 Ella Elizabeth Doornaert1
Ella Elizabeth Doornaert1 Alaa El-Cheikh Mohamad1Gurwinder Johal1
Alaa El-Cheikh Mohamad1Gurwinder Johal1 Brian Leonard Allman1Dorit Möhrle1,2†
Brian Leonard Allman1Dorit Möhrle1,2† Susanne Schmid1,3*†
Susanne Schmid1,3*†- 1Anatomy and Cell Biology, Schulich School of Medicine & Dentistry, University of Western Ontario, London, ON, Canada
- 2Faculty of Veterinary Medicine, University of Calgary, Calgary, AB, Canada
- 3Department of Psychology, University of Western Ontario, London, ON, Canada
The homozygous Cntnap2 knockout (KO) rat is a well-established genetic model for neurodevelopmental disorders, exhibiting core features of autism spectrum disorder (ASD), including impaired sensory processing and sensorimotor gating. Recent findings indicate that the severity of ASD-like phenotypes in Cntnap2 KO offspring is influenced by the parental genotype, with more pronounced impairments observed in KO rats bred from homozygous pairs compared to heterozygous pairs (Cntnap2 HET). However, it is unclear to what extent this is due to in utero versus postnatal effects. We, therefore, investigated how early postnatal environmental factors, shaped by differences in parental and littermate genotypes, influence auditory processing and sensorimotor gating in Cntnap2 KO rats. To examine this, we cross-fostered Cntnap2 KO pups bred from Cntnap2 KO rats to be reared with litters of Cntnap2 HET dams. Cross-fostering Cntnap2 KO rats reversed or partially reversed delayed hearing sensitivity maturation, heightened acoustic startle responses, and deficits in prepulse inhibition of the acoustic startle response. However, cross-fostering also exacerbated deficits in the neural responsiveness and conductivity in the auditory brainstem, as well as in gap-induced prepulse inhibition of the acoustic startle response. These results emphasize the importance of considering the postnatal environment and breeding strategies in preclinical genetic models of neuropsychiatric disorders. More importantly, they also demonstrate that ASD-like traits, including alterations in brainstem sensory processing, are not strictly determined by genetic factors, but remain malleable by environmental factors during early postnatal development.
Introduction
Autism spectrum disorder (ASD) is a neurodevelopmental condition caused by genetic, environmental, or most often, a combination of both factors (Lord et al., 2018). One genetic risk factor of ASD are variants in the contactin-associated protein-like 2 (CNTNAP2) gene, and a complete CNTNAP2 loss-of-function causes a syndromic disorder with core symptoms of autism (Strauss et al., 2006; Arking et al., 2008). CNTNAP2 encodes for the neurexin CASPR2, a transmembrane protein highly expressed in sensory pathways during the critical period of brain development (Gordon et al., 2016). The homozygous Cntnap2 knockout (KO) rat is a validated preclinical model of ASD that consistently exhibits ASD-like traits, including sensory processing alterations similar to the ones often observed in autistic individuals (Scott et al., 2018, 2020, 2022; Möhrle et al., 2021; El-Cheikh Mohamad et al., 2023; Doornaert et al., 2024). Auditory processing disruptions in ASD are at least partly due to delayed development of the auditory brainstem (Roth et al., 2012; Miron et al., 2016, 2018). Correspondingly, Cntnap2 KO rats showed delayed maturation of the auditory brainstem response (ABR), resulting in lower and slower ABR waves (Scott et al., 2018). Like many autistic individuals, Cntnap2 KO rats also exhibit heightened acoustic startle responses to sudden loud noises and reduced prepulse inhibition (PPI), the attenuation of the startle response when a less intense stimulus precedes the startle stimulus (Scott et al., 2018, 2020; Möhrle et al., 2021; El-Cheikh Mohamad et al., 2023; Doornaert et al., 2024).
“Critical periods” of brain development are time windows of increased plasticity in early postnatal development when neural circuits demonstrate a heightened sensitivity to environmental inputs (Inguaggiato et al., 2017). During critical periods, sensory deprivation can result in significant and often irreversible changes in neural circuitry (Kral et al., 2001; Chang and Merzenich, 2003; Bi et al., 2006; Bures et al., 2010; Bureš et al., 2017). Conversely, enriched sensory environments can enhance plasticity and promote optimal development of sensory pathways, highlighting the critical role of sensory experiences during early life (Engineer et al., 2004; Percaccio et al., 2007; Bureš et al., 2014; Svobodová Burianová and Syka, 2020). Indeed, evidence suggests that environmental factors during the critical period of auditory development can modulate phenotypic outcomes in Cntnap2 KO rats. Möhrle et al. (2023) examined the maternal isolation-induced ultrasound vocalizations of Cntnap2 KO pups bred from both heterozygous Cntnap2 knockout (Cntnap2 HET) and Cntnap2 KO pairings. Whereas the ultrasound vocalizations of Cntnap2 KO pups bred from heterozygous parents (KOhet) were only slightly different from wildtype controls, Cntnap2 KO pups from homozygous KO pairings (KOhom) exhibited a higher number of calls with different temporal structure characteristics, altered call pitch, and less consistency in transitioning between call types. Similarly, in vitro patch-clamp electrophysiology revealed differences in the intrinsic neuronal properties and synaptic activity in auditory cortex pyramidal neurons between Cntnap2 KOhet and KOhom rats (Scott et al., 2022; Mann et al., 2023). Lastly, delayed ABR maturation was observed in Cntnap2 KOhom rats but not in Cntnap2 KOhet pups (Scott et al., 2018; Zheng et al., 2023).
In summary, evidence suggests that brain alterations caused by the Cntnap2 mutation may depend on differences between heterozygous and homozygous breeding. These differences could emerge prenatally, due to the maternal in utero environment and/or maternal mitochondrial DNA, or postnatally, caused, e.g., by differences in maternal care or sibling social and communicative interactions. It also remains unclear to what extent the developmental consequences resulting from the Cntnap2 mutation are still malleable and sensitive to early environmental conditions. To examine this, the current study used a cross-fostering paradigm: Cntnap2 KOhom pups were cross-fostered to be reared by a heterozygous dam in her litter prior to the critical period of auditory development (KOCF). The ABR, startle reactivity, and PPI were compared between Cntnap2 KOCF animals and non-cross-fostered KOhom and KOhet animals. We hypothesized that cross-fostering benefits the development of neural circuits underlying auditory processing, potentially restoring hearing sensitivity maturation and reducing heightened acoustic startle response and PPI deficits.
Materials and methods
Experimental design
Animals
This study was conducted using male (M) and female (F) Sprague–Dawley wildtype rats (Cntnap2 WT) and Cntnap2 KO rats. A total of 14 litters were used in this study consisting of 9 litters from Cntnap2 HET breeders (Cntnap2 HET × Cntnap2 HET) and 5 litters from Cntnap2 KO breeders (Cntnap2 KO × Cntnap2 KO). Original Cntnap2 HET breeders (Cntnap2em1Sage) were obtained from Horizon Discovery (Boyertown, PA, United States). Cntnap2 KO breeders were obtained from in-house breeding of Cntnap2 HET pairs. Rats were housed in open cages and given ad libitum food and water. Holding rooms were temperature-controlled and kept on a 12-h light/dark cycle. All behavioral testing occurred during this cycle’s light phase (from 7:00 to 19:00 h). All procedures were approved by the University of Western Ontario Animal Care Committee and followed the guidelines provided by the Canadian Council on Animal Care.
Five breeding rounds were initially conducted with at least 2 weeks in between. One breeding round involved pairing two Cntnap2 HET males with two Cntnap2 HET females each, as well as one Cntnap2 KO male with two Cntnap2 KO females. Pairings used males aged 13 weeks to 11 months and females aged 11 weeks to 11 months. Unrelated males and females (non-siblings) were used for pairings. All females that were impregnated were only used once as first-time dams. All males that impregnated a female were only used once, except for one Cntnap2 KO male that impregnated two females from consecutive rounds.
Rats were paired for 48 h to ensure that pregnant dams would have a similar date of birth within a breeding round. Females paired together for breeding were separated into single cages three to 4 days before their expected date of giving birth. After birth, toe clips were taken for offspring between postnatal (PND) 5 and 6 to genotype Cntnap2. Between PND 10 and 11, litters from Cntnap2 HET pairings were culled by euthanizing all the Cntnap2 HET pups so that only Cntnap2 WT and Cntnap2 KO (KOhet) offspring remained (Figure 1A; Cntnap2 WT animals are represented in blue, Cntnap2 KOhet animals are represented in red). At that time, 2 to 5 Cntnap2 KO pups from Cntnap2 KO pairings were transferred to these culled litters of Cntnap2 HET pairings (group Cntnap2 KOCF; represented in green). The remainder of the Cntnap2 KO offspring from the Cntnap2 KO pairings remained with the Cntnap2 KO dam for rearing (group Cntnap2 KOhom; represented in purple). The cross-fostering time point of PND 10 to 11 was chosen balancing the time it took to receive the genotyping results with the start of the critical period of auditory development upon opening of the meatus at around PND 12 (Figure 1B). It is expected that the pups did not receive functional auditory input until this point (Bi et al., 2006).
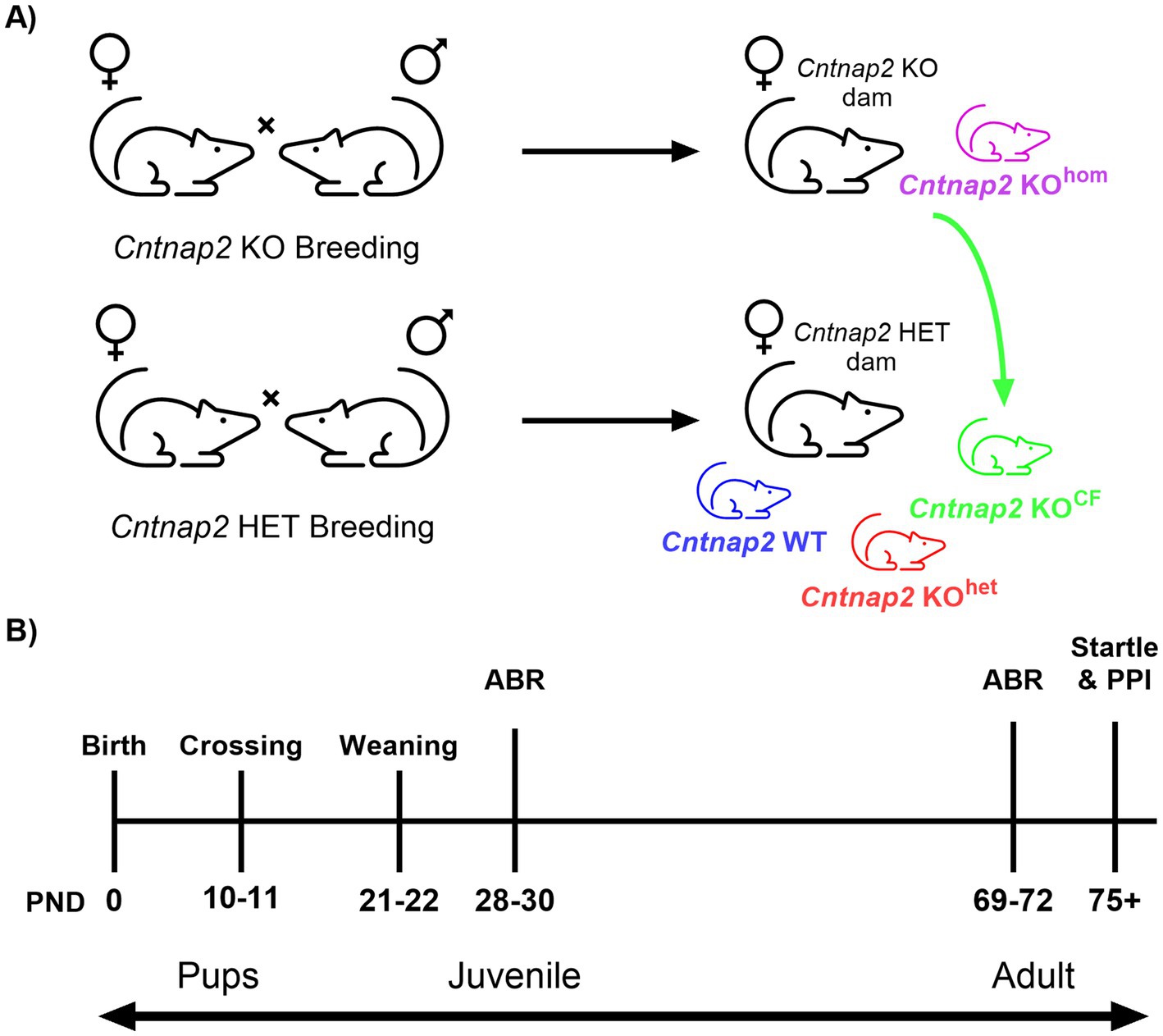
Figure 1. Cross-fostering illustration and summary of experimental timeline. (A) A breeding round consisted of breeding Cntnap2 KO (Cntnap2- KO × Cntnap2 KO) and Cntnap2 HET (Cntnap2 HET × Cntnap2 HET) rats. Experimental groups included: Cntnap2 WT animals bred by Cntnap2 HET rats and reared by a Cntnap2 HET dam (Cntnap2 WT; blue), Cntnap2 KO animals bred by Cntnap2 HET rats and reared by a Cntnap2 HET dam (Cntnap2 KOhet; red), Cntnap2 KO animals bred by Cntnap2 KO rats and reared by a Cntnap2 KO dam (Cntnap2 KOhom; purple), and Cntnap2 KO animals bred by Cntnap2 KO rats and cross-fostered to be reared by a Cntnap2 HET dam (Cntnap2 KOCF; green). (B) Experimental timeline outlining the postnatal days (PND) for cross-fostering and behavioral assessments. The auditory brainstem response (ABR) was assessed during the juvenile period and adulthood, while the acoustic startle response and prepulse inhibition (PPI) were evaluated in adulthood.
