
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Neurosci., 11 January 2024
Sec. Translational Neuroscience
Volume 17 - 2023 | https://doi.org/10.3389/fnins.2023.1286665
This article is part of the Research TopicTranslational Applications of NeuroimagingView all 12 articles
 Apurva Watve1
Apurva Watve1 Amelie Haugg2
Amelie Haugg2 Nada Frei2
Nada Frei2 Yury Koush3
Yury Koush3 David Willinger2,4,5
David Willinger2,4,5 Annette Beatrix Bruehl1,6
Annette Beatrix Bruehl1,6 Philipp Stämpfli1
Philipp Stämpfli1 Frank Scharnowski1,5,7,8
Frank Scharnowski1,5,7,8 Ronald Sladky1,9*
Ronald Sladky1,9*Introduction: Maladaptive functioning of the amygdala has been associated with impaired emotion regulation in affective disorders. Recent advances in real-time fMRI neurofeedback have successfully demonstrated the modulation of amygdala activity in healthy and psychiatric populations. In contrast to an abstract feedback representation applied in standard neurofeedback designs, we proposed a novel neurofeedback paradigm using naturalistic stimuli like human emotional faces as the feedback display where change in the facial expression intensity (from neutral to happy or from fearful to neutral) was coupled with the participant’s ongoing bilateral amygdala activity.
Methods: The feasibility of this experimental approach was tested on 64 healthy participants who completed a single training session with four neurofeedback runs. Participants were assigned to one of the four experimental groups (n = 16 per group), i.e., happy-up, happy-down, fear-up, fear-down. Depending on the group assignment, they were either instructed to “try to make the face happier” by upregulating (happy-up) or downregulating (happy-down) the amygdala or to “try to make the face less fearful” by upregulating (fear-up) or downregulating (fear-down) the amygdala feedback signal.
Results: Linear mixed effect analyses revealed significant amygdala activity changes in the fear condition, specifically in the fear-down group with significant amygdala downregulation in the last two neurofeedback runs as compared to the first run. The happy-up and happy-down groups did not show significant amygdala activity changes over four runs. We did not observe significant improvement in the questionnaire scores and subsequent behavior. Furthermore, task-dependent effective connectivity changes between the amygdala, fusiform face area (FFA), and the medial orbitofrontal cortex (mOFC) were examined using dynamic causal modeling. The effective connectivity between FFA and the amygdala was significantly increased in the happy-up group (facilitatory effect) and decreased in the fear-down group. Notably, the amygdala was downregulated through an inhibitory mechanism mediated by mOFC during the first training run.
Discussion: In this feasibility study, we intended to address key neurofeedback processes like naturalistic facial stimuli, participant engagement in the task, bidirectional regulation, task congruence, and their influence on learning success. It demonstrated that such a versatile emotional face feedback paradigm can be tailored to target biased emotion processing in affective disorders.
Worldwide, nearly a billion people suffer from mental health problems in some form of mood disorder that significantly affects their quality of life (World Health Organization, 2022). Among the most common mood disorders are affective disorders, such as major depressive disorder (MDD), and anxiety disorder (AD), which have the highest global prevalence rate and cause a social and economic burden (Steel et al., 2014). Epidemiological data from Europe suggest that AD (14.0%) has the highest 12-month prevalence rate with MDD (6.9%) in the third position (Wittchen et al., 2011). They have become even more prominent with the emergence of the COVID-19 pandemic in 2020 (Santomauro et al., 2021). In addition to disrupting emotional well-being and behavior, affective disorders also affect somatic health (Momen et al., 2020), which can lead to reduced life expectancy (Nordentoft et al., 2013). They are substantially associated with high treatment costs, disability, and chronicity (Cuijpers et al., 2012; Baxter et al., 2014). Considerable financial and scientific resources have been invested in the development of efficient and cost-effective treatments for MDD and AD in the fields of neuropsychopharmacology, brain stimulation, and psychotherapy. However, further research into different treatment approaches for the improvement of AD and MDD symptoms is warranted due to the significant number of non-responders.
In terms of symptomatology, affective disorders are mainly characterized by affective biases, dysfunctional self-belief (Hofmann et al., 2012), and deficits in emotion regulation (Joormann and Siemer, 2014). Established research suggests that the maladaptive functioning of cortico-limbic regions, particularly the amygdala, is involved in the development and propagation of these symptoms (Disner et al., 2011). Altered amygdala responses to affective stimuli trigger emotion dysregulation, manifesting hallmark MDD symptoms such as anhedonia, i.e., diminished positive affectivity (McMakin et al., 2012), and amplified negative emotional responses, resulting in dysphoria, i.e., feelings of fear, anxiety, distress, etc. (Siegle et al., 2007). Neural signatures of MDD reveal that these affective biases are associated with decreased amygdala activity in response to positive stimuli and increased reactivity to negative stimuli (Suslow et al., 2010; Victor et al., 2010; Young et al., 2016). Reduced amygdala reactivity to happy faces in MDD patients correlates with higher anhedonia scores and poorer or inappropriate salience attributions to positive environmental cues (Victor et al., 2010). Such impaired processing of positive affect is also associated with an increased bias toward sad faces, which correlates with MDD severity (Suslow et al., 2010).
In anxiety disorders such as posttraumatic stress disorder (PTSD) and social anxiety disorder (SAD), attentional and emotion-processing biases lead to exaggerated responses to negative stimuli that are perceived as threatening. Fear is one of the core symptoms of anxiety disorders, e.g., fear of negative social evaluation and speech anxiety in SAD (Tillfors et al., 2002; Davies et al., 2017). Studies suggest that hyperactivation of the amygdala is associated with the biased processing of negative emotions that trigger fear responses in individuals with AD. There is evidence that amygdala activity scales with the intensity of emotional images (Karlsson et al., 2010) and faces (Wang et al., 2017). Amygdala hyperactivity has been observed in response to negative (Etkin and Wager, 2007) and neutral facial expressions in AD (Cooney et al., 2006). For example, patients suffering from SAD showed increased amygdala activity compared to healthy controls when presented with emotional faces (Stein et al., 2002; Phan et al., 2006; Schneier et al., 2009).
These findings of impaired neural processing of affective stimuli, such as emotional faces, are replicated in depression and anxiety disorders (Stein et al., 2002; Campbell et al., 2007; Etkin and Wager, 2007). Although the amygdala is not the only brain region relevant to the perception and regulation of mood and emotion (Pessoa and Adolphs, 2010; Grupe and Nitschke, 2013), its critical involvement in these processes remains undisputed. Current empirical evidence confirms that the amygdala not only plays a critical role in the detection and processing of emotionally salient stimuli but is also involved in emotional memory formation, fear conditioning, social cognition, and reward processing (Bickart et al., 2011; Inman et al., 2020; Domínguez-Borràs and Vuilleumier, 2022). These neural mechanisms are necessary for appropriately processing emotional cues and subsequent behavioral responses. Thus, the ability to regulate amygdala responses may indicate successful context-dependent affect processing. Unfortunately, its location deep within the temporal lobe limits the applicability of non-invasive exogenous brain stimulation methods such as transcranial magnetic and electrical stimulation. Therefore, a new form of non-invasive and individualized treatment is needed to address the heterogeneous symptoms of affective disorders.
The application of a novel form of endogenous brain stimulation method, such as real-time functional magnetic resonance imaging-based neurofeedback (rt-fMRI NF), could be a promising alternative or adjunct to current treatment regimens for affective disorders. Using this technique, participants can learn to voluntarily control their neural responses by modulating their brain activity, which is presented in the form of a real-time feedback signal. Previous rtfMRI-NF studies of amygdala self-regulation have already demonstrated the benefits of this new non-invasive approach in improving emotion regulation in healthy individuals and various psychiatric disorders (e.g., Linden et al., 2012; Brühl et al., 2014; Zotev et al., 2016; Young et al., 2017; Koush et al., 2017b; Paret et al., 2018; Zaehringer et al., 2020).
However, these previous studies used a symbolic representation of the feedback signal, such as a thermometer-like scale. Here, we investigated the applicability of a novel feedback approach using adaptive naturalistic face stimuli. Instead of a discrete and abstract representation of the neurofeedback signal, we used a sequence of human faces displaying smooth transformations of varying intensities of neutral to happy or neutral to fearful facial emotions that were generated using face-morphing algorithms. This novel feedback modality might have several advantages over conventional feedback. First, social feedback is highly relevant because it reflects interactions that humans encounter in their natural habitat, and thus emotional faces are more ecologically valid than abstract symbolic feedback representations (Mathiak et al., 2015). Second, previous studies simultaneously presented the visual cue and the feedback signal at different locations or subsequently, in order to modulate stimulus-induced activity through neurofeedback training (Brühl et al., 2014; Paret et al., 2014; Hartwell et al., 2016). Such dual-task interference between stimulus perception and feedback monitoring can be avoided by combining amygdala-relevant pictorial cues (Geissberger et al., 2020) and the feedback signal. Third, interactive emotional faces (i.e., a fearful face that gradually becomes less fearful, and a neutral face that becomes happier) are more socially motivating as the participant’s regulation effort has real motivational consequences in terms of approach and avoidance of the task-relevant feedback signal (Carver, 2006; Ihssen et al., 2017).
A few neurofeedback studies have used interactive feedback interfaces such as human avatars that dynamically change emotional expressions corresponding to the targeted neural activity (Mathiak et al., 2015; Direito et al., 2019, 2021). The current study paradigm included a more realistic and dynamic feedback presentation of human face stimuli. We argue that this type of innovative feedback could be more naturalistic, ecologically valid, and socially rewarding as compared to virtual avatars (Philip et al., 2018). Finally, this setup also allows for more complex research designs inspired by closed-loop control theory to shape brain dynamics through positive (i.e., a strong brain response causes a stronger stimulus) and negative (i.e., a strong brain response causes a weaker stimulus) feedback loops that increase or decrease neural responsivity, respectively (Toates, 1975; Mulholland and Eberlin, 1977; Mulholland et al., 1979; Pope et al., 1995).
In this study, we tested the efficacy of this novel closed-loop neurofeedback approach in healthy participants. The prospective goal is to use this tailored adaptive experimental therapy to target affective biases and associated symptoms in psychiatric disorders. Participants were randomly assigned to four groups based on the emotional face stimuli and task congruency. Participants were instructed to make the neutral face happier by increasing their amygdala activity in the happy-up (task-congruent) and decreasing it in the happy-down (task-incongruent) groups. Whereas participants were instructed to reduce the fearfulness of the face by downregulating their amygdala activity in the fear-down (task-congruent) group and upregulating it in the fear-up (task-incongruent) group, respectively (Figure 1). Perceiving facial expressions can help regulate emotional responses, such as enhancing positive mood or reducing negative affective state, especially when the expression of the face stimulus matches one’s intentions, in other words, is congruent with one’s regulatory efforts. For example, in the happy-up (task-congruent) group, the task goal was to increase the happiness of the face stimulus by upregulating amygdala activity, i.e., the face stimulus is rewarding the inherent human tendency to perceive happy faces by upregulating the amygdala. On the other hand, being exposed to emotional stimuli that are not in alignment with one’s regulatory efforts can lead to task incongruency. For example, in the happy-down group, the goal was to increase the happiness of the face stimulus by decreasing amygdala activity. In this condition, the rewarding face stimuli (happy faces) contradict the task goal of downregulating the amygdala. This type of feedback setup may require greater cognitive effort due to internal conflict of pursuing a reward by engaging contrasting neural patterns and may provide a training opportunity akin to exposure therapy. With such a versatile and comprehensive feedback design, we hypothesized that healthy participants would learn to upregulate and downregulate their amygdala activity in the task-congruent groups, i.e., happy-up and fear-down, respectively, as compared to the respective task-incongruent groups (happy-down and fear-up).
