 Yuki Ito
Yuki Ito Sayaka Nagamoto
Sayaka Nagamoto Tetsuya Takano
Tetsuya Takano
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
MINI REVIEW article
Front. Mol. Neurosci., 25 April 2024
Sec. Neuroplasticity and Development
Volume 17 - 2024 | https://doi.org/10.3389/fnmol.2024.1361956
This article is part of the Research TopicSpatio-temporal Molecular Mechanisms Regulating Synapse Function and Neural Circuit DynamicsView all 8 articles
Synapses play a pivotal role in forming neural circuits, with critical implications for brain functions such as learning, memory, and emotions. Several advances in synaptic research have demonstrated the diversity of synaptic structure and function, which can form thousands of connections depending on the neuronal cell types. Moreover, synapses not only interconnect neurons but also establish connections with glial cells such as astrocytes, which play a key role in the architecture and function of neuronal circuits in the brain. Emerging evidence suggests that dysfunction of synaptic proteins contributes to a variety of neurological and psychiatric disorders. Therefore, it is crucial to determine the molecular networks within synapses in various neuronal cell types to gain a deeper understanding of how the nervous system regulates brain function. Recent advances in synaptic proteome approaches, such as fluorescence-activated synaptosome sorting (FASS) and proximity labeling, have allowed for a detailed and spatial analysis of many cell-type-specific synaptic molecules in vivo. In this brief review, we highlight these novel spatial proteomic approaches and discuss the regulation of synaptic formation and function in the brain. This knowledge of molecular networks provides new insight into the understanding of many neurological and psychiatric disorders.
A human brain consists of approximately 86 billion neurons, interconnected through an intricate network of around 100 trillion synapses. In addition to neurons, glial cells such as astrocytes are involved in the formation of synapses, particularly evident in structures termed “tripartite synapses.” These cellular networks are crucial for the establishment of neural circuits underlying highly complex brain functions such as memory, learning, and emotion. Despite their incredibly small volume within neural cells, synapses play a key role in determining neuronal functionality. These synapses exhibit diverse functions as well as shapes, depending on the composition of thousands of proteins that vary across different neuronal types. Importantly, recent advancements in gene profiling have demonstrated that the abnormality of the synaptic protein is highly associated with psychiatric and neurological disorders, thus defining it as a “synaptopathies” (Lepeta et al., 2016; Luo et al., 2018; Yoshino et al., 2021). Thus, it is crucial to identify spatial molecular networks within synapses across specific neuronal cell types to understand how the nervous system governs the variety of brain functions. However, it remains a significant challenge to comprehend how a wide variety of synaptic molecules control each neuronal connectivity and functioning in the brain.
Over the past several decades, numerous researchers have attempted to isolate synapses to uncover the intricate functions orchestrated by synaptic molecules. Synaptic molecules have been isolated and analyzed using a variety of biochemical approaches such as cell fractionation, density gradient centrifugation (e.g., synaptosomes), immunoprecipitation, and affinity chromatography followed by liquid chromatography–tandem mass spectrometry (LC–MS/MS). These proteomic analyses have shed light on the components of synaptic proteins, as demonstrated by a comprehensive study that identified 2,876 proteins across 41 in vivo interactomes from mouse cerebral cortex, revealing the extensive landscape of the core-scaffold machinery within the postsynaptic density (Li et al., 2017). In particular, the identification of several key components of synaptic molecules, such as the NMDA receptor complex and PSD95 complex, has emerged as crucial discoveries in current synaptic research (Husi et al., 2000; Dosemeci et al., 2007; Fernández et al., 2009; Delint-Ramirez et al., 2010; Xu et al., 2021). Other major synaptic protein complexes were later elucidated, including SHANKs (Lee et al., 2017), SynGAP (Krapivinsky et al., 2004), PSD-93 (Zhang et al., 2020), FMRP (Schenck et al., 2001; Pasciuto and Bagni, 2014; Zhang et al., 2019), and metabotropic glutamate receptors (Farr et al., 2004; Francesconi et al., 2009; Ramos et al., 2012; Pandya et al., 2016). Additionally, further research has revealed that protein networks of small GTPase proteins control synapse formation (Wilkinson et al., 2017). Despite the effectiveness of these approaches, deciphering the molecular composition of synapses at the cellular level in vivo has been technically challenging due to the inability to maintain the spatial resolution indicating their originating cell types. In recent years, innovative approaches such as fluorescence-activated synaptosome sorting (FASS) and proximity labeling (BioID and APEX) approaches have been developed to facilitate the analysis of the spatial proteome, unveiling the cell-type-specific molecular composition within specific subcellular sites such as synapse and synaptic cleft in vivo (Biesemann et al., 2014; Han et al., 2018; Takano and Soderling, 2021). Additionally, we have recently developed novel in vivo cell-surface BioID approaches, namely TurboID-surface and Split-TurboID. These techniques have proven highly effective in elucidating the molecular network of cell-type-specific connections such as neurons and astrocytes (Takano et al., 2020). Here, this mini-review emphasizes the application of cutting-edge spatial proteomic approaches to investigate the molecular networks of cell-type-specific synapse compartments in the brain. Additionally, we discuss the current insights into synapse formation and function in vivo. These fascinating studies will contribute to our understanding of individual neural circuit structure and function as well as the pathomechanisms of psychiatric and neurological disorders.
One of the most significant breakthroughs in the synaptic proteome has been the isolation of synaptosomes from the brain and the subsequent comprehensive proteomic analysis of their molecular components. In 1964, Dr. Whittaker and his colleagues first successfully purified the synaptic compartment termed “synaptosome” from brain tissue through density-gradient centrifugation (Whittaker et al., 1964). The purification of synaptosomes has enabled extensive studies on the molecular composition and structure of synapses. However, these conventional synaptosome studies faced difficulties in purifying cell-type-specific synaptic compartments in vivo.
