
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
SYSTEMATIC REVIEW article
Front. Neurosci. , 07 March 2024
Sec. Gut-Brain Axis
Volume 18 - 2024 | https://doi.org/10.3389/fnins.2024.1341656
This article is part of the Research Topic Insights in Gut-Brain Axis: 2023 View all 5 articles
 Valentina Caputi1*†
Valentina Caputi1*† Lee Hill2,3†
Lee Hill2,3† Melanie Figueiredo2,4
Melanie Figueiredo2,4 Jelena Popov2,5,6
Jelena Popov2,5,6 Emily Hartung2,7Kara Gross Margolis8,9,10Kanish Baskaran2,4Papiha Joharapurkar2Michal Moshkovich2,4
Emily Hartung2,7Kara Gross Margolis8,9,10Kanish Baskaran2,4Papiha Joharapurkar2Michal Moshkovich2,4 Nikhil Pai2,11,12,13
Nikhil Pai2,11,12,13Introduction: Critical phases of neurodevelopment and gut microbiota diversification occur in early life and both processes are impacted by genetic and environmental factors. Recent studies have shown the presence of gut microbiota alterations in neurodevelopmental disorders. Here we performed a systematic review of alterations of the intestinal microbiota composition and function in pediatric and adult patients affected by autism spectrum disorder (ASD), attention-deficit/hyperactivity disorder (ADHD), and Rett syndrome (RETT).
Methods: We searched selected keywords in the online databases of PubMed, Cochrane, and OVID (January 1980 to December 2021) with secondary review of references of eligible articles. Two reviewers independently performed critical appraisals on the included articles using the Critical Appraisal Skills Program for each study design.
Results: Our systematic review identified 18, 7, and 3 original articles describing intestinal microbiota profiles in ASD, ADHD, and RETT, respectively. Decreased Firmicutes and increased Bacteroidetes were observed in the gut microbiota of individuals affected by ASD and ADHD. Proinflammatory cytokines, short-chain fatty acids and neurotransmitter levels were altered in ASD and RETT. Constipation and visceral pain were related to changes in the gut microbiota in patients affected by ASD and RETT. Hyperactivity and impulsivity were negatively correlated with Faecalibacterium (phylum Firmicutes) and positively correlated with Bacteroides sp. (phylum Bacteroidetes) in ADHD subjects. Five studies explored microbiota-or diet-targeted interventions in ASD and ADHD. Probiotic treatments with Lactobacillus sp. and fecal microbiota transplantation from healthy donors reduced constipation and ameliorated ASD symptoms in affected children. Perinatal administration of Lactobacillus sp. prevented the onset of Asperger and ADHD symptoms in adolescence. Micronutrient supplementation improved disease symptomatology in ADHD without causing significant changes in microbiota communities’ composition.
Discussion: Several discrepancies were found among the included studies, primarily due to sample size, variations in dietary practices, and a high prevalence of functional gastrointestinal symptoms. Further studies employing longitudinal study designs, larger sample sizes and multi-omics technologies are warranted to identify the functional contribution of the intestinal microbiota in developmental trajectories of the human brain and neurobehavior.
Systematic review registration: https://clinicaltrials.gov/, CRD42020158734.
Over the last decade, the field of neuroscience has benefited from the rapid evolution of microbiology and microbiome science. This includes new insights into the pathophysiology of neurodevelopmental disorders. The Developmental Origins of Health and Disease (DOHaD) hypothesis proposes that early life influences can dramatically impact the health of infants. A combination of genetic predisposition and environmental triggers may have deleterious effects on infants’ neurodevelopmental programming, predisposing individuals to long-lasting behavioral and cognitive alterations. This has been described in children with conditions such as autism spectrum disorders (ASD) (Santos et al., 2022), attention deficit hyperactivity disorder (ADHD) (Hare et al., 2022; Makris et al., 2022) and Rett syndrome (RETT) (Hoffbuhr et al., 2002).
ASD comprises a spectrum of neurodevelopmental disorders characterized by deficits in social interactions and communication skills. Affected individuals are often male and may present with unusual repetitive behaviors, restricted interests, difficulties with communication and marked impairments in daily functioning. Personal, social, school, and work life are often affected (Harris, 2018). The spectrum of ASD includes autism (AD), Asperger’s Syndrome (AS) and, prior to the 2013 edition of Diagnostic and Statistical Manual of Mental Disorders (DSM) (American Psychiatric Association, 2013), Pervasive Developmental Disorder Not Otherwise Specified (PDD-NOS) (Haglund and Källén, 2011; Tateno et al., 2011).
ADHD is a frequently diagnosed condition in children and adults. ADHD affects approximately 7.2% of children worldwide and up to 3-10% of school-aged children (Lange et al., 2010; Polanczyk et al., 2014; Thomas et al., 2015). The underlying pathophysiology of this neurodevelopmental disorder involves disturbances in neural responses to dopamine and noradrenaline stimulation and deficits in reward processing via the ventral striatum. Genome-wide association studies have identified pathognomonic abnormalities in serotonin, dopamine, and noradrenaline-related genes (Guimarães et al., 2007; Ghosh et al., 2013; Abraham et al., 2020).
RETT is a rare genetic disorder, principally affecting females, caused by loss-of-function mutations of the X-linked methyl-CpG binding protein 2 (MeCP2) gene (Borghi and Vignoli, 2019). Individuals with RETT are characterized by loss of purposeful hand use and spoken communication skills, as well as the development of stereotypic hand movements and gait abnormalities after a period of apparently normal development. RETT shares several common features with ASD, yet clear differences exist between the two diseases (Percy, 2011).
Patients with ASD, ADHD and RETT have a high prevalence of functional gastrointestinal disorders (FGID) (Adams et al., 2011; Motil et al., 2012; Severance et al., 2015; Bermudez et al., 2019; Kedem et al., 2020), now referred as disorders of gut brain interaction (DGBI) (Camilleri, 2023; for the purposes of consistency with literature assessed in our systematic review, we will continue to use FGDI) and stereotypic dietary habits (Wong et al., 2021; Bourne et al., 2022; Salvat et al., 2022). These observations have led to increased research toward identifying differences in the gastrointestinal (GI) system across these neurodevelopmental disorders with a particular focus on the microbiota. Gut microbes and their collective genome (the microbiome) may be considered an essential organ for maintaining the homeostasis of intestinal and extraintestinal body sites throughout the lifespan (Cryan et al., 2020). This complex microbial community has been recognized to influence brain neurophysiology and behavior by communicating with the central nervous system (CNS) through multiple neuronal, immune, and endocrine signaling pathways involved in the microbiota-gut-brain axis (MGBA) (Luczynski et al., 2016; Cryan et al., 2019). In response, the brain establishes appropriate physiological responses to direct mental and physical functions (Fung et al., 2017; Cryan et al., 2019; Bioque et al., 2021).
Human perinatal and early neurodevelopment are characterized by sensitive periods or “critical windows” in which the biological systems involved in brain development (i.e., neurogenesis, neuronal differentiation and migration, synaptic rearrangement) show significant plasticity. These periods are uniquely susceptible and responsive to environmental stimuli (i.e., hypoxia, trauma, infection or inflammation; Cowan et al., 2020; Lopatina et al., 2021). The gut microbiota changes and evolves rapidly during these same temporal windows, and this parallels numerous physiologic trajectories (immunity, metabolism, stress responsivity, and neuroglial formation) within the MGBA (Borre et al., 2014; Cowan et al., 2020; Bioque et al., 2021). Thus, these changes may have long-lasting consequences for brain function, behavior and psychiatric well-being (Cryan et al., 2019; Margolis et al., 2021).
The central nervous system’s maturation begins in the first trimester of gestation with the formation of the neural tube, which becomes organized into the brain and spinal cord. Concurrent development of the blood brain barrier (BBB) begins at gestational week eight and continues throughout pregnancy. In the third trimester of pregnancy, the human brain doubles in size and oligodendrocytes proliferate to start the process of myelination (Borre et al., 2014). Critical events for normal brain development, such as hippocampal neurogenesis and gliogenesis, occur during these early gestation phases and continue through the postnatal period up to 3 years of age (Borre et al., 2014; Lopatina et al., 2021). The BBB, while functionally active by week 14 of gestation, continues to undergo maturation into an individuals’ third decade (Saili et al., 2017).
The assembly of gut microbial communities in early life then occurs as a dynamic process that is significantly influenced by diet as babies shift from exclusive breastmilk or formula intake to complementary solid foods (Ratsika et al., 2021). By the age of 3 years, the microbiota composition reaches a more stable, adult-like phenotype (Tanaka and Nakayama, 2017).
Emerging evidence shows that host-microbiota crosstalk through the immune system is fundamental to preserving the integrity of the intestinal and peripheral epithelial-immune-endothelial barriers (such as the BBB). Within the central nervous system, bacterial microbes help regulate the development of the microglia and other resident immune cells which help sustain and protect neuronal function (Fung et al., 2017). Intestinal microbes can produce a variety of neurotransmitters, as well as microbial-derived neuroactive compounds such as short-chain fatty acids (SCFA) that modulate the peripheral and central nervous system, and immune system (Fung et al., 2017; Erny et al., 2021; Popov et al., 2021).
Disturbances of the microbiota-immune axis in early life affect intestinal barrier permeability, leading to systemic translocation of bacterial-derived products that may cross the BBB. The infant BBB is still immature until at least 4 months of age, leading to microglia hyperactivation within the CNS and neuroinflammation (Hill et al., 2021). In the setting of intestinal inflammation and increased intestinal permeability, the translocation of bacterial products into the systemic circulation further increases. Ordinarily, endothelial structures outside the BBB (e.g., choroid plexus) block the passage of substrates from the periphery to the brain, thus protecting the CNS from the progressive increase of pro-inflammatory factors (Carloni et al., 1979). This response compromises the bidirectional flow of key signaling molecules and hormones between the gut-brain components of the MGBA (Carloni et al., 1979). Pathogens that have molecular mimicry with host antigens can exert effects by causing immunological cross-reactions and autoimmunity. These have been associated with both neuroinflammation and behavioral affective disorders (Hill et al., 2021).
Causal relationships between changes in the microbiome and behavioral outcomes remain unclear (Laue et al., 2021; Yap et al., 2021). Studies of the microbiome in pediatric neurodevelopmental disorders face persistent questions of temporal dynamics of behavioral symptoms, and the independent impacts of key variables known to alter microbiota composition, including dietary intake, GI comorbidities, and/or sex differences (Baikie et al., 2014; Laue et al., 2021; Yap et al., 2021; Tarui et al., 2022). Nevertheless, recent advances in sequencing technologies and bioinformatics are facilitating the concomitant examination of subtle microbiome changes with behavioral outcomes in animal models or human subjects affected by pediatric neurodevelopmental disorders (Berg et al., 2020; Morais et al., 2021; Socała et al., 2021).
In this systematic review, we provide a comprehensive overview of research on the diverse presence and potential roles of the gut microbiome in ASD, ADHD and RETT. Our protocol identified case–control and cohort studies investigating the intestinal microbiome in pediatric patients affected by ASD, and pediatric and adult patients’ affected by ADHD (Hill et al., 2021) and Rett syndrome. We also summarize other critical factors that play essential roles in brain-gut crosstalk including molecular metabolites, immunologic markers, and neurodevelopmental and GI symptoms. We discuss potential changes in host-microbe immune and endocrine interactions, alterations in microbial-derived metabolite production and inflammatory and other biochemical pathways associated with these relevant disorders. Finally, we explore microbiota-targeted interventions that have been shown to improve symptoms.
We have previously published our protocol for this systematic review, consistent with the Preferred Reporting Items for Systematic review and Meta-analysis (PRISMA) protocols checklist (Liberati et al., 2009; Shamseer et al., 2015). The systematic review protocol (Hill et al., 2021) is registered on PROSPERO (ID: CRD42020158734).
We appraised available evidence surrounding the gut microbiota in ASD, ADHD, and RETT (Hill et al., 2021). Relevant studies were identified using PubMed, Cochrane Library, and OVID electronic databases. Articles published between January 1980 and December 2021 were considered for inclusion and analysis.
We implemented a rigorous search strategy that involved two rounds of literature review, using a three-step method in each round (Hill et al., 2021). The first round involved a review of articles from the database search results; the second round involved a review of works cited by those articles identified in the first round. Titles, abstracts, and full texts were independently screened by at least two authors (LH, JP, MF, VC, EH, MM, or NP). Articles were excluded if they were unrelated or met exclusion criteria (Supplementary Table S1). Inclusion/exclusion criteria were slightly modified for ADHD and RETT studies. The age of the patients was >18 for three of the seven included ADHD studies. In all included studies on RETT, the age range was between 5 and 36 years. We decided to include these studies as they met other inclusion criteria, and they were the only available studies that included human participants (Supplementary Table S1). Two reviewers (MF and LH) completed data extraction using a pre-piloted Microsoft Excel data extraction form. Data extraction was subsequently checked for accuracy and completeness by members of the study team (NP, JP, EH, MM, and VC). One further study was included upon consultation with external content experts (KM and EA) (Luna et al., 2017).
We assessed quality of the included studies using the appropriate Critical Appraisal Skills Program (CASP) checklist based on study methodology. CASP was selected for its quality of appraisal of health-related studies, assessment of transparency of research practice and reporting standards (Long et al., 2020). Level of evidence was assessed using established methods and a “level of evidence score” was established (Obremskey et al., 2005). Two independent reviewers assessed each study and, if discrepancies occurred, a third independent reviewer was included. We applied a semi-quantitative appraisal of CASP checklists based on an approach by Nichols et al. (2020) where study quality was scored out of 20.
The initial search strategy outlined in our protocol (Hill et al., 2021) identified 166 articles. After initial screening and removal of duplicates, 131 articles were screened for inclusion. Finally, after review with external content experts 33 original studies that described the association between intestinal microbiota and ASD, ADHD or RETT in children and adults were included (Figure 1). Twenty-six studies were assessed using the CASP Checklist for Case–Control Studies (Supplementary Table S2), five using the CASP Checklist for Cohort Studies (Supplementary Table S3), and two for RCT studies (Supplementary Table S4).

Figure 1. PRISMA-ScR systematic flowchart of the systematic review process.
Scores for each study ranged from 7 (low methodological quality) to 20 (high methodological quality). Two studies (6.9%) had a total score below 10, suggesting weak methodology (Zhang et al., 2018; Ahmed et al., 2020). The majority of studies (75.9%) had a total score between 15 and 20, and eight studies achieved the highest score for methodological quality (De Angelis et al., 2013; Pärtty et al., 2015; Tomova et al., 2015; Strati et al., 2016, 2017; Kang et al., 2018; Zhai et al., 2019; Wan et al., 2020).
Eighteen case–control studies assessed the intestinal microbiota in ASD, seven in ADHD (four cohort studies, three case–control), and three in RETT (one cohort, two case–control; Tables 1–4; Supplementary Tables S5–S8). Geographically, the studies included patients from different countries including the United States, China, India, Australia, Egypt, Germany, Italy, and The Netherlands. The studies included in our review encompassed a total of 2,251 subjects with ASD (among which 10 subjects were diagnosed with PDD-NOS; Supplementary Tables S5, S6), 272 subjects with ADHD (Supplementary Table S7) and 101 subjects with RETT (Supplementary Table S8). Control groups included 1,059 sex- and age- matched neurotypical (NT) subjects in studies of ASD and PDD-NOS, 321 NT subjects in studies of ADHD, and 60 NT subjects in studies of RETT (Supplementary Tables S5–S8). Ages of cases and controls spanned 2 to 18 years old for studies of ASD (Tables 1, 2; Supplementary Tables S5, S6), 6 to 44 years for studies of ADHD (Table 3; Supplementary Table S7), and 5 to 36 years for studies of RETT (Table 4; Supplementary Table S8). Subjects with ASD, PDD-NOS, ADHD and respective NT controls were males and females, while subjects with RETT and their NT controls were all females.
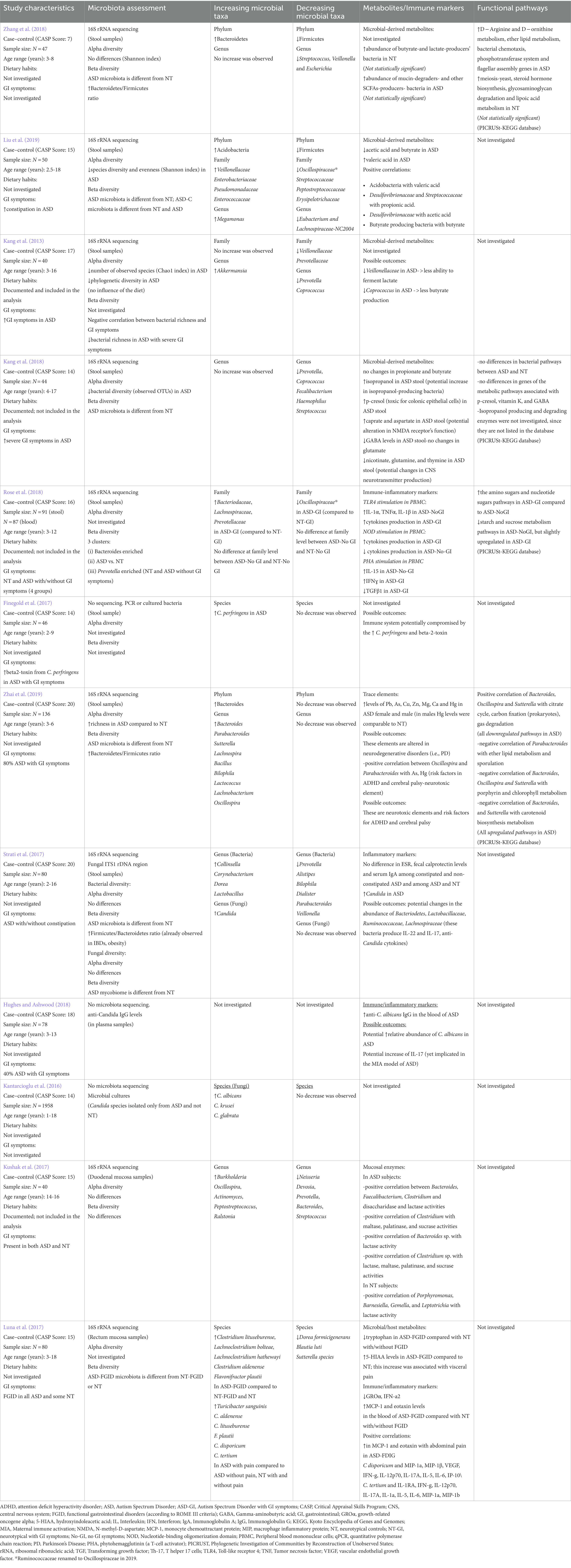
Table 1. Evidence of microbiota alterations in ASD compared to unrelated NT controls.
