 Virginie Vlaeminck-Guillem
Virginie Vlaeminck-Guillem
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Oncol. , 13 June 2018
Sec. Genitourinary Oncology
Volume 8 - 2018 | https://doi.org/10.3389/fonc.2018.00222
This article is part of the Research Topic Updates, Emerging Methodologies, and Tools for Prostate Cancer Precision Medicine View all 7 articles
Extracellular vesicles (EVs), especially exosomes, are now well recognized as major ways by which cancer cells interact with each other and stromal cells. The meaningful messages transmitted by the EVs are carried by all components of the EVs, i.e., the membrane lipids and the cargo (DNAs, RNAs, microRNAs, long non-coding RNAs, proteins). They are clearly part of the armed arsenal by which cancer cells obtain and share more and more advantages to grow and conquer new spaces. Identification of these messages offers a significant opportunity to better understand how a cancer occurs and then develops both locally and distantly. But it also provides a powerful means by which cancer progression can be detected and monitored. In the last few years, significant research efforts have been made to precisely identify how the EV trafficking is modified in cancer cells as compared to normal cells and how this trafficking is altered during cancer progression. Prostate cancer has not escaped this trend. The aim of this review is to describe the results obtained when assessing the meaningful content of prostate cancer- and stromal-derived EVs in terms of a better comprehension of the cellular and molecular mechanisms underlying prostate cancer occurrence and development. This review also deals with the use of EVs as powerful tools to diagnose non-indolent prostate cancer as early as possible and to accurately define, in a personalized approach, its present and potential aggressiveness, its response to treatment (androgen deprivation, chemotherapy, radiation, surgery), and the overall patients’ prognosis.
Cancer is largely thought to occur as a result of one or several genetic events intrinsically appearing in a cell and providing it with a survival and/or growing advantage. The concept that a malignant tumor only occurs and develops by itself has, however, been challenged for many years. It is now well accepted that environmental factors can be key inducers of both cancer appearance and development (1, 2). By a mirror effect, cancer cells are also able to modify the behavior of the cells that constitute their microenvironment (i.e., the stroma) (3). This continuous interplay between cancer and stromal cells requires specific means of communication including direct cell–cell contacts and secreted factors (growth factors, peptides, etc.) that act via paracrine pathways (4). Extracellular vesicles (EVs) more recently proved to be another powerful means used by cells to communicate with each other (3, 5).
The field of EVs is extremely fast-moving (6, 7) (1) because of a clear fashion trend, (2) because of the huge hope that the reliable biomarkers we still need for several pathological conditions including cancers will finally emerge from their exploration, and (3) because it is a relative recent concept. The term “extracellular vesicle” was introduced as a generic term to describe any type of membrane-enclosed particles released by any type of cell (including microorganisms) in the extracellular space (8, 9). This definition covers in fact several terms, sometimes used indifferently in publications and including notably exosomes, microvesicles, microparticles, apoptotic particles, apoptotic bodies, oncosomes, etc. and, in the specific field of the prostate gland, prostasomes (10). Exosomes were first described in the 1980s as a means to recycle the transferrin receptor from the plasma membrane through the endocytic compartments (11–14). They have now become the most studied EV portion because of their strong implication in both physiological and pathological processes including cancers. Thought, at their discovery, as artifacts (15) or as a trash bin for unnecessary and redundant proteins (16), they eventually exhibit a high functional ability: secreted by all kinds of cells, they are able to carry and transfer their proteic, lipidic, and nucleotidic content (the “cargo”) to target cells (5, 8, 9).
In this review, we will focus on the role of exosomes in prostate cancer, but also of other EVs because of the persistent difficulty to distinguish between the different types of EVs (17). This difficulty relies on the criteria used to characterize the diverse EV subpopulations. They can be based on size, density, subcellular origin (in other words biogenesis), function, content (the cargo), membrane markers, etc. (17). And these criteria are deeply influenced by both the orientation of the research team and the methods used to detect and/or characterize the isolated EVs (18, 19). Fundamental research will focus on the biological functions of the EVs: whether the EVs bud from the endosomal compartment (endosomes) or the plasma membrane (ectosomes) is consequently extremely important. This in fact goes back to the question of the specific content of the EVs, which is closely related to the EV biogenesis as will be detailed later. Transfer research, which aims to identify biomarkers, will focus on the tissue or biological fluid where the EVs have to be isolated (for example, prostasomes are only found in prostatic fluid and seminal plasma) and the methods to detect and/or quantify the EVs are a major concern for the researchers. In this regard, a definition of exosomes based on size will be preferentially used when isolating EVs by centrifugation-based protocols, while a definition based on the membrane markers will be put forth when using immunodetection methods. Of course, several motivations can animate the researchers at the same time and several definition criteria and methods are usually used simultaneously or sequentially for a specific research. Furthermore, it is not always of major importance to ascertain which types of EVs have been detected or used at the bench if the goal is to develop a reproducible method that allow reliable detection of EV-derived biomarkers or to demonstrate that a type of cells influence the behavior of another one by the means of EVs Because of these general considerations, it should always kept in mind that despite many efforts there is no real consensual nomenclature of EVs widely used in routine research and that comparisons between the numerous publications dealing with EVs is a huge challenge that should take into account an attentive analysis of the “Material and Methods” part of the papers.
We will first attempt to provide the readers some keys to understand EV classifications according size, biogenesis, and function. We will next explore how studies on EVs shed light on prostate cancer occurrence and progression and at last their potential value as diagnostic, prognostic, or theranostic biomarkers of prostate cancer.
Exosomes consist of a lipid bilayer membrane trapping a small amount of cytosol that do not contain any organelles (8, 9, 20). Exosomes are classically considered as 30–100 nm EVs originating from intracellular budding from multivesicular bodies (MVBs) (9). They are, therefore, part of the endosomal compartment and strongly differ from the two other main classes of EVs: microvesicles (100–1,000 nm, ectosomal vesicles produced by direct budding from the plasma membrane) and apoptotic bodies (0.5–5 mm, vesicles obtained by cell lysis during the late stages of apoptosis) (21, 22) (Figure 1).
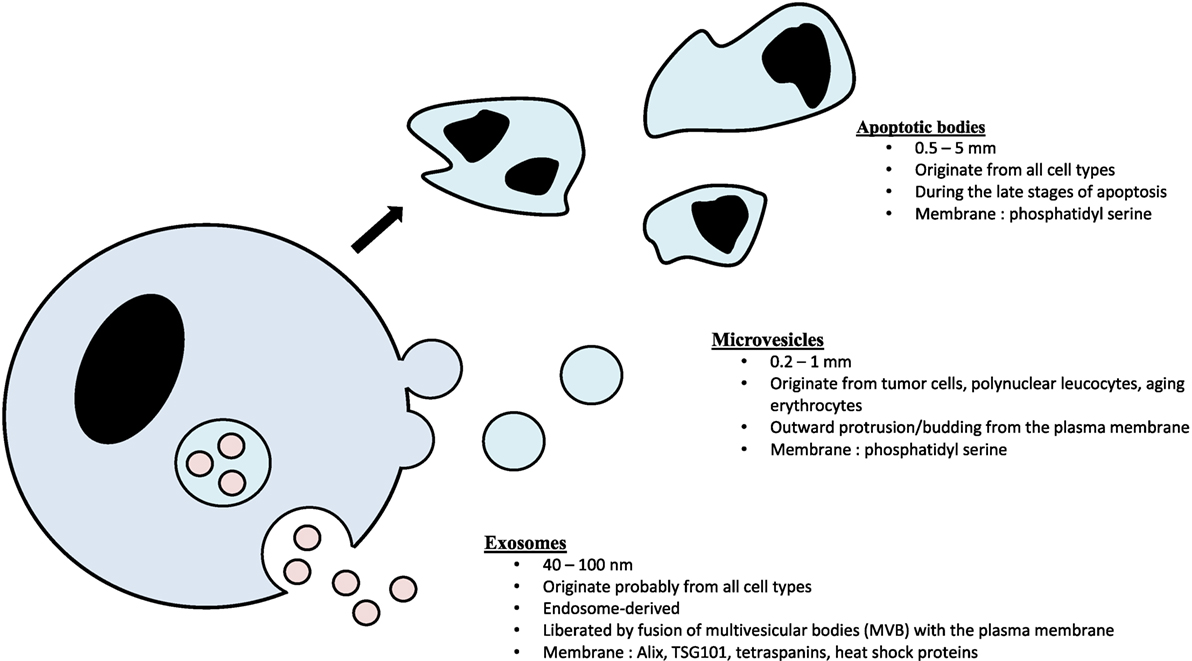
Figure 1. The three main classes of extracellular vesicles.
The biogenesis of exosomes begins within the endocytic pathway with the formation of late endosomes, the MVBs containing multiple intraluminal bilayered organelles (8, 9). Most of the MVBs fuse to lysosomes and are, therefore, degraded. Some of the MVBs fuse to plasma membrane and release their vesicular content in the extracellular space. The mechanisms by which MVBs are directed toward degradation or fusion to plasma membrane are not well understood. The formation of intraluminal organelles within MVBs has been mostly related to a specific complex of about 30 proteins called endosomal sorting complex required for transport (ESCRT) (8, 9). Specific subpopulations of ESCRT members are responsible for the sequestration of ubiquitinated transmembrane proteins in the endosomal membrane, the membrane deformation into buds and the vesicle scission (23). Accessory proteins, not included in but associated with the ESCRT, are also involved, including ALIX (8). Several studies helped in determining how exosome content is selected and in demonstrating that it is a deliberate, tightly tuned process (9). The membrane constituents of the early endosomes are key elements that govern exosomal content. For example, tetraspanins are proteins of the endosomal membrane able to determine which proteins will be loaded in the intraluminal organelles of the MVBs (9). The lipid composition of the membrane is another major indicator of exosomal content. Likewise, exosome release from parental cells is also governed by specific factors such as the presence of Rab-GTPases, the specific organization of the cytoskeleton and/or the intracellular Ca2+ levels (8).
Exosomes are internalized by target cells through different mechanisms including receptor-mediated endocytosis, pinocytosis (invagination of the plasma membrane leading to the formation of a vesicle that contains a more or less high amount of extracellular space), phagocytosis (similar to the pinocytosis but a direct contact is necessary via a receptor), and direct fusion with the plasma membrane (8, 9). Like exosome biogenesis and release by parental cells, exosome capture, and internalization by target cells is highly regulated and depends on the local conditions (e.g., microenvironmental pH) (24) and on both the parental and target cell types (9). For example, despite quite similar cargo contents, exosomes derived from the PC3 prostate cancer cell line are taken up by normal epithelial cells at a higher level than LNCaP-derived exosomes (25).
Exosomes contain lipids (essentially in their membranes), proteins, and RNAs (9), or even genomic double-stranded DNA (26). Omics studies in several cell types (including benign and malignant cells) provided exhaustive data about these components, in prostate cancer as an example of all others (25, 27–38) (Table 1). As a consequence of their endosomal origin, exosomes do not exhibit proteins from the nucleus, the Golgi apparatus, the endoplasmic reticulum or the mitochondria. They are rather enriched in proteins from the cytoplasm (including cytoskeletal components such as actin and tubulin), the membrane of the endosomes and the plasma membrane (39). Endosome-associated proteins are Rab-GTPases, SNAREs, annexins, flotillin, and those involved in MVB biogenesis (e.g., ALIX, Tsg101), in exosome sorting (e.g., tetraspanins) or in lipid rafting (e.g., glycosylphosphatidyl inositol-anchored proteins) (9).
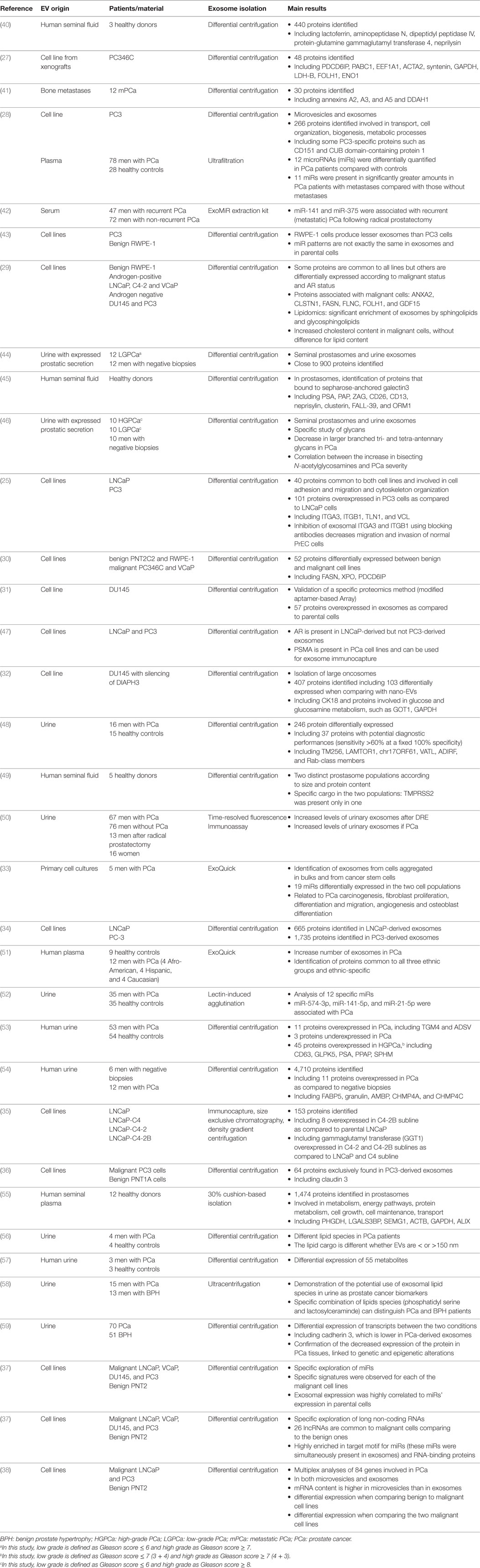
Table 1. High-throughput studies exploring prostate extracellular vesicles (EVs).
As compared to plasma membranes, exosomal membranes are enriched in cholesterol, sphingomyelin, and hexosylceramides, while being bald in phosphatidylcholine and phosphatidylethanolamine. This accounts for the exosome formation within MVBs (60).
A turning point has in fact been marked with the demonstration that exosomes contain mRNAs and microRNAs (miRs) that can be eventually translated into functional proteins upon capture by target cells (5, 61). In other words, exosomes can be considered as intercellular messengers able to transmit biologically active messages from parental cells to target cells (5, 62). The biological behavior of the target cells can, therefore, be distally modified to provide specific advantages in terms of favorable microenvironment (e.g., production of growth factors, neoangiogenesis, remodeling of the extracellular matrix, etc.) (5). Of note, when transfer of oncogenic material is able to induce transformation of the recipient cells (a typical demonstration of a biological significance!), exosomes are called oncosomes (63). Recent studies have also demonstrated that exosomes also contain other varieties of RNA including small or long non-coding RNAs, structural RNAs, tRNAs, and small interfering RNAs, all species with functional interactive abilities despite the absence of protein coding ability (8, 62).
Normal prostate cells have been shown to produce EVs for a long time. It is worthy to note that the first prostate-originating EVs, called prostasomes, were reported in the late 70s as extracellular nanosized membrane-surrounded particles found in seminal plasma (64–66). Whether prostasomes are really exosomes remains matter of debate (67, 68). There are similarities such as stockade in the MVBs, release after MVB fusion with the plasma membrane (69), presence of exosomal markers such as CD9 and CD63 (70), and exchange of biologically significant information (71). Nevertheless, prostasomes are usually bigger than exosomes (50–500 nm, the mean diameter being 150 nm) (72), they can have a multilayer membrane (40, 73), their lipidic content is slightly different (higher concentration of cholesterol and sphingomyelin, and higher cholesterol/phospholipid ratio) (70, 72, 74), and they can contain chromosomal DNA (seldom reported in exosomes) (69, 75). As a normal component of the seminal plasma, prostasomes have been shown to interact with spermatozoa and are strongly involved in male fertility and reproduction (76, 77). They are indeed implicated in spermatozoa motility, capacitation, acrosome reaction, modulation of immunological attacks by the female’s immune cells, antioxidant, and antibacterial capacities (69, 71).