Fluorescence activated synaptosome sorting (FASS) is a powerful approach that improves the purification of conventional synaptosomes (Biesemann et al., 2014). The synaptosomes are fluorescently labeled by a transgenic approach with fluorescent proteins or the application of dyes specifically targeting synaptic membranes. After fluorescent labeling, they are isolated by flow cytometry and flowed by LC–MS/MS (Biesemann et al., 2014; Apóstolo et al., 2020; Paget-Blanc et al., 2022). Thus, FASS technique allows for the analysis of cell-type-specific synaptic compartments (Biesemann et al., 2014). Using VGLUT1VENUS knock-in mice, fluorescent glutamatergic synaptosomes (VGLUT1-positive excitatory synapse) were selectively purified and identified 163 synaptic proteins in the mouse forebrain (Biesemann et al., 2014). A notable finding of this study is the discovery of FXYD6 and Tpd52 as novel glutamatergic synapse components (Biesemann et al., 2014). Moreover, FASS approach was utilized to analyze the cell-surface protein composition of mossy fiber synapses in the hippocampus that govern microcircuit connectivity related to cognitive function (Apóstolo et al., 2020). Mossy fiber synaptosomes were purified by fluorescent labeling with Nectin-3 and FM4-64 membrane dye, followed by isolation using a fluorescent cell sorter (Apóstolo et al., 2020). Based on the approach, 77 proteins were identified as cell surface proteins of the mossy fiber synapse in vivo (Apóstolo et al., 2020). Among these proteins, this study reveals that IgSF8 is a crucial regulator of CA3 microcircuit connectivity and function (Apóstolo et al., 2020). FASS has recently also been employed for the synaptic molecular mapping of dopaminergic neurons in vivo (Paget-Blanc et al., 2022). Dopaminergic synapses were labeled by the expression of adeno-associated viral (AAV) vectors carrying Cre-dependent EGFP in the dopaminergic neurons (DAT-Cre transgenic mouse). EGFP-labeled dopaminergic synaptosomes were purified and then sorted using fluorescent cell sorters. It was found that 57 proteins specifically enriched at the dopaminergic synapse (Paget-Blanc et al., 2022). Interestingly, these dopaminergic synaptosomes contain multiple synaptic compartments including glutamatergic, GABAergic, or cholinergic synapses, that form “dopamine hub synapses” (Paget-Blanc et al., 2022). The dopamine hub synapses potentially modulate dopamine volume transmission, which is crucial for reward processing and motor control. Thus, FASS approach enables spatial synaptic proteome analysis through improved isolation specificity for particular cell types in comparison to conventional synaptosome purification.
Proximity labeling is a highly innovative approach to evaluating protein components present in specific cell types and subcellular localizations within cultured neurons as well as brain tissue (Han et al., 2018; Takano and Soderling, 2021). Proximity labeling is classified as either peroxidase-based proximity labeling or biotin ligase-based proximity labeling (BioID) (Han et al., 2018). Peroxidase-based proximity labeling utilizes ascorbate peroxidase derivatives like APEX or horseradish peroxidase (HRP) (Honke and Kotani, 2011; Rhee et al., 2013). By fusing these enzymes to a bait protein and introducing them into cells, they label tyrosine residues in proteins within a 10–20 nm range with biotin (Figure 1A). Following affinity purification with avidin-coated beads, the biotinylated proteins can be analyzed in more detail using LC–MS/MS to discover the spatial protein composition. Since the biotinylation labeling by HRP occurs in 1 min through biotin-phenol radical catalysis in the presence of hydrogen peroxide (H2O2), it is suitable for the analysis of spatial proteomes in living cells in vitro and ex vivo. Also, APEX catalyzes the oxidation and deposition of diaminobenzidine (DAB) and can also be used for electron microscopy imaging. BioID approach first utilized an Escherichia coli-derived mutant biotin ligase (BirA*-R118G), which produces reactive biotin (biotinoyl-5′-AMP) with an enhanced off-rate such that biotin covalently binds to exposed lysine residues of neighboring proteins within approximately 10 nm (Roux et al., 2012; Kim et al., 2016). There are several biotin ligases currently used in BioID approach, including BioID2, BASU, miniTurbo, TurboID, ultraID, microID, MicroID2, and AirID (Kim et al., 2016; Branon et al., 2018; Ramanathan et al., 2018; Kido et al., 2020; Johnson et al., 2022; Kubitz et al., 2022). Recently, BioID has been extensively applied to spatial proteome analysis in the brain and referred to as “in vivo BioID (iBioID)” (Uezu et al., 2016). iBioID possesses biocompatibility, high detection sensitivity, and spatial resolution capabilities, making it one of the most effective approaches for uncovering molecular landscapes specific to neuronal cell types and synapses. Therefore, we have summarized these approaches to elucidate the spatial proteome, which includes the molecular landscapes of synapses and the subcellular compartments of neurons and astrocytes, as well as the molecular mechanisms controlling synaptic formation and function in the brain (Figure 1B).
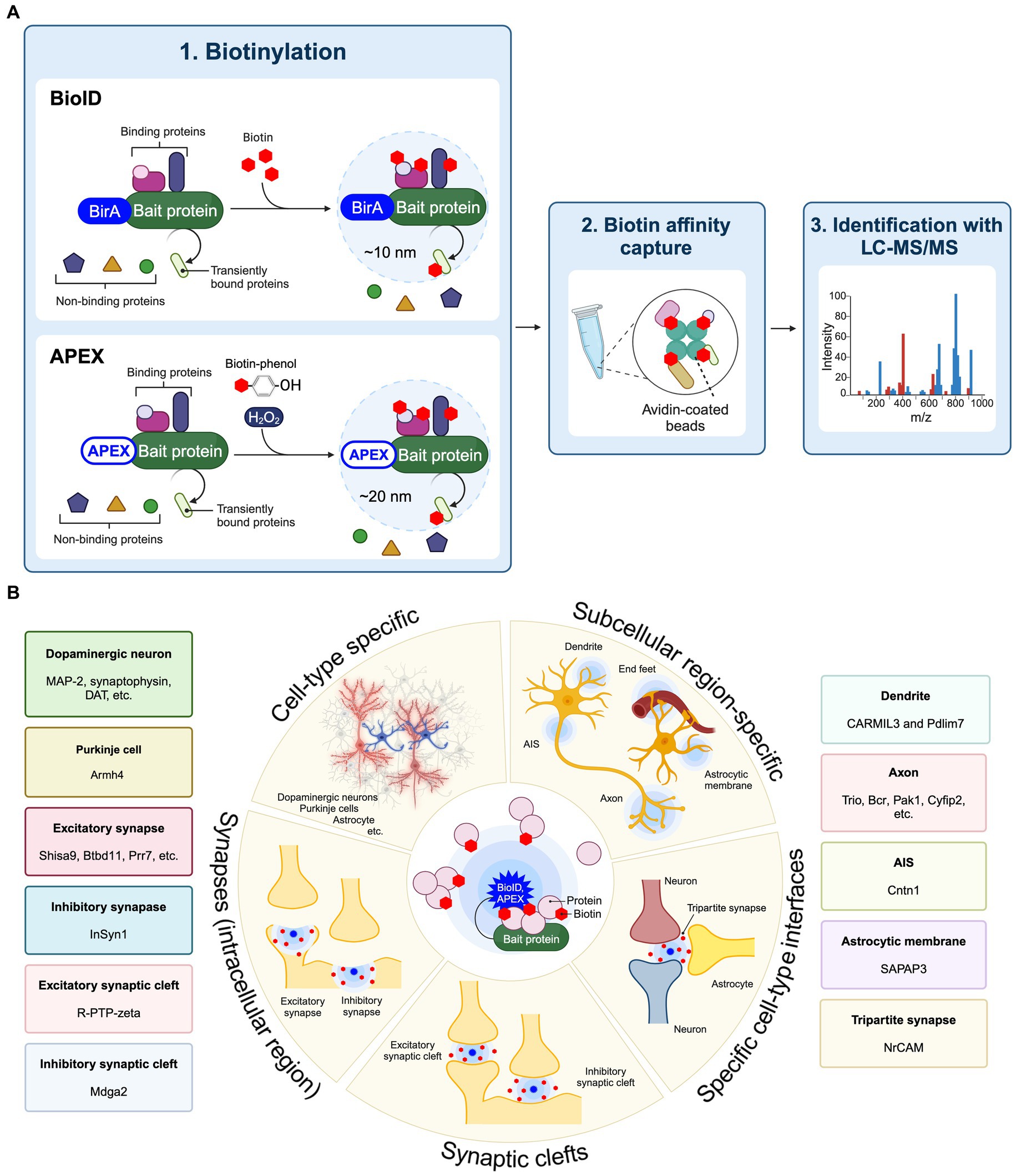
Figure 1. Application of proximity labeling to identify molecular landscape at specific cell type and subcellular site in vivo. (A) A schematic diagram and application of BioID and APEX. The protein of interest (bait) is fused with BirA*-R118G or APEX and expressed in cells. The enzyme biotinylates neighboring proteins of the bait protein. The biotinylated proteins are purified using avidin beads. Mass spectrometry is then utilized to identify proteins around the bait protein. (B) Proximity labeling approaches such as BioID and APEX enable the spatial proteomic analysis at cell-type-specific levels combined with subcellular localization, including individual types of synapses from neurons and astrocytes in the brain, using cell-type-specific AAVs or transgenic mouse lines. Proximity labeling can specifically target distinct cell types such as neurons and astrocytes to identify protein components through the cell-type-specific expression approaches in vivo. Additionally, proximity labeling can find protein networks within particular subcellular compartments, including the cell surface, dendritic spine, axon, axon initial segment (AIS), astrocytic membrane, end feet, and synapses by utilizing protein fusion targeted to these subcellular regions. At synapses, proximity labeling is capable of labeling intracellular synaptic proteins or synaptic cleft proteins by fusion with scaffold proteins or cell adhesion proteins. Split-TurboID also can decipher protein compositions between different cell types in the brain. Representative molecular landscapes that identified by these cell-type specific BioID or APEX are also shown.