To cross-foster the pups from the litter of a Cntnap2 KO to a Cntnap2 HET dam, both dams were removed from their home cages and a heating pad was placed under the cages to keep the pups warm. Before crossing, Cntnap2 KOCF pups were covered and rubbed in the nesting material of the Cntnap2 HET dam for 3 min to make them smell like the pups of the Cntnap2 HET dam. This process of rubbing the animals with bedding from their final cage was performed on all the animals to maintain consistency. Gloves were changed between the handling of animals of different litters to reduce the scents of foreign litters from carrying over. Then, dams were returned to their home cages and left undisturbed for 48 h (cages not changed). Across all breeding rounds, all cross-fostered pups were accepted by their foster dam. Animals were weaned between PND 21 and 22 and housed in cages of two to three sex-matched siblings from their final litter. When possible, Cntnap2 KOCF animals were housed with one or two each of Cntnap2 WT and KOhet animals.
After five breeding rounds, an additional two rounds were conducted with only Cntnap2 HET pairings to increase the sample size for the Cntnap2 WT and KOhet animal groups. For these rounds, the procedure of cross-fostering animals into these litters was mimicked by introducing Cntnap2 HET animals from other Cntnap2 HET pairings.
Auditory brainstem response
ABR recordings were conducted during the juvenile stage and adulthood. 90 juvenile rats were tested at PND 28–30: 26 Cntnap2 WT rats (13 male/13 female), 21 Cntnap2 KOhet (11 male/10 female), 23 Cntnap2 KOhom (12 male/11 female), and 20 Cntnap2 KOCF (10 male/10 female). A total of 81 adult rats were tested at PND 69–72: 21 Cntnap2 WT rats (11 male/10 female), 18 Cntnap2 KOhet (10 male/ 8 female), 22 Cntnap2 KOhom (12 male/10 female), and 20 Cntnap2 KOCF (10 male/10 female). Rats were anesthetized with ketamine (juvenile: 40 mg/kg or adult: 80 mg/kg) and xylazine (juvenile: 2.5 mg/kg or adult: 5 mg/kg). For juveniles, dosing intervals were all IM. and administered as follows: initial full dose +1/4 dose, 1/4 dose after 5 min, 1/3 dose after 20–30 min, and 1/4 dose every 30 min after as necessary. For adults, the initial full dose +1/4 dose was administered IP. and supplemental doses were administered IM as follows: 1/3 dose after 15 min and 1/4 dose every 30 min as necessary. For both juveniles and adults, supplemental doses were switched to ketamine only after administering the supplemental doses totaling the initial full dose. Injections of 0.9% saline were provided every hour (10 mL/kg) and eye lubrication was applied before testing. Throughout the recordings, body temperature was maintained at approximately 37°C using a homeothermic heating pad (ATC-2000; World Precision Instruments).
Subdermal electrodes (27 gauge; Rochester) were positioned over the right mastoid process (active electrode), the vertex (reference electrode), and the midback (ground electrode). The acoustic stimuli used in the ABR assessment consisted of a click and 4 tones (4 kHz, 11 kHz, 20 kHz, and 32 kHz; 10 ms duration; 1 ms rise/fall time), which were generated using a Tucker-Davis Technologies RZ6 processing module and sampled at 100 kHz. A magnetic speaker (MF1; Tucker-Davis Technologies) positioned 10 cm from the animal’s right ear was used to deliver the stimuli and its left ear was blocked with a custom foam plug. Before the ABR assessment, the acoustic stimuli were calibrated with custom MATLAB software (The Math-Works) using a 1/4-inch microphone (2,530; Larson Davis) and preamplifier (2,221; Larson Davis). The acoustic stimuli were each presented 750 times (21 times/s) at decreasing intensities from 90 to 0 dB sound pressure level (SPL) in 5 dB SPL steps. The sound-evoked activity associated with the ABR assessment was preamplified and digitized using a Medusa4Z BioAmp (Tucker-Davis Technologies) set to a 12 kHz sampling frequency. A fiber-optic cable sent the digitized signal to the RZ6 processing module. The signal was filtered (300–3,000 Hz) and averaged using BioSig software (Tucker-Davis Technologies). The positive and negative peak amplitudes of each of the characteristic waves of the rat ABR were measured in microvolts in reference to the baseline (0 V) and the latency of each of these peaks was determined from the stimulus onset.
The whole trace data were exported to MATLAB R2022a to determine the hearing sensitivity (ABR threshold) for the 5 sound stimuli as well as extract the values for the amplitudes and latencies for waves I, II, and IV resulting from the click stimuli (Figure 2A). Consistent with previous studies, each rat’s ABR threshold for the click and tonal stimuli was determined using the criterion of just noticeable deflection of the averaged electrical activity within a 10 ms window following the click (Schormans et al., 2017; Scott et al., 2018). Wave amplitudes were calculated by subtracting the negative peak from the positive peak amplitude (peak-to-peak). Central compensation of neuronal responsiveness with age was calculated by the ratio of ABR wave IV to I amplitudes (Möhrle et al., 2016). ABR peak latencies and interpeak latencies were based on the negative peak amplitudes. To normalize the amplitude and latency measures across animals, these measures were compared at sound levels above the animals’ thresholds (20 dB to 60 dB above threshold). The experimenter was blinded to the animal’s genotype for all analyses associated with the ABR assessment.
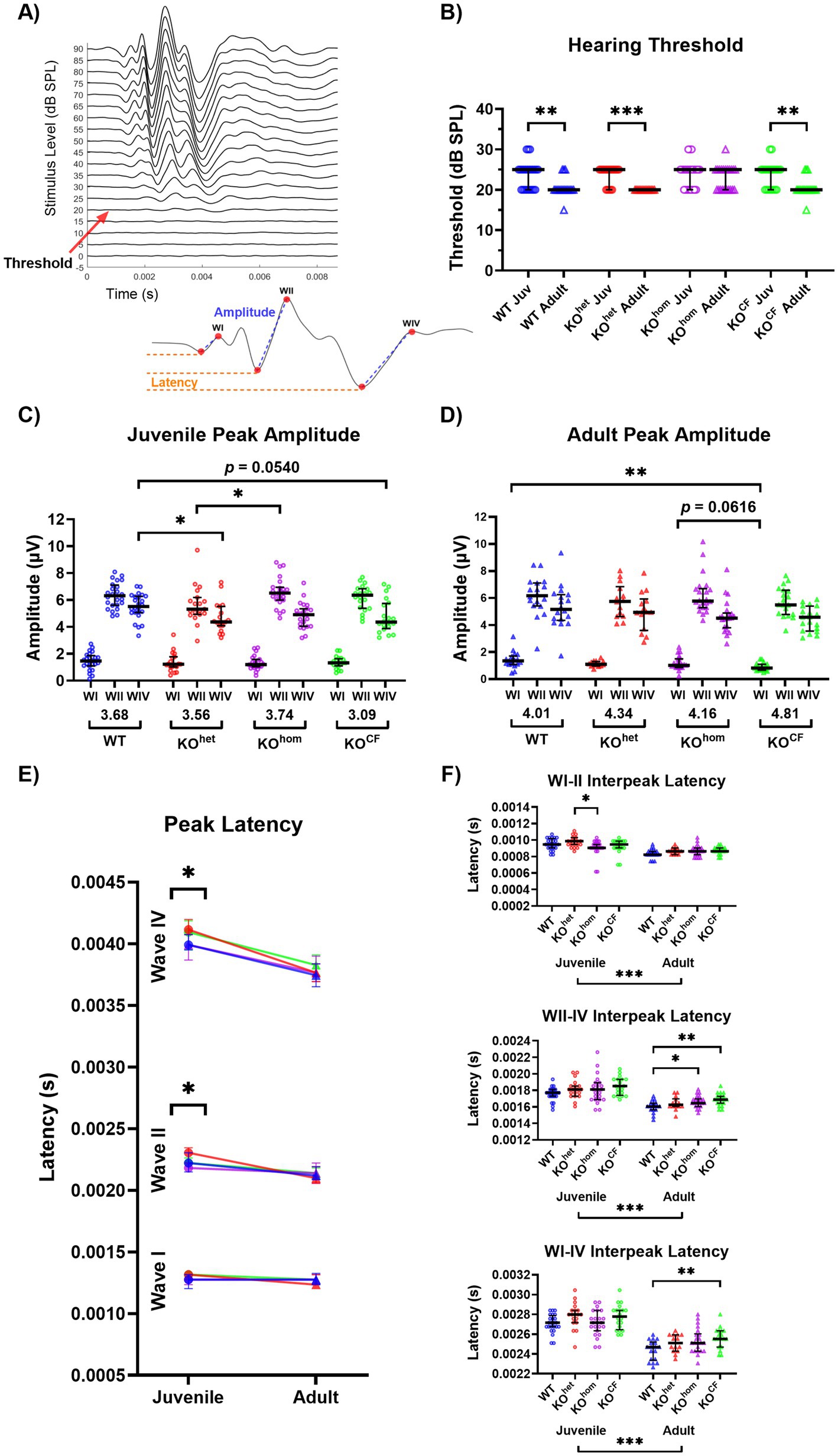
Figure 2. Cross-fostered Cntnap2 knockout rats have improved development of hearing sensitivity but reduced neural responsiveness and conduction within the auditory brainstem pathway. (A) An example of the acoustically evoked ABRs from a rat in response to click stimuli of increasing sound level with a red arrow to indicate the ABR threshold. Below is a magnification of an ABR trace from the same animal in response to a 70 dB SPL click stimulus. Waves I, II, and IV reflect the synchronized neural activity in the auditory nerve, cochlear nucleus, and inferior colliculus/lateral lemniscus, respectively. Measurements show how amplitude (from negative to positive peak; blue) and peak latency (negative peak; orange) were determined. (B) Change in ABR threshold to click stimuli with age. The hearing threshold of Cntnap2 KOhom rats does not improve between juvenile age and adulthood as observed in WT and KOhet rats. Cntnap2 KOCF rats exhibit typical maturation of hearing threshold with age, suggesting they have restored development of hearing sensitivity. (C) Juvenile ABR peak amplitudes. Group WIV/I ratios are indicated under the x-axis. Juvenile Cntnap2 KOhom and KOhet rats exhibit reduced peak amplitudes for WII and WIV, respectively, compared to WT rats. The WIV amplitude of Cntnap2 KOCF rats is trending toward being lower than WT rats. (D) Adult ABR peak amplitudes. Group WIV/I ratios are indicated under the x-axis. The reduced peak amplitudes observed in juvenile Cntnap2 KOhom and KOhet rats do not persist in adulthood. In contrast, Cntnap2 KOCF rats display reduced WI amplitude compared to WT rats. (E) ABR peak latencies across age. Only juvenile Cntnap2 KOhet rats show slower WII and WIV latencies. This is normalized by adulthood. (F) ABR interpeak latencies. In adulthood, Cntnap2 KOhom and KOCF exhibit slower WII-IV interpeak latencies. Adult Cntnap2 KOCF rats also show a slower WI-IV interpeak latency, indicative of overall slower conduction of the ABR. *p < 0.05, **p < 0.01, ***p < 0.0001.