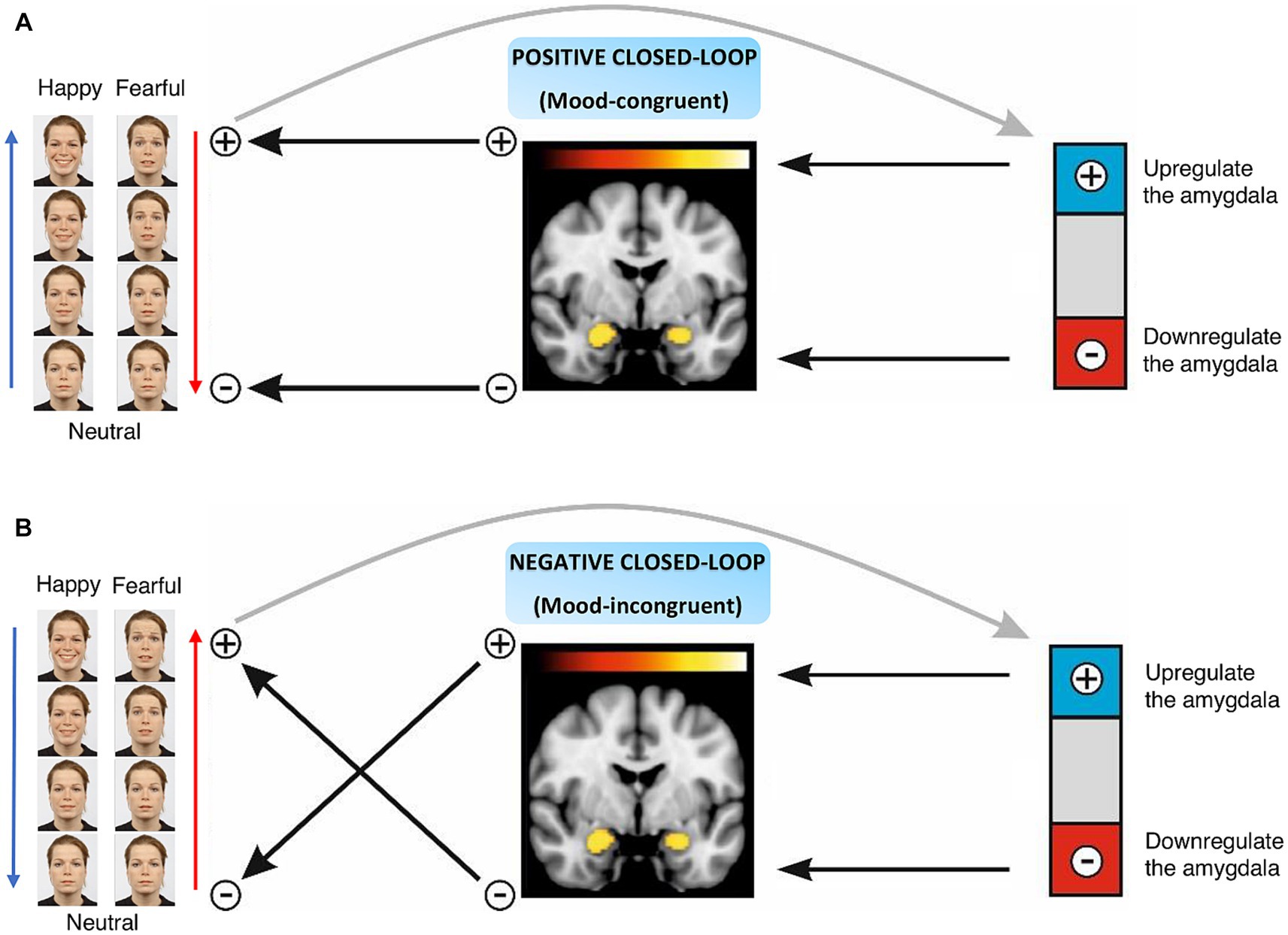
Figure 1. Naturalistic closed-loop neurofeedback design. (A) Positive closed-loop includes task-congruent conditions, i.e., happy-up and fear-down groups, where an increase in the amygdala activity enhances the intensity of the facial affect. (B) Negative closed-loop characterizes task-incongruent conditions, i.e., happy-down and fear-up groups where an increase in the amygdala activity reduces the intensity of the facial affect and vice versa.
Next, we hypothesized that the task-dependent effective connectivity between the amygdala and the prefrontal cortex and/or face-sensitive regions in the temporal lobe would increase over the course of the neurofeedback training session. In the context of affective disorders, maladaptive emotional responses are manifested due to dysfunctional inhibitory control of higher order cognitive top-down processes on the maladaptive bottom-up pathways regulated by subcortical regions like the amygdala (Nicholson et al., 2017). Previous work has already demonstrated the role of the fusiform face area (FFA) and the medial orbitofrontal cortex (mOFC) in face processing and amygdala regulation (Kanwisher et al., 1997; Almeida et al., 2009; de Almeida et al., 2011; Sabatinelli et al., 2011; Roy et al., 2012; Sladky et al., 2015). We employed dynamic causal modeling (DCM; Friston et al., 2003), a Bayesian framework, to assess the directionality of neural dynamics involved in emotion processing (Krylova et al., 2021; Sladky et al., 2022). Specifically, we hypothesized that upregulation of the amygdala would result in positive (facilitatory), and downregulation would entail negative (inhibitory) connectivity between the mOFC and the amygdala. Finally, we assessed the intervention-induced mood changes in the participants using self-rated psychometric questionnaires such as the Positive And Negative Affect Schedule (PANAS, Watson et al., 1988) and the Self-rating Depression Scale (SDS, Zung, 1965). We hypothesized that the naturalistic closed-loop training would improve participants’ positive affectivity scores on the PANAS scale while decreasing PANAS negative affectivity scores, and SDS scores post-training, specifically, in the task-congruent groups (happy-up and fear-down).
Sixty-four healthy adults between 18 and 65 years of age, fluent in German, right-handed with normal vision and without any MRI contraindications such as pregnancy, claustrophobia, metallic implants, clinically significant somatic diseases, brain surgery, neurological disorders, and substance abuse, were recruited for the study. The participants (mean age = 25.07 ± 4.46 years) were assigned to one of the four age- and gender-matched neurofeedback intervention groups with 16 participants in each group, i.e., happy-up (mean age = 26.59 ± 4.87 years, gender = 8 m:8f), happy-down (mean age = 25.93 ± 5.93 years, gender = 8 m:8f), fear-up (mean age = 23.85 ± 3.54 years, gender = 7 m:9f), and fear-down (mean age = 23.63 ± 2.85 years, gender = 8 m:8f). They were instructed to either up or downregulate the amygdala neurofeedback signal while they were presented with neutral to happy or neutral to fearful human faces (Figure 2). Participants were asked to abstain from psychotropic substances for a minimum of 3 days and from alcohol for at least 24 h prior and nicotine/caffeine 1 h prior to the imaging session. All participants provided written informed consent in accordance with the Declaration of Helsinki and were compensated for their participation. The study was approved by the local ethics committee of the canton Zurich.
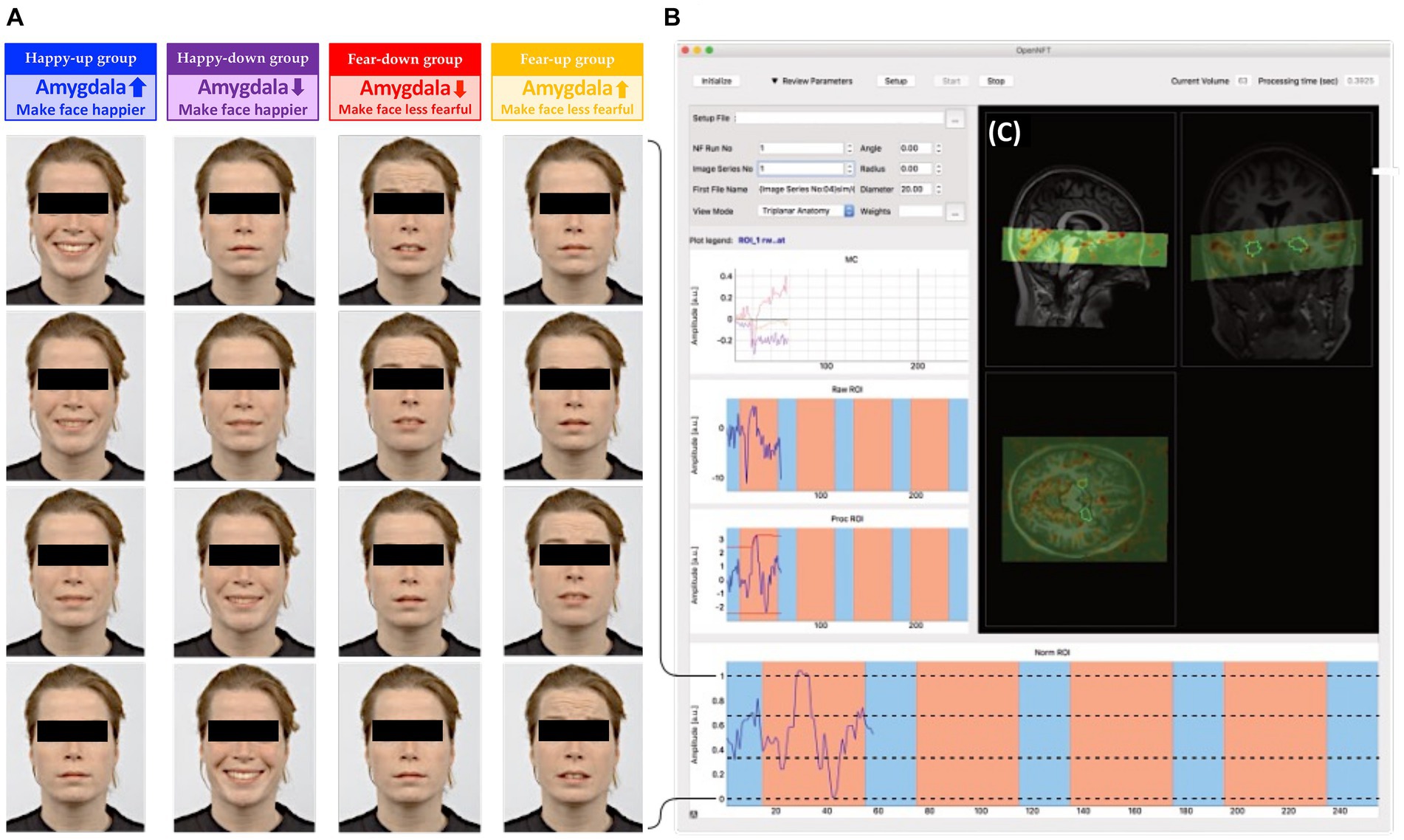
Figure 2. (A) Dynamic emotional face stimuli: Faces were digitally morphed in 30 steps from neutral to happy (happy groups) or neutral to fearful (fear groups). Subjects were instructed to either upregulate (up groups) or downregulate (down groups) the amygdala, and thus change the valence of the faces according to the group assignment. (B) OpenNFT display: Feedback from bilateral amygdala was continuously estimated using OpenNFT, an open-source framework for neurofeedback training (http://www.opennft.org, Koush et al., 2017a). (C) The field of view (green) projected on a participant’s structural scan covered the mOFC, temporal lobe (amygdala, fusiform gyrus) and parts of the visual cortex. The stimulus intensities were calculated based on an anatomical mask of the bilateral amygdala (Tyszka and Pauli, 2016), and were presented as feedback using PsychoPy.