Proximity labeling allows for the isolation of protein components within excitatory and inhibitory synapses in vivo. Notably, the structural characteristics of inhibitory synapses make it challenging to access via conventional synaptosomal purification. Dr. Soderling and his colleagues first established in vivo BioID (iBioID) to investigate the proteomes of excitatory and inhibitory synapses in the brain using BirA*-R118G fused with postsynaptic scaffold proteins PSD-95 and gephyrin (Uezu et al., 2016) (Figure 1B; Table 1). In this study, AAV vectors carrying PSD95-BirA or gephyrin-BirA were expressed in mouse cortical and hippocampal neurons, and the biotin-labeled synaptic proteins were analyzed by LC–MS/MS. These proteomic approaches successfully identified 121 excitatory synaptic proteins and 181 inhibitory synaptic proteins, including both known and novel proteins. The important finding is that a novel inhibitory synaptic protein InSyn1 plays a critical role in inhibitory synapse formation and function through the interaction with the dystrophin complex in vivo (Uezu et al., 2016). This iBioID approach has facilitated the spatial proteome analysis within various neuronal intracellular compartments (Figure 1B). For example, the analysis of the dendritic spine proteome has been investigated using Rac-GAP (Wrp)-BirA and BioID2-synaptopodin (Spence et al., 2019; Falahati et al., 2022), while the axonal proteome has been explored using BioID2-Synapsin (O’Neil et al., 2021). Recently, the proteome of axon initial segment (AIS) has been detected using Neurofascin-HRP (Ogawa et al., 2023) (Figure 1B; Table 1). iBioID has also been applied to extensive interactome analyses of molecules associated with autism spectrum disorders (ASD) (Gao et al., 2023) (Table 1). In this study, a high-throughput genome-editing approach was used to target 14 high-confidence ASD genes (Anks1b, Syngap1, Shank2, Shank3, Nckap1, Nbea, Ctnnb1, Lrrc4c, Iqsec2, Arhgef9, Ank3, Scn2a, Scn8a, and Hnrnpu) for the insertion of a highly active mutant biotin ligase TurboID. These genes were selected for an iBioID-based proteome as spatially localized proteins at neuronal compartments such as the synapse, AIS, and nucleus. Comparative interactome analysis revealed a significant overlap of protein interactions among the targeted genes, with enrichment for ASD-related cellular functions. Additionally, this genome-editing mediated iBioID approach enables the investigation of native proteomes without relying on any overexpression methods in vivo (Gao et al., 2023). Therefore, these approaches will enable comprehensive analyses of synaptic-related molecules in physiological states and pathological conditions that lead to neurological disorders. iBioID approach has also been utilized for the cell-type-specific proteomic analysis of glutamatergic neurons and astrocytes in the brain using cell-type-specific AAV vectors or transgenic mouse lines (Sun et al., 2022) (Table 1). By using two different cell-type-specific promotors to express TurboID into the glutamatergic neurons (CaMKII promoter) and astrocytes (GFAP promoter) in the brain, more than 10,000 proteins were identified with excellent reproducibility (Sun et al., 2022). In addition to AAV-based expression approaches with TurboID or BioID2, neuron-specific (Rosa26TurboID/wt/CaMK2a) and astrocyte-specific (Rosa26TurboID/wt/Aldh1l1) TurboID knock-in mouse lines were established to investigate the cell-type-specific proteome in vivo (Rayaprolu et al., 2022) (Table 1). They identified over 2,000 proteins in each cell type, and more than 200 proteins that are different between neurons and astrocytes. Notably, MAPK signaling was elevated in neurons compared to astrocytes. These findings suggest the clarification of unique protein networks in CaMK2a-positive glutamatergic neurons and Aldh1l1-positive astrocytes, which are associated with their distinct cellular functions (Rayaprolu et al., 2022). Recently, BV2 microglial cells expressing cytoplasmic TurboID were analyzed to investigate their intracellular proteome (Sunna et al., 2023) (Table 1). Under lipopolysaccharide (LPS)-stimulated inflammatory conditions, more than 500 proteins showed differential abundance out of the 2,350 identified, demonstrating an increase in inflammatory proteins (Il1a, Irg1, Oasl1) and a decrease in anti-inflammatory proteins (Arg1, Mgl2) (Sunna et al., 2023). Moreover, the proteome specific to different cell types and subcellular locations was assessed using AAV vectors carrying either cytosolic-BioID2 or plasma membrane-BioID2 in vivo (Soto et al., 2023) (Figure 1B; Table 1). They were expressed in mouse striatal neurons (AAV-hSynI-BioID2 and AAV-hSynI-Lck-BioID2) and astrocytes (AAV-GfaABC1D-BioID2 and AAV-GfaABC1D-Lck-BioID2) (Soto et al., 2023). Using these approaches, they discovered a range of 500–1,800 proteins in the cytoplasm and plasma membrane of neurons and astrocytes. Furthermore, the subcellular proteomes of five distinct astrocyte subcellular compartments were explored in vivo (Soto et al., 2023) (Table 1). These include the end feet (AQP4-BioID2), astrocytic process (EZR-BioID2), extracellular glutamate uptake site (GLT1-BioID2), extracellular K+ homeostasis site (KIR4.1-BioID2) and astrocyte-astrocyte contacts (CX43-BioID2). These approaches were highly successful, revealing that SAPAP3 is specifically localized to the astrocytic process and involved in obsessive-compulsive disorder (OCD) and repetitive behaviors (Soto et al., 2023).
