 Maroun Bou Zerdan
Maroun Bou Zerdan Hazem I. Assi
Hazem I. Assi
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Mol. Neurosci., 05 October 2021
Sec. Molecular Signalling and Pathways
Volume 14 - 2021 | https://doi.org/10.3389/fnmol.2021.722396
Anaplastic oligodendrogliomas are a type of glioma that occurs primarily in adults but are also found in children. These tumors are genetically defined according to the mutations they harbor. Grade II and grade III tumors can be differentiated most of the times by the presence of anaplastic features. The earliest regimen used for the treatment of these tumors was procarbazine, lomustine, and vincristine. The treatment modalities have shifted over time, and recent studies are considering immunotherapy as an option as well. This review assesses the latest management modalities along with the pathways involved in the pathogenesis of this malignancies.
Oligodendroglial tumors are rare tumors that constitute part of the neuro epithelial tumors of the central nervous system. Accounting to up to 5% of all neuroepithelial tumors (Ostrom et al., 2017), oligodendroglial tumors have an incidence rate of around 1,000 new cases per year in the United States. Oligodendroglial tumors can be divided into two groups based on the classification of the world health organization (WHO): grade II oligodendroglioma and grade III (anaplastic) oligodendroglioma. Most commonly occurring between 25 and 45 years of age, grade III oligodendrogliomas tend to present 10 years later than grade II tumors and can rarely develop in younger and older populations. Oligodendroglioma is genetically defined as a tumor confirmed to harbor either an IDH1 or IDH2 mutation along with co-deletion of chromosome arms 1p and 19q. Histologically, oligodendroglial tumors show sheets of isomorphic round nuclei with a clear cytoplasm—the classic “fried egg” appearance. Grade III oligodendroglioma show a worse prognosis than grade II tumors due to the presence of anaplastic features such as nuclear atypia, necrosis, microvascular proliferation, high cell density and number of mitotic figures. It is believed that anaplastic oligodendroglioma (AO) can progress from a lower grade oligodendroglioma after the acquisition of specific genetic alterations (Youssef and Miller, 2020). However, a clear distinction of both grades is not always possible. Increasing interest has been projected toward favorable molecular markers which oligodendrogliomas harbor. In this review article, we describe the clinical management of AO and summarize the different molecular pathways that drive the development, maintenance, and treatment response of these tumors.
To establish the diagnosis of AO, a pathological sample is crucial. Hence, surgeons should biopsy patients suspected to have AO and attempt tumor resection, as with all other high-grade gliomas. In a study by Shin et al. (2020), gross tumor resection (GTR) was done in 43 of 88 patients. Upon multivariate analysis, median progression free survival (PFS) was 41.1 vs. 23.9 months along with a hazard ratio (HR) of 0.58 with a 95% CI 0.35–0.97 (p = 0.038) compared to patients who had no GTR (Shin et al., 2020). However, upon multivariate analysis there was no significant difference in overall survival (OS). Similarly in a retrospective study by Fujii et al. (2017) patients with anaplastic astrocytoma or anaplastic oligoastrocytoma but not AO had a significant survival advantage when resection of at least 53% of the preoperative T2-weighted high-signal intensity volume was done. Alattar et al. (2018) conducted a Surveillance, Epidemiology, and End Results (SEER)-based analysis in 2017 and showed that GTR was not associated with improved survival in patients with WHO grade II and grade III oligodendrogliomas compared to patients with anaplastic astrocytomas and glioblastomas. This was attributed to the sensitivity of oligodendrogliomas to chemotherapy compared to astrocytomas (Alattar et al., 2018).
Although surgery can help relieve symptoms by decreasing the mass effect of the tumor, the tumor’s predilection to the frontal lobe hinders its maximal resection. This comes with a risk of sacrificing important brain centers and hence compromising functionality and quality of life. Retrospective studies have revealed that the post-operative seizure-free rate is 67–80% (Luyken et al., 2003; Zaatreh et al., 2003; Benifla et al., 2006; Chang et al., 2008; Englot et al., 2011). Despite utilizing a multimodal approach in nearly all patients, refractory seizures can still be seen in patients suffering from epilepsy in 50% of the cases before the initial surgery and 15–40% of cases following surgery and anticonvulsant therapy (Smits and Duffau, 2011; You et al., 2011; Calatozzolo et al., 2012). Two plausible hypotheses to explain treatment resistance in oligodendrogliomas exist. The first is the presence of alterations in drug targets affecting antiepileptic drugs’ binding. The second is diminished intracellular drug transport through the overexpression of ATP-binding cassette transporter proteins such as P-gp (MDR1), MRP1, and MRP5 (Calatozzolo et al., 2012; Alms et al., 2014).
Postoperative radiotherapy (XRT) to a total dose of roughly 60 Gy over 30 fractions is recommended (Blakeley and Grossman, 2008). Although one survey showed that 34% of neuro-oncologists suggested delaying XRT in patients with 1p19 co-deletions (Abrey et al., 2007), clinical trials addressing the efficacy of delayed XRT in this subset of patients are needed.
The earliest reported results of the chemotherapy regimens, procarbazine, lomustine (CCNU), and vincristine (PCV), in AO were reported by Cairncross et al. (1994) and showed that the median time to progression for patients was at least 25.2 months for complete responders, 14.2 months for partial responders and 6.8 months for stable patients. Afterward in 2001, Chinot et al. (2001) reported that 16.7% of patients experienced a complete response and 27.1% experienced a partial response when receiving temozolomide (TMZ) after previous PCV. The radiation Therapy Oncology Group (RTOG) also explored the use of pre-irradiation TMZ followed by concurrent TMZ and radiotherapy in a phase 2 study (RTOGBR013) (Vogelbaum et al., 2009).
Accordingly, the treatment approach is tailored according to the presence of 1p19q co-deletion, which characterizes oligodendrogliomas. Patients harboring co-deleted tumors can receive either PCV or TMZ. The European Organization for Research and Treatment of Cancer study 26951 (EORTC26951) and RTOG9402 showed an increase in OS and PFS when PCV is added to radiotherapy (RT) in patients with 1p19q co-deleted oligodendrogliomas (Cairncross et al., 2013; van den Bent et al., 2013a). With almost 12 years of follow-up, patients harboring tumors with 1p19q co-deletions showed an improved survival when treated with PCV and RT as compared to RT alone (EORTC26951: 157 vs. 50 months; RTOG9402: 14.7 vs. 7.3 years). Moreover, treatment of these patients with PCV demonstrated an improved OS in both groups when compared to RT alone. In patients with astrocytic tumors, only PFS was prolonged in patients treated with XRT who received up-front PCV vs. PCV at the time of recurrence (Pan-Weisz, 2019; Tork and Atkinson, 2020). Consequently, and in terms of improvement in quality of life (QOL), the EORTC study showed no difference between the two groups, and PCV toxicity contributed to a decreased QOL for a prolonged period.
The response of tumors harboring IDH mutations to PCV therapy has also been described in a subset analysis and follow-up study of RTOG9402 trial. As expected, patients with an IDH mutation and 1p19q co-deletion showed significant benefit in OS. While IDH-WT tumors retained a poor prognosis and showed no benefit from PCV treatment, improved OS was seen in IDH mutant non-co-deleted tumors, and astrocytic tumors when treated with PCV plus RT.
A subset analysis of patients with other methylation profiles, such as CpGisland hypermethylated phenotype (CIMP) and MGMT promoter methylation (MGMT-STP27) status, was also conducted by van den Bent et al. (2013b). It was found that CIMP + or MGMT-STP27 methylated tumors had a superior OS 1.05 vs. 6.46 years and 1.06 vs. 3.8 years (both P < 0.0001) for CIMP and MGMT-STP27 status, respectively. CIMP + and MGMT-STP27 methylated tumors had a clear benefit from adjuvant PCV; the median OS in the RT and RT-PCV arms was 3.27 vs. 9.51 years (P < 0.0033), respectively for CIMP + tumors and 1.98 vs. 8.65 years (P < 0.0001) for MGMT-STP27 methylated tumors (van den Bent et al., 2013b). There was however no such benefit for CIMP- or for MGMT-STP27 unmethylated tumors.