Acoustic startle response, PPI, gap-PPI
The acoustic startle response, PPI, and gap-induced prepulse inhibition of startle (gap-PPI) were evaluated in adulthood (PND 75+). A total of 81 rats were tested, including 21 Cntnap2 WT rats (11 male/10 female), 19 Cntnap2 KOhet (10 male/9 female), 21 Cntnap2 KOhom (11 male/10 female), and 20 Cntnap2 KOCF (10 male/10 female), using the Med Associates (Vermont, United States) startle system with protocols modified from Doornaert et al. (2024), El-Cheikh Mohamad et al. (2023) and Miller et al. (2021). In brief, animals were placed in plexiglass tubes on weight-transducing platforms in sound-attenuating startle boxes (Med Associates). Animals were initially acclimated to the experimental procedure by undergoing three 5-min sessions in the startle box with only background noise (65 dB sound pressure level, SPL, white noise). Then, a session to determine the startle reactivity (I/O function) was conducted consisting of 12 startle stimuli ranging from 65 dB to 120 dB in 5 dB increments (20 ms white noise) plus the two prepulse intensities used in the PPI protocol, 75 dB and 85 dB (4 ms white noise). Stimuli were presented in pseudorandomized order. This was done before testing to adjust the gain of the platform transducer signal to ensure optimal spread over the dynamic range of the system for each animal.
Testing of the acoustic startle response and PPI involved two sessions a day for five consecutive days. Each testing session consisted of three distinct blocks: acclimation, habituation, and PPI. The first block was 5 min of acclimation to the startle box with only background noise (65 dB white noise). The habituation block presented animals with 12 trials of a 110 dB startle stimulus (20 ms white noise; 10–15 s variable intertrial intervals). The data were only collected from the third block, in which PPI was assessed by pairing a non-startling prepulse (75 dB or 85 dB; 4 ms white noise) and a startle stimulus (70 dB, 80 dB, 90 dB, 100 dB, 110 dB, or 120 dB; 20 ms white noise) with a 100 ms fixed interstimulus interval. Startle-alone stimuli without prepulses were also presented. Trials had variable intertrial intervals of 10–15 s and were presented in a pseudorandomized order. Each condition was repeated 4 times per testing session, resulting in 84 trials per testing session and 40 repetitions per trial type across all 10 testing sessions.
Following PPI testing and a day of break without testing, gap-PPI testing was conducted over 3 consecutive days. The first day consisted of a 5-min acclimation session to the new background noise used for the gap-PPI protocol (75 dB, white noise). Then, a session to assess gap-PPI was done on days 2 and 3. These sessions paired gap periods (2 ms, 5 ms, 10 ms, 20 ms, 40 ms, 50 ms, 75 ms, or 100 ms) and a 115 dB startle stimulus (20 ms white noise; 100 ms fixed interstimulus interval). A startle-alone stimulus was also presented. Again, trials had variable intertrial intervals of 10–15 s and were presented in a pseudorandomized order. Each condition was repeated 10 times per testing session, resulting in 90 trials per testing session and 20 sessions total per trial type combined over the two sessions.
The startle magnitude was defined as the maximum peak-to-peak value of the response waveform. Before statistical analyses, each rat’s startle response values were corrected for the gain setting. The subsequent analysis followed the methods outlined by Martin-Iverson and Stevenson (2005) and Miller et al. (2021). Startle reactivity across the range of stimuli was assessed by fitting each animal’s responses to a sigmoidal regression function using GraphPad Prism 9.3.1 (San Diego, California, United States; Non-linear regression; Method: Sigmoidal, 4PL, X is concentration; Method: Least squares regression; Initial values: choose automatically; Confidence: Unstable parameters and ambiguous fits as Neither option; Diagnostics: default values including Adjusted R Squared, RMSE, and tests of normality, see also Möhrle et al., 2021) with the following equation:
where Y represents the startle response magnitude, Top is the maximum startle response magnitude and Bottom is the minimum response magnitude. X denotes the startle stimulus intensity (dB SPL) necessary to elicit a certain Y value (in arbitrary units). ES50 is the sound intensity (dB SPL) required to maintain the half-maximum response. Hillslope is the slope of the curve. Parameters of interest were derived from the equation to evaluate and compare differences in baseline startle and PPI. This includes the maximum startle response (Top), startle threshold (10% of maximum threshold), ES50, and saturation point (90% of maximum startle). In this sigmoidal regression analysis, GraphPad Prism provided the standard error of regression using Sy.x, which serves as an estimate of the goodness-of-fit for models involving two or more parameters.
Startle scaling was assessed through changes to the maximum startle response, or Top (El-Cheikh Mohamad et al., 2023; Doornaert et al., 2024). Each animal’s responses were fitted to a sigmoidal regression function as previously described (including Constrain: Bottom is constant equal to 0). Sound scaling was determined by changes to the threshold, ES50, and saturation point (Martin-Iverson and Stevenson, 2005; Möhrle et al., 2021; El-Cheikh Mohamad et al., 2023; Doornaert et al., 2024). For this, startle responses for each animal and prepulse condition were scaled between 0 and 1. To scale the startle magnitudes at each startle stimulus intensity (X), we used the following equation: (startle magnitude at X—startle magnitude at 70 dB startle stimulus) / (startle magnitude at 120 dB startle stimulus—startle magnitude at 70 dB startle stimulus). The scaled values were then fitted to the sigmoidal regression function using the same procedure as above (except Constrain: Bottom is constant equal to 0 and Top is constant equal to 1). ES50 was provided by the regression, as well as Hillslope, which can be used as a metric of reflex efficiency (Martin-Iverson and Stevenson, 2005). The threshold and saturation point were calculated in MATLAB R2022a by rearranging the above equation to solve for X (below). The threshold Y value was set to 10% of the Top, and the saturation point Y value was set to 90% of the Top.
For PPI analysis, the percent PPI was calculated using the startle magnitudes from the 10 PPI sessions:
Similarly, the percent gap-PPI was obtained using the startle magnitude from the 2 gap-PPI sessions:
Statistical analysis
The data are presented as group medians with errors indicating interquartile range (IQR). Outlier analysis was performed in IBM SPSS (version 26) for each testing component in this study: ABR, startle/PPI, and gap-PPI. Through boxplot assessment, extreme outliers were identified as those exceeding 3 IQR from the group median. Table 1 shows the number of animals per group for each behavioral test after outlier exclusion. Two additional Cntnap2 KOhet females were excluded from the adult ABR analyses because their trace data were too noisy and illegible to extract reliable measures.
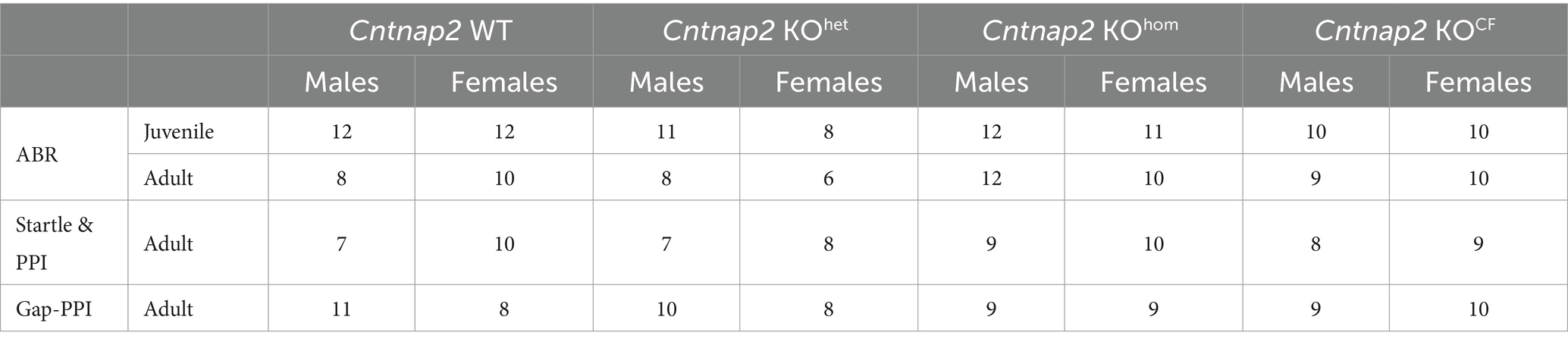
Table 1. The number of animals per group for each behavioral experiment after outlier exclusion.
Subsequent statistical analyses were performed in GraphPad Prism 9.3.1 and RStudio 2022.07.2, and figures were generated in GraphPad Prism 9.3.1. To determine the main effects and interactions, we employed ARTool (Aligned Rank Transform, ART) to align-and-rank data for nonparametric factorial ANOVA, and ART-C for post-hoc pairwise comparisons (Wobbrock et al., 2011; Elkin et al., 2021). Statistical tests following the ART were based on the experimental design and included univariant analysis of variance [x-way ANOVA, repeated measures (RM) ANOVA, or Mixed-effects model, as appropriate], followed by multiple comparison tests with correction for type 1 error after Tukey’s method or Sidak’s multiple comparison test when appropriate. For measures in which there was no effect of sex or an interaction effect involving sex, the data were collapsed across sex to examine the effect of genotype and/or age. The chosen statistical significance level was α = 0.05. Resulting p values are reported in the figure captions using: no asterisk or ns for non-significance, *p < 0.05; **p < 0.01; ***p < 0.0001.
Results
Cross-fostering improves the maturation of hearing sensitivity in Cntnap2 KO rats but exacerbates ABR responsiveness and latency impairments
We compared ABR thresholds to click stimuli from juvenile age and adulthood to assess the development of hearing sensitivity (Figures 2A,B). We found a significant effect of age on the threshold [p < 0.0001, F(1, 63) = 43.41]. Considering there was no effect of sex, nor interaction effects involving sex, the data across sex were collapsed for further analysis. After collapsing across sex, the effect of age on the threshold persisted, in addition to the appearance of an interaction effect between genotype and age [age p < 0.0001, F(1, 67) = 18.99; genotype × age p = 0.0011, F(3, 67) = 2.866]. Post-hoc testing revealed disrupted maturation of hearing sensitivity in Cntnap2 KOhom rats: whereas Cntnap2 WT and KOhet rats exhibited a decrease in their ABR thresholds between the juvenile and adult stages, thresholds did not decrease in Cntnap2 KOhom [Cntnap2 WT p = 0.0067, Cntnap2 KOhet < 0.0001, Cntnap2 KOhom p = 0.6323; Figure 2B]. Interestingly, Cntnap2 KOCF rats displayed a decrease in threshold with age (p = 0.0011), indicating restored maturation of the brainstem upon cross-fostering. Please note that this finding was only observed for the threshold of click stimuli, whereas there were only subtle group differences for the threshold for tone stimuli (see Supplementary Figure 1).
Next, we examined the peak-to-peak amplitude and negative peak latency of WI, WII, and WIV of the ABR, representing sound processing in the auditory nerve, cochlear nucleus, and inferior colliculus/lateral lemniscus, respectively (Figures 2A,C–E). Moreover, we examined the interpeak latencies between WI-II, WII-IV, and WI-IV (Figure 2F). The analysis was first conducted across sound intensities 20 dB to 60 dB above the threshold and is shown in Supplementary Figures 2–4. This analysis revealed subtle group differences for all measures, and the sound level substantially interacted with all other factors in the analysis. Consequently, we chose to examine these measurements at the sound level 60 dB above the threshold, as this is where group differences appeared to emerge but were undetected by our first analysis. Again, the data were collapsed across sex for further analysis because there were no effects of sex nor interaction effects involving sex on these measures.