The neurofeedback session consisted of four training runs with each neurofeedback run having four 40-s regulation blocks interleaved with four 20-s baseline blocks, during which participants were instructed to mentally count slowly (starting from one in increments of one) while fixating on a dot. During each regulation block, a new face from the Radboud Faces Database (Langner et al., 2010) was presented continuously. The faces were randomized, unique (i.e., novel for the participant), and counterbalanced for gender. Based on the group assignment, the faces expressed some degree of positive (i.e., happy) or negative (i.e., fearful) affective state. The intensity of the emotional expression was proportional to the mean blood oxygenation level dependent (BOLD) signal intensity of the participant’s bilateral amygdala. The varying degrees of emotional intensity were achieved by smoothly blending faces with different emotional intensity using a face morphing software written in Python1 (Willinger et al., 2019). Thus, participants were presented with highly naturalistic face stimuli that dynamically changed depending on their amygdala activity (Figure 2). Depending on the emotional stimulus and regulation condition, participants were assigned to one of the four experimental groups and received group-specific instructions as described below:
1. happy-up: “Try to make the face happier by up-regulating the amygdala.”
2. happy-down: “Try to make the face happier by down-regulating the amygdala.”
3. fear-down: “Try to make the face less fearful by down-regulating the amygdala.”
4. fear-up: “Try to make the face less fearful by up-regulating the amygdala.”
For all groups, ongoing bilateral amygdala activity was translated into facial expression intensity either from neutral to happy (first and second groups), or from fearful to neutral (third and fourth groups), as depicted in Figure 2. Participants were informed about the amygdala’s involvement in affect processing and that the change in the expression of the face stimulus was based on their amygdala activity. They were given the freedom to use the emotion regulation strategies of their choice and to modify the strategies between runs. They were also asked to verbalize the experimental mental strategy beforehand and to remain focused during the task. Repeated sampling of participants’ feedback between each run also ensured that participants remained engaged in the task.
The naturalistic face stimuli consisted of human faces of 30 Caucasian models (15 females) from the Radboud Face Database depicting neutral, fearful, and happy emotions (Langner et al., 2010). A face morphing algorithm developed in Python was used to create dynamic emotional faces with gradually changing facial expressions in 30 steps,2 such that 0 corresponded to the lowest valence or neutral emotion (0%) and 30 to the highest valence (100%), i.e., fearful or happy (smiling) expressions. The intensity of the emotional valence was coupled to the average BOLD signal of the participant’s bilateral amygdalae.
The estimation and presentation of the feedback signal was achieved using the Open NeuroFeedback Training (OpenNFT) software, an open-source neurofeedback framework implemented using Python and Matlab (Koush et al., 2017a). The feedback was scaled to the normalized amygdala time-series using OpenNFT’s default dynamic range to estimate the maximum and minimum limits of the scaling (Koush et al., 2017a) by using the average of the 5% highest and lowest signal intensities observed so far. The preprocessed amygdala signal was then mapped to the intensity of the emotional expression ranging between the lowest and the highest valence of the face stimulus that served as the feedback signal.
A high spatial resolution, three-dimensional, probabilistic in vivo anatomical mask of the bilateral amygdala used in the current study was based on the California Institute of Technology (CIT168) human brain templates (Tyszka and Pauli, 2016), and was co-registered non-linearly in individual MNI space (Figure 2C). The template mask was created using the SPM normalization function through inverse warping.
MRI acquisitions were performed using an Achieva 3-Tesla MRI scanner (Philipps Healthcare, Best, The Netherlands) and the manufacturer’s 32 channel head coil at the MR Centre of the Psychiatric Hospital, University of Zurich. We acquired 265 volumes covering the mOFC, temporal lobe (amygdala, fusiform gyrus) and parts of the visual cortex per training run using a gradient echo T2*-weighted echo planar imaging (EPI) sequence with the following parameters: TR = 1,000 ms, TE = 35 ms, 15 interleaved ascending axial slices, 2 × 2 × 2 mm3 voxel size, 1 mm slice gap, 112 × 110 matrix, field of view (FOV) = 224 × 224 × 44 mm3, flip angle of 65°, SENSE factor 2. The first five volumes were discarded as dummy scans. Additionally, a whole brain EPI volume (same parameters as above except 70 slices and a TR = 5,000 ms) and a whole brain T1-weighted structural scan (TR = 9 ms, TE = 5 ms, 160 coronal slices, 1 × 1 × 1 mm3 voxel size, 240 × 240 matrix, FOV = 240 × 160 × 240 mm3, flip angle = 8°) were acquired for online and offline data processing and to localize the bilateral amygdala. For rt-fMRI NF, brain images were exported in real-time to a high-performance computer using the proprietary Philips DRIN export system.
A spatial transformation Statistical Parametric Mapping (SPM12, v7771, https://www.fil.ion.ucl.ac.uk/spm/software/spm12/) batch pipeline was used to transform the participant’s structural MRI scan, whole brain EPI volume, test EPI volume with the FOV, and a bilateral amygdala mask (Tyszka and Pauli, 2016) into the individual’s space to serve as a volume of interest for neurofeedback training. The test EPI volume was used as a reference for online realignment.
Online rt-fMRI data analysis and neurofeedback signal calculation were performed using OpenNFT (Koush et al., 2017a). OpenNFT’s default online preprocessing pipeline was used which comprised real-time realignment for motion correction and spatial smoothing (Gaussian kernel, 5 mm FWHM). Temporal data processing included spike removal using a Kalman filter, drift removal using a cumulative general linear model (GLM), a first-order autoregressive model AR (1) to account for serial correlations, and OpenNFT’s default dynamical range scaling. The extracted, preprocessed amygdala timeseries data were used to dynamically scale the intensity of the emotional expression in the faces and served as a feedback signal for the participant.
Offline data processing and analysis of the four neurofeedback runs were performed in SPM12 and comprised slice-timing correction (Sladky et al., 2013), realignment, coregistration to the participant’s whole brain EPI volume and T1-weighted structural MRI image, normalization to MNI space, and spatial smoothing with a 6 mm FWHM Gaussian kernel.
A subject-level analyses were performed using SPM12-based GLM only on the first neurofeedback run due to the groups’ different regulation instructions. We aimed at differentiating the individual contributions of the experimental manipulations. We expected that the task-relevant brain responses could be modeled using three orthogonalized regressors: x1 models the effects of the fMRI task condition in general, i.e., the blocks where participants were asked to regulate their amygdala response (Figure 3A, green) by convolving the box car function that encodes the regulation blocks (20s off, 40s on period) with SPM’s canonical hemodynamic response function (cHRF). x2 modeled the subject’s response to the emotional face that scaled proportionally to the intensity of the neurofeedback signal (Figure 3A, orange), calculated by convolving the online-extracted, normalized amygdala BOLD signal (i.e., OpenNFT’s Norm ROI signal, which was also used for neurofeedback stimulus presentation) with the cHRF, which corresponds to a parametric modulator of the block design. x3 was used to model the source of the neurofeedback signal (Figure 3A, magenta), which is confounded by both the regulation task (x1) and the response to the stimulus intensity (x2). To account for this, x1 and x2 were regressed out of the normalized amygdala BOLD signal, i.e., yielding a regressor (x3) that corresponds to the residual amygdala signal independent of the BOLD response to the neurofeedback stimulus intensity (x2) and regulation blocks (x1). Six realignment parameter estimates were included as nuisance regressors to account for residual movement artifacts not accounted for by the realignment preprocessing step. This design matrix captured the circularity that is inherent to the closed-loop paradigm of this study, where the stimulus intensity depends on the activity of the target region, which in turn is influenced by the stimulus intensity.
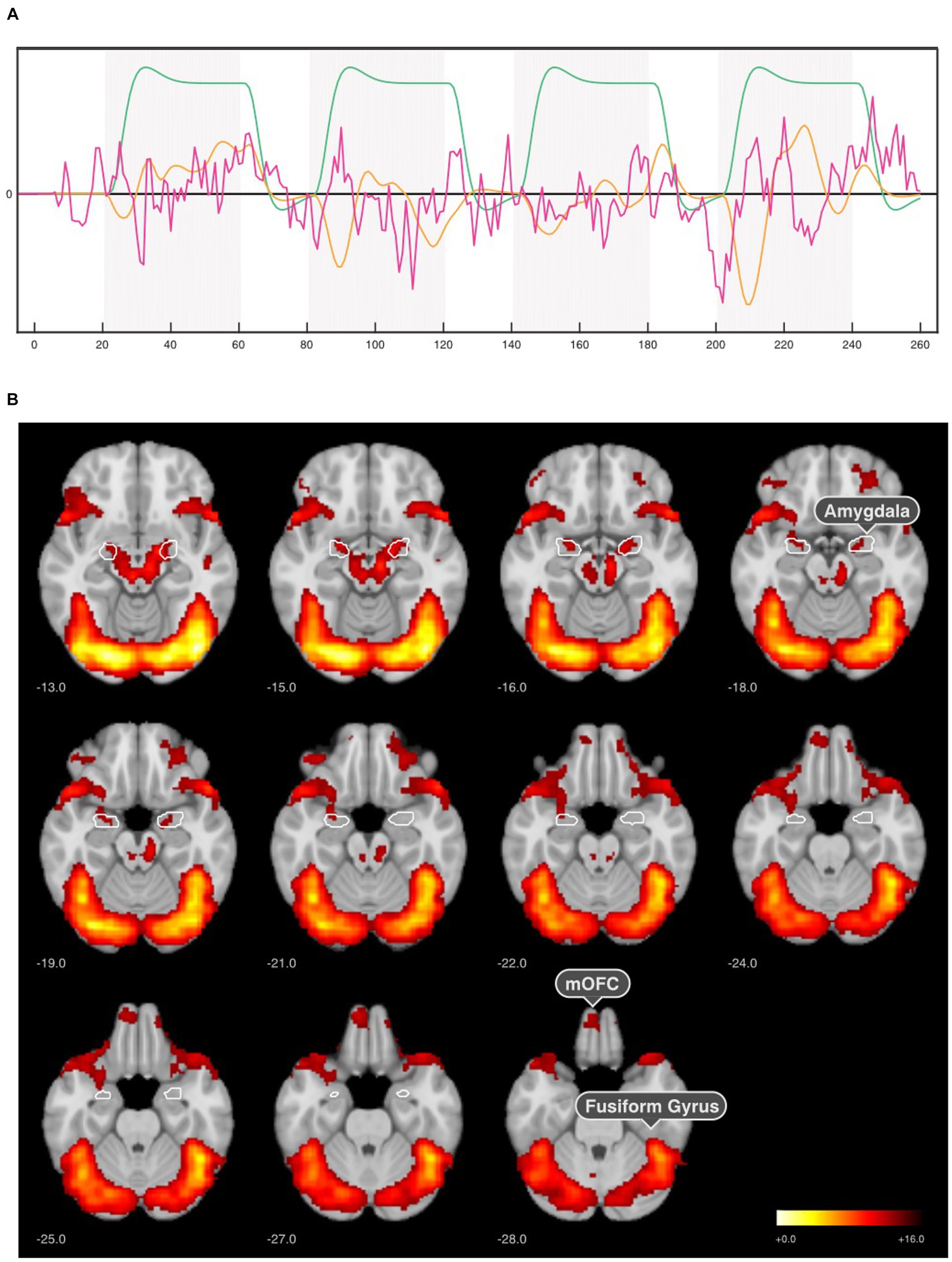
Figure 3. SPM-based GLM analysis results for the first neurofeedback run. (A) GLM model comprising three orthogonalized regressors of interest: x1—regulation blocks (green), x2 -neurofeedback response (amygdala signal x cHRF, orange), and x3—neurofeedback signal (residual amygdala signal, magenta). (B) Regulation > Baseline contrasts for x1 regressor in the first neurofeedback run across all groups (n = 64) revealed activation in the amygdala, medial orbitofrontal cortex (mOFC), fusiform gyrus (fusiform face area). The results were corrected for p < 0.001 voxel-wise threshold and p < 0.05 cluster-level family wise error (FWE).