Adjuvant TMZ has also been shown to be effective with better tolerability and less toxicity (van den Bent et al., 2003; Brandes et al., 2006). A randomized clinical trial is currently in progress to compare the efficacy of PCV or TMZ when combined with RT in 1p19q co-deleted tumors (CODEL: NCT00887146). Preliminary results are mentioned toward the end of the manuscript. For patients with astrocytic tumors, EORTC26951 and RTOG9402 did not show any benefit of PCV with RT. A trial of adjuvant TMZ with RT in patients harboring this tumor subtype showed a significantly improved PFS and OS (van den Bent et al., 2017). While increasing the risk of toxicity, concurrent TMZ is currently being assessed in comparison to adjuvant treatment in astrocytic tumors (van den Bent et al., 2017). The interim report from the RTOG0131 trial suggests that combination therapy with TMZ and XRT is well tolerated in patients with AO being treated with neoadjuvant TMZ for 6 months, followed by TMZ and concurrent XRT (Tork and Atkinson, 2020). Apart from RTOG9402 and EORTC26951, Wick et al., 2016 conducted NOA-4, a randomized phase 3 trial of sequential RT followed by chemotherapy against anaplastic glioma with PCV or TMZ (Vogelbaum et al., 2009). In this trial, MGMT hypermethylation was associated with prolonged PFS in both arms (Wick et al., 2009; Tork and Atkinson, 2020).
All in all, patients with 1p19q co-deleted tumors should be treated with RT and adjuvant PCV while those lacking this co-deletion should receive adjuvant TMZ. PCV and TMZ are also used in cases of recurrence but result in lower response rates and disease-free survival. Other agents have also been investigated for recurring disease including paclitaxel, irinotecan, carboplatin, etoposide, and cisplatin (Poisson et al., 1991; Yung et al., 1991; Warnick et al., 1994; Chamberlain and Kormanik, 1995, 1999; Fulton et al., 1996; Macdonald et al., 1996; Friedman et al., 1999; Chang et al., 2001; Cloughesy et al., 2003; Batchelor et al., 2004; Ascierto et al., 2016). However, no results have proven enough benefit for treating patients with recurrent AO. Table 1 outlines some information related to the major drugs used in treatment.
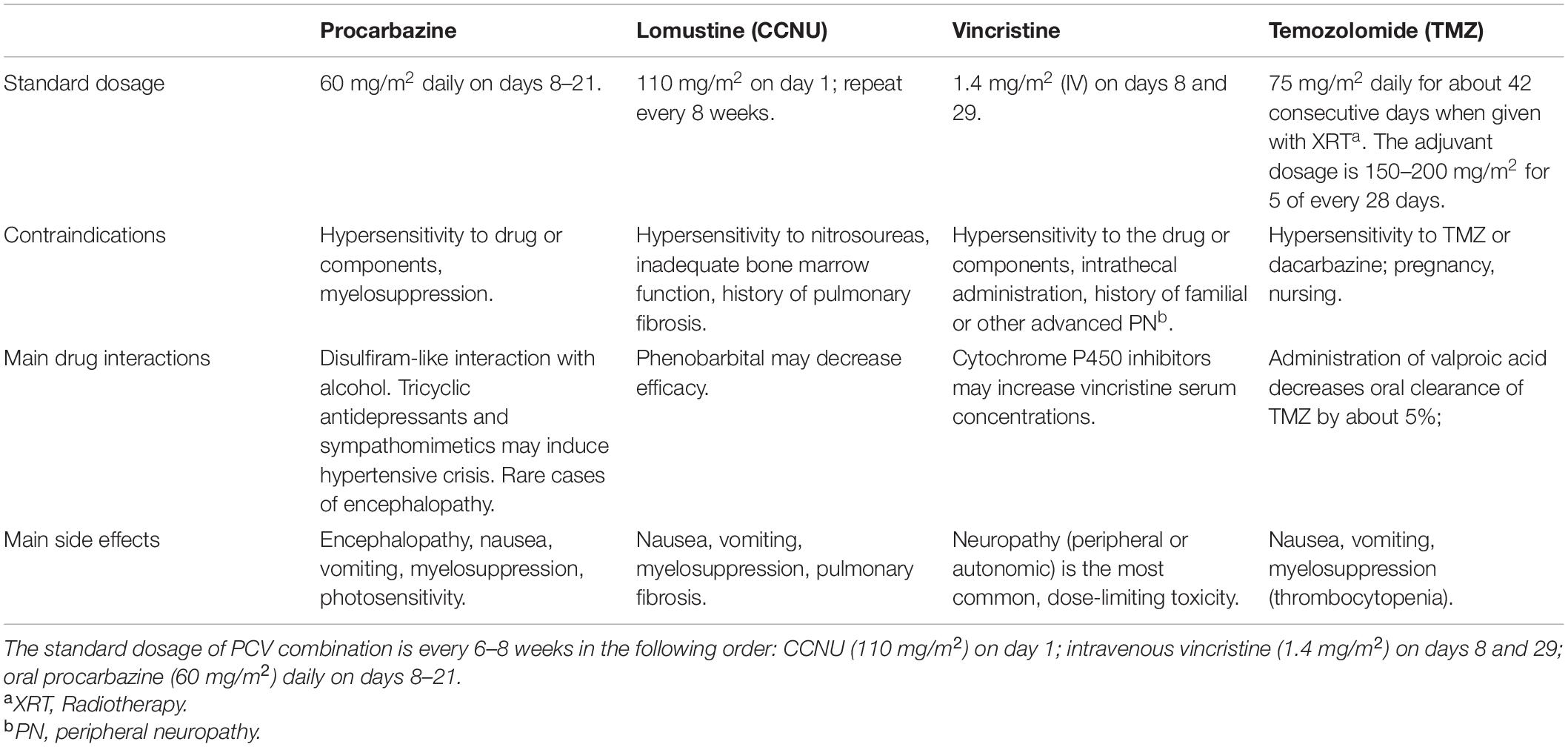
Table 1. Major drugs utilized in the treatment of AO.
Recently, immunotherapy has been explored as a potential treatment modality. Elens et al. (2012) reported the survival benefit of immunotherapy in patients with relapsed AO enrolled in the HGG-IMMUNO-2003 trial. The PFS and OS were 3.4 and 18.8 months, respectively. In a recent case report by Yu et al. (2021) a patient who had multiple tumor recurrences, following several regimens was started eventually on nivolumab. On magnetic resonance imaging, he was considered to have disease progression. Upon surgical debulking and pathological diagnosis, he was found to have recurrent diseases. However, tumor samples collected from enhancing and non-enhancing areas for a scRNAseq analysis revealed an abundance of immune cells. Infiltration of these cells might have been perceived as the increased mass on MRI. The patient sustained a disease-free response to nivolumab at least 12 months after surgery. This highlights the importance in incorporating novel techniques to better understand the tumor microenvironment (Yu et al., 2021).
The main function of the IDH1 and IDH2 enzymes is the oxidative decarboxylation of isocitrate to alpha-ketoglutarate. This reaction promotes the formation of NADPH, the reduced form of NADP+, which helps in protecting the cell from oxidative radicals that can damage DNA (Soffietti et al., 1998; van den Bent et al., 1998). In the cytosol, the product of the reaction catalyzed by IDH1, alpha-KG, has been reported to be involved in multiple cellular pathways including hypoxia sensing, lipogenesis and epigenetic modification through its action on alpha-KG dependent dioxygenases such as TET and JmjC and other enzymes (Mason et al., 1996; Buckner et al., 2003; Abrey et al., 2006; Taliansky-Aronov et al., 2006). The role of IDH2, on the other hand, is limited to the mitochondria where it catalyzes the same reaction as part of the tricarboxylic acid cycle (TCA).
