCs (57, 58). In adults, Sox2 can be detected in the different layers of the SVZ with decreasing numbers toward the parenchyma indicating the presence of NSC in the adult SVZ (59). However, when considering the expression of the immature neuronal markers such as doublecortin, only rare NSCs are found in the adult human brain and only in layer III (60). Finally, in the SVZ, the proliferative marker Ki67 decreases during aging (61) with a limited number of Ki67 positive cells detected in layer III in the adult SVZ, reflecting very few cycling cells (60). However, it is to note that whereas proliferative cells in juvenile SVZ correspond to different cell types including immature cells, in the adult SVZ, Ki67 is exclusively expressed by microglia (60), the primary resident immune cells of the brain (62).
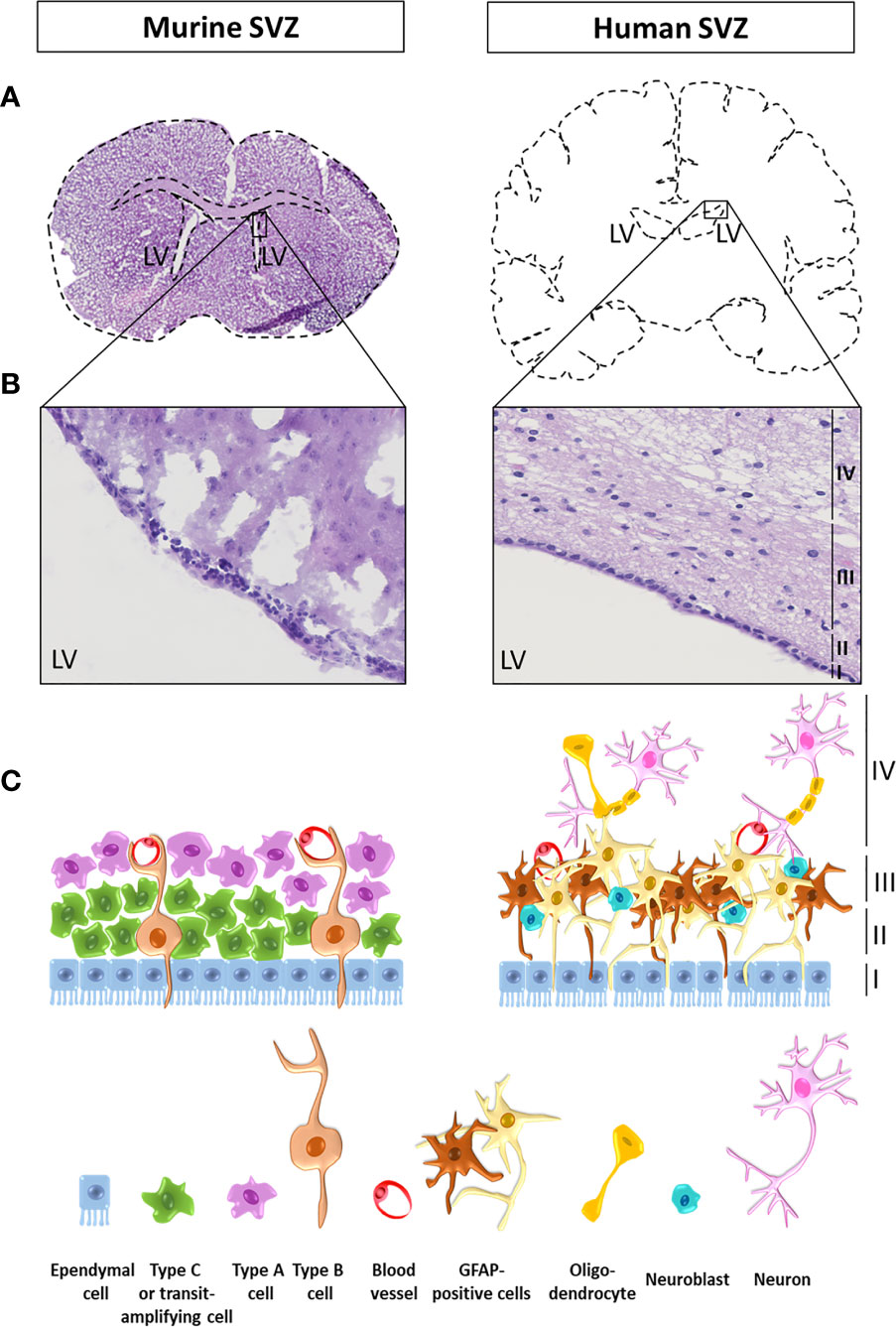
Figure 1 Illustration and schematic representation of a mouse and a human subventricular zone. (A) Coronal sections of a mouse (left) and a human (right) brain at the level of the lateral ventricles (LV). (B) Zoomed images of a mouse and a human brain subventricular zone stained with haematoxylin and eosin. The four human layers are indicated as layers I to IV from the lumen of the LV toward the parenchyma. (C) Schematic representation of the cellular composition of the mouse and the human subventricular zone (SVZ). In adult rodents, neural stem cells (NSCs), also called type B cells, are separated from the LV lumen by ependymal cells. Type B cells undergo asymmetrical cell divisions to give rise to a new type of B cell population with self-renewal properties (one of the hallmarks of a stem cell), as well as transit amplifying progenitor cells, also known as type C cells. Type C cells then migrate to become neural precursor cells including migrating neuroblasts (type A cells) or oligodendrocytes precursor cells. In the SVZ, type B cells display a double contact, one with the ventricle and one with the basal lamina of blood vessels, where the blood-brain barrier (BBB) is not complete. The human SVZ is composed of four distinct layers going from I to IV from the innermost toward the outermost layer. Layer I runs alongside the ventricular cavity and is a monolayer of ependymal cells responsible for the production and secretion of cerebrospinal fluid. Layer II is known as the hypocellular space as it contains cellular processes with only very few cell bodies. Layer III is a cellular ribbon mainly comprising cells expressing glial fibrillary acidic protein (GFAP) and neuroblasts. Finally, layer IV, the outermost layer adjacent to the parenchyma, is a transition zone mainly composed of myelinated axons and oligodendrocytes.
Differences in the cellular composition exist between the SVZ found in children and the adult counterpart, with more proliferative cells in children, indicating higher neurogenesis under the age of four (60, 63). It is also worth to note that in addition to a continuum between children and adult HGGs, there is a progression in the cellular and molecular properties of NSCs hosted in the SVZ from the embryo, through childhood to adulthood (64).
Variations also exist in the organization of the different SVZ layers between species. In adult rodents, NSCs, also called type B1 cells, are separated from the LV lumen by ependymal cells. Type B1 cells undergo asymmetrical cell divisions to give rise to a new type of B1 cell population with self-renewal properties (one of the hallmarks of a stem cell), as well as transit amplifying progenitor cells, also known as type C cells. Type C cells then migrate to become neural precursor cells including migrating neuroblasts (type A cells) or oligodendrocytes precursor cells. In the SVZ, type B cells display a double contact, one with the ventricle and one with the basal lamina of blood vessels, where the blood-brain barrier (BBB) is not complete. B cells are able to form C cells, which in turn divide in A cells that finally migrate and integrate in the olfactory bulb in mice (Figure 1). Note that, in human, the SVZ present a dense layer of B cells while there are just a few of A and C cells. For a complete review on neural stem cells in the adult mammalian brain and a good schematic representation of the SVZ in rodent, we would refer readers to the article from Obernier and Alvarez-Buylla (64, 65).
In non-tumoral brains, white matter tracts can act as a guide, or a motorway, for the migration of NSCs or glial progenitor cells. Multiple evidence highlights a similar pattern of migration for GBM cells. The first evidence of GBM invasion through the white matter tracts was formulated by Scherer and collaborators in 1938 after they studied 100 patients with gliomas. They also demonstrated GBM cell migration through other routes like blood vessels, the neural parenchyma and the subarachnoid space (66). Later, NSCs were transformed to gain tumorigenic capacities before being implanted in mouse brains to model GBM tumor growth and brain invasion. Grafted mice successfully recapitulated GBM tumor development four weeks post-injection. More importantly, two weeks after injection, few GBM cells were detected in the corpus callosum, consisting of white matter tracts connecting the two cerebral hemispheres (67). As reported by our team, the injection of human GSCs in the mouse striatum led to the formation of a tumor. Furthermore, some GSCs left the tumor mass and migrated through the corpus callosum to reach the ipsi- and controlateral SVZ (Figure 2). GSCs were also detected in the olfactory bulb, demonstrating their migration from the SVZ through the rostral migratory stream (45), a structure containing a high density of parallel blood vessels classically used as a scaffold for neuroblasts (68). Another interesting study by Kakita et al. revealed the migration of labeled glial progenitors from the neonatal SVZ through the corpus callosum to the contralateral hemispheres, which correlates with the pattern of migration described by Kroonen et al. (45, 69). More recently, diffusion and magnetic resonance performed on seven glioma and six control patients showed that the human corpus callosum also act as a GBM cell migration track (70). In children, Caretti et al. analyzed a series of autopsies from 16 patients with DMG and observed that tumoral cells spread to the SVZ in 63% of the cases (19). For the last ten years, neurogenic niches have received more and more attention as not only it is the largest site for stem cells persisting in adulthood, but also, as discussed above, it can be a preferred destination site for GSCs (23, 45–47).