The second-level random effects group analysis consisted of a one-sample t-test of the individual contrasts between faces and baseline to identify neurofeedback-selective brain regions in the first neurofeedback run. Significance threshold for the resulting statistical parametric map was set to p < 0.001 (voxel-wise threshold) and p < 0.05 FWE cluster-level corrected.
Amygdala time-series analyses were based on OpenNFT logs from the neurofeedback experiment. We used the pre-processed amygdala signal (i.e., kalmanProc data). Each neurofeedback run consisted of four regulation blocks (dynamic face stimulus) interleaved with the four baseline blocks (a dot). Preprocessed data were modeled using GLM analysis with four regressors for the four individual regulation blocks to estimate the mean amygdala BOLD signal in the regulation condition. Contrasts were designed as a boxcar function with the 20s off (baseline) and 40s on (regulation) period convolved with SPM’s cHRF. Average values (beta weights) were extracted from the ROI beta maps associated with each regulation block and utilized in subsequent statistical analyses.
Amygdala activity changes over four neurofeedback runs were analyzed based on linear mixed-effect model (LMM) using “lme4” package (version 1.1–31, Bates et al., 2015) in R (version 4.2.0, R Core Team, 2022). LMM analysis was performed separately for the happy (happy-up and happy-down groups) and fear (fear-up and fear-down groups) conditions. For each condition, we modeled neurofeedback runs (4 runs) and groups (congruent and incongruent) categorical variables that served as fixed effects of interest with the participant as a random effect factor. For example, in the “Happy” condition, we analyzed amygdala activity changes over four runs in task-congruent (happy-up, N = 16) and task-incongruent (happy-down, N = 16) groups with a run x group interaction. The statistical model was fitted with restricted maximum likelihood (REML) using Satterthwaite’s method. A similar analysis was performed for the “Fear” condition by testing an interaction between training runs and groups (fear-down, fear-up) to assess amygdala activity changes.
Based on previous research (de Almeida et al., 2011; Sabatinelli et al., 2011; Sladky et al., 2022), we assumed that the amygdala (Amy) receives task-relevant input from the fusiform face area (FFA; Kanwisher et al., 1997) and the medial orbitofrontal cortex (mOFC; Roy et al., 2012). The primary visual cortex (V1) was considered to be the input for the experimental perturbation, i.e., the visual stimulation by emotional faces, and connected to the other regions. FFA is part of the ventral visual processing stream originating from V1 extending to the amygdala. In addition, the amygdala and mOFC also receive low-level visual input from other sources, e.g., thalamic pathways. This means in this case the direct connections from V1 do not reflect direct anatomical connectivity but a relevant task-specific, functional relationship. Omitting these connections would bias the model toward FFA-centered processing of emotional stimuli.
Dynamic causal modeling (DCM12 as implemented in SPM12, build 6,906) was used to estimate effective connectivity between task regions. Temporally filtered and detrended time courses of V1, bilateral FFA, bilateral amygdala, and medial orbitofrontal cortex were extracted for each subject using SPM’s volume of interest (VOI) extraction batch script (single-subject significance threshold p < 0.05, first eigenvariate used as summary statistic, adjusted for effect of interest). For the amygdala, we used the same mask that was used in the neurofeedback experiment. For the other regions, 8 mm spheres were centered around the local maximum in the group-level SPMs: V1 x/y/z = −0.0/−87.0/−6.0 mm [MNI], rFFA x/y/z = +42.0/−51.0/−24.0 mm [MNI], left FFA x/y/z = −45.0/−45.0/−25.0 mm [MNI], and mOFC x/y/z = −8.00/56.00/−26.00 mm [MNI].
The model space comprised Amy, V1, FFA, and mOFC. It was assumed a priori that V1 receives the driving input (i.e., modeled as the neurofeedback face regressor). Different DCM models were created where forward connections between V1 and Amy, FFA, and mOFC as well es bidirectional connections between amygdala and mOFC and FFA were considered, which resulted in 128 models. All models were estimated and analyzed using random effects Bayesian model averaging (BMA; Penny et al., 2010) to estimate the mean effective connectivity for each connection, depending on the neurofeedback run (i.e., 1–4) and group assignment (happy-up, happy-down, fear-up, fear-down).
The influence of closed-loop amygdala neurofeedback on participants’ affectivity was investigated using self-reporting psychometric questionnaires. Participants’ positive and negative affective state changes were measured using 20-item Positive And Negative Affect Schedule (PANAS, Watson et al., 1988). The Self-rating Depression Scale (SDS, Zung, 1965) was used to screen the symptoms relating to depression. These questionnaires were filled out by the participants on a desktop computer immediately before and after their MRI measurement. Training-induced behavioral changes were assessed by fitting a LMM with PANAS and SDS scores as the outcome variable, time (pre- vs. post-training) and group (task-congruent vs. incongruent) as a fixed effect of interest, and an intercept for each participant as random effect in happy and fear conditions.
The Consensus on Reporting and Experimental Design of Clinical and Cognitive-Behavioral Neurofeedback Studies checklist (CRED-nf, Ros et al., 2020), which summarizes the current rtfMRI-NF study design, is provided in Supplementary material.
Based on SPM’s GLM analysis, the three regressors of interest were analyzed (x1—regulation blocks, x2—neurofeedback response, and x3—neurofeedback signal). For the face regulation blocks (x1), highest activation was observed in the visual cortex (occipital lobe) and fusiform face area (inferior temporal lobe) in the first neurofeedback run. The peak MNI coordinates [42, −78, −10] implied an activation in the occipital face-selective area. Besides the obvious maximum in the bilateral amygdala, this model indicated a strong BOLD response in a distributed network encompassing the mOFC, temporal, and occipital lobe in the first neurofeedback run across all participants (Figure 3B).
Neural correlates of neurofeedback training were examined using LMM analysis by modeling run (1–4) and group (congruent and incongruent) as the fixed effects and a random intercept for participant separately in the happy and fear conditions. We observed a significant effect of run [F(3,90) = 5.28, p = 0.0002] in the fear condition with decreased amygdala activity in the third run [β = −0.34, SE = 0.11, t(90) = −3.04, p = 0.003], and the fourth run [β = −0.34, SE = 0.11, t(90) = −3.02, p = 0.003] compared to the first neurofeedback run in the fear-down group. We did not observe a significant effect of group (fear-down, fear-up) or the interaction between fixed effects, i.e., run x group. In the happy condition, LMM analyses revealed no significant effect of run, group (happy-up, happy-down), or their interaction. Amygdala activity changes over four training runs in all groups are depicted in Figure 4.
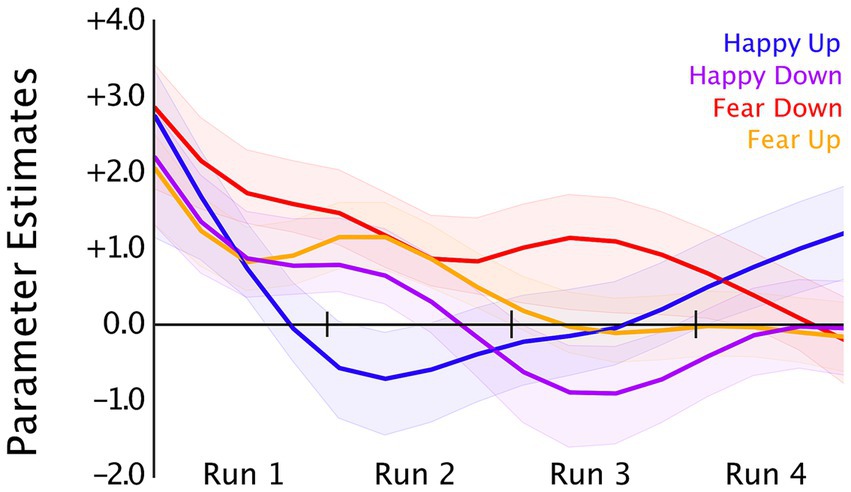
Figure 4. Bilateral amygdala time courses averaged (OpenNFT’s kalmanProc timeseries) over four neurofeedback training runs in the happy-up (blue, n = 16), happy-down (purple, n = 16), fear-down (red, n = 16), and fear-up (orange, n = 16) groups. OpenNFT’s filtered timeseries were modeled using a GLM with each regulation blocks as regressors, which resulted in a parameter estimate that represents the single subject mean of the block’s time course corrected for the hemodynamic delay. For display purposes, the plot of group means is smoothed using a Gaussian window (window length = 6).
Potential amygdala habituation effects within runs were investigated using a linear regression model of mean amygdala BOLD signal for each block in all participants. The resulting parameter estimates for habituation effects of each participant and run were further analyzed on a group level. No significant within-run habituation effects were observed for the happy-up and the happy-down groups. However, amygdala habituation effects were present in the last runs in the fear-down and fear-up groups. In the fear-down group, a statistically significant amygdala habituation occurred mainly in the third [Run 3: β = −0.449 ± 0.770, t(15) = −2.33, p = 0.034] and fourth runs [Run 4: β = −0.799 ± 0.853, t(15) = −3.75, p = 0.002]. Participants in the fear-up group exhibited an amygdala upregulation in the first three training runs with a significant habituation effect in the fourth run [Run 4: β = −0.622 ± 0.740, t(15) = −3.36, p = 0.004]. The results are summarized in Table 1.
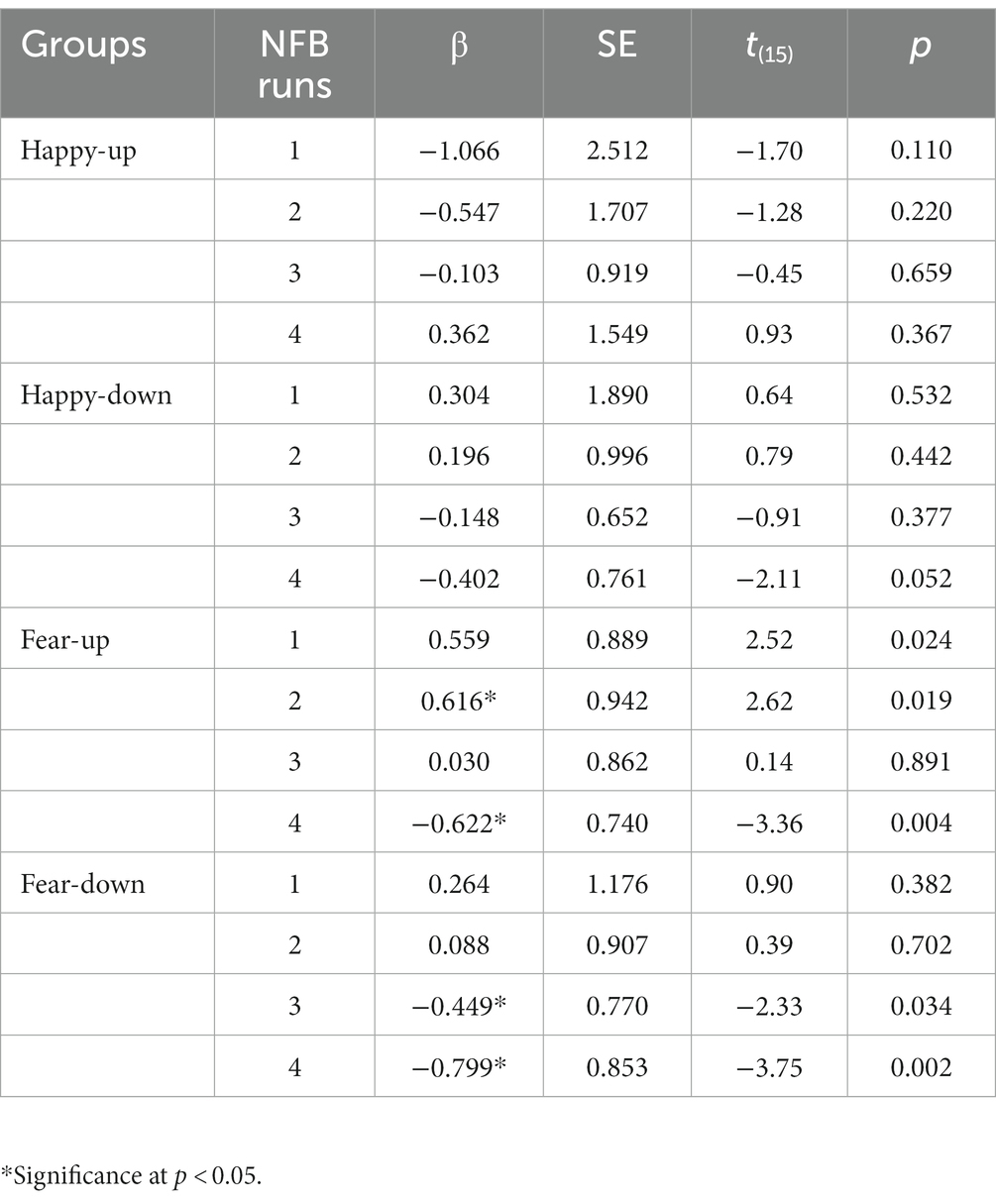
Table 1. Estimated coefficients (β), standard errors (SE), t (degrees of freedom) and p values of the change in the amygdala activity within four training runs in four experimental groups.