IDH mutations identified in gliomas tend to occur at the active site of the enzyme at arginine 132 and 172 in IDH1 and IDH2, respectively. IDH mutations can dominantly inhibit WT-IDH when heterozygous through the formation of enzymatically inactive heterodimers (Zhao et al., 2009). It was shown by Uhm (2010) that IDH mutations lead to the acquisition of a new enzymatic function that catalyzes the formation of D-2HG from alpha-KG. 2-HG can inhibit alpha-KG dependent dioxygenases and cause epigenetic alterations (Xu et al., 2011). It can also stimulate the activity of EGLN leading to decreased HIF levels. This in turn allows tumor proliferation in low oxygen conditions (Zhao et al., 2009; Koivunen et al., 2012). It has also been reported that 2-HG can inhibit p53 via microRNA activated by HIF-2α, driving tumorigenesis (Jiang et al., 2018). As a result of the disruption of IDH’s enzymatic function, 2-HG tilts off the NADP/NADPH balance thereby increasing the production of ROS and leading to DNA damage and tumor formation (Latini et al., 2003; Rinaldi et al., 2016). JmjC demethylases are one of the many dioxygenases regulated by α-KG and inhibited by 2-HG. They are responsible for histone methylation on lysine residues. It has been observed that in IDH-mutant cell lines, repressive histone methylation precedes global DNA hypermethylation. This in turn provides evidence that IDH mutations could allow cells to remain in a vulnerable state, and prone to additional DNA alterations. Mutant IDH1 has also been shown to inhibit the ALkB family DNA repair enzymes further contributing to erroneous DNA replication (Wang et al., 2015; Rinaldi et al., 2016).
High mutant allele fractions have been found in patient samples at diagnosis and recurrence in tumor evolution studies. IDH1 mutations seem to be at the core of this tumorigenesis (Johnson et al., 2014). IDH mutated enzymes can promote proliferation and colony formation through its end metabolite 2-HG (Koivunen et al., 2012; Bittinger et al., 2013). Turcan et al. (2012) showed that an IDH1 mutation can induce a methylation profile known as the G-CIMP signature, which is a glioma specific methylation pattern at CpG islands. Interestingly, an in vitro treatment of cells with D-2HG also induced a similar methylation pattern (Lu et al., 2012) which further supports the vital role of this metabolite in epigenetic alteration and tumor formation. Additionally, hypermethylation caused by IDH1 mutations was shown to occur at CTCF-binding sites that normally insulate and prevent the interaction between different parts of the genome (Flavahan et al., 2016). Methylation of these sites promotes the interaction of enhancers with new genes (Flavahan et al., 2016).
IDH mutations have also been implicated in the regulation of the recruitment of inflammatory cells to tumor sites, specifically through D-2HG. Evidence from in vivo models have demonstrated reduced levels of STAT1 and CXCL10 in IDH-mutant gliomas. Infiltration of immune cells, specifically T cells, were also reduced in these tumors (Amankulor et al., 2017; Kohanbash et al., 2017). Additionally, the mTOR pathway has been identified at a potential target for treatment due its activation in IDH-mutant gliomas. This occurs via 2-HG’s inhibition of KDM4A, an α-KG dependent deoxygenase, and destabilization of DEPTOR, a negative regulator of mTORC1/2, resulting in mTOR pathway activation (Carbonneau et al., 2016). This activation is of special interest since it has been shown that mTOR and its downstream effectors are implicated in tumorigeneses in brain malignancies (Fan and Weiss, 2010; Ryskalin et al., 2017).
The RTOG 9802 trial, which included non-molecularly stratified patients harboring grade II gliomas, demonstrated a 5.5-year survival benefit of PCV administration (Shaw et al., 2008). Results of this trial raise the possibility that the chemosensitivity seen in these tumors might be due to the IDH mutation that is common to both oligodendroglial and low-grade astrocytic gliomas. Upon reanalysis of RTOG 9802 after molecular classification, AO patients with IDH-mutated tumors actually showed a survival benefit when treated with PCV chemotherapy (Cairncross et al., 2014). However, analysis of other trials such as the EORTC 26951 did not reveal a correlation between IDH mutations and survival in patients with astrocytic tumors (grade II) (van den Bent et al., 2010, 2013a). Appropriate design of future clinical trials can help in determining better correlations with molecular subclasses.
The unbalanced translocation of the centromeric regions of chromosomes 1p and 19q attribute to the loss of the whole arm on both chromosomes. This co-deletion, along with the IDH mutation, enables a tumor to be classified as an oligodendroglioma according to the WHO 2016 criteria (Louis et al., 2016). Patients with co-deleted tumors demonstrate favorable prognoses (Smith et al., 2000a; Ino et al., 2001; Cairncross et al., 2006; Kaloshi et al., 2007; Cairncross et al., 2013). The mechanism by which this co-deletion leads to chemosensitivity remains unclear and data showing the implication of other genes in this chemosensitivity is emerging.
Following the stratification of AO according to 1p/19q co-deletion status, an in-depth genetic analysis of 1p/19q co-deleted tumors revealed inactivating mutations affecting the FUBP1 gene on chromosome 1p and the CIC gene on chromosome 19 (Bettegowda, 2000; Sahm et al., 2012; Yip et al., 2012). CIC normally functions as a reversible repressor by binding to the DNA regulatory elements downstream of growth factor signaling pathways (Ajuria et al., 2011). Acting as a tumor-suppressor gene, missense mutations affecting CIC are mostly found within the DNA-binding domain thereby inhibiting its binding to regulatory elements. In a population of patients with oligodendroglial tumors, four cases exhibited absent CIC expression with no detectable mutations, suggesting that alterations affecting CIC could occur through other unidentified mechanisms (Chan et al., 2014). Another DNA-binding protein found mutated in AO is FUBP1. The Far Upstream Element (FUSE) Binding Protein 1 (FUBP1) is known to regulate several cell cycle regulators such as MYC and p21. While often found upregulated in many tumors, FUBP1 acts as a tumor suppressor gene due to its inactivating mutations reported in around 15% of oligodendroglial tumors (Baumgarten et al., 2014). As for the clinical relevance of these molecular markers, inactivating mutations affecting FUBP1 have correlated with a shorter time to recurrence and CIC mutations have been associated with worse prognosis, especially in those patients with 1p/19q co-deleted oligodendrogliomas (Chan et al., 2014; Michaud et al., 2018). Nevertheless, further studies are needed to elucidate the role of CIC/FUBP1 alterations in the pathogenesis of AO and oligodendrogliomas, in general.
In addition to the aforementioned pathways, homozygous and the less common hemizygous losses of 9p21 have been reported with high frequencies in gliomas, and up to 55% in AO (Maruno et al., 1996; Perry et al., 1999; Rasheed et al., 2002; Ohgaki and Kleihues, 2009; Michaud et al., 2018). These alterations have correlated with a shorter event free survival (EFS; 29 vs. 53 months, p < 0.0001) and OS (48 vs. 83 months, p < 0.0001). At the molecular level, 9p losses result in the loss of the cyclin-dependent kinase inhibitor CDKN2A gene, which normally inhibits cellular division. CDKN2A inhibits the interaction between the cyclin dependent-kinases CDK4 or CDK6 and D-type cyclins, preventing both the phosphorylation of the retinoblastoma (RB1) protein and the release of the elongation factor (EF2) (Weinberg, 1995; Sherr and Roberts, 1999). Hence, cellular proliferation and dysregulation of pro-apoptotic pathways ensues (Ruas and Peters, 1998). Although 9p losses can be found in many gliomas, they more commonly occur in higher grade tumors (grades 3 and 4), which make the CDKN2A gene or p16 protein (CDKN2A product) potential players involved in the malignant progression and anaplastic transformation of low-grade gliomas into higher grades (He et al., 1995; Ueki et al., 1996; Watanabe et al., 2001). Interestingly, some tumors exhibited p16 hyperexpression without any chromosome 9p alterations and this was associated with a shorter EFS and OS. Cyclin D1 expression was also significantly higher in AO and was associated with a shorter EFS (Michaud et al., 2018).