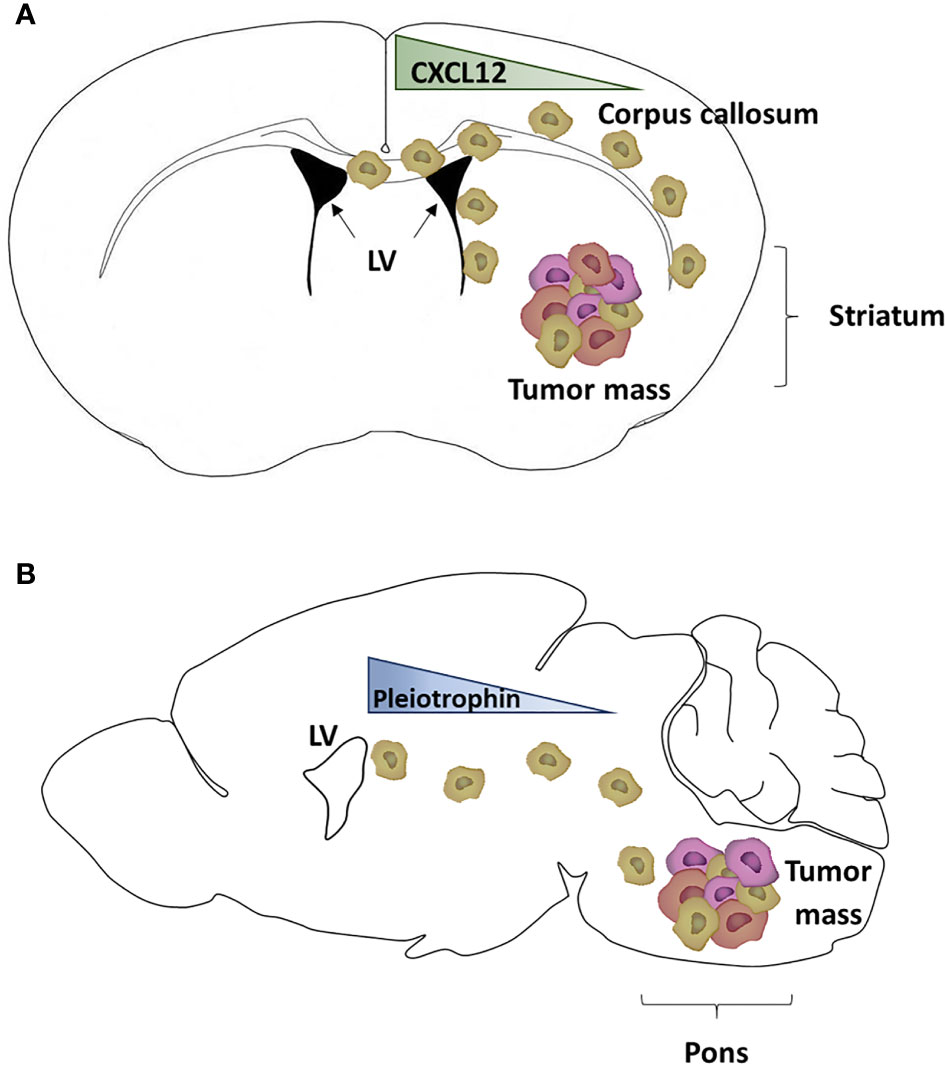
Figure 2 Mouse model of glioblastoma cell invasion into the subventricular zone. (A) Schematic representation of adult glioblastoma (GBM) cells grafted into the right striatum of a mouse brain (schematically drawn as a coronal section ahead of the hippocampus) and generating a tumor mass. Some GBM cells expressing CXCR4 (light brown cells) migrated through the corpus callosum to reach the subventricular zone (SVZ) of the lateral ventricles (LV), following a CXCL12 gradient (45, 46). (B) Schematic representation of pediatric diffuse midline glioma (DMG) cells grafted into the pons of a mouse brain (schematically drawn as a sagittal section) and generating a tumor mass. Some DMG cells (light brown cells) have migrated out of the tumor mass and reached the SVZ, following a gradient of proteins including pleiotrophin (47).
The SVZ secretes various factors including chemokines and other proteins regulating cell migration (71). Some of the pathways involved in the migration pattern of GSCs from the tumor mass toward the SVZ have already been identified (46, 47).
The first axis, CXCL12/CXCR4, has been identified by our team in 2015 (46). CXCL12 is a chemokine acting on two main receptors, CXCR4 and CXCR7 (72–75). We have demonstrated that CXCL12 is a key player in the migration of CXCR4 positive adult GSCs from the tumor mass toward the SVZ (21). In addition to its chemokine activity, CXCL12 is involved in many biological activities (76) including the regulation of cell proliferation and tumor growth (75), favors an epithelio-mesenchymal transition, regulates the expression of GSC cell markers (77), and increases resistance to both radiotherapy and chemotherapy (21, 22, 78, 79). Furthermore, CXCL12 increases cell survival and facilitates DNA double strands break repairs through the recruitment and phosphorylation of nuclear MAP kinase phosphatase 1. It is also interesting to note that both effects induced by CXCL12 (oriented migration and DNA repair) are dependent on a CXCR4 signalization (78). A potential role for CXCR7 in the mediation of CXCL12 effects on GSCs remains to be investigated since these cells also express that receptor (76, 78).
A second migratory axis has been identified by Qin et al. (47) who showed that pleiotrophin, along with three required binding partners (secreted protein acidic and rich in cysteine (SPARC), SPARC-like protein 1 and heat shock protein 90B) is secreted by SVZ NPC and triggers the migration of DMG and adult GBM cells to the SVZ, through activation of the Rho/ROCK pathway (47).
Additional chemokines are known to be expressed in the SVZ-environment and could also contribute to the recruitment of GSCs (21, 46, 47). Using an array, we studied mouse SVZ conditioned media which led to the identification of several chemokines, including CXCL12 already described above. Amongst the other chemokines, we detected CXCL1, CCL5, CXCL10, and CXCL2. Furthermore, a gene expression profiling analysis was performed using real-time PCR Arrays on total RNA extract obtained from microdissected mouse SVZs. Genes were classified into high, basal and low mRNA levels. Amongst 14 genes highly expressed in the SVZ, we identified CXCL12 as well as CX3CL1 (also known as fractalkine), CCL19 and CCL12 which is the homologue of Monocyte Chemoattractant Protein-1 (MCP1/CCL2) in human (46). In a later study, chemokines were detected in condition media obtained from a human SVZ (21). This technical approach allowed the identification of eight chemokines in the conditioned media, within which Macrophage Inflammatory Protein-3 Alpha (MIP-3α/CCL20) was detected at the highest level, followed by interleukin 8 IL-8/CXCL8, MIP-1α/CCL3, Neutrophil-activating peptide 2 (NAP-2/CXCL7), and MCP-1. Interestingly, at least three chemokines detected in the conditioned media obtained from mouse SVZ were also detected in the human one, namely MCP1, Tazarotene-induced gene 2 protein (TIG2/chemerin) and IL16 (21, 46).
To the best of our knowledge, studies from our team were the only one that employed mouse and human SVZ-conditioned media to identify chemokines released by the SVZ. However, the identification of previously described chemokines was based on targeted techniques. Beside our work, numerous transcriptomic and proteomic studies of the SVZ have been performed; however, without specific focus on SVZ-secreted proteins. Indeed, these studies were based on isolated cells from the SVZ or whole SVZ extracts (61, 80–84). Recently, whole proteins forming the extracellular matrix and their associated proteins, respectively named the “matrisome” and “matrisome-associated proteins”, were extracted from 8 to 12-week-old murine SVZ. In the later study, S100 proteins and Serpins were identified as highly soluble in the SVZ-matrisome. An in-depth analysis of the identified SVZ-associated soluble proteins could potentially lead to the discovery of new migratory-related soluble factors (80). In humans, the composition of the SVZ has mainly been studied during development and is based on the characterization of proteins and/or mRNA expression in specific cell-types and/or on whole SVZs (85, 86). Moreover, databases are now available to study mRNA expression in human age-related SVZ: i.e., BrainSpan Atlas of the Developing Human Brain (83, 87). These databases deserve attention as they could help identify potential SVZ-chemokines. In 2016, the comparison of the secretome of human NSC and GSC cell lines, identified 138 proteins differentially expressed (86). Although this analysis was based on cell lines, the identification of NSC-secreted proteins could help interpret and/or validate future large-scale studies based on SVZ-secreted proteins. Indeed, after reviewing the current literature, it is clear that large-scale analysis of human SVZ-chemokines or secreted proteins are still required.
To conclude, various chemokines are expressed and tightly regulated in the SVZ environment. Numerous studies on human SVZ-secreted proteins would highlight new migratory-related factors and/or confirm the one shown in murine SVZ. Interestingly, some of these secreted proteins would be responsible for specific GSCs migration toward this neurogenic niche. Delocalization of GSCs would be responsible for their maintenance even after tumor resection and their role in HGG recurrences. The identification of specific chemokines, the analyses of their role in GSCs migration capacity and the study of their targeting is therefore of interest to better understand and fight HGG recurrences.
The association of GSCs with non-tumoral cells together with soluble factors provides specific intra-tumoral microenvironments known as niches. The niche concept can be described as an environment able to maintain NSC stemness (88). This concept has been transposed to gliomas with GSC maintenance, division and differentiation in specific GBM localisations. Whereas perivascular, perinecrotic and invasive niches clearly exist in GBM, these structures are dynamic and are not always easily distinguishable one from another (89). In addition to GBM stem cells, the cellular components of these tumors include lymphocytes, macrophages, fibroblasts, pericytes, astrocytes, microglial cells, and neurons. GBM heterogeneity also occurs in different part of the tumor mass with niches not clearly distinct from each other. Niches tightly regulate GBM pathogenesis such as GBM cell survival, invasion, immune escape, and metabolic needs as well as stemness maintenance. A hypoxic environment can give rise to necrotic areas surrounded by hypoxic palisading GBM and immune cells. GBM cells can also hijack abnormal blood vessels to constitute an angiogenic niche or use blood vessels to invade surrounding brain parenchyma in the invasive niche.
This section of the review will describe the SVZ microenvironment and give an overview of the benefits that the SVZ can provide for GSCs. As going into details for each of these advantages is beyond the scope of this article, we will provide general information on the subject and briefly discuss some aspects of the SVZ environment that is beneficial for SVZ-nested GSCs.
The SVZ encompasses various cell types involved in the maintenance of endogenous NSCs and provides all soluble factors, nutrients and oxygen required for the regulation of their biological processes (90). Even the composition of the extracellular matrix of the SVZ plays a major role in the regulation of neurogenesis, cell proliferation and migration (80). Thus, this brain region is an interesting niche for GSCs as by providing an environment adapted for NSCs it also gives the same advantages to SVZ-nested GSCs. The interaction of GSCs with the tumor microenvironment is in fact key for the maintenance of their malignancy (91). In addition to the different cell types composing the SVZ (see above), neurogenic niches encompass other cell types able to secrete soluble factors which can directly act on GSCs and regulate major biological processes involved in the development of the pathology. These cells include microglia, NPCs, and cells composing the architecture of a large vasculature network with specialized properties (91). Indeed, the BBB in the SVZ consists of a vasculature lacking astrocyte end-feet and pericyte coverage at sites. In GBM, those specialized blood vessels are altered which leads to the dysregulation of numerous factors in the brain (92, 93).