DCM Bayesian model averaging over 128 models is illustrated in Figure 5. The extrinsic inputs into V1 were similar in all groups. Positive connections were observed from V1 to FFA, Amygdala, and mOFC between groups. Task-dependent effective connectivity between FFA to amygdala and the mOFC to amygdala during the first and the fourth runs and in between group comparisons were performed using t-tests. The results revealed that participants in the happy-up group exhibited a significant task-dependent (i.e., intrinsic connectivity [A] + modulation by faces [B1] and the intensity of the emotion [B2]) upregulation of the amygdala induced by the FFA and mOFC in the last run as compared to the first run. On the contrary, the fear-down group exhibited strong upregulation of the amygdala in the first run, which is counteracted by inhibitory feedback of the mOFC. This downregulation effect persisted until the last run. While participants from happy-up group maintained the positive influence of FFA and mOFC on the amygdala over runs, connectivity in the fear-down group reduced over time. This could indicate a training-related learning mechanism in the fear-down group. Other groups did not show significant connectivity changes. The group differences in the effective connectivity between V1, FFA, amygdala and mOFC are depicted in Table 2.
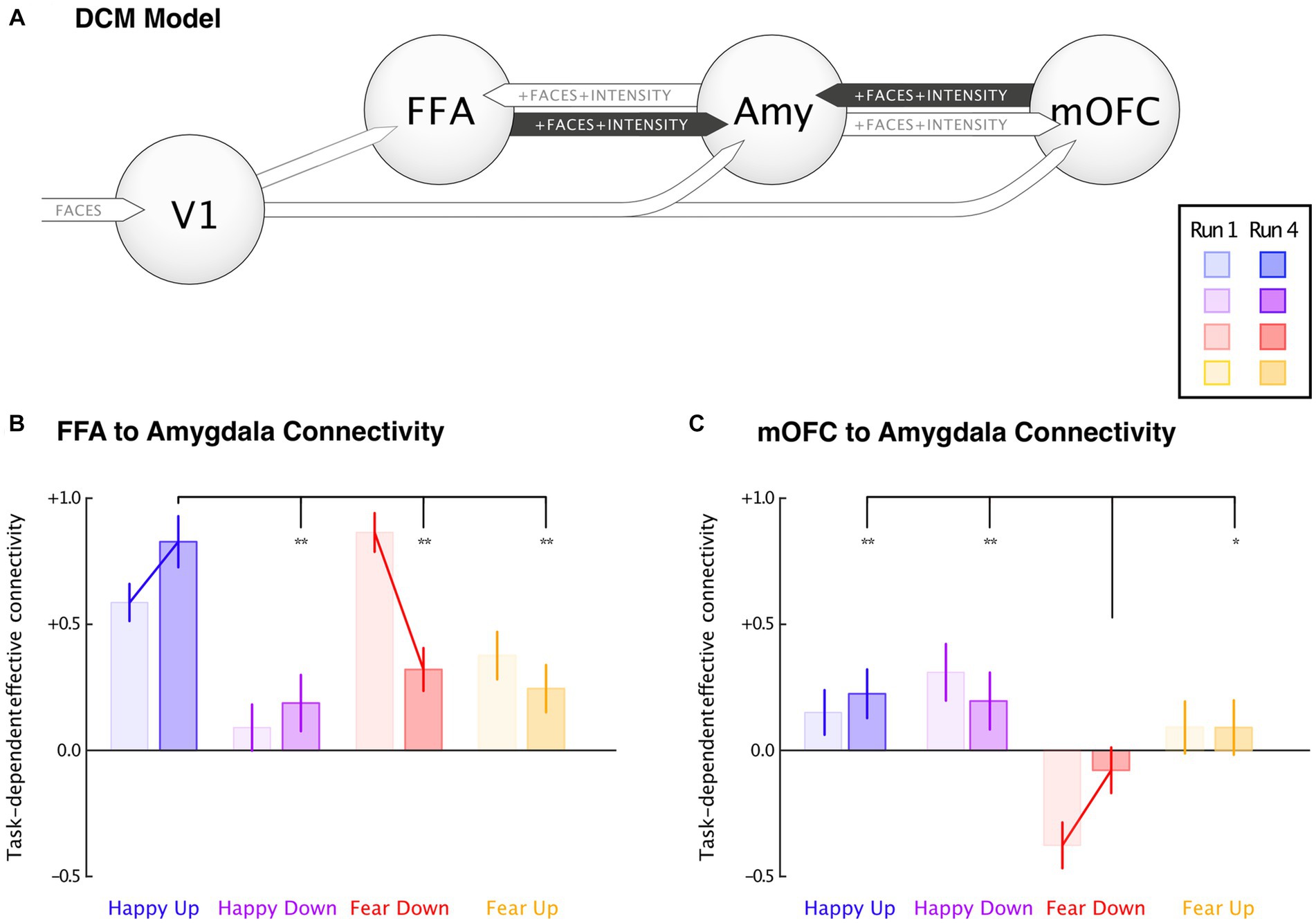
Figure 5. Dynamic causal modeling. Task-dependent connectivity changes were depicted between the first (light-colored bar) and the fourth (dark-colored bar) runs, and between groups. (A) A DCM model comprising primary visual cortex (V1), fusiform face area (FFA), medial orbitofrontal cortex (mOFC), and bilateral amygdala (Amy). (B) Effective connectivity changes between FFA to amygdala revealed a significant increase in the happy-up and decrease in the fear-down group, and (C) mOFC to amygdala connectivity showed significant negative responses in the fear-down group. Significantly decreased negative connectivity parallels reduced input from FFA. Error bars indicate 95% confidence interval, and the connecting lines denote mean significance computed using paired t-test. *Significance at p < 0.05, and **Significance at p < 0.001.

Table 2. DCM Bayesian Model Averaging results.
Within groups significant changes for FFA and amygdala connectivity were observed in happy-up (M = 0.83 ± 0.21) vs. happy-down [M = 0.19 ± 0.23, t(15) = 8.30, p < 0.001], happy-up (M = 0.83 ± 0.21) vs. fear-down [M = 0.32 ± 0.17, t(15) = 7.50, p < 0.001], and happy-up (M = 0.83 ± 0.21) vs. fear-up [M = 0.25 ± 0.19, t(15) = 8.26, p < 0.001]. Whereas, the significant mOFC and amygdala connectivity changes were observed in the happy-up (M = 0.22 ± 0.19) vs. fear-down [M = −0.08 ± 0.19, t(15) = 4.48, p < 0.001], happy-down (M = 0.19 ± 0.23) vs. fear-down [M = −0.08 ± 0.18, t(15) =3.71, p < 0.001], and fear-down (M = 0.08 ± 0.18) vs. fear-up [M = 0.09 ± 0.22, t(15) = −2.35, p = 0.03].
LMM analyses of psychometric measures revealed no significant effect of time, group, and interaction on PANAS and SDS scores in any group in the happy and fear conditions.
In the current study, we introduced an innovative neurofeedback design using dynamically adapting naturalistic face stimuli (happy and fearful faces) as a feedback signal coupled to the participant’s ongoing amygdala activity. Healthy participants were trained in four groups to upregulate or downregulate their amygdala explicitly by modulating the valence of the face stimulus. Participants in the happy condition were instructed to make the face happier either by upregulating (happy-up group, task-congruent) or downregulating (happy-down group, task-incongruent) their amygdala activity. Meanwhile, participants in the fear condition were instructed to reduce fearfulness of the face through amygdala downregulation (fear-down group, task-congruent) and through upregulation (fear-up group, task-incongruent). Thus, with such a versatile neurofeedback design, we investigated the effects of the stimulus (happy vs. fearful faces), regulation condition (upregulation vs. downregulation), and task congruency (task-congruent vs. incongruent) on learning success.
We assessed changes in the amygdala activity across four runs using LMM analysis which revealed a significant effect of run, mainly in the fear-down group (Figure 3). Higher amygdala activity was observed in all groups during the first training run which may contribute to the novelty of the emotionally salient facial stimulus (Balderston et al., 2011). Particularly, we observed a decrease in the amygdala activity in the fear-down group in the last two runs as compared to the first run. Decreasing amygdala activity in the fear-down group is congruent with the task instruction, but not in the fear-up group. It is possible that a task-congruent condition may have facilitated the perception of reward which might have encouraged the participant’s regulatory effort. Whereas, in the fear-up group, a conflict between the high-level goal of amygdala upregulation (by making the face less fearful) and intrinsic desire to perceive neutral or less intense faces as rewarding (by implicitly downregulating the amygdala) may have contributed to unsuccessful upregulation.
However, contrary to our hypothesis, we did not observe a significant increase in the amygdala BOLD signal in the happy-up group, where the task instructions were congruent. A possible explanation is a ceiling effect, as reported by Paret et al. (2018), where healthy participants could not achieve successful amygdala upregulation while exposed to aroused pictures.
Some of the observed effects, particularly in the fear conditions, could be attributed to amygdala habituation. Similar effects have been observed previously (Plichta et al., 2012; Geissberger et al., 2020), however, these studies did not address the internal regulatory mechanisms and goal-directed behavior of the participants. We propose that the amygdala is not a passive processor of emotional stimuli that becomes habituated when the stimuli are no longer novel. Instead, it is part of a complex network that is also influenced by the participant’s ongoing motivational goals, expectations, and other cognitive processes. We observed that the amygdala habituated in the fear-down and fear-up groups, but not in the happy-up and happy-down groups. This suggests that the participant’s goal-directed behavior and the valence of the stimulus influence potential amygdala habituation effects.
We also observed amygdala effective connectivity changes using DCM that were specific to the experimental condition. We found that FFA to amygdala connectivity increased in the happy-up group and decreased in the fear-down group over four training runs. Consistent with our hypothesis, amygdala upregulation is associated with positive (i.e., facilitatory) and downregulation with negative (i.e., inhibitory) connectivity between the mOFC and the amygdala. This finding is supported by the previous effective connectivity studies suggesting that amygdala downregulation is mediated by the mOFC when participants view emotional faces (Sladky et al., 2015; Minkova et al., 2017). In the first run, the amygdala was strongly downregulated by the mOFC in the fear-down group as compared to the other groups. This downregulation effect persisted until the last run. It appears that during the first run most of the regulation occurs top-down via the mOFC, while in the last run, the bottom-up influence of the FFA is increased. While the present study cannot provide a definitive answer for the underlying mechanisms, we can speculate that upregulating (happy-up) or downregulating (fear-down) the influence of the external stimulus would be more efficient than prefrontal regulation, both neuronally (i.e., because of long-range connections) and psychologically (i.e., because of cognitive effort). Conceptualizing neurofeedback learning as a metabolically efficient adaptation of hierarchical regulation loops contrasts the view that the amygdala is a statically mapped, stimulus-driven system. Instead, viewing the amygdala as a dynamic control circuit would be in line with an active inference interpretation of amygdala function, where amygdala communication occurs via flexible perceptual predictions and resulting prediction errors (Sladky et al., 2023).