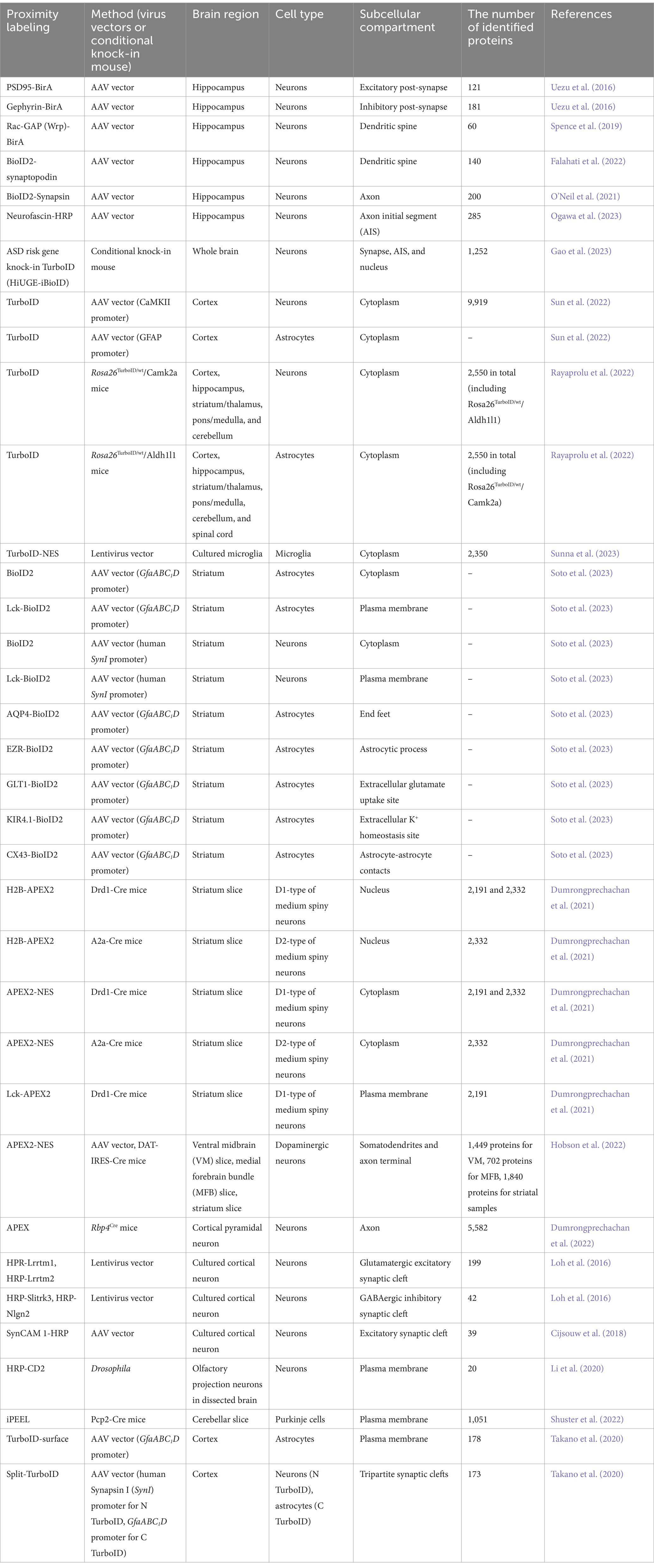
Table 1. Summary of proximity labeling approaches to decode molecular landscape in the brain.
APEX has also been used to determine cell-type-specific subcellular protein composition ex vivo (Dumrongprechachan et al., 2021) (Table 1). Utilizing a combination of AAV vectors and cell-type-specific conditional mouse lines, APEX2 was expressed in D1 or D2 types of medium spiny neurons of the striatum using Drd1-Cre and A2a-Cre mice, respectively (Dumrongprechachan et al., 2021). In addition, D1-type neurons were analyzed for their subcellular compartment-specific proteomic profiles using nucleus-APEX2 (H2B-APEX2), cytoplasm-APEX2 (APEX2-NES), and plasma membrane-APEX2 (Lck-APEX2). Consequently, this study successfully isolated cell-type-specific subcellular proteomic data from brain tissue samples, unveiling dynamic changes in the proteome of striatal neurons following chemogenetic activation of Gαq-coupled signaling cascades (Dumrongprechachan et al., 2021). Another research group employed a similar APEX-based approach to elucidate the protein compositions in midbrain dopaminergic neurons (Hobson et al., 2022) (Figure 1B; Table 1). APEX was expressed in dopaminergic (DA) neurons within the midbrain of DAT-IRES-Cre mouse using AAV vector. Subsequently, the cellular compartments such as the somatodendrites and axon terminal were isolated and analyzed by microdissection (Hobson et al., 2022). This approach determined the DA-specific presynaptic enriched proteins including DA neurotransmission and metabolism from the midbrain (Hobson et al., 2022). The application of APEX has further enabled the mapping of the cell-type-specific proteome and phosphoproteome of corticostriatal axons during neuronal development (Dumrongprechachan et al., 2022) (Table 1). APEX2 expression was induced in layer 5 cortical pyramidal neurons using Rbp4Cre mice (Dumrongprechachan et al., 2022). Subsequently, the axonal proteome was isolated from the striatal lysates at various developmental stages (Dumrongprechachan et al., 2022). This study identified co-regulated proteins and phosphorylations in corticostriatal axons during neurodevelopment, which are linked to genetic risk for human brain disorders (Dumrongprechachan et al., 2022). Taken together, these findings demonstrated that iBioID and APEX approaches are highly effective for determining the cell-type-specific subcellular protein compositions in vivo and ex vivo.
Integrated neuronal systems communicate extensively through signaling in the synaptic cleft. This cell-surface signaling is pivotal in controlling nearly all aspects of neuronal development and physiology in the brain. Additionally, astrocytes and neurons form a unique cell adhesion structure called a “tripartite synapse,” suggesting that astrocytes’ direct involvement in synaptic formation and function (Allen and Eroglu, 2017; Takano and Soderling, 2021). However, comprehensive identification of molecular compositions at the cell-type specific interfaces in vivo has encountered significant limitations. This limitation stems from conventional biological approaches’ inability to capture protein components at synaptic clefts. Moreover, the characteristic attributes of cell-surface proteins, such as their low abundance, high hydrophobicity, and heterogeneity, further complicate this issue (Kuhlmann et al., 2018; Li et al., 2019).