NSCs from the SVZ present an increased resistance to TMZ and radiation therapy (21, 23). This radioresistance can be explained in part by high expression of the anti-apoptotic proteins Bcl2 and Mcl1 (94). Interestingly, GSCs nested in the SVZ differ from those which remained in the tumor mass, with SVZ-nested GSCs presenting an increased resistance to irradiation (21). This increased resistance is at least partially regulated by the presence of high levels of CXCL12 in the SVZ (95). Indeed, our group has demonstrated that in addition to attracting GSCs in the SVZ, CXCL12 had a protective effect against irradiation (21). The addition of SVZ-conditioned media to human GBM cells led to a decrease in histone variant H2AX phosphorylation on Ser-139 (γH2AX), a reliable molecular marker of DNA damage repair (21). One of the mechanism involved, is the recruitment and the phosphorylation of MKP1, regulated by CXCL12, which in turn regulates DNA strand breaks repair (78). A radioprotective effect of CXCL12 is supported by a recent study by Rajendiran et al. showing that the ubiquitous overexpression of CXCL12 in a mouse model led to a significant increase in the number of multipotent progenitors and increased radioresistance by promoting quiescence (96). Piccirillo et al. studied human GSCs isolated from the tumor mass or the SVZ and found that most GSCs, isolated from different patients, were resistant to TMZ no matter their origin. GSCs were also resistant to cisplatin, an agent previously used for the treatment of GBM (23).
It is evident that multiple other soluble factors present in the SVZ could promote GSC survival. For example, CX3CL1 highly expressed in the adult SVZ (46) promotes NPC survival (97). Interestingly, CXC3L1 and its receptor are both increased in high grade gliomas, with higher CXC3CL1 being associated with shorter overall survival (OS) (98). CXCL1, also secreted by the SVZ, is overexpressed in GBM tumors, provides radioresistance and is associated with a poorer prognosis for patients affected by the disease (99).
Soluble factors secreted in the SVZ also tightly regulate the balance of NSCs between quiescence and proliferation. These extrinsic signals act through the presence of receptor at the surface of NSCs (100). Amongst receptors enriched in quiescent NSCs, there is for example cadherin 2 which has recently been suggested as a biomarker for the prognosis of GBM and as a predictive factor for the response of gliomas to TMZ (101). In addition to soluble cytokines, oxygen, and nutrients can influence the biology of NSC and thus, of GSCs. The importance of oxygen concentrations for the maintenance of stem cell normal physiology has already been reviewed (102). The level of oxygen found in the SVZ and the SGZ is higher than in the other parts of the brain including the cortex and the thalamus (103). This is interesting since higher oxygen level would help maintain NSC, as well as GSC which have reached the SVZ, in a quiescent and undifferentiated states (104). NSCs in the SVZ are in close contact with the BBB which constantly expose them to circulating molecules and nutrients (105). As already mentioned above, the BBB is often altered in GBM brains (92), which could lead to blood vessel leakage and nutrients unbalance in the SVZ and consequently influence GSC quiescence state (105).
More than 10 years ago, a retrospective clinical study described that a GBM directly in contact with the SVZ at diagnosis was associated with invasiveness and multifocal disease in adults (106). The authors also described four patterns allowing a better characterization of the SVZ involvement: SVZ+/Cortex+ (I), SVZ+/Cortex- (II), SVZ-/Cortex+ (III), SVZ-/Cortex- (IV). A first observational study of 69 patients reported a poorer OS in patients with GBM contacting the LV in comparison to patients with GBM not bordering the LV (107). Multiple studies then confirmed that the SVZ involvement at diagnosis or at recurrence was associated with poorer OS (108–110). Recently, Mistry et al. confirmed in a meta-analysis of fifteen studies and in a retrospective study of 207 adult patients, that GBM contact with the LV was associated with lower OS and can be considered as an independent factor of survival (111, 112). In comparison, there was no decreased survival in case of SGZ involvement or corpus callosum invasion (111). Importantly, the proximity with the SVZ does not allow assessing the origin of GBM. Indeed, Han et al. reported that GSCs display similar stem gene expression in GBM with and without LV contact (113). In the same way, Mistry et al. reported that SVZ contact is not associated with molecular signatures in GBM bulk tumor (114). Finally, in the latest study by Comas et al., an analysis of GBM progression in 133 adult patients with primary GBM treated with the standard TMZ-based adjuvant radiochemotherapy showed that GBM in contact with the SVZ appears to be an independent prognostic factor for poorer progression free-survival (PFS) but not for OS. They also reported that SVZ-contacting tumors were associated with a higher rate of contralateral relapses and more aggressive recurrences which they defined as relapses occurring in patients presenting a sudden worsening of their clinical condition before it could be detected by the follow up MRIs taken every 3 months. They concluded that a direct contact of GBM tumors with the SVZ could be used as a prognostic factor (115).
A similar retrospective analysis was recently conducted in 63 children and adolescents (median age of 12.3 years) diagnosed with HGG (116). Tumors contacting the SVZ were found in 54% of the patients and were usually larger than tumors not in contact with the SVZ. Furthermore, patients with SVZ-associated tumors had a decreased survival time (HR = 1.94, 95% CI 1.03–3.64, p = 0.04). Thus, similarly to adult findings, these data suggest that in children and adolescents, the presence of HGG attached to the SVZ is associated with a poorer prognosis (116). Targeting the SVZ could therefore be a common therapeutic target for adult GBM and pediatric HGG.
While more and more studies have highlighted the importance of Gross Total Resection, nay Supratotal Resection in regard to OS (117, 118), it remains unclear how large the resection should be when a GBM tumor touches the SVZ. Some surgeons are indeed reluctant to open the ventricle in order to obtain a complete resection of the tumor, as it has been associated with communicating hydrocephalus or tumor spread among the ependyma or via the cerebro-spinal fluid (119). However, in a retrospective study of 229 GBM adult patients, Behling et al. showed that ventricular opening was not associated with a reduced OS in a multivariate analysis and could therefore be considered to achieve gross total resection (120). Moreover, Saito et al. recently performed a retrospective study with 111 GBM adult patients and reported that a wide ventricle opening (>23.2 mm) is a strong predictive factor for longer OS (121). Information on surgical intervention for pediatric glioma tumors contacting the SVZ is lacking. Thus, retrospective and prospective studies are undeniably needed to confirm those results in children.
As the SVZ involvement worsens the prognosis, it seems sensible to find a way to interfere with it. In this context, many studies have considered the specific ipsilateral, nay bilateral irradiation of the SVZ, even in absence of neuroradiological clues of the presence of tumoral cells in this brain region. In 2016, Smith et al. reviewed the different studies that investigated the advantages of irradiating the SVZ to improve the OS for adult GBM patients (122). We updated their findings to include the latest publications on that topic (Table 1). Evers et al. published the first retrospective study revealing an improvement of PFS after bilateral irradiation of the SVZ with a median dose superior to 43 Gy (PFS: 15.0 vs 7.2 months, p = 0.028) for patients suffering from grade III/IV gliomas (123). In a similar way, another retrospective study of 40 patients reported a better OS if a dose equal or superior to 53.6 Gy was delivered to the ipsilateral SVZ (iSVZ) (124). Inversely, Slotman et al. used the same cut-off of 43 Gy for bilateral SVZ irradiation and did not observe any difference in OS or PFS in their retrospective study. However, and importantly, they reported less distant recurrences in case of a delivered dose greater than 43 Gy to the contralateral ventricle (131). However, the conclusions of those three studies suffer from a limited number of patients (55, 40, and 40, respectively) (123, 124, 131).
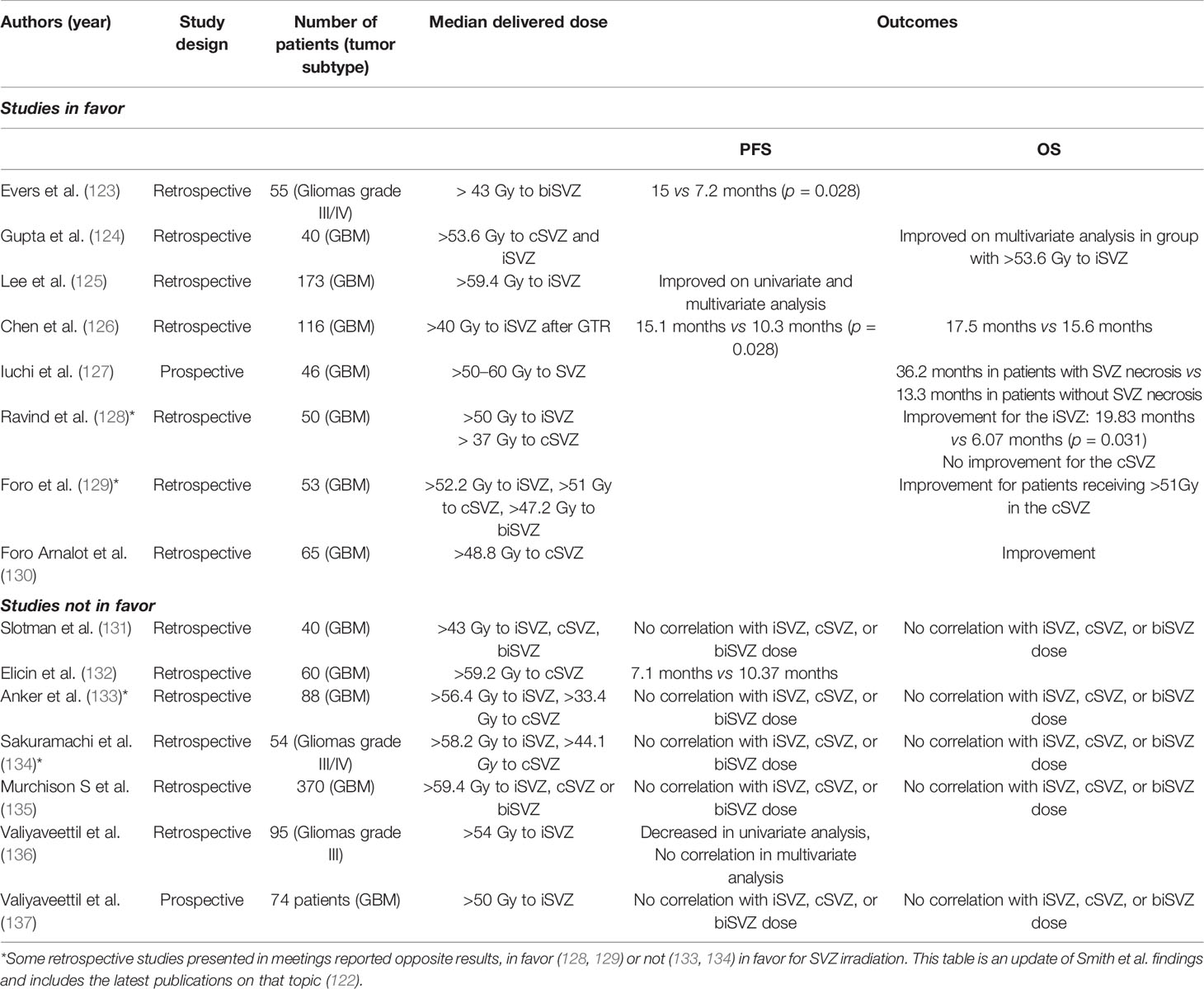
Table 1 Summary of the advantages of irradiating the subventricular zone to improve the overall survival for adult glioblastoma (GBM) patients.