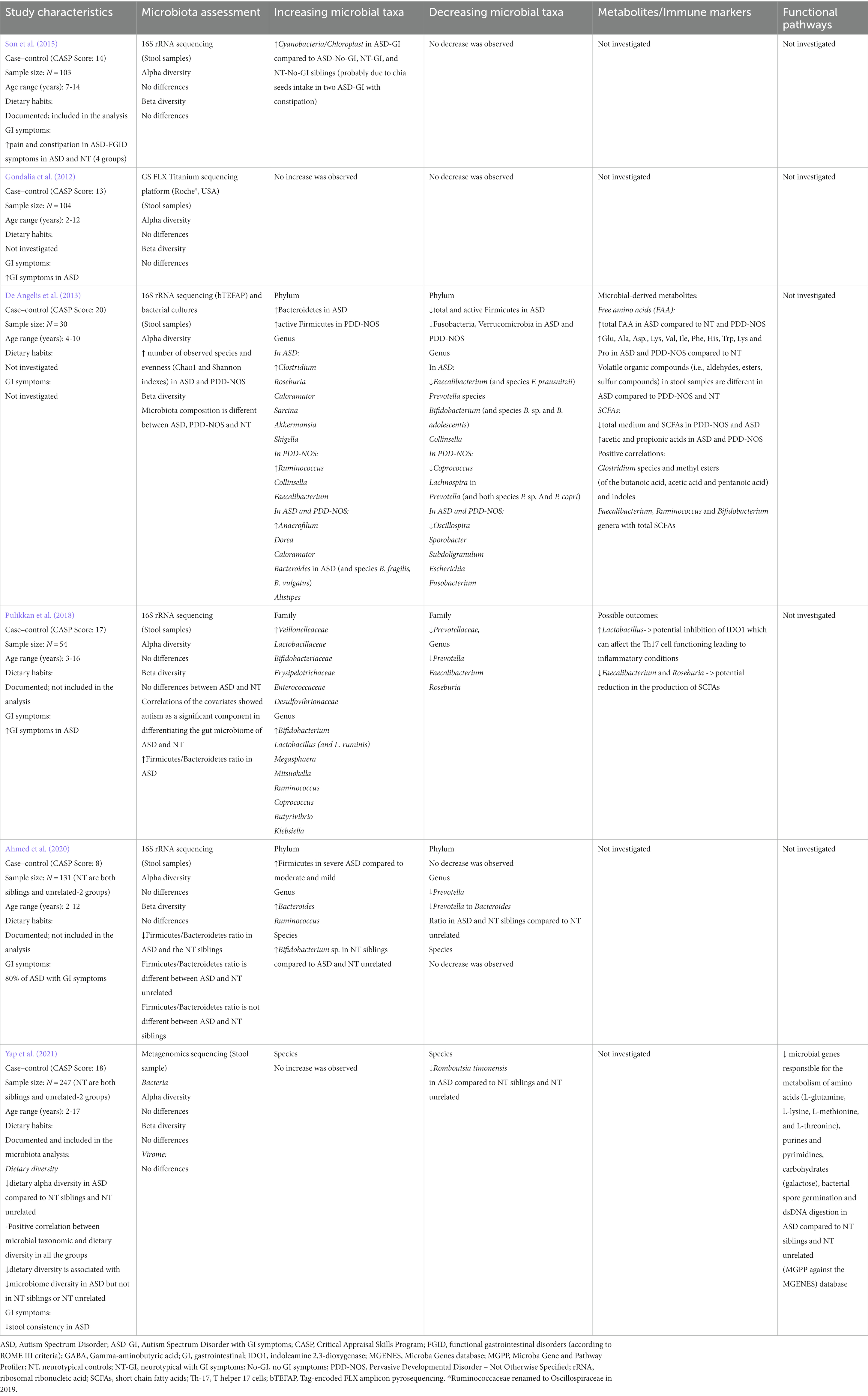
Table 2. Evidence of microbiota alterations in ASD compared to NT siblings.

Table 3. Evidence of microbiota alterations in ADHD.
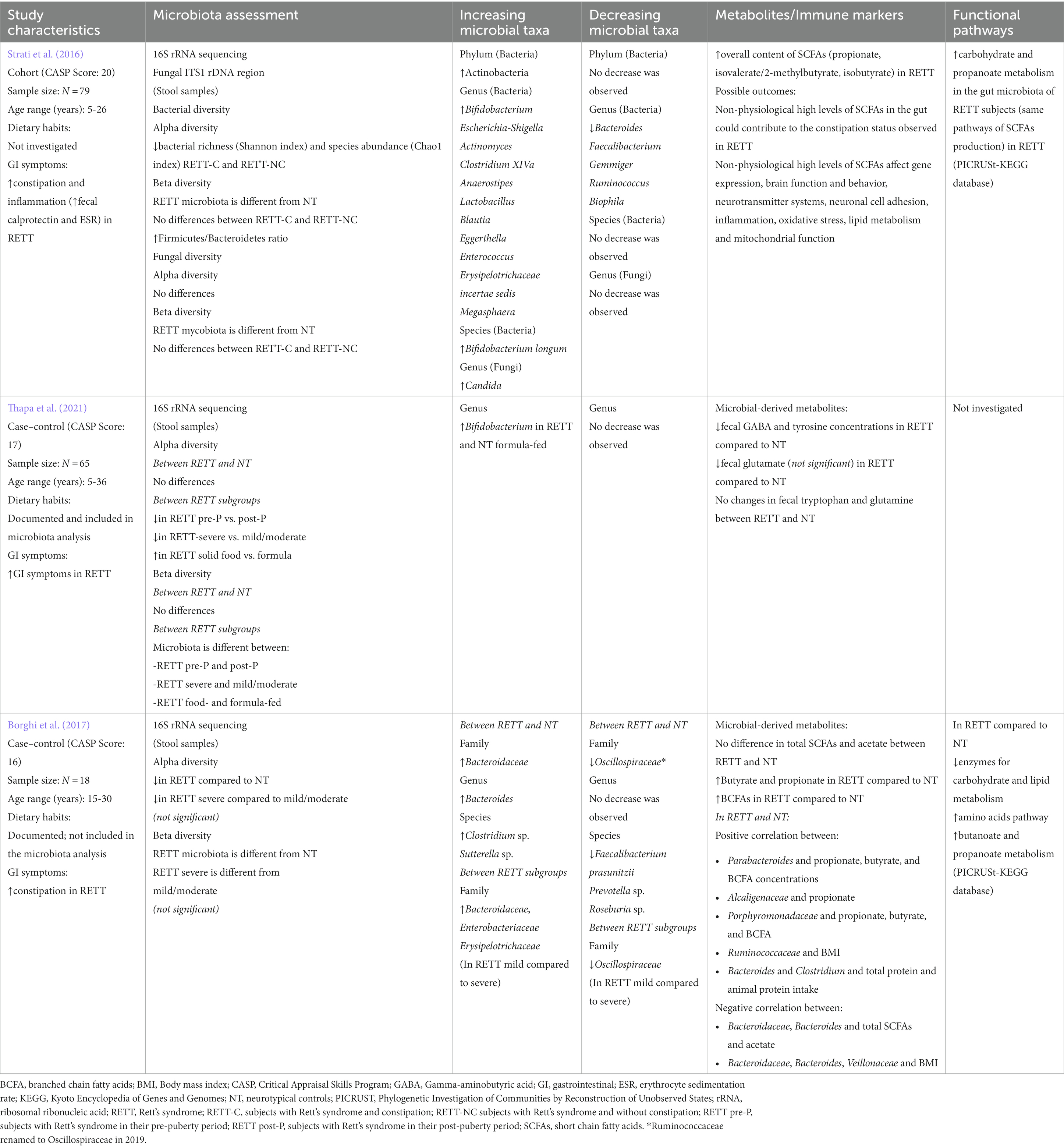
Table 4. Evidence of microbiota alterations in Rett syndrome.
Studies examining the effects of microbiota- or diet-targeted interventions on behavioral outcomes and microbiota composition in subjects with neurodevelopmental disorders are summarized (Table 5; Supplementary Table S9). Two studies assessed the effect of probiotic administration (Tomova et al., 2015; Shaaban et al., 2018), and one study explored the effects of FMT (Kang et al., 2017) on gut microbiota, GI symptoms, and neurodevelopmental outcomes in individuals with ASD. One study assessed the effect of dietary micronutrient supplementation on gut microbiota composition and behavioral symptomatology in subjects with ADHD (Stevens et al., 2019). One study assessed the effect of perinatal administration of probiotic Lactobacillus rhamnosus GG in preventing the onset of ASD or ADHD in childhood (Pärtty et al., 2015). The characteristics of subjects included in the intervention trials are summarized (Supplementary Table S9).
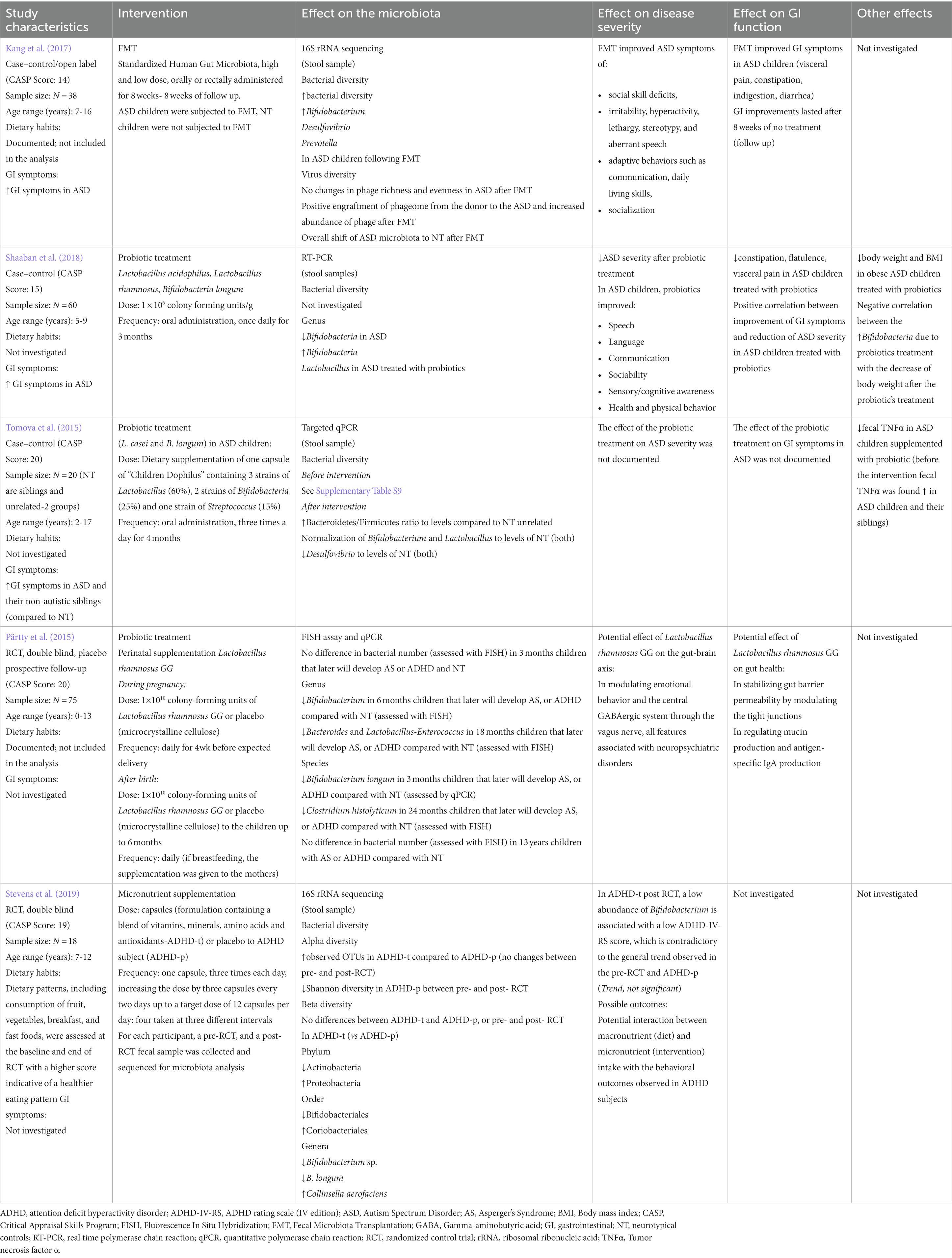
Table 5. Intestinal microbiota- or diet-targeted interventions in neurodevelopmental disorders.
Samples used in the selected studies were derived from stool (31 studies), rectal tissue (1 study), and duodenal tissue (1 study). Samples from seven studies were stored at 4°C for ≤24 h before being frozen for storage (Pärtty et al., 2015; Kantarcioglu et al., 2016; Aarts et al., 2017; Prehn-Kristensen et al., 2018; Stevens et al., 2019; Wang et al., 2020; Szopinska-Tokov et al., 2021). All other studies’ samples were frozen immediately upon collection. Samples from five studies were stored with preservation agents at the time of collection (De Angelis et al., 2013; Son et al., 2015; Rose et al., 2018; Richarte et al., 2021; Thapa et al., 2021). Further details about DNA extraction, sequencing technique, reference database and sequence corrections are described (Supplementary Table S10).
Measurements of neurodevelopmental outcomes in ASD, ADHD, and RETT are summarized in Supplementary Tables S5–S9.
Ten studies compared stool-associated bacterial communities of ASD subjects to NT controls (with no first-degree family relationships to ASD subjects, Table 1), while four studies used NT first-degree siblings of ASD subjects, and two studies included both NT first-degree siblings of ASD subjects and unrelated NT controls (Table 2). Bacterial α-diversity indices were evaluated in five of the 10 studies in which NT controls were unrelated to ASD individuals. Four studies reported reduced, and one study reported increased bacterial α-diversity of ASD-associated microbiota (Table 1). Six of the 10 studies, in which NT controls were unrelated to ASD individuals, evaluated β-diversity indices and showed differences in gut bacterial profiles between ASD subjects and NT controls (Table 1).
Conflicting α- and β-diversity outcomes occurred in three of the four studies in which NT controls were siblings of ASD children as no differences in bacterial α-diversity or composition (β-diversity) were found between groups in these studies (Table 2). No differences in α- or β-diversity were again reported by Yap et al. (2021) and Ahmed et al. (2020), who compared bacterial α- and β-diversities of ASD subjects with both NT siblings and unrelated NT controls. De Angelis et al. (2013) reported significantly increased α-diversity in both ASD and PDD-NOS subjects, and differences in relative abundance of bacterial communities between subjects with ASD, PDD-NOS, and their NT first-degree siblings (Table 2).
The two studies evaluating mucosal biopsy-associated microbiota included only unrelated NT controls (Table 1). Kushak et al. (2017) did not report any differences in bacterial diversity of duodenal mucosa-associated microbiota between ASD subjects and NT controls (Kushak et al., 2017). Luna et al. (2017) reported differences in β-diversity between rectal mucosa-associated microbiota of ASD and NT controls (Luna et al., 2017).
Altered bacterial diversity in ASD children compared to unrelated NT controls might suggest the presence of an ASD-related gut microbiota phenotype (Brito et al., 2019). However further studies with larger sample sizes and with both siblings and family unrelated NT controls are warranted to determine whether children with ASD have an altered microbiota due to the disease.
Multiple studies reported distinctive alterations in bacterial taxa among subjects with ASD; however, these results were not consistent. A reduction in the relative abundance of the genus Prevotella in ASD was reported across five studies assessing stool-associated microbiota (two studies included NT siblings), and in the study by Kushak et al. (2017), which investigated duodenal mucosa-associated microbiota (Tables 1, 2). The results from bacterial cultures in the study by De Angelis et al. (2013), reported increased relative abundance of Prevotella in ASD compared to NT subjects (Supplementary Table S6). Faecalibacterium and Streptococcus levels were decreased in ASD subjects in three (De Angelis et al., 2013; Kang et al., 2018; Pulikkan et al., 2018) and four studies (De Angelis et al., 2013; Kushak et al., 2017; Kang et al., 2018; Zhang et al., 2018), respectively (Tables 1, 2). Bacterial culture, 16S rDNA sequencing, and targeted PCR revealed higher levels of genus Clostridium in stool samples of ASD patients in the studies by De Angelis et al. (2013) (Table 2; Supplementary Table S6) and Tomova et al. (2015) (Table 5; Supplementary Table S9). In ASD-associated microbiota, Lactobacillus was increased in three studies (Tomova et al., 2015; Strati et al., 2017; Pulikkan et al., 2018) and decreased in De Angelis et al. (2013) (Tables 1, 2, 5). Three studies reported increased abundance of genus Bacteroides in ASD stool samples (De Angelis et al., 2013; Zhai et al., 2019; Ahmed et al., 2020), while Kushak et al. (2017) showed decreased abundance of Bacteroides in duodenal mucosa-associated microbiota from ASD subjects (Tables 1, 2).