Recent advancements in cell-surface proteomic profiling have enabled significant insights into neuronal synapse formation and function in vitro and ex vivo using HRP-based proximity labeling approaches (Loh et al., 2016; Cijsouw et al., 2018; Li et al., 2020; Shuster et al., 2022). For instance, HRP fused to known synaptic cleft proteins such as Lrrtm1, Lrrtm2, Slitrk3, or Neuroligin-2 (Nlgn2), have been employed for analysis of excitatory and inhibitory synaptic cleft proteomes in cultured neurons (Loh et al., 2016) (Figure 1B; Table 1). These studies have uncovered 199 glutamatergic and 42 GABAergic synaptic cleft proteins in vitro. Importantly, they found that a novel synaptic cleft protein Mdga2 plays a key role in inhibitory synapse formation by recruiting Nlgn2 to the postsynaptic site (Loh et al., 2016). Additionally, HRP fused to an excitatory cell adhesion molecule SynCAM 1 has deciphered several excitatory synaptic cleft proteins including receptor-type tyrosine phosphatase zeta (R-PTP-zeta) in cultured cortical neurons (Cijsouw et al., 2018) (Figure 1B; Table 1). In another study, the cell-type-specific expression of HRP fused to the N-terminal extracellular domain of CD2 (HRP-CD2) has been employed to discover novel cell surface molecules in Drosophila olfactory projection neurons ex vivo (Li et al., 2020) (Table 1). Recently, a new approach for cell-type-specific cell surface proteomics, termed in situ cell surface proteome extraction using extracellular labeling (iPEEL), has been developed (Shuster et al., 2022) (Figure 1B; Table 1). This approach extends the HRP-CD2-based proteome analysis by utilizing cell-type-specific conditional mouse lines. Using a transgenic mouse line (Pcp2-Cre), iPEEL identified 1,051 cell-surface proteins in Purkinje cells. Interestingly, it was revealed that mature cerebellar Purkinje cells and developing Purkinje cells exhibit substantial proteomic overlap. Additionally, different classes of cell-surface proteins exhibited selective enrichment at different developmental stages. Furthermore, they found that Armh4 plays an essential role in Purkinje cell dendrite morphogenesis by regulating endocytosis (Shuster et al., 2022).
A new approach known as TurboID-surface and Split-TurboID has emerged more recently for spatial proteomics on cell-type-specific cell surfaces and interfaces in vivo including astrocytic cell surfaces and tripartite synapses (Takano et al., 2020; Takano and Soderling, 2021) (Figure 1B; Table 1). TurboID-surface, fused to a glycosylphosphatidylinositol (GPI) anchor, has been delivered to cortical astrocytes using cell-type-specific AAV vectors (GfaABC1D promoter), facilitating the comprehensive profiling of astrocytic membrane proteins in vivo. Split-TurboID is composed of the N-terminal (N-TurboID) and C-terminal (C-TurboID) fragments of the TurboID surface. The cell-type-specific AAV vectors carrying N-TurboID and C-TurboID were expressed in mouse cortical neurons (human Synapsin I (SynI) promoter) and astrocytes (GFAP promoter), respectively. A functional TurboID is formed at the tripartite synaptic clefts once N-TurboID and C-TurboID fragments merge at the interface between neurons and astrocytes (Takano et al., 2020; Takano and Soderling, 2021). Super-resolution Stimulated Emission Depletion (STED) microscopy showed that astrocytic TurboID-surface and Split-TurboID were localized at the peri-synaptic sites in vivo (Takano et al., 2020). Using label-free quantitative LC–MS/MS analysis, TurboID-surface and Split-TurboID identified 178 and 173 extracellular proteins, respectively. One hundred and eighteen proteins were found to be common among these proteins, forming a tripartite proteome synapse with a high level of confidence (Takano et al., 2020). Notably, these spatial proteome approaches have unveiled a novel role for astrocytes in directly regulating the formation and function of inhibitory synapses through a new player NrCAM in vivo. Together, these studies provide novel insight into neuronal connectivity and brain function by mapping the protein composition in cell-type-specific synaptic cleft.
Deciphering the synaptic molecular landscape in specific cell types and synapses is crucial for understanding neural circuit formation and their roles in brain functions. Additionally, it’s essential for developing new therapeutic strategies based on the pathophysiology of neurological disorders characterized by synaptic dysfunction. However, given the considerable heterogeneity of neurons and glial cells in the brain, determining the molecular composition of specific cell types, along with their subcellular structures and synapses, continues to be a significant challenge in neuroscience.
Recent advancements in spatial proteomics, including FASS, APEX, and iBoID, have facilitated the analysis of protein components at targeted cellular and subcellular levels within the brain. These highly innovative approaches have uncovered numerous uncharacterized molecules in individual types of neurons, astrocytes, and specific synapses within brain tissue, each exhibiting unique molecular mechanisms. Employing these spatial proteome strategies will enhance our comprehension of various synaptic types, though they have a few restrictions to overcome. FASS approach shows a high level of specificity in selecting particular subcellular components, effectively reducing contamination (Biesemann et al., 2014; van Oostrum et al., 2023). This specificity is a consequence of synaptosome sorting properties, although it may result in lower throughput. Since neurons typically receive input from a wide variety of different presynaptic partners, the current FASS approach is restrained to targeting neurons at a cellular level, and may lead to an analysis that reflects only an averaged synaptic proteome. In contrast, APEX and iBioID allow spatial proteomic analysis at cell-type-specific levels combined with subcellular localization, including individual types of synapses from neurons and astrocytes in the brain, using cell-type-specific AAVs or transgenic mouse lines (Figure 1B). However, the in vivo application of both APEX and iBioID is constrained by certain limitations. The enzyme activity of APEX necessitates H2O2, which is cytotoxic and less amenable to labeling reactions in tissue. With iBioID, exogenous biotin administration for 7 days is necessary to achieve significant biotinylation labeling (Uezu et al., 2016; Spence et al., 2019). In addition, the enzyme activity of all BirA variants depends on ATP, which is typically present in low extracellular concentrations in vivo (Takano and Soderling, 2021). Despite these limitations, APEX and iBioID approaches have enabled successfully the identification of protein compositions in various neuron types—including glutamatergic neurons, dopaminergic neurons, and Purkinje cells as well as astrocytes (Table 1). The knowledge would provide deeper insight into how these different protein networks regulate individual neural circuits modulation and control various brain functions such as memory, learning, and emotion. Notably, iBioID accomplished subcellular protein profiling without causing cellular toxicity (Takano et al., 2020; Takano and Soderling, 2021). Furthermore, TurboID-surface and Split-TurboID have efficiently enabled the profiling of spatial protein networks in cell-type-specific synaptic clefts, showing particularly notable results in tripartite synapses (Takano et al., 2020; Takano and Soderling, 2021). Recent research has reported metabolic labeling methods as novel approaches for labeling nascent proteomes with cell type specificity in the mouse brain, such as MetRS and SORT (Alvarez-Castelao et al., 2017; Krogager et al., 2017; Tang and Chen, 2023). These methods utilize a methionyl-tRNA synthetase and an orthogonal pyrrolysyl-tRNA synthetase-tRNAXXX pair, thereby tagging newly synthesized proteins with non-canonical amino acids and click chemistry (Alvarez-Castelao et al., 2017; Krogager et al., 2017; Tang and Chen, 2023). The combination of these approaches with proximity labeling will offer more precise insights into the cell-type-specific synaptic proteome.