Later, using a larger cohort of 173 patients, Lee et al. retrospectively showed an increased PFS for an ipsilateral SVZ irradiation with a delivered dose superior to 59.4 Gy (125) Interestingly, Chen et al. showed that an increased iSVZ irradiation (superior or equal to 40 Gy) after GTR was associated to a better PFS and OS (126). Another retrospective study showed a poor PFS if the dose delivered to the contro-lateral SVZ (cSVZ) was superior to 59.2Gy (132). Those studies are in fact rather difficult to compare as they are retrospective studies and do not control for important variables such as (i) patient selection, (ii) irradiation dose, (iii) cut-off values, or (iv) importantly, the delineation of the SVZ. Moreover, classical prognostic factors such as MGMT or IDH status have frequently not been considered. Besides, a prospective study initially designed to test hypofractionated high-dose intensity modulated radiation therapy reported a better OS in case of radionecrosis in the SVZ (127).
In 2017, Foro Arnalot et al. reported another retrospective study of 65 patients showing an improvement in PFS but not in OS if the cSVZ received a dose superior or equal to 48.8 Gy (130). Khalifa et al. showed that a dose inferior to 20 Gy for bilateral SVZ irradiation was associated with poor prognosis (138). In 2018, Murchison et al. did not found any correlation between SVZ dose and PFS/OS in a large retrospective study of 370 GBM patients (135). Recently, in a retrospective study of 95 patients suffering from anaplastic gliomas, Valiyaveettil et al. reported no correlation between SVZ dose and PFS/OS in multivariate analysis (136). In a prospective study including 74 GBM patients, the same team did not found any correlation between SVZ dose and PFS/OS (137). Finally, in a short prospective study of 30 GBM patients, the sparing irradiation of neurogenic niches, including SVZ, did not modify PFS or OS in comparison to a matched historical control (139).
In this context, a phase II clinical trial combining standard radio- and chemotherapy to deliver irradiation to the ipsilateral (60 Gy) and the contralateral (46 Gy) SVZ is ongoing (NCT02177578). This study will provide valuable information on benefits that targeting the SVZ could offer for the treatment of GBM. It has to be said that the major constraint to investigate SVZ irradiation more deeply, or even to consider SGZ irradiation, is the fear that it might hasten neurocognitive decline as healthy NSCs would not be spared by the treatment and would suffer along with tumor cells. While some of the cited studies did not show a correlation between the delivered irradiation dose to SVZ and changes in performance status, Iuchi et al. found that high dose radiations deliver to the SVZ, leads to radionecrosis and better OS. However, it also results in progressive decline in Karnofsky performance status which measures the ability for a patient to carry out daily tasks (127). Henceforth, it is not surprising that some clinical trials focused on sparing the SVZ and the hippocampus.
In children, retrospective and prospective studies suggest an association between neurocognitive deficits and radiation dose to the hippocampus hosting the SGZ, but not the SVZ. Due to the rarity of HGG in children, the link between SVZ irradiation and survival has not yet been investigated (126, 140–143).
Many current researches aim at targeting GSCs. To do so in an efficient manner, it is important to know what to target. The most common pathways involved in GSC maintenance include the Wnt, the Sonic hedgehog (SHH) and the Notch pathways. As recently discussed by Sharifzad et al., targeting these pathways could help eradicating GSCs or increase chemotherapy efficiency (144). To target GSCs nested in the SVZ, it has been suggested to use perphenazine, an inhibitor of the dopamine receptor D3, in order to block the migration of GBM cells to the SVZ (145). As the activation of the dopamine receptor D3 in SVZ cells is associated with their proliferation in vitro (146), its blockade by perphenazine could also maintain GSC in a quiescent state in the SVZ. Bardella et al. reviewed another interesting approach which consists of interfering with the SVZ inflammatory environment as it might predispose cells to mutations and worsen cancer phenotypes (147). However, while it has been largely reported that microglia participate in GBM progression locally by adopting an anti-inflammatory state (148), their interaction and effects on SVZ-nested GSCs remain to be proven.
HGG account for a high proportion of death resulting from cancer, both in adults and children. Unfortunately, survival has not been significantly improved over the last decades. Both bench and bedside evidences strongly support the involvement of GSCs and SVZ in HGG recurrences. We and others, previously demonstrated that GSCs migrate from the tumor mass toward the SVZ, through the CXCL12/CXCR4 axis or through a pleiotrophin-driven axis. Once hosted in the SVZ, GSCs benefit from a protective environment providing increased resistance to irradiation and chemotherapy, before these cells get reactivated by a still unknown mechanism and recolonize the TM or invade other sites. In adults, the benefit/risk balance of targeting the SVZ by surgery and/or radiotherapy was investigated in clinical settings; however, the review of the current literature does not permit a clear conclusion yet. In children, it has not been evaluated and should be further investigated. Other technical approaches to target the SVZ also remain to be explored. Blocking the migration of GSCs toward the SVZ is probably not an option, given that the cells would already have migrated out at the time of the diagnosis. Other possibilities could be to decrease or to block the recolonization of the TM. The cancer cell trap approach is another interesting and original concept that exploits the migratory potential of cancer cells in order to concentrate them toward specific locations (149). This approach has been showed to reduce the metastatic potential of human breast cancer cells implanted in female mice, through biomaterial scaffolds implanted in peritoneal fat pads (150, 151). This kind of approach should definitely be further investigated in the context of pediatric and adult HGG and DMG and could be combined with local targeted therapies.
In conclusion, there is strong evidence that the migration of GSCs toward the SVZ is implicated in HGG recurrences, both in adults and in children. The exact mechanisms supporting this process should be further investigated with the perspective of specifically targeting this particular cell population.
AL, MD, and CD performed the literature review, and wrote the manuscript. BR edited the manuscript. CP and NC conceived, supervised, and edited the manuscript. All authors contributed to the article and approved the submitted version.
AL received the Clinical Researcher of the FNRS-Belgium. MD is a PhD student funded by the TELEVIE-FNRS. CD is PhD student supported by the EUROMA funds (Fondation Léon Frédéricq, University of Liège). NC is supported by the Neurological Foundation of New Zealand (Philip Wrightson Postdoctoral Fellowship). BR research group is supported by the FNRS, the University of Liège and the Fonds Léon Frédéricq.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The authors would like to thank the National Fund for Scientific Research (F.N.R.S.), the Télévie sub-organization, the Anti-Cancer Centre Léon Frédéricq Foundation and the Neurological Foundation of New Zealand, for their financial support.