At the species level, increased Bacteroides fragilis and B. vulgatus (De Angelis et al., 2013), Clostridium sp. (De Angelis et al., 2013; Finegold et al., 2017)., Lactobacillus sp. (Tomova et al., 2015). and reduced Faecalibacterium prausnitzii (De Angelis et al., 2013), Romboutsia timonensis (Yap et al., 2021), Prevotella sp. (De Angelis et al., 2013) were detected in ASD-associated microbiota (Tables 1, 2). Conflicting results were reported for Bifidobacterium abundance in ASD subjects compared to NT first-degree siblings. Decreased and increased abundance of the genus Bifidobacterium in ASD subjects compared to NT siblings were reported by De Angelis et al. (2013) and Pulikkan et al. (2018), respectively (Table 2). At the species level, reduced B. adolescentis, and Bifidobacterium sp. were reported by De Angelis et al. (2013) and Ahmed et al. (2020) (Table 2), whereas Tomova et al. (2015) observed an increased abundance of Bifidobacterium sp. in their cohort of ASD children (Table 5; Supplementary Table S9). In all studies there was no difference in the abundance of Bifidobacterium between children with ASD and unrelated NT subjects.
In mucosa-associated microbiota from duodenal biopsies of subjects with ASD, significant alterations occurred at the genus level, with reduced abundance of Prevotella, Bacteroides, Streptococcus, Neisseria, and increased abundance of Devosia, Oscillospira, Actinomyces, and Peptostreptococcus. These findings recapitulate some of the changes observed in the stool-associated microbiota from ASD subjects (Kushak et al., 2017). Positive correlations between Bacteroides, Faecalibacterium, and Clostridium and the activity of duodenal enzymes such as maltase, lactase and sucrase were also observed in ASD (Kushak et al., 2017; Table 1).
The study of Luna et al. (2017) reported increased abundance of bacterial species from Clostridiales in rectal mucosa-associated microbiota from ASD subjects. In particular, bacterial species Clostridium lituseburense, Lachnoclostridium bolteae, Lachnoclostridium hathewayi, Clostridium aldenense, and Flavonifractor plautii (Table 1), were significantly more abundant in ASD microbiota compared to NT controls. This study also observed a decrease in Dorea formicigenerans and Blautia luti (Table 1) as well as reduced Sutterella species (Table 1) in ASD-associated microbiota. Together these data showed major perturbations in the abundance of bacterial genera and species belonging to the phyla Firmicutes and Bacteroidetes in ASD-associated microbiota.
Few studies assessed changes in inferred bacterial function using 16S profiles and Phylogenetic Investigation of Communities by Reconstruction of Unobserved States (PICRUSt) to predict the presence of Kyoto Encyclopedia of Genes and Genomes (KEGG) orthologs across functional and metabolic pathways (Box A1; Kanehisa, 2000). Zhang et al. (2018) observed significant enrichments of several bacterial genes associated with D − Arginine and D − ornithine metabolism, ether lipid metabolism, bacterial chemotaxis, phosphotransferase system (PTS) and flagellar assembly genes in ASD subjects. Steroid hormone biosynthesis, glycosaminoglycan degradation and lipoic acid metabolism were more abundant in NT controls (Zhang et al., 2018; Table 1). Conversely, Kang et al. (2018) assessed genes in the metabolic pathways associated with p-cresol, vitamin K, and GABA but did not find any significant differences between ASD subjects and NT controls (Table 1). The study by Rose et al. (2018) differentiated NT or ASD subjects with GI symptoms (NT-GI and ASD-GI, respectively), compared with NT or ASD without GI symptoms (NT-no-GI and ASD-no-GI, respectively; Supplementary Table S5). Amino sugars and nucleotide sugars pathways related to the gut microbiota were significantly upregulated in ASD-GI compared to ASD-no-GI subjects. Starch and sucrose metabolism pathways were downregulated in ASD-no-GI subjects compared to ASD-GI subjects (Rose et al., 2018; Table 1). Zhai et al. (2019) described 40 metabolic pathways found to be statistically significantly different between subjects with ASD and NT controls. Bacteroides, Oscillospira and Sutterella were positively correlated with carbon fixation pathways in prokaryotes, the citrate cycle, and gas degradation (pathways downregulated in ASD), and negatively correlated with porphyrin and chlorophyll metabolism (pathways upregulated in ASD). Ether lipid metabolism and sporulation were upregulated in ASD and negatively associated with Parabacteroides. A negative correlation was found between Bacteroides and Sutterella and carotenoid biosynthesis metabolism (Table 1). Yap et al. (2021) assessed the potential functional profiling of ASD-associated microbiota by using the Microba Gene and Pathway Profiler (MGPP) tool against the Microba Genes (MGENES) database. The study assessed metabolic pathways associated with Romboutsia timonensis, as this species was found to be less abundant in ASD subjects. The predictive analysis showed reduced abundance of microbial genes responsible for the metabolism of amino acids (L-glutamine, L-lysine, L-methionine, and L-threonine), purines and pyrimidines, carbohydrates (galactose), bacterial spore germination and dsDNA digestion in ASD subjects compared to NT siblings and unrelated NT controls (Table 2). These results suggest potential alterations in the transcription of microbial genes associated with ASD. Further analyses (e.g., metatranscriptomics) will be required to evaluate protein expression associated with these microbial differences.
Two studies investigated possible correlations between intestinal microbiota changes and ASD severity. Ahmed et al. (2020) assessed ASD subjects with mild–moderate (n = 35) and severe ASD (n = 6). No statistically significant differences were found across most taxa, except for Firmicutes taxa which were significantly higher in patients with severe ASD (Figure 2; Table 2). This was also described by Tomova et al. (2015), who found a lower Bacteroidetes to Firmicutes ratio in children with severe ASD, compared to children with milder ASD and NT controls, respectively (Supplementary Table S9). This study also found children with severe ASD had higher Desulfovibrio and Clostridia compared to children with a milder ASD phenotype (Tomova et al., 2015) (Supplementary Table S9). A strong correlation between Desulfovibrio and the ADI restricted/repetitive behavior subscale score (a measure of ASD severity) was reported by Tomova et al. (2015) (Figure 2; Supplementary Table S9). These studies suggest a possible association between the phylum Firmicutes and ASD severity.

Figure 2. Microbiota-gut-brain axis alterations in ASD, ADHD, and Rett syndrome. Schematic representation of the main findings emerged from the studies included in our systematic review, portraited as alterations across the microbiota-gut-brain axis in ASD (left panel), ADHD (middle panel), and Rett’s syndrome (RETT; right panel). Changes in the intestinal microbiota of subjects with ASD are associated with a high prevalence of GI symptoms and low-quality diet. The family household might have an influence in shaping gut microbiota communities, as ASD and family unrelated NT subjects have a different microbiota, however no differences are observed when ASD subjects are compared with their siblings. RETT subjects present intestinal inflammation (indicated by increased ESR and fecal calprotectin) and constipation and there are changes in the gut microbiota depending upon the type of diet (solid food vs. formula). No information is provided on the presence of GI disorders in ADHD subjects, however differences in dietary habits between ADHD subjects and NT controls have been shown even though their impact on the gut microbiota has not been investigated. Intestinal dysbiosis is mainly characterized by decreased Faecalibacterium (phylum Firmicutes) and Prevotella (phylum Bacteroides) in all three disorders. Increased Bacteroides sp. (phylum Bacteroidetes) were identified in ASD and ADHD. Lactobacillus (phylum Firmicutes) was increased in ASD and RETT. Bifidobacterium (phylum Actinobacteria) increased in ADHD and RETT and decreased in ASD. Desulfovibrio (phylum Proteobacteria) increased in ASD and decreased in RETT. Escherichia (phylum Proteobacteria) increased in RETT and decreased in ASD. Increased abundances of Candida sp. were identified whithin the mycobiota of ASD and RETT. Microbiota associated metabolites such as SCFAs and gut-brain neurotransmitters GABA, glutamine, tryptophan, and tyrosine are altered in ASD and RETT, however they have not been explored in ADHD studies. Increased p-cresol, isopropanol in the stool and trace elements As, Hg, Pb, Cu, Zn, Mg, Ca in the hair were found in ASD subjects. ASD is also associated with a dysregulation of the immune system (and potential inflammation), as blood cytokines IL-1α, TNFα, IL-1β, IL-15, IFNɣ, MCP-1, eotaxin are increased and GROα, IFN-a2, TGFβ1 are reduced. Conversely, no sign of inflammation was reported by studies investigating ADHD. Possible correlation between changes in microbial taxa and disease severity can be extrapolated across the three disorders. ASD severity is positively correlated with Desulfovibrio (phylum Proteobacteria). Hyperactivity and impulsivity (ADHD symptoms) are negatively correlated with Faecalibacterium (phylum Firmicutes) and positively correlated with Bacteroides sp. (phylum Bacteroidetes) in ADHD. A reduced alpha-diversity and increased beta diversity are associated with severe RETT (compared to mild/moderate). CDT was increased in the ADHD bacteriome. CDT is responsible of producing phenylalanine, an essential AA that crosses the BBB and acts as a precursor for dopamine and noradrenaline, key neurotransmitters involved in modulating behavior-related neuronal pathways in the brain. AA, amino acid; ADI-RRB, autism diagnostic interview-restricted and repetitive behaviors; ADHD, attention deficit hyperactivity disorder; ASD, Autism Spectrum Disorder; BBB, blood brain barrier; BCFAs, branched chain fatty acids; CDT, cyclohexadienyl dehydratase; GABA, Gamma-aminobutyric acid; GI, gastrointestinal; GROα, growth-related oncogene alpha; IL, Interleukin; IFN, Interferon; MCP-1, monocyte chemoattractant protein; NT, neurotypical controls; RETT, Rett syndrome; SCFAs, short chain fatty acids.
Three studies investigated alterations in fungi composition of stool samples from ASD subjects (Table 1). Strati et al. (2016) observed a significant difference in the composition of the gut mycobiome between subjects with ASD and unrelated NT controls with enrichment in relative abundance of the genus, Candida in ASD. The study by Kantarcioglu et al. (2016) also reported increased levels of Candida species in stool specimens from ASD subjects, and Hughes and Ashwood (2018) observed an increased concentration of anti-C. albicans immunoglobulin (IgG) in serum samples (Hughes and Ashwood, 2018; Figure 2). Further studies are required to assess the role of fungi in ASD.
Five studies did not find any significant differences in bacterial α- or β-diversities indices between ADHD subjects and NT controls, including those studies where the age of the subjects was >18 years (Aarts et al., 2017; Zhai et al., 2019; Szopinska-Tokov et al., 2020; Wan et al., 2020; Richarte et al., 2021; Szopinska-Tokov et al., 2021; Table 3). Prehn-Kristensen et al. (2018) observed significantly decreased richness and evenness in the gut microbiota of ADHD subjects and their mothers compared to unrelated NT controls. The same study observed significant differences in bacterial profiles between ADHD subjects and NT controls, and between ADHD subjects and their mothers (Prehn-Kristensen et al., 2018; Table 3). Wang et al. (2020) found changes in bacterial α-diversity in ADHD subjects and NT controls, with no significant differences in bacterial composition overall (Table 3). Only two studies evaluated first-degree NT siblings of ADHD subjects, however NT siblings were mixed with unrelated NT controls and data were analyzed as a single group (Aarts et al., 2017; Szopinska-Tokov et al., 2021). Diversity measures showed no differences in bacterial communities between ADHD subjects and NT controls. Changes in α-diversity observed in the studies by Prehn-Kristensen et al. (2018) and Wang et al. (2020) need further validation using larger cohorts.
The principal phyla characterizing the bacterial kingdom of the human microbiota, Firmicutes, Bacteroidetes, Actinobacteria, and Proteobacteria, were identified across all ADHD studies. One study found a significant increase of Actinobacteria, accompanied by a decrease of Firmicutes in ADHD gut microbiota (Aarts et al., 2017). Further taxonomic analyses showed that taxa from families and genera belonging to the phyla Firmicutes and Bacteroidetes were primarily associated with ADHD diagnosis (Figure 2; Table 3). At least two studies reported reduced abundance of bacteria from the family Oscillospiraceae (Aarts et al., 2017; Wan et al., 2020) (renamed from Ruminococcaceae (Tindall, 2019)), and the genera Faecalibacterium (Jiang et al., 2018; Wan et al., 2020) and Coprococcus (e.g., Coprococcus_2) (Aarts et al., 2017; Szopinska-Tokov et al., 2021) within the ADHD gut microbiota (Table 3). Within the phylum Bacteroidetes, the abundance of bacteria from the genera Bacteroides, Alistipes, Odoribacter, and Parabacteroides, as well as from the family Bacteroidaceae, were reported to be increased in ADHD subjects (Table 3). Increased abundance of Egghertella, Neisseria (phylum Proteobacteria), Bifidobacterium (phylum Actinobacteria), and Fusobacterium (phylum Fusobacteria) were also observed in the gut microbiota of ADHD subjects. At the species level, two studies reported increased amounts of Bacteroides uniformis and Bacteroides ovatus in ADHD gut microbiota (Aarts et al., 2017; Wang et al., 2020). Increased relative abundance of Bacteroides vulgatus, Bacteroides caccae, Paraprevotella xylaniphila, Odoribacter splanchnicus (phylum Bacteroidetes) as well as Bifidobacterium longum, Bifidobacterium adolescentis (phylum Actinobacteria), and Sutterella stercoricanis (phylum Proteobacteria) were observed in ADHD-associated microbiota (Table 3). Each set of findings were reported in only one study; further validation across larger samples is needed.
Changes in inferred metabolic function of the intestinal microbiota in ADHD subjects were assessed by two studies using PICRUSt-KEGG analysis. Wan et al. (2020) found an enrichment in the enzymatic pathways involved in dopamine turnover at the postsynaptic neuron as well as in genes encoding the catalytic subunit of protein phosphatase-1 (PP1), threonine synthase, and 6-pyruvoyl-5,6,7,8-tetrahydropterin, and depletion of the gene encoding 4-hydroxy threonine-4-phosphate dehydrogenase in ADHD-associated microbiota (Wan et al., 2020; Table 3). PP1 catalytic subunit has been described to increase synaptic sodium ion flux and may impact transmembrane sodium/chloride-dependent transporter dopamine receptors (Navratna and Gouaux, 2019). Abnormal Enterococcus and Odoribacter were found to be associated with alterations in the dopamine metabolic pathway in ADHD patients compared to NT controls (Wan et al., 2020). Aarts et al. (2017) reported increased abundance of the enzyme cyclohexadienyl dehydratase (CDT) in the gut microbiota of ADHD subjects. CDT is responsible for the production of phenylalanine, a precursor for the synthesis of dopamine and noradrenaline. Predicted CDT was found to be positively correlated with Bifidobacterium in ADHD. Reward anticipation, a key symptom in ADHD mediated by dopamine, was negatively correlated with predicted CDT in ADHD subjects (Aarts et al., 2017; Table 3). This study suggests that Bifidobacterium may enhance production of phenylalanine via the enzyme CDT in patients with ADHD. Several studies have assessed the role of phenylalanine in ADHD with conflicting results, showing decreased concentrations of phenylalanine in ADHD and no association (Bornstein et al., 1990; Baker et al., 1991; Bergwerff et al., 2016). In contrast, among patients with the metabolic condition, phenylketonuria, toxic phenylalanine build-up may lead to ADHD symptoms (Stevenson and McNaughton, 2013; Ashe et al., 2019; Beckhauser et al., 2020).
None of the studies in our review stratified the microbiota data according to the ADHD severity. Some correlations between bacterial taxa and ADHD symptoms, however, were reported. The relative abundance of B. uniformis, B. ovatus, and S. stercoricanis correlated with ADHD symptoms (Wang et al., 2020), and Coproccocus_2 with inattention symptoms, even though this association was not statistically significant (p = 0.055) (Szopinska-Tokov et al., 2020) (Figure 2; Table 3). A negative correlation occurred between hyperactivity scores and α-diversity in ADHD patients (Prehn-Kristensen et al., 2018), and between hyperactivity and impulsivity and Faecalibacterium levels (Figure 2; Table 3). No correlations were identified between microbiota changes and attention deficit and impulsivity (parental rating assessed by the German ADHD Rating Scale, FBB-HKS), or clinical symptoms assessed by the CBCL. A positive correlation between Bacteroides and levels of hyperactivity and impulsivity was observed (Figure 2; Table 3). The authors further assessed intestinal microbiota of mothers of ADHD patients and found patterns of similarly reduced α-diversity, which correlated with maternal ADHD symptoms as compared to NT maternal controls. There were no differences between α-diversity and ADHD symptoms between fathers of ADHD patients and fathers of NT controls (Prehn-Kristensen et al., 2018). The study of Richarte et al. (2021) did not find any correlation between bacterial taxa (family, genus) and ADHD rating scale scores; however, some correlations between bacterial taxa among ADHD subjects were identified (Table 3). Together these findings suggest relationships between ADHD symptoms and Bacteroides, yet the biological mechanisms behind these remain unknown.
Two studies from Italy showed a reduction in α- and β-diversity between RETT subjects and NT controls (Strati et al., 2016; Borghi et al., 2017). The study by Borghi et al. (2017) did not reach statistical significance. The authors suggested that this result was affected by the small sample size (Table 4). A third study, conducted in United States, did not find any differences in α- and β-diversities between RETT and NT controls; however, changes in bacterial richness, diversity, and composition were observed within RETT subjects based on pubertal stage, severity of disease and dietary habits (Thapa et al., 2021) (Table 4). These studies indicate a possible change in bacterial diversity associated with RETT. However further analyses in larger cohorts are required to confirm these results.
Most changes in the abundance of bacterial taxa associated with RETT occurred within the Firmicutes and Bacteroidetes phyla, followed by Actinobacteria and Proteobacteria phyla (Figure 2; Table 4). In general, RETT microbiota was characterized by reduced abundance of bacterial genera from the family Oscillospiraceae (Ruminococcus, Faecalibacterium, Oscillibacter, Sporobacter) and several genera from the phylum Bacteroidetes (Prevotella, Barnesiella, Alistipes, Odoribacter, Butyricimonas).