Even if most of the studies exploring EVs in prostate cancer focused on “true” prostate exosomes, it should be kept in mind that prostasomes are also produced by prostate cancer cells (78–81), including metastatic ones (79, 80), using the same processes than normal cells (80). Although demonstration is still lacking, their roles in prostate carcinogenesis could be reminiscent of those observed in human reproduction including escape from the immune system or induction of migration (82). Of note, one study suggested that production of prostasomes in prostate cancer cells is inversely correlated with the Gleason score (83). Other studies demonstrated differential expression in prostasomes derived from normal or cancer prostate cells (44–46, 49, 55).
Prostate cancer is the tissue in which a new class of EVs has been described in the late 2000s (84, 85). They have been called large oncosomes because of their size (1–10 µm) and their primitive description from cancer cells. Large oncosomes production results from the shedding of non-apoptotic plasma membrane blebs and can be induced by overexpression of oncoproteins (84) or silencing of the cytoskeletal regulator DIAPH3 (85, 86).
Like exosomes, prostate large oncosomes carry abundant bioactive molecules, including signaling factors implicated in cell growth, cell metabolism, cell motility or RNA processing (84), but their cargo seems specific, at least different from that of the nanosized prostate EVs (32). Proteins differentially expressed are involved in cancer-associated metabolic processes such as glucose, glutamine, and amino acid metabolism. Of interest, cancer cells exposed to large oncosomes but not to exosomes disclosed altered glutamine metabolism, suggesting that large oncosomes exert specific functional abilities (32). This specific content may result from their specific release by cells transitioning to an amoeboid phenotype, a phenotype used by some invasive cancer cells to migrate in the peritumoral space. Amoeboid cells affect an elliptical blebbing morphology, which would favor the scission of large oncosomes. As such, large oncosomes could represent markers of invasive, migrating cancer cells (10).
Although distinction with other nanosized EVs is usually not fully achieved, prostate exosomes constitute by far the most studied portion of prostate EVs (10, 67). Large-scale profiling experiments have been widely performed, using various origins such as cell lines, plasma, serum, urine, and different methods depending on the part of the content to be targeted (proteins, lipids, metabolites, RNAs, etc.) (Table 1). Comparisons have been made between different cell lines (for example androgen-sensitive or androgen-independent; aggressive or less aggressive, etc.) or between different clinical conditions (benign prostate hypertrophy, localized cancer, metastatic cancer, castration-resistant cancer, etc.). Quite all studies demonstrate that such specific culture or clinical situations can be related to specific expression patterns, fueling the hope to find valuable biomarkers (see below). Along with other tissues, the resulting datasets have generally been introduced in large databases such as Exocarta, Vesiclepedia, and EVpedia (21, 87, 88).
In prostate cancer as in all other tissues (benign or malignant), exosomes (or at least EVs) have been found to be produced by all studied cell types. In other words, they are secreted not only by cancer cells but also by all the cells constituting its microenvironment, i.e., cells from the adjacent normal and malignant epithelium, as well as cells from the tumor stroma, such as fibroblasts, immune cells, and endothelial cells (8). All these cell types can interplay with each other by producing EVs and we here successively present the effects of cancer cell-derived exosomes on stromal cells, the effects of stromal cell-derived exosomes on cancer cells, and then the effects of cancer cell-derived exosomes on adjacent epithelial benign or malignant cells.
For cancer cells, the aim of targeting stromal cells by biologically significant exosomes is clearly to modify the microenvironment to favor local and distant extension (2). One of the major changes during cancer and development is the transition from a simple stroma (with only supportive abilities) to a reactive stroma able to favor cancer cell proliferation and metastatic spread (89). This transition is accompanied by changes in all the stromal components: nature and proportion of immune cells, proteic components of the extracellular matrix, etc. All these modifications have been correlated to prostate cancer differentiation (Gleason score) or survival (90). Among these modifications, a fundamental one is the occurrence of CAFs (91), producing several growth factors (TGF-beta, SDF-1, CXCL14, HIF-alpha, etc.) that target cancer cells (92). Conversely, epithelial cells produce several signals able to stimulate CAF occurrence (TGF-beta, PDGF, FGF2, hedgehog, etc.) (92). Other communication means include exosomes, as demonstrated in several cancers.
In this regard, exosomes derived from the prostate cancer cell lines DU145 and PC3 (and to a less extent the LNCaP) contain TGF-beta that can induce transformation of fibroblasts to myofibroblasts (a feature of CAFs) via the activation of the TGF-beta/SMAD signaling (93, 94). Indeed, while LNCaP cells produce exosomes with low levels of TGF-beta, other cell types such as PC3 and DU-145 produce exosomes with high TGF-beta levels. Expression of the transmembrane proteoglycan betaglycan at the exosome surface appears to be necessary for TGF-beta loading in the exosomes (93). This TGF-beta is able to trigger the SMAD3-dependant intracellular signaling pathway of target cells (93). TGF-beta could thus induce both the fibroblastic expression of alpha-smooth actin (SMA; at the protein level)—a key marker of myofibroblastic differentiation—and its filamentous structures (93, 94). Fibronectin containing the alternatively spliced EDA exon, another myofibroblast biomarker, also proved to be influenced by exosomal TGF-beta (94). In addition, exosome-derived TGF-beta triggers an alteration of the immediate peri-fibroblastic microenvironment that is also specific for myoblast differentiation: the formation of a pericellular coat largely composed of hyaluronic acid (93). Other responses classically obtained with soluble TGF-beta were also observed (such as autocrine production of TGF-beta by fibroblasts), but it was suggested that exosomal TGF-beta could exert specific intracellular effects such as increased FGF2 expression (93). Similar results were obtained when normal prostate stromal cells (PrSC) were cocultured with prostate cancer cells (92). Of interest these results obtained from fibroblast cultures could be reproduced using normal and activated stromas obtained from fresh tissues (patients undergoing prostatectomies) (94): normal stromal cells do not express alpha-SMA (but gained this ability upon exposition to exosomal TGF-beta), while cells isolated from reactive stroma exhibit heterogeneous but clear alpha-SMA expression that could not by reinforced by stimulation. To ascertain the importance of exosomal delivery over soluble TGF-beta secretion, the use of Rab27a, a regulator of exosome production, allowed clear demonstration that cells selectively deficient in exosome production could not trigger similar responses (94).
Although the underlying mechanisms are not fully understood, hypoxia in prostate cancers has been associated with an aggressive phenotype and a poor prognosis (95). A recent paper showed that exosomes extracted from LNCaP and PC3 cells cultured in hypoxic conditions induced a CAF phenotype in normal human PrSC (96). Expression of the alpha-smooth muscle actin was indeed found to be enhanced (96). It is likely that the exosomal content is modified under hypoxic conditions since hypoxic prostate cancer exosomes are loaded with a significantly higher amount of triglycerides (97).
It has been well demonstrated that CAFs can be obtained from several cell lineages and not only from normal stromal fibroblasts. For example, mesenchymal stem cells (MSCs) are pluripotent cells, able to differentiate into several connective tissues, including myocytes, neurons, osteoblasts, chondrocytes, adipocytes, and fibroblasts (98). The roles of MSCs include migration into inflammatory or injured tissues and their repair. Intratumoral MSC migration is also considered, notably in prostate cancer (99) with tumor-promoting effects similar to those of cancer-associated stroma. A recent experiment suggests that cancer cell-derived exosomes are able to induce differentiation of MSCs into myofibroblasts (100). Bone marrow MSCs exposed to DU145-derived exosomes despite pro-adipogenic culture conditions underwent differentiation into myofibroblasts with expression of alpha-smooth muscle actin and secretion of both growth factors (VEGF-A, HGF) and extracellular matrix regulators (MMP-1, MMP-3, and MMP-13). These properties allow promotion of both angiogenesis and tumor progression (proliferation, migration, and invasion). Again, this reprogramming effect of cancer-derived exosomes was dependent on the presence of exosomal TGF-beta, while soluble TGF-beta did not produce similar effects (100).
It is worthy to note that the way in which fibroblasts engage upon EV stimulation may not be unequivocal and could depend on the nature of the EVs Pioneer study suggested that large oncosomes extracted from the tumorigenic RWPE-2 cell line can induce CAF migration (101). The same team later showed that these oncosomes harbor sustained AKT1 kinase activity that induced prostate fibroblast reprogramming (102). The reprogramming was molecularly defined by the increased expression of alphaSMA, Il-6, and matrix-metalloprotease 9, without any change in TGF-beta levels, suggesting a provascularization phenotype rather than an epithelial-to-mesenchymal transition (EMT) phenotype (102). In fact reprogrammed fibroblasts were effectively able to stimulate endothelial tube formation in vitro and tumor growth in mice (102). In these experiments, activation of stromal MYC was found to be the critical event for fibroblast reprogramming.
Other changes induced in the stroma by cancer-derived MVs include alterations of the extracellular matrix (96, 103) and promotion of angiogenesis (94, 103). Prostate cancer-derived microvesicles indeed proved to express matrix metalloproteinases (MMP) such as MMP9 (PC3 and LNCaP cells) and MMP14 (PC3 cells only), possibly through stimulation of ERK1/2 phosphorylation (103). MVs released by prostate cancer tissues seem, therefore, able to degrade collagen IV and the basal membrane, suggesting a role in promoting both cancer cell protection from apoptosis (specially anoikis, i.e., the specific apoptotic process that results from the isolation of a migrating cell devoid of cell–cell contacts) and mobility of both fibroblasts and cancer cells (103). Of note, prostasomes extracted from PC3 and LNCaP (as compared to prostasomes extracted from healthy ejaculates) could degrade the extracellular matrix through modifications of their surface proteolytic enzymes: decrease in dipeptidyl peptidase IV and increase in UPa (urokinase plasminogen activator) and cathepsin B were observed (104). Since MMPs are also involved in angiogenesis (96, 103), an effect of MV release by prostate cancer cells on neoangiogenesis is also suggested, and consistent with the proangiogenic (although indirect) effect of cancer cell-derived exosomes observed by Webber et al. (94): they found that myofibroblasts activated by cancer-derived exosomal TGF-beta can modulate the behavior of endothelial cells, leading to more branching and elaborated networks of vessel-like structures connected to neighboring vessels (94). Similar effects on cultured endothelial cell proliferation and migration were observed with MVs derived from DU-145 (105). Likewise, stromal cells cocultured with prostate cancer cells also exhibited an increase in VEGF secretion, suggesting a potential proangiogenic effect (92). By contrast, prostasomes obtained from postvasectomy human semen were unable to influence endothelial cell survival and rather inhibited the development of capillary-like networks (106, 107). This discrepancy between prostasomes and other prostate MVs may result from the prostasome specific lipid content, which proved to be enriched in sphingomyelin and cholesterol (see above), even if sphingomyelin was suggested by others as a key mediator of the proangiogenic effects of DU145-derived MVs (105).
Immune cells present in the tumor stroma are involved in the host mechanisms of defense against cancer occurrence and development (9). Lundholm et al. demonstrated that prostate cancer-derived exosomes could help cancer cells to evade the immune responses (108). Using flow cytometry experiments, the authors found that ULBP1-2 and MICA/B, ligands for NKG2D [a membrane receptor expressed at the membrane of natural killer cells (NK) and CD8+ T cells that activate target cell death] are expressed at the surface of cancer-derived exosomes. Bound to the NKG2D receptors present on the NK and CD8+ T cells, these exosomal ligands are able to induce downregulation of NKG2D receptors and consequently to impair the cytotoxic function of immune cells (108). Of interest, these in vitro results are consistent with the in vivo observations that patients with castration-resistant prostate cancer exhibit a significantly decreased surface expression of NKG2D on circulating NK and CD8+ T cells and that cancer-derived exosomes extracted from plasma or serum of these patients induced NKG2D downregulation in cultured lymphocytes (108). A recent study suggested that prostate cancer-derived exosomes may affect the action of CD8+ T cells, through altering the ability of dendritic cells to interact with them (109). The fact that prostate EVs can protect cells from lymphocyte-mediated lysis was previously observed for prostasomes (110). At a molecular level, it was demonstrated that prostasomes express CD59 (also named membrane inhibitor of reactive lysis), an important regulator of the complement cytotoxicity (111). The transfer of exosomal CD59 from prostasomes to red cells lacking CD59 (rabbit erythrocytes and human erythrocytes from patients with paroxysmal nocturnal hemoglobinuria) indeed proved to induce protection against complement-mediated hemolysis (111). Prostasomes produced by PC3 and DU-145 cells proved to contain high CD59 levels as compared to those derived from LNCaP, normal seminal fluid or normal prostate tissues (112). All these prostasomes could, however, be transferred to red cells and decrease complement-mediated hemolysis (112). Prostate large oncosomes are also able to influence immune response, by suppressing proliferation of human macrophages and peripheral blood mononuclear cells (86). This immune suppressive effect could be related to the reduced expression of Akt1 in these tumor-infiltrating immune cells, in response to the miR-125a present in the EVs produced by cancer cells (86).
Fibroblasts are not the only stromal cell type that undergoes transformation upon exosomal stimulation by cancer cells. Adipose-tissue derived stem cells (ASCs), a subtype of MSCs, can be isolated from a perivascular niche of fat tissue and have the ability to differentiate into multiple cell lineages (113). In an intriguing study, ASCs obtained from the fat of the Retzius space of patients undergoing radical prostatectomy for prostate cancer and selected for their high migratory ability were exposed to the culture media of benign (RWPE-1) and malignant cell lines (PC3 and C4-2B). Transplanted in the flanks of athymic nude mice, ASCs exposed to malignant cell culture media developed subcutaneous nodules with neoplastic histopathological features reminiscent of prostate cancer. Expression of epithelial and prostate cancer-specific biomarkers (CK8, CK5/18, and AMACR) fueled the hypothesis that ASCs can form prostate-like neoplastic lesions upon specific stimulation. Further in vitro experiments suggested that prostate cancer cell-derived exosomes are responsible for this prostate tumorigenic mimicry and that the ASC reprogramming is linked to the presence of oncogenic factors within exosomes such as H-ras, K-ras, other members of the Ras superfamily of GTPases and some miRs (miR-125b, miR-130b, miR155) (113). These experiments suggest that MSCs, such as ASCs, can transform into tumor cells, an ability that has in fact not been fully elucidated (114).
Although the effects of exosomes are usually measured in the close microenvironment of cancer cells, there is now strong evidence that they can be released in several body fluids including the blood, as observed for prostate cancer (115). As such, whether they can participate to the metastatic spread remains to be precisely determined but several recent papers favor this hypothesis. As prostate cancer progresses, osteoblastic bone metastases are frequently observed and many experiments suggest that the bone tropism of prostate cancer cells results from a vicious cycle established between cancer and bone cells (osteoblasts, osteoclasts) (116, 117). While prostate cancer cells supply osteoblastic and osteoclastic factors to bone cells, these cells, thereby activated, in turn supply prostate cancer cells with growth factors (116, 117).
Some recent papers suggest that cancer-derived EVs could both favor osteoblast differentiation, impair osteoclast differentiation and favor prostate mimicry in bone cells. For example, exposition of murine pro-osteoblastic MC3T3-E1 cells to the MVs extracted from the prostate cancer cell lines PC3 and DU-145 proved to facilitate osteoblast differentiation (118). Of note, ETS1, a transcription factor known to be involved in many metastatic processes, can be detected in the MVs of PC3 and DU-145 cells, but not in those extracted from LNCaP cells that failed to facilitate osteoblast differentiation (118). Similarly, when exposed to exosomes extracted from the murine prostate cancer cell line TRAMP-C1, monocytic osteoclast precursors (monocyte cell line RAW264-7 and bone marrow-derived osteoclast precursors) failed to fusion and differentiate into mature multinucleated osteoclasts despite induction by the osteoclastogenic factor RANKL (119). Consistently osteoclastic markers (DC-STAMP, TRAP, cathepsin K, MMP9) were significantly less expressed upon exosome exposition. Another study suggested the same mechanism: high amounts of miR-141-3-p present in the MDA-PCa-2b cells (PCa bone metastasis cell line) can be transferred to osteoblasts where they directly target DLC1, a protein involved in the regulation of Rho GTPases (120). The p38MAPK pathway is thereby influenced, inducing increased proliferation, increased calcium deposition and increased expression of markers of osteoblastic differentiation (120). In vivo experiments showed that intravenously injected fluorescent-tagged MDA-PCa-2b-derived exosomes were preferentially captured by bone, where they induced osteoblastic activity (120). Similarly, MDA-PCa-2b cells transfected with miR-141-3-p and injected into mice developed bone metastases that reduced the animals’ survival (120).