1. Ostrom QT, Gittleman H, Fulop J, Liu M, Blanda R, Kromer C, et al. CBTRUS statistical Report: primary brain and central nervous system tumors diagnosed in the United States in 2008-2012. Neuro Oncol (2015) 17:iv1–iv62. doi: 10.1093/neuonc/nov189
2. Louis DN, Perry A, Reifenberger G, von Deimling A, Figarella-Branger D, Cavenee WK, et al. The 2016 World Health Organization Classification of Tumors of the Central Nervous System: a summary. Acta Neuropathol (2016) 131:803–20. doi: 10.1007/s00401-016-1545-1
3. Johung T, Monje M. Diffuse Intrinsic Pontine Glioma: New Pathophysiological Insights and Emerging Therapeutic Targets. Curr Neuropharmacol (2017) 15:88–97. doi: 10.2174/1570159x14666160509123229
4. Sturm D, Bender S, Jones DTW, Lichter P, Grill J, Becher O, et al. Paediatric and adult glioblastoma: Multiform (epi)genomic culprits emerge. Nat Rev Cancer (2014) 14:92–107. doi: 10.1038/nrc3655
5. Ebrahimi A, Skardelly M, Schuhmann MU, Ebinger M, Reuss D, Neumann M, et al. High frequency of H3 K27M mutations in adult midline gliomas. J Cancer Res Clin Oncol (2019) 145:839–50. doi: 10.1007/s00432-018-02836-5
6. Luo Y, Zeng L, Xie XQ, Wang F, Liu YZ, Kang JB, et al. H3K27M mutant diffuse midline glioma: A case report. Eur Rev Med Pharmacol Sci (2020) 24:2579–84. doi: 10.26355/eurrev_202003_20527
7. Schroeder KM, Hoeman CM, Becher OJ. Children are not just little adults: Recent advances in understanding of diffuse intrinsic pontine glioma biology. Pediatr Res (2014) 75:205–9. doi: 10.1038/pr.2013.194
8. Stupp R, Mason WP, van den Bent MJ, Weller M, Fisher B, Taphoorn MJB, et al. Radiotherapy plus Concomitant and Adjuvant Temozolomide for Glioblastoma. N Engl J Med (2005) 352:987–96. doi: 10.1056/NEJMoa043330
9. Lara-Velazquez M, Al-Kharboosh R, Jeanneret S, Vazquez-Ramos C, Mahato D, Tavanaiepour D, et al. Advances in brain tumor surgery for glioblastoma in adults. Brain Sci (2017) 7:166–82. doi: 10.3390/brainsci7120166
10. David H. Rudolf Virchow and Modern Aspects of Tumor Pathology. Pathol Res Pract (1988) 183:356–64. doi: 10.1016/S0344-0338(88)80138-9
11. Yi Y, Hsieh IY, Huang X, Li J, Zhao W. Glioblastoma stem-like cells: Characteristics, microenvironment, and therapy. Front Pharmacol (2016) 7:477–91. doi: 10.3389/fphar.2016.00477
12. Galli R, Binda E, Orfanelli U, Cipelletti B, Gritti A, De Vitis S, et al. Isolation and characterization of tumorigenic, stem-like neural precursors from human glioblastoma. Cancer Res (2004) 64:7011–21. doi: 10.1158/0008-5472.CAN-04-1364
13. Lapidot T, Sirard C, Vormoor J, Murdoch B, Hoang T, Caceres-Cortes J, et al. A cell initiating human acute myeloid leukaemia after transplantation into SCID mice. Nature (1994) 367:645–8. doi: 10.1038/367645a0
14. Singh SK, Hawkins C, Clarke ID, Squire JA, Bayani J, Hide T, et al. Identification of human brain tumour initiating cells. Nature (2004) 432:396–401. doi: 10.1038/nature03128
15. Brescia P, Richichi C, Pelicci G. Current strategies for identification of glioma stem cells: adequate or unsatisfactory? J Oncol (2012) 2012:376894. doi: 10.1155/2012/376894
16. Gimple RC, Bhargava S, Dixit D, Rich JN. Glioblastoma stem cells: Lessons from the tumor hierarchy in a lethal cancer. Genes Dev (2019) 33:591–609. doi: 10.1101/gad.324301.119
17. Suvà ML, Tirosh I. The Glioma Stem Cell Model in the Era of Single-Cell Genomics. Cancer Cell (2020) 37:630–6. doi: 10.1016/j.ccell.2020.04.001
18. Song W-S, Yang Y-P, Huang C-S, Lu K-H, Liu W-H, Wu W-W, et al. Sox2, a stemness gene, regulates tumor-initiating and drug-resistant properties in CD133-positive glioblastoma stem cells. J Chin Med Assoc (2016) 79:538–45. doi: 10.1016/j.jcma.2016.03.010
19. Caretti V, Bugiani M, Freret M, Schellen P, Jansen M, van Vuurden D, et al. Subventricular spread of diffuse intrinsic pontine glioma. Acta Neuropathol (2014) 128:605–7. doi: 10.1007/s00401-014-1307-x
20. Hemmati HD, Nakano I, Lazareff JA, Masterman-Smith M, Geschwind DH, Bronner-Fraser M, et al. Cancerous stem cells can arise from pediatric brain tumors. Proc Natl Acad Sci U S A (2003) 100:15178–83. doi: 10.1073/pnas.2036535100
21. Goffart N, Lombard A, Lallemand F, Kroonen J, Nassen J, Di Valentin E, et al. CXCL12 mediates glioblastoma resistance to radiotherapy in the subventricular zone. Neuro Oncol (2017) 19:66–77. doi: 10.1093/neuonc/now136
22. Wang S, Chen C, Li J, Xu X, Chen W, Li F. The CXCL12/CXCR4 axis confers temozolomide resistance to human glioblastoma cells via up-regulation of FOXM1. J Neurol Sci (2020) 414:116837. doi: 10.1016/j.jns.2020.116837
23. Piccirillo SGM, Spiteri I, Sottoriva A, Touloumis A, Ber S, Price SJ, et al. Contributions to drug resistance in glioblastoma derived from malignant cells in the sub-ependymal zone. Cancer Res (2015) 75:194–202. doi: 10.1158/0008-5472.CAN-13-3131
24. Dedobbeleer M, Willems E, Freeman S, Lombard A, Goffart N, Rogister B. Phosphatases and solid tumors: Focus on glioblastoma initiation, progression and recurrences. Biochem J (2017) 474:2903–24. doi: 10.1042/BCJ20170112
25. Blough MD, Westgate MR, Beauchamp D, Kelly JJ, Stechishin O, Ramirez AL, et al. Sensitivity to temozolomide in brain tumor initiating cells. Neuro Oncol (2010) 12:756–60. doi: 10.1093/neuonc/noq032
26. Bao S, Wu Q, McLendon RE, Hao Y, Shi Q, Hjelmeland AB, et al. Glioma stem cells promote radioresistance by preferential activation of the DNA damage response. Nature (2006) 444:756. doi: 10.1038/nature05236
27. Chen J, Li Y, Yu TS, McKay RM, Burns DK, Kernie SG, et al. A restricted cell population propagates glioblastoma growth after chemotherapy. Nature (2012) 488:522–6. doi: 10.1038/nature11287
28. Fan X, Khaki L, Zhu TS, Soules ME, Talsma CE, Gul N, et al. NOTCH pathway blockade depletes CD133-positive glioblastoma cells and inhibits growth of tumor neurospheres and xenografts. Stem Cells (2010) 28:5–16. doi: 10.1002/stem.254
29. Imayoshi I, Sakamoto M, Yamaguchi M, Mori K, Kageyama R. Essential roles of Notch signaling in maintenance of neural stem cells in developing and adult brains. J Neurosci (2010) 30:3489–98. doi: 10.1523/JNEUROSCI.4987-09.2010
30. Lim DA, Tramontin AD, Trevejo JM, Herrera DG, García-Verdugo JM, Alvarez-Buylla A. Noggin antagonizes BMP signaling to create a niche for adult neurogenesis. Neuron (2000) 28:713–26. doi: 10.1016/S0896-6273(00)00148-3
31. Lee J, Son MJ, Woolard K, Donin NM, Li A, Cheng CH, et al. Epigenetic-mediated dysfunction of the bone morphogenetic protein pathway inhibits differentiation of glioblastoma-initiating cells. Cancer Cell (2008) 13:69–80. doi: 10.1016/j.ccr.2007.12.005
32. Kaur N, Chettiar S, Rathod S, Rath P, Muzumdar D, Shaikh ML, et al. Wnt3a mediated activation of Wnt/β-catenin signaling promotes tumor progression in glioblastoma. Mol Cell Neurosci (2013) 54:44–57. doi: 10.1016/j.mcn.2013.01.001
33. Ming G, Song H. Adult neurogenesis in the mammalian central nervous system. Annu Rev Neurosci (2005) 28:223–50. doi: 10.1146/annurev.neuro.28.051804.101459
34. Bar EE, Chaudhry A, Lin A, Fan X, Schreck K, Matsui W, et al. Cyclopamine-mediated hedgehog pathway inhibition depletes stem-like cancer cells in glioblastoma. Stem Cells (2007) 25:2524–33. doi: 10.1634/stemcells.2007-0166
35. Ahn S, Joyner AL. In vivo analysis of quiescent adult neural stem cells responding to Sonic hedgehog. Nature (2005) 437:894. doi: 10.1038/nature03994
36. de la Iglesia N, Konopka G, Puram SV, Chan JA, Bachoo RM, You MJ, et al. Identification of a PTEN-regulated STAT3 brain tumor suppressor pathway. Genes Dev (2008) 22:0. doi: 10.1101/gad.1606508
37. Sherry MM, Reeves A, Wu JK, Cochran BH. STAT3 is required for proliferation and maintenance of multipotency in glioblastoma stem cells. Stem Cells (2009) 27:2383–92. doi: 10.1002/stem.185
38. Zahonero C, Sánchez-Gómez P. EGFR-dependent mechanisms in glioblastoma: towards a better therapeutic strategy. Cell Mol Life Sci (2014) 71:3465–88. doi: 10.1007/s00018-014-1608-1
39. Jiang L, Wu J, Yang Y, Liu L, Song L, Li J, et al. Bmi-1 promotes the aggressiveness of glioma via activating the NF-kappaB/MMP-9 signaling pathway. BMC Cancer (2012) 12:406. doi: 10.1186/1471-2407-12-406
40. Wang J, Wang H, Li Z, Wu Q, Lathia JD, McLendon RE, et al. c-Myc is required for maintenance of glioma cancer stem cells. PloS One (2008) 3:e3769. doi: 10.1371/journal.pone.0003769
41. Feng R, Zhou S, Liu Y, Song D, Luan Z, Dai X, et al. Sox2 protects neural stem cells from apoptosis via up-regulating survivin expression. Biochem J (2013) 450:459–68. doi: 10.1042/BJ20120924
42. Ligon KL, Huillard E, Mehta S, Kesari S, Liu H, Alberta JA, et al. Olig2-Regulated Lineage-Restricted Pathway Controls Replication Competence in Neural Stem Cells and Malignant Glioma. Neuron (2007) 53:503–17. doi: 10.1016/j.neuron.2007.01.009