Conflicting findings were reported for the genus Bacteroides (Strati et al., 2016; Borghi et al., 2017). An increased relative abundance of the genera Lactobacillus, Enterococcus (phylum Firmicutes), Egghertella, Bifidobacterium, Actinomyces (phylum Actinobacteria) were observed in the gut microbiota of RETT subjects. Strati et al. reported decreased abundance of bacteria from the genus Bilophila and increased levels of bacteria in genera Escherichia and Shigella within phylum Proteobacteria (Strati et al., 2016). At the species level, subjects with RETT in the study by Borghi et al. (2017) showed decreased abundance of F. prausnitzii, Roseburia sp., Prevotella sp. and an increased abundance of Clostridium sp., and Sutterella sp., compared to NT controls. The study by Strati et al. (2016) also reported increase abundance of Bifidobacterium longum in RETT-associated microbiota (Table 4). Overall, these results suggest the presence of bacterial changes across multiple bacterial phyla that are associated with RETT.
Two studies investigated the potential metabolic function of gut bacteria using PICRUSt-KEGG analysis. Strati et al. (2016) observed an enrichment in carbohydrate and propanoate metabolism (pathways of SCFA production) in the gut microbiota of RETT subjects. In the study by Borghi et al. (2017), enzymes for carbohydrate and lipid metabolism were depleted, whereas amino acids pathways, butanoate and propanoate metabolism were enriched in RETT subjects (Table 4). These data suggest that the bacterial metabolic pathways responsible for SCFA production are altered in RETT. As these findings were only derived from two studies, larger cohorts of patients are required for further validation.
Specific correlations between intestinal microbial dysbiosis and disease severity in RETT were only assessed by Thapa et al. (2021), who found a significant reduction in α-diversity and differences in β-diversity between subjects with severe RETT compared to mild and moderate phenotypes (Figure 2; Table 4). Further studies are needed to understand whether there is a link between the composition of bacterial communities and RETT severity.
One study investigated alterations in the gut mycobiota associated with RETT (Strati et al., 2016). Fungal β-diversity measurements showed a significant difference between RETT subjects and NT controls (Table 4). Metataxonomic analysis led to the identification of 77 fungal genera that differed between groups, including Candida, Penicillium, Aspergillus, Malassezia. Among identified genera, only Candida was increased in RETT subjects compared with NT controls (Figure 2; Table 4).
Several studies assessed the concentration of microbial-derived metabolites in stool samples from ASD subjects and evaluated possible correlations with changes in microbial taxa associated with ASD. De Angelis et al. (2013) found that the metabolism of free amino acids (FAA) differed between patients with PDD-NOS and ASD compared to NT first-degree sibling controls. Levels of FAA and volatile organic compounds (VOCs) of subjects with PDD-NOS were more similar to NT controls than those of ASD on principal component analysis (PCA) plots (De Angelis et al., 2013).
Glutamine was highest in fecal samples of subjects with ASD compared to NT controls while indole, a microbial metabolite of tryptophan (which converts to serotonin) and 3-methylindole were increased in ASD and PDD-NOS subjects (De Angelis et al., 2013; Table 2). Kang et al. (2018) found trimethylamine, malonate, methylamine, choline, and 3- phenylpropionate weakly correlated with age in ASD subjects but were not statistically significant after correction for multiple hypothesis testing. Fecal concentrations of GABA, a major inhibitory neurotransmitter, were lower in stool of children with ASD in this cohort, although differences were only marginally significant. Concentrations of glutamate, a major excitatory neurotransmitter, were comparable between groups. GABA to glutamate ratio was observed to be marginally lower in stool of ASD subjects (Kang et al., 2018; Table 1).
Phenol compounds (phenol, 4-(1,1- dimethylethyl)-phenol, and para-Cresol) and methyl esters were increased in the stool of both PDD-NOS and ASD subjects compared to NT controls (De Angelis et al., 2013). This was also reported by Kang et al. (2018), who found significantly higher concentrations of p-cresol in stool of children with ASD compared to neurotypical controls. p-Cresol is introduced to the body either through skin, inhalation, or microbially produced in the intestine from dietary tyrosine or toluene substrates. The authors found no significant correlation between p-cresol and tyrosine in subjects’ stool (Spearman correlation, r = 0.08, p = 0.62) (Kang et al., 2018). p-cresol concentrations were also found to be significantly and negatively correlated with age within the ASD group (Spearman correlation, r = 0.47, p = 0.02), whereas no significant correlation was observed within NT subjects (Spearman correlation, r = 0.22, p = 0.34) (Kang et al., 2018) (Supplementary Table S5).
The study of Kang et al. (2018) involved multiple testing corrections (permutation-based non-parametric significance test, adjusted p = 0.022) among 59 metabolites and found that isopropanol was the only metabolite that was significantly different between ASD and NT controls after multiple testing corrections. In this same study, metabolite combinations from the 25 most differently abundant metabolites were evaluated to determine if they could support differentiation of ASD subjects from NT controls. Only five out of 23 children with ASD (78% sensitivity), and four out of 21 neurotypical controls children would not be classified as belonging to their true group (81% specificity). Further, using a more stringent threshold (>30% taxa membership in ASD subjects), cross-validation showed 70% sensitivity and 95% specificity (Kang et al., 2018). This analysis showed higher concentrations of caprate and aspartate in stool of subjects with ASD, while nicotinate, glutamine, and thymine were lower.
Differences in SCFA have also been described between subjects with ASD and NT controls (Figure 2; Tables 1, 2) Children with ASD showed significantly lower levels of total SCFA compared to NT, except for propionic acid which was increased (De Angelis et al., 2013). The proportion of SCFA correlated with levels of Faecalibacterium, Ruminococcus and Bifidobacterium (De Angelis et al., 2013). Liu et al. (2019) found correlations between microbial taxa and decreased fecal SCFA in ASD. Ruminococcaceae, Peptostreptococcaceae, Lactobacillales, Streptococcaceae, Eubacterium and Lachnospiraceae-NC2004 all correlated with decreased fecal concentrations of butyrate. A correlation was found between relative Acidobacteria levels and increased levels of fecal valerate (Spearman correlation, rs = 0.349, p = 0.013), Desulfovibrionaceae and increased fecal acetate (r = 0.360, p = 0.01), and Streptococcaceae and increased propionate (r = 0.368, p = 0.009) (Liu et al., 2019). Subjects in both cohorts were thought to have similar dietary contributions of butyrate and valerate, suggesting that differences in microbial taxa were the primary determinant of SCFA changes (Liu et al., 2019). Kang et al. (2018) found that fecal concentrations of propionate and butyrate were not different between ASD and NT controls. Other carbohydrate fermentation products of intestinal bacteria, formate and lactate, were detected at relatively lower concentrations in children with ASD, but their differences were not significant after multiple testing corrections (Table 1).
The study by Zhai et al. (2019) described high levels of Pb, As, Cu, Zn, Mg, Ca and Hg in children with ASD (Figure 2; Table 1). In particular, As and Hg were positively correlated with Oscillospira and Parabacteroides in ASD. Luna et al. (2017) assessed tryptophan and serotonin metabolite concentrations in supernatants of rectal biopsies obtain from ASD patients (all displaying FGID) and correlated these data with bacterial taxa that were abundant in biopsy preparations from an ASD cohort. Decreased tryptophan and increased 5-hydroxyindoleacetic acid (5-HIAA) levels were found in ASD rectal biopsies compared to NT control rectal biopsy preparations (Luna et al., 2017). Erysipelotrichaceae, C lituseburense, and Terrisporobacter species correlated with tryptophan concentrations from the rectal biopsies of the ASD cohort. L. bolteae, L. hathewayi, and F. plautii correlated with levels of serotonin in ASD tissue biopsy specimens (Luna et al., 2017; Table 1; Supplementary Table S5).
Microbial-derived metabolites were not directly investigated in ADHD studies.
Among RETT subjects, Borghi et al. (2017) found fecal butyrate and propionate to be significantly increased, as well as iso-butyrate and iso-valerate. This was found in conjunction with a greater abundance of Bacteroidetes and Veillonaceae, major propionate producers (Borghi et al., 2017; Table 4). Similar findings were described in a study by Strati et al. (2016), who found that the fecal content of SCFAs in RETT subjects was significantly enriched in propionate, isobutyrate and 2- methylbutyl isovalerate (Table 4). Concentrations of selected protein end-products of the gut bacterial metabolome, including GABA, tyrosine, and glutamate, were reduced in RETT subjects as compared to NT (Thapa et al., 2021; Table 4).
Overall, these studies show changes in microbial-host metabolites, neurotransmitters and trace elements in stool and intestinal mucosa of ASD and RETT patients that correlated with bacterial taxa altered in ASD patients. These results require further validation to infer a causal relationship between microbiota changes and neurodevelopmental outcomes.
Few studies assessed correlations between immune dysregulation and changes in microbial taxa associated with ASD (three studies) or RETT (one study). No studies assessed these relationships in individuals with ADHD. Hughes and Ashwood (2018) reported that children with ASD have elevations in IgG against Candida albicans with concomitant elevations of interleukin (IL)-17 (Table 1). Rose et al. (2018) also found that peripheral blood mononuclear cells (PBMC), isolated from the blood of subjects with ASD and GI symptoms (ASD-GI), produced elevated levels of interleukin (IL)-17, as well as IL-5 and IL-15, after exposure to the toll-like receptor (TLR)-4 agonist lipopolysaccharide (LPS), a component of gram-negative bacteria cellular walls. This elevation specifically occurred in PBMC isolated from ASD-GI subjects, while those with ASD-no-GI showed increased levels of IL-1α, IL-1β and Tumor Necrosis Factor (TNF)-α after TLR-4 stimulation of PBMC with LPS (Rose et al., 2018). Moreover, production of Transforming Growth Factor (TGF)-β1 was consistently decreased in ASD-GI subjects, as compared to subjects with ASD-no-GI and NT controls. The study by Luna et al. (2017) measured cytokines levels in the blood and biopsy supernatants from ASD-GI and NT controls, with or without GI symptoms (Figure 2; Table 1). In serum, decreased growth related oncogene alpha (GROα) and Interferon (IFN)-a2, and increased monocyte chemoattractant protein (MCP) and eotaxin levels were observed in ASD-GI subjects compared with NT-GI and NT controls. Moreover, increases in MCP-1 and eotaxin were found in ASD-GI subjects with abdominal pain. Luna et al. (2017) reported positive correlations between serum/mucosal cytokines and bacterial species associated with ASD-GI group (Table 1) and also between cytokines and bacterial species associated with pain in the ASD-GI group. C. disporicum correlated with macrophage inflammatory protein 1 alpha (MIP-1a) and beta (MIP-1β), vascular endothelial growth factor (VEGF), IFN-γ, IL12p70, IL17A, IL5, IL6 and IP-10. C. tertium correlated with IL1RA, IFN-γ, IL12p70, IL17A, IL1a, IL5, IL6, MIP-1a, and MIP-1b; P. excrementihominis correlated with GROα, and Tyzzerella nexilis with soluble CD40 ligand (Luna et al., 2017). The study by Tomova et al. (2015), which assessed probiotic supplementation in ASD subjects, identified a significant decrease in relative abundance of Firmicutes, Bifidobacterium and Desulfovibrio that correlated with decreased fecal TNFα levels in probiotic-treated ASD subjects (Table 5; Supplementary Table S9). Oxytocin was significantly lower in the plasma of subjects with ASD and NT first-degree sibling controls compared to unrelated NT controls. A positive correlation was found between plasma oxytocin levels and ASD severity (ADI) (r = 0.71, p < 0.05) (Tomova et al., 2015). Plasma levels of dehydroepiandrosterone sulfate (DHEA-S) were significantly lower in children with ASD, compared to NT first-degree sibling controls or unrelated NT controls. Testosterone levels did not show significant differences between groups but were positively correlated with ASD severity (as measured by ADI scores) (Tomova et al., 2015; Supplementary Table S9).
Inflammatory markers were not investigated in ADHD studies.
Strati et al. (2016) found that subjects with RETT had elevated levels of fecal calprotectin and serum erythrocyte sedimentation rate (ESR), which correlated with levels of serum IgA antibodies (Table 4). These increased inflammatory markers were hypothesized to be associated with RETT subjects’ lower intestinal bacterial microbiota richness and diversity as compared to NT controls (Strati et al., 2016).
GI symptoms were assessed using the Questionnaire on Pediatric Gastrointestinal Symptoms-Rome III (QPGS-RIII) in five studies (Son et al., 2015; Luna et al., 2017; Strati et al., 2017; Hughes and Ashwood, 2018; Rose et al., 2018) and using the 6-item Gastrointestinal Severity Index (6-GSI) in four studies (Kang et al., 2013, 2018; Liu et al., 2019; Ahmed et al., 2020; Supplementary Tables S5, S6). In two studies, GI symptoms were evaluated based on parental questionnaires (Gondalia et al., 2012; Tomova et al., 2015; Supplementary Tables S5, S6), and four studies did not assess the presence of GI symptoms in their patient cohorts (De Angelis et al., 2013; Pulikkan et al., 2018; Zhang et al., 2018; Zhai et al., 2019). Many ASD subjects included in these studies manifested GI symptoms including constipation, diarrhea, abdominal pain, and others.
Liu et al. (2019) observed that only constipation was significantly increased in ASD subjects compared to NT controls. ASD subjects with constipation displayed stool enrichments in Fusobacterium, Barnesiella, Coprobacter and valeric acid-associated taxa (Actinomycetaceae), and reduced butyrate-producing taxa (Liu et al., 2019). The study by Strati et al. (2017) found that taxa belonging to Clostridium cluster XVIII and pro-inflammatory bacteria Escherichia/Shigella positively correlated with ASD constipation status. Finegold et al. (2017) did not report their methodology for assessment of GI symptoms. This study found that samples from subjects with ASD have significantly higher counts of β2-toxin expressing-Clostridium perfringens compared to NT controls. 79% of the 33 ASD subjects showed the presence of β2- toxin gene in stool, compared to 38% of the 13 NT controls. Those subjects with ASD also showed more severe GI symptoms, although the correlation between β2- toxin gene levels and GI symptoms severity was not assessed. Bacterial richness was negatively correlated with GI symptoms in ASD subjects in the study by Kang et al. (2013) (Table 1). Almost 40% of anti-C. albicans+ IgG-or anti-C. albicans-I IgG- ASD subjects had GI symptoms in the study by Hughes and Ashwood (2018). Son et al. (2015) observed increased abdominal pain and functional constipation in ASD subjects compared with their NT siblings, but no significant correlation was identified between ASD, FGIDs or gut bacteria composition (Son et al., 2015).
In a study evaluating 50 individuals with ASD, Rose et al. (2018) showed that the relative abundance of Bacteriodaceae, Lachnospiraceae, Ruminococcaceae and Prevocellaceae differed between ASD-GI subjects and NT-GI controls. No differences were found among these taxa between ASD-no-GI subjects, and NT-no-GI controls. ASD-GI subjects had more irritability and agitation, social withdrawal, lethargy, and hyperactivity compared to ASD-no-GI. Further analyses showed that microbiota-related biological pathways were differentially represented between groups and that alterations were related to the presence of ASD, and not GI symptoms independently (Rose et al., 2018). In contrast, the studies by Tomova et al. (2015) and Kang et al. (2018) reported positive correlations between GI symptoms and ASD severity in their cohort of patients. Luna et al. (2017) found positive correlations between bacterial taxa (from mucosal biopsies) and GI symptoms in both ASD and NT subjects. Six bacterial taxa were higher in ASD with abdominal pain compared to ASD without abdominal pain, as well as NT controls with and without abdominal pain (Luna et al., 2017; Table 1).
Ahmed et al. (2020) showed that the 6-GSI score was not correlated with changes in microbiota profiles among subjects with ASD. This was corroborated by Gondalia et al. (2012), who also found no differences in intestinal microbial profiles between ASD-GI and ASD-no-GI subjects.
In the study by Yap et al. (2021), reduced stool consistency, as measured on one stool sample using the Bristol Stool Chart, was associated with reduced dietary and microbiota diversity in ASD patients. Kushak et al. (2017) reported constipation and gastroesophageal reflux in ASD subjects assessed for GI symptoms through upper GI endoscopy. However, the authors did not observe any effects of GI symptoms on mucosal-associated microbiota or duodenal enzyme activity (Kushak et al., 2017). Overall, these findings confirm the presence of GI comorbidities in subjects with ASD. Nevertheless, relationships between gut microbes, GI symptoms and ASD-related neurobehavioral outcomes remain unclear.
The study by Wan et al. (2020) was the only study included in our systematic review to describe GI comorbidities in ADHD patients. An association was reported between bacterial changes and constipation symptoms in the ADHD cohort (Supplementary Table S7).
All individuals with RETT suffered from constipation, although relationships between microbiota alterations or disease severity with GI symptoms were not investigated (Table 4; Supplementary Table S8).
Eight out of 17 studies on ASD documented dietary patterns of patients. The effect of diet in shaping the gut microbiota composition, however, was evaluated only by Yap et al. (2021) (Table 2; Supplementary Table S6). The investigators in this study performed stool metagenomics in a large (n = 247) cohort of ASD and NT controls from a closely observed population from the Australian Autism Biobank and Queensland Twin Adolescent Brain project. ASD subjects had greater restriction in food intake and a less-diverse diet, which was associated with lower microbial α-diversity (Yap et al., 2021). This study found little evidence for a direct association between intestinal microbiota and ASD; rather, any observed changes were found to be most associated with influences of stereotypic dietary intakes in ASD subjects compared to NT controls (Yap et al., 2021).
A strictly controlled cross-sectional cohort study was conducted by Prehn-Kristensen et al. (2018) where subjects with ADHD were matched with NT controls with similar BMI and food intake (meat, fruits/vegetables, yogurt, milk products, or fast food during the month prior to microbiota analysis; Table 3; Supplementary Table S7). Differences were observed in α-diversity between subjects with ADHD and NT, and authors speculated that microbial changes in ADHD individuals were due to the underlying disease and not dietary patterns. In a cohort study by Wang et al. (2020), diets of ADHD subjects and NT controls were compared. ADHD subjects were found to have higher intake of refined grains (p = 0.027) and a lower proportion of dairy (p = 0.020) and vitamin B2 (p = 0.033) intake (Wang et al., 2020). In this study, S. stercoricanis was significantly associated with dairy, nut, seed and legume intake, and B. uniformis correlated with fat and carbohydrate intake. S. stercoricanis and B. ovatus also positively correlated with ADHD symptoms. These findings correlate with findings from Aarts et al. (2017), who demonstrated greater abundance of B. uniformis and B. ovatus in ADHD patients. Bacteroides sp. have been associated with the development of the frontal lobe, cerebellum and hippocampus (Aarts et al., 2017; Table 3; Supplementary Table S7). Overall, these data suggest that changes in the microbiota may correlate with dietary intake and not the actual disease state.