Cell plasticity can be defined as the ability of a cell to undergo reversible phenotypic changes during tissue invasion. Epithelial cells are particularly able to adopt plasticity as a means to favor distant progression. Prostate cancer cells for example can transit to a mesenchyme-like phenotype (EMT) like many cancer cells. More specifically, evolution toward neuroendocrine differentiation (NED) (121, 122) and osteomimicry (123, 124) was reported in prostate cancer cells, respectively, as a response to androgen deprivation or radiotherapy, and as a means to favor bone metastasis. Prostate cancer cells expressing bone markers (osteomimicry) better adapt to the bone microenvironment. Conversely, bone cells could also produce prostate-specific biomarkers that could help prostate cancer cell integration into the bone tissue. Renzulli et al. demonstrated that human bone marrow freshly harvested from healthy volunteers can express prostate-specific markers (TMPRSS2, PSA, PSCA, ERG, ETV1, KLK3) when cocultured with 1-cm2 wide pieces of human prostate cancers (125). MVs isolated from these prostate cancer specimens provoked similar effects.
Studies dealing with the EV-mediated interplays between prostate cancer cells and stromal cells essentially addressed the functional properties of cancer-derived exosomes. Analyses of stroma-derived EVs are dramatically less frequent but provide significant results.
One of the studies showed that stromal fibroblasts produce EVs measuring 1–30 µm in size (126). These EVs contain miR-409 which has been previously shown to promote tumorigenesis, EMT, stemness, and bone metastasis of human prostate cancer (127). Stromal-derived EVs can be efficiently internalized by the prostate cancer cell lines ARCaPE and C4-2B, in which they induce specific changes including decreased expression of miR-409 target genes (Stag2, RSU1, PHC3), increased proliferation, morphological EMT, biochemical EMT (decreased expression of E-cadherin and increased vimentin expression), and tumorigenesis in athymic nude mice (126). Subcutaneous coinjection of ARCaPE cells together with miR-409-expressing stromal fibroblasts targeted development of bigger tumors, with higher proliferative rate, than injection of the sole ARCaPE cells. Expression of miR-409 by ARCaPE cells (naturally devoid of it) in the tumors obtained after subcutaneous coinjection was checked, suggesting that promotion of tumor development by miR-409 can effectively be obtained through exosomal transfer of the miR from stromal cells to cancer cells. Whether stromal exosomes-mediated stimulation can also favor prostate cancer occurrence remains to be determined but the authors also demonstrated that orthotopic implantation of miR-409-expressing stromal fibroblasts in mouse prostate induced prostatic luminal expansion, epithelial atrophy, increased inflammation, and both epithelial and stromal hyperplasia (126).
Another study used menstrual stem cells (MenSCs), that are MSCs isolated from menstrual blood (128). These cells behave as multipotent cells with finely tuned angiogenic properties necessary for the hormonal cycle of the endometrium. The MenSCs were shown to produce exosomes that can be internalized by PC3 cells. This incorporation targeted specific changes in cultured PC3 cells including decreased expression of VEGF (measured at both RNA and protein levels and in the culture media) and decreased production of reactive oxygen species (ROS). The authors further demonstrated that MenSC-derived exosomes suppress the secretion of proangiogenic factors (such as HIF1-alpha) by PC3 cells in a ROS-dependent manner, leading to decreased hemoglobin content and vascular density within the prostate tumors and thereby a reduction of tumor growth (128).
The last study explored EVs derived from pre-osteoblasts (considering as non-mineralizing since they are enable to deposit calcium) and mature osteoblasts (mineralizing and obtained from pre-osteoblasts upon dexamethasone stimulation) (129). The two EV populations disclosed differential protein expression depending on mineralization and could be internalized by PC3 cells. In the prostate cancer cells, RNA profiling analyses suggested that EVs internalization significantly induced transcriptional changes specific of the mineralization status. Although different in their nature, both transcriptional programs induced by (pre)osteoblast-derived EVs converged on pathways involved in cell survival and growth, a result consistent with the observed increase in PC3 cell growth under stimulation by the two EV populations (129).
Among cells that constitute the immediate microenvironment of a cancer cell are the neighboring cancer cells, on which exosomes can act as paracrine factors. If a specific intrinsic, genetic, or epigenetic, event occurs in a cancer cell and provide it a survival or growth advantage, it will be “vertically” transmitted to daughter cells. Exosomes are a means to transmit this advantage to “sister” cells, as a kind of “horizontal” transfer (130).
This phenomenon has been described for several cancers including the prostate cancer. Halin Bergstrom et al. (131) extracted EVs from the rat Dunning G (low-growing, androgen-sensitive, non-metastatic) and MML (rapidly growing, androgen-insensitive, metastatic) prostate cancer cell lines and injected them into the ventral prostate of naïve rats. Dunning G cells were then injected 72 h later at the same place. Tumors developed that were bigger when EVs were previously injected in rat prostates, in relation with a higher macrophage infiltration but also with an increased proliferation of rat normal epithelial cells (131). In other studies, normal epithelial prostate cell lines RWPE-1 and PNT-2 and prostate cancer cell lines LNCaP and DU-145 were exposed to EVs derived from the supernatants of primary cultures of PCa and BPH tissues as well as EVs extracted from plasma of PCa patients and healthy controls (132) or to exosomes extracted from LNCaP and DU-145 cells (133). Significant changes were observed in treated cells such as decreased apoptosis, increased migration and proliferation, increased secretion of interleukin-8 [a proinflammatory chemokine associated with the promotion and progression of several cancers (134)], alterations in gene expression suggestive of EMT (132, 133). Of note, observed effects differed according to both parental and target cells: LNCaP-derived exosomes had lower effect than DU-145-derived exosomes, while apoptosis was only reduced in LNCaP and proliferation was only increased in LNCaP and DU-145 cells and not RWPE-1 cells (133).
Integrins are membrane proteins necessary for cell-to-cell interactions and cell–cell junctions. Epithelial cells express integrins, whether they are malignant or not and prostate cancer cells do not escape this rule. Integrins can be found in the exosomes of prostate cancer cell lines. It has been demonstrated that integrins αυβ6 and αυβ3 are present in exosomes extracted from PC3 cells (and αυβ3 from CwR22 cells) (135, 136). They can be internalized by other cancer cells (DU145, C4-2B) that do not express these integrins, in which the proteins are then detected, without any change in their mRNA counterparts. A direct horizontal transfer of an immediately active biological molecule is, therefore, suggested that confers to recipient cells an increased ability to adhere and migrate (135, 136). Of interest, exosome-mediated transfer of αυβ3 could also be observed into non-malignant BPH-1 prostate epithelial cells (136). Whether the presence of these integrins could participate in prostate cancer progression and metastatic spread is even suggested by the demonstration of expression of αυβ3 in circulating exosomes extracted from the plasma of TRAMP mice (136).
CD9 is a member of the transmembrane 4 superfamily, also known as the tetraspanin family, and is considered as a key marker of exosomes. Soekmadji et al. recently demonstrated that exposure of LNCaP and DUCaP to dihydrotestosterone (DHT) increased the secretion of CD9-expressing exosomes (without any increase in the total number of secreted exosomes) (137). Proteomic analyses showed that DHT treatment, culture in androgen-free medium and treatment with the antiandrogen enzalutamide are effectively able to modify exosomes contents (137, 138). When CD9-enriched exosomes obtained after DHT treatment were inserted in the culture media of androgen-deprived LNCaP cells, an increased proliferation was observed. Whether this phenomenon could be observed in vivo as a means for cancer cells to escape or reduce the effects of androgen deprivation remains to be determined (137). It was recently demonstrated that AR can be loaded in EVs from LNCaP and 22Rv1 cells and thereby transferred to AR-null PC3 cells (139). In these recipient cells, AR proved to be functional through translocation to the nucleus and activation of target genes such as the PSA one (139). Of interest, EV-derived nuclear AR stimulated the proliferation of recipient cells in the absence of androgen, suggesting a new possible mechanism of resistance to androgen deprivation.
The interplays between AR and exosome pathways has been also highlighted in a recent experiment that used a variant of 22Rv1cells (140). Obtained by stable expression of the CRIPTO protein (an epidermal growth factor-related protein), this variant named Mes-PCa cells displays mesenchymal features that are accompanied by an increased released of EVs when comparing to parental cells. Androgen-dependent VCaP cells exposed to Mes-PCa-EVs showed decrease in both AR expression and transcriptional activity, activation of TGF-beta pathway, features of EMT, increased migrative, and invasive abilities as well as resistance to enzalutamide. Mes-PCa-EVs were found to express miRs known to target AR such as miR-21, miR-31 and miR-145 (140). EV-mediated cell plasticity, as observed for EMT, has also been described for NED (141). Although NED has frequently been reported as a result of androgen deprivation, the precise underlying mechanisms are still poorly understood (121, 122). Among soluble factors (cytokines, neuromediators, etc.) potentially involved, Il-6 proved to induce NED in PCa cells in association with lipid accumulation and induction of PPARgamma and adipocyte differentiation-related protein (ADRP) (141). In this study, C4-2B cells treated by Il-6 produced exosomes enriched in ADRP and able to be internalized by non-treated cells. Dendrite-like expression was, therefore, observed, both in PCa cells (DU-145 and C4-2B) and normal RWPE-1 cells. Of note, similar release of ADRP-containing exosomes were observed for cells treated by enzalutamide, confirming the potential induction of NED by androgen-deprivation therapy (ADT) (141).
It is unknown whether the exosome-mediated interactions between cancer cells could be used as a therapeutic tool (see below). Two independent studies nevertheless demonstrated that the oncosuppressor PTEN is loaded into exosomes and can thereby be transferred from PTEN-positive cells to PTEN-negative cells (142, 143). Moreover, they consistently disclosed decreased cell growth upon PTEN-containing exosome intake.
Association between resistance to chemotherapy and EVs shedding by cancer cells was first experimentally suggested by Shedden et al. (144): a correlation was found between chemosensitivity to anticancer agents and expression of membrane shedding-related genes. Anticancer drug expulsion via shed vesicles was suggested since doxorubicin accumulated in vesicles to an amount higher than its cytoplasmic concentrations (144). More recently, Kosgodage et al. consistently demonstrated an increase in 5-fluorouracyl-mediated apoptosis of PC3 cells in the presence of two inhibitors of EV release (chloramidine and bisindolylmaleimide-I) (145).
More interestingly, EVs are now also thought to transfer resistance to chemotherapy. Several research teams established prostate cancer cell lines resistant to doxorubicine (146), camptothecin (147), docetaxel (148), or paclitaxel (149). The pioneer experiments showed that exosomes obtained from DU145 cells resistant to doxorubicine were able to confer chemoresistance to parental DU145 cells but also to LNCaP and 22Rv1 cells (146). Similar results were obtained with exosomes from doxorubicine-resistant 22Rv1 cells (146). An increased expression of p-glycoprotein (multidrug resistance MDR-1, or p-gp) was observed in the exosomes of resistant cells, confirming the hypothesis that exosomes could behave, under chemotherapy conditions, as an efflux means for anticancer agent (144). Similar results were observed with camptothecin with decreased apoptosis when exposition to exosomes from resistant cells (147). Similarly, even if exosome-mediated transfer of chemoresistance was not strictly observed, uptake of exosomes extracted from DU145 cells resistant to docetaxel by parental sensitive DU145 cells proved to induce an increase in anchorage-independent growth (soft agar colony formation) (148). It even seems that chemoresistance could be transferred by exosomes from malignant epithelial cells to stromal fibroblasts (103).
The molecular mechanisms that underlie exosome-mediated transfer of chemoresistance remain to be fully understood but exposition to chemotherapy expectedly proved to induce changes in the exosomal contents, as evaluated both for individual proteins or miR (146, 150, 151) and by high-throughput analyses (152, 153). For example, exosomes from taxane-resistant PC3 cells contain integrin β4 and vinculin in an extent similar to parental cells but to a higher amount as compared to exosomes from sensitive cells (150). Transcriptomic analyses of the culture media of DU145 cells treated by fludarabin showed a global increase in the miR production (153). In surviving cells, expression of miR-485-3p was found to be enhanced, leading to a downregulation of the transcriptional repressor NF-YB and the subsequent overexpression of genes involved in chemoresistance and/or survival, such as topoisomerase IIa, p-gp, and cyclin B2 (153).
Of interest, impaired sensitivity to chemotherapy can be reversed: when DU145 cells resistant to camptothecin were exposed to exosomes from the benign prostate cell line PrEC, anchorage-independent growth decreased (147). Similar results were observed for DU145 cells resistant to paclitaxel and exposed to exosomes from the benign prostate cell line RWPE (149). Sensitivity to paclitaxel was even partly recovered (149). It is worthy to note that the same effect was reported when resistant DU145 cells were exposed to exosomes from the human mesenchymal stem cells, a non-prostatic cell line (149), a result reminiscent of the controlling role of stroma on cancer cell growth. Proteomic analyses can be performed to explore the molecular pathways that sustain the phenotypic shift between induction and reversal of chemoresistance (147). Exosomes extracted from the sera of prostate cancer patients submitted or resistant to chemotherapy proved to decrease sensitivity to chemotherapy in naïve prostate cancer cell lines (146). Whether these fundamental researches could be translated in routine practice remains to be determined but consistent data suggest that exosome-mediated chemoresistance could be efficient in vivo.
Radiation therapy is one of the recommended treatment for localized PCa. It has been demonstrated that it acts by inducing premature senescence rather than apoptotic cell death (154). This premature senescence is associated with an increase in exosome secretion (155–157). Activation of exosome secretion in irradiated cells is regulated by TSAP6 protein (transmembrane protein tumor suppressor), which belongs to the p53 pathway (157, 158). The composition of exosomes is also modified by radiation. Irradiated prostate cancer cells have been shown in vitro to produce exosomes with elevated levels of B7-H3 (CD276) (155). Similarly, levels of exosome-derived Hsp72 were found elevated in the sera of 13 patients undergoing radiation therapy for PCa (159). Of note, tissue expression of B7-H3 has been linked to AR pathway and immune response (160) and evaluated as a prognostic or theranostic marker of PCa (160–164). Radiation therapy does not only influence exosome release, it also influences exosome internalization by recipient cells: it has been demonstrated that radiation of human bone marrow-derived MSCs induces an increased uptake of exosomes (165).
Secreted exosomes proved to influence the behavior of recipient cells as demonstrated in glioblastoma: irradiated cells secrete exosomes enriched in molecules involved in cell migration such as Connective Tissue Growth Factor and Insulin-like Growth Factor-Binding Protein 2 (166). After exosome incorporation into recipient cells, proteins are activated that are also involved in cell migration such as Src (166, 167), which has been shown to be enriched in prostate cancer cell exosomes (168). An interesting issue is the fact that effects of radiation are not limited to irradiated cells but is also communicated to adjacent cells, a phenomenon known as the radiation-induced bystander effect (RIBE) (157, 169, 170). Observed RIBEs are either beneficial (senescence of non-irradiated cancer cells) or deleterious (side-effects on non-malignant cells) and can be observed in cells adjacent to irradiated cells or in distant cells. The underlying mechanisms are incompletely understood but are likely to imply soluble clastogenic factors such as exosomes (157, 169, 170). First studies about the exosome involvement in RIBE initiation came very recently (171–173). As a demonstrating example, exosomes were isolated from the supernatant medium of BEP2D cells cultures (normal human bronchial epithelial cell line) and the expression profile of miRs were compared between exosomes collected from 2 Gy-irradiated cells and from non-irradiated cells (174). Several miRs were found to be differentially expressed, including miR-7-5p. Exosomes containing miR-7-5p proved to be internalized by non-irradiated BEP2D cells which, therefore, entered in autophagy, by reduced expression of EGFR, a known target of miR-7-5p (174). A distant bystander effect was also demonstrated in vivo (175). After focal brain irradiation of mice, exosomes were obtained from irradiated astrocytes and found to express high levels of miR-7. Injected into the tail vein of non-irradiated mice, they were able to induce autophagy of lung tissues by directly targeting Bcl-2 (175). Other miRs may have similar effects, such as miR-145, which proved to sensitize PCa cells to radiation (176). Whether these recent advances in RIBE mechanisms would be translated in clinical practice to enhance both tumor cell kill and normal tissue protection (for example through blockade of exosomal secretion) remains to be determined.