TCF12 protein is a transcription factor and member of the basic helix-loop-helix (bHLH) E-protein family. Through the formation of homo- and hetero-dimers with other bHLH transcription factors, TCF12 modulates the transcription of specific genes that are intrinsic to the oligodendrocyte lineage (Fu et al., 2009) and are involved in neural development (Uittenbogaard and Chiaramello, 2002). Two main alterations affecting the TCF12 protein have been reported in AO: absence of bHLH DNA-binding domain and single amino acid substitutions such as R602M within the bHLH domain. Both types of alterations have been shown to drastically impact the ability of TCF12 to function as a transcription factor and interact with other bHLH proteins, eventually leading to mutant protein accumulation (Labreche et al., 2015). Patients harboring TCF12 mutations or LOH exhibited a shorter median OS. The frequency of these alterations was much higher in grade III AO as compared to grade II oligodendroglioma. This suggests that TCF12 alterations play a role in dictating an aggressive phenotype in AO. One analysis looking at the downstream effect of TCF12 alterations showed a downregulation of TCF21, EZH2, and BMI1 pathway and especially CDH1 (E-cadherin), which has been shown to be implicated in tumor characteristics and metastasis (Lee et al., 2012). Interestingly, it has been reported that TCF12 may have a haploinsufficient tumor suppressor role which increases the risk of developing AO in those patients harboring a TCF12 germline mutation.
Human telomerase reverse transcriptase (TERT) mutations have been found to be present in 77% of grades II and III oligodendrogliomas and 82% of tumors with 1p19q co-deletion (Koelsche et al., 2013). Telomerase reverse transcriptase is a subunit of the enzyme telomerase that protects the overall integrity and length of telomeres. Telomerase normally functions to regenerate chromosomal ends (telomeres) thereby allowing DNA replication and mitosis. While usually unexpressed in mature cells, cancer cells make use of this enzyme to promote their survival and increase proliferation. TERT mutations in glioma are often found within the promoter region. This results in the opening of a binding site for the E26 transformation-specific transcription factors (Killela et al., 2013). TERT reactivation then takes place when GA-binding protein (GABP) transcription factor binds to the mutant TERT promoter (Dahlin et al., 2016).
In addition to being a surrogate for oligodendroglial lineage, TERT mutations seem to have some prognostic significance (Dahlin et al., 2016). Pekmezci et al. (2017) studied the status of both TERT and ATRX mutations along with their prognostic values in cohorts including grade II/III astrocytomas. The wildtype (WT) TERT group was associated with good prognosis only in IDH1/IDH2 WT (IDH-WT) grade II/III astrocytomas. However, in those groups with IDH mutations, including AO, TERT promoter mutation status was not a statistically significant prognostic factor (Dahlin et al., 2016). Thus, prognostic markers should be assessed while accounting for other genetic alterations.
Lately, IDH 1 and 2, which are known to generate nicotinamide adenine dinucleotide phosphate (NADPH), have been heavily observed. Their predictive value stems from their close relationship to human gliomas. Zou et al. (2013) was the first to conduct a meta-analysis on PFS and OS in gliomas based on IDH mutations. A better outcome was associated with IDH mutations and a combined HR estimate for OS and PFS was 0.33 (95% CI: 0.25–0.42) and 0.38 (95% CI: 0.21–0.68) for patients with gliomas harboring IDH mutation (Zou et al., 2013). A study by Kaminska et al. (2019) depicted how the mutant IDH1 (R132H) blocks cellular differentiation and contributes to antitumor immunity. Although a mutated IDH1 cannot generate NADPH since it has lost its normal catalytic activity, it gains the function of producing D-(R)-2-hydroxyglutarate. When the latter is overproduced in cancer cells, it inhibits histone and DNA methylases and interferes with cellular metabolism. The end result is DNA hypermethylation and thus the blockage of cellular differentiation (Kaminska et al., 2019).
The platelet-derived growth factor (PDGF) signaling system has been associated with the development and malignant progression of AO. Overexpression of PDGF system components, particularly the α subtype receptor (PDGFRα), was detected in Southern and Fluorescence in situ hybridization (FISH) analyses 4/41 AO. Although these tumors were not examined for correspondence between PDGFRα expression and PDGFRα gene amplification, application of the same methodology on studies involving EGFR indicate that a high level of protein expression is to be expected in the future (Smith et al., 2000b).
Finally, even though PTEN gene alterations have an unclear association with AO, their function in the control of cellular proliferation could explain their role in pathogenesis of AO. Sasaki et al. (2001) showed that 7/72 AO had PTEN gene alterations; 2 had homozygous DMBT1 deletions, but at least one reflected unmasking of a germline DMBT1 deletion. Moreover, no mutations were found in ERCC6 exon 2 and only two patients had a chemotherapeutic response, but with unexpected short survival times. Therefore, PTEN is a target of 10q loss, and PTEN alterations are associated with aggressive tumor phenotypes regardless of chemosensitivity (Sasaki et al., 2001).
There are 11 ongoing clinical trials recruiting patients with AO. NCT03971734 aims to determine the optimal dose of Regadenoson which alters the integrity of the Blood-Brain-Barrier in patients with high grade gliomas. Another phase 2 clinical trial (NCT04623931) is assessing chemotherapy and RT for the treatment of IDH wildtype gliomas or non-histological glioblastomas in approximately 40 patients. In an ongoing phase 3 study (NCT00887146), patients with AO or low-grade gliomas were split into two arms. Patients in arm A received RT with concomitant TMZ followed by adjuvant TMZ. Patients in arm B received RT first followed by PCV chemotherapy. Another clinical trial is a pediatric long-term follow-up and rollover phase 4 study (NCT03975829), whereby approximately 250 participants will be treated with dabrafenib and/or tametinib. NCT03434262 is a phase 1 study assessing the efficacy of different drugs on children and young adults. Each stratum has different combination treatments and targeted patient populations. While ribociclib is included as a treatment regimen across all strata, gemcitabine, trametinib, and sonidgib are included in strata A, B, and C, respectively. Elsewhere, another phase 1 study (NCT02644291) is assessing the use of mebendazole in recurrent/progressive pediatric brain tumors of 21 participants. Periclinal laboratory models have shown the efficacy of mebendazole against high grade gliomas and medulloblastomas.
NCT01849952 is another clinical trial that will evaluate the expression levels of microRNA-10b in patients with AO, although it will not involve any new therapeutic regimens. Investigators of this trial will be testing the in vitro sensitivity of individual primary tumors to anti-mir-10b treatment. Another currently ongoing phase I study (NCT04135807) is assessing the efficacy of an implantable microdevice in the brain before tumor resection is initiated. This microdevice will be used for 8 intratumor drugs: TMZ, Lomustine, Irinotecan, Carboplatin, Lapatinib, Osimertinib, Abenaciclib, and Everolimus. NCT04708548 is an ongoing European cross-sectional study that is looking at health-related quality of parameters and outcomes in survivors after being treated with surgery, chemotherapy and/or RT. The estimated completion date is August 2022.
NCT04541082 is another ongoing phase 1 study aiming to determine the maximum tolerated dose of the oral drug ONC206, a member of the imipridone class of anti-cancer small molecules which target G protein-coupled receptors. This trial aims to determine the maximum tolerated dose of ONC206. The efficacy and safety of other novel therapeutic drugs such as rQNestin34.5v.2 (an oncolytic viral vector) is also being assessed. As part of an ongoing phase 1 trial to treat recurrent malignant gliomas (NCT03152318), investigators hope that the rQNestin34.5v.2 drug will spread to a glioma cell, kill it, and then make a copy of itself and spread again. With approximately 108 participants included in this study, the estimated completion date is July 2022.
The CODEL study is a phase 3 study whereby 36 patients with newly diagnosed grade III oligodendrogliomas were randomized to receive RT alone (Arm A), RT with concomitant and adjuvant TMZ (Arm B) or TMZ alone (Arm C) (Jaeckle et al., 2021). At a median follow up of 7.5 years, around 80% (n = 10) patients in Arm C progressed vs. approximately 40% (n = 9) in the other arms. The HR was 3.12 with a 95% CI of 1.26–7.19 (P = 0.014) (Jaeckle et al., 2021). Even though there wasn’t any difference in OS, the PFS remained shorter for patients not receiving any RT; even after adjusting for IDH status and RT treatment status. The PFS HR was 3.33 with a 95% CI 1.31–8.45 (P = 0.011) while the OS HR was 2.78 with a 95% CI 0.58–13.22 (P = 0.20) (Jaeckle et al., 2021).
Lastly, it is worth noting that there are approximately 230 other clinical trials which involve oligodendrogliomas but are not actively recruiting patients.