The influence of diet on gut microbiota profiles among NT controls and RETT subjects was only investigated by Thapa et al. (2021) (Table 4; Supplementary Table S8). Differences in bacterial composition and increased α-diversity were observed in RETT subjects who consumed table foods alone or combined with formula compared with RETT subjects who received exclusive formula. However, the effect of the diet in influencing disease severity was not investigated (Thapa et al., 2021).
Two studies examined the effects of probiotic treatment on gut microbiota profiles, disease outcomes and GI symptoms in patients with ASD (Tomova et al., 2015; Shaaban et al., 2018; Table 5; Supplementary Table S9). Both studies used a mix of Lactobacillus and Bifidobacteria strains, orally administered once (Shaaban et al., 2018) or three times per day (Tomova et al., 2015) for one month (Table 5). In both studies, NT controls were siblings of ASD subjects. Shaaban et al. (2018) found that decreases in the amount of Bifidobacterium in stool from ASD subjects recovered to levels observed in NT controls following probiotic treatment (Table 5). Conversely, Tomova et al. (2015) showed higher amounts of Bifidobacterium and increased Clostridium, Desulfovibrio, and Lactobacillus in ASD subjects. After probiotic treatment, levels of Bifidobacterium and Lactobacillus of ASD subjects decreased, reaching levels observed in NT controls (Tomova et al., 2015).
Tomova et al. (2015) did not assess the effect of probiotic treatment on disease severity. Shaaban et al. (2018) observed a significant decrease in ASD severity in probiotic-treated patients, represented by improvements in ATEC scores (Table 5).
In the study by Pärtty et al. (2015), Lactobacillus rhamnosus GG was administered to mothers daily, from four weeks before delivery up to six months of life (Table 5). Subjects were followed for 13 years to assess whether probiotic administration in early life reduced the risk of developing AS and ADHD in adolescence (Pärtty et al., 2015). ADHD or AS was diagnosed in 17.1% of subjects in the placebo group vs. 0% of the probiotic-supplemented group by age 13. There were no differences in bacterial abundance between ADHD subjects and NT controls, by 3 months or 13 years. Decreases in Bifidobacterium, Clostridium histolyticum, Bacteroides, Lactobacillus, and Enterococcus observed in 2-year-old children treated with placebo were associated with the development of AS or ADHD at 13 years old. These taxa differences did not persist at age 13 years (Pärtty et al., 2015; Table 5).
In an open-label study, Kang et al. (2017) showed that fecal microbiota transfer (FMT) to patients with ASD, using stool derived from healthy controls (Standardized Human Gut Microbiota) (Hamilton et al., 2012) (Table 5; Supplementary Table S9) increased bacterial diversity compared to NT controls and remained higher than baseline bacterial diversity 8 weeks after final administration of FMT (Kang et al., 2017). In ASD patients, the relative abundance of Bifidobacterium, Desulfovibrio, and Prevotella increased following FMT. Median microbial richness at week 18 was not statistically different between ASD and NT control groups (p > 0.05). A positive engraftment of phageome from the healthy donor to ASD subjects and increased abundance of phage was also observed after FMT (Table 5). Similar to the probiotic treatment described by Shaaban et al., FMT significantly improved ASD symptoms by improving social skill deficits, stereotypy, communication, daily living skills and socialization (Kang et al., 2017) (Table 5).
Probiotic and FMT intervention have demonstrated beneficial effects in improving GI symptoms in ASD patients (Table 5). Among recipients of probiotic supplementation, improvements in GI symptoms (assessed by the 6-GSI) were also strongly correlated with improvements in autism severity (assessed by the ATEC) (r = 0.674, p = 0.0001; Shaaban et al., 2018). Specifically, these subjects had a significant reduction in constipation scores (p < 0.01), stool consistency (p < 0.023), flatulence (p < 0.037) and abdominal pain (p < 0.002). Further, a significant correlation was found between probiotic administration, stool colony counts of Bifidobacteria, and reduction of constipation severity score (r = −0.441, p < 0.015; Shaaban et al., 2018). In the FMT study by Kang et al. (2017), 89% of ASD subjects had improvement in GI symptoms as reflected by an 80% reduction in the Gastrointestinal Symptom Rating Scale (GSRS). Further, improvements persisted up to 8 weeks after FMT (follow up) though similar changes in microbial richness were also observed in one of two non-responders (subjects whose GI symptoms improved less than 50% on the GSRS; Kang et al., 2017).
Stevens et al. (2019) assessed the effects of dietary supplementation on the modulation of ADHD symptom expression and gut microbiota composition in a subset of participants from a large RCT (Table 5; Supplementary Table S9). Participants ages seven to 12 years with a diagnosis of ADHD were randomized to receive a 10-week course of broad-spectrum micronutrients containing a mixture of vitamins, minerals, amino acids, and antioxidants, or inert placebo control. Subjects receiving the micronutrient supplementation had improved symptoms of attention, emotional regulation, and aggression relative to placebo. Eighteen children from this cohort (n = 8 placebo, n = 10 micronutrient group) had their fecal microbiota assessed and showed a decrease in Bifidobacterium genera by 25% in the micronutrient treatment group while Coriobacteriales increased though these changes were not significant (Stevens et al., 2019).
The gut microbiota contributes to the maintenance of human health by modulating the function of numerous organs (Al Nabhani and Eberl, 2020; Beckhauser et al., 2020; Wang et al., 2021), including the central nervous system (Ratsika et al., 2023). Changes in gut microbial communities have been described in children and adults with neurodevelopmental disorders. These patients display altered dietary patterns and GI comorbidities, particularly constipation, visceral sensitivity, and intestinal inflammation. These factors collectively play a critical role in shaping gut microbiota composition, especially in early life. Given this, a key unresolved question is whether changes in the microbiota dynamics are part of the biomolecular alterations that cause neurodevelopmental disorders or are merely a consequence of gastrointestinal dysfunctions or food selectivity more commonly associated with these conditions.
We present the first systematic review to date assessing the influence of altered gut microbiota composition on the expression of three highly prevalent pediatric and adult neurodevelopmental disorders: ASD, ADHD and RETT. Herein, we highlight the implications of intestinal microbiota changes on diverse aspects of host–microbe interactions that may contribute to underlying neuropathology. Our inclusion criteria identified studies that explore intestinal microbiota dynamics in stool and intestinal mucosal biopsy samples of children and adults affected by ASD, ADHD, and RETT. Associations between microbial changes, disease-related behavioral and gastrointestinal outcomes were evaluated across 13 ASD studies, five ADHD studies, and one RETT study, per our predefined review criteria (Hill et al., 2021). Further, nine ASD and three RETT studies also sought to evaluate associations between gut microbiota composition and alterations in microbial-derived metabolites, trace elements, enzymes, and/or immune-inflammatory markers. Lastly, we assessed studies that sought to explore the role of microbial- or dietary-mediated interventions in restoring gut eubiosis and behavioral outcomes.
The main finding that emerged from our review is the disproportionate abundance of bacterial taxa from the phyla Firmicutes and Bacteroidetes in the microbiota of individuals affected by ASD, ADHD, and RETT. These taxa could possibly alter host-microbe homeostasis toward a pro-inflammatory phenotype, ultimately contributing to the neurological symptoms typical of these three diseases. The majority of ASD and RETT subjects are affected by GI symptoms, and RETT subjects have intestinal inflammation. ASD subjects display changes in microbiota-derived metabolites (SCFAs, neurotransmitters, FAA), inflammatory markers such as interleukins, interferons, and other proinflammatory and regulatory cytokines. Increased levels of As and Hg, trace elements with potential neurotoxic activity were also found in ASD subjects. Symptoms of ADHD, particularly hyperactivity and impulsivity were negatively correlated with Faecalibacterium (phylum Firmicutes) and positively correlated with Bacteroides sp. (phylum Bacteroidetes). Individuals with RETT are mostly constipated and have intestinal dysbiosis, associated with increased SCFAs and reduced GABA and glutamate levels. Increased abundances of Candida sp. were observed in ASD and RETT subjects. Our review shows that microbiota-targeted interventions with probiotics or FMT are capable of ameliorating or preventing ASD and ADHD symptoms. However, despite such evidence, many inconsistencies were identified across the included studies, primarily due to the small sample sizes, a wide range of subjects’ age, cross-sectional design with single time-point sampling, different sequencing techniques and other many confounding factors including, but not limited to, GI dysmotility, differences in diet and sex. Therefore, further research in studies with larger sample sizes and longitudinal design is necessary to determine the extent of the contribution of the gut microbiota to the neurobehavioral alterations of neurodevelopmental disorders.
At first, we assessed bacterial diversity between neurodivergent and NT subjects. Our systematic review identified changes in α- and β-diversity between subjects affected by ASD or RETT, compared to NT controls. However, these changes were inconsistent among selected studies. A significant discrepancy occurred in studies comparing the gut microbiota of ASD individuals with NT first-degree sibling controls or unrelated NT controls. Most studies in which NT controls were first-degree siblings of ASD subjects did not detect any differences in α- or β-diversity of gut bacterial populations (Gondalia et al., 2012; Son et al., 2015; Pulikkan et al., 2018) but one study, which examined the gut microbiota of subjects with ASD and PDD-NOS, found that both α- and β- diversities were higher in ASD subjects than their NT first-degree siblings (De Angelis et al., 2013). Three studies (Tomova et al., 2015; Ahmed et al., 2020; Yap et al., 2021) compared the gut microbiota of ASD subjects with both NT first-degree siblings and unrelated NT controls. In these studies, no differences in gut microbiota diversity were found between the three groups. Distinct bacterial changes were observed only between ASD and unrelated NT controls, except for Bifidobacterium, which differed [lower (Ahmed et al., 2020) and higher (Tomova et al., 2015)] compared to NT first-degree siblings. Other studies enrolling unrelated NT controls have found significant differences in bacterial diversity between NT and ASD-associated microbiota. The same results emerged in studies of RETT subjects, where only unrelated NT controls were used for microbiota comparison (Kang et al., 2013, 2018; Strati et al., 2016, 2017; Borghi et al., 2017; Zhang et al., 2018; Liu et al., 2019; Zhai et al., 2019; Thapa et al., 2021).
The most plausible explanation for these discrepancies is that first-degree siblings of neurodivergent children possess similar alterations in their intestinal microbiome secondary to shared environmental exposures that include dietary habits from shared cooking, living conditions, and host genetic factors (Orsmond and Seltzer, 2007; Wade et al., 2015). Further, a higher relative prevalence of first-degree siblings of ASD subjects have been shown to display some neurocognitive alterations even without a diagnosis of ASD (Wade et al., 2015), suggesting that some NT siblings may manifest a disease subtype, with a different host-microbe phenotype, compared to unrelated NT individuals (Son et al., 2015).
Our review showed no difference in α- or β-diversities of the gut microbiota between individuals with ADHD and NT first-degree siblings, or unrelated NT controls.
Monozygotic twin studies have shown significant heritability of ADHD (h2 = 0.77-0.82), yet up to 20% discordance in diagnosis remains (Sprich et al., 2000; Thapar et al., 2013). Only one study in our systematic review found differences in gut microbiota composition between ADHD subjects and unrelated NT controls. Interestingly, the mothers of those ADHD individuals displayed reduced bacterial α-diversity compared to their offspring. This suggests that the microbiota could be partially involved in the intergenerational transfer of an altered gut-brain phenotype responsible for the disease (Prehn-Kristensen et al., 2018), a finding that has also been described in mouse models of inflammatory bowel disease. However, these studies’ cross-sectional design precludes firm conclusions.
The sample sizes of most of the studies included in our systematic review were small (N = 10-50), and this might have contributed to some of the variability in microbiota measurements. We included two studies that assessed microbiota changes in ASD (Yap et al., 2021) and ADHD (Richarte et al., 2021) in larger sample sizes (>100 individuals per group). Yap et al. (2021) performed shotgun metagenomics analyses in stool samples from a large cohort of ASD children (approximately 100 ASD subjects and unrelated NT controls, and 50 pairs of ASD subjects and NT first-degree siblings). This study found a significant association between microbiota changes in ASD and ASD-related dietary preferences, and no changes in the microbiota of ASD subjects as compared to NT controls (siblings, or unrelated healthy controls) (Yap et al., 2021).
Richarte et al. (2021) examined the gut microbiota of 100 medication-naïve ADHD subjects. Changes were found in relative abundance of several microbial taxa, but no differences were found in α- or β-diversity indices between ADHD and NT controls. This confirmed the findings of other studies with smaller sample sizes (Richarte et al., 2021). Conflicting results between the study by Yap et al. (2021) and other ASD studies might be secondary to differing methodologies used for microbial sequencing and bioinformatics analyses.
The choices of sequencing techniques, genome databases and sample sizes and other factors that may also shape the gut microbiota during early childhood development (e.g., GI dysfunction, dietary impact, and geographical locations) are crucial for determining the influence of microbial changes on neurobehavioral pathology (Hamady and Knight, 2009; Chavaria et al., 2022; Morton et al., 2022).
The two major bacterial phyla of the human gut microbiome, Firmicutes and Bacteroidetes, were evaluated across all studies. Our systematic review shows that differences in the relative abundance of both phyla, between ASD, RETT, and ADHD subjects and NT controls are not consistent, and the ratio between these two phyla does not seem to be an adequate differentiator between these neurodevelopmental disorders and NT control populations.
Taxa belonging to phylum Firmicutes are involved in the intestinal fermentation of carbohydrates undigestible by the host, which may result in the production of SCFAs, one of the most characterized families of microbial-derived metabolites that have also been recognized to influence brain function through different biological pathways within the MGBA (Plöger et al., 2012; Bourassa et al., 2016; Erny et al., 2021; O’Riordan et al., 2022). Increasing evidence suggests that SCFAs, such as butyrate, may be linked to neurodevelopment. Besides being an energy substrate for intestinal colonocytes and strengthening the intestinal epithelial barrier, butyrate participates in early life immune programming and modulates gene expression of brain neurotransmitters and structural components of the BBB (Zhang et al., 2018; Deleu et al., 2021; O’Riordan et al., 2022). Lower levels of butyrate or dysbiosis of butyrate-producing bacteria could be a potential contributor to the neuroimmune alterations observed in neurodevelopmental disorders such as ASD. Higher levels of propionate-producing bacteria have also been associated with the development of ASD in few studies (Finegold et al., 2002, 2010; Finegold, 2011). Propionate is a SCFA involved in gluconeogenesis and energy production in the liver, and studies in animals have shown that injection of propionic acid in the rat brain can induce neurobehavioral alterations characteristic of ASD (MacFabe et al., 2008; Abuaish et al., 2022). Our systematic review shows lower levels of butyrate and acetate, and increased levels of valeric acid and propionate in fecal samples from ASD individuals (De Angelis et al., 2013; Liu et al., 2019). These fluctuations of SCFAs levels are paralleled by reduced abundance of butyrate- and acetate-producing bacteria (Zhang et al., 2018) and increases in the proportion of Desulfovibrio (Finegold, 2011), Clostridium (Tomova et al., 2015) and Bacteroides (De Angelis et al., 2013), bacterial groups primarily involved in the production of propionate and acetate (Zhou et al., 2021; O’Riordan et al., 2022). Nevertheless, data on the effects of SCFA in ASD that emerged from our review warrant further exploration in larger sample sizes, as they do not correspond with what has been previously reported in literature (Adams et al., 2011; Wang et al., 2012),or show trends that are not statistically significant (Kang et al., 2018).
RETT is associated with high levels of propionate, isobutyrate, and isovalerate in the stool of affected subjects enrolled in the studies by Strati et al. (2016) and Borghi et al. (2017). The authors have linked the enrichment of Bifidobacterium, Bacteroides, Clostridium, Enterococcus, Escherichia/Shigella and Lactobacillus in RETT microbiota with increased production of SCFAs and have also demonstrated the role of SCFAs in slowing intestinal motility and causing constipation, a frequent GI comorbidity in RETT subjects (Strati et al., 2016; Borghi et al., 2017).
Distinct changes in bacterial taxa in subjects with ASD compared to NT controls were reported across multiple studies, with no specific taxa consistently identified. In ASD, decreases in Faecalibacterium, Streptococcus (Phylum Firmicutes), Prevotella (Phylum Bacteroidetes), Bifidobacterium (phylum Actinobacteria), and increases in Clostridium, Lactobacillus (phylum Firmicutes), and Bacteroides (phylum Bacteroidetes) were observed across studies that assessed bacterial taxonomic composition using the 16S rRNA gene (De Angelis et al., 2013; Kang et al., 2013, 2018; Strati et al., 2017; Pulikkan et al., 2018; Zhang et al., 2018; Liu et al., 2019; Zhai et al., 2019), by performing qPCR for specific bacterial taxa (Son et al., 2015; Tomova et al., 2015) or through quantification from bacterial cultures (De Angelis et al., 2013). Reduction in the genus Prevotella in children with ASD was one of the most consistent findings emerging from our systematic review, as well as from previous work (Ho et al., 2020). However, the role of this bacterial group in affecting host neurobiology is still unknown. In adults, a Western-style diet poor in fiber content and rich in saturated fat has been found to affect Prevotella and Bacteroides abundance (Derrien et al., 2019; Tett et al., 2021). As suggested by Yap et al. (2021), ASD is associated with reduced dietary diversity, which in turn can impact microbiome diversity and GI function. The altered dietary behaviors of ASD individuals have a significant impact on the developmental trajectory of the microbiome in early life, and a dysbiotic microbiome affects host physiology, metabolism, and brain function which may contribute to some of the behavioral traits seen in ASD.