Several high-throughput analyses comparing body fluids from prostate cancer patients and controls demonstrated that the cargo of EVs is specific for the parental cells and the conditions in which they produce them (44, 46, 48, 53, 54, 59). Exploring whether EVs could be used as new providers of potential cancer biomarkers is the next logical step. Many factors influence EVs shedding and a first approach can, therefore, consist in evaluating the sole enumeration of EVs as a diagnostic tool, as first described in melanoma (177). As an easily available body fluid, urine was soon used as a means to detect EVs (83, 178) and demonstration has been made that prior prostate massage enhance the efficiency of urinary EVs detection (50, 179, 180). Urine from prostate cancer patients was, therefore, characterized by elevated exosomes secretion (50, 69, 178) and a decrease was observed after cancer treatment such as ADT (178). In plasma, feasibility was rapidly demonstrated through both in vivo mice experiments and direct blood sampling in human patients. Large oncosomes were detected in the plasma of TRAMP mice as caveolin-positive large vesicles and their number was found to be higher in mice with lymph node involvement or lung distant metastases (85). In Humans, studies consistently confirmed that plasma EVs are more numerous in prostate cancer patients than in healthy volunteers or patients with benign prostate hyperplasia (47, 51, 181–185). Diminution was observed after radical prostatectomy (183). A correlation was also observed with cancer aggressiveness, metastatic spread, and/or Gleason score (181–183). African-American patients were also demonstrated to have significantly higher amounts of EVs in their plasma and sera than European-American patients and, within their EVs, higher amounts of inhibitors of apoptosis proteins such as survivin, XIAP, and cIAP-2 (186).
Besides EVs enumeration, a specific EV component can also be specifically measured to help prostate cancer diagnosis. Again, urines and blood were tested and all molecules present in EVs can be evaluated (lipids, proteins, RNAs, miRs, long non-coding RNAs, etc.). As an example, a high-throughput mass spectrometry quantitative lipidomics analysis recently showed significantly different levels of nine lipid species including phosphatidylserine and lactosylceramide (58). Among exosomal proteins evaluated for their potential diagnostic value, PSMA, PSA, PTEN, survivin, p-gp, claudin3, FABP5, gammaglutamyl transferase, EGFR, ITGA3, ITGB1, TM256, and TGM4-ADSV were found elevated in the blood (plasma or serum) or the urine from prostate cancer patients (25, 35, 36, 48, 53, 54, 142, 151, 178, 181, 187–189) (Table 2). More generally, specific posttranscriptional modifications can also be explored: assessment of N-glycosylation pattern from urine EVs showed for example a decreased in overall biantennary core-fucosylation in prostate cancer patients (190).
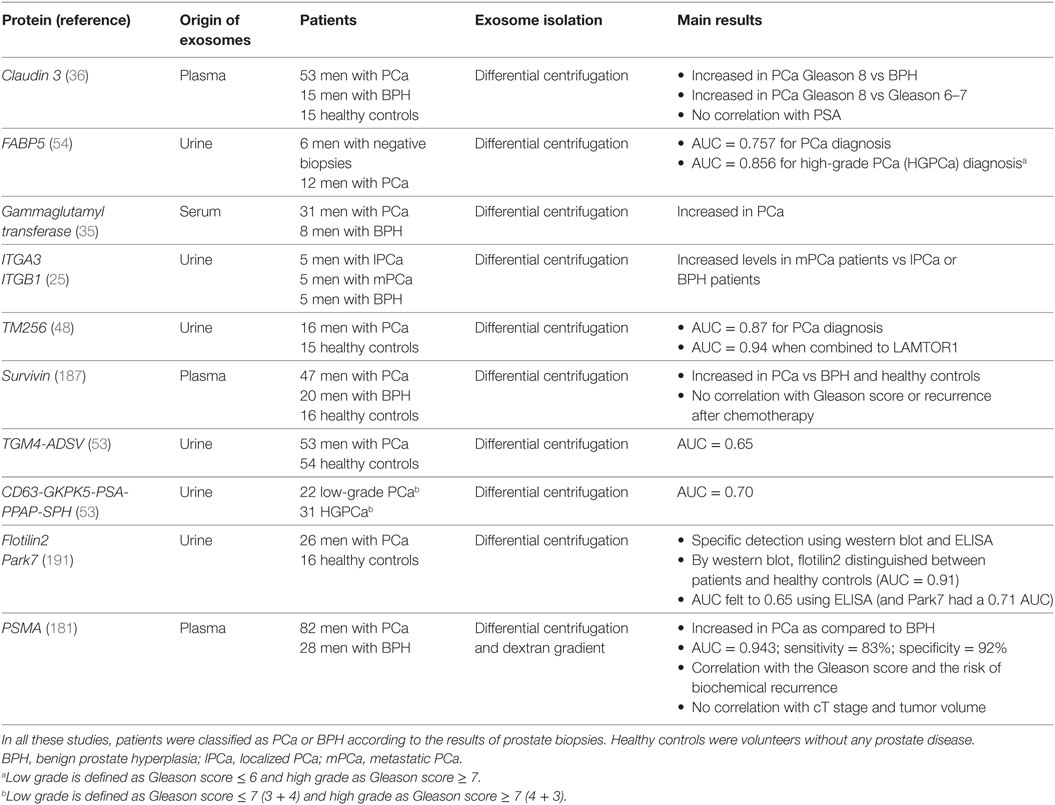
Table 2. Clinical studies exploring exosomal proteins as potential prostate cancer biomarkers.
PCA3 is a long non-coding RNA and has been proposed as urinary biomarker prostate cancer since 2003 (192). It has been detected in urine exosomes as soon as 2009 (83) and a specific assay based on the measure of PCA3 and ERG in urinary exosomes without prostate examination was developed to allow detection between Gleason score 6 vs Gleason score 7–10 prostate cancers (193). The real value of this specific exosomal test remains to be determined: although detection is more robust in the exosomes than in urine sediments (179, 180), it seems to be less efficient than the measure in the whole urine extracts (194). Another long non-coding RNA, lincRNA-p21, was identified in urine exosomes and appeared more abundant in prostate cancer patients than in patients with benign prostate hyperplasia (195). Other exosomal RNA species with suggestive diagnostic values are TMPRSS2:ERG fusion transcript, BIRC5, ERG, and TMPRSS2 (180, 196) (Table 3). In fact, a clear priority has been given to miRs because of fashion trend, because they represent the most abundant exosomal RNA species, and because exosome shedding and miR production are intrinsically related. Whether miRs circulate in blood or are present in body fluids (such as urines) freely (i.e., free of cells and cell compartments) or encapsulated in vesicles remains controversial, and this could depend on the nature of the body fluid itself (197–200). Because of this ambiguity, literature search using “EV” or “exosome” as keyword when looking for miRs as diagnostic tool for prostate cancer diagnosis dramatically reduces the number of published studies. Numerous papers reported detection and quantification of miRs in plasma or urines without any notion on whether the evaluated miRs were exosome-derived or not. We here focused on those papers where the exosomal origin was clearly specified while other reviews can be read to explore more largely the diagnostic value of miRs in prostate cancer (201–203).
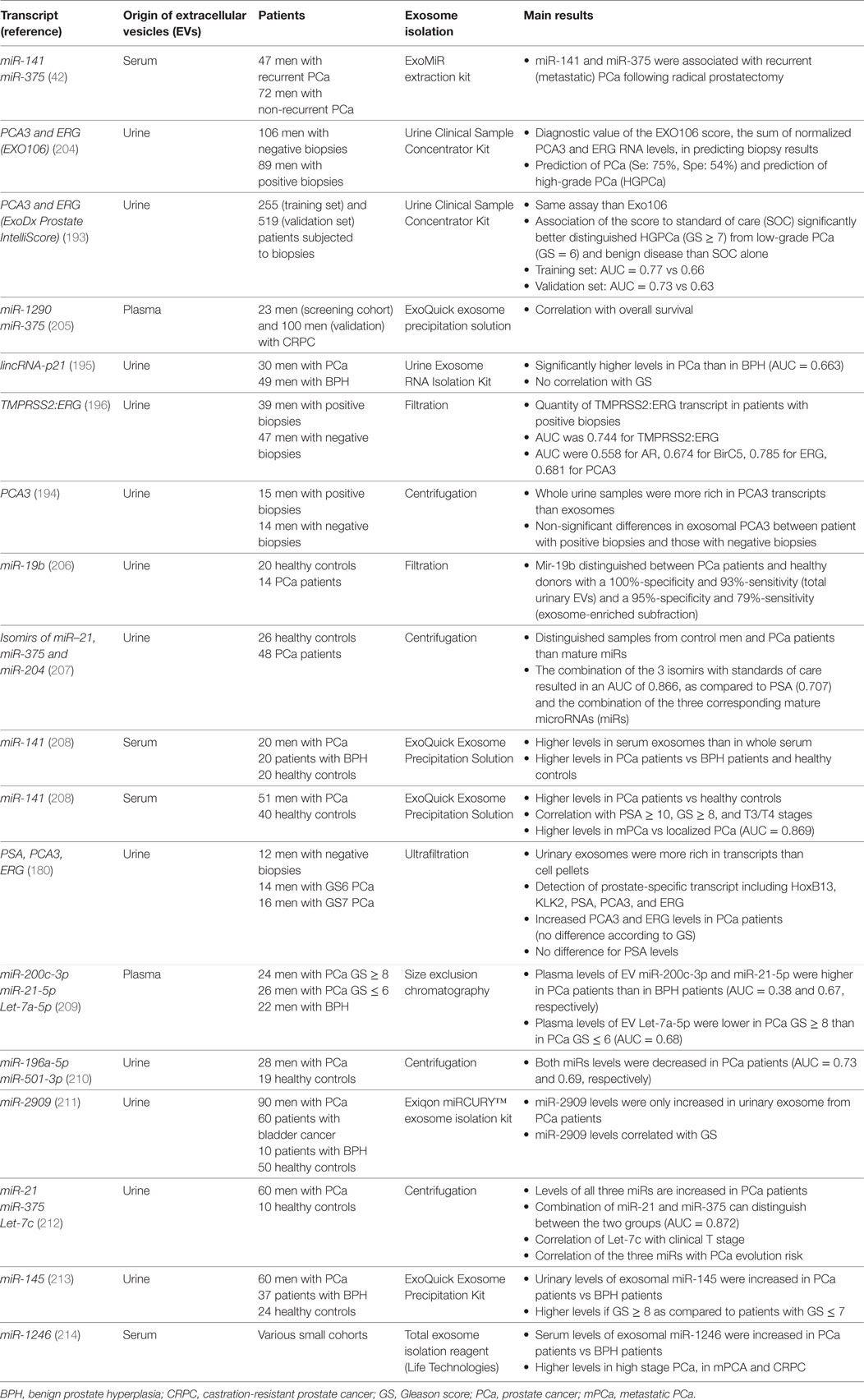
Table 3. Clinical studies exploring exosomal RNA species as potential prostate cancer biomarkers.
Several papers identified some exosomal miRs as potential diagnostic biomarkers of prostate cancer (Table 3). Among them, miR-141 is one of the most studied (in fact also for several cancers) because it belongs to the miR-200 family (along with miR-200a, b and c and miR-429). This family is involved in fundamental events of epithelial carcinogenesis such as cell transformation, EMT, or metastatic spread. In a cornerstone study about prostate cancer, miR-141 was identified (along with miR-375) among the exosomal miRs differentially expressed when comparing the expression of a panel of 742 miRs in plasma- or serum-derived circulating EVs in 78 prostate cancer patients and 28 healthy controls (42). Expression of miR-141 was found to be correlated with Gleason score and tumor progression, as well as the metastatic spread. Results in serum exosomes were independently confirmed (208, 215) while upregulation of miR-141 was also observed in urinary exosomes (52). Other exosomal miRs with potential diagnostic or prognostic values include miR-19b, miR-21, miR-34a, miR-375, miR-483-5p, miR-575, miR-1290, miR-1246, and let-7c (42, 52, 152, 205, 206, 212, 216). Serum exosomal let-7c has been correlated with Gleason score (212), miR-1290 and miR-375 to overall survival (205) while the decrease in plasma exosomal miR-34a was predictive of prostate cancer progression and poor response to docetaxel (152).
The situation in fact becomes more and more complicated. For example, recent studies deal with isomirs, the various miR isoforms generated through the miR processing and maturation process from a single miR locus (217). Koppers-Lalic et al. showed that the use of miR-21, miR-375, and miR-204 isomirs could provide a better diagnostic tool than that of the mature miRs (207).
Responses to ADT and to radiotherapy may also be predicted by the use of exosomal markers. A recent study suggests that some specific miRs in serum exosomes, particularly miR-654-3p and miR-379-5p, could be able to predict response to radiotherapy (218). As regards ADT, AR-V7 is an androgen receptor splice variant, which is constitutively active because of the truncation of the ligand-binding domain (219). The expression of AR-V7 constitutes a means by which prostate cancer cells escape to androgen-deprivation and Antonarakis et al. demonstrated a link between the expression of AR-V7 in prostate cancer circulating cells and resistance to the second generation antiandrogens enzalutamide and abiraterone (220). AR–V7 can be detected in plasma prostate cancer-derived exosomes (139, 221). Similarly to CTC-derived AR–V7, exosome-derived AR-V7 was found to be a reliable predictor of resistance to castration (221).
As well as several cancers, a great hope emerged from studies demonstrating the role of exosomes, and more broadly EVs, in both prostate carcinogenesis, prostate cancer progression, and prostate cancer diagnosis. Numerous fundamental studies suggested that the various exosome contents could be used as a powerful tool to better understand how EV-mediated cell–cell interactions, as integral part of the reciprocal talk between malignant cells and their neighboring stromal cells, and constitute major events for cancer occurrence and progression. Numerous high-throughput analyses provided tens of proteins, RNAs, miRs as potential diagnostic and prognostic markers. Efforts are still warranted to translate these results into routine practice. Efforts have to be made to standardize EV classification and identification and to optimize the detection and isolation techniques. While the use of dedicated kits remains controversial (222), the development of specific and simple devices is, therefore, warranted (223, 224). Likewise, integrative approaches are needed that explored simultaneously the different exosomal contents. By exploring from the same samples both the miR and the long non-coding RNA repertories in prostate cancer cell lines, Ahadi et al. demonstrated that exosomal long non-coding RNAs are highly enriched in target motifs for the miRs that are also present in the same exosomes (37). This suggests that exosome sorting is an extraordinary organized process that allows selection of both specific biologically significant messages (miRs) and their specific conveyors (long non-coding RNAs).
Implication of exosomes in the prediction of the response to treatment (146, 152), in the prostate cancer prognosis or the patients’ overall survival (205), is highly promising of the announced shift toward personalized medicine. Whether exosomes could serve by themselves as therapeutic agents is also suggested (5, 225, 226). Their use as vectors of anticancer agents is advocated because they are cell-type specific (specific of both the parental and the target cells), lipophilic, and non-immunogenic by themselves, because they can cross membranes (including the blood–brain barrier), are not filtered by the glomerulus (long life in the circulation) and can carry very diverse molecules (proteins, lipids, RNAs, miRs, siRNAs, etc.) from diverse compartments (cell surface, cytosol) (227). EVs as transporters of anticancer agents have, therefore, been evaluated in prostate cancer although preclinical studies remain necessary before clinical applications (228). EVs can even function as cell-free vaccines to decrease or stabilize tumor growth (229). Since exosomes are able to significantly participate to cancer local progression and distal spread, it is also tempting to develop strategies able to block exosome-mediated procarcinogenetic effects: efforts are currently made to either block exosome release by parental cells, exosome circulation in the bloodstream and exosome interactions with target cells (125).