A recent study utilizing spatial transcriptomics has accomplished the single-cell tracing of developmental gene programs, leading to the differentiation of diverse cell types in the brain (Lein et al., 2017; Ratz et al., 2022). Similarly, these conceptual approaches are currently being adapted and optimized for proteomics. Integrating these applications with proximity labeling will advance spatial proteomic analysis, allowing for the tracking of the proteome within a single synapse in the future. These breakthroughs would enable the discovery of molecular mechanisms governing single neural circuit formation and function. Furthermore, recent evidence has increasingly indicated that the dysfunction of many synaptic genes significantly contributes to psychiatric and neurological disorders. Therefore, a comparative analysis of cell-type-specific synaptic proteomes is becoming increasingly essential in understanding the diverse pathogenesis of many neurological disorders. A large-scale interactome analysis of 14 risk genes including synaptic genes associated with ASD, using TurboID, has revealed a wealth of common cellular functions and molecular interactions related to the pathomechanisms of ASD (Gao et al., 2023). This dataset is expected to contribute to the establishment of new diagnostic and therapeutic methods targeting common molecules in ASD pathology. Taken together, advancing these studies further could eventually involve investigations exploring variations in cell-type-specific synaptic proteomes among different states of neuronal cells and glial cells in vivo. The consolidation of this accumulated data into a database is expected to substantially progress our comprehension of the brain’s information processing mechanisms and pharmaceutical development.
YI: Conceptualization, Visualization, Writing – original draft, Writing – review & editing. SN: Writing – review & editing. TT: Conceptualization, Funding acquisition, Visualization, Writing – original draft, Writing – review & editing.
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This work was supported by a Grant-Aid for Scientific Research B (21380936) from the JSPS (TT), a PRESTO (21461219) from JST (TT), The Memorial Foundation for Medical and Pharmaceutical Research (TT), and The Takeda Science Foundation (TT).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Allen, N. J., and Eroglu, C. (2017). Cell biology of astrocyte-synapse interactions. Neuron 96, 697–708. doi: 10.1016/J.NEURON.2017.09.056
Alvarez-Castelao, B., Schanzenbächer, C. T., Hanus, C., Glock, C., tom Dieck, S., Dörrbaum, A. R., et al. (2017). Cell-type-specific metabolic labeling of nascent proteomes in vivo. Nat. Biotechnol. 35, 1196–1201. doi: 10.1038/nbt.4016
Apóstolo, N., Smukowski, S. N., Vanderlinden, J., Condomitti, G., Rybakin, V., ten Bos, J., et al. (2020). Synapse type-specific proteomic dissection identifies IgSF8 as a hippocampal CA3 microcircuit organizer. Nat. Commun. 11, 5171–5121. doi: 10.1038/s41467-020-18956-x
Biesemann, C., Grønborg, M., Luquet, E., Wichert, S. P., Bernard, V., Bungers, S. R., et al. (2014). Proteomic screening of glutamatergic mouse brain synaptosomes isolated by fluorescence activated sorting. EMBO J. 33, 157–170. doi: 10.1002/embj.201386120
Branon, T. C., Bosch, J. A., Sanchez, A. D., Udeshi, N. D., Svinkina, T., Carr, S. A., et al. (2018). Efficient proximity labeling in living cells and organisms with TurboID. Nat. Biotechnol. 36, 880–887. doi: 10.1038/nbt.4201
Cijsouw, T., Ramsey, A. M., Lam, T. K. T., Carbone, B. E., Blanpied, T. A., and Biederer, T. (2018). Mapping the proteome of the synaptic cleft through proximity labeling reveals new cleft proteins. Proteomes 6:48. doi: 10.3390/PROTEOMES6040048
Delint-Ramirez, I., Fernández, E., Bayés, A., Kicsi, E., Komiyama, N. H., and Grant, S. G. N. (2010). In vivo composition of NMDA receptor signaling complexes differs between membrane subdomains and is modulated by PSD-95 and PSD-93. J. Neurosci. 30, 8162–8170. doi: 10.1523/JNEUROSCI.1792-10.2010
Dosemeci, A., Makusky, A. J., Jankowska-Stephens, E., Yang, X., Slotta, D. J., and Markey, S. P. (2007). Composition of the synaptic PSD-95 complex. Mol. Cell. Proteomics 6, 1749–1760. doi: 10.1074/mcp.M700040-MCP200
Dumrongprechachan, V., Salisbury, R. B., Butler, L., Macdonald, M. L., and Kozorovitskiy, Y. (2022). Dynamic proteomic and phosphoproteomic atlas of corticostriatal axons in neurodevelopment. eLife 11:e78847. doi: 10.7554/ELIFE.78847
Dumrongprechachan, V., Salisbury, R. B., Soto, G., Kumar, M., MacDonald, M. L., and Kozorovitskiy, Y. (2021). Cell-type and subcellular compartment-specific APEX2 proximity labeling reveals activity-dependent nuclear proteome dynamics in the striatum. Nat. Commun. 12, 4855–4816. doi: 10.1038/s41467-021-25144-y
Falahati, H., Wu, Y., Feuerer, V., Simon, H. G., and De Camilli, P. (2022). Proximity proteomics of synaptopodin provides insight into the molecular composition of the spine apparatus of dendritic spines. Proc. Natl. Acad. Sci. USA 119:e2203750119. doi: 10.1073/pnas.2203750119
Farr, C. D., Gafken, P. R., Norbeck, A. D., Doneanu, C. E., Stapels, M. D., Barofsky, D. F., et al. (2004). Proteomic analysis of native metabotropic glutamate receptor 5 protein complexes reveals novel molecular constituents. J. Neurochem. 91, 438–450. doi: 10.1111/J.1471-4159.2004.02735.X
Fernández, E., Collins, M. O., Uren, R. T., Kopanitsa, M. V., Komiyama, N. H., Croning, M. D. R., et al. (2009). Targeted tandem affinity purification of PSD-95 recovers core postsynaptic complexes and schizophrenia susceptibility proteins. Mol. Syst. Biol. 5:269. doi: 10.1038/msb.2009.27
Francesconi, A., Kumari, R., and Suzanne Zukin, R. (2009). Proteomic analysis reveals novel binding partners of metabotropic glutamate receptor 1. J. Neurochem. 108, 1515–1525. doi: 10.1111/J.1471-4159.2009.05913.X