43. Mehta S, Huillard E, Kesari S, Maire CL, Golebiowski D, Harrington EP, et al. The central nervous system-restricted transcription factor Olig2 opposes p53 responses to genotoxic damage in neural progenitors and malignant glioma. Cancer Cell (2011) 19:359–71. doi: 10.1016/j.ccr.2011.01.035
44. Vermeulen L, de Sousa e Melo F, Richel DJ, Medema JP. The developing cancer stem-cell model: clinical challenges and opportunities. Lancet Oncol (2012) 13:e83–9. doi: 10.1016/S1470-2045(11)70257-1
45. Kroonen J, Nassen J, Boulanger Y-G, Provenzano F, Capraro V, Bours V, et al. Human glioblastoma-initiating cells invade specifically the subventricular zones and olfactory bulbs of mice after striatal injection. Int J Cancer (2011) 129:574–85. doi: 10.1002/ijc.25709
46. Goffart N, Kroonen J, Di Valentin E, Dedobbeleer M, Denne A, Martinive P, et al. Adult mouse subventricular zones stimulate glioblastoma stem cells specific invasion through CXCL12/CXCR4 signaling. Neuro Oncol (2015) 17:81–94. doi: 10.1093/neuonc/nou144
47. Qin EY, Cooper DD, Abbott KL, Lennon J, Nagaraja S, Mackay A, et al. Neural Precursor-Derived Pleiotrophin Mediates Subventricular Zone Invasion by Glioma. Cell (2017) 170:845–59.e19. doi: 10.1016/j.cell.2017.07.016
48. Yeh AC, Ramaswamy S. Mechanisms of cancer cell dormancy-another hallmark of cancer? Cancer Res (2015) 75:5014–22. doi: 10.1158/0008-5472.CAN-15-1370
49. Marlier Q, Verteneuil S, Vandenbosch R, Malgrange B. Mechanisms and functional significance of stroke-induced neurogenesis. Front Neurosci (2015) 9:458. doi: 10.3389/fnins.2015.00458
50. Barnabé-Heider F, Göritz C, Sabelström H, Takebayashi H, Pfrieger FW, Meletis K, et al. Origin of new glial cells in intact and injured adult spinal cord. Cell Stem Cell (2010) 7:470–82. doi: 10.1016/j.stem.2010.07.014
51. Pellegrino G, Trubert C, Terrien J, Pifferi F, Leroy D, Loyens A, et al. A comparative study of the neural stem cell niche in the adult hypothalamus of human, mouse, rat and gray mouse lemur (Microcebus murinus). J Comp Neurol (2018) 526:1419–43. doi: 10.1002/cne.24376
52. Chesler DA, Berger MS, Quinones-Hinojosa A. The potential origin of glioblastoma initiating cells. Front Biosci - Schol Ed (2012) 4 S:190–205. doi: 10.2741/s261
53. Quiñones-Hinojosa A, Chaichana K. The human subventricular zone: A source of new cells and a potential source of brain tumors. Exp Neurol (2007) 205:313–24. doi: 10.1016/j.expneurol.2007.03.016
54. Dulken BW, Leeman DS, Boutet SC, Hebestreit K, Brunet A. Single-Cell Transcriptomic Analysis Defines Heterogeneity and Transcriptional Dynamics in the Adult Neural Stem Cell Lineage. Cell Rep (2017) 18:777–90. doi: 10.1016/j.celrep.2016.12.060
55. Kriegstein A, Alvarez-Buylla A. The glial nature of embryonic and adult neural stem cells. Annu Rev Neurosci (2009) 32:149–84. doi: 10.1146/annurev.neuro.051508.135600
56. Wang S, Chandler-Militello D, Lu G, Roy NS, Zielke A, Auvergne R, et al. Prospective identification, isolation, and profiling of a telomerase-expressing subpopulation of human neural stem cells, using sox2 enhancer-directed fluorescence-activated cell sorting. J Neurosci (2010) 30:14635–48. doi: 10.1523/JNEUROSCI.1729-10.2010
57. Codega P, Silva-Vargas V, Paul A, Maldonado-Soto AR, DeLeo AM, Pastrana E, et al. Prospective Identification and Purification of Quiescent Adult Neural Stem Cells from Their In Vivo Niche. Neuron (2014) 82:545–59. doi: 10.1016/j.neuron.2014.02.039
58. Michaelidesová A, Konířová J, Bartůněk P, Zíková M. Effects of Radiation Therapy on Neural Stem Cells. Genes (Basel) (2019) 10:640. doi: 10.3390/genes10090640
59. Brazel CY, Limke TL, Osborne JK, Miura T, Cai J, Pevny L, et al. Sox2 expression defines a heterogeneous population of neurosphere-forming cells in the adult murine brain. Aging Cell (2005) 4:197–207. doi: 10.1111/j.1474-9726.2005.00158.x
60. Dennis CV, Suh LS, Rodriguez ML, Kril JJ, Sutherland GT. Human adult neurogenesis across the ages: An immunohistochemical study. Neuropathol Appl Neurobiol (2016) 42:621–38. doi: 10.1111/nan.12337
61. Barry G, Guennewig B, Fung S, Kaczorowski D, Weickert CS. Long non-coding RNA expression during aging in the human subependymal zone. Front Neurol (2015) 6:45–55. doi: 10.3389/fneur.2015.00045
62. Smith AM, Dragunow M. The human side of microglia. Trends Neurosci (2014) 37:125–35. doi: 10.1016/j.tins.2013.12.001
63. Coletti AM, Singh D, Kumar S, Shafin TN, Briody PJ, Babbitt BF, et al. Characterization of the ventricular-subventricular stem cell niche during human brain develovnpment. Dev (2018) 145:145–57. doi: 10.1242/dev.170100
64. Obernier K, Alvarez-Buylla A. Neural stem cells: Origin, heterogeneity and regulation in the adult mammalian brain. Dev (2019) 146. doi: 10.1242/dev.156059
65. Ponti G, Obernier K, Guinto C, Jose L, Bonfanti L, Alvarez-Buylla A. Cell cycle and lineage progression of neural progenitors in the ventricular-subventricular zones of adult mice. Proc Natl Acad Sci U S A (2013) 110:1045–54. doi: 10.1073/pnas.1219563110
66. Scherer HJ. Structural development in gliomas. Am J Cancer (1938) 34:333–51. doi: 10.1158/ajc.1938.333
67. Sampetrean O, Saga I, Nakanishi M, Sugihara E, Fukaya R, Onishi N, et al. Invasion precedes tumor mass formation in a malignant brain tumor model of genetically modified neural stem cells. Neoplasia (2011) 13:784–91. doi: 10.1593/neo.11624
68. Cuddapah VA, Robel S, Watkins S, Sontheimer H. A neurocentric perspective on glioma invasion. Nat Rev Neurosci (2014) 15:455–65. doi: 10.1038/nrn3765
69. Kakita A, Zerlin M, Takahashi H, Goldman JE. Some glial progenitors in the neonatal subventricular zone migrate through the corpus callosum to the contralateral cerebral hemisphere. J Comp Neurol (2003) 458:381–8. doi: 10.1002/cne.10597
70. Sharifi G, Pajavand AM, Nateghinia S, Meybodi TE, Hasooni H. Glioma Migration Through the Corpus Callosum and the Brainstem Detected by Diffusion and Magnetic Resonance Imaging: Initial Findings. Front Hum Neurosci (2020) 13:472. doi: 10.3389/fnhum.2019.00472
71. Dillen Y, Kemps H, Gervois P, Wolfs E, Bronckaers A. Adult Neurogenesis in the Subventricular Zone and Its Regulation After Ischemic Stroke: Implications for Therapeutic Approaches. Transl Stroke Res (2020) 11:60–79. doi: 10.1007/s12975-019-00717-8
72. Meyrath M, Szpakowska M, Zeiner J, Massotte L, Merz MP, Benkel T, et al. The atypical chemokine receptor ACKR3/CXCR7 is a broad-spectrum scavenger for opioid peptides. Nat Commun (2020) 11:1–16. doi: 10.1038/s41467-020-16664-0
73. Yang L, Jackson E, Woerner BM, Perry A, Piwnica-Worms D, Rubin JB. Blocking CXCR4-mediated cyclic AMP suppression inhibits brain tumor growth in vivo. Cancer Res (2007) 67:651–8. doi: 10.1158/0008-5472.CAN-06-2762
74. Rubin JB, Kung AL, Klein RS, Chan JA, Sun YP, Schmidt K, et al. A small-molecule antagonist of CXCR4 inhibits intracranial growth of primary brain tumors. Proc Natl Acad Sci U S A (2003) 100:13513–8. doi: 10.1073/pnas.2235846100
75. Lee CC, Lai JH, Hueng DY, Ma HI, Chung YC, Sun Yy, et al. Disrupting the CXCL12/CXCR4 axis disturbs the characteristics of glioblastoma stem-like cells of rat RG2 glioblastoma. Cancer Cell Int (2013) 13:85. doi: 10.1186/1475-2867-13-85
76. Würth R, Bajetto A, Harrison JK, Barbieri F, Florio T. CXCL12 modulation of CXCR4 and CXCR7 activity in human glioblastoma stem-like cells and regulation of the tumor microenvironment. Front Cell Neurosci (2014) 8:1–19. doi: 10.3389/fncel.2014.00144
77. Zhu Y, Yang P, Wang Q, Hu J, Xue J, Li G, et al. The effect of CXCR4 silencing on epithelial-mesenchymal transition related genes in glioma U87 cells. Anat Rec (2013) 296:1850–6. doi: 10.1002/ar.22821
78. Dedobbeleer M, Willems E, Lambert J, Lombard A, Digregorio M, Lumapat PN, et al. MKP1 phosphatase is recruited by CXCL12 in glioblastoma cells and plays a role in DNA strand breaks repair. Carcinogenesis (2019) 41(4):417–29. doi: 10.1093/carcin/bgz151
79. Willems E, Dedobbeleer M, Digregorio M, Lombard A, Goffart N, Lumapat PN, et al. Aurora A plays a dual role in migration and survival of human glioblastoma cells according to the CXCL12 concentration. Oncogene (2019) 38:73–87. doi: 10.1038/s41388-018-0437-3
80. Kjell J, Fischer-Sternjak J, Thompson AJ, Friess C, Sticco MJ, Salinas F, et al. Defining the Adult Neural Stem Cell Niche Proteome Identifies Key Regulators of Adult Neurogenesis. Cell Stem Cell (2020) 26:277–93.e8. doi: 10.1016/j.stem.2020.01.002
81. Mcginn MJ, Colello RJ, Sun D. Age-related proteomic changes in the subventricular zone and their association with neural stem/progenitor cell proliferation. J Neurosci Res (2012) 90:1159–68. doi: 10.1002/jnr.23012
82. Fietz SA, Lachmann R, Brandl H, Kircher M, Samusik N, Lakshmanaperumal N, et al. Transcriptomes of germinal zones of human and mouse fetal neocortex suggest a role of extracellular matrix in progenitor self-renewal. Proc Natl Acad Sci U S A (2012) 109:11836–41. doi: 10.1073/pnas.1209647109