The author confirms being the sole contributor of this work and approved it for publication.
The author declares that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
1. Hanahan D, Weinberg RA. Hallmarks of cancer: the next generation. Cell (2011) 144(5):646–74. doi:10.1016/j.cell.2011.02.013
2. Shiao SL, Chu GC, Chung LW. Regulation of prostate cancer progression by the tumor microenvironment. Cancer Lett (2016) 380(1):340–8. doi:10.1016/j.canlet.2015.12.022
3. Naito Y, Yoshioka Y, Yamamoto Y, Ochiya T. How cancer cells dictate their microenvironment: present roles of extracellular vesicles. Cell Mol Life Sci (2017) 74(4):697–713. doi:10.1007/s00018-016-2346-3
4. Roma-Rodrigues C, Fernandes AR, Baptista PV. Exosome in tumour microenvironment: overview of the crosstalk between normal and cancer cells. Biomed Res Int (2014) 2014:179486. doi:10.1155/2014/179486
5. Zocco D, Ferruzzi P, Cappello F, Kuo WP, Fais S. Extracellular vesicles as shuttles of tumor biomarkers and anti-tumor drugs. Front Oncol (2014) 4:267. doi:10.3389/fonc.2014.00267
6. Atay S, Godwin AK. Tumor-derived exosomes: a message delivery system for tumor progression. Commun Integr Biol (2014) 7(1):e28231. doi:10.4161/cib.28231
7. Azmi AS, Bao B, Sarkar FH. Exosomes in cancer development, metastasis, and drug resistance: a comprehensive review. Cancer Metastasis Rev (2013) 32(3–4):623–42. doi:10.1007/s10555-013-9441-9
8. Colombo M, Raposo G, Thery C. Biogenesis, secretion, and intercellular interactions of exosomes and other extracellular vesicles. Annu Rev Cell Dev Biol (2014) 30:255–89. doi:10.1146/annurev-cellbio-101512-122326
9. Yanez-Mo M, Siljander PR, Andreu Z, Zavec AB, Borras FE, Buzas EI, et al. Biological properties of extracellular vesicles and their physiological functions. J Extracell Vesicles (2015) 4:27066. doi:10.3402/jev.v4.27066
10. Minciacchi VR, Freeman MR, Di Vizio D. Extracellular vesicles in cancer: exosomes, microvesicles and the emerging role of large oncosomes. Semin Cell Dev Biol (2015) 40:41–51. doi:10.1016/j.semcdb.2015.02.010
11. Harding C, Heuser J, Stahl P. Receptor-mediated endocytosis of transferrin and recycling of the transferrin receptor in rat reticulocytes. J Cell Biol (1983) 97(2):329–39. doi:10.1083/jcb.97.2.329
12. Johnstone RM, Adam M, Hammond JR, Orr L, Turbide C. Vesicle formation during reticulocyte maturation. Association of plasma membrane activities with released vesicles (exosomes). J Biol Chem (1987) 262(19):9412–20.
13. Pan BT, Johnstone RM. Fate of the transferrin receptor during maturation of sheep reticulocytes in vitro: selective externalization of the receptor. Cell (1983) 33(3):967–78. doi:10.1016/0092-8674(83)90040-5
14. Pan BT, Teng K, Wu C, Adam M, Johnstone RM. Electron microscopic evidence for externalization of the transferrin receptor in vesicular form in sheep reticulocytes. J Cell Biol (1985) 101(3):942–8. doi:10.1083/jcb.101.3.942
15. Cocucci E, Racchetti G, Meldolesi J. Shedding microvesicles: artefacts no more. Trends Cell Biol (2009) 19(2):43–51. doi:10.1016/j.tcb.2008.11.003
16. Thery C, Zitvogel L, Amigorena S. Exosomes: composition, biogenesis and function. Nat Rev Immunol (2002) 2(8):569–79. doi:10.1038/nri855
17. Gould SJ, Raposo G. As we wait: coping with an imperfect nomenclature for extracellular vesicles. J Extracell Vesicles (2013) 2(1):20389. doi:10.3402/jev.v2i0.20389
18. Royo F, Zuniga-Garcia P, Sanchez-Mosquera P, Egia A, Perez A, Loizaga A, et al. Different EV enrichment methods suitable for clinical settings yield different subpopulations of urinary extracellular vesicles from human samples. J Extracell Vesicles (2016) 5:29497. doi:10.3402/jev.v5.29497
19. Royo F, Diwan I, Tackett MR, Zuniga P, Sanchez-Mosquera P, Loizaga-Iriarte A, et al. Comparative miRNA analysis of urine extracellular vesicles isolated through five different methods. Cancers (Basel) (2016) 8(12):E112. doi:10.3390/cancers8120112
20. Thery C. Exosomes: secreted vesicles and intercellular communications. F1000 Biol Rep (2011) 3:15. doi:10.3410/B3-15
21. Kalra H, Simpson RJ, Ji H, Aikawa E, Altevogt P, Askenase P, et al. Vesiclepedia: a compendium for extracellular vesicles with continuous community annotation. PLoS Biol (2012) 10(12):e1001450. doi:10.1371/journal.pbio.1001450
22. Raposo G, Stoorvogel W. Extracellular vesicles: exosomes, microvesicles, and friends. J Cell Biol (2013) 200(4):373–83. doi:10.1083/jcb.201211138
23. Hanson PI, Cashikar A. Multivesicular body morphogenesis. Annu Rev Cell Dev Biol (2012) 28:337–62. doi:10.1146/annurev-cellbio-092910-154152
24. Parolini I, Federici C, Raggi C, Lugini L, Palleschi S, De Milito A, et al. Microenvironmental pH is a key factor for exosome traffic in tumor cells. J Biol Chem (2009) 284(49):34211–22. doi:10.1074/jbc.M109.041152
25. Bijnsdorp IV, Geldof AA, Lavaei M, Piersma SR, van Moorselaar RJ, Jimenez CR. Exosomal ITGA3 interferes with non-cancerous prostate cell functions and is increased in urine exosomes of metastatic prostate cancer patients. J Extracell Vesicles (2013) 2(1):22097. doi:10.3402/jev.v2i0.22097
26. Lazaro-Ibanez E, Sanz-Garcia A, Visakorpi T, Escobedo-Lucea C, Siljander P, Ayuso-Sacido A, et al. Different gDNA content in the subpopulations of prostate cancer extracellular vesicles: apoptotic bodies, microvesicles, and exosomes. Prostate (2014) 74(14):1379–90. doi:10.1002/pros.22853
27. Jansen FH, Krijgsveld J, van Rijswijk A, van den Bemd GJ, van den Berg MS, van Weerden WM, et al. Exosomal secretion of cytoplasmic prostate cancer xenograft-derived proteins. Mol Cell Proteomics (2009) 8(6):1192–205. doi:10.1074/mcp.M800443-MCP200
28. Sandvig K, Llorente A. Proteomic analysis of microvesicles released by the human prostate cancer cell line PC-3. Mol Cell Proteomics (2012) 11(7):M111012914. doi:10.1074/mcp.M111.012914
29. Hosseini-Beheshti E, Pham S, Adomat H, Li N, Tomlinson Guns ES. Exosomes as biomarker enriched microvesicles: characterization of exosomal proteins derived from a panel of prostate cell lines with distinct AR phenotypes. Mol Cell Proteomics (2012) 11(10):863–85. doi:10.1074/mcp.M111.014845
30. Duijvesz D, Burnum-Johnson KE, Gritsenko MA, Hoogland AM, Vredenbregt-van den Berg MS, Willemsen R, et al. Proteomic profiling of exosomes leads to the identification of novel biomarkers for prostate cancer. PLoS One (2013) 8(12):e82589. doi:10.1371/journal.pone.0082589
31. Webber J, Stone TC, Katilius E, Smith BC, Gordon B, Mason MD, et al. Proteomics analysis of cancer exosomes using a novel modified aptamer-based array (SOMAscan) platform. Mol Cell Proteomics (2014) 13(4):1050–64. doi:10.1074/mcp.M113.032136
32. Minciacchi VR, You S, Spinelli C, Morley S, Zandian M, Aspuria PJ, et al. Large oncosomes contain distinct protein cargo and represent a separate functional class of tumor-derived extracellular vesicles. Oncotarget (2015) 6(13):11327–41. doi:10.18632/oncotarget.3598
33. Sanchez CA, Andahur EI, Valenzuela R, Castellon EA, Fulla JA, Ramos CG, et al. Exosomes from bulk and stem cells from human prostate cancer have a differential microRNA content that contributes cooperatively over local and pre-metastatic niche. Oncotarget (2016) 7(4):3993–4008. doi:10.18632/oncotarget.6540
34. Rauschenberger L, Staar D, Thom K, Scharf C, Venz S, Homuth G, et al. Exosomal particles secreted by prostate cancer cells are potent mRNA and protein vehicles for the interference of tumor and tumor environment. Prostate (2016) 76(4):409–24. doi:10.1002/pros.23132
35. Kawakami K, Fujita Y, Matsuda Y, Arai T, Horie K, Kameyama K, et al. Gamma-glutamyltransferase activity in exosomes as a potential marker for prostate cancer. BMC Cancer (2017) 17(1):316. doi:10.1186/s12885-017-3301-x
36. Worst TS, von Hardenberg J, Gross JC, Erben P, Schnolzer M, Hausser I, et al. Database-augmented mass spectrometry analysis of exosomes identifies claudin 3 as a putative prostate cancer biomarker. Mol Cell Proteomics (2017) 16(6):998–1008. doi:10.1074/mcp.M117.068577
37. Ahadi A, Brennan S, Kennedy PJ, Hutvagner G, Tran N. Long non-coding RNAs harboring miRNA seed regions are enriched in prostate cancer exosomes. Sci Rep (2016) 6:24922. doi:10.1038/srep24922
38. Lazaro-Ibanez E, Lunavat TR, Jang SC, Escobedo-Lucea C, Oliver-De La Cruz J, Siljander P, et al. Distinct prostate cancer-related mRNA cargo in extracellular vesicle subsets from prostate cell lines. BMC Cancer (2017) 17(1):92. doi:10.1186/s12885-017-3087-x
39. Thery C, Boussac M, Veron P, Ricciardi-Castagnoli P, Raposo G, Garin J, et al. Proteomic analysis of dendritic cell-derived exosomes: a secreted subcellular compartment distinct from apoptotic vesicles. J Immunol (2001) 166(12):7309–18. doi:10.4049/jimmunol.166.12.7309
40. Poliakov A, Spilman M, Dokland T, Amling CL, Mobley JA. Structural heterogeneity and protein composition of exosome-like vesicles (prostasomes) in human semen. Prostate (2009) 69(2):159–67. doi:10.1002/pros.20860
41. Ronquist KG, Ronquist G, Larsson A, Carlsson L. Proteomic analysis of prostate cancer metastasis-derived prostasomes. Anticancer Res (2010) 30(2):285–90.
42. Bryant RJ, Pawlowski T, Catto JW, Marsden G, Vessella RL, Rhees B, et al. Changes in circulating microRNA levels associated with prostate cancer. Br J Cancer (2012) 106(4):768–74. doi:10.1038/bjc.2011.595
43. Hessvik NP, Phuyal S, Brech A, Sandvig K, Llorente A. Profiling of microRNAs in exosomes released from PC-3 prostate cancer cells. Biochim Biophys Acta (2012) 1819(11–12):1154–63. doi:10.1016/j.bbagrm.2012.08.016
44. Principe S, Jones EE, Kim Y, Sinha A, Nyalwidhe JO, Brooks J, et al. In-depth proteomic analyses of exosomes isolated from expressed prostatic secretions in urine. Proteomics (2013) 13(10–11):1667–71. doi:10.1002/pmic.201200561
45. Kovak MR, Saraswati S, Goddard SD, Diekman AB. Proteomic identification of galectin-3 binding ligands and characterization of galectin-3 proteolytic cleavage in human prostasomes. Andrology (2013) 1(5):682–91. doi:10.1111/j.2047-2927.2013.00099.x
46. Nyalwidhe JO, Betesh LR, Powers TW, Jones EE, White KY, Burch TC, et al. Increased bisecting N-acetylglucosamine and decreased branched chain glycans of N-linked glycoproteins in expressed prostatic secretions associated with prostate cancer progression. Proteomics Clin Appl (2013) 7(9–10):677–89. doi:10.1002/prca.201200134
47. Mizutani K, Terazawa R, Kameyama K, Kato T, Horie K, Tsuchiya T, et al. Isolation of prostate cancer-related exosomes. Anticancer Res (2014) 34(7):3419–23.