Reduced effective connectivity between the OFC and the amygdala has also been observed during increased cognitive workload (Minkova et al., 2017) and in patients with social anxiety disorder (Sladky et al., 2015). The orbitofrontal cortex appears to play a central role in social anhedonia (Germine et al., 2011). Consistent with this model, MDD patients showed a decreased amygdala upregulation by the orbitofrontal cortex when exposed to happy faces as compared to healthy controls (de Almeida et al., 2009). In our study, significant affect-specific changes were observed only in the task-congruent groups such as happy-up and fear-down, supporting an innate tendency of neural processes to increase positive and decrease negative affectivity. This may suggest an adaptive learning process guided by mOFC activity during the training. It is highly plausible that the connectivity between mOFC and amygdala is rapidly updated during task performance, as mOFC is thought to be involved in regulation mechanisms that enable adaptive behavior (Lichtenberg et al., 2017).
We did not observe significant changes in the psychometric questionnaire scores of the participants in any of the groups. As this feasibility study was conducted in healthy participants to test the efficacy of the naturalistic feedback approach, it is less likely that the training will produce lasting or strong changes in the affectivity in participants without mood impairments. Therefore, the plausibility of this novel training approach in reducing negative mood and anxiety symptoms should be evaluated in a clinical sample such as MDD and AD patients. In addition, the pragmatic utility of the amygdala dynamic face feedback training for improving affective responses in real-life environment needs to be investigated in future studies.
This innovative approach allowed participants to train self-regulation of their amygdala by changing the emotional expression of facial stimuli. Previous rtfMRI-NF studies have reported successful amygdala downregulation in healthy participants while viewing negative emotional faces (Brühl et al., 2014) or aversive situations (Sarkheil et al., 2015; Paret et al., 2016; Herwig et al., 2018), and in clinical populations such as patients suffering with PTSD (Nicholson et al., 2016) and bipolar disorder (Zaehringer et al., 2020). Amygdala upregulation was also achieved through positive autobiographical memory recall in healthy subjects (Zotev et al., 2016; Hellrung et al., 2018) and MDD patients (Young et al., 2017). Moreover, Paret et al. (2018) demonstrated concurrent upregulation and downregulation of the amygdala in healthy female participants while viewing emotional images. Our new experimental setup extends these previous findings by providing an ecologically valid, naturalistic face feedback training that could be a versatile neuroscience research tool and a novel experimental therapy.
Compared to the abstract feedback representations used in these studies, the use of emotional faces is highly relevant to studies investigating how we perceive emotions and regulate subsequent neural responses. Social cognition involves the processing of verbal and nonverbal social cues by identifying, perceiving, and interpreting other people’s behaviors, intentions, feelings, and beliefs (Frith and Frith, 2006). Faces are an important source of information relevant to the perception of the mental state of others (Weightman et al., 2014). As a result, humans are highly trained to discriminate static (e.g., identity, gender, age) and dynamic features (e.g., gaze direction and emotional expression) of other peoples’ faces (Bruce and Young, 1986). While static features are processed in the fusiform gyrus (Haxby et al., 2000), their dynamicity is processed in the superior temporal sulcus (STS) with the amygdala and insula for processing of emotional expressions (Haxby et al., 2000). Thus, using a facial feedback with dynamically adapting affect is more realistic, socially rewarding and increases the sense of agency as compared to the symbolic feedback representation. Furthermore, it reduces the cognitive load associated with participants’ explicit regulatory efforts by preventing dual-task interference between visual cues and feedback signal (Ihssen et al., 2017; Sitaram et al., 2017).
Naturalistic face feedback could lead to more rewarding social interactions which is specifically relevant for autism spectrum disorders (Dichter, 2018), SAD (Sladky et al., 2012), and addiction disorders, where the hedonic value of social rewards is reduced. Regarding the latter, most studies address the “wanting” component of addiction (e.g., compulsive thoughts; Koob and Le Moal, 1997; Sulzer et al., 2013; Kirschner et al., 2018), which is thought to be mediated by the dopaminergic reward network. Amygdala neurofeedback using naturalistic social rewards could be a complementary method to address the “liking” component of hedonic experience (Robinson and Berridge, 2000; Haugg et al., 2022). Thus, presenting social feedback in the form of emotional faces may be beneficial in training healthy individuals and psychiatric patients to improve social cognition.
Given the variety of methodological and psychological questions that can be addressed with such an adaptive facial feedback design, its use is not solely restricted to training a single brain region such as the amygdala. It can be tailored to address other affect processing brain regions like the anterior cingulate cortex (ACC; Mathiak et al., 2010, 2015) or even functional networks (Ramot et al., 2017; Koush et al., 2017b; Zich et al., 2020; Taylor et al., 2022) to regulate neural responses that govern emphatic behavior and social cognition. Thus, dynamic emotional faces serve as suitable naturalistic feedback stimuli for shaping clinically relevant amygdala activity.
This feasibility study has several limitations. We used a high-resolution anatomical mask to define the region of interest (ROI), i.e., bilateral amygdalae (Tyszka and Pauli, 2016). The amygdala is a small subcortical structure that is functionally dynamic and anatomically diverse (Tyszka and Pauli, 2016; Wang et al., 2017), so using a predefined mask to generate the feedback signal could be a limitation. Comparatively, a functional localizer task may have higher accuracy in defining participant-specific ROI that facilitates neurofeedback learning, but there is currently no supporting evidence (Haugg et al., 2021). A more refined ROI definition based on amygdala sub-nuclei may improve the quality of the neurofeedback signal which can be achieved with ultra-high field imaging at 7-Tesla fMRI, which provides a higher signal-to-noise ratio, improving spatial specificity in the subcortex (Hahn et al., 2013; Sladky et al., 2018).
Another limitation could be a restricted field of view (temporal lobe, visual cortex, and mOFC), which hindered understanding the potential influence of affect processing brain regions other than the amygdala, specifically, the cortical areas and other face processing areas such as STS and occipital face area (OFA). We restricted our data acquisition to the ventral brain because we wanted to investigate the involvement of the FFA and mOFC in addition to the amygdala. While prefrontal regions such as dorsolateral prefrontal cortex (DLPFC) play an important role in reappraisal mechanisms (Morawetz et al., 2016), we previously identified the mOFC as the most relevant causal influence on amygdala regulation (Sladky et al., 2022). However, other PFC regions engaged through the participants’ explicit cognitive regulatory efforts and emotion perception regions like STS which is responsible for social cognition (Direito et al., 2021) are highly relevant and require further exploration. Future studies using faster acquisition methods such as multiband EPI sequences and higher field strengths will allow for better brain coverage without compromising the image resolution.
The main limiting factor could be the short duration of the neurofeedback training. In this proof-of-concept study, participants completed only four training runs within a single session of neurofeedback training. It is possible that such short-duration training is not enough to induce learning and subsequent behavioral changes. Hence, it is difficult to make any causal inferences regarding the effectiveness of the novel naturalistic face neurofeedback design. Currently, there is no consensus on the optimal number of neurofeedback runs or sessions required to achieve desired neural and behavioral changes in healthy and in clinical populations (Paret et al., 2019). Furthermore, recent evidence suggests that patients are more successful than healthy participants at learning self-regulation of brain activity (Haugg et al., 2021). Therefore, enrolling more participants in intensive longitudinal neurofeedback training and testing this novel approach in psychiatric disorders may lead to clinically relevant neural and behavioral changes.
Another main limitations of the study are the lack of a control group and the no-feedback practice (i.e., baseline) and transfer runs. The inclusion of a control condition is necessary to improve the specificity of the training and to rule out non-specific factors influencing the regulation success (Sorger et al., 2019). Thus, without a control group in the current study, it is difficult to ascertain causal effects of the naturalistic face neurofeedback training. A recent meta-analysis has reported a positive correlation between the learning success and pre-training practice run without feedback (Haugg et al., 2021). While a practice run may enhance the effectiveness of the chosen regulation strategies during the actual training, a post-training no-feedback run, or a transfer run is a prerequisite for improving the transferability of the learned behavior to real life. These aspects of neurofeedback training are not addressed in the current study.
The type of adaptive emotional face feedback used in the current study may pose a challenge in determining whether the observed changes in amygdala activity are attributed to participants’ conscious regulatory efforts based on the feedback they receive (feedback-driven) or are solely a response to the facial stimuli (stimulus-driven). This is because face feedback not only represents the brain’s response to external stimuli (e.g., dynamic facial emotions) but also simultaneously influences the person’s ability to regulate brain activity (Direito et al., 2019). This distinction is crucial for ascertaining the effectiveness of neurofeedback training. However, disentangling these two aspects is methodologically complex given the small sample size and beyond the scope of the current study. Larger and more diverse study samples (e.g., including clinical populations) might allow investigating the individual differences in emotion and amygdala regulation success at the level of effective connectivity differences. Furthermore, follow-up studies could employ specifically designed transfer runs or sham feedback conditions. Future neurofeedback studies using an adaptive feedback interface should aim to investigate the stimulus-driven and self-regulation-driven changes in brain activity.
The unavailability of the data on subjective performance during the training could be another confound. Participants included in the study were given the freedom to use emotion regulation strategies of their choice and were briefed about the amygdala neurofeedback and subsequent training goals (i.e., to make the face happier or less fearful). They were asked to verbalize the mental strategy that they intended to use before each neurofeedback run to avoid mind-wandering and rumination during the training. The most common strategies reported by the participants to make the face happier were mentally telling a joke, imagining positive memories, imagining tickling a person etc., whereas, to reduce the fearfulness of the face, calming the person down, imagining walking by the lake, hugging a person etc., were used. However, we did not include the individual regulation strategies used by the participants during the training in our analysis as we did not perform a structured qualitative interview. The use of appropriate and effective mental strategies may be central to the cognitive learning and may have an influence on neural processes and subsequent behavior, especially, in patients with psychiatric disorders (Mac Duffie et al., 2018; Herwig et al., 2019). Hence, future neurofeedback studies should consider including subjective data to address regulatory success.
In conclusion, the results of this proof-of-concept study in healthy participants have demonstrated that the dynamic emotional face neurofeedback design offers a powerful experimental approach for investigating complex interactions between stimuli, regulation conditions, and adaptive feedback signals to efficiently shape brain activity. Although the present study did not confirm the clinical efficacy of closed-loop naturalistic face neurofeedback, the literature has demonstrated the therapeutic benefits of interactive feedback interfaces such as dynamic face stimuli in clinical settings (Direito et al., 2021). In this context, such a novel naturalistic face feedback design could be an extension of currently developing adaptive neurofeedback protocols for affective and other psychiatric disorders.
The raw data supporting the conclusions of this article will be made available by the authors on request, without undue reservation.
The studies involving humans were approved by Kantonale Ethikkommission Zürich. The studies were conducted in accordance with the local legislation and institutional requirements. The participants provided their written informed consent to participate in this study.