ADHD and RETT are characterized by alterations in bacterial taxa belonging to the phyla Firmicutes and Bacteroidetes, and similar to ASD, these changes were not consistent across the included studies. A recent meta-analysis that analyzed some of the studies included in our systematic review identified a significant increase of Blautia (phylum Firmicutes) in ADHD subjects (Wang et al., 2022). This bacterial genus has been associated with various human diseases, such as depression, obesity, and diabetes (Wang et al., 2022), yet the link between Blautia and neurodevelopment is still undefined. Correlations between bacterial groups and disease symptoms, or bacteria-associated functional pathways related to dopamine or norepinephrine turnover have been described across different ADHD studies (Aarts et al., 2017; Jiang et al., 2018; Prehn-Kristensen et al., 2018; Wang et al., 2020).
Besides changes in microbial composition, our systematic review also identified other elements that link the microbiota with host neuronal function. For example, decreased GABA levels were observed in stool from ASD (De Angelis et al., 2013) and RETT (Thapa et al., 2021) subjects. Decreased tryptophan and increased 5-HIAA, the primary metabolite of serotonin, were observed in ASD intestinal tissues, and such changes were correlated with specific bacterial taxa. GABA, glutamine, serotonin, and tryptophan are neurotransmitters of the PNS and CNS that play a critical role in modulating neuronal activity within the gut-brain axis. Bacteria in the gut can synthesize and metabolize such neurotransmitters, using them as host-microbe interkingdom signaling molecules and thus contributing to the neuro-hormonal functions of the host (Burton et al., 2002; Nzakizwanayo et al., 2015). Nevertheless, the impact of host-microbiota signaling in influencing neurobehavioral trajectories and its implication in neurodevelopmental disorders remains unclear. One study described high levels of minerals such as As and Hg and their correlation with Oscillospira and Parabacteroides in children with ASD. Changes in the levels of trace elements have been associated with the pathophysiology of many neurological disorders such as Huntington’s disease, amyotrophic lateral sclerosis, Parkinson’s disease, Alzheimer’s disease and others (Bayir, 2015).
One of the critical factors that can influence both microbiota dynamics and nervous system integrity in neurodevelopmental disorders is the aberrant activity of the immune system and the consequent local or systemic inflammation. Increases in proinflammatory cytokine production upon TLR4 stimulation in blood cells (Rose et al., 2018), or changes in serum cytokine levels (Luna et al., 2017) have been identified in ASD subjects. High fecal calprotectin and ESR were found in RETT subjects. Only the study of Luna et al. (2017) correlated the altered serum cytokines profiles of their ASD subjects with the microbiota changes observed in intestinal tissues. These preliminary data suggest a possible link between microbiota changes and neuroimmune alterations that may affect gut-brain axis signaling and behavior in ASD and RETT. However, only some studies included in our review evaluated host-microbial metabolites, neurotransmitters, or cytokines together with microbiota analyses. For instance, all the studies included in our systematic review on ADHD assessed microbiota composition and function without investigating other factors that may contribute to host-microbiota interactions associated with the disease. Our systematic review provides clear evidence of how microbiota data in neurodevelopmental disorders may be strengthened, by conducting clinical studies using larger cohorts and including concomitant neuronal and inflammatory markers analysis.
The GI tract is colonized not only by bacteria, but also fungi, viruses, and other microorganisms that reside in mutually beneficial relationships with each other and their hosts. Perturbations in the composition of commensal bacteria can alter this inter-kingdom homeostasis, favoring the overgrowth of other microbial species that may become potentially pathogenic to the host. Strati et al. found alterations in the gut mycobiome of RETT (Strati et al., 2016) and ASD (Strati et al., 2017). Both pathologies are associated with higher abundance of intestinal Candida and Aspergillus. These results are supported by the identification of three Candida species in stool samples of a large cohort of pediatric ASD subjects in Turkey (Kantarcioglu et al., 2016), and anti-Candida albicans IgG in the blood of ASD subjects from a cohort in California, United States (Hughes and Ashwood, 2018).
Subjects from all studies evaluating the presence of GI comorbidities in ASD and RETT in our systematic review experienced constipation and abdominal pain (Kang et al., 2013, 2018; Son et al., 2015; Tomova et al., 2015; Strati et al., 2016, 2017; Borghi et al., 2017; Pulikkan et al., 2018; Rose et al., 2018; Liu et al., 2019; Zhai et al., 2019; Thapa et al., 2021). GI symptoms pose a significant burden in children with neurodevelopmental disorders (Ming et al., 2008; Adams et al., 2011). GI symptoms correlated with severity of ASD symptoms in three of 19 included studies (18 case–control studies and one case–control study with intervention). Notably, in these studies both ASD and GI symptoms were measured using three different scoring tools (Tomova et al., 2015; Kang et al., 2018; Rose et al., 2018). In agreement with previous findings (Adams et al., 2011), Kang et al. (2018) reported that GI symptoms, assessed by the modified 6-GSI score, correlated with ASD symptom severity assessed by the ATEC, which is completed by parents. However, when a physician assessed GI symptoms in ASD subjects there was no correlation between ASD symptom severity with GI symptoms (Kang et al., 2013; Son et al., 2015). Bacterial taxa were associated with constipation (Liu and Zhu, 2018; Zhou et al., 2022) or visceral pain (Luna et al., 2017) in ASD subjects.
Our review shows the presence of GI symptoms in ASD and RETT, but not in ADHD. Few studies examined the possible correlations between GI symptoms and disease severity, however, no signature of disease- or GI symptom-driven change in gut microbial communities could be identified. GI tract dysfunction could be the cause of altered microbial communities in subjects with ASD or RETT; however, this link remains unclear.
Dietary habits of NT and ASD, ADHD, and RETT patients were documented in most of the studies included in our systematic review but not included in the microbiota analysis.
Food is the primary source providing nutrients that are fundamental precursors of neurotransmitters and neuromodulators, therefore diet has a critical impact on brain health (Marrone and Coccurello, 2019). The study by Yap et al. (2021) is the only one assessing dietary and microbiota diversity in ASD subjects compared to NT siblings and unrelated NT controls. ASD subjects seem to have less dietary diversity rather than microbiota diversity. Indeed, the study reports a lower quality of food and reduced meat intake in ASD compared to NT subjects. Such dietary imbalance, especially during child development, can significantly affect the quality and quantity of macronutrient intake, for instance lipids, thus reducing the production of bioactive lipid molecules, important mediators of brain signaling pathways (Coccurello et al., 2022). The study by Thapa et al. (2021) shows that solid food intake increases intestinal bacterial diversity in RETT subjects compared to those who are fed with only formula. This functional connection between diet and microbiome needs to be further explored as could set the basis for nutritional interventions targeting the microbiota-gut ecosystem.
It is still not known if therapeutic modulation of microbiota can improve brain health and behavioral outcomes in neurodevelopmental disorders. Our systematic review identified several clinical studies investigating the effect of probiotics or FMT in ASD and ADHD. Supplementation of Lactobacillus species (Shaaban et al., 2018) or FMT from NT controls (Kang et al., 2017) in open-label studies have been shown to significantly improve social skills, language and communication, irritability, stereotypy, as well as to reduce constipation in children with ASD. In a follow-up study by Kang et al. (2018), 18 patients from their initial FMT-treated ASD cohort were observed for an additional 2 years. The authors describe most improvements in GI symptoms were maintained among this cohort (58% reduction in GSRS relative to baseline), and autism-related symptoms improved even further after FMT treatment ended (47% reduction in CARS at two-year follow-up, compared to 23% lower at final FMT treatment). This study was not included in our systematic review due to not meeting inclusion criteria. Further control studies are needed to discern the potential for placebo effect on GI symptoms reporting.
Pärtty et al. (2015) showed that perinatal Lactobacillus rhamnosus GG supplementation may reduce the risk of ADHD or ASD development in later childhood, possibly through mechanisms involving interactions between the probiotic and neuroimmune component of the MGBA such as the vagus nerve. Epidemiological data have suggested that probiotic treatment in early life may reduce risks of ADHD diagnosis at 13 years of age, while insults that promote microbial dysbiosis like antibiotic exposure and cesarean section delivery may increase risks (Curran et al., 2015; Pärtty et al., 2015; Slykerman et al., 2017). These probiotics have been extensively studied in preclinical models of gut-brain axis anomalies such as anxiety (Zhou et al., 2022), depression (Kochalska et al., 2020), and stress-induced visceral pain (McVey Neufeld et al., 2020). This may offer a promising approach to understanding the biological mechanisms linking gut microbes with neurodevelopment, and to develop novel early therapeutic interventions for neurodevelopmental disorders in the future.
This comprehensive systematic review highlights important milestones that may guide future research on the MGBA in pediatric neurodevelopmental disorders. The most salient limitation that emerged from this review is the sample size of our included studies. Pediatric and adult-focused studies in this area remain limited, and results of our review propose valuable hypotheses that may be better assessed in larger cohorts.
The study of microbial dynamics in neurodevelopmental disorders must consider several factors beyond the primary research question. Age, stress across the lifespan, differences in dietary habits, presence of concomitant GI dysfunction, geographic location, host genetics and physiology all can influence gut microbiota composition, confounding establishment of relationships between microbes and host neuroimmune homeostasis. For example, the dietary habits of the participants were documented in only eight (of eighteen) ASD studies, three of seven ADHD studies, and two of three RETT studies. Only Yap et al. (2021) and Thapa et al. (2021) explored the link between altered dietary patterns in ASD or RETT subjects and microbiota changes. The stereotypical diet of neurodivergent subjects may play a determining role in shaping the microbiota communities, and therefore diet should always be included in the analysis.
We assessed quality of evidence by applying a semi-quantitative appraisal with CASP checklists, a widely used tool for quality appraisal in health-related studies. While informative, the CASP tool is limited by its assessment of what has been reported by a given study only (Long et al., 2020). Our use of the CASP tool offered a clear, consistent appraisal of the literature by assessing individual aspects of quality (Harrison et al., 2017).
Overall, the studies included in our search did not have a longitudinal design except the study of Pärtty et al. (2015) Thus, it is unknown if microbiome changes are a precursor to disease manifestation, or if the relationship between specific microbes and behavioral symptoms persist over time and occurred secondarily.
The choice of the most appropriate control group is fundamental to draw appropriate comparisons and represents a challenge across many microbiome studies. Differences existed across studies with use of first-degree sibling NT controls, or unrelated NT controls. A further limitation of clinical microbiome research is the impact of sampling procedure and bioinformatics analytic techniques. The 16S-rRNA gene sequencing provides information on taxonomic composition with limited resolution and poor functional readout, whereas shotgun metagenomics leads to superior sequencing and functional profiling of metabolic capacity of gut microbes (Jovel et al., 2016). Inconsistencies in sample handling, variability in sequencing methodologies and resolution, and differences in bioinformatic databases used between studies all may contribute to variability between study conclusions.
In summary, our systematic review describes the presence of alterations in the gut microbiota of patients with common neurodevelopmental disorders such as ASD, ADHD, and RETT. However, these findings are inconsistent and do not support causal inferences between microbiome changes and neurodevelopmental outcomes. Future studies are required to investigate the functional role of the gut microbiota in the pathophysiology of neurodevelopmental disorders and such studies should include longitudinal design, host- and microbial-derived metabolites, dietary and physiological characteristics of the patients, and more powerful tools to sequence and analyze microbial communities.
The original contributions presented in the study are included in the article/Supplementary material, further inquiries can be directed to the corresponding author/s.
VC: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Project administration, Supervision, Validation, Writing – original draft, Writing – review & editing. LH: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Project administration, Supervision, Validation, Writing – original draft, Writing – review & editing. MF: Conceptualization, Data curation, Investigation, Writing – original draft. JP: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Writing – original draft. EH: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Writing – original draft. KM: Supervision, Writing – original draft. KB: Data curation, Visualization, Writing – original draft. PJ: Data curation, Visualization, Writing – original draft. MM: Data curation, Investigation, Writing – original draft. NP: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Project administration, Supervision, Validation, Writing – original draft, Writing – review & editing.
The author(s) declare that no financial support was received for the research, authorship, and/or publication of this article.
The authors wish to thank Evdokia Anagnostou for her critical review and comments on the initial drafts of the submitted manuscript.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fnins.2024.1341656/full#supplementary-material
Aarts, E., Ederveen, T. H. A., Naaijen, J., Zwiers, M. P., Boekhorst, J., Timmerman, H. M., et al. (2017). Gut microbiome in ADHD and its relation to neural reward anticipation. PLoS One 12, 1–17. doi: 10.1371/journal.pone.0183509
Abraham, E., Scott, M. A., and Blair, C. (2020). Catechol-O-methyltransferase Val158Met genotype and early-life family adversity interactively affect attention-deficit hyperactivity symptoms across childhood. Front. Genet. 11:724. doi: 10.3389/fgene.2020.00724
Abuaish, S., Al-Otaibi, N. M., Aabed, K., Abujamel, T. S., Alzahrani, S. A., Alotaibi, S. M., et al. (2022). The efficacy of fecal transplantation and Bifidobacterium supplementation in ameliorating propionic acid-induced behavioral and biochemical autistic features in juvenile male rats. J. Mol. Neurosci. 72, 372–381. doi: 10.1007/s12031-021-01959-8
Adams, J. B., Johansen, L. J., Powell, L. D., Quig, D., and Rubin, R. A. (2011). Gastrointestinal flora and gastrointestinal status in children with autism – comparisons to typical children and correlation with autism severity. BMC Gastroenterol. 11:22. doi: 10.1186/1471-230X-11-22
Ahmed, S. A., Elhefnawy, A. M., Azouz, H. G., Roshdy, Y. S., Ashry, M. H., Ibrahim, A. E., et al. (2020). Study of the gut microbiome profile in children with autism Spectrum disorder: a single tertiary hospital experience. J. Mol. Neurosci. 70, 887–896. doi: 10.1007/s12031-020-01500-3
Al Nabhani, Z., and Eberl, G. (2020). Imprinting of the immune system by the microbiota early in life. Mucosal Immunol. 13, 183–189. doi: 10.1038/s41385-020-0257-y
American Psychiatric Association. (2013). Diagnostic and statistical manual of mental disorders. 5th Edn. Virginia, US: American Psychiatric Association.
Ashe, K., Kelso, W., Farrand, S., Panetta, J., Fazio, T., de Jong, G., et al. (2019). Psychiatric and cognitive aspects of phenylketonuria: the limitations of diet and promise of new treatments. Front. Psychiatry 10:10. doi: 10.3389/fpsyt.2019.00561
Baikie, G., Ravikumara, M., Downs, J., Naseem, N., Wong, K., Percy, A., et al. (2014). Gastrointestinal Dysmotility in Rett syndrome. J. Pediatr. Gastroenterol. Nutr. 58, 237–244. doi: 10.1097/MPG.0000000000000200
Baker, G. B., Bornstein, R. A., Rouget, A. C., Ashton, S. E., van Muyden, J. C., and Coutts, R. T. (1991). Phenylethylaminergic mechanisms in attention-deficit disorder. Biol. Psychiatry 29, 15–22. doi: 10.1016/0006-3223(91)90207-3
Bayir, A. (2015). “The effects of trace element deficiencies on neurological disease and treatment with trace element support” in Bioactive nutraceuticals and dietary supplements in neurological and brain disease. eds. R. R. Watson and V. R. Preedy (Amsterdam, Netherlands: Elsevier), 153–159.
Beckhauser, M. T., Vieira, M. B. M., Iser, B. M., De Luca, G. R., and Masruha, M. R. (2020). Attention deficit disorder with hyperactivity symptoms in early-treated phenylketonuria patients. Iran J. Child. Neurol. 14, 93–103.