During the juvenile stage, there was a main effect of genotype on the amplitude of WII and WIV in response to a 60 dB click [WII p < 0.0001, F(1, 67) = 18.9; WIV p < 0.0001, F(1, 67) = 18.9; Figure 2C]. Post-hoc testing revealed that Cntnap2 KOhet animals had a lower WII amplitude than KOhom animals (p = 0.0323). Cntnap2 KOhet animals also had a lower WIV amplitude than WT animals, and KOCF animals showed a similar trend (Cntnap2 KOhet p = 0.0418, Cntnap2 KOCF p = 0.0540). In adulthood, an effect of genotype on amplitude was only found in WI [p = 0.0036, F(3, 69) = 4.959; Figure 2D], where Cntnap2 KOCF animals had lower amplitudes than WT animals and were trending toward having lower amplitude than Cntnap2 KOhom (Cntnap2 WT p = 0.0018, Cntnap2 KOhom p = 0.0616).
Based on these age-related findings, we calculated the change in amplitude with age for each wave by dividing each animal’s juvenile response by the adult response and taking its percentage. A significant effect of genotype was found only in WI [p = 0.038, F(3, 67) = 2.96; Supplementary Figure 6A], where Cntnap2 KOCF animals had a greater change in amplitude than WT animals (p = 0.0434).
We also calculated the WIV/I ratio, a measure reflecting the maturation of neural processing in the auditory brainstem (Scott et al., 2018; Möhrle et al., 2019). A higher ratio typically indicates a more mature auditory system, while a lower ratio suggests immaturity or delayed development. Across all the sound levels above threshold, only subtle group differences were found, except that Cntnap2 KOCF females showed greater WIV/I ratios in adulthood compared to juvenile age at the higher sound intensities (Supplementary Figure 5). At 60 dB above the threshold, there was no effect of genotype within the juvenile or adult stage for the WIV/I ratio [juvenile p = 0.6968, F(3, 82) = 0.4803; adult p = 0.1525, F(3, 69) = 1.815; group ratios are indicated under the x-axes in Figures 2C,D]. We calculated the percent change in WIV/I between juvenile age and adulthood and found no significant effect of genotype [p = 0.1001, F(3, 67) = 2.16; Supplementary Figure 6B].
We observed a significant effect of genotype on peak latencies in the juvenile age for WII and WIV [WII p = 0.0157, F(3, 82) = 3.662; WIV p = 0.0116, F(3, 82) = 3.907; Figure 2E]. Post-hoc testing revealed that Cntnap2 KOhet animals had a greater WII latency than KOhom animals and a greater WIV latency than WT animals (WII p = 0.0057, WIV p = 0.0393). By adulthood, this slower latency appeared to normalize, as no significant genotype effects were found for any of the waves [WI p = 0.3311, F(3, 69) = 1.161; WII p = 0.8178, F(3, 69) = 0.3103; WIV p = 0.1541, F(3, 69) = 1.806].
When examining interpeak latencies, there was a significant effect of genotype for WI-II in juvenile animals [p = 0.0458, F(3, 82) = 2.788], where Cntnap2 KOhom animals showed a lower WI-II interpeak latency than KOhet animals (p = 0.0306; Figure 2F). In adulthood, there was an effect of genotype on WII-IV and WI-IV interpeak latencies [WII-IV p = 0.0063, F(3, 69) = 4.466; WI-IV p = 0.0147, F(3, 69) = 3.755]. Interestingly, both adult Cntnap2 KOhom and KOCF animals had greater WII-IV interpeak latency than WT animals (Cntnap2 KOhom p = 0.0459, Cntnap2 KOCF p = 0.0042), and only KOCF animals had a greater WI-IV interpeak latency than WT animals (Cntnap2 KOhom p = 0.1178, Cntnap2 KOCF p = 0.0045). We calculated the percent change in interpeak latency with age and found a significant effect of genotype for WI-II [p = 0.044, F(3, 67) = 2.815; Supplementary Figure 6C]. Cntnap2 KOhom animals had a lower change in WI-II interpeak latency with age compared to WT animals (p = 0.0388).
In summary, we found cross-fostering to have beneficial effects on the impaired maturation of hearing sensitivity as it restored the age-related decrease of hearing threshold in Cntnap2 KOCF rats upon reaching adulthood. However, cross-fostering did not benefit the changes in neural responsiveness or conductivity measured by the ABR. Rather on the contrary, cross-fostered animals appeared to have greater ABR disruptions as evidenced by a reduction in WI amplitude and a slower WI-IV interpeak latency.
Cross-fostering reduces heightened startle response in Cntnap2 KO males
After establishing a restoration of the ABR threshold upon maturation in cross-fostered animals and otherwise rather subtle changes in the ABR, we proceeded to test the effect of cross-fostering on baseline startle magnitude using the extracted parameters of the baseline startle I/O curves: maximum startle response (Top; Figures 3A,B), threshold, ES50, saturation point, and slope (Figures 3C–G). There were significant effects of sex for the threshold, ES50, and saturation [threshold p < 0.0001, F(1, 60) = 23.65; ES50 p < 0.0001, F(1, 60) = 31.61; saturation p = 0.0176, F(1, 60) = 5.959], as well as interaction effects between sex and genotype for the Top, threshold, ES50, and saturation [Top p = 0.0290, F(3, 60) = 3.218; threshold p = 0.0001, F(3, 60) = 8.004; ES50 p < 0.0001, ‑F(3, 60) = 11.63; saturation p = 0.0115, F(3, 60) = 4.007]. Therefore, for all parameters, the data were analyzed separately for males and females to assess the effect of genotype within the sexes.
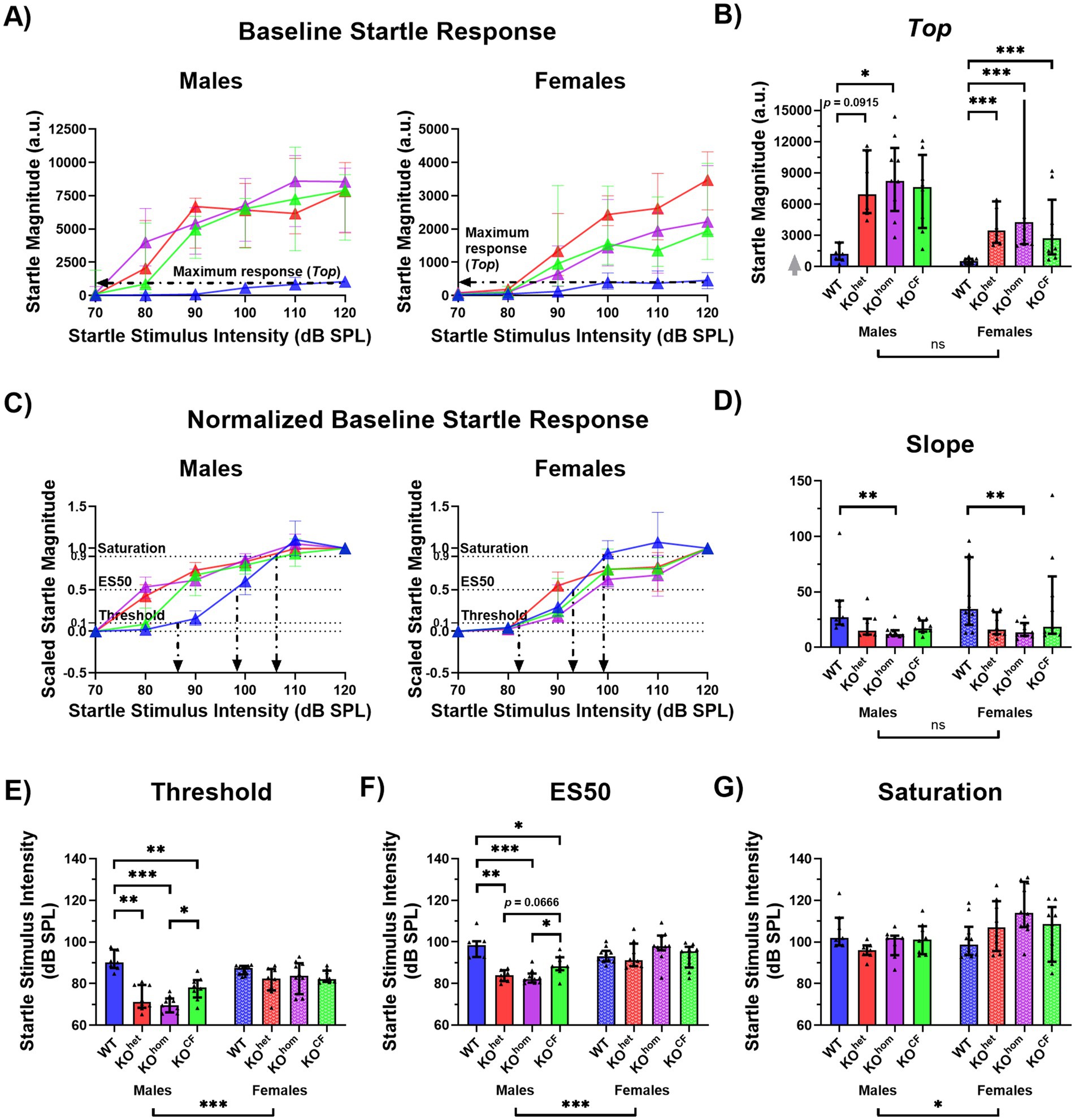
Figure 3. Cross-fostered Cntnap2 knockout males show less exaggerated startle responses. (A) Baseline startle response curves. Black arrows point to the maximum startle response value (Top) for WT animals. Goodness of fit Sy.x: male WT = 449.7, male KOhet = 2,485, male KOhom = 2,413, male KOCF = 2,736, female WT = 181.1, female KOhet = 841.6, female KOhom = 932.0, female KOCF = 2,223. (B) Maximum startle response (Top). The gray arrow indicates that there are values outside the limits of the y-axis, but the graph was zoomed in to visualize the data more clearly. Unlike male Cntnap2 KOhom rats, KOCF males do not have a statistically higher Top than WT males. This is not apparent in females where all Cntnap2 KO rats have a higher Top than WT rats. (C) Scaled startle response curves. Black arrows point to the threshold, ES50, and saturation point for WT animals. Goodness of fit Sy.x: male WT = 0.2048, male KOhet = 0.1263, male KOhom = 0.1311, male KOCF = 0.1673, female WT = 0.1956, female KOhet = 0.1961, female KOhom = 0.1823, female KOCF = 0.2003. (D) Slope. In males and females, Cntnap2 KOhom rats but not KOCF rats have a lower slope than WT rats. (E) Threshold. Male Cntnap2 KOCF rats have a higher threshold than KOhom rats, but still a lower threshold than WT rats. (F) ES50. Male Cntnap2 KOCF rats have a higher ES50 than KOhom rats, but still a lower ES50 than WT rats. (G) Saturation. There are no group differences in saturation point. *p < 0.05, **p < 0.01, ***p < 0.0001, ns indicates non-significance of the comparison.
In terms of maximum startle response, there was a significant effect of genotype for males and females [males p = 0.0337, F(3, 27) = 3.348; females p < 0.0001, F(3, 33) = 16.29]. Male Cntnap2 KOhom rats showed a higher Top value than WT rats (p = 0.0303), but Cntnap2 KOCF did not differ from WT rats (p = 0.13339; Figures 3A,B). All female Cntnap2 KO rats showed a higher Top value than WT rats (Cntnap2 KOhet p < 0.0001, Cntnap2 KOhom p < 0.0001, Cntnap2 KOCF p = 0.0009).