AW: Data curation, Formal analysis, Project administration, Writing – original draft, Methodology. AH: Data curation, Writing – review & editing. NF: Data curation, Writing – review & editing. YK: Software, Writing – review & editing. DW: Software, Writing – review & editing. AB: Conceptualization, Methodology, Writing – review & editing. PS: Software, Writing – review & editing. FS: Conceptualization, Funding acquisition, Investigation, Methodology, Resources, Supervision, Validation, Writing – review & editing. RS: Conceptualization, Data curation, Formal analysis, Funding acquisition, Investigation, Methodology, Project administration, Supervision, Writing – original draft, Writing – review & editing.

The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This study was supported by the Swiss National Science Foundation (BSSG10_155915, 100014_178841, and 32003B_166566), the Foundation for Research in Science and the Humanities at the University of Zurich (STWF-17-012), and the Baugarten Stiftung.
The authors would like to thank Bingjie Cheng, Margrith Kramis Jordi, Cindy Lor, and Vivian Steiger for their assistance during data acquisition, recruitment, and preparation. We would like to thank Lydia Hellrung and Hanne Scheerer for their valuable research inputs.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fnins.2023.1286665/full#supplementary-material
Almeida, J. R. C., Mechelli, A., Hassel, S., Versace, A., Kupfer, D. J., and Phillips, M. L. (2009). Abnormally increased effective connectivity between parahippocampal gyrus and ventromedial prefrontal regions during emotion labeling in bipolar disorder. Psychiatry Res. Neuroimaging 174, 195–201. doi: 10.1016/j.pscychresns.2009.04.015
Balderston, N. L., Schultz, D. H., and Helmstetter, F. J. (2011). The human amygdala plays a stimulus specific role in the detection of novelty. Neuroimage 55, 1889–1898. doi: 10.1016/j.neuroimage.2011.01.034
Bates, D., Mächler, M., Bolker, B., and Walker, S. (2015). Fitting linear mixed-effects models using lme 4. J. Stat. Softw. 67, 1–48. doi: 10.18637/jss.v067.i01
Baxter, A. J., Vos, T., Scott, K. M., Ferrari, A. J., and Whiteford, H. A. (2014). The global burden of anxiety disorders in 2010. Psychol. Med. 44, 2363–2374. doi: 10.1017/S0033291713003243
Bickart, K. C., Wright, C. I., Dautoff, R. J., Dickerson, B. C., and Barrett, L. F. (2011). Amygdala volume and social network size in humans. Nat. Neurosci. 14, 163–164. doi: 10.1038/nn.2724
Bruce, V., and Young, A. (1986). Understanding face recognition. Br. J. Psychol. 77, 305–327. doi: 10.1111/j.2044-8295.1986.tb02199.x
Brühl, A. B., Scherpiet, S., Sulzer, J., Stämpfli, P., Seifritz, E., and Herwig, U. (2014). Real-time neurofeedback using functional MRI could improve down-regulation of amygdala activity during emotional stimulation: a proof-of-concept study. Brain Topogr. 27, 138–148. doi: 10.1007/s10548-013-0331-9
Campbell, D. W., Sareen, J., Paulus, M. P., Goldin, P. R., Stein, M. B., and Reiss, J. P. (2007). Time-varying amygdala response to emotional faces in generalized social phobia. Biol. Psychiatry 62, 455–463. doi: 10.1016/j.biopsych.2006.09.017
Carver, C. S. (2006). Approach, avoidance, and the self-regulation of affect and action. Motiv. Emot. 30, 105–110. doi: 10.1007/s11031-006-9044-7
Cooney, R. E., Atlas, L. Y., Joormann, J., Eugène, F., and Gotlib, I. H. (2006). Amygdala activation in the processing of neutral faces in social anxiety disorder: is neutral really neutral? Psychiatry Res. 148, 55–59. doi: 10.1016/j.pscychresns.2006.05.003
Cuijpers, P., Beekman, A. T. F., and Reynolds, C. F. (2012). Preventing depression: a global priority. JAMA 307, 1033–1034. doi: 10.1001/jama.2012.271
Davies, C. D., Young, K., Torre, J. B., Burklund, L. J., Goldin, P. R., Brown, L. A., et al. (2017). Altered time course of amygdala activation during speech anticipation in social anxiety disorder. J. Affect. Disord. 209, 23–29. doi: 10.1016/j.jad.2016.11.014
de Almeida, J. R. C., Kronhaus, D. M., Sibille, E. L., Langenecker, S. A., Versace, A., LaBarbara, E. J., et al. (2011). Abnormal left-sided Orbitomedial prefrontal cortical-amygdala connectivity during happy and fear face processing: a potential neural mechanism of female MDD. Front. Psych. 2:69. doi: 10.3389/fpsyt.2011.00069
de Almeida, J. R. C., Versace, A., Mechelli, A., Hassel, S., Quevedo, K., Kupfer, D. J., et al. (2009). Abnormal amygdala-prefrontal effective connectivity to happy faces differentiates bipolar from major depression. Biol. Psychiatry 66, 451–459. doi: 10.1016/j.biopsych.2009.03.024
Dichter, G. S. (2018). Motivational impairments in autism may be broader than previously thought. JAMA Psychiatry 75:773. doi: 10.1001/jamapsychiatry.2018.1078
Direito, B., Lima, J., Simões, M., Sayal, A., Sousa, T., Lührs, M., et al. (2019). Targeting dynamic facial processing mechanisms in superior temporal sulcus using a novel fMRI neurofeedback target. Neuroscience 406, 97–108. doi: 10.1016/j.neuroscience.2019.02.024
Direito, B., Mouga, S., Sayal, A., Simões, M., Quental, H., Bernardino, I., et al. (2021). Training the social brain: clinical and neural effects of an 8-week real-time functional magnetic resonance imaging neurofeedback phase IIa clinical trial in autism. Autism 25, 1746–1760. doi: 10.1177/13623613211002052
Disner, S. G., Beevers, C. G., Haigh, E. A. P., and Beck, A. T. (2011). Neural mechanisms of the cognitive model of depression. Nat. Rev. Neurosci. 12, 467–477. doi: 10.1038/nrn3027
Domínguez-Borràs, J., and Vuilleumier, P. (2022). Amygdala function in emotion, cognition, and behavior. Handb. Clin. Neurol. 187:8. doi: 10.1016/b978-0-12-823493-8.00015-8
Etkin, A., and Wager, T. D. (2007). Functional neuroimaging of anxiety: a Meta-analysis of emotional processing in PTSD, social anxiety disorder, and specific phobia. Am. J. Psychiatry 164, 1476–1488. doi: 10.1176/appi.ajp.2007.07030504
Friston, K. J., Harrison, L., and Penny, W. (2003). Dynamic causal modelling. Neuroimage 19, 1273–1302. doi: 10.1016/S1053-8119(03)00202-7
Frith, C. D., and Frith, U. (2006). How we predict what other people are going to do. Brain Res. 1079, 36–46. doi: 10.1016/j.brainres.2005.12.126
Geissberger, N., Tik, M., Sladky, R., Woletz, M., Schuler, A.-L., Willinger, D., et al. (2020). Reproducibility of amygdala activation in facial emotion processing at 7T. Neuroimage 211:116585. doi: 10.1016/j.neuroimage.2020.116585
Germine, L. T., Garrido, L., Bruce, L., and Hooker, C. (2011). Social anhedonia is associated with neural abnormalities during face emotion processing. Neuroimage 58, 935–945. doi: 10.1016/j.neuroimage.2011.06.059
Grupe, D. W., and Nitschke, J. B. (2013). Uncertainty and anticipation in anxiety: an integrated neurobiological and psychological perspective. Nat. Rev. Neurosci. 14, 488–501. doi: 10.1038/nrn3524
Hahn, A., Kranz, G. S., Seidel, E. M., Sladky, R., Kraus, C., Küblböck, M., et al. (2013). Comparing neural response to painful electrical stimulation with functional MRI at 3 and 7T. Neuroimage 82, 336–343. doi: 10.1016/j.neuroimage.2013.06.010
Hartwell, K. J., Hanlon, C. A., Li, X., Borckardt, J. J., Canterberry, M., Prisciandaro, J. J., et al. (2016). Individualized real-time fMRI neurofeedback to attenuate craving in nicotine-dependent smokers. J. Psychiatry Neurosci. 41, 48–55. doi: 10.1503/jpn.140200
Haugg, A., Manoliu, A., Sladky, R., Hulka, L. M., Kirschner, M., Brühl, A. B., et al. (2022). Disentangling craving- and valence-related brain responses to smoking cues in individuals with nicotine use disorder. Addict. Biol. 27:83. doi: 10.1111/adb.13083
Haugg, A., Renz, F. M., Nicholson, A. A., Lor, C., Götzendorfer, S. J., Sladky, R., et al. (2021). Predictors of real-time fMRI neurofeedback performance and improvement – a machine learning mega-analysis. Neuroimage 237:118207. doi: 10.1016/j.neuroimage.2021.118207
Haxby, J. V., Hoffman, E. A., and Gobbini, M. I. (2000). The distributed human neural system for face perception. Trends Cogn. Sci. 4, 223–233. doi: 10.1016/s1364-6613(00)01482-0
Hellrung, L., Dietrich, A., Hollmann, M., Pleger, B., Kalberlah, C., Roggenhofer, E., et al. (2018). Intermittent compared to continuous real-time fMRI neurofeedback boosts control over amygdala activation. Neuroimage 166, 198–208. doi: 10.1016/j.neuroimage.2017.10.031
Herwig, U., Lutz, J., Scherpiet, S., Scheerer, H., Kohlberg, J., Opialla, S., et al. (2019). Training emotion regulation through real-time fMRI neurofeedback of amygdala activity. Neuroimage 184, 687–696. doi: 10.1016/j.neuroimage.2018.09.068
Herwig, U., Opialla, S., Cattapan, K., Wetter, T. C., Jäncke, L., and Brühl, A. B. (2018). Emotion introspection and regulation in depression. Psychiatry Res. Neuroimaging 277, 7–13. doi: 10.1016/j.pscychresns.2018.04.008
Hofmann, S. G., Ellard, K. K., and Siegle, G. J. (2012). Neurobiological correlates of cognitions in fear and anxiety: a cognitive–neurobiological information-processing model. Cognit. Emot. 26, 282–299. doi: 10.1080/02699931.2011.579414
Ihssen, N., Sokunbi, M. O., Lawrence, A. D., Lawrence, N. S., and Linden, D. E. J. (2017). Neurofeedback of visual food cue reactivity: a potential avenue to alter incentive sensitization and craving. Brain Imaging Behav. 11, 915–924. doi: 10.1007/s11682-016-9558-x
Inman, C. S., Bijanki, K. R., Bass, D. I., Gross, R. E., Hamann, S., and Willie, J. T. (2020). Human amygdala stimulation effects on emotion physiology and emotional experience. Neuropsychologia 145:106722. doi: 10.1016/j.neuropsychologia.2018.03.019
Joormann, J., and Siemer, M. (2014). “Emotion regulation in mood disorders” in Handbook of emotion regulation. ed. J. J. Gross. 2nd ed (New York, NY: Guilford Press), 3–20.