48. Overbye A, Skotland T, Koehler CJ, Thiede B, Seierstad T, Berge V, et al. Identification of prostate cancer biomarkers in urinary exosomes. Oncotarget (2015) 6(30):30357–76. doi:10.18632/oncotarget.4851
49. Chiasserini D, Mazzoni M, Bordi F, Sennato S, Susta F, Orvietani PL, et al. Identification and partial characterization of two populations of prostasomes by a combination of dynamic light scattering and proteomic analysis. J Membr Biol (2015) 248(6):991–1004. doi:10.1007/s00232-015-9810-0
50. Duijvesz D, Versluis CY, van der Fels CA, Vredenbregt-van den Berg MS, Leivo J, Peltola MT, et al. Immuno-based detection of extracellular vesicles in urine as diagnostic marker for prostate cancer. Int J Cancer (2015) 137(12):2869–78. doi:10.1002/ijc.29664
51. Turay D, Khan S, Diaz Osterman CJ, Curtis MP, Khaira B, Neidigh JW, et al. Proteomic profiling of serum-derived exosomes from ethnically diverse prostate cancer patients. Cancer Invest (2016) 34(1):1–11. doi:10.3109/07357907.2015.1081921
52. Samsonov R, Shtam T, Burdakov V, Glotov A, Tsyrlina E, Berstein L, et al. Lectin-induced agglutination method of urinary exosomes isolation followed by mi-RNA analysis: application for prostate cancer diagnostic. Prostate (2016) 76(1):68–79. doi:10.1002/pros.23101
53. Sequeiros T, Rigau M, Chiva C, Montes M, Garcia-Grau I, Garcia M, et al. Targeted proteomics in urinary extracellular vesicles identifies biomarkers for diagnosis and prognosis of prostate cancer. Oncotarget (2017) 8(3):4960–76. doi:10.18632/oncotarget.13634
54. Fujita K, Kume H, Matsuzaki K, Kawashima A, Ujike T, Nagahara A, et al. Proteomic analysis of urinary extracellular vesicles from high Gleason score prostate cancer. Sci Rep (2017) 7:42961. doi:10.1038/srep42961
55. Yang C, Guo WB, Zhang WS, Bian J, Yang JK, Zhou QZ, et al. Comprehensive proteomics analysis of exosomes derived from human seminal plasma. Andrology (2017) 5(5):1007–15. doi:10.1111/andr.12412
56. Yang JS, Lee JC, Byeon SK, Rha KH, Moon MH. Size dependent lipidomic analysis of urinary exosomes from patients with prostate cancer by flow field-flow fractionation and nanoflow liquid chromatography-tandem mass spectrometry. Anal Chem (2017) 89(4):2488–96. doi:10.1021/acs.analchem.6b04634
57. Puhka M, Takatalo M, Nordberg ME, Valkonen S, Nandania J, Aatonen M, et al. Metabolomic profiling of extracellular vesicles and alternative normalization methods reveal enriched metabolites and strategies to study prostate cancer-related changes. Theranostics (2017) 7(16):3824–41. doi:10.7150/thno.19890
58. Skotland T, Ekroos K, Kauhanen D, Simolin H, Seierstad T, Berge V, et al. Molecular lipid species in urinary exosomes as potential prostate cancer biomarkers. Eur J Cancer (2017) 70:122–32. doi:10.1016/j.ejca.2016.10.011
59. Royo F, Zuniga-Garcia P, Torrano V, Loizaga A, Sanchez-Mosquera P, Ugalde-Olano A, et al. Transcriptomic profiling of urine extracellular vesicles reveals alterations of CDH3 in prostate cancer. Oncotarget (2016) 7(6):6835–46. doi:10.18632/oncotarget.6899
60. Kowal J, Tkach M, Thery C. Biogenesis and secretion of exosomes. Curr Opin Cell Biol (2014) 29:116–25. doi:10.1016/j.ceb.2014.05.004
61. Valadi H, Ekstrom K, Bossios A, Sjostrand M, Lee JJ, Lotvall JO. Exosome-mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells. Nat Cell Biol (2007) 9(6):654–9. doi:10.1038/ncb1596
62. Kahlert C, Kalluri R. Exosomes in tumor microenvironment influence cancer progression and metastasis. J Mol Med (Berl) (2013) 91(4):431–7. doi:10.1007/s00109-013-1020-6
63. Al-Nedawi K, Meehan B, Micallef J, Lhotak V, May L, Guha A, et al. Intercellular transfer of the oncogenic receptor EGFRvIII by microvesicles derived from tumour cells. Nat Cell Biol (2008) 10(5):619–24. doi:10.1038/ncb1725
64. Ronquist G, Hedstrom M. Restoration of detergent-inactivated adenosine triphosphatase activity of human prostatic fluid with concanavalin A. Biochim Biophys Acta (1977) 483(2):483–6. doi:10.1016/0005-2744(77)90078-X
65. Ronquist G, Brody I, Gottfries A, Stegmayr B. An Mg2+ and Ca2+-stimulated adenosine triphosphatase in human prostatic fluid – part II. Andrologia (1978) 10(6):427–33. doi:10.1111/j.1439-0272.1978.tb03064.x
66. Ronquist G, Brody I, Gottfries A, Stegmayr B. An Mg2+ and Ca2+ -stimulated adenosine triphosphatase in human prostatic fluid: part I. Andrologia (1978) 10(4):261–72. doi:10.1111/j.1439-0272.1978.tb03030.x
67. Giusti I, Dolo V. Extracellular vesicles in prostate cancer: new future clinical strategies? Biomed Res Int (2014) 2014:561571. doi:10.1155/2014/561571
68. Zijlstra C, Stoorvogel W. Prostasomes as a source of diagnostic biomarkers for prostate cancer. J Clin Invest (2016) 126(4):1144–51. doi:10.1172/JCI81128
69. Ronquist GK, Larsson A, Stavreus-Evers A, Ronquist G. Prostasomes are heterogeneous regarding size and appearance but affiliated to one DNA-containing exosome family. Prostate (2012) 72(16):1736–45. doi:10.1002/pros.22526
70. Arvidson G, Ronquist G, Wikander G, Ojteg AC. Human prostasome membranes exhibit very high cholesterol/phospholipid ratios yielding high molecular ordering. Biochim Biophys Acta (1989) 984(2):167–73. doi:10.1016/0005-2736(89)90212-5
71. Ronquist G. Prostasomes are mediators of intercellular communication: from basic research to clinical implications. J Intern Med (2012) 271(4):400–13. doi:10.1111/j.1365-2796.2011.02487.x
72. Duijvesz D, Luider T, Bangma CH, Jenster G. Exosomes as biomarker treasure chests for prostate cancer. Eur Urol (2011) 59(5):823–31. doi:10.1016/j.eururo.2010.12.031
73. Stridsberg M, Fabiani R, Lukinius A, Ronquist G. Prostasomes are neuroendocrine-like vesicles in human semen. Prostate (1996) 29(5):287–95. doi:10.1002/(SICI)1097-0045(199611)29:5<287::AID-PROS3>3.0.CO;2-7
74. Arienti G, Carlini E, Polci A, Cosmi EV, Palmerini CA. Fatty acid pattern of human prostasome lipid. Arch Biochem Biophys (1998) 358(2):391–5. doi:10.1006/abbi.1998.0876
75. Olsson I, Ronquist G. Nucleic acid association to human prostasomes. Arch Androl (1990) 24(1):1–10. doi:10.3109/01485019008986852
76. Burden HP, Holmes CH, Persad R, Whittington K. Prostasomes – their effects on human male reproduction and fertility. Hum Reprod Update (2006) 12(3):283–92. doi:10.1093/humupd/dmi052
77. Aalberts M, Stout TA, Stoorvogel W. Prostasomes: extracellular vesicles from the prostate. Reproduction (2014) 147(1):R1–14. doi:10.1530/REP-13-0358
78. Schrimpf SP, Hellman U, Carlsson L, Larsson A, Ronquist G, Nilsson BO. Identification of dipeptidyl peptidase IV as the antigen of a monoclonal anti-prostasome antibody. Prostate (1999) 38(1):35–9. doi:10.1002/(SICI)1097-0045(19990101)38:1<35::AID-PROS4>3.0.CO;2-J
79. Carlsson L, Nilsson O, Larsson A, Stridsberg M, Sahlen G, Ronquist G. Characteristics of human prostasomes isolated from three different sources. Prostate (2003) 54(4):322–30. doi:10.1002/pros.10189
80. Sahlen GE, Egevad L, Ahlander A, Norlen BJ, Ronquist G, Nilsson BO. Ultrastructure of the secretion of prostasomes from benign and malignant epithelial cells in the prostate. Prostate (2002) 53(3):192–9. doi:10.1002/pros.10126
81. Sahlen G, Ahlander A, Frost A, Ronquist G, Norlen BJ, Nilsson BO. Prostasomes are secreted from poorly differentiated cells of prostate cancer metastases. Prostate (2004) 61(3):291–7. doi:10.1002/pros.20090
82. Ronquist G, Nilsson BO. The Janus-faced nature of prostasomes: their pluripotency favours the normal reproductive process and malignant prostate growth. Prostate Cancer Prostatic Dis (2004) 7(1):21–31. doi:10.1038/sj.pcan.4500684
83. Nilsson J, Skog J, Nordstrand A, Baranov V, Mincheva-Nilsson L, Breakefield XO, et al. Prostate cancer-derived urine exosomes: a novel approach to biomarkers for prostate cancer. Br J Cancer (2009) 100(10):1603–7. doi:10.1038/sj.bjc.6605058
84. Di Vizio D, Kim J, Hager MH, Morello M, Yang W, Lafargue CJ, et al. Oncosome formation in prostate cancer: association with a region of frequent chromosomal deletion in metastatic disease. Cancer Res (2009) 69(13):5601–9. doi:10.1158/0008-5472.CAN-08-3860
85. Di Vizio D, Morello M, Dudley AC, Schow PW, Adam RM, Morley S, et al. Large oncosomes in human prostate cancer tissues and in the circulation of mice with metastatic disease. Am J Pathol (2012) 181(5):1573–84. doi:10.1016/j.ajpath.2012.07.030
86. Kim J, Morley S, Le M, Bedoret D, Umetsu DT, Di Vizio D, et al. Enhanced shedding of extracellular vesicles from amoeboid prostate cancer cells: potential effects on the tumor microenvironment. Cancer Biol Ther (2014) 15(4):409–18. doi:10.4161/cbt.27627
87. Mathivanan S, Simpson RJ. ExoCarta: a compendium of exosomal proteins and RNA. Proteomics (2009) 9(21):4997–5000. doi:10.1002/pmic.200900351
88. Kim DK, Kang B, Kim OY, Choi DS, Lee J, Kim SR, et al. EVpedia: an integrated database of high-throughput data for systemic analyses of extracellular vesicles. J Extracell Vesicles (2013) 2(1):20384. doi:10.3402/jev.v2i0.20384
89. Tripathi M, Billet S, Bhowmick NA. Understanding the role of stromal fibroblasts in cancer progression. Cell Adh Migr (2012) 6(3):231–5. doi:10.4161/cam.20419
90. Hagglof C, Bergh A. The stroma—a key regulator in prostate function and malignancy. Cancers (Basel) (2012) 4(2):531–48. doi:10.3390/cancers4020531
91. Ao M, Franco OE, Park D, Raman D, Williams K, Hayward SW. Cross-talk between paracrine-acting cytokine and chemokine pathways promotes malignancy in benign human prostatic epithelium. Cancer Res (2007) 67(9):4244–53. doi:10.1158/0008-5472.CAN-06-3946
92. Ishii K, Mizokami A, Tsunoda T, Iguchi K, Kato M, Hori Y, et al. Heterogenous induction of carcinoma-associated fibroblast-like differentiation in normal human prostatic fibroblasts by co-culturing with prostate cancer cells. J Cell Biochem (2011) 112(12):3604–11. doi:10.1002/jcb.23291
93. Webber J, Steadman R, Mason MD, Tabi Z, Clayton A. Cancer exosomes trigger fibroblast to myofibroblast differentiation. Cancer Res (2010) 70(23):9621–30. doi:10.1158/0008-5472.CAN-10-1722
94. Webber JP, Spary LK, Sanders AJ, Chowdhury R, Jiang WG, Steadman R, et al. Differentiation of tumour-promoting stromal myofibroblasts by cancer exosomes. Oncogene (2015) 34(3):290–302. doi:10.1038/onc.2013.560
95. Deep G, Panigrahi GK. Hypoxia-induced signaling promotes prostate cancer progression: exosomes role as messenger of hypoxic response in tumor microenvironment. Crit Rev Oncog (2015) 20(5–6):419–34. doi:10.1615/CritRevOncog.v20.i5-6.130
96. Ramteke A, Ting H, Agarwal C, Mateen S, Somasagara R, Hussain A, et al. Exosomes secreted under hypoxia enhance invasiveness and stemness of prostate cancer cells by targeting adherens junction molecules. Mol Carcinog (2015) 54(7):554–65. doi:10.1002/mc.22124
97. Schlaepfer IR, Nambiar DK, Ramteke A, Kumar R, Dhar D, Agarwal C, et al. Hypoxia induces triglycerides accumulation in prostate cancer cells and extracellular vesicles supporting growth and invasiveness following reoxygenation. Oncotarget (2015) 6(26):22836–56. doi:10.18632/oncotarget.4479
98. Sung SY, Hsieh CL, Wu D, Chung LW, Johnstone PA. Tumor microenvironment promotes cancer progression, metastasis, and therapeutic resistance. Curr Probl Cancer (2007) 31(2):36–100. doi:10.1016/j.currproblcancer.2006.12.002
99. Brennen WN, Chen S, Denmeade SR, Isaacs JT. Quantification of mesenchymal stem cells (MSCs) at sites of human prostate cancer. Oncotarget (2013) 4(1):106–17. doi:10.18632/oncotarget.805
100. Chowdhury R, Webber JP, Gurney M, Mason MD, Tabi Z, Clayton A. Cancer exosomes trigger mesenchymal stem cell differentiation into pro-angiogenic and pro-invasive myofibroblasts. Oncotarget (2015) 6(2):715–31. doi:10.18632/oncotarget.2711
101. Morello M, Minciacchi VR, de Candia P, Yang J, Posadas E, Kim H, et al. Large oncosomes mediate intercellular transfer of functional microRNA. Cell Cycle (2013) 12(22):3526–36. doi:10.4161/cc.26539
102. Minciacchi VR, Spinelli C, Reis-Sobreiro M, Cavallini L, You S, Zandian M, et al. MYC mediates large oncosome-induced fibroblast reprogramming in prostate cancer. Cancer Res (2017) 77(9):2306–17. doi:10.1158/0008-5472.CAN-16-2942
103. Castellana D, Zobairi F, Martinez MC, Panaro MA, Mitolo V, Freyssinet JM, et al. Membrane microvesicles as actors in the establishment of a favorable prostatic tumoral niche: a role for activated fibroblasts and CX3CL1-CX3CR1 axis. Cancer Res (2009) 69(3):785–93. doi:10.1158/0008-5472.CAN-08-1946
104. Bellezza I, Aisa MC, Palazzo R, Costanzi E, Mearini E, Minelli A. Extracellular matrix degrading enzymes at the prostasome surface. Prostate Cancer Prostatic Dis (2005) 8(4):344–8. doi:10.1038/sj.pcan.4500828
105. Kim CW, Lee HM, Lee TH, Kang C, Kleinman HK, Gho YS. Extracellular membrane vesicles from tumor cells promote angiogenesis via sphingomyelin. Cancer Res (2002) 62(21):6312–7.