Berg, G., Rybakova, D., Fischer, D., Cernava, T., Vergès, M.-C. C., Charles, T., et al. (2020). Microbiome definition re-visited: old concepts and new challenges. Microbiome. 8:875. doi: 10.1186/s40168-020-00875-0
Bergwerff, C. E., Luman, M., Blom, H. J., and Oosterlaan, J. (2016). No tryptophan, tyrosine and phenylalanine abnormalities in children with attention-deficit/hyperactivity disorder. PLoS One 11:e0151100. doi: 10.1371/journal.pone.0151100
Bermudez, B. E. B. V., Oliveira, C. M., Lima Cat, M. N., Magdalena, N. I. R., and Celli, A. (2019). Gastrointestinal disorders in down syndrome. Am. J. Med. Genet. A :61258. doi: 10.1002/ajmg.a.61258
Bioque, M., González-Rodríguez, A., Garcia-Rizo, C., Cobo, J., Monreal, J. A., Usall, J., et al. (2021). Targeting the microbiome-gut-brain axis for improving cognition in schizophrenia and major mood disorders: A narrative review. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 105:110130. doi: 10.1016/j.pnpbp.2020.110130
Borghi, E., Borgo, F., Severgnini, M., Savini, M. N., Casiraghi, M. C., and Vignoli, A. (2017). Rett syndrome: A focus on gut microbiota. Int. J. Mol. Sci. 18, 1–17. doi: 10.3390/ijms18020344
Borghi, E., and Vignoli, A. (2019). Rett syndrome and other neurodevelopmental disorders share common changes in gut microbial community: A descriptive review. Int. J. Mol. Sci. 20:160. doi: 10.3390/ijms20174160
Bornstein, R. A., Baker, G. B., Carroll, A., King, G., Wong, J. T. F., and Douglass, A. B. (1990). Plasma amino acids in attention deficit disorder. Psychiatry Res. 33, 301–306. doi: 10.1016/0165-1781(90)90046-8
Borre, Y. E., O’Keeffe, G. W., Clarke, G., Stanton, C., Dinan, T. G., and Cryan, J. F. (2014). Microbiota and neurodevelopmental windows: implications for brain disorders. Trends Mol. Med. 20, 509–518. doi: 10.1016/j.molmed.2014.05.002
Bourassa, M. W., Alim, I., Bultman, S. J., and Ratan, R. R. (2016). Butyrate, neuroepigenetics and the gut microbiome: can a high fiber diet improve brain health? Neurosci. Lett. 625, 56–63. doi: 10.1016/j.neulet.2016.02.009
Bourne, L., Mandy, W., and Bryant-Waugh, R. (2022). Avoidant/restrictive food intake disorder and severe food selectivity in children and young people with autism: A scoping review. Dev. Med. Child Neurol. 2022:139. doi: 10.1111/dmcn.15139
Brito, I. L., Gurry, T., Zhao, S., Huang, K., Young, S. K., Shea, T. P., et al. (2019). Transmission of human-associated microbiota along family and social networks. Nat. Microbiol. 4, 964–971. doi: 10.1038/s41564-019-0409-6
Burton, C. L., Chhabra, S. R., Swift, S., Baldwin, T. J., Withers, H., Hill, S. J., et al. (2002). The growth response of Escherichia coli to neurotransmitters and related catecholamine drugs requires a functional Enterobactin biosynthesis and uptake system. Infect. Immun. 70, 5913–5923. doi: 10.1128/IAI.70.11.5913-5923.2002
Camilleri, M. (2023). Abnormal gastrointestinal motility is a major factor in explaining symptoms and a potential therapeutic target in patients with disorders of gut–brain interaction. Gut 72, 2372–2380. doi: 10.1136/gutjnl-2023-330542
Carloni, S., Bertocchi, A., Mancinelli, S., Bellini, M., Erreni, M., Borreca, A., et al. (1979). Identification of a choroid plexus vascular barrier closing during intestinal inflammation. Science 374, 439–448. doi: 10.1126/science.abc6108
Chavaria, A., Wang, E. H., and Mills, R. H. (2022). Meta-analysis of the autism gut microbiome identifies factors influencing study discrepancies and machine learning classification. bioRxiv 2020:910. doi: 10.1101/2022.03.18.484910
Coccurello, R., Marrone, M. C., and Maccarrone, M. (2022). The endocannabinoids-microbiota Partnership in gut-Brain Axis Homeostasis: implications for autism Spectrum disorders. Front. Pharmacol. 13:869606. doi: 10.3389/fphar.2022.869606
Cowan, C. S. M., Dinan, T. G., and Cryan, J. F. (2020). Annual research review: critical windows – the microbiota–gut–brain axis in neurocognitive development. J. Child Psychol. Psychiatry 61, 353–371. doi: 10.1111/jcpp.13156
Cryan, J. F., O’Riordan, K. J., Cowan, C. S. M., Sandhu, K. V., Bastiaanssen, T. F. S., Boehme, M., et al. (2019). The microbiota-gut-brain Axis. Physiol. Rev. 99, 1877–2013. doi: 10.1152/physrev.00018.2018
Cryan, J. F., O’Riordan, K. J., Sandhu, K., Peterson, V., and Dinan, T. G. (2020). The gut microbiome in neurological disorders. Lancet Neurol. 19, 179–194. doi: 10.1016/S1474-4422(19)30356-4
Curran, E. A., O’Neill, S. M., Cryan, J. F., Kenny, L. C., Dinan, T. G., Khashan, A. S., et al. (2015). Research review: birth by caesarean section and development of autism spectrum disorder and attention-deficit/hyperactivity disorder: a systematic review and meta-analysis. J. Child Psychol. Psychiatry 56, 500–508. doi: 10.1111/jcpp.12351
De Angelis, M., Piccolo, M., Vannini, L., Siragusa, S., De Giacomo, A., Serrazzanetti, D. I., et al. (2013). Fecal microbiota and metabolome of children with autism and pervasive developmental disorder not otherwise specified. PLoS One 8, 1–18. doi: 10.1371/journal.pone.0076993
Deleu, S., Machiels, K., Raes, J., Verbeke, K., and Vermeire, S. (2021). Short chain fatty acids and its producing organisms: an overlooked therapy for IBD? EBioMedicine 66:103293. doi: 10.1016/j.ebiom.2021.103293
Derrien, M., Alvarez, A.-S., and de Vos, W. M. (2019). The gut microbiota in the first decade of life. Trends Microbiol. 27, 997–1010. doi: 10.1016/j.tim.2019.08.001
Erny, D., Dokalis, N., Mezö, C., Castoldi, A., Mossad, O., Staszewski, O., et al. (2021). Microbiota-derived acetate enables the metabolic fitness of the brain innate immune system during health and disease. Cell Metab. 33, 2260–2276.e7. doi: 10.1016/j.cmet.2021.10.010
Finegold, S. M. (2011). Desulfovibrio species are potentially important in regressive autism. Med. Hypotheses 77, 270–274. doi: 10.1016/j.mehy.2011.04.032
Finegold, S. M., Dowd, S. E., Gontcharova, V., Liu, C., Henley, K. E., Wolcott, R. D., et al. (2010). Pyrosequencing study of fecal microflora of autistic and control children. Anaerobe 16, 444–453. doi: 10.1016/j.anaerobe.2010.06.008
Finegold, S. M., Molitoris, D., Song, Y., Liu, C., Vaisanen, M., Bolte, E., et al. (2002). Gastrointestinal microflora studies in late-onset autism. Clin. Infect. Dis. 35, S6–S16. doi: 10.1086/341914
Finegold, S. M., Summanen, P. H., Downes, J., Corbett, K., and Komoriya, T. (2017). Detection of Clostridium perfringens toxin genes in the gut microbiota of autistic children. Anaerobe 45, 133–137. doi: 10.1016/j.anaerobe.2017.02.008
Fung, T. C., Olson, C. A., and Hsiao, E. Y. (2017). Interactions between the microbiota, immune and nervous systems in health and disease. Nat. Neurosci. 20, 145–155. doi: 10.1038/nn.4476
Ghosh, P., Sarkar, K., Bhaduri, N., Ray, A., Sarkar, K., Sinha, S., et al. (2013). Catecholaminergic gene variants: contribution in ADHD and associated comorbid attributes in the eastern Indian Probands. Biomed. Res. Int. 2013, 1–12. doi: 10.1155/2013/918410
Gondalia, S. V., Palombo, E. A., Knowles, S. R., Cox, S. B., Meyer, D., and Austin, D. W. (2012). Molecular characterisation of gastrointestinal microbiota of children with autism (with and without gastrointestinal dysfunction) and their Neurotypical siblings. Autism Res. 5, 419–427. doi: 10.1002/aur.1253
Guimarães, A. P. M., Zeni, C., Polanczyk, G. V., Genro, J. P., Roman, T., Rohde, L. A., et al. (2007). Serotonin genes and attention deficit/hyperactivity disorder in a Brazilian sample: preferential transmission of the HTR2A 452His allele to affected boys. Am. J. Med. Genet. B Neuropsychiatr. Genet. 144B, 69–73. doi: 10.1002/ajmg.b.30400
Haglund, N. G. S., and Källén, K. B. M. (2011). Risk factors for autism and Asperger syndrome. Autism 15, 163–183. doi: 10.1177/1362361309353614
Hamady, M., and Knight, R. (2009). Microbial community profiling for human microbiome projects: tools, techniques, and challenges. Genome Res. 19, 1141–1152. doi: 10.1101/gr.085464.108
Hamilton, M. J., Weingarden, A. R., Sadowsky, M. J., and Khoruts, A. (2012). Standardized frozen preparation for transplantation of fecal microbiota for recurrent Clostridium difficile infection. Am. J. Gastroenterol. 107, 761–767. doi: 10.1038/ajg.2011.482
Hare, M. M., Dick, A. S., and Graziano, P. A. (2022). Adverse childhood experiences predict neurite density differences in young children with and without attention deficit hyperactivity disorder. Dev. Psychobiol. 64:234. doi: 10.1002/dev.22234
Harris, J. (2018). Leo Kanner and autism: a 75-year perspective. Int. Rev. Psychiatry 30, 3–17. doi: 10.1080/09540261.2018.1455646
Harrison, J. K., Reid, J., Quinn, T. J., and Shenkin, S. D. (2017). Using quality assessment tools to critically appraise ageing research: a guide for clinicians. Age Ageing 46, 359–365. doi: 10.1093/ageing/afw223
Hill, L., Popov, J., Figueiredo, M., Caputi, V., Hartung, E., Moshkovich, M., et al. (2021). Protocol for a systematic review on the role of the gut microbiome in paediatric neurological disorders. Acta Neuropsychiatr. 33, 211–216. doi: 10.1017/neu.2021.8
Ho, L. K. H., Tong, V. J. W., Syn, N., Nagarajan, N., Tham, E. H., Tay, S. K., et al. (2020). Gut microbiota changes in children with autism spectrum disorder: a systematic review. Gut Pathog. 12:6. doi: 10.1186/s13099-020-0346-1
Hoffbuhr, K. C., Moses, L. M., Jerdonek, M. A., Naidu, S., and Hoffman, E. P. (2002). Associations betweenmeCP2 mutations, x-chromosome inactivation, and phenotype. Ment. Retard. Dev. Disabil. Res. Rev. 8, 99–105. doi: 10.1002/mrdd.10026
Hughes, H. K., and Ashwood, P. (2018). Anti-Candida albicans IgG antibodies in children with autism spectrum disorders. Front. Psych. 9, 1–5. doi: 10.3389/fpsyt.2018.00627
Jiang, H., Zhou, Y., Zhou, G., Li, Y., Yuan, J., Li, X., et al. (2018). Gut microbiota profiles in treatment-naïve children with attention deficit hyperactivity disorder. Behav. Brain Res. 347, 408–413. doi: 10.1016/j.bbr.2018.03.036
Jovel, J., Patterson, J., Wang, W., Hotte, N., O'Keefe, S., Mitchel, T., et al. (2016). Characterization of the gut microbiome using 16S or shotgun metagenomics. Front. Virol. 7:459. doi: 10.3389/fmicb.2016.00459
Kanehisa, M. (2000). KEGG: Kyoto encyclopedia of genes and genomes. Nucleic Acids Res. 28, 27–30. doi: 10.1093/nar/28.1.27
Kang, D. W., Adams, J. B., Gregory, A. C., Borody, T., Chittick, L., Fasano, A., et al. (2017). Microbiota transfer therapy alters gut ecosystem and improves gastrointestinal and autism symptoms: an open-label study. Microbiome 5, 1–16. doi: 10.1186/s40168-016-0225-7
Kang, D.-W., Ilhan, Z. E., Isern, N. G., Hoyt, D. W., Howsmon, D. P., Shaffer, M., et al. (2018). Differences in fecal microbial metabolites and microbiota of children with autism spectrum disorders. Anaerobe 49, 121–131. doi: 10.1016/j.anaerobe.2017.12.007
Kang, D.-W., Park, J. G., Ilhan, Z. E., Wallstrom, G., LaBaer, J., Adams, J. B., et al. (2013). Reduced incidence of Prevotella and other fermenters in intestinal microflora of autistic children. PLoS One 8:322. doi: 10.1371/journal.pone.0068322
Kantarcioglu, A. S., Kiraz, N., and Aydin, A. (2016). Microbiota–gut–brain Axis: yeast species isolated from stool samples of children with suspected or diagnosed autism Spectrum disorders and in vitro susceptibility against nystatin and fluconazole. Mycopathologia 181, 1–7. doi: 10.1007/s11046-015-9949-3
Kedem, S., Yust-Katz, S., Carter, D., Levi, Z., Kedem, R., Dickstein, A., et al. (2020). Attention deficit hyperactivity disorder and gastrointestinal morbidity in a large cohort of young adults. World J. Gastroenterol. 26, 6626–6637. doi: 10.3748/wjg.v26.i42.6626
Kochalska, K., Oakden, W., Słowik, T., Chudzik, A., Pankowska, A., Łazorczyk, A., et al. (2020). Dietary supplementation with Lactobacillus rhamnosus JB-1 restores brain neurochemical balance and mitigates the progression of mood disorder in a rat model of chronic unpredictable mild stress. Nutr. Res. 82, 44–57. doi: 10.1016/j.nutres.2020.06.019
Kushak, R. I., Winter, H. S., Buie, T. M., Cox, S. B., Phillips, C. D., and Ward, N. L. (2017). Analysis of the duodenal microbiome in autistic individuals: association with carbohydrate digestion. J. Pediatr. Gastroenterol. Nutr. 64, e110–e116. doi: 10.1097/MPG.0000000000001458
Lange, K. W., Reichl, S., Lange, K. M., Tucha, L., and Tucha, O. (2010). The history of attention deficit hyperactivity disorder. Atten Defic Hyperact Disord 2, 241–255. doi: 10.1007/s12402-010-0045-8
Laue, H. E., Karagas, M. R., Coker, M. O., Bellinger, D. C., Baker, E. R., Korrick, S. A., et al. (2021). Sex-specific relationships of the infant microbiome and early-childhood behavioral outcomes. Pediatr. Res. 92, 580–591. doi: 10.1038/s41390-021-01785-z
Liberati, A., Altman, D. G., Tetzlaff, J., Mulrow, C., Gøtzsche, P. C., Ioannidis, J. P. A., et al. (2009). The PRISMA statement for reporting systematic reviews and meta-analyses of studies that evaluate health care interventions: explanation and elaboration. PLoS Med. 6:100. doi: 10.1371/journal.pmed.1000100
Liu, S., Li, E., Sun, Z., Fu, D., Duan, G., Jiang, M., et al. (2019). Altered gut microbiota and short chain fatty acids in Chinese children with autism spectrum disorder. Sci Rep 9, 1–9. doi: 10.1038/s41598-018-36430-z
Liu, L., and Zhu, G. (2018). Gut–brain Axis and mood disorder. Front. Psych. 9:223. doi: 10.3389/fpsyt.2018.00223
Long, H. A., French, D. P., and Brooks, J. M. (2020). Optimising the value of the critical appraisal skills programme (CASP) tool for quality appraisal in qualitative evidence synthesis. Res. Methods Med. Health Sci. 1, 31–42. doi: 10.1177/2632084320947559
Lopatina, O. L., Panina, Y. A., Malinovskaya, N. A., and Salmina, A. B. (2021). Early life stress and brain plasticity: from molecular alterations to aberrant memory and behavior. Rev. Neurosci. 32, 131–142. doi: 10.1515/revneuro-2020-0077
Luczynski, P., McVey Neufeld, K.-A., Oriach, C. S., Clarke, G., Dinan, T. G., and Cryan, J. F. (2016). Growing up in a bubble: using germ-free animals to assess the influence of the gut microbiota on brain and behavior. Int. J. Neuropsychopharmacol. 19:pyw020. doi: 10.1093/ijnp/pyw020
Luna, R. A., Oezguen, N., Balderas, M., Venkatachalam, A., Runge, J. K., Versalovic, J., et al. (2017). Distinct microbiome-Neuroimmune signatures correlate with functional abdominal pain in children with autism Spectrum disorder. Cell. Mol. Gastroenterol. Hepatol. 3, 218–230. doi: 10.1016/j.jcmgh.2016.11.008
MacFabe, D. F., Rodriguez-, K., Hoffman, J. E., Franklin, A. E., Mohammad-A, Y., Taylor, A. R., et al. (2008). A novel rodent model of autism: intraventricular infusions of propionic acid increase locomotor activity and induce Neuroinflammation and oxidative stress in discrete regions of adult rat brain. Am. J. Biochem. Biotechnol. 4, 146–166. doi: 10.3844/ajbbsp.2008.146.166
Makris, G., Eleftheriades, A., and Pervanidou, P. (2022). Early life stress, hormones, and neurodevelopmental disorders. Horm. Res. Paediatr 96, 17–24. doi: 10.1159/000523942
Margolis, K. G., Cryan, J. F., and Mayer, E. A. (2021). The microbiota-gut-brain Axis: from motility to mood. Gastroenterology 160, 1486–1501. doi: 10.1053/j.gastro.2020.10.066
Marrone, M. C., and Coccurello, R. (2019). Dietary fatty acids and microbiota-brain communication in neuropsychiatric diseases. Biomol. Ther. 10:12. doi: 10.3390/biom10010012
McVey Neufeld, K.-A., Strain, C. R., Pusceddu, M. M., Waworuntu, R. V., Manurung, S., Gross, G., et al. (2020). Lactobacillus rhamnosus GG soluble mediators ameliorate early life stress-induced visceral hypersensitivity and changes in spinal cord gene expression. Neuronal. Signals 4:7. doi: 10.1042/NS20200007
Ming, X., Brimacombe, M., Chaaban, J., Zimmerman-Bier, B., and Wagner, G. C. (2008). Autism Spectrum disorders: concurrent clinical disorders. J. Child Neurol. 23, 6–13. doi: 10.1177/0883073807307102
Morais, L. H., Schreiber, H. L., and Mazmanian, S. K. (2021). The gut microbiota–brain axis in behaviour and brain disorders. Nat. Rev. Microbiol. 19, 241–255. doi: 10.1038/s41579-020-00460-0
Morton, J. T., Jin, D., Mills, R. H., Shao, Y., Rahman, G., McDonald, D., et al. (2022). Multi-omic analysis along the gut-brain axis points to a functional architecture of autism. bioRxiv 2022:2050. doi: 10.1101/2022.02.25.482050
Motil, K. J., Caeg, E., Barrish, J. O., Geerts, S., Lane, J. B., Percy, A. K., et al. (2012). Gastrointestinal and nutritional problems occur frequently throughout life in girls and women with Rett syndrome. J. Pediatr. Gastroenterol. Nutr. 55, 292–298. doi: 10.1097/MPG.0b013e31824b6159
Navratna, V., and Gouaux, E. (2019). Insights into the mechanism and pharmacology of neurotransmitter sodium symporters. Curr. Opin. Struct. Biol. 54, 161–170. doi: 10.1016/j.sbi.2019.03.011
Nichols, V. P., Toye, F., Eldabe, S., Sandhu, H. K., Underwood, M., and Seers, K. (2020). Experiences of people taking opioid medication for chronic non-malignant pain: a qualitative evidence synthesis using meta-ethnography. BMJ Open 10:e032988. doi: 10.1136/bmjopen-2019-032988