Kanwisher, N., McDermott, J., and Chun, M. M. (1997). The fusiform face area: a module in human Extrastriate cortex specialized for face perception. J. Neurosci. 17, 4302–4311. doi: 10.1523/JNEUROSCI.17-11-04302.1997
Karlsson, K. A., Windischberger, C., Gerstl, F., Mayr, W., Siegel, J. M., and Moser, E. (2010). Modulation of hypothalamus and amygdalar activation levels with stimulus valence. Neuro Image 51, 324–328. doi: 10.1016/j.neuroimage.2010.02.029
Kirschner, M., Sladky, R., Haugg, A., Stämpfli, P., Jehli, E., Hodel, M., et al. (2018). Self-regulation of the dopaminergic reward circuit in cocaine users with mental imagery and neurofeedback. EBioMedicine 37, 489–498. doi: 10.1016/j.ebiom.2018.10.052
Koob, G. F., and Le Moal, M. (1997). Drug abuse: hedonic homeostatic dysregulation. Science 278, 52–58. doi: 10.1126/science.278.5335.52
Koush, Y., Ashburner, J., Prilepin, E., Sladky, R., Zeidman, P., Bibikov, S., et al. (2017a). Open NFT: an open-source Python/Matlab framework for real-time fMRI neurofeedback training based on activity, connectivity and multivariate pattern analysis. Neuroimage 156, 489–503. doi: 10.1016/j.neuroimage.2017.06.039
Koush, Y., Meskaldji, D. E., Pichon, S., Rey, G., Rieger, S. W., Linden, D. E. J., et al. (2017b). Learning control over emotion networks through connectivity-based neurofeedback. Cereb. Cortex 27, 1193–1202. doi: 10.1093/cercor/bhv311
Krylova, M., Skouras, S., Razi, A., Nicholson, A. A., Karner, A., Steyrl, D., et al. (2021). Progressive modulation of resting-state brain activity during neurofeedback of positive-social emotion regulation networks. Sci. Rep. 11:23363. doi: 10.1038/s41598-021-02079-4
Langner, O., Dotsch, R., Bijlstra, G., Wigboldus, D. H. J., Hawk, S. T., and van Knippenberg, A. (2010). Presentation and validation of the Radboud faces database. Cognit. Emot. 24, 1377–1388. doi: 10.1080/02699930903485076
Lichtenberg, N. T., Pennington, Z. T., Holley, S. M., Greenfield, V. Y., Cepeda, C., Levine, M. S., et al. (2017). Basolateral amygdala to orbitofrontal cortex projections enable Cue-triggered reward expectations. J. Neurosci. 37, 8374–8384. doi: 10.1523/JNEUROSCI.0486-17.2017
Linden, D. E. J., Habes, I., Johnston, S. J., Linden, S., Tatineni, R., Subramanian, L., et al. (2012). Real-time self-regulation of emotion networks in patients with depression. PloS One 7:e38115. doi: 10.1371/journal.pone.0038115
Mac Duffie, K. E., Mac Innes, J., Dickerson, K. C., Eddington, K. M., Strauman, T. J., and Adcock, R. A. (2018). Single session real-time fMRI neurofeedback has a lasting impact on cognitive behavioral therapy strategies. Neuro Image. Clin 19, 868–875. doi: 10.1016/j.nicl.2018.06.009
Mathiak, K. A., Alawi, E. M., Koush, Y., Dyck, M., Cordes, J. S., Gaber, T. J., et al. (2015). Social reward improves the voluntary control over localized brain activity in fMRI-based neurofeedback training. Front. Behav. Neurosci. 9:136. doi: 10.3389/fnbeh.2015.00136
Mathiak, K. A., Koush, Y., Dyck, M., Gaber, T. J., Alawi, E. A., Zepf, F. D., et al. (2010). Social reinforcement can regulate localized brain activity. Eur. Arch. Psychiatry Clin. Neurosci. 260, 132–136. doi: 10.1007/s00406-010-0135-9
McMakin, D. L., Olino, T. M., Porta, G., Dietz, L. J., Emslie, G., Clarke, G., et al. (2012). Anhedonia predicts poorer recovery among youth with selective serotonin reuptake inhibitor treatment-resistant depression. J. Am. Acad. Child Adolesc. Psychiatry 51, 404–411. doi: 10.1016/j.jaac.2012.01.011
Minkova, L., Sladky, R., Kranz, G. S., Woletz, M., Geissberger, N., Kraus, C., et al. (2017). Task-dependent modulation of amygdala connectivity in social anxiety disorder. Psychiatry Res. Neuroimaging 262, 39–46. doi: 10.1016/j.pscychresns.2016.12.016
Momen, N. C., Plana-Ripoll, O., Agerbo, E., Benros, M. E., Børglum, A. D., Christensen, M. K., et al. (2020). Association between mental disorders and subsequent medical conditions. N. Engl. J. Med. 382, 1721–1731. doi: 10.1056/NEJMoa1915784
Morawetz, C., Bode, S., Baudewig, J., Kirilina, E., and Heekeren, H. R. (2016). Changes in effective connectivity between dorsal and ventral prefrontal regions moderate emotion regulation. Cereb. Cortex 26, 1923–1937. doi: 10.1093/cercor/bhv005
Mulholland, T., Boudrot, R., and Davidson, A. (1979). Feedback delay and amplitude threshold and control of the occipital EEG. Biofeedback Self Regul. 4, 93–102. doi: 10.1007/BF01007104
Mulholland, T., and Eberlin, P. (1977). Effect of feedback contingencies on the control of occipital alpha. Biofeedback Self Regul. 2, 43–57. doi: 10.1007/BF01001719
Nicholson, A. A., Friston, K. J., Zeidman, P., Harricharan, S., McKinnon, M. C., Densmore, M., et al. (2017). Dynamic causal modeling in PTSD and its dissociative subtype: bottom–up versus top–down processing within fear and emotion regulation circuitry. Hum. Brain Mapp. 38, 5551–5561. doi: 10.1002/hbm.23748
Nicholson, A. A., Ros, T., Frewen, P. A., Densmore, M., Théberge, J., Kluetsch, R. C., et al. (2016). Alpha oscillation neurofeedback modulates amygdala complex connectivity and arousal in posttraumatic stress disorder. Neuroimage Clin 12, 506–516. doi: 10.1016/j.nicl.2016.07.006
Nordentoft, M., Wahlbeck, K., Hällgren, J., Westman, J., Ösby, U., Alinaghizadeh, H., et al. (2013). Excess mortality, causes of death and life expectancy in 270,770 patients with recent onset of mental disorders in Denmark, Finland and Sweden. PloS One 8:e55176. doi: 10.1371/journal.pone.0055176
Paret, C., Goldway, N., Zich, C., Keynan, J. N., Hendler, T., Linden, D., et al. (2019). Current progress in real-time functional magnetic resonance-based neurofeedback: methodological challenges and achievements. Neuroimage 202:116107. doi: 10.1016/j.neuroimage.2019.116107
Paret, C., Kluetsch, R., Ruf, M., Demirakca, T., Hoesterey, S., Ende, G., et al. (2014). Down-regulation of amygdala activation with real-time fMRI neurofeedback in a healthy female sample. Front. Behav. Neurosci. 8:299. doi: 10.3389/fnbeh.2014.00299
Paret, C., Ruf, M., Gerchen, M. F., Kluetsch, R., Demirakca, T., Jungkunz, M., et al. (2016). fMRI neurofeedback of amygdala response to aversive stimuli enhances prefrontal–limbic brain connectivity. Neuroimage 125, 182–188. doi: 10.1016/j.neuroimage.2015.10.027
Paret, C., Zähringer, J., Ruf, M., Gerchen, M. F., Mall, S., Hendler, T., et al. (2018). Monitoring and control of amygdala neurofeedback involves distributed information processing in the human brain. Hum. Brain Mapp. 39, 3018–3031. doi: 10.1002/hbm.24057
Penny, W. D., Stephan, K. E., Daunizeau, J., Rosa, M. J., Friston, K. J., Schofield, T. M., et al. (2010). Comparing families of dynamic causal models. PLoS Comput. Biol. 6:e1000709. doi: 10.1371/journal.pcbi.1000709
Pessoa, L., and Adolphs, R. (2010). Emotion processing and the amygdala: from a “low road” to “many roads” of evaluating biological significance. Nat. Rev. Neurosci. 11, 773–782. doi: 10.1038/nrn2920
Phan, K. L., Fitzgerald, D. A., Nathan, P. J., and Tancer, M. E. (2006). Association between amygdala hyperactivity to harsh faces and severity of social anxiety in generalized social phobia. Biol. Psychiatry 59, 424–429. doi: 10.1016/j.biopsych.2005.08.012
Philip, L., Martin, J.-C., and Clavel, C. (2018). Rapid facial reactions in response to facial expressions of emotion displayed by real versus virtual faces. Iperception 9:204166951878652. doi: 10.1177/2041669518786527
Plichta, M. M., Schwarz, A. J., Grimm, O., Morgen, K., Mier, D., Haddad, L., et al. (2012). Test–retest reliability of evoked BOLD signals from a cognitive–emotive fMRI test battery. Neuroimage 60, 1746–1758. doi: 10.1016/j.neuroimage.2012.01.129
Pope, A. T., Bogart, E. H., and Bartolome, D. S. (1995). Biocybernetic system evaluates indices of operator engagement in automated task. Biol. Psychol. 40, 187–195. doi: 10.1016/0301-0511(95)05116-3
R Core Team, (2022). R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. Available at: https://www.R-project.org/.
Ramot, M., Kimmich, S., Gonzalez-Castillo, J., Roopchansingh, V., Popal, H., White, E., et al. (2017). Direct modulation of aberrant brain network connectivity through real-time neuro feedback. Elife 6:e28974. doi: 10.7554/eLife.28974
Robinson, T. E., and Berridge, K. C. (2000). The psychology and neurobiology of addiction: an incentive–sensitization view. Addiction 95, 91–117. doi: 10.1080/09652140050111681
Ros, T., Enriquez-Geppert, S., Zotev, V., Young, K. D., Wood, G., Whitfield- Gabrieli, S., et al. (2020). Conensus on the reporting and experimental design of clinical and cognitive behavioural neurofeedback studies (CRED-nf checklist). Brain. 143, 1674–1685. doi: 10.1093/brain/awaa009
Roy, M., Shohamy, D., and Wager, T. D. (2012). Ventromedial prefrontal-subcortical systems and the generation of affective meaning. Trends Cogn. Sci. 16, 147–156. doi: 10.1016/j.tics.2012.01.005
Sabatinelli, D., Fortune, E. E., Li, Q., Siddiqui, A., Krafft, C., Oliver, W. T., et al. (2011). Emotional perception: Meta-analyses of face and natural scene processing. Neuroimage 54, 2524–2533. doi: 10.1016/j.neuroimage.2010.10.011
Santomauro, D. F., Mantilla Herrera, A. M., Shadid, J., Zheng, P., Ashbaugh, C., Pigott, D. M., et al. (2021). Global prevalence and burden of depressive and anxiety disorders in 204 countries and territories in 2020 due to the COVID-19 pandemic. Lancet 398, 1700–1712. doi: 10.1016/S0140-6736(21)02143-7
Sarkheil, P., Zilverstand, A., Kilian-Hütten, N., Schneider, F., Goebel, R., and Mathiak, K. (2015). fMRI feedback enhances emotion regulation as evidenced by a reduced amygdala response. Behav. Brain Res. 281, 326–332. doi: 10.1016/j.bbr.2014.11.027
Schneier, F. R., Kent, J. M., Star, A., and Hirsch, J. (2009). Neural circuitry of submissive behavior in social anxiety disorder: a preliminary study of response to direct eye gaze. Psychiatry Res. Neuroimaging 173, 248–250. doi: 10.1016/j.pscychresns.2008.06.004
Siegle, G. J., Thompson, W., Carter, C. S., Steinhauer, S. R., and Thase, M. E. (2007). Increased amygdala and decreased dorsolateral prefrontal BOLD responses in unipolar depression: related and independent features. Biol. Psychiatry 61, 198–209. doi: 10.1016/j.biopsych.2006.05.048
Sitaram, R., Ros, T., Stoeckel, L., Haller, S., Scharnowski, F., Lewis-Peacock, J., et al. (2017). Closed-loop brain training: the science of neurofeedback. Nat. Rev. Neurosci. 18, 86–100. doi: 10.1038/nrn.2016.164
Sladky, R., Baldinger, P., Kranz, G. S., Tröstl, J., Höflich, A., Lanzenberger, R., et al. (2013). High-resolution functional MRI of the human amygdala at 7T