There was also a significant effect of genotype on startle thresholds in male, but not female rats [males p < 0.0001, F(3, 27) = 16.96; females p = 0.1196, F(3, 33) = 2.096]. All male Cntnap2 KO groups showed a lower threshold than WT males (Cntnap2 KOhet p = 0.0001, Cntnap2 KOhom p < 0.0001, Cntnap2 KOCF p = 0.0075), however, thresholds were less reduced in male Cntnap2 KOCF than in Cntnap2 KOhom males, as they significantly differed (p = 0.0136; Figures 3C,E). Similarly, there was a significant effect of genotype in male but not female rats in ES50 [males p < 0.0001, F(3, 27) = 14.76; females p = 0.1610, F(3, 33) = 1.829]. Again, whereas all male Cntnap2 KO showed a lower ES50 than WT males (Cntnap2 KOhet p = 0.0001, Cntnap2 KOhom p < 0.0001, Cntnap2 KOCF p = 0.0333), male Cntnap2 KOCF rats showed a significantly higher ES50 than male Cntnap2 KOhom rats (p = 0.0211; Figures 3C,F). For saturation, there was no effect of genotype for males or females [males p = 0.0979, F(3, 27) = 2.318; females p = 0.0696, F(3, 33) = 2.587; Figures 3C,G]. Lastly, there was a significant effect of genotype on the slope of the startle I/O function in males [p = 0.0056, F(3, 27) = 5.240] and females [p = 0.0291, F(3, 33) = 3.399]. Both Cntnap2 KOhom males and females showed lower slopes than their WT counterparts (males p = 0.0026, females p = 0.0207; Figure 3D).
Taken together, we found that male Cntnap2 KOCF compared to male Cntnap2 KOhom exhibited a less exaggerated startle response as evidenced by a decreased maximum response (Top) as well as a less decreased threshold and ES50 of the startle response I/O curves. Cross-fostering does not appear to have affected the startle response of Cntnap2 KO females.
Cross-fostering improves PPI deficit in Cntnap2 KO rats
PPI was assessed across the entire range of startle stimulus levels (70 dB, 80 dB, 90 dB, 100 dB, 110 dB, and 120 dB) with a 75 dB and 85 dB prepulse (Figures 4A,B). With a 75 dB prepulse, there was an interaction effect between sex and startle stimulus level and between sex, genotype, and startle stimulus level [sex × startle stimulus p < 0.0001, F(5, 300) = 14.29; sex × genotype × startle stimulus p = 0.0013, F(15, 300) = 2.554]. With an 85 dB prepulse, these same interaction effects were found as well as a significant effect of sex and an interaction effect between sex and genotype [sex p < 0.0001, F(1, 60) = 286.9; sex × genotype p < 0.0001, F(3, 60) = 162.0; sex × startle stimulus p < 0.0001, F(5, 300) = 95.79; sex × genotype × startle stimulus p < 0.0001, F(15, 300) = 32.64]. Therefore, for both prepulse levels, males and females were separated for further analysis.

Figure 4. Cross-fostered Cntnap2 knockout males and females show improved PPI. (A) PPI (% inhibition) across startle stimulus intensity levels with the 75 dB prepulse. # denotes startle stimulus intensities where there were group differences in PPI. Stimulus levels highlighted by gray rectangles indicate significant improvements by cross-fostering and are presented in the bottom panel. (B) PPI analysis across startle stimulus intensity with the 85 dB prepulse. (C) PPI of males with 100 dB startle stimulus with a 75 dB prepulse. PPI in male Cntnap2 KOCF is not different from WT. (D) PPI in males at the 110 dB startle stimulus with a 75 dB prepulse. Again, PPI of male Cntnap2 KOCF is not different from WT. (E) PPI of females at the 110 dB startle stimulus with a 75 dB prepulse. PPI of Cntnap2 KOCF females is not different from WT females. (F) PPI of males at the 100 dB startle stimulus with an 85 dB prepulse. PPI of Cntnap2 KOCF males is not different from WT males. *p < 0.05, **p < 0.01, ***p < 0.0001.
With a 75 dB prepulse, there was a significant effect of startle stimulus level and an interaction effect of genotype and startle stimulus level for males [startle stimulus level p < 0.0001, F(5, 135) = 33.89; genotype × startle stimulus p < 0.0001, F(15, 135) = 7.992] and females [startle stimulus level p < 0.0001, F(5, 165) = 43.22; genotype × startle stimulus p < 0.0001, F(15, 135) = 3.370; Figure 4A]. Post-hoc tests revealed PPI differences in males at the 80 dB, 100 dB, and 110 dB startle stimulus. At the 80 dB stimulus, all Cntnap2 KO males showed greater PPI than WT males (Cntnap2 KOhet p < 0.0001, Cntnap2 KOhom p = 0.0002, Cntnap2 KOCF p = 0.0011). Interestingly, whereas Cntnap2 KOhet and KOhom males exhibited lower PPI than WT males at the 100 dB stimulus (Cntnap2 KOhet p = 0.0337, Cntnap2 KOhom p = 0.0009) and 110 dB stimulus (Cntnap2 KOhet p = 0.0425, Cntnap2 KOhom p = 0.0077), PPI in KOCF males did not differ from WT males (100 dB p = 0.2216, 110 dB p = 0.1439; Figures 4C,D).
For females with a 75 dB prepulse, similar effects were found at the 110 dB startle stimulus: whereas Cntnap2 KOhet and KOhom females showed lower PPI than WT females (Cntnap2 KOhet p = 0.0112, Cntnap2 KOhom p = 0.0399), PPI in KOCF females did not differ from WT females (p = 0.3114; Figure 4E). Cntnap2 KOCF females also showed greater PPI than WT at the 80 dB startle stimulus (p = 0.0039), but lower PPI at the 120 dB stimulus (p = 0.0465).
With an 85 dB prepulse, there was a main effect of genotype and startle stimulus intensity, as well as an interaction effect for males [genotype p < 0.0001, F(3, 27) = 55.97; startle stimulus level p < 0.0001, F(5, 135) = 27.32; genotype × startle stimulus p < 0.0001, F(15, 135) = 12.90] and females [genotype p = 0.0033, F(3, 33) = 5.563; startle stimulus level p < 0.0001, F(5, 165) = 92/57; genotype × startle stimulus p = 0.0013, F(15, 165) = 2.644; Figure 4B]. Similarly to what was observed with the 75 dB prepulse, at the 100 dB startle stimulus Cntnap2 KOhet and KOhom males showed lower PPI than WT males (Cntnap2 KOhet p = 0.0116, Cntnap2 KOhom p = 0.0021), but KOCF males did not differ from WT males (p = 0.2259; Figure 4F). With the 120 dB startle stimulus, Cntnap2 KOhom but not KOCF males had a lower PPI than WT males (Cntnap2 KOhom p = 0.0252, Cntnap2 KOCF p = 0.2206). With the 85 dB prepulse, females only differed at the 90 dB stimulus, where Cntnap2 KOCF females exhibited even greater PPI than WT females (p = 0.0177).
In summary, cross-fostering showed positive effects on PPI at both prepulse intensities for males and females. Cross-fostered males and females exhibited ameliorated PPI impairment at the higher startle stimulus intensities. In addition, Cntnap2 KOCF females showed even better PPI than WT animals at lower startle stimulus intensities.
We further examined how cross-fostering affects PPI by assessing how the baseline I/O curve changes with adding a prepulse. As described by Doornaert et al. and El-Cheikh Mohamad et al., the entire baseline I/O curve can be scaled by two different components: startle and sound scaling (2023; 2024). Startle scaling, as shown by a downward shift in the I/O curve, results from a reduction in response amplitude, whereas sound scaling, as shown by a rightward shift in the I/O curve, results from a reduction in sound sensitivity. Like our previous study, here we show that Cntnap2 KO rats have intact startle scaling but altered sound scaling compared to WT rats (Doornaert et al., 2024). However, Cntnap2 KOCF animals did not differ significantly from KOhom animals in our measures of startle and sound scaling (see Supplementary Figures 7, 8). Therefore, we cannot use our differentiation of startle and sound scale components to explain the improved PPI observed in Cntnap2 KOCF.
Cross-fostering does not alleviate the gap-PPI impairment in Cntnap2 KO rats; rather, it may exacerbate the impairment
Gap-PPI was assessed using small gaps in the background noise (2 ms, 5 ms, and 10 ms; Figure 5A) and long gap durations (20 ms, 40 ms, 60 ms, 75 ms, and 100 ms; Figure 5B). For small and long gaps, there was a significant effect of sex on PPI, and therefore, males and females were separated for further analysis [short p = 0.0100, F(1, 66) = 7.031; long p < 0.0001, F(1, 66) = 39.99].
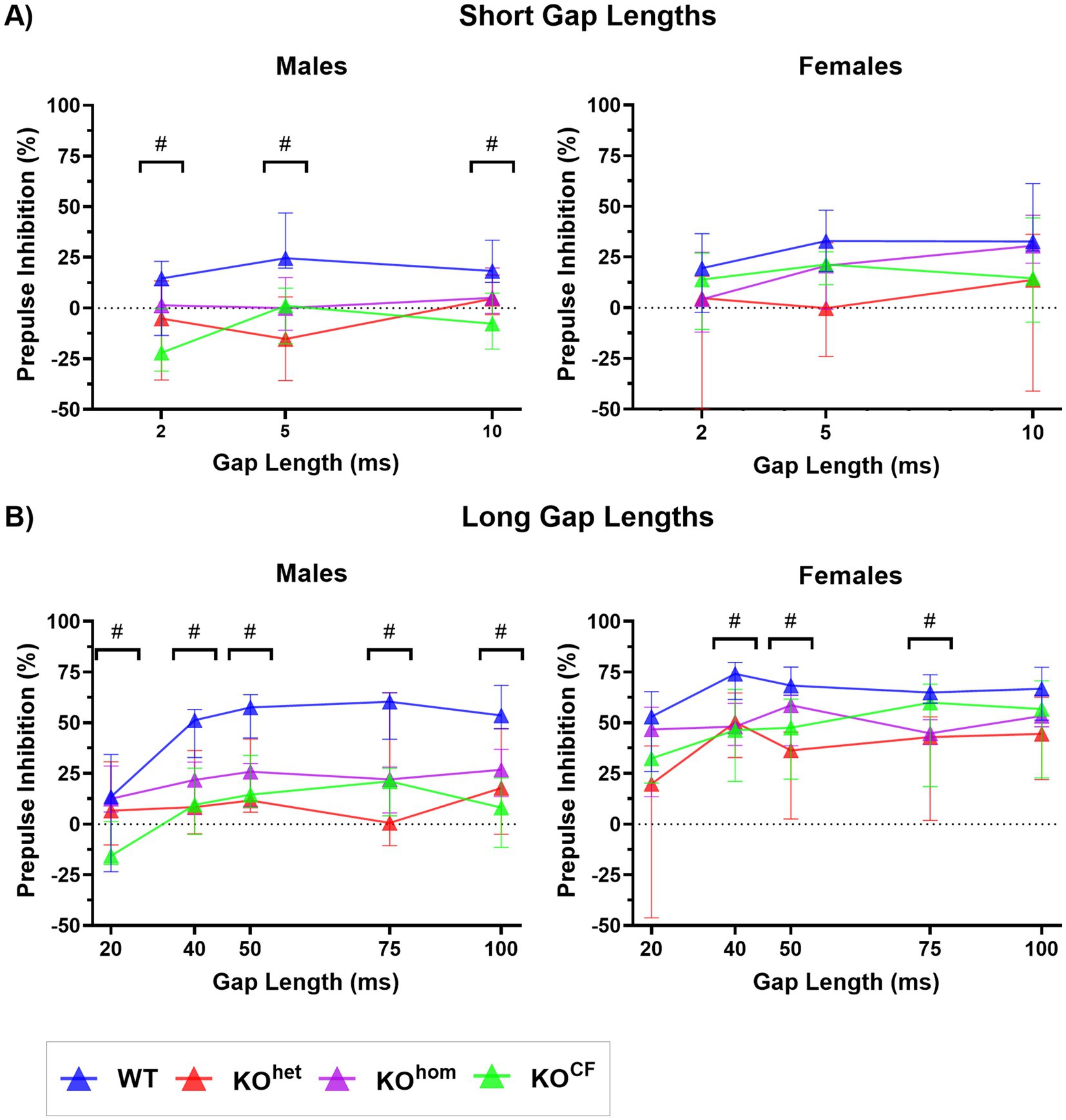
Figure 5. Gap-PPI impairment found in Cntnap2 knockout rats is not improved by cross-fostering. (A) PPI (% inhibition) with short gap lengths (2 ms, 5 ms, and 10 ms). Male Cntnap2 KOhom showed a lower PPI with a 5 ms gap than WT rats, whereas KOCF rats had a lower %PPI than WT rats at all short gap lengths. PPI did not differ between female Cntnap2 WT and KO rats for short gap lengths. (B) PPI with long gap lengths (20 ms, 40 ms, 50 ms, 75 ms, and 100 ms). All Cntnap2 KO males had a lower PPI than WT males for gap lengths between 40 ms and 100 ms. At 20 ms, Cntnap2 KOCF males had a lower PPI than KOhom males. Cntnap2 KOhom and KOCF females had lower PPI than WT females with a 40 ms gap length. Female Cntnap2 KOCF but not KOhom showed a lower PPI than WT with a 50 ms gap length, and female Cntnap2 KOhom but not KOCF had a lower PPI than WT with a 75 ms gap length. # denotes gap lengths where there were group differences in PPI.