83. Miller JA, Ding SL, Sunkin SM, Smith KA, Ng L, Szafer A, et al. Transcriptional landscape of the prenatal human brain. Nature (2014) 508:199–206. doi: 10.1038/nature13185
84. Oldham MC, Konopka G, Iwamoto K, Langfelder P, Kato T, Horvath S, et al. Functional organization of the transcriptome in human brain. Nat Neurosci (2008) 11:1271–82. doi: 10.1038/nn.2207
85. Tome-Garcia J, Tejero R, Nudelman G, Yong RL, Sebra R, Wang H, et al. Prospective Isolation and Comparison of Human Germinal Matrix and Glioblastoma EGFR+ Populations with Stem Cell Properties. Stem Cell Rep (2017) 8:1421–9. doi: 10.1016/j.stemcr.2017.03.019
86. Okawa S, Gagrica S, Blin C, Ender C, Pollard SM, Krijgsveld J. Proteome and Secretome Characterization of Glioblastoma-Derived Neural Stem Cells. Stem Cells (2017) 35:967–80. doi: 10.1002/stem.2542
87. Vied CM, Freudenberg F, Wang Y, Raposo AASF, Feng D, Nowakowski RS. A multi-resource data integration approach: Identifcation of candidate genes regulating cell proliferation during neocortical development. Front Neurosci (2014) 8:257. doi: 10.3389/fnins.2014.00257
88. Mirzadeh Z, Merkle FT, Soriano-Navarro M, Garcia-Verdugo JM, Alvarez-Buylla A. Neural Stem Cells Confer Unique Pinwheel Architecture to the Ventricular Surface in Neurogenic Regions of the Adult Brain. Cell Stem Cell (2008) 3:265–78. doi: 10.1016/j.stem.2008.07.004
89. Schiffer D, Annovazzi L, Casalone C, Corona C, Mellai M. Glioblastoma: Microenvironment and niche concept. Cancers (Basel) (2019) 11:5–23. doi: 10.3390/cancers11010005
90. Sinnaeve J, Mobley BC, Ihrie RA. Space Invaders: Brain Tumor Exploitation of the Stem Cell Niche. Am J Pathol (2018) 188:29–38. doi: 10.1016/j.ajpath.2017.08.029
91. Goffart N, Kroonen J, Rogister B. Glioblastoma-initiating cells: Relationship with neural stem cells and the micro-environment. Cancers (Basel) (2013) 5:1049–71. doi: 10.3390/cancers5031049
92. Arvanitis CD, Ferraro GB, Jain RK. The blood–brain barrier and blood–tumour barrier in brain tumours and metastases. Nat Rev Cancer (2020) 20:26–41. doi: 10.1038/s41568-019-0205-x
93. Xu C, Liu X, Geng Y, Bai Q, Pan C, Sun Y, et al. Patient-derived DIPG cells preserve stem-like characteristics and generate orthotopic tumors. Oncotarget (2017) 8:76644–55. doi: 10.18632/oncotarget.19656
94. Cameron BD, Traver G, Roland JT, Brockman AA, Dean D, Johnson L, et al. Bcl2-Expressing Quiescent Type B Neural Stem Cells in the Ventricular-Subventricular Zone Are Resistant to Concurrent Temozolomide/X-Irradiation. Stem Cells (2019) 37:1629–39. doi: 10.1002/stem.3081
95. Kokovay E, Goderie S, Wang Y, Lotz S, Lin G, Sun Y, et al. Adult SVZ lineage cells home to and leave the vascular niche via differential responses to SDF1/CXCR4 signaling. Cell Stem Cell (2010) 7:163–73. doi: 10.1016/j.stem.2010.05.019
96. Rajendiran S, Smith-Berdan S, Kunz L, Risolino M, Selleri L, Schroeder T, et al. Ubiquitous overexpression of CXCL12 confers radiation protection and enhances mobilization of hematopoietic stem and progenitor cells. Stem Cells (2020) 38(9)1159–74. doi: 10.1002/stem.3205
97. Krathwohl MD, Kaiser JL. Chemokines promote quiescence and survival of human neural progenitor cells. Stem Cells (2004) 22:109–18. doi: 10.1634/stemcells.22-1-109
98. Erreni M, Solinas G, Brescia P, Osti D, Zunino F, Colombo P, et al. Human glioblastoma tumours and neural cancer stem cells express the chemokine CX3CL1 and its receptor CX3CR1. Eur J Cancer (2010) 46:3383–92. doi: 10.1016/j.ejca.2010.07.022
99. Alafate W, Li X, Zuo J, Zhang H, Xiang J, Wu W, et al. Elevation of CXCL1 indicates poor prognosis and radioresistance by inducing mesenchymal transition in glioblastoma. CNS Neurosci Ther (2020) 26:475–85. doi: 10.1111/cns.13297
100. Morizur L, Chicheportiche A, Gauthier LR, Daynac M, Boussin FD, Mouthon MA. Distinct Molecular Signatures of Quiescent and Activated Adult Neural Stem Cells Reveal Specific Interactions with Their Microenvironment. Stem Cell Rep (2018) 11:565–77. doi: 10.1016/j.stemcr.2018.06.005
101. Chen Q, Cai J, Jiang C. CDH2 expression is of prognostic significance in glioma and predicts the efficacy of temozolomide therapy in patients with glioblastoma. Oncol Lett (2018) 15:7415–22. doi: 10.3892/ol.2018.8227
102. Mohyeldin A, Garzón-Muvdi T, Quiñones-Hinojosa A. Oxygen in stem cell biology: A critical component of the stem cell niche. Cell Stem Cell (2010) 7:150–61. doi: 10.1016/j.stem.2010.07.007
103. Zhang K, Zhou Y, Zhao T, Wu L, Huang X, Wu K, et al. Reduced Cerebral Oxygen Content in the DG and SVZ In Situ Promotes Neurogenesis in the Adult Rat Brain In Vivo. Plos ONE (2015) 10:e0140035. doi: 10.1371/journal.pone.0140035
104. Mas-Bargues C, Sanz-Ros J, Román-Domínguez A, Inglés M, Gimeno-Mallench L, El Alami M, et al. Relevance of oxygen concentration in stem cell culture for regenerative medicine. Int J Mol Sci (2019) 20:1195–222. doi: 10.3390/ijms20051195
105. Cavallucci V, Fidaleo M, Pani G. Neural Stem Cells and Nutrients: Poised Between Quiescence and Exhaustion. Trends Endocrinol Metab (2016) 27:756–69. doi: 10.1016/j.tem.2016.06.007
106. Lim DA, Cha S, Mayo MC, Chen M-H, Keles E, VandenBerg S, et al. Relationship of glioblastoma multiforme to neural stem cell regions predicts invasive and multifocal tumor phenotype. Neuro Oncol (2007) 9:424–9. doi: 10.1215/15228517-2007-023
107. Chaichana KL, McGirt MJ, Frazier J, Attenello F, Guerrero-Cazares H, Quinones-Hinojosa A. Relationship of glioblastoma multiforme to the lateral ventricles predicts survival following tumor resection. J Neurooncol (2008) 89:219–24. doi: 10.1007/s11060-008-9609-2
108. Sonoda Y, Saito R, Kanamori M, Kumabe T, Uenohara H, Tominaga T. The association of subventricular zone involvement at recurrence with survival after repeat surgery in patients with recurrent glioblastoma. Neurol Med Chir (Tokyo) (2014) 54:302–9. doi: 10.2176/nmc.oa.2013-0226
109. Adeberg S, Bostel T, König L, Welzel T, Debus J, Combs SE. A comparison of long-term survivors and short-term survivors with glioblastoma, subventricular zone involvement: a predictive factor for survival? Radiat Oncol (2014) 9:95. doi: 10.1186/1748-717X-9-95
110. Jafri NF, Clarke JL, Weinberg V, Barani IJ, Cha S. Relationship of glioblastoma multiforme to the subventricular zone is associated with survival. Neuro Oncol (2013) 15:91–6. doi: 10.1093/neuonc/nos268
111. Mistry AM, Dewan MC, White-Dzuro GA, Brinson PR, Weaver KD, Thompson RC, et al. Decreased survival in glioblastomas is specific to contact with the ventricular-subventricular zone, not subgranular zone or corpus callosum. J Neurooncol (2017) 132:341–9. doi: 10.1007/s11060-017-2374-3
112. Mistry AM, Hale AT, Chambless LB, Weaver KD, Thompson RC, Ihrie RA. Influence of glioblastoma contact with the lateral ventricle on survival: a meta-analysis. J Neurooncol (2017) 131:125–33. doi: 10.1007/s11060-016-2278-7
113. Han S, Li X, Qiu B, Jiang T, Wu A. Can lateral ventricle contact predict the ontogeny and prognosis of glioblastoma? J Neurooncol (2015) 124:45–55. doi: 10.1007/s11060-015-1818-x
114. Mistry AM, Wooten DJ, Davis LT, Mobley BC, Quaranta V, Ihrie RA. Ventricular-Subventricular Zone Contact by Glioblastoma is Not Associated with Molecular Signatures in Bulk Tumor Data. Sci Rep (2019) 9:1–12. doi: 10.1038/s41598-018-37734-w
115. Comas S, Luguera E, Molero J, Balaña C, Estival A, Castañer S, et al. Influence of glioblastoma contact with the subventricular zone on survival and recurrence patterns. Clin Transl Oncol (2020), 1–11. doi: 10.1007/s12094-020-02448-x
116. Mistry AM, Mummareddy N, CreveCoeur TS, Lillard JC, Vaughn BN, Gallant J-N, et al. Association between supratentorial pediatric high-grade gliomas involved with the subventricular zone and decreased survival: a multi-institutional retrospective study. J Neurosurg Pediatr (2020) 1–7:288–94. doi: 10.3171/2020.3.peds19593
117. Pessina F, Navarria P, Cozzi L, Ascolese AM, Simonelli M, Santoro A, et al. Maximize surgical resection beyond contrast-enhancing boundaries in newly diagnosed glioblastoma multiforme: is it useful and safe? A single institution retrospective experience. J Neurooncol (2017) 135:129–39. doi: 10.1007/s11060-017-2559-9
118. Jackson C, Choi J, Khalafallah AM, Price C, Bettegowda C, Lim M, et al. A systematic review and meta-analysis of supratotal versus gross total resection for glioblastoma. J Neurooncol (2020) 148:1–13. doi: 10.1007/s11060-020-03556-y
119. Elliott JP, Keles GE, Waite M, Temkin N, Berger MS. Ventricular entry during resection of malignant gliomas: effect on intracranial cerebrospinal fluid tumor dissemination. J Neurosurg (1994) 80:834–9. doi: 10.3171/jns.1994.80.5.0834