Nzakizwanayo, J., Dedi, C., Standen, G., Macfarlane, W. M., Patel, B. A., and Jones, B. V. (2015). Escherichia coli Nissle 1917 enhances bioavailability of serotonin in gut tissues through modulation of synthesis and clearance. Sci. Rep. 5:17324. doi: 10.1038/srep17324
O’Riordan, K. J., Collins, M. K., Moloney, G. M., Knox, E. G., Aburto, M. R., Fülling, C., et al. (2022). Short chain fatty acids: microbial metabolites for gut-brain axis signalling. Mol. Cell. Endocrinol. 546:111572. doi: 10.1016/j.mce.2022.111572
Obremskey, W. T., Pappas, N., Attallah-Wasif, E., Tornetta, P., and Bhandari, M. (2005). Level of evidence in orthopaedic journals. J. Bone Joint Surg. Am. 87, 2632–2638. doi: 10.2106/JBJS.E.00370
Orsmond, G. I., and Seltzer, M. M. (2007). Siblings of individuals with autism spectrum disorders across the life course. Ment. Retard. Dev. Disabil. Res. Rev. 13, 313–320. doi: 10.1002/mrdd.20171
Pärtty, A., Kalliomäki, M., Wacklin, P., Salminen, S., and Isolauri, E. (2015). A possible link between early probiotic intervention and the risk of neuropsychiatric disorders later in childhood: A randomized trial. Pediatr. Res. 77, 823–828. doi: 10.1038/pr.2015.51
Plöger, S., Stumpff, F., Penner, G. B., Schulzke, J.-D., Gäbel, G., Martens, H., et al. (2012). Microbial butyrate and its role for barrier function in the gastrointestinal tract. Ann. N. Y. Acad. Sci. 1258, 52–59. doi: 10.1111/j.1749-6632.2012.06553.x
Polanczyk, G. V., Willcutt, E. G., Salum, G. A., Kieling, C., and Rohde, L. A. (2014). ADHD prevalence estimates across three decades: an updated systematic review and meta-regression analysis. Int. J. Epidemiol. 43, 434–442. doi: 10.1093/ije/dyt261
Popov, J., Caputi, V., Nandeesha, N., Rodriguez, D. A., and Pai, N. (2021). Microbiota-immune interactions in ulcerative colitis and colitis associated Cancer and emerging microbiota-based therapies. Int. J. Mol. Sci. 22:11365. doi: 10.3390/ijms222111365
Prehn-Kristensen, A., Zimmermann, A., Tittmann, L., Lieb, W., Schreiber, S., Baving, L., et al. (2018). Reduced microbiome alpha diversity in young patients with ADHD. PLoS One 13:e0200728. doi: 10.1371/journal.pone.0200728
Pulikkan, J., Maji, A., Dhakan, D. B., Saxena, R., Mohan, B., Anto, M. M., et al. (2018). Gut microbial Dysbiosis in Indian children with autism Spectrum disorders. Microb. Ecol. 76, 1102–1114. doi: 10.1007/s00248-018-1176-2
Ratsika, A., Codagnone, M. C., O’Mahony, S., Stanton, C., and Cryan, J. F. (2021). Priming for life: early life nutrition and the microbiota-gut-brain Axis. Nutrients 13:423. doi: 10.3390/nu13020423
Ratsika, A., Cruz Pereira, J. S., Lynch, C. M. K., Clarke, G., and Cryan, J. F. (2023). Microbiota-immune-brain interactions: A lifespan perspective. Curr. Opin. Neurobiol. 78:102652. doi: 10.1016/j.conb.2022.102652
Richarte, V., Sánchez-Mora, C., Corrales, M., Fadeuilhe, C., Vilar-Ribó, L., Arribas, L., et al. (2021). Gut microbiota signature in treatment-naïve attention-deficit/hyperactivity disorder. Transl. Psychiatry 11:382. doi: 10.1038/s41398-021-01504-6
Rose, D. R., Yang, H., Serena, G., Sturgeon, C., Ma, B., Careaga, M., et al. (2018). Differential immune responses and microbiota profiles in children with autism spectrum disorders and co-morbid gastrointestinal symptoms. Brain Behav. Immun. 70, 354–368. doi: 10.1016/j.bbi.2018.03.025
Saili, K. S., Zurlinden, T. J., Schwab, A. J., Silvin, A., Baker, N. C., Hunter, E. S., et al. (2017). Blood-brain barrier development: systems modeling and predictive toxicology. Birth Defects Res. 109, 1680–1710. doi: 10.1002/bdr2.1180
Salvat, H., Mohammadi, M. N., Molavi, P., Mostafavi, S. A., Rostami, R., and Salehinejad, M. A. (2022). Nutrient intake, dietary patterns, and anthropometric variables of children with ADHD in comparison to healthy controls: a case-control study. BMC Pediatr. 22:70. doi: 10.1186/s12887-022-03123-6
Santos, J. X., Rasga, C., Marques, A. R., Martiniano, H., Asif, M., Vilela, J., et al. (2022). A role for gene-environment interactions in autism Spectrum disorder is supported by variants in genes regulating the effects of exposure to xenobiotics. Front. Neurosci. 16:315. doi: 10.3389/fnins.2022.862315
Severance, E. G., Prandovszky, E., Castiglione, J., and Yolken, R. H. (2015). Gastroenterology issues in schizophrenia: why the gut matters. Curr. Psychiatry Rep. 17:27. doi: 10.1007/s11920-015-0574-0
Shaaban, S. Y., El Gendy, Y. G., Mehanna, N. S., El-Senousy, W. M., El-Feki, H. S. A., Saad, K., et al. (2018). The role of probiotics in children with autism spectrum disorder: A prospective, open-label study. Nutr. Neurosci. 21, 676–681. doi: 10.1080/1028415X.2017.1347746
Shamseer, L., Moher, D., Clarke, M., Ghersi, D., Liberati, A., Petticrew, M., et al. (2015). Preferred reporting items for systematic review and meta-analysis protocols (PRISMA-P) 2015: elaboration and explanation. BMJ 349:g7647. doi: 10.1136/bmj.g7647
Slykerman, R. F., Thompson, J., Waldie, K. E., Murphy, R., Wall, C., and Mitchell, E. A. (2017). Antibiotics in the first year of life and subsequent neurocognitive outcomes. Acta Paediatr. 106, 87–94. doi: 10.1111/apa.13613
Socała, K., Doboszewska, U., Szopa, A., Serefko, A., Włodarczyk, M., Zielińska, A., et al. (2021). The role of microbiota-gut-brain axis in neuropsychiatric and neurological disorders. Pharmacol. Res. 172:105840. doi: 10.1016/j.phrs.2021.105840
Son, J. S., Zheng, L. J., Rowehl, L. M., Tian, X., Zhang, Y., Zhu, W., et al. (2015). Comparison of fecal microbiota in children with autism Spectrum disorders and Neurotypical siblings in the Simons simplex collection. PLoS One :e0137725. doi: 10.1371/journal.pone.0137725
Sprich, S., Biederman, J., Crawford, M. H., Mundy, E., and Faraone, S. V. (2000). Adoptive and biological families of children and adolescents with ADHD. J. Am. Acad. Child Adolesc. Psychiatry 39, 1432–1437. doi: 10.1097/00004583-200011000-00018
Stevens, A. J., Purcell, R. V., Darling, K. A., Eggleston, M. J. F. F., Kennedy, M. A., and Rucklidge, J. J. (2019). Human gut microbiome changes during a 10 week randomised control trial for micronutrient supplementation in children with attention deficit hyperactivity disorder. Sci. Rep. 9:146. doi: 10.1038/s41598-019-46146-3
Stevenson, M., and McNaughton, N. (2013). A comparison of phenylketonuria with attention deficit hyperactivity disorder: do markedly different aetiologies deliver common phenotypes? Brain Res. Bull. 99, 63–83. doi: 10.1016/j.brainresbull.2013.10.003
Strati, F., Cavalieri, D., Albanese, D., De Felice, C., Donati, C., Hayek, J., et al. (2016). Altered gut microbiota in Rett syndrome. Microbiome 4:41. doi: 10.1186/s40168-016-0185-y
Strati, F., Cavalieri, D., Albanese, D., De Felice, C., Donati, C., Hayek, J., et al. (2017). New evidences on the altered gut microbiota in autism spectrum disorders. Microbiome 5, 1–11. doi: 10.1186/s40168-017-0242-1
Szopinska-Tokov, J., Dam, S., Naaijen, J., Konstanti, P., Rommelse, N., Belzer, C., et al. (2020). Investigating the gut microbiota composition of individuals with attention-deficit/hyperactivity disorder and association with symptoms. Microorganisms 8:406. doi: 10.3390/microorganisms8030406
Szopinska-Tokov, J., Dam, S., Naaijen, J., Konstanti, P., Rommelse, N., Belzer, C., et al. (2021). Correction: Szopinska-Tokov et al. investigating the gut microbiota composition of individuals with attention-deficit/hyperactivity disorder and association with symptoms. Microorganisms 2020, 8, 406. Microorganisms 9:1358. doi: 10.3390/microorganisms9071358
Tanaka, M., and Nakayama, J. (2017). Development of the gut microbiota in infancy and its impact on health in later life. Allergol. Int. 66, 515–522. doi: 10.1016/j.alit.2017.07.010
Tarui, T., Rasool, A., and O’Tierney-Ginn, P. (2022). How the placenta-brain lipid axis impacts the nutritional origin of child neurodevelopmental disorders: focus on attention deficit hyperactivity disorder and autism spectrum disorder. Exp. Neurol. 347:113910. doi: 10.1016/j.expneurol.2021.113910
Tateno, M., Kikuchi, S., Uehara, K., Fukita, K., Uchida, N., Sasaki, R., et al. (2011). Pervasive developmental disorders and autism Spectrum disorders: are these disorders one and the same? Psychiatry Investig. 8, 67–70. doi: 10.4306/pi.2011.8.1.67
Tett, A., Pasolli, E., Masetti, G., Ercolini, D., and Segata, N. (2021). Prevotella diversity, niches and interactions with the human host. Nat. Rev. Microbiol. 19, 585–599. doi: 10.1038/s41579-021-00559-y
Thapa, S., Venkatachalam, A., Khan, N., Naqvi, M., Balderas, M., Runge, J. K., et al. (2021). Assessment of the gut bacterial microbiome and metabolome of girls and women with Rett syndrome. PLoS One :e0251231. doi: 10.1371/journal.pone.0251231
Thapar, A., Cooper, M., Eyre, O., and Langley, K. (2013). Practitioner review: what have we learnt about the causes of ADHD? J. Child Psychol. Psychiatry 54, 3–16. doi: 10.1111/j.1469-7610.2012.02611.x
Thomas, R., Sanders, S., Doust, J., Beller, E., and Glasziou, P. (2015). Prevalence of attention-deficit/hyperactivity disorder: A systematic review and Meta-analysis. Pediatrics 135, e994–e1001. doi: 10.1542/peds.2014-3482
Tindall, B. J. (2019). The names Hungateiclostridium Zhang et al. 2018, Hungateiclostridium thermocellum (Viljoen et al. 1926) Zhang et al. 2018, Hungateiclostridium cellulolyticum (Patel et al. 1980) Zhang et al. 2018, Hungateiclostridium aldrichii (Yang et al. 1990) Zhang et al. 2018, Hungateiclostridium alkalicellulosi (Zhilina et al. 2006) Zhang et al. 2018, Hungateiclostridium clariflavum (Shiratori et al. 2009) Zhang et al. 2018, Hungateiclostridium straminisolvens (Kato et al. 2004) Zhang et al. 2018 and Hungateiclostridium saccincola (Koeck et al. 2016) Zhang et al. 2018 contravene rule 51b of the international code of nomenclature of prokaryotes and require replacement names in the genus Acetivibrio Patel et al. 1980. Int. J. Syst. Bacteriol. 69, 3927–3932. doi: 10.1099/ijsem.0.003685
Tomova, A., Husarova, V., Lakatosova, S., Bakos, J., Vlkova, B., Babinska, K., et al. (2015). Gastrointestinal microbiota in children with autism in Slovakia. Physiol. Behav. 138, 179–187. doi: 10.1016/j.physbeh.2014.10.033
Wade, M., Prime, H., and Madigan, S. (2015). Using sibling designs to understand neurodevelopmental disorders: from genes and environments to prevention programming. Biomed. Res. Int. 2015, 1–16. doi: 10.1155/2015/672784
Wan, L., Ge, W.-R., Zhang, S., Sun, Y.-L., Wang, B., and Yang, G. (2020). Case-control study of the effects of gut microbiota composition on neurotransmitter metabolic pathways in children with attention deficit hyperactivity disorder. Front. Neurosci. 14:127. doi: 10.3389/fnins.2020.00127
Wang, L., Christophersen, C. T., Sorich, M. J., Gerber, J. P., Angley, M. T., and Conlon, M. A. (2012). Elevated fecal short chain fatty acid and Ammonia concentrations in children with autism Spectrum disorder. Dig. Dis. Sci. 57, 2096–2102. doi: 10.1007/s10620-012-2167-7
Wang, N., Gao, X., Zhang, Z., and Yang, L. (2022). Composition of the gut microbiota in attention deficit hyperactivity disorder: A systematic review and Meta-analysis. Front. Endocrinol. 13:838941. doi: 10.3389/fendo.2022.838941
Wang, R., Tang, R., Li, B., Ma, X., Schnabl, B., and Tilg, H. (2021). Gut microbiome, liver immunology, and liver diseases. Cell. Mol. Immunol. 18, 4–17. doi: 10.1038/s41423-020-00592-6
Wang, L. J., Yang, C. Y., Chou, W. J., Lee, M. J., Chou, M. C., Kuo, H. C., et al. (2020). Gut microbiota and dietary patterns in children with attention-deficit/hyperactivity disorder. Eur. Child Adolesc. Psychiatry 29, 287–297. doi: 10.1007/s00787-019-01352-2
Wong, L. C., Chen, Y., Tsai, S., Lin, Y., Hsu, C., Wang, H., et al. (2021). Dietary intake and growth deficits in Rett syndrome—A cross-section study. Autism Res. 14, 1512–1521. doi: 10.1002/aur.2508
Yap, C. X., Henders, A. K., Alvares, G. A., Wood, D. L. A., Krause, L., Tyson, G. W., et al. (2021). Autism-related dietary preferences mediate autism-gut microbiome associations. Cell 184, 5916–5931. doi: 10.1016/j.cell.2021.10.015
Zhai, Q., Cen, S., Jiang, J., Zhao, J., Zhang, H., and Chen, W. (2019). Disturbance of trace element and gut microbiota profiles as indicators of autism spectrum disorder: A pilot study of Chinese children. Environ. Res. 171, 501–509. doi: 10.1016/j.envres.2019.01.060
Zhang, M., Ma, W., Zhang, J., He, Y., and Wang, J. (2018). Analysis of gut microbiota profiles and microbe-disease associations in children with autism spectrum disorders in China. Sci. Rep. 8, 1–9. doi: 10.1038/s41598-018-32219-2
Zhou, B., Jin, G., Pang, X., Mo, Q., Bao, J., Liu, T., et al. (2022). Lactobacillus rhamnosus GG colonization in early life regulates gut-brain axis and relieves anxiety-like behavior in adulthood. Pharmacol. Res. 177:106090. doi: 10.1016/j.phrs.2022.106090
Zhou, P., Manoil, D., Belibasakis, G. N., and Kotsakis, G. A. (2021). Veillonellae: beyond bridging species in Oral biofilm ecology. Front. Oral Health :774115:2. doi: 10.3389/froh.2021.774115
Key concepts in microbiome classification and measurement.
Microbiota – Term used to describe all bacteria, fungi, and viruses in a given ecological space. Individual microbiota is commonly called, “taxon” or “taxa” (plural).
Microbiome – Term used to describe genome of all bacteria, fungi, and viruses in a given ecological space.
Microbial Culture – Technique used to detect specific microbiota using selective, known culture conditions.
16S rRNA Sequencing – Technique used to identify bacteria by PCR amplifying specific regions (i.e., V4) of the bacterial DNA, and matching amplified regions against a known reference library of “amplicons.” Resolution is often limited to genus level of taxonomic classification.
Shotgun Metagenomics – Technique used to detect specific organisms by sequencing all regions of the microbial genome, and matching available genes against known genomic libraries. Offers greater specificity to species and strain levels of taxonomic classification.
Functional Metagenomics – Term used to describe a group of techniques to determine the “functional capacity” of a microbiome. Includes: metatranscriptomics (measuring transcribed mRNA in a sample), metaproteomics (measuring available proteins in a sample), and metabolomics (measuring non-protein small molecules in a sample). Each technique provides analyses of different metabolic products produced by a microbial community and offers progressively deeper insights into how the microbiota interacts with their host.
Inferred Metagenomics – Similar to functional metagenomics, computational technique that uses the results of 16S rRNA Sequencing or shotgun metagenomics data to predict the function of a microbiota. Identified taxa are evaluated against a known library of genes for each taxon to make an indirect prediction of their functional capacity. While significantly cheaper than measures of functional metagenomics, this approach has less accuracy.
α-diversity – Term used to describe microbial diversity within a given ecological space. Techniques include describing the number of species found within a particular environment or describing the balance of species within the environment (or “evenness”). Examples of common α-diversity measures include Shannon, Simpson, and Chao1 indices.
β-diversity – Term used to describe diversity between different ecological spaces. Techniques include describing differences in number of species between two (or more) environments or describing the balance of species within the environments. β-diversity is commonly expressed visually as a principal component analysis (PCA) plot, where taxa are plotted across two to three spatial dimensions to visualize their distribution relative to one another.
Keywords: autism spectrum disorder, attention deficit hyperactivity disorder, Rett syndrome, pediatric, children, microbiota, metabolites, neurodevelopment
Citation: Caputi V, Hill L, Figueiredo M, Popov J, Hartung E, Margolis KG, Baskaran K, Joharapurkar P, Moshkovich M and Pai N (2024) Functional contribution of the intestinal microbiome in autism spectrum disorder, attention deficit hyperactivity disorder, and Rett syndrome: a systematic review of pediatric and adult studies. Front. Neurosci. 18:1341656. doi: 10.3389/fnins.2024.1341656
Edited by:
Paul Forsythe, University of Alberta, CanadaReviewed by:
Roberto Coccurello, National Research Council (CNR), ItalyCopyright © 2024 Caputi, Hill, Figueiredo, Popov, Hartung, Margolis, Baskaran, Joharapurkar, Moshkovich and Pai. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Valentina Caputi, dmNhcHV0aUB1YXJrLmVkdQ==
†These authors have contributed equally to this work and share first authorship
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.