106. Delves GH, Stewart AB, Lwaleed BA, Cooper AJ. In vitro inhibition of angiogenesis by prostasomes. Prostate Cancer Prostatic Dis (2005) 8(2):174–8. doi:10.1038/sj.pcan.4500790
107. Delves GH, Goyal A, Lwaleed BA, Cooper AJ. Seminal prostasomes inhibit the angiogenesis activity of rat aortic rings. Prostate Cancer Prostatic Dis (2006) 9(4):444–7. doi:10.1038/sj.pcan.4500896
108. Lundholm M, Schroder M, Nagaeva O, Baranov V, Widmark A, Mincheva-Nilsson L, et al. Prostate tumor-derived exosomes down-regulate NKG2D expression on natural killer cells and CD8+ T cells: mechanism of immune evasion. PLoS One (2014) 9(9):e108925. doi:10.1371/journal.pone.0108925
109. Salimu J, Webber J, Gurney M, Al-Taei S, Clayton A, Tabi Z. Dominant immunosuppression of dendritic cell function by prostate-cancer-derived exosomes. J Extracell Vesicles (2017) 6(1):1368823. doi:10.1080/20013078.2017.1368823
110. Kelly RW, Holland P, Skibinski G, Harrison C, McMillan L, Hargreave T, et al. Extracellular organelles (prostasomes) are immunosuppressive components of human semen. Clin Exp Immunol (1991) 86(3):550–6. doi:10.1111/j.1365-2249.1991.tb02968.x
111. Babiker AA, Ronquist G, Nilsson UR, Nilsson B. Transfer of prostasomal CD59 to CD59-deficient red blood cells results in protection against complement-mediated hemolysis. Am J Reprod Immunol (2002) 47(3):183–92. doi:10.1034/j.1600-0897.2002.1o023.x
112. Babiker AA, Nilsson B, Ronquist G, Carlsson L, Ekdahl KN. Transfer of functional prostasomal CD59 of metastatic prostatic cancer cell origin protects cells against complement attack. Prostate (2005) 62(2):105–14. doi:10.1002/pros.20102
113. Abd Elmageed ZY, Yang Y, Thomas R, Ranjan M, Mondal D, Moroz K, et al. Neoplastic reprogramming of patient-derived adipose stem cells by prostate cancer cell-associated exosomes. Stem Cells (2014) 32(4):983–97. doi:10.1002/stem.1619
114. Lugini L, Valtieri M, Federici C, Cecchetti S, Meschini S, Condello M, et al. Exosomes from human colorectal cancer induce a tumor-like behavior in colonic mesenchymal stromal cells. Oncotarget (2016) 7(31):50086–98. doi:10.18632/oncotarget.10574
115. Vlaeminck-Guillem V. When prostate cancer circulates in the bloodstream. Diagnostics (Basel) (2015) 5(4):428–74. doi:10.3390/diagnostics5040428
116. Hegemann M, Maas M, Rausch S, Walz S, Bedke J, Stenzl A, et al. Current concepts and trends in the treatment of bone metastases in patients with advanced prostate cancer. Asian J Androl (2017). doi:10.4103/aja.aja_59_17
117. Cook LM, Shay G, Araujo A, Lynch CC. Integrating new discoveries into the “vicious cycle” paradigm of prostate to bone metastases. Cancer Metastasis Rev (2014) 33(2–3):511–25. doi:10.1007/s10555-014-9494-4
118. Itoh T, Ito Y, Ohtsuki Y, Ando M, Tsukamasa Y, Yamada N, et al. Microvesicles released from hormone-refractory prostate cancer cells facilitate mouse pre-osteoblast differentiation. J Mol Histol (2012) 43(5):509–15. doi:10.1007/s10735-012-9415-1
119. Karlsson T, Lundholm M, Widmark A, Persson E. Tumor cell-derived exosomes from the prostate cancer cell line TRAMP-C1 impair osteoclast formation and differentiation. PLoS One (2016) 11(11):e0166284. doi:10.1371/journal.pone.0166284
120. Ye Y, Li SL, Ma YY, Diao YJ, Yang L, Su MQ, et al. Exosomal miR-141-3p regulates osteoblast activity to promote the osteoblastic metastasis of prostate cancer. Oncotarget (2017) 8(55):94834–49. doi:10.18632/oncotarget.22014
121. Hu CD, Choo R, Huang J. Neuroendocrine differentiation in prostate cancer: a mechanism of radioresistance and treatment failure. Front Oncol (2015) 5:90. doi:10.3389/fonc.2015.00090
122. Vlachostergios PJ, Papandreou CN. Targeting neuroendocrine prostate cancer: molecular and clinical perspectives. Front Oncol (2015) 5:6. doi:10.3389/fonc.2015.00006
123. Hagberg Thulin M, Jennbacken K, Damber JE, Welen K. Osteoblasts stimulate the osteogenic and metastatic progression of castration-resistant prostate cancer in a novel model for in vitro and in vivo studies. Clin Exp Metastasis (2014) 31(3):269–83. doi:10.1007/s10585-013-9626-1
124. Koeneman KS, Yeung F, Chung LW. Osteomimetic properties of prostate cancer cells: a hypothesis supporting the predilection of prostate cancer metastasis and growth in the bone environment. Prostate (1999) 39(4):246–61. doi:10.1002/(SICI)1097-0045(19990601)39:4<246::AID-PROS5>3.0.CO;2-U
125. Renzulli JF II, Del Tatto M, Dooner G, Aliotta J, Goldstein L, Dooner M, et al. Microvesicle induction of prostate specific gene expression in normal human bone marrow cells. J Urol (2010) 184(5):2165–71. doi:10.1016/j.juro.2010.06.119
126. Josson S, Gururajan M, Sung SY, Hu P, Shao C, Zhau HE, et al. Stromal fibroblast-derived miR-409 promotes epithelial-to-mesenchymal transition and prostate tumorigenesis. Oncogene (2015) 34(21):2690–9. doi:10.1038/onc.2014.212
127. Josson S, Gururajan M, Hu P, Shao C, Chu GY, Zhau HE, et al. miR-409-3p/-5p promotes tumorigenesis, epithelial-to-mesenchymal transition, and bone metastasis of human prostate cancer. Clin Cancer Res (2014) 20(17):4636–46. doi:10.1158/1078-0432.CCR-14-0305
128. Alcayaga-Miranda F, Gonzalez PL, Lopez-Verrilli A, Varas-Godoy M, Aguila-Diaz C, Contreras L, et al. Prostate tumor-induced angiogenesis is blocked by exosomes derived from menstrual stem cells through the inhibition of reactive oxygen species. Oncotarget (2016) 7(28):44462–77. doi:10.18632/oncotarget.9852
129. Morhayim J, van de Peppel J, Demmers JA, Kocer G, Nigg AL, van Driel M, et al. Proteomic signatures of extracellular vesicles secreted by nonmineralizing and mineralizing human osteoblasts and stimulation of tumor cell growth. FASEB J (2015) 29(1):274–85. doi:10.1096/fj.14-261404
130. Ramachandran S, Palanisamy V. Horizontal transfer of RNAs: exosomes as mediators of intercellular communication. Wiley Interdiscip Rev RNA (2012) 3(2):286–93. doi:10.1002/wrna.115
131. Halin Bergstrom S, Hagglof C, Thysell E, Bergh A, Wikstrom P, Lundholm M. Extracellular vesicles from metastatic rat prostate tumors prime the normal prostate tissue to facilitate tumor growth. Sci Rep (2016) 6:31805. doi:10.1038/srep31805
132. Souza AG, IB BS, Campos-Fernandez E, Marangoni K, VA FB, Alves PT, et al. Extracellular vesicles as drivers of epithelial-mesenchymal transition and carcinogenic characteristics in normal prostate cells. Mol Carcinog (2017) 57(4):503–11. doi:10.1002/mc.22775
133. Hosseini-Beheshti E, Choi W, Weiswald LB, Kharmate G, Ghaffari M, Roshan-Moniri M, et al. Exosomes confer pro-survival signals to alter the phenotype of prostate cells in their surrounding environment. Oncotarget (2016) 7(12):14639–58. doi:10.18632/oncotarget.7052
134. Waugh DJ, Wilson C. The interleukin-8 pathway in cancer. Clin Cancer Res (2008) 14(21):6735–41. doi:10.1158/1078-0432.CCR-07-4843
135. Fedele C, Singh A, Zerlanko BJ, Iozzo RV, Languino LR. The alphavbeta6 integrin is transferred intercellularly via exosomes. J Biol Chem (2015) 290(8):4545–51. doi:10.1074/jbc.C114.617662
136. Singh A, Fedele C, Lu H, Nevalainen MT, Keen JH, Languino LR. Exosome-mediated transfer of alphavbeta3 integrin from tumorigenic to nontumorigenic cells promotes a migratory phenotype. Mol Cancer Res (2016) 14(11):1136–46. doi:10.1158/1541-7786.MCR-16-0058
137. Soekmadji C, Riches JD, Russell PJ, Ruelcke JE, McPherson S, Wang C, et al. Modulation of paracrine signaling by CD9 positive small extracellular vesicles mediates cellular growth of androgen deprived prostate cancer. Oncotarget (2017) 8(32):52237–55. doi:10.18632/oncotarget.11111
138. Soekmadji C, Rockstroh A, Ramm GA, Nelson CC, Russell PJ. Extracellular vesicles in the adaptive process of prostate cancer during inhibition of androgen receptor signaling by enzalutamide. Proteomics (2017) 17(23–24). doi:10.1002/pmic.201600427
139. Read J, Ingram A, Al Saleh HA, Platko K, Gabriel K, Kapoor A, et al. Nuclear transportation of exogenous epidermal growth factor receptor and androgen receptor via extracellular vesicles. Eur J Cancer (2017) 70:62–74. doi:10.1016/j.ejca.2016.10.017
140. El-Sayed IY, Daher A, Destouches D, Firlej V, Kostallari E, Maille P, et al. Extracellular vesicles released by mesenchymal-like prostate carcinoma cells modulate EMT state of recipient epithelial-like carcinoma cells through regulation of AR signaling. Cancer Lett (2017) 410:100–11. doi:10.1016/j.canlet.2017.09.010
141. Lin LC, Gao AC, Lai CH, Hsieh JT, Lin H. Induction of neuroendocrine differentiation in castration resistant prostate cancer cells by adipocyte differentiation-related protein (ADRP) delivered by exosomes. Cancer Lett (2017) 391:74–82. doi:10.1016/j.canlet.2017.01.018
142. Gabriel K, Ingram A, Austin R, Kapoor A, Tang D, Majeed F, et al. Regulation of the tumor suppressor PTEN through exosomes: a diagnostic potential for prostate cancer. PLoS One (2013) 8(7):e70047. doi:10.1371/journal.pone.0070047
143. Putz U, Howitt J, Doan A, Goh CP, Low LH, Silke J, et al. The tumor suppressor PTEN is exported in exosomes and has phosphatase activity in recipient cells. Sci Signal (2012) 5(243):ra70. doi:10.1126/scisignal.2003084
144. Shedden K, Xie XT, Chandaroy P, Chang YT, Rosania GR. Expulsion of small molecules in vesicles shed by cancer cells: association with gene expression and chemosensitivity profiles. Cancer Res (2003) 63(15):4331–7.
145. Kosgodage US, Trindade RP, Thompson PR, Inal JM, Lange S. Chloramidine/bisindolylmaleimide-I-mediated inhibition of exosome and microvesicle release and enhanced efficacy of cancer chemotherapy. Int J Mol Sci (2017) 18(5):E1007. doi:10.3390/ijms18051007
146. Corcoran C, Rani S, O’Brien K, O’Neill A, Prencipe M, Sheikh R, et al. Docetaxel-resistance in prostate cancer: evaluating associated phenotypic changes and potential for resistance transfer via exosomes. PLoS One (2012) 7(12):e50999. doi:10.1371/journal.pone.0050999
147. Panagopoulos K, Cross-Knorr S, Dillard C, Pantazatos D, Del Tatto M, Mills D, et al. Reversal of chemosensitivity and induction of cell malignancy of a non-malignant prostate cancer cell line upon extracellular vesicle exposure. Mol Cancer (2013) 12(1):118. doi:10.1186/1476-4598-12-118
148. Kharaziha P, Chioureas D, Rutishauser D, Baltatzis G, Lennartsson L, Fonseca P, et al. Molecular profiling of prostate cancer derived exosomes may reveal a predictive signature for response to docetaxel. Oncotarget (2015) 6(25):21740–54. doi:10.18632/oncotarget.3226
149. Wang JQ, DeChalus A, Chatterjee DN, Keller ET, Mizokami A, Camussi G, et al. Extracellular vesicle-mediated reversal of paclitaxel resistance in prostate cancer. Crit Rev Oncog (2015) 20(5–6):407–17. doi:10.1615/CritRevOncog.v20.i5-6.120
150. Kawakami K, Fujita Y, Kato T, Mizutani K, Kameyama K, Tsumoto H, et al. Integrin beta4 and vinculin contained in exosomes are potential markers for progression of prostate cancer associated with taxane-resistance. Int J Oncol (2015) 47(1):384–90. doi:10.3892/ijo.2015.3011
151. Kato T, Mizutani K, Kameyama K, Kawakami K, Fujita Y, Nakane K, et al. Serum exosomal P-glycoprotein is a potential marker to diagnose docetaxel resistance and select a taxoid for patients with prostate cancer. Urol Oncol (2015) 33(9):385.e15–20. doi:10.1016/j.urolonc.2015.04.019
152. Corcoran C, Rani S, O’Driscoll L. miR-34a is an intracellular and exosomal predictive biomarker for response to docetaxel with clinical relevance to prostate cancer progression. Prostate (2014) 74(13):1320–34. doi:10.1002/pros.22848
153. Lucotti S, Rainaldi G, Evangelista M, Rizzo M. Fludarabine treatment favors the retention of miR-485-3p by prostate cancer cells: implications for survival. Mol Cancer (2013) 12(1):52. doi:10.1186/1476-4598-12-52
154. Lehmann BD, McCubrey JA, Terrian DM. Radiosensitization of prostate cancer by priming the wild-type p53-dependent cellular senescence pathway. Cancer Biol Ther (2007) 6(8):1165–70. doi:10.4161/cbt.6.8.4544
155. Lehmann BD, Paine MS, Brooks AM, McCubrey JA, Renegar RH, Wang R, et al. Senescence-associated exosome release from human prostate cancer cells. Cancer Res (2008) 68(19):7864–71. doi:10.1158/0008-5472.CAN-07-6538
156. Malla B, Zaugg K, Vassella E, Aebersold DM, Dal Pra A. Exosomes and exosomal microRNAs in prostate cancer radiation therapy. Int J Radiat Oncol Biol Phys (2017) 98(5):982–95. doi:10.1016/j.ijrobp.2017.03.031
157. Jelonek K, Widlak P, Pietrowska M. The influence of ionizing radiation on exosome composition, secretion and intercellular communication. Protein Pept Lett (2016) 23(7):656–63. doi:10.2174/0929866523666160427105138
158. Yu X, Harris SL, Levine AJ. The regulation of exosome secretion: a novel function of the p53 protein. Cancer Res (2006) 66(9):4795–801. doi:10.1158/0008-5472.CAN-05-4579
159. Hurwitz MD, Kaur P, Nagaraja GM, Bausero MA, Manola J, Asea A. Radiation therapy induces circulating serum Hsp72 in patients with prostate cancer. Radiother Oncol (2010) 95(3):350–8. doi:10.1016/j.radonc.2010.03.024
160. Benzon B, Zhao SG, Haffner MC, Takhar M, Erho N, Yousefi K, et al. Correlation of B7-H3 with androgen receptor, immune pathways and poor outcome in prostate cancer: an expression-based analysis. Prostate Cancer Prostatic Dis (2017) 20(1):28–35. doi:10.1038/pcan.2016.49
161. Parker AS, Heckman MG, Sheinin Y, Wu KJ, Hilton TW, Diehl NN, et al. Evaluation of B7-H3 expression as a biomarker of biochemical recurrence after salvage radiation therapy for recurrent prostate cancer. Int J Radiat Oncol Biol Phys (2011) 79(5):1343–9. doi:10.1016/j.ijrobp.2010.01.061
162. Chavin G, Sheinin Y, Crispen PL, Boorjian SA, Roth TJ, Rangel L, et al. Expression of immunosuppresive B7-H3 ligand by hormone-treated prostate cancer tumors and metastases. Clin Cancer Res (2009) 15(6):2174–80. doi:10.1158/1078-0432.CCR-08-2262
163. Zang X, Thompson RH, Al-Ahmadie HA, Serio AM, Reuter VE, Eastham JA, et al. B7-H3 and B7x are highly expressed in human prostate cancer and associated with disease spread and poor outcome. Proc Natl Acad Sci U S A (2007) 104(49):19458–63. doi:10.1073/pnas.0709802104
164. Roth TJ, Sheinin Y, Lohse CM, Kuntz SM, Frigola X, Inman BA, et al. B7-H3 ligand expression by prostate cancer: a novel marker of prognosis and potential target for therapy. Cancer Res (2007) 67(16):7893–900. doi:10.1158/0008-5472.CAN-07-1068
165. Hazawa M, Tomiyama K, Saotome-Nakamura A, Obara C, Yasuda T, Gotoh T, et al. Radiation increases the cellular uptake of exosomes through CD29/CD81 complex formation. Biochem Biophys Res Commun (2014) 446(4):1165–71. doi:10.1016/j.bbrc.2014.03.067
166. Arscott WT, Tandle AT, Zhao S, Shabason JE, Gordon IK, Schlaff CD, et al. Ionizing radiation and glioblastoma exosomes: implications in tumor biology and cell migration. Transl Oncol (2013) 6(6):638–48. doi:10.1593/tlo.13640
167. Vlaeminck-Guillem V, Gillet G, Rimokh R. SRC: marker or actor in prostate cancer aggressiveness. Front Oncol (2014) 4:222. doi:10.3389/fonc.2014.00222
168. DeRita RM, Zerlanko B, Singh A, Lu H, Iozzo RV, Benovic JL, et al. c-Src, insulin-like growth factor I receptor, G-protein-coupled receptor kinases and focal adhesion kinase are enriched into prostate cancer cell exosomes. J Cell Biochem (2017) 118(1):66–73. doi:10.1002/jcb.25611
169. Najafi M, Fardid R, Hadadi G, Fardid M. The mechanisms of radiation-induced bystander effect. J Biomed Phys Eng (2014) 4(4):163–72.