AO remains an understudied tumor with several unclear pathogenic pathways. Several genetic and protein alterations have been identified in AO. Some of these alterations have correlated with prognosis and response to treatment. Re-analysis of some trials prior to the 2016 WHO brain tumor classification has given further insight into some molecular pathways that were previously poorly defined or investigated. More studies, however, are needed to explore molecular pathways in oligodendroglioma and AO specifically after the 2016 classification.
MB drafted the manuscript and contributed to the discussion section. HA conceived the idea for the manuscript. Both authors have read and approved the final manuscript.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Abrey, L. E., Childs, B. H., Paleologos, N., Kaminer, L., Rosenfeld, S., Salzman, D., et al. (2006). High-dose chemotherapy with stem cell rescue as initial therapy for anaplastic oligodendroglioma: long-term follow-up. Neuro Oncol. 8, 183–188. doi: 10.1215/15228517-2005-009
Abrey, L. E., Louis, D. N., Paleologos, N., Lassman, A. B., Raizer, J. J., Mason, W., et al. (2007). Survey of treatment recommendations for anaplastic oligodendroglioma. Neuro Oncol. 9, 314–318.
Ajuria, L., Nieva, C., Winkler, C., Kuo, D., Samper, N., Andreu, M. J., et al. (2011). Capicua DNA-binding sites are general response elements for RTK signaling in Drosophila. Development. 138, 915–924. doi: 10.1242/dev.057729
Alattar, A. A., Brandel, M. G., Hirshman, B. R., Dong, X., Carroll, K. T., Ali, M. A., et al. (2018). Oligodendroglioma resection: a Surveillance, Epidemiology, and End Results (SEER) analysis. J. Neurosurg. JNS 128, 1076–1083. doi: 10.3171/2016.11.jns161974
Alms, D., Fedrowitz, M., Römermann, K., Noack, A., and Löscher, W. (2014). Marked differences in the effect of antiepileptic and cytostatic drugs on the functionality of P-glycoprotein in human and rat brain capillary endothelial cell lines. Pharm. Res. 31, 1588–1604. doi: 10.1007/s11095-013-1264-4
Amankulor, N. M., Kim, Y., Arora, S., Kargl, J., Szulzewsky, F., Hanke, M., et al. (2017). Mutant IDH1 regulates the tumor-associated immune system in gliomas. Genes Dev. 31, 774–786. doi: 10.1101/gad.294991.116
Ascierto, P. A., McArthur, G. A., Dréno, B., Atkinson, V., Liszkay, G., Di Giacomo, A. M., et al. (2016). Cobimetinib combined with vemurafenib in advanced BRAFV600-mutant melanoma (coBRIM): updated efficacy results from a randomised, double-blind, phase 3 trial. Lancet Oncol. 17, 1248–1260. doi: 10.1016/s1470-2045(16)30122-x
Batchelor, T. T., Gilbert, M. R., Supko, J. G., Carson, K. A., Nabors, L. B., Grossman, S. A., et al. (2004). Phase 2 study of weekly irinotecan in adults with recurrent malignant glioma: final report of NABTT 97-11. Neuro Oncol. 6, 21–27. doi: 10.1215/s1152851703000218
Baumgarten, P., Harter, P., Tönjes, M., Capper, D., Blank, A. E., Sahm, F., et al. (2014). Loss of FUBP1 expression in gliomas predicts FUBP1 mutation and is associated with oligodendroglial differentiation, IDH1 mutation and 1p/19q loss of heterozygosity. Neuropathol. Appl. Neurobiol. 40, 205–216. doi: 10.1111/nan.12088
Benifla, M., Otsubo, H., Ochi, A., Weiss, S. K., Donner, E. J., Shroff, M., et al. (2006). Temporal lobe surgery for intractable epilepsy in children: an analysis of outcomes in 126 children. Neurosurgery 59, 1203–1214.
Bittinger, M., Su, S., Fantin, V., Zhong, C., Huang, W., Ding, J., et al. (2013). An Inhibitor of Mutant IDH1 Delays. Science 340, 626–630.
Blakeley, J., and Grossman, S. (2008). Anaplastic oligodendroglioma. Curr. Treat. Options Neurol. 10, 295–307.
Brandes, A. A., Tosoni, A., Cavallo, G., Reni, M., Franceschi, E., Bonaldi, L., et al. (2006). Correlations Between O6-Methylguanine DNA Methyltransferase Promoter Methylation Status, 1p and 19q Deletions, and Response to Temozolomide in Anaplastic and Recurrent Oligodendroglioma: A. J. Clin. Oncol. 24, 4746–4753. doi: 10.1200/jco.2006.06.3891
Buckner, J. C., Gesme, D. Jr., O’Fallon, J. R., Hammack, J. E., Stafford, S., Brown, P. D., et al. (2003). Phase II trial of procarbazine, lomustine, and vincristine as initial therapy for patients with low-grade oligodendroglioma or oligoastrocytoma: efficacy and associations with chromosomal abnormalities. J. Clin. Oncol. 21, 251–255. doi: 10.1200/jco.2003.06.023
Cairncross, G., Berkey, B., Shaw, E., Jenkins, R., Scheithauer, B., Brachman, D., et al. (2006). Phase III trial of chemotherapy plus radiotherapy compared with radiotherapy alone for pure and mixed anaplastic oligodendroglioma: Intergroup Radiation Therapy Oncology Group Trial 9402. J. Clin. Oncol. 24, 2707–2714.
Cairncross, G., Macdonald, D., Ludwin, S., Lee, D., Cascino, T., Buckner, J., et al. (1994). Chemotherapy for anaplastic oligodendroglioma. National Cancer Institute of Canada Clinical Trials Group. J. Clin. Oncol. 12, 2013–2021. doi: 10.1200/JCO.1994.12.10.2013
Cairncross, G., Wang, M., Shaw, E., Jenkins, R., Brachman, D., Buckner, J., et al. (2013). Phase III trial of chemoradiotherapy for anaplastic oligodendroglioma: long-term results of RTOG 9402. J. Clin. Oncol. 31:337. doi: 10.1200/jco.2012.43.2674
Cairncross, J. G., Wang, M., Jenkins, R. B., Shaw, E. G., Giannini, C., Brachman, D. G., et al. (2014). Benefit from procarbazine, lomustine, and vincristine in oligodendroglial tumors is associated with mutation of IDH. J. Clin. Oncol. 32:783. doi: 10.1200/jco.2013.49.3726
Calatozzolo, C., Pollo, B., Botturi, A., Dinapoli, L., Carosi, M., Salmaggi, A., et al. (2012). Multidrug resistance proteins expression in glioma patients with epilepsy. J. Neurooncol. 110, 129–135. doi: 10.1007/s11060-012-0946-9
Carbonneau, M., Gagné, L. M., Lalonde, M.-E., Germain, M.-A., Motorina, A., Guiot, M.-C., et al. (2016). The oncometabolite 2-hydroxyglutarate activates the mTOR signalling pathway. Nat. Commun. 7, 1–12. doi: 10.1038/ncomms12700
Chamberlain, M. C., and Kormanik, P. (1995). Salvage chemotherapy with paclitaxel for recurrent primary brain tumors. J. Clin. Oncol. 13, 2066–2071. doi: 10.1200/jco.1995.13.8.2066
Chamberlain, M. C., and Kormanik, P. (1999). Salvage chemotherapy with taxol for recurrent anaplastic astrocytomas. J. Clin. Oncol. 43, 71–78.