120. Behling F, Kaltenstadler M, Noell S, Schittenhelm J, Bender B, Eckert F, et al. The prognostic impact of ventricular opening in glioblastoma surgery: a retrospective single center analysis. World Neurosurg (2017) 106:615–24. doi: 10.1016/j.wneu.2017.07.034
121. Saito T, Muragaki Y, Maruyama T, Komori T, Nitta M, Tsuzuki S, et al. Influence of wide opening of the lateral ventricle on survival for supratentorial glioblastoma patients with radiotherapy and concomitant temozolomide-based chemotherapy. Neurosurg Rev (2019) 1:1–11. doi: 10.1093/noajnl/vdz039.087
122. Smith AW, Mehta MP, Wernicke AG. Neural stem cells, the subventricular zone and radiotherapy: implications for treating glioblastoma. J Neurooncol (2016) 128:207–16. doi: 10.1007/s11060-016-2123-z
123. Evers P, Lee PP, DeMarco J, Agazaryan N, Sayre JW, Selch M, et al. Irradiation of the potential cancer stem cell niches in the adult brain improves progression-free survival of patients with malignant glioma. BMC Cancer (2010) 10:1–7. doi: 10.1186/1471-2407-10-384
124. Gupta T, Nair V, Paul SN, Kannan S, Moiyadi A, Epari S, et al. Can irradiation of potential cancer stem-cell niche in the subventricular zone influence survival in patients with newly diagnosed glioblastoma? J Neurooncol (2012) 109:195–203. doi: 10.1007/s11060-012-0887-3
125. Lee P, Eppinga W, Lagerwaard F, Cloughesy T, Slotman B, Nghiemphu PL, et al. Evaluation of high ipsilateral subventricular zone radiation therapy dose in glioblastoma: A pooled analysis. Int J Radiat Oncol Biol Phys (Elsevier) (2013) 86:609–15. doi: 10.1016/j.ijrobp.2013.01.009
126. Chen L, Guerrero-Cazares H, Ye X, Ford E, McNutt T, Kleinberg L, et al. Increased subventricular zone radiation dose correlates with survival in glioblastoma patients after gross total resection. Int J Radiat Oncol Biol Phys (Elsevier) (2013) 86:616–22. doi: 10.1016/j.ijrobp.2013.02.014
127. Iuchi T, Hatano K, Kodama T, Sakaida T, Yokoi S, Kawasaki K, et al. Phase 2 trial of hypofractionated high-dose intensity modulated radiation therapy with concurrent and adjuvant temozolomide for newly diagnosed glioblastoma. Int J Radiat Oncol Biol Phys (2014) 88:793–800. doi: 10.1016/j.ijrobp.2013.12.011
128. Ravind RR, Prameela CG, Dinesh M. P0111 Sub-ventricular zone irradiation in glioblastoma: Can it increase survival? Eur J Cancer (2015) 51:e23. doi: 10.1016/j.ejca.2015.06.069
129. Foro P, Pera O, De Dios NR, Sanz J, Castillejo AR, Conejo IM, et al. Relationship Between Subventricular Zone Dose and Survival in Patients With Glioblastoma Multiforme Treated With Surgery Followed Radiation Chemotherapy. Int J Radiat Oncol (2015) 93:E81. doi: 10.1016/j.ijrobp.2015.07.752
130. Foro Arnalot P, Pera O, Rodriguez N, Sanz X, Reig A, Membrive I, et al. Influence of incidental radiation dose in the subventricular zone on survival in patients with glioblastoma multiforme treated with surgery, radiotherapy, and temozolomide. Clin Transl Oncol (2017) 19:1225–31. doi: 10.1007/s12094-017-1659-5
131. Slotman BJ, Eppinga WSC, de Haan PF, Lagerwaard FJ. Is Irradiation of Potential Cancer Stem Cell Niches in the Subventricular Zones Indicated in GBM? Int J Radiat Oncol (2011) 81:S184. doi: 10.1016/j.ijrobp.2011.06.328
132. Elicin O, Inac E, Uzel EK, Karacam S, Uzel OE. Relationship between survival and increased radiation dose to subventricular zone in glioblastoma is controversial. J Neurooncol (2014) 118:413–9. doi: 10.1007/s11060-014-1424-3
133. Anker CJ, Bagshaw HP, Sarkar V, Dritto M, Boucher K, Jensen RL, et al. Impact of Subventricular Zone Dose and Relationship to Glioblastoma Tumor Location on Outcomes. Int J Radiat Oncol (2015) 93:E110–1. doi: 10.1016/j.ijrobp.2015.07.829
134. Sakuramachi M, Igaki H, Nomoto A, Sekiya N, Takahashi W, Sakumi A, et al. Radiation Dose to Ipsilateral Subventricular Zone as a Prognostic Factor in Malignant Glioma Patients. Int J Radiat Oncol (2015) 93:E68. doi: 10.1016/j.ijrobp.2015.07.716
135. Murchison SC, Wiksyk B, Gossman S, Jensen B, Sayers D, Lesperance M, et al. Subventricular Zone Radiation Dose and Outcome for Glioblastoma Treated Between 2006 and 2012. Cureus (2018) 10:3618–31. doi: 10.7759/cureus.3618
136. Valiyaveettil D, Malik M, Joseph DM. Effect of radiation dose to the periventricular zone and subventricular zone on survival in anaplastic gliomas. Ecancermedicalscience (2019) 13. doi: 10.3332/ecancer.2019.956
137. Valiyaveettil D, Malik M, Syed K, Fayaz Ahmed S, Joseph DM. Prospective study to assess the survival outcomes of planned irradiation of ipsilateral subventricular and periventricular zones in glioblastoma. Ecancermedicalscience (2020) 14:1021–32. doi: 10.3332/ecancer.2020.1021
138. Khalifa J, Tensaouti F, Lusque A, Plas B, Lotterie JA, Benouaich-Amiel A, et al. Subventricular zones: New key targets for glioblastoma treatment. Radiat Oncol (2017) 12:1–11. doi: 10.1186/s13014-017-0791-2
139. Gui C, Vannorsdall TD, Kleinberg LR, Assadi R, Moore JA, Hu C, et al. A Prospective Cohort Study of Neural Progenitor Cell-Sparing Radiation Therapy Plus Temozolomide for Newly Diagnosed Patients With Glioblastoma. Neurosurgery (2020) 87:E31–40. doi: 10.1093/neuros/nyaa107
140. Hsiao KY, Yeh SA, Chang CC, Tsai PC, Wu JM, Gau JS. Cognitive Function Before and After Intensity-Modulated Radiation Therapy in Patients With Nasopharyngeal Carcinoma: A Prospective Study. Int J Radiat Oncol Biol Phys (2010) 77:722–6. doi: 10.1016/j.ijrobp.2009.06.080
141. Jalali R, Mallick I, Dutta D, Goswami S, Gupta T, Munshi A, et al. Factors Influencing Neurocognitive Outcomes in Young Patients With Benign and Low-Grade Brain Tumors Treated With Stereotactic Conformal Radiotherapy. Int J Radiat Oncol Biol Phys (2010) 77:974–9. doi: 10.1016/j.ijrobp.2009.06.025
142. Armstrong GT, Jain N, Liu W, Merchant TE, Stovall M, Srivastava DK, et al. Region-specific radiotherapy and neuropsychological outcomes in adult survivors of childhood CNS malignancies. Neuro Oncol (2010) 12:1173–86. doi: 10.1093/neuonc/noq104
143. Khatua S, Dhall G, O’Neil S, Jubran R, Villablanca JG, Marachelian A, et al. Treatment of primary CNS germinomatous germ cell tumors with chemotherapy prior to reduced dose whole ventricular and local boost irradiation. Pediatr Blood Cancer (2010) 55:n/a–a. doi: 10.1002/pbc.22468
144. Sharifzad F, Ghavami S, Verdi J, Mardpour S, Sisakht MM, Azizi Z, et al. Glioblastoma cancer stem cell biology: Potential theranostic targets Corresponding author at: Paediatric Stem Cell Transplant. Drug Resist Update (2019) 42:35–45. doi: 10.1016/j.drup.2018.03.003
145. Kast RE, Ellingson BM, Marosi C, Halatsch ME. Glioblastoma treatment using perphenazine to block the subventricular zone’s tumor trophic functions. J Neurooncol (2014) 116:207–12. doi: 10.1007/s11060-013-1308-y
146. O’Keeffe GC, Tyers P, Aarsland D, Dalley JW, Barker RA, Caldwellf MA. Dopamine-induced proliferation of adult neural precursor cells in the mammalian subventricular zone is mediated through EGF. Proc Natl Acad Sci U S A (2009) 106:8754–9. doi: 10.1073/pnas.0803955106
147. Bardella C, Al-Shammari AR, Soares L, Tomlinson I, O’Neill E, Szele FG. The role of inflammation in subventricular zone cancer. Prog Neurobiol (2018) 170:37–52. doi: 10.1016/j.pneurobio.2018.04.007
148. Roesch S, Rapp C, Dettling S, Herold-Mende C. When immune cells turn bad—tumor-associated microglia/macrophages in glioma. Int J Mol Sci (2018) 19:436–56. doi: 10.3390/ijms19020436
149. Van Der Sanden B, Appaix F, Berger F, Selek L, Issartel JP, Wion D. Translation of the ecological trap concept to glioma therapy: The cancer cell trap concept. Future Oncol (2013) 9:817–24. doi: 10.2217/fon.13.30
150. Azarin SM, Yi J, Gower RM, Aguado BA, Sullivan ME, Goodman AG, et al. In vivo capture and label-free detection of early metastatic cells. Nat Commun (2015) 6:1–9. doi: 10.1038/ncomms9094
Keywords: glioblastoma, recurrence, subventricular zone, glioma stem cell, cancer stem cell, diffuse intrinsic pontine glioma, high grade glioma, diffuse midline glioma
Citation: Lombard A, Digregorio M, Delcamp C, Rogister B, Piette C and Coppieters N (2021) The Subventricular Zone, a Hideout for Adult and Pediatric High-Grade Glioma Stem Cells. Front. Oncol. 10:614930. doi: 10.3389/fonc.2020.614930
Received: 07 October 2020; Accepted: 09 December 2020;
Published: 26 January 2021.
Edited by:
Natanael Zarco, Mayo Clinic, United StatesReviewed by:
Constantinos G. Hadjipanayis, Mount Sinai Health System, United StatesCopyright © 2021 Lombard, Digregorio, Delcamp, Rogister, Piette and Coppieters. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Bernard Rogister, QmVybmFyZC5Sb2dpc3RlckB1bGllZ2UuYmU=
†These authors have contributed equally to this work
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.