Gao, Y., Trn, M., Shonai, D., Zhao, J., Soderblom, E. J., Garcia-Moreno, S. A., et al. (2023). Proximity analysis of native proteomes reveals Interactomes predictive of phenotypic modifiers of autism and related neurodevelopmental conditions. BioRxiv, 2022.10.06.511211. doi: 10.1101/2022.10.06.511211
Han, S., Li, J., and Ting, A. Y. (2018). Proximity labeling: spatially resolved proteomic mapping for neurobiology. Curr. Opin. Neurobiol. 50, 17–23. doi: 10.1016/J.CONB.2017.10.015
Hobson, B. D., Choi, S. J., Mosharov, E. V., Soni, R. K., Sulzer, D., and Sims, P. A. (2022). Subcellular proteomics of dopamine neurons in the mouse brain. eLife 11:e70921. doi: 10.7554/ELIFE.70921
Honke, K., and Kotani, N. (2011). The enzyme-mediated activation of radical source reaction: a new approach to identify partners of a given molecule in membrane microdomains. J. Neurochem. 116, 690–695. doi: 10.1111/J.1471-4159.2010.07027.X
Husi, H., Ward, M. A., Choudhary, J. S., Blackstock, W. P., and Grant, S. G. N. (2000). Proteomic analysis of NMDA receptor–adhesion protein signaling complexes. Nat. Neurosci. 3, 661–669. doi: 10.1038/76615
Johnson, B. S., Chafin, L., Farkas, D., Adair, J., Elhance, A., Farkas, L., et al. (2022). MicroID2: a novel biotin ligase enables rapid proximity-dependent proteomics. Mol. Cell. Proteomics 21:100256. doi: 10.1016/j.mcpro.2022.100256
Kido, K., Yamanaka, S., Nakano, S., Motani, K., Shinohara, S., Nozawa, A., et al. (2020). Airid, a novel proximity biotinylation enzyme, for analysis of protein–protein interactions. eLife 9:e54983. doi: 10.7554/ELIFE.54983
Kim, D. I., Jensen, S. C., Noble, K. A., Kc, B., Roux, K. J. K. H. K. J., Motamedchaboki, K., et al. (2016). An improved smaller biotin ligase for BioID proximity labeling. Mol. Biol. Cell 27, 1188–1196. doi: 10.1091/mbc.E15-12-0844
Krapivinsky, G., Medina, I., Krapivinsky, L., Gapon, S., and Clapham, D. E. (2004). SynGAP-MUPP1-CaMKII synaptic complexes regulate p38 MAP kinase activity and NMDA receptor- dependent synaptic AMPA receptor potentiation. Neuron 43, 563–574. doi: 10.1016/j.neuron.2004.08.003
Krogager, T. P., Ernst, R. J., Elliott, T. S., Calo, L., Beránek, V., Ciabatti, E., et al. (2017). Labeling and identifying cell-specific proteomes in the mouse brain. Nat. Biotechnol. 36, 156–159. doi: 10.1038/nbt.4056
Kubitz, L., Bitsch, S., Zhao, X., Schmitt, K., Deweid, L., Roehrig, A., et al. (2022). Engineering of ultraID, a compact and hyperactive enzyme for proximity-dependent biotinylation in living cells. Commun. Biol. 5, 657–614. doi: 10.1038/s42003-022-03604-5
Kuhlmann, L., Cummins, E., Samudio, I., and Kislinger, T. (2018). Cell-surface proteomics for the identification of novel therapeutic targets in cancer. Expert Rev. Proteomics 15, 259–275. doi: 10.1080/14789450.2018.1429924
Lee, Y., Ryu, J. R., Kang, H., Kim, Y., Kim, S., Zhang, Y., et al. (2017). Characterization of the zinc-induced Shank3 interactome of mouse synaptosome. Biochem. Biophys. Res. Commun. 494, 581–586. doi: 10.1016/J.BBRC.2017.10.143
Lein, E., Borm, L. E., and Linnarsson, S. (2017). The promise of spatial transcriptomics for neuroscience in the era of molecular cell typing. Science 358, 64–69. doi: 10.1126/science.aan6827
Lepeta, K., Lourenco, M. V., Schweitzer, B. C., Martino Adami, P. V., Banerjee, P., Catuara-Solarz, S., et al. (2016). Synaptopathies: synaptic dysfunction in neurological disorders – a review from students to students. J. Neurochem. 138, 785–805. doi: 10.1111/JNC.13713
Li, J., Han, S., Li, H., Udeshi, N. D., Svinkina, T., Mani, D. R., et al. (2020). Cell-surface proteomic profiling in the Fly brain uncovers wiring regulators. Cell 180, 373–386.e15. doi: 10.1016/j.cell.2019.12.029
Li, Y., Wang, Y., Mao, J., Yao, Y., Wang, K., Qiao, Q., et al. (2019). Sensitive profiling of cell surface proteome by using an optimized biotinylation method. J. Proteome 196, 33–41. doi: 10.1016/J.JPROT.2019.01.015
Li, J., Zhang, W., Yang, H., Howrigan, D. P., Wilkinson, B., Souaiaia, T., et al. (2017). Spatiotemporal profile of postsynaptic interactomes integrates components of complex brain disorders. Nat. Neurosci. 20, 1150–1161. doi: 10.1038/nn.4594
Loh, K. H., Stawski, P. S., Draycott, A. S., Udeshi, N. D., Lehrman, E. K., Wilton, D. K., et al. (2016). Proteomic analysis of unbounded cellular compartments: synaptic clefts. Cell 166, 1295–1307.e21. doi: 10.1016/j.cell.2016.07.041
Luo, J., Norris, R. H., Gordon, S. L., and Nithianantharajah, J. (2018). Neurodevelopmental synaptopathies: insights from behaviour in rodent models of synapse gene mutations. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 84, 424–439. doi: 10.1016/J.PNPBP.2017.12.001
Ogawa, Y., Lim, B. C., George, S., Oses-Prieto, J. A., Rasband, J. M., Eshed-Eisenbach, Y., et al. (2023). Antibody-directed extracellular proximity biotinylation reveals that Contactin-1 regulates axo-axonic innervation of axon initial segments. Nat. Commun. 14, 6797–6718. doi: 10.1038/s41467-023-42273-8
O’Neil, S. D., Rácz, B., Brown, W. E., Gao, Y., Soderblom, E. J., Yasuda, R., et al. (2021). Action potential-coupled rho gtpase signaling drives presynaptic plasticity. eLife 10:e63756. doi: 10.7554/ELIFE.63756
Paget-Blanc, V., Pfeffer, M. E., Pronot, M., Lapios, P., Angelo, M. F., Walle, R., et al. (2022). A synaptomic analysis reveals dopamine hub synapses in the mouse striatum. Nat. Commun. 13, 3102–3119. doi: 10.1038/s41467-022-30776-9
Pandya, N. J., Klaassen, R. V., van der Schors, R. C., Slotman, J. A., Houtsmuller, A., Smit, A. B., et al. (2016). Group 1 metabotropic glutamate receptors 1 and 5 form a protein complex in mouse hippocampus and cortex. Proteomics 16, 2698–2705. doi: 10.1002/PMIC.201500400
Pasciuto, E., and Bagni, C. (2014). SnapShot: FMRP interacting proteins. Cell 159, 218–218.e1. doi: 10.1016/j.cell.2014.08.036