170. Yahyapour R, Motevaseli E, Rezaeyan A, Abdollahi H, Farhood B, Cheki M, et al. Reduction-oxidation (redox) system in radiation-induced normal tissue injury: molecular mechanisms and implications in radiation therapeutics. Clin Transl Oncol (2018). doi:10.1007/s12094-017-1828-6
171. Al-Mayah AH, Irons SL, Pink RC, Carter DR, Kadhim MA. Possible role of exosomes containing RNA in mediating nontargeted effect of ionizing radiation. Radiat Res (2012) 177(5):539–45. doi:10.1667/RR2868.1
172. Al-Mayah A, Bright S, Chapman K, Irons S, Luo P, Carter D, et al. The non-targeted effects of radiation are perpetuated by exosomes. Mutat Res (2015) 772:38–45. doi:10.1016/j.mrfmmm.2014.12.007
173. Jella KK, Rani S, O’Driscoll L, McClean B, Byrne HJ, Lyng FM. Exosomes are involved in mediating radiation induced bystander signaling in human keratinocyte cells. Radiat Res (2014) 181(2):138–45. doi:10.1667/RR13337.1
174. Song M, Wang Y, Shang ZF, Liu XD, Xie DF, Wang Q, et al. Bystander autophagy mediated by radiation-induced exosomal miR-7-5p in non-targeted human bronchial epithelial cells. Sci Rep (2016) 6:30165. doi:10.1038/srep30165
175. Cai S, Shi GS, Cheng HY, Zeng YN, Li G, Zhang M, et al. Exosomal miR-7 mediates bystander autophagy in lung after focal brain irradiation in mice. Int J Biol Sci (2017) 13(10):1287–96. doi:10.7150/ijbs.18890
176. Gong P, Zhang T, He D, Hsieh JT. MicroRNA-145 modulates tumor sensitivity to radiation in prostate cancer. Radiat Res (2015) 184(6):630–8. doi:10.1667/RR14185.1
177. Logozzi M, De Milito A, Lugini L, Borghi M, Calabro L, Spada M, et al. High levels of exosomes expressing CD63 and caveolin-1 in plasma of melanoma patients. PLoS One (2009) 4(4):e5219. doi:10.1371/journal.pone.0005219
178. Mitchell PJ, Welton J, Staffurth J, Court J, Mason MD, Tabi Z, et al. Can urinary exosomes act as treatment response markers in prostate cancer? J Transl Med (2009) 7:4. doi:10.1186/1479-5876-7-4
179. Dijkstra S, Birker IL, Smit FP, Leyten GH, de Reijke TM, van Oort IM, et al. Prostate cancer biomarker profiles in urinary sediments and exosomes. J Urol (2014) 191(4):1132–8. doi:10.1016/j.juro.2013.11.001
180. Pellegrini KL, Patil D, Douglas KJS, Lee G, Wehrmeyer K, Torlak M, et al. Detection of prostate cancer-specific transcripts in extracellular vesicles isolated from post-DRE urine. Prostate (2017) 77(9):990–9. doi:10.1002/pros.23355
181. Park YH, Shin HW, Jung AR, Kwon OS, Choi YJ, Park J, et al. Prostate-specific extracellular vesicles as a novel biomarker in human prostate cancer. Sci Rep (2016) 6:30386. doi:10.1038/srep30386
182. Tavoosidana G, Ronquist G, Darmanis S, Yan J, Carlsson L, Wu D, et al. Multiple recognition assay reveals prostasomes as promising plasma biomarkers for prostate cancer. Proc Natl Acad Sci U S A (2011) 108(21):8809–14. doi:10.1073/pnas.1019330108
183. Biggs CN, Siddiqui KM, Al-Zahrani AA, Pardhan S, Brett SI, Guo QQ, et al. Prostate extracellular vesicles in patient plasma as a liquid biopsy platform for prostate cancer using nanoscale flow cytometry. Oncotarget (2016) 7(8):8839–49. doi:10.18632/oncotarget.6983
184. Logozzi M, Angelini DF, Iessi E, Mizzoni D, Di Raimo R, Federici C, et al. Increased PSA expression on prostate cancer exosomes in in vitro condition and in cancer patients. Cancer Lett (2017) 403:318–29. doi:10.1016/j.canlet.2017.06.036
185. Lance RS, Drake RR, Troyer DA. Multiple recognition assay reveals prostasomes as promising plasma biomarkers for prostate cancer. Expert Rev Anticancer Ther (2011) 11(9):1341–3. doi:10.1586/era.11.134
186. Khan S, Simpson J, Lynch JC, Turay D, Mirshahidi S, Gonda A, et al. Racial differences in the expression of inhibitors of apoptosis (IAP) proteins in extracellular vesicles (EV) from prostate cancer patients. PLoS One (2017) 12(10):e0183122. doi:10.1371/journal.pone.0183122
187. Khan S, Jutzy JM, Valenzuela MM, Turay D, Aspe JR, Ashok A, et al. Plasma-derived exosomal survivin, a plausible biomarker for early detection of prostate cancer. PLoS One (2012) 7(10):e46737. doi:10.1371/journal.pone.0046737
188. Liu T, Mendes DE, Berkman CE. Functional prostate-specific membrane antigen is enriched in exosomes from prostate cancer cells. Int J Oncol (2014) 44(3):918–22. doi:10.3892/ijo.2014.2256
189. Kharmate G, Hosseini-Beheshti E, Caradec J, Chin MY, Tomlinson Guns ES. Epidermal growth factor receptor in prostate cancer derived exosomes. PLoS One (2016) 11(5):e0154967. doi:10.1371/journal.pone.0154967
190. Vermassen T, D’Herde K, Jacobus D, Van Praet C, Poelaert F, Lumen N, et al. Release of urinary extracellular vesicles in prostate cancer is associated with altered urinary N-glycosylation profile. J Clin Pathol (2017) 70(10):838–46. doi:10.1136/jclinpath-2016-204312
191. Wang L, Skotland T, Berge V, Sandvig K, Llorente A. Exosomal proteins as prostate cancer biomarkers in urine: from mass spectrometry discovery to immunoassay-based validation. Eur J Pharm Sci (2017) 98:80–5. doi:10.1016/j.ejps.2016.09.023
192. Hessels D, Klein Gunnewiek JM, van Oort I, Karthaus HF, van Leenders GJ, van Balken B, et al. DD3(PCA3)-based molecular urine analysis for the diagnosis of prostate cancer. Eur Urol (2003) 44(1):8–15; discussion 15–6. doi:10.1016/S0302-2838(03)00201-X
193. McKiernan J, Donovan MJ, O’Neill V, Bentink S, Noerholm M, Belzer S, et al. A novel urine exosome gene expression assay to predict high-grade prostate cancer at initial biopsy. JAMA Oncol (2016) 2(7):882–9. doi:10.1001/jamaoncol.2016.0097
194. Hendriks RJ, Dijkstra S, Jannink SA, Steffens MG, van Oort IM, Mulders PF, et al. Comparative analysis of prostate cancer specific biomarkers PCA3 and ERG in whole urine, urinary sediments and exosomes. Clin Chem Lab Med (2016) 54(3):483–92. doi:10.1515/cclm-2015-0599
195. Isin M, Uysaler E, Ozgur E, Koseoglu H, Sanli O, Yucel OB, et al. Exosomal lncRNA-p21 levels may help to distinguish prostate cancer from benign disease. Front Genet (2015) 6:168. doi:10.3389/fgene.2015.00168
196. Motamedinia P, Scott AN, Bate KL, Sadeghi N, Salazar G, Shapiro E, et al. Urine exosomes for non-invasive assessment of gene expression and mutations of prostate cancer. PLoS One (2016) 11(5):e0154507. doi:10.1371/journal.pone.0154507
197. Turchinovich A, Weiz L, Langheinz A, Burwinkel B. Characterization of extracellular circulating microRNA. Nucleic Acids Res (2011) 39(16):7223–33. doi:10.1093/nar/gkr254
198. Arroyo JD, Chevillet JR, Kroh EM, Ruf IK, Pritchard CC, Gibson DF, et al. Argonaute2 complexes carry a population of circulating microRNAs independent of vesicles in human plasma. Proc Natl Acad Sci U S A (2011) 108(12):5003–8. doi:10.1073/pnas.1019055108
199. Gallo A, Tandon M, Alevizos I, Illei GG. The majority of microRNAs detectable in serum and saliva is concentrated in exosomes. PLoS One (2012) 7(3):e30679. doi:10.1371/journal.pone.0030679
200. Cheng L, Sun X, Scicluna BJ, Coleman BM, Hill AF. Characterization and deep sequencing analysis of exosomal and non-exosomal miRNA in human urine. Kidney Int (2014) 86(2):433–44. doi:10.1038/ki.2013.502
201. Endzelins E, Melne V, Kalnina Z, Lietuvietis V, Riekstina U, Llorente A, et al. Diagnostic, prognostic and predictive value of cell-free miRNAs in prostate cancer: a systematic review. Mol Cancer (2016) 15(1):41. doi:10.1186/s12943-016-0523-5
202. Filella X, Foj L. miRNAs as novel biomarkers in the management of prostate cancer. Clin Chem Lab Med (2017) 55(5):715–36. doi:10.1515/cclm-2015-1073
203. Luu HN, Lin HY, Sorensen KD, Ogunwobi OO, Kumar N, Chornokur G, et al. miRNAs associated with prostate cancer risk and progression. BMC Urol (2017) 17(1):18. doi:10.1186/s12894-017-0206-6
204. Donovan MJ, Noerholm M, Bentink S, Belzer S, Skog J, O’Neill V, et al. A molecular signature of PCA3 and ERG exosomal RNA from non-DRE urine is predictive of initial prostate biopsy result. Prostate Cancer Prostatic Dis (2015) 18(4):370–5. doi:10.1038/pcan.2015.40
205. Huang X, Yuan T, Liang M, Du M, Xia S, Dittmar R, et al. Exosomal miR-1290 and miR-375 as prognostic markers in castration-resistant prostate cancer. Eur Urol (2015) 67(1):33–41. doi:10.1016/j.eururo.2014.07.035
206. Bryzgunova OE, Zaripov MM, Skvortsova TE, Lekchnov EA, Grigor’eva AE, Zaporozhchenko IA, et al. Comparative study of extracellular vesicles from the urine of healthy individuals and prostate cancer patients. PLoS One (2016) 11(6):e0157566. doi:10.1371/journal.pone.0157566
207. Koppers-Lalic D, Hackenberg M, de Menezes R, Misovic B, Wachalska M, Geldof A, et al. Noninvasive prostate cancer detection by measuring miRNA variants (isomiRs) in urine extracellular vesicles. Oncotarget (2016) 7(16):22566–78. doi:10.18632/oncotarget.8124
208. Li Z, Ma YY, Wang J, Zeng XF, Li R, Kang W, et al. Exosomal microRNA-141 is upregulated in the serum of prostate cancer patients. Onco Targets Ther (2016) 9:139–48. doi:10.2147/OTT.S95565
209. Endzelins E, Berger A, Melne V, Bajo-Santos C, Sobolevska K, Abols A, et al. Detection of circulating miRNAs: comparative analysis of extracellular vesicle-incorporated miRNAs and cell-free miRNAs in whole plasma of prostate cancer patients. BMC Cancer (2017) 17(1):730. doi:10.1186/s12885-017-3737-z
210. Rodriguez M, Bajo-Santos C, Hessvik NP, Lorenz S, Fromm B, Berge V, et al. Identification of non-invasive miRNAs biomarkers for prostate cancer by deep sequencing analysis of urinary exosomes. Mol Cancer (2017) 16(1):156. doi:10.1186/s12943-017-0726-4
211. Wani S, Kaul D, Mavuduru RS, Kakkar N, Bhatia A. Urinary-exosomal miR-2909: a novel pathognomonic trait of prostate cancer severity. J Biotechnol (2017) 259:135–9. doi:10.1016/j.jbiotec.2017.07.029
212. Foj L, Ferrer F, Serra M, Arevalo A, Gavagnach M, Gimenez N, et al. Exosomal and non-exosomal urinary miRNAs in prostate cancer detection and prognosis. Prostate (2017) 77(6):573–83. doi:10.1002/pros.23295
213. Xu Y, Qin S, An T, Tang Y, Huang Y, Zheng L. MiR-145 detection in urinary extracellular vesicles increase diagnostic efficiency of prostate cancer based on hydrostatic filtration dialysis method. Prostate (2017) 77(10):1167–75. doi:10.1002/pros.23376
214. Bhagirath D, Yang TL, Bucay N, Sekhon K, Majid S, Shahryari V, et al. MicroRNA-1246 is an exosomal biomarker for aggressive prostate cancer. Cancer Res (2018) 78(7):1833–44. doi:10.1158/0008-5472.CAN-17-2069
215. Brase JC, Johannes M, Schlomm T, Falth M, Haese A, Steuber T, et al. Circulating miRNAs are correlated with tumor progression in prostate cancer. Int J Cancer (2011) 128(3):608–16. doi:10.1002/ijc.25376
216. Korzeniewski N, Tosev G, Pahernik S, Hadaschik B, Hohenfellner M, Duensing S. Identification of cell-free microRNAs in the urine of patients with prostate cancer. Urol Oncol (2015) 33(1):16.e17–22. doi:10.1016/j.urolonc.2014.09.015
218. Yu Q, Li P, Weng M, Wu S, Zhang Y, Chen X, et al. Nano-vesicles are a potential tool to monitor therapeutic efficacy of carbon ion radiotherapy in prostate cancer. J Biomed Nanotechnol (2018) 14(1):168–78. doi:10.1166/jbn.2018.2503
219. Hu R, Dunn TA, Wei S, Isharwal S, Veltri RW, Humphreys E, et al. Ligand-independent androgen receptor variants derived from splicing of cryptic exons signify hormone-refractory prostate cancer. Cancer Res (2009) 69(1):16–22. doi:10.1158/0008-5472.CAN-08-2764
220. Antonarakis ES, Lu C, Wang H, Luber B, Nakazawa M, Roeser JC, et al. AR-V7 and resistance to enzalutamide and abiraterone in prostate cancer. N Engl J Med (2014) 371(11):1028–38. doi:10.1056/NEJMoa1315815
221. Del Re M, Biasco E, Crucitta S, Derosa L, Rofi E, Orlandini C, et al. The detection of androgen receptor splice variant 7 in plasma-derived exosomal RNA strongly predicts resistance to hormonal therapy in metastatic prostate cancer patients. Eur Urol (2017) 71(4):680–7. doi:10.1016/j.eururo.2016.08.012
222. Brett SI, Lucien F, Guo C, Williams KC, Kim Y, Durfee PN, et al. Immunoaffinity based methods are superior to kits for purification of prostate derived extracellular vesicles from plasma samples. Prostate (2017) 77(13):1335–43. doi:10.1002/pros.23393
223. Yasui T, Yanagida T, Ito S, Konakade Y, Takeshita D, Naganawa T, et al. Unveiling massive numbers of cancer-related urinary-microRNA candidates via nanowires. Sci Adv (2017) 3(12):e1701133. doi:10.1126/sciadv.1701133
224. Bijnsdorp IV, Maxouri O, Kardar A, Schelfhorst T, Piersma SR, Pham TV, et al. Feasibility of urinary extracellular vesicle proteome profiling using a robust and simple, clinically applicable isolation method. J Extracell Vesicles (2017) 6(1):1313091. doi:10.1080/20013078.2017.1313091
225. Lener T, Gimona M, Aigner L, Borger V, Buzas E, Camussi G, et al. Applying extracellular vesicles based therapeutics in clinical trials – an ISEV position paper. J Extracell Vesicles (2015) 4:30087. doi:10.3402/jev.v4.30087
226. Fais S, O’Driscoll L, Borras FE, Buzas E, Camussi G, Cappello F, et al. Evidence-based clinical use of nanoscale extracellular vesicles in nanomedicine. ACS Nano (2016) 10(4):3886–99. doi:10.1021/acsnano.5b08015
227. Zhang G, Guo B, Wu H, Tang T, Zhang BT, Zheng L, et al. A delivery system targeting bone formation surfaces to facilitate RNAi-based anabolic therapy. Nat Med (2012) 18(2):307–14. doi:10.1038/nm.2617
228. Saari H, Lazaro-Ibanez E, Viitala T, Vuorimaa-Laukkanen E, Siljander P, Yliperttula M. Microvesicle- and exosome-mediated drug delivery enhances the cytotoxicity of Paclitaxel in autologous prostate cancer cells. J Control Release (2015) 220(Pt B):727–37. doi:10.1016/j.jconrel.2015.09.031
Keywords: extracellular vesicles, prostate cancer, microenvironment, diagnosis, theranostic, exosomes
Citation: Vlaeminck-Guillem V (2018) Extracellular Vesicles in Prostate Cancer Carcinogenesis, Diagnosis, and Management. Front. Oncol. 8:222. doi: 10.3389/fonc.2018.00222
Received: 09 December 2017; Accepted: 29 May 2018;
Published: 13 June 2018
Edited by:
Stéphane Terry, Institut Gustave Roussy, FranceReviewed by:
Stefano Fais, Istituto Superiore di Sanità, ItalyCopyright: © 2018 Vlaeminck-Guillem. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Virginie Vlaeminck-Guillem, dmlyZ2luaWUudmxhZW1pbmNrLWd1aWxsZW1AdW5pdi1seW9uMS5mcg==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.