Chan, A. K.-Y., Pang, J. C.-S., Chung, N. Y.-F., Li, K. K.-W., Poon, W. S., Chan, D. T.-M., et al. (2014). Loss of CIC and FUBP1 expressions are potential markers of shorter time to recurrence in oligodendroglial tumors. Mod. Pathol. 27, 332–342. doi: 10.1038/modpathol.2013.165
Chang, E. F., Potts, M. B., Keles, G. E., Lamborn, K. R., Chang, S. M., Barbaro, N. M., et al. (2008). Seizure characteristics and control following resection in 332 patients with low-grade gliomas. J. Neurosurg. 108, 227–235. doi: 10.3171/JNS/2008/108/2/0227
Chang, S. M., Kuhn, J. G., Robins, H. I., Schold, S. C. Jr., Spence, A. M., Berger, M. S., et al. (2001). A Phase II study of paclitaxel in patients with recurrent malignant glioma using different doses depending upon the concomitant use of anticonvulsants: a North American Brain Tumor Consortium report. Cancer 91, 417–422. doi: 10.1002/1097-0142(20010115)91:2<417::aid-cncr1016>3.0.co;2-9
Chinot, O.-L., Honore, S., Dufour, H., Barrie, M., Figarella-Branger, D., Muracciole, X., et al. (2001). Safety and efficacy of temozolomide in patients with recurrent anaplastic oligodendrogliomas after standard radiotherapy and chemotherapy. J. Clin. Oncol. 19, 2449–2455. doi: 10.1200/JCO.2001.19.9.2449
Cloughesy, T. F., Filka, E., Kuhn, J., Nelson, G., Kabbinavar, F., Friedman, H., et al. (2003). Two studies evaluating irinotecan treatment for recurrent malignant glioma using an every-3-week regimen. Cancer Interdiscipl. Int. J. Am. Cancer Soc. 97, 2381–2386. doi: 10.1002/cncr.11306
Dahlin, A. M., Wibom, C., Ghasimi, S., Brännström, T., Andersson, U., and Melin, B. (2016). Relation between established glioma risk variants and DNA methylation in the tumor. PLoS One 11:e0163067. doi: 10.1371/journal.pone.0163067
Elens, I., De Vleeschouwer, S., Pauwels, F., and Van Gool, S. (2012). Resection and immunotherapy for recurrent grade III glioma. Int. Scholarly Res. Notices 2012:530179.
Englot, D. J., Berger, M. S., Barbaro, N. M., and Chang, E. F. (2011). Predictors of seizure freedom after resection of supratentorial low-grade gliomas: a review. J. Neurosurg. 115, 240–244. doi: 10.3171/2011.3.JNS1153
Fan, Q.-W., and Weiss, W. A. (2010). Targeting the RTK-PI3K-mTOR axis in malignant glioma: overcoming resistance. Phosphoinositide 3-Kinase Health Dis. 2010, 279–296. doi: 10.1007/82_2010_67
Flavahan, W. A., Drier, Y., Liau, B. B., Gillespie, S. M., Venteicher, A. S., and Stemmer-Rachamimov, A. O. (2016). Insulator dysfunction and oncogene activation in IDH mutant gliomas. Nature 529, 110–114. doi: 10.1038/nature16490
Friedman, H., Petros, W. P., Friedman, A. H., Schaaf, L. J., Kerby, T., Lawyer, J., et al. (1999). Irinotecan therapy in adults with recurrent or progressive malignant glioma. J. Clin. Oncol. 17, 1516–1525. doi: 10.1200/jco.1999.17.5.1516
Fu, H., Cai, J., Clevers, H., Fast, E., Gray, S., Greenberg, R., et al. (2009). A genome-wide screen for spatially restricted expression patterns identifies transcription factors that regulate glial development. J. Neurosci. 29, 11399–11408. doi: 10.1523/JNEUROSCI.0160-09.2009
Fujii, Y., Muragaki, Y., Maruyama, T., Nitta, M., Saito, T., Ikuta, S., et al. (2017). Threshold of the extent of resection for WHO Grade III gliomas: retrospective volumetric analysis of 122 cases using intraoperative MRI. J. Neurosurg. 129, 1–9. doi: 10.3171/2017.3.JNS162383
Fulton, D., Urtasun, R., and Forsyth, P. (1996). Phase II study of prolonged oral therapy with etoposide (VP16) for patients with recurrent malignant glioma. J. Neurooncol. 27, 149–155. doi: 10.1007/BF00177478
He, J., Olson, J. J., and James, C. D. (1995). Lack of p16INK4 or retinoblastoma protein (pRb), or amplification-associated overexpression of cdk4 is observed in distinct subsets of malignant glial tumors and cell lines. Cancer Res. 55, 4833–4836.
Ino, Y., Betensky, R. A., Zlatescu, M. C., Sasaki, H., Macdonald, D. R., Stemmer-Rachamimov, A. O., et al. (2001). Molecular subtypes of anaplastic oligodendroglioma: implications for patient management at diagnosis. Clin. Cancer Res. 7, 839–845.
Jaeckle, K. A., Ballman, K. V., van den Bent, M., Giannini, C., Galanis, E., Brown, P. D., et al. (2021). CODEL: phase III study of RT, RT+ TMZ, or TMZ for newly diagnosed 1p/19q codeleted oligodendroglioma. Analysis from the initial study design. Neuro Oncol. 23, 457–467. doi: 10.1093/neuonc/noaa168
Jiang, B., Zhao, W., Shi, M., Zhang, J., Chen, A., Ma, H., et al. (2018). IDH1 Arg-132 mutant promotes tumor formation through down-regulating p53. J. Biol. Chem. 293, 9747–9758. doi: 10.1074/jbc.RA117.001385
Johnson, B. E., Mazor, T., Hong, C., Barnes, M., Aihara, K., McLean, C. Y., et al. (2014). Mutational analysis reveals the origin and therapy-driven evolution of recurrent glioma. Science 343, 189–193. doi: 10.1126/science.1239947
Kaloshi, G., Benouaich-Amiel, A., Diakite, F., Taillibert, S., Lejeune, J., Laigle-Donadey, F., et al. (2007). Temozolomide for low-grade gliomas: predictive impact of 1p/19q loss on response and outcome. Neurology 68, 1831–1836. doi: 10.1212/01.wnl.0000262034.26310.a2
Kaminska, B., Czapski, B., Guzik, R., Król, S. K., and Gielniewski, B. (2019). Consequences of IDH1/2 mutations in gliomas and an assessment of inhibitors targeting mutated IDH proteins. Molecules 24:968. doi: 10.3390/molecules24050968
Killela, P. J., Reitman, Z. J., Jiao, Y., Bettegowda, C., Agrawal, N., and Diaz, L. A. Jr. et al. (2013). TERT promoter mutations occur frequently in gliomas and a subset of tumors derived from cells with low rates of self-renewal. Proc. Natl. Acad. Sci. U. S. A. 110, 6021–6026. doi: 10.1073/pnas.1303607110
Koelsche, C., Sahm, F., Capper, D., Reuss, D., Sturm, D., Jones, D. T., et al. (2013). Distribution of TERT promoter mutations in pediatric and adult tumors of the nervous system. Acta Neuropathol. 126, 907–915. doi: 10.1007/s00401-013-1195-5
Kohanbash, G., Carrera, D. A., Shrivastav, S., Ahn, B. J., Jahan, N., Mazor, T., et al. (2017). Isocitrate dehydrogenase mutations suppress STAT1 and CD8+ T cell accumulation in gliomas. J. Clin. Investigat. 127, 1425–1437. doi: 10.1172/JCI90644
Koivunen, P., Lee, S., Duncan, C. G., Lopez, G., Lu, G., Ramkissoon, S., et al. (2012). Transformation by the (R)-enantiomer of 2-hydroxyglutarate linked to EGLN activation. Nature 483, 484–488. doi: 10.1038/nature10898
Labreche, K., Simeonova, I., Kamoun, A., Gleize, V., Chubb, D., Letouzé, E., et al. (2015). TCF12 is mutated in anaplastic oligodendroglioma. Nat. Commun. 6, 1–9.
Latini, A., Scussiato, K., Rosa, R. B., Llesuy, S., Belló-Klein, A., Dutra-Filho, C. S., et al. (2003). D-2-hydroxyglutaric acid induces oxidative stress in cerebral cortex of young rats. Eur. J. Neurosci. 17, 2017–2022. doi: 10.1046/j.1460-9568.2003.02639.x
Lee, C.-C., Chen, W.-S., Chen, C.-C., Chen, L.-L., Lin, Y.-S., Fan, C.-S., et al. (2012). TCF12 protein functions as transcriptional repressor of E-cadherin, and its overexpression is correlated with metastasis of colorectal cancer. J. Biol. Chem. 287, 2798–2809. doi: 10.1074/jbc.M111.258947
Louis, D., Perry, A., Reifenberger, G., von, D. A., Figarella-Branger, D., Cavenee, W. K., et al. (2016). The 2016 World Health Organization classification of tumors of the central nervous system: A summary. Acta Neuropathol. 131, 803–820. doi: 10.1007/s00401-016-1545-1
Lu, C., Ward, P., Kapoor, G., Rohle, D., Turcan, S., Abdel-Wahab, O., et al. (2012). O, Rourke, DM; Berger, SL; Chan, TA; Levine, R. L.; Mellinghoff, IK; Thompson, CB IDH mutation impairs histone demethylation and results in a block to cell differentiation. Nature 483, 474–478. doi: 10.1038/nature10860
Luyken, C., Blümcke, I., Fimmers, R., Urbach, H., Elger, C. E., Wiestler, O. D., et al. (2003). The Spectrum of Long-term Epilepsy–associated Tumors: Long-term Seizure and Tumor Outcome and Neurosurgical Aspects. Epilepsia 44, 822–830. doi: 10.1046/j.1528-1157.2003.56102.x
Macdonald, D., Cairncross, G., Stewart, D., Forsyth, P., Sawka, C., Wainman, N., et al. (1996). Phase II study of topotecan in patients with recurrent malignant glioma. Ann. Oncol. 7, 205–207.