Ramanathan, M., Majzoub, K., Rao, D. S., Neela, P. H., Zarnegar, B. J., Mondal, S., et al. (2018). RNA–protein interaction detection in living cells. Nat. Methods 15, 207–212. doi: 10.1038/nmeth.4601
Ramos, C., Chardonnet, S., Marchand, C. H., Decottignies, P., Ango, F., Daniel, H., et al. (2012). Native presynaptic metabotropic glutamate receptor 4 (mGluR4) interacts with exocytosis proteins in rat cerebellum. J. Biol. Chem. 287, 20176–20186. doi: 10.1074/jbc.M112.347468
Ratz, M., von Berlin, L., Larsson, L., Martin, M., Westholm, J. O., La Manno, G., et al. (2022). Clonal relations in the mouse brain revealed by single-cell and spatial transcriptomics. Nat. Neurosci. 25, 285–294. doi: 10.1038/s41593-022-01011-x
Rayaprolu, S., Bitarafan, S., Santiago, J. V., Betarbet, R., Sunna, S., Cheng, L., et al. (2022). Cell type-specific biotin labeling in vivo resolves regional neuronal and astrocyte proteomic differences in mouse brain. Nat. Commun. 13, 2927–2918. doi: 10.1038/s41467-022-30623-x
Rhee, H. W., Zou, P., Udeshi, N. D., Martell, J. D., Mootha, V. K., Carr, S. A., et al. (2013). Proteomic mapping of mitochondria in living cells via spatially restricted enzymatic tagging. Science 339, 1328–1331. doi: 10.1126/science.1230593
Roux, K. J., Kim, D. I., Raida, M., and Burke, B. (2012). A promiscuous biotin ligase fusion protein identifies proximal and interacting proteins in mammalian cells. J. Cell Biol. 196, 801–810. doi: 10.1083/JCB.201112098
Schenck, A., Bardoni, B., Moro, A., Bagni, C., and Mandel, J. L. (2001). A highly conserved protein family interacting with the fragile X mental retardation protein (FMRP) and displaying selective interactions with FMRP-related proteins FXR1P and FXR2P. Proc. Natl. Acad. Sci. USA 98, 8844–8849. doi: 10.1073/pnas.151231598
Shuster, S. A., Li, J., Chon, U., Sinantha-Hu, M. C., Luginbuhl, D. J., Udeshi, N. D., et al. (2022). In situ cell-type-specific cell-surface proteomic profiling in mice. Neuron 110, 3882–3896.e9. doi: 10.1016/j.neuron.2022.09.025
Soto, J. S., Jami-Alahmadi, Y., Chacon, J., Moye, S. L., Diaz-Castro, B., Wohlschlegel, J. A., et al. (2023). Astrocyte–neuron subproteomes and obsessive–compulsive disorder mechanisms. Nature 616, 764–773. doi: 10.1038/s41586-023-05927-7
Spence, E. F., Dube, S., Uezu, A., Locke, M., Soderblom, E. J., and Soderling, S. H. (2019). In vivo proximity proteomics of nascent synapses reveals a novel regulator of cytoskeleton-mediated synaptic maturation. Nat. Commun. 10, 386–316. doi: 10.1038/s41467-019-08288-w
Sun, X., Sun, H., Han, X., Chen, P. C., Jiao, Y., Wu, Z., et al. (2022). Deep single-cell-type proteome profiling of mouse brain by nonsurgical AAV-mediated proximity labeling. Anal. Chem. 94, 5325–5334. doi: 10.1021/acs.analchem.1c05212
Sunna, S., Bowen, C., Zeng, H., Rayaprolu, S., Kumar, P., Bagchi, P., et al. (2023). Cellular proteomic profiling using proximity labeling by TurboID-NES in microglial and neuronal cell lines. Mol. Cell. Proteom. 22:100546. doi: 10.1016/j.mcpro.2023.100546
Takano, T., and Soderling, S. H. (2021). Tripartite synaptomics: cell-surface proximity labeling in vivo. Neurosci. Res. 173, 14–21. doi: 10.1016/J.NEURES.2021.05.002
Takano, T., Wallace, J. T., Baldwin, K. T., Purkey, A. M., Uezu, A., Courtland, J. L., et al. (2020). Chemico-genetic discovery of astrocytic control of inhibition in vivo. Nature 588, 296–302. doi: 10.1038/s41586-020-2926-0
Tang, Q., and Chen, X. (2023). Nascent proteomics: chemical tools for monitoring newly synthesized proteins. Angew. Chem. Int. Ed. 62:e202305866. doi: 10.1002/ANIE.202305866
Uezu, A., Kanak, D. J., Bradshaw, T. W. A., Soderblom, E. J., Catavero, C. M., Burette, A. C., et al. (2016). Identification of an elaborate complex mediating postsynaptic inhibition. Science 353, 1123–1129. doi: 10.1126/science.aag0821
van Oostrum, M., Blok, T. M., Giandomenico, S. L., tom Dieck, S., Tushev, G., Fürst, N., et al. (2023). The proteomic landscape of synaptic diversity across brain regions and cell types. Cell 186, 5411–5427.e23. doi: 10.1016/J.CELL.2023.09.028
Whittaker, V. P., Michaelson, I. A., and Kirkland, R. J. (1964). The separation of synaptic vesicles from nerve-ending particles (‘synaptosomes’). Biochem. J. 90, 293–303. doi: 10.1042/BJ0900293
Wilkinson, B., Li, J., and Coba, M. P. (2017). Synaptic GAP and GEF complexes cluster proteins essential for GTP signaling. Sci. Rep. 7, 5272–5212. doi: 10.1038/s41598-017-05588-3
Xu, Y., Song, X., Wang, D., Wang, Y., Li, P., and Li, J. (2021). Proteomic insights into synaptic signaling in the brain: the past, present and future. Mol. Brain 14, 37–18. doi: 10.1186/S13041-021-00750-5
Yoshino, Y., Roy, B., Kumar, N., Shahid Mukhtar, M., and Dwivedi, Y. (2021). Molecular pathology associated with altered synaptic transcriptome in the dorsolateral prefrontal cortex of depressed subjects. Transl. Psychiatry 11, 73–16. doi: 10.1038/s41398-020-01159-9
Zhang, J., Wang, G., He, W. W., Losh, M., Berry-Kravis, E., and Funk, W. E. (2019). Expression and characterization of human fragile X mental retardation protein isoforms and interacting proteins in human cells. Proteom. Insights 10, 1–9. doi: 10.1177/1178641818825268
Keywords: synapse, BioID, APEX, proteomics, astrocyte, neuron, TurboID, Split-TurboID
Citation: Ito Y, Nagamoto S and Takano T (2024) Synaptic proteomics decode novel molecular landscape in the brain. Front. Mol. Neurosci. 17:1361956. doi: 10.3389/fnmol.2024.1361956
Edited by:
Kwok-On Lai, City University of Hong Kong, Hong Kong SAR, ChinaReviewed by:
Dan Ohtan Wang, New York University Abu Dhabi, United Arab EmiratesCopyright © 2024 Ito, Nagamoto and Takano. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Tetsuya Takano, dGV0c3V5YS50YWthbm9AYmlvcmVnLmt5dXNodS11LmFjLmpw
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.