For the short gaps, there was a significant effect of genotype and gap length on PPI in males [genotype p = 0.0002, F(3, 35) = 8.533; gap length p = 0.0134, F(2, 70) = 4.591]. Only male Cntnap2 KOCF rats had a lower PPI than WT males with a 2 ms and 10 ms gap (2 ms p = 0.0360, 10 ms p = 0.0062; Figure 5A), whereas with a 5 ms gap, all male KO rats regardless of group showed lower PPI than WT males (Cntnap2 KOhet p = 0.0017, Cntnap2 KOhom p = 0.0140, Cntnap2 KOCF p = 0.0191). Although females also had a significant effect of genotype and gap length on PPI for short gaps, there were no significant differences found between groups following post-hoc testing [genotype p = 0.0481, F(3, 31) = 2.948; gap length p = 0.0197, F(2, 62) = 4.186].
For long gaps, there was a significant effect of genotype and gap length on PPI, as well as an interaction effect between genotype and gap length [genotype p = 0.0003, F(3, 35) = 8.333; gap length p < 0.0001, F(4, 140) = 12.23; genotype × gap length p = 0.0053, F(12, 140) = 2.502]. For the 20 ms gap, male Cntnap2 KOCF rats showed lower PPI than male WT animals (p = 0.0139; Figure 5B). At all longer gap lengths, all male Cntnap2 KO rats regardless of group had lower PPI than WT males (40 ms: Cntnap2 KOhet p = 0.0498, Cntnap2 KOhom p = 0.0198, Cntnap2 KOCF p = 0.0117; 50 ms: Cntnap2 KOhet p = 0.0207, Cntnap2 KOhom p = 0.0076, Cntnap2 KOCF p = 0.0028; 75 ms: Cntnap2 KOhet p = 0.0259, Cntnap2 KOhom p = 0.0015, Cntnap2 KOCF p = 0.0014; 100 ms: Cntnap2 KOhet p = 0.0418, Cntnap2 KOhom p = 0.0206, Cntnap2 KOCF p = 0.0011). For females, there was again a significant effect of genotype and gap length on PPI [genotype p = 0.0059, F(3, 31) = 5.028; gap length p = 0.0003, F(4, 124) = 5.66]. With a 40 ms gap, female Cntnap2 KOhom and KOCF rats had lower PPI than WT rats (Cntnap2 KOhom p = 0.0102, Cntnap2 KOCF p = 0.0315). With a 50 ms gap length, Cntnap2 KOCF but not KOhom showed lower PI than female WT rats (Cntnap2 KOhom p = 0.1098, Cntnap2 KOCF p = 0.0127). In contrast, Cntnap2 KOhom, but not KOCF, had a lower PPI than WT females with a 75 ms gap (Cntnap2 KOhom p = 0.0008, Cntnap2 KOCF p = 0.2151). Overall, it appears that cross-fostering did not reverse the gap-PPI impairment observed in Cntnap2 KO rats but rather exacerbated the deficit at certain gap lengths.
Discussion
This study investigated the impact of early postnatal environmental factors on long-term auditory processing and sensorimotor gating in the Cntnap2 KO rat model. Cntnap2 KOCF rats showed partial restoration of auditory maturation, evidenced by improved hearing sensitivity, a less exaggerated startle response, and enhanced PPI compared to Cntnap2 KOhom rats. These results indicate that early environmental interventions can positively modulate some deficits associated with the loss-of-function of Cntnap2. Despite these improvements, Cntnap2 KOCF rats also displayed more pronounced changes in the ABR, including reduced neural responsiveness and slower conduction times, along with persistent gap-PPI impairments. These findings highlight that while the early postnatal environment can modify certain effects of the loss of Cntnap2, it may also exacerbate other deficits, illustrating the complex nature of gene–environment interactions in neurodevelopment.
Cross-fostering has both beneficial and consequential effects on ABR
Hearing sensitivity, measured by the threshold to a click stimulus, typically increases from the postnatal period to adulthood as the auditory pathway matures (Smith and Kraus, 1987; Hang et al., 2016). Notably, this maturation of hearing sensitivity was not observed in Cntnap2 KOhom animals but was restored in KOCF animals. While the ABR threshold reflects overall auditory pathway function, wave amplitudes and latencies provide more insights into neural activity and conduction. Adult Cntnap2 KOCF animals exhibited smaller wave I amplitudes than WT, suggesting reduced auditory nerve activity, and slower interpeak latencies, indicating impaired conduction between brainstem regions. Hence, cross-fostering reversed hearing sensitivity deficits, but neural responsiveness and conduction of the auditory brainstem remained impaired.
Cntnap2 plays an important role in neural development, myelination, and synaptic function, which is important particularly in the development of pathways requiring precise timing (Truong et al., 2015; Gordon et al., 2016; Poot, 2017). Cross-fostering during the critical period exposes Cntnap2 KO animals to an enriched sensory/auditory environment, likely promoting neural plasticity, and potentially improving ABR thresholds through mechanisms like synaptic upscaling or increased neurotransmitter release in central auditory structures. However, the lower wave I amplitude suggests that these brainstem and/or cortical compensatory mechanisms may strain peripheral auditory nerve function perhaps through increased efferent feedback (Boothalingam et al., 2023). The slower interpeak latencies in our Cntnap2 KO animals reflect impaired synaptic transmission and/or myelination, a common observation in Cntnap2 KO models (Varea et al., 2015; Scott et al., 2019, 2022). These neural conduction deficits persisted despite cross-fostering, suggesting that environmental factors cannot fully compensate for these Cntnap2-related impairments.
The finding that cross-fostered animals exhibited greater ABR disruptions, particularly reduced wave I amplitude and prolonged wave I–IV interpeak latency, suggests that cross-fostering exerts a stronger influence on central auditory plasticity than on peripheral conduction integrity. The reduced wave I amplitude indicates diminished auditory nerve responsiveness, possibly due to increased efferent modulation from central auditory structures, such as the medial olivocochlear system, which regulates peripheral auditory excitability (Boothalingam et al., 2023). This could reflect a compensatory mechanism aimed at reducing auditory hyperexcitability at higher processing levels. However, the persistent delays in interpeak latencies suggest that environmental enrichment is insufficient to overcome myelination-dependent deficits in synaptic transmission and conduction velocity within the auditory brainstem. While cross-fostering may enhance auditory plasticity by altering central processing, it does not fully restore the timing-dependent aspects of auditory function that rely on intact myelination.
Cross-fostering reduces heightened startle in Cntnap2 KO males and improves PPI deficit in Cntnap2 KO males and females
Cross-fostering alleviated increased startle responses and PPI deficits in Cntnap2 KO rats, with improvements in startle responses found in males and in PPI observed in both males and females. These changes likely result from enhanced neural plasticity during the critical period of auditory development. The startle response is regulated by a well-characterized brainstem pathway involving cochlear root neurons, the caudal pontine reticular nucleus (PnC), and spinal cord motor neurons (for review see Koch, 1999; Zheng and Schmid, 2023). PPI is thought to be mediated by a feed-forward inhibitory circuit starting with cochlear neurons projecting to the inferior colliculus and the pedunculopontine tegmental nucleus (PPTg; Koch, 1999). Cntnap2 is prominently expressed in the neural circuits governing startle and PPI, and its dysfunction likely disrupts the balance of excitatory and inhibitory signaling in these pathways (Gordon et al., 2016; Scott et al., 2018). Previous studies have shown elevated levels of excitatory and inhibitory neurotransmitters in the PnC of Cntnap2 KO rats and altered firing rates in response to startling sounds that likely account for the increased startle (Möhrle et al., 2021; Zheng et al., 2023). Besides these changes in the primary startle pathway, minimal differences were observed in PPTg firing rates and PnC inhibition from the PPTg in Cntnap2 KO rats, indicating that these structures do not account for the reduced PPI (Zheng et al., 2023). Cross-fostering may mitigate increased startle and impaired PPI by decreasing PnC hyperexcitability due to enriched sensory input during the critical period, possibly through increased GABAergic signaling or reduced glutamatergic activity. Similar effects have been observed with pharmacological treatments like R-Baclofen, which dampens hyperexcitability in the startle pathway (Möhrle et al., 2021). Cross-fostering may also induce structural changes in the PnC, such as increased dendritic branching, enhanced spine density, and improved myelination, all of which are influenced by Cntnap2 expression (Anderson et al., 2012; Scott et al., 2019).
The observed sex differences in startle response and PPI in Cntnap2 KO rats may be attributed to distinct underlying neural mechanisms. Prior research suggests that heightened startle in male and female Cntnap2 KO rats arises from different factors: males exhibit both increased startle magnitude and a leftward shift in the startle I/O function, indicating heightened sound sensitivity, whereas females show only increased startle magnitude (Doornaert et al., 2024). In vivo electrophysiological recordings further support this distinction, as female Cntnap2 KO rats display increased firing rates of PnC neurons in response to startle stimuli, while males exhibit enhanced recruitment of startle-mediating neurons, potentially accounting for their increased sound sensitivity (Zheng et al., 2023, 2024). Given these sex-specific differences, cross-fostering may impact males and females differently by modulating PnC hyperexcitability through distinct pathways. For instance, in males, cross-fostering could reduce the recruitment of startle-mediating neurons, thereby normalizing the leftward shift in the I/O function, while in females, it may directly attenuate PnC hyperactivity, reducing startle magnitude.
These findings align with studies of the effects of environmental enrichment on startle and PPI, which have shown beneficial effects in the valproic acid rat model and mGlur5 KO mice (Schneider et al., 2006; Burrows et al., 2015). Cross-fostering may act as a form of environmental enrichment by providing increased maternal care, exposure to social ultrasonic vocalizations, and varied social interactions with littermates. These changes could modulate sensory processing in Cntnap2 KO rats and improve the function of startle and PPI networks. Future studies will address the effect of housing Cntnap2 KO rats in sensory-enriched environments during early development on sensory processing.
Gap-PPI deficit revealed in Cntnap2 KO rat; not improved by cross-fostering
This study is the first to evaluate gap-PPI in the Cntnap2 KO rat, which assesses an animal’s ability to perceive and respond to brief gaps in continuous noise, reflecting auditory temporal resolution (Fournier and Hébert, 2016). This capability is crucial for speech and sound processing, especially in distinguishing rapid changes in sound patterns (Poldrack et al., 2001). Unlike traditional PPI, gap-PPI involves greater engagement of the primary auditory cortex (Bowen et al., 2003; Weible et al., 2014; Moreno-Paublete et al., 2017). While gap-PPI has often been used to assess tinnitus (Galazyuk and Hébert, 2015), recent findings suggest it may also serve as a biomarker for ASD (Foss-Feig et al., 2018). In fact, Cntnap2 KO mice have shown deficits in gap-PPI, specifically exhibiting a significantly higher detection threshold for short gaps compared to control mice (Truong et al., 2015). We observed a similar pattern of impairment in Cntnap2 KO rats, with short gaps affected primarily in males and longer gaps impaired in both males and females. These deficits are likely due to immature and hyperexcitable auditory cortex neurons (Scott et al., 2022), which may elevate “neuronal noise,” making it harder to distinguish relevant stimuli from background noise.