Maruno, M., Yoshimine, T., Muhammad, A. G., Tokiyoshi, K., and Hayakawa, T. (1996). Loss of heterozygosity of microsatellite loci on chromosome 9p in astrocytic tumors and its prognostic implications. J. Neurooncol. 30, 19–24. doi: 10.1007/BF00177439
Mason, W. P., Krol, G. S., and DeAngelis, L. M. (1996). Low-grade oligodendroglioma responds to chemotherapy. Neurology 46, 203–207. doi: 10.1212/wnl.46.1.203
Michaud, K., de Tayrac, M., D’Astous, M., Paquet, C., Gould, P. V., and Saikali, S. (2018). Impact of 9p deletion and p16, Cyclin D1, and Myc hyperexpression on the outcome of anaplastic oligodendrogliomas. PLoS One 13:e0193213. doi: 10.1371/journal.pone.0193213
Ohgaki, H., and Kleihues, P. (2009). Genetic alterations and signaling pathways in the evolution of gliomas. Cancer Sci. 100, 2235–2241. doi: 10.1111/j.1349-7006.2009.01308.x
Ostrom, Q. T., Gittleman, H., Liao, P., Vecchione-Koval, T., Wolinsky, Y., Kruchko, C., et al. (2017). CBTRUS statistical report: primary brain and other central nervous system tumors diagnosed in the United States in 2010–2014. Neuro Oncol. 19, (Suppl._5) v1–v88. doi: 10.1093/neuonc/nox158
Pan-Weisz, T. M. (2019). Health-Related Quality of Life and Executive Functioning among Primary Brain Tumor Patients: Associations and Causal Pathways: University of California, San Diego; 2019. California: University of California.
Pekmezci, M., Rice, T., Molinaro, A. M., Walsh, K. M., Decker, P. A., Hansen, H., et al. (2017). Adult infiltrating gliomas with WHO 2016 integrated diagnosis: additional prognostic roles of ATRX and TERT. Acta Neuropathol. 133, 1001–1016. doi: 10.1007/s00401-017-1690-1
Perry, A., Anderl, K., Borell, T. J., Kimmel, D. W., Wang, C. H., O’Fallon, J. R., et al. (1999). Detection of p 16, RB, CDK4, and p53 gene deletion and amplification by fluorescence in situ hybridization in 96 gliomas. Am. J. Clin. Pathol. 112, 801–809. doi: 10.1093/ajcp/112.6.801
Poisson, M., Pereon, Y., Chiras, J., and Delattre, J. (1991). Treatment of recurrent malignant supratentorial gliomas with carboplatin (CBDCA). J. Neurooncol. 10, 139–144. doi: 10.1007/bf00146875
Rasheed, A., Herndon, J. E., Stenzel, T. T., Raetz, J. G., Kendelhardt, J., Friedman, H. S., et al. (2002). Molecular markers of prognosis in astrocytic tumors. Cancer 94, 2688–2697. doi: 10.1002/cncr.10544
Rinaldi, M., Caffo, M., Minutoli, L., Marini, H., Abbritti, R. V., Squadrito, F., et al. (2016). ROS and brain gliomas: an overview of potential and innovative therapeutic strategies. Int. J. Mol. Sci. 17:984. doi: 10.3390/ijms17060984
Ruas, M., and Peters, G. (1998). The p16INK4a/CDKN2A tumor suppressor and its relatives. Biochim. Biophys. Acta 1378, F115–F177.
Ryskalin, L., Lazzeri, G., Flaibani, M., Biagioni, F., Gambardella, S., Frati, A., et al. (2017). mTOR-dependent cell proliferation in the brain. BioMed Res. Int. 2017:7082696.
Sahm, F., Koelsche, C., Meyer, J., Pusch, S., Lindenberg, K., Mueller, W., et al. (2012). CIC and FUBP1 mutations in oligodendrogliomas, oligoastrocytomas and astrocytomas. Acta Neuropathol. 123, 853–860. doi: 10.1007/s00401-012-0993-5
Sasaki, H., Zlatescu, M. C., Betensky, R. A., Ino, Y., Cairncross, J. G., and Louis, D. N. (2001). PTEN is a target of chromosome 10q loss in anaplastic oligodendrogliomas and PTEN alterations are associated with poor prognosis. Am. J. Pathol. 159, 359–367. doi: 10.1016/S0002-9440(10)61702-6
Shaw, E. G., Berkey, B., Coons, S. W., Bullard, D., Brachman, D., Buckner, J. C., et al. (2008). Recurrence following neurosurgeon-determined gross-total resection of adult supratentorial low-grade glioma: results of a prospective clinical trial. J. Neurosurg. 109, 835–841. doi: 10.3171/JNS/2008/109/11/0835
Sherr, C. J., and Roberts, J. M. (1999). CDK inhibitors: positive and negative regulators of G1-phase progression. Genes Dev. 13, 1501–1512. doi: 10.1101/gad.13.12.1501
Shin, D.-W., Lee, S., Song, S. W., Cho, Y. H., Hong, S. H., Kim, J. H., et al. (2020). Survival outcome and prognostic factors in anaplastic oligodendroglioma: a single-institution study of 95 cases. Sci. Rep. 10:20162.
Smith, J. S., Perry, A., Borell, T. J., Lee, H. K., O’Fallon, J., Hosek, S. M., et al. (2000a). Alterations of chromosome arms 1p and 19q as predictors of survival in oligodendrogliomas, astrocytomas, and mixed oligoastrocytomas. J. Clin. Oncol. 18:636. doi: 10.1200/jco.2000.18.3.636
Smith, J. S., Wang, X.-Y., Qian, J., Hosek, S. M., Scheithauer, B. W., Jenkins, R. B., et al. (2000b). Amplification of the platelet-derived growth factor receptor-A (PDGFRA) gene occurs in oligodendrogliomas with grade IV anaplastic features. J. Neuropathol. Exp. Neurol. 59, 495–503. doi: 10.1093/jnen/59.6.495
Smits, A., and Duffau, H. (2011). Seizures and the natural history of World Health Organization Grade II gliomas: a review. Neurosurgery 68, 1326–1333. doi: 10.1227/neu.0b013e31820c3419
Soffietti, R., Rudà, R., Bradac, G. B., and Schiffer, D. (1998). PCV chemotherapy for recurrent oligodendrogliomas and oligoastrocytomas. Neurosurgery 43, 1066–1073. doi: 10.1097/00006123-199811000-00035
Taliansky-Aronov, A., Bokstein, F., Lavon, I., and Siegal, T. (2006). Temozolomide treatment for newly diagnosed anaplastic oligodendrogliomas: a clinical efficacy trial. J. Neurooncol. 79, 153–157. doi: 10.1007/s11060-005-9020-1
Turcan, S., Rohle, D., Goenka, A., Walsh, L. A., Fang, F., Yilmaz, E., et al. (2012). IDH1 mutation is sufficient to establish the glioma hypermethylator phenotype. Nature 483, 479–483. doi: 10.1038/nature10866
Ueki, K., Ono, Y., Henson, J. W., Efird, J. T., von Deimling, A., and Louis, D. N. (1996). CDKN2/p16 or RB alterations occur in the majority of glioblastomas and are inversely correlated. Cancer Res. 56, 150–153.
Uhm, J. (2010). Cancer-associated IDH1 mutations produce 2-hydroxyglutarate Dang L, White DW, Gross S, et al. (Agios Pharmaceuticals, Cambridge, MA) Nature 462: 739-744, 2009. Year Book Neurol. Neurosurg. 2010, 111–112. doi: 10.1038/nature08617
Uittenbogaard, M., and Chiaramello, A. (2002). Expression of the bHLH transcription factor Tcf12 (ME1) gene is linked to the expansion of precursor cell populations during neurogenesis. Gene Expression Patterns 1, 115–121. doi: 10.1016/s1567-133x(01)00022-9
van den Bent, M. J., Baumert, B., Erridge, S. C., Vogelbaum, M. A., Nowak, A. K., Sanson, M., et al. (2017). Interim results from the CATNON trial (EORTC study 26053-22054) of treatment with concurrent and adjuvant temozolomide for 1p/19q non-co-deleted anaplastic glioma: a phase 3, randomised, open-label intergroup study. Lancet 390, 1645–1653. doi: 10.1016/S0140-6736(17)31442-3
van den Bent, M. J., Brandes, A. A., Taphoorn, M. J., Kros, J. M., Kouwenhoven, M. C., Delattre, J.-Y., et al. (2013a). Adjuvant procarbazine, lomustine, and vincristine chemotherapy in newly diagnosed anaplastic oligodendroglioma: long-term follow-up of EORTC brain tumor group study 26951. J. Clin. Oncol. 31, 344–350. doi: 10.1200/JCO.2012.43.2229
van den Bent, M. J., Dubbink, H. J., Marie, Y., Brandes, A. A., Taphoorn, M. J., Wesseling, P., et al. (2010). IDH1 and IDH2 mutations are prognostic but not predictive for outcome in anaplastic oligodendroglial tumors: a report of the European Organization for Research and Treatment of Cancer Brain Tumor Group. Clin. Cancer Res. 16, 1597–1604.
van den Bent, M. J., Erdem-Eraslan, L., Idbaih, A., de Rooi, J., Eilers, P. H., Spliet, W. G., et al. (2013b). MGMT-STP27 methylation status as predictive marker for response to PCV in anaplastic oligodendrogliomas and oligoastrocytomas. A report from EORTC study 26951. Clin. Cancer Res. 19, 5513–5522. doi: 10.1158/1078-0432.CCR-13-1157
van den Bent, M., Kros, J., Heimans, J., Pronk, L., Van Groeningen, C., Krouwer, H., et al. (1998). Response rate and prognostic factors of recurrent oligodendroglioma treated with procarbazine, CCNU, and vincristine chemotherapy. Neurology 51, 1140–1145. doi: 10.1212/wnl.51.4.1140
van den Bent, M., Taphoorn, M., Brandes, A., Menten, J., Stupp, R., Frenay, M., et al. (2003). Phase II study of first-line chemotherapy with temozolomide in recurrent oligodendroglial tumors: the European Organization for Research and Treatment of Cancer Brain Tumor Group Study 26971. J. Clin. Oncol. 21, 2525–2528. doi: 10.1200/JCO.2003.12.015
Vogelbaum, M. A., Berkey, B., Peereboom, D., Macdonald, D., Giannini, C., Suh, J. H., et al. (2009). Phase II trial of preirradiation and concurrent temozolomide in patients with newly diagnosed anaplastic oligodendrogliomas and mixed anaplastic oligoastrocytomas: RTOG BR0131. Neuro Oncol. 11, 167–175. doi: 10.1215/15228517-2008-073
Wang, P., Wu, J., Ma, S., Zhang, L., Yao, J., Hoadley, K. A., et al. (2015). Oncometabolite D-2-hydroxyglutarate inhibits ALKBH DNA repair enzymes and sensitizes IDH mutant cells to alkylating agents. Cell Rep. 13, 2353–2361. doi: 10.1016/j.celrep.2015.11.029
Warnick, R. E., Prados, M. D., Mack, E. E., Chandler, K. L., Doz, F., Rabbitt, J. E., et al. (1994). A phase II study of intravenous carboplatin for the treatment of recurrent gliomas. J. Neurooncol. 19, 69–74. doi: 10.1007/bf01051050
Watanabe, T., Yokoo, H., Yokoo, M., Yonekawa, Y., Kleihues, P., and Ohgaki, H. (2001). Concurrent inactivation of RB1 and TP53 pathways in anaplastic oligodendrogliomas. J. Neuropathol. Exp. Neurol. 60, 1181–1189. doi: 10.1093/jnen/60.12.1181
Wick, W., Hartmann, C., Engel, C., Stoffels, M., Felsberg, J., Stockhammer, F., et al. (2009). NOA-04 randomized phase III trial of sequential radiochemotherapy of anaplastic glioma with procarbazine, lomustine, and vincristine or temozolomide. J. Clin. Oncol. 27:5874. doi: 10.1200/jco.2009.23.6497
Wick, W., Roth, P., Hartmann, C., Hau, P., Nakamura, M., Stockhammer, F., et al. (2016). Neurooncology Working Group (NOA) of the German Cancer Society. Long-term analysis of the NOA-04 randomized phase III trial of sequential radiochemotherapy of anaplastic glioma with PCV or temozolomide. Neuro. Oncol. 18, 1529–1537. doi: 10.1093/neuonc/now133
Xu, W., Yang, H., Liu, Y., Yang, Y., Wang, P., Kim, S.-H., et al. (2011). Oncometabolite 2-hydroxyglutarate is a competitive inhibitor of α-ketoglutarate-dependent dioxygenases. Cancer Cell 19, 17–30. doi: 10.1016/j.ccr.2010.12.014
Yip, S., Butterfield, Y. S., Morozova, O., Chittaranjan, S., Blough, M. D., An, J., et al. (2012). Concurrent CIC mutations, IDH mutations, and 1p/19q loss distinguish oligodendrogliomas from other cancers. J. Pathol. 226, 7–16. doi: 10.1002/path.2995
You, G., Sha, Z.-Y., Yan, W., Zhang, W., Wang, Y.-Z., Li, S.-W., et al. (2011). Seizure characteristics and outcomes in 508 Chinese adult patients undergoing primary resection of low-grade gliomas: a clinicopathological study. Neuro Oncol. 14, 230–241. doi: 10.1093/neuonc/nor205
Yu, G., Butler, M. K., Abdelmaksoud, A., Pang, Y., Su, Y.-T., Rae, Z., et al. (2021). Case Report: Single-Cell Transcriptomic Analysis of an Anaplastic Oligodendroglioma Post Immunotherapy. Front. Oncol. 10:2935. doi: 10.3389/fonc.2020.601452
Yung, W., Mechtler, L., and Gleason, M. J. (1991). Intravenous carboplatin for recurrent malignant glioma: a phase II study. J. Clin. Oncol. 9, 860–864. doi: 10.1200/jco.1991.9.5.860
Zaatreh, M. M., Firlik, K. S., Spencer, D. D., and Spencer, S. S. (2003). Temporal lobe tumoral epilepsy: characteristics and predictors of surgical outcome. Neurology 61, 636–641. doi: 10.1212/01.wnl.0000079374.78589.1b
Zhao, S., Lin, Y., Xu, W., Jiang, W., Zha, Z., Wang, P., et al. (2009). Glioma-derived mutations in IDH1 dominantly inhibit IDH1 catalytic activity and induce HIF-1α. Science 324, 261–265. doi: 10.1126/science.1170944
Keywords: anaplastic oligodendroglioma, PCV, IDH mutation, procarbazine, lomustine, vincristine, temozolomide
Citation: Bou Zerdan M and Assi HI (2021) Oligodendroglioma: A Review of Management and Pathways. Front. Mol. Neurosci. 14:722396. doi: 10.3389/fnmol.2021.722396
Received: 08 June 2021; Accepted: 15 September 2021;
Published: 05 October 2021.
Edited by:
Verena Tretter, Medical University of Vienna, AustriaReviewed by:
Ryuta Saito, Tohoku University School of Medicine, JapanCopyright © 2021 Bou Zerdan and Assi. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Hazem I. Assi, aGExNTdAYXViLmVkdS5sYg==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.