Cross-fostering did not improve gap-PPI and even exacerbated impairments at certain gap lengths. This suggests that gap-PPI, which relies heavily on cortical processing (Bowen et al., 2003; Weible et al., 2014; Moreno-Paublete et al., 2017), may not benefit from cross-fostering, unlike startle and prepulse inhibition, which are largely brainstem-dependent (Wright and Barnes, 1972; Hammond, 1974; Davis and Gendelman, 1977; Hunter and Willott, 1993). The finding that cross-fostering exacerbated gap-PPI deficits highlights the complexity of gene–environment interactions in auditory processing. Given that early environmental interventions can influence neurodevelopment by altering sensory input and plasticity, it is possible that increased environmental stimulation further disrupted auditory cortical processing rather than normalizing it. This could be due to excessive neural excitability in the auditory cortex of Cntnap2 KO rats (Scott et al., 2022; Mann et al., 2023) where additional sensory input amplifies rather than mitigates cortical dysregulation. Alternatively, cross-fostering might have affected the development of inhibitory circuits in a way that selectively worsened temporal resolution deficits. Future studies examining cortical excitability, inhibitory signaling, and synaptic plasticity in the auditory cortex following cross-fostering could help elucidate the mechanisms underlying this unexpected outcome.
Limitations and future directions
This study is limited in its ability to identify the specific environmental factors being manipulated by the cross-fostering paradigm. As noted earlier, variations in maternal care may exist between Cntnap2 KO and HET dams. Maternal behaviors, such as grooming and milk quality, are known to significantly impact pup development (Champagne et al., 2003; Qiu et al., 2020; DeRosa et al., 2022). Another factor to consider is the genetic composition of the litter. While homozygous breeding results in exclusively homozygous offspring, heterozygous breeding produces a mix of homozygous, heterozygous, and wildtype pups. Consequently, cross-fostered animals are exposed to different social environments, which could influence social communication and interactions, ultimately shaping their developmental outcomes. Moreover, the process of cross-fostering could induce stress on the pups, again potentially affecting development (Vorhees et al., 2024). Notwithstanding the challenges of identifying the crucial factors, our findings demonstrate that environmental influences induced by cross-fostering can significantly alter developmental outcomes regarding auditory brainstem processing.
Another limitation of this study is the one-way cross-fostering design as we did not cross-foster Cntnap2 KO pups bred from a Cntnap2 HET dam to a KO dam. Cross-fostering was limited to litters born within 48 h to align with the critical period for early auditory development. Given the need for many more litters to be born within this period and the low reproductive success rate of homozygous KO dams, implementing reciprocal cross-fostering was impractical. As a result, while the benefits observed in the heterozygous environment are noted, the potential positive, negative, or neutral impacts of the homozygous environment on sensory processing remain undetermined.
Future research should investigate prenatal factors that may differ between heterozygous and homozygous pairings. Variations in dam health, including hormonal, metabolic, and stress responses, could influence fetal development and brain function (Fatima et al., 2017; Baud and Berkane, 2019). Homozygous KO dams may experience distinct prenatal environments, such as altered uterine blood flow, oxygen levels, and placental function, which could impact critical physiological processes during pregnancy (Hufnagel et al., 2022). Additionally, maternal mitochondrial DNA (mtDNA), which is inherited maternally, could play a role in neurodevelopmental outcomes, especially since mitochondrial dysfunction has been repeatedly linked to autism (Wang et al., 2022; Al-Kafaji et al., 2023). Lastly, investigating the role of parental vocalizations during the critical period could further enhance understanding of how early auditory experiences, such as exposure to biological parental sounds during cross-fostering, shape sensory processing and developmental outcomes in ASD models.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary material, further inquiries can be directed to the corresponding author.
Ethics statement
The animal study was approved by Animal Care Committee, University of Western Ontario. The study was conducted in accordance with the local legislation and institutional requirements.
Author contributions
ED: Data curation, Formal analysis, Investigation, Methodology, Software, Visualization, Writing – original draft, Writing – review & editing. AM: Investigation, Methodology, Validation, Writing – review & editing. GJ: Data curation, Investigation, Writing – review & editing. BA: Conceptualization, Methodology, Supervision, Validation, Writing – review & editing. DM: Conceptualization, Funding acquisition, Methodology, Resources, Software, Supervision, Validation, Writing – review & editing. SS: Conceptualization, Data curation, Funding acquisition, Project administration, Resources, Supervision, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research, authorship, and/or publication of this article. This study was supported by a project grant of the Canadian Institute of Health Research (CIHR, PJF168866) and the Natural Sciences and Engineering Council Discovery grant (NSERC 04472-2018RGPIN) to SS, the Deutsche Forschungsgemeinschaft Fellowship (DFG 442662585) and a 2021C. Kingsley Allison Research Grant to DM.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
Generative AI statement
The authors declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fnins.2025.1565919/full#supplementary-material
References
Al-Kafaji, G., Jahrami, H. A., Alwehaidah, M. S., Alshammari, Y., and Husni, M. (2023). Mitochondrial DNA copy number in autism spectrum disorder and attention deficit hyperactivity disorder: a systematic review and meta-analysis. Front. Psychol. 14:1196035. doi: 10.3389/fpsyt.2023.1196035
Anderson, G. R., Galfin, T., Xu, W., Aoto, J., Malenka, R. C., and Südhof, T. C. (2012). Candidate autism gene screen identifies critical role for cell-adhesion molecule CASPR2 in dendritic arborization and spine development. Proc. Natl. Acad. Sci. USA 109, 18120–18125. doi: 10.1073/pnas.1216398109
Arking, D. E., Cutler, D. J., Brune, C. W., Teslovich, T. M., West, K., Ikeda, M., et al. (2008). A common genetic variant in the Neurexin superfamily member CNTNAP2 increases familial risk of autism. Am. J. Hum. Genet. 82, 160–164. doi: 10.1016/j.ajhg.2007.09.015
Baud, O., and Berkane, N. (2019). Hormonal changes associated with intra-uterine growth restriction: impact on the developing brain and future neurodevelopment. Front Endocrinol (Lausanne) 10:179. doi: 10.3389/fendo.2019.00179
Bi, C., Cui, Y., Mao, Y., Dong, S., Zhang, J., and Sun, X. (2006). The effect of early auditory deprivation on the age-dependent expression pattern of NR2B mRNA in rat auditory cortex. Brain Res. 1110, 30–38. doi: 10.1016/j.brainres.2006.06.056
Boothalingam, S., Peterson, A., Powell, L., and Easwar, V. (2023). Auditory brainstem mechanisms likely compensate for self-imposed peripheral inhibition. Sci. Rep. 13:12693. doi: 10.1038/s41598-023-39850-8
Bowen, G. P., Lin, D., Taylor, M. K., and Ison, J. R. (2003). Auditory cortex lesions in the rat impair both temporal acuity and noise increment thresholds, revealing a common neural substrate. Cereb. Cortex 13, 815–822. doi: 10.1093/cercor/13.8.815
Bureš, Z., Bartošová, J., Lindovský, J., Chumak, T., Popelář, J., and Syka, J. (2014). Acoustical enrichment during early postnatal development changes response properties of inferior colliculus neurons in rats. Eur. J. Neurosci. 40, 3674–3683. doi: 10.1111/ejn.12732
Bures, Z., Grécová, J., Popelár, J., and Syka, J. (2010). Noise exposure during early development impairs the processing of sound intensity in adult rats. Eur. J. Neurosci. 32, 155–164. doi: 10.1111/j.1460-9568.2010.07280.x
Bureš, Z., Popelář, J., and Syka, J. (2017). The effect of noise exposure during the developmental period on the function of the auditory system. Hear. Res. 352, 1–11. doi: 10.1016/j.heares.2016.03.008
Burrows, E. L., McOmish, C. E., Buret, L. S., Van den Buuse, M., and Hannan, A. J. (2015). Environmental enrichment ameliorates behavioral impairments modeling schizophrenia in mice lacking metabotropic glutamate receptor 5. Neuropsychopharmacology 40, 1947–1956. doi: 10.1038/npp.2015.44
Champagne, F. A., Francis, D. D., Mar, A., and Meaney, M. J. (2003). Variations in maternal care in the rat as a mediating influence for the effects of environment on development. Physiol. Behav. 79, 359–371. doi: 10.1016/S0031-9384(03)00149-5
Chang, E. F., and Merzenich, M. M. (2003). Environmental noise retards auditory cortical development. Science 300, 498–502. doi: 10.1126/science.1082163
Davis, M., and Gendelman, P. M. (1977). Plasticity of the acoustic startle response in the acutely decerebrate rat. J. Comp. Physiol. Psychol. 91, 549–563. doi: 10.1037/h0077345
DeRosa, H., Caradonna, S. G., Tran, H., Marrocco, J., and Kentner, A. C. (2022). Got Milk? Maternal immune activation during the mid-lactational period affects nutritional milk quality and adolescent offspring sensory processing in male and female rats. Mol. Psychiatry 27, 4829–4842. doi: 10.1038/s41380-022-01744-y
Doornaert, E. E., Mohamad, A. E.-C., Johal, G., Allman, B., Möhrle, D., and Schmid, S. (2024). Not a deficit, just different - Prepulse inhibition disruptions in autism depend on startle stimulus intensities. eNeuro. 11, ENEURO.0179–ENEU24.2024. doi: 10.1523/ENEURO.0179-24.2024
El-Cheikh Mohamad, A., Möhrle, D., Haddad, F. L., Rose, A., Allman, B. L., and Schmid, S. (2023). Assessing the Cntnap2 knockout rat prepulse inhibition deficit through prepulse scaling of the baseline startle response curve. Transl. Psychiatry 13, 321–311. doi: 10.1038/s41398-023-02629-6
Elkin, L. A., Kay, M., Higgins, J. J., and Wobbrock, J. O. (2021). “An aligned rank transform procedure for multifactor contrast tests.” in The 34th annual ACM symposium on user Interface software and technology, (Virtual Event USA: ACM), 754–768.
Engineer, N. D., Percaccio, C. R., Pandya, P. K., Moucha, R., Rathbun, D. L., and Kilgard, M. P. (2004). Environmental enrichment improves response strength, threshold, selectivity, and latency of auditory cortex neurons. J. Neurophysiol. 92, 73–82. doi: 10.1152/jn.00059.2004
Fatima, M., Srivastav, S., and Mondal, A. C. (2017). Prenatal stress and depression associated neuronal development in neonates. Int. J. Dev. Neurosci. 60, 1–7. doi: 10.1016/j.ijdevneu.2017.04.001
Foss-Feig, J. H., Stavropoulos, K. K. M., McPartland, J. C., Wallace, M. T., Stone, W. L., and Key, A. P. (2018). Electrophysiological response during auditory gap detection: biomarker for sensory and communication alterations in autism spectrum disorder? Dev. Neuropsychol. 43, 109–122. doi: 10.1080/87565641.2017.1365869
Fournier, P., and Hébert, S. (2016). The gap-startle paradigm to assess auditory temporal processing: bridging animal and human research. Psychophysiology 53, 759–766. doi: 10.1111/psyp.12620
Galazyuk, A., and Hébert, S. (2015). Gap-Prepulse inhibition of the acoustic startle reflex (GPIAS) for tinnitus assessment: current status and future directions. Front. Neurol. 6:88. doi: 10.3389/fneur.2015.00088
Gordon, A., Salomon, D., Barak, N., Pen, Y., Tsoory, M., Kimchi, T., et al. (2016). Expression of Cntnap2 (Caspr2) in multiple levels of sensory systems. Mol. Cell. Neurosci. 70, 42–53. doi: 10.1016/j.mcn.2015.11.012
Hammond, G. R. (1974). Frontal cortical lesions and prestimulus inhibition of the rat’s acoustic startle reaction. Psychobiology 2, 151–156. doi: 10.3758/BF03333016
Hang, J., Pan, W., Chang, A., Li, S., Li, C., Fu, M., et al. (2016). Synchronized progression of Prestin expression and auditory brainstem response during postnatal development in rats. Neural Plast. 2016, 4545826–4545810. doi: 10.1155/2016/4545826
Hufnagel, A., Grant, I. D., and Aiken, C. E. M. (2022). Glucose and oxygen in the early intrauterine environment and their role in developmental abnormalities. Semin. Cell Dev. Biol. 131, 25–34. doi: 10.1016/j.semcdb.2022.03.041
Hunter, K. P., and Willott, J. F. (1993). Effects of bilateral lesions of auditory cortex in mice on the acoustic startle response. Physiol. Behav. 54, 1133–1139. doi: 10.1016/0031-9384(93)90337-F
Inguaggiato, E., Sgandurra, G., and Cioni, G. (2017). Brain plasticity and early development: implications for early intervention in neurodevelopmental disorders. Neuropsychiatr. Enfance Adolesc. 65, 299–306. doi: 10.1016/j.neurenf.2017.03.009
Koch, M. (1999). The neurobiology of startle. Prog. Neurobiol. 59, 107–128. doi: 10.1016/S0301-0082(98)00098-7
Kral, A., Hartmann, R., Tillein, J., Heid, S., and Klinke, R. (2001). Delayed maturation and sensitive periods in the auditory cortex. Audiol. Neurootol. 6, 346–362. doi: 10.1159/000046845
Lord, C., Elsabbagh, M., Baird, G., and Veenstra-Vanderweele, J. (2018). Autism spectrum disorder. Lancet 392, 508–520. doi: 10.1016/S0140-6736(18)31129-2
Mann, R. S., Allman, B. L., and Schmid, S. (2023). Developmental changes in electrophysiological properties of auditory cortical neurons in the Cntnap2 knockout rat. J. Neurophysiol. 129, 937–947. doi: 10.1152/jn.00029.2022
Martin-Iverson, M. T., and Stevenson, K. N. (2005). Apomorphine effects on emotional modulation of the startle reflex in rats. Psychopharmacology 181, 60–70. doi: 10.1007/s00213-005-2217-3
Miller, E. A., Kastner, D. B., Grzybowski, M. N., Dwinell, M. R., Geurts, A. M., and Frank, L. M. (2021). Robust and replicable measurement for prepulse inhibition of the acoustic startle response. Mol. Psychiatry 26, 1909–1927. doi: 10.1038/s41380-020-0703-y
Miron, O., Ari-Even Roth, D., Gabis, L. V., Henkin, Y., Shefer, S., Dinstein, I., et al. (2016). Prolonged auditory brainstem responses in infants with autism. Autism Res. 9, 689–695. doi: 10.1002/aur.1561
Miron, O., Beam, A. L., and Kohane, I. S. (2018). Auditory brainstem response in infants and children with autism spectrum disorder: a meta-analysis of wave V. Autism Res. 11, 355–363. doi: 10.1002/aur.1886
Möhrle, D., Hofmeier, B., Amend, M., Wolpert, S., Ni, K., Bing, D., et al. (2019). Enhanced central neural gain compensates acoustic trauma-induced Cochlear impairment, but unlikely correlates with tinnitus and Hyperacusis. Neuroscience 407, 146–169. doi: 10.1016/j.neuroscience.2018.12.038
Möhrle, D., Ni, K., Varakina, K., Bing, D., Lee, S. C., Zimmermann, U., et al. (2016). Loss of auditory sensitivity from inner hair cell synaptopathy can be centrally compensated in the young but not old brain. Neurobiol. Aging 44, 173–184. doi: 10.1016/j.neurobiolaging.2016.05.001
Möhrle, D., Wang, W., Whitehead, S. N., and Schmid, S. (2021). GABAB receptor agonist R-baclofen reverses altered auditory reactivity and filtering in the Cntnap2 Knock-out rat. Front. Integr. Neurosci. 15:710593. doi: 10.3389/fnint.2021.710593
Möhrle, D., Yuen, M., Zheng, A., Haddad, F. L., Allman, B. L., and Schmid, S. (2023). Characterizing maternal isolation-induced ultrasonic vocalizations in a gene–environment interaction rat model for autism. Genes Brain Behav. 22:e12841. doi: 10.1111/gbb.12841
Moreno-Paublete, R., Canlon, B., and Cederroth, C. R. (2017). Differential neural responses underlying the inhibition of the startle response by pre-pulses or gaps in mice. Front. Cell. Neurosci. 11:19. doi: 10.3389/fncel.2017.00019
Percaccio, C. R., Pruette, A. L., Mistry, S. T., Chen, Y. H., and Kilgard, M. P. (2007). Sensory experience determines enrichment-induced plasticity in rat auditory cortex. Brain Res. 1174, 76–91. doi: 10.1016/j.brainres.2007.07.062
Poldrack, R. A., Temple, E., Protopapas, A., Nagarajan, S., Tallal, P., Merzenich, M., et al. (2001). Relations between the neural bases of dynamic auditory processing and phonological processing: evidence from fMRI. J. Cogn. Neurosci. 13, 687–697. doi: 10.1162/089892901750363235
Poot, M. (2017). Intragenic CNTNAP2 deletions: a bridge too far? Mol Syndromol 8, 118–130. doi: 10.1159/000456021
Qiu, J., Singh, P., Pan, G., de Paolis, A., Champagne, F. A., Liu, J., et al. (2020). Defining the relationship between maternal care behavior and sensory development in Wistar rats: auditory periphery development, eye opening and brain gene expression. PLoS One 15:e0237933. doi: 10.1371/journal.pone.0237933
Roth, D. A.-E., Muchnik, C., Shabtai, E., Hildesheimer, M., and Henkin, Y. (2012). Evidence for atypical auditory brainstem responses in young children with suspected autism spectrum disorders. Dev. Med. Child Neurol. 54, 23–29. doi: 10.1111/j.1469-8749.2011.04149.x
Schneider, T., Turczak, J., and Przewłocki, R. (2006). Environmental enrichment reverses behavioral alterations in rats prenatally exposed to Valproic acid: issues for a therapeutic approach in autism. Neuropsychopharmacology 31, 36–46. doi: 10.1038/sj.npp.1300767
Schormans, A. L., Scott, K. E., Vo, A. M. Q., Tyker, A., Typlt, M., Stolzberg, D., et al. (2017). Audiovisual temporal processing and synchrony perception in the rat. Front. Behav. Neurosci. 10:246. doi: 10.3389/fnbeh.2016.00246
Scott, K. E., Kazazian, K., Mann, R. S., Möhrle, D., Schormans, A. L., Schmid, S., et al. (2020). Loss of Cntnap2 in the rat causes autism-related alterations in social interactions, stereotypic behavior, and sensory processing. Autism Res. 13, 1698–1717. doi: 10.1002/aur.2364
Scott, K. E., Mann, R. S., Schormans, A. L., Schmid, S., and Allman, B. L. (2022). Hyperexcitable and immature-like neuronal activity in the auditory cortex of adult rats lacking the language-linked CNTNAP2 gene. Cereb. Cortex 32, 4797–4817. doi: 10.1093/cercor/bhab517
Scott, R., Sánchez-Aguilera, A., van Elst, K., Lim, L., Dehorter, N., Bae, S. E., et al. (2019). Loss of Cntnap2 causes axonal excitability deficits, developmental delay in cortical myelination, and abnormal stereotyped motor behavior. Cereb. Cortex 29, 586–597. doi: 10.1093/cercor/bhx341
Scott, K. E., Schormans, A. L., Pacoli, K. Y., De Oliveira, C., Allman, B. L., and Schmid, S. (2018). Altered auditory processing, filtering, and reactivity in the Cntnap2 Knock-out rat model for neurodevelopmental disorders. J. Neurosci. 38, 8588–8604. doi: 10.1523/JNEUROSCI.0759-18.2018
Smith, D. I., and Kraus, N. (1987). Postnatal development of the auditory brainstem response (ABR) in the unanesthetized gerbil. Hear. Res. 27, 157–164. doi: 10.1016/0378-5955(87)90016-5
Strauss, K. A., Puffenberger, E. G., Huentelman, M. J., Gottlieb, S., Dobrin, S. E., Parod, J. M., et al. (2006). Recessive symptomatic focal epilepsy and mutant contactin-associated protein-like 2. N. Engl. J. Med. 354, 1370–1377. doi: 10.1056/NEJMoa052773
Svobodová Burianová, J., and Syka, J. (2020). Postnatal exposure to an acoustically enriched environment alters the morphology of neurons in the adult rat auditory system. Brain Struct. Funct. 225, 1979–1995. doi: 10.1007/s00429-020-02104-8
Truong, D. T., Rendall, A. R., Castelluccio, B. C., Eigsti, I.-M., and Fitch, R. H. (2015). Auditory processing and morphological anomalies in medial geniculate nucleus of Cntnap2 mutant mice. Behav. Neurosci. 129, 731–743. doi: 10.1037/bne0000096
Varea, O., Martin-de-Saavedra, M. D., Kopeikina, K. J., Schürmann, B., Fleming, H. J., Fawcett-Patel, J. M., et al. (2015). Synaptic abnormalities and cytoplasmic glutamate receptor aggregates in contactin associated protein-like 2/Caspr2 knockout neurons. Proc. Natl. Acad. Sci. USA 112, 6176–6181. doi: 10.1073/pnas.1423205112
Vorhees, C. V., Amos-Kroohs, R. M., and Williams, M. T. (2024). Long-term effects of Preweaning environmental impoverishment on neurobehavioral and neurocognitive outcomes in Sprague Dawley rats: an early environmental stress model. Neurotoxicol. Teratol. 103:107356. doi: 10.1016/j.ntt.2024.107356
Wang, Y., Guo, X., Hong, X., Wang, G., Pearson, C., Zuckerman, B., et al. (2022). Association of mitochondrial DNA content, heteroplasmies and inter-generational transmission with autism. Nat. Commun. 13:3790. doi: 10.1038/s41467-022-30805-7
Weible, A. P., Moore, A. K., Liu, C., DeBlander, L., Wu, H., Kentros, C., et al. (2014). Perceptual gap detection is mediated by gap termination responses in auditory cortex. Curr. Biol. 24, 1447–1455. doi: 10.1016/j.cub.2014.05.031
Wobbrock, J. O., Findlater, L., Gergle, D., and Higgins, J. J. (2011). “The aligned rank transform for nonparametric factorial analyses using only anova procedures.” in Proceedings of the SIGCHI Conference on Human Factors in Computing Systems, (Vancouver BC Canada: ACM), 143–146.
Wright, C. G., and Barnes, C. D. (1972). Audio-spinal reflex responses in decerebrate and chloralose anesthetized cats. Brain Res. 36, 307–331. doi: 10.1016/0006-8993(72)90737-8
Zheng, A., Mann, R. S., Solaja, D., Allman, B. L., and Schmid, S. (2024). Properties of the caudal pontine reticular nucleus neurons determine the acoustic startle response in Cntnap2 KO rats. J. Integr. Neurosci. 23:63. doi: 10.31083/j.jin2303063
Zheng, A., and Schmid, S. (2023). A review of the neural basis underlying the acoustic startle response with a focus on recent developments in mammals. Neurosci. Biobehav. Rev. 148:105129. doi: 10.1016/j.neubiorev.2023.105129
Zheng, A., Scott, K. E., Schormans, A. L., Mann, R., Allman, B. L., and Schmid, S. (2023). Differences in startle and Prepulse inhibition in Contactin-associated protein-like 2 Knock-out rats are associated with sex-specific alterations in brainstem neural activity. Neuroscience 513, 96–110. doi: 10.1016/j.neuroscience.2023.01.020
Keywords: CNTNAP2, auditory processing, startle, prepulse inhibition, gap detection, rat, auditory brainstem response, neurodevelopmental disorders
Citation: Doornaert EE, Mohamad AE-C, Johal G, Allman BL, Möhrle D and Schmid S (2025) Postnatal environment affects auditory development and sensorimotor gating in a rat model for autism spectrum disorder. Front. Neurosci. 19:1565919. doi: 10.3389/fnins.2025.1565919
Edited by:
Ioanna Zioga, Radboud University, NetherlandsReviewed by:
Binliang Tang, Zhejiang University, ChinaYuko Tamaoki, New York University, United States
Copyright © 2025 Doornaert, Mohamad, Johal, Allman, Möhrle and Schmid. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Susanne Schmid, U3VzYW5uZS5zY2htaWRAc2NodWxpY2gudXdvLmNh
†These authors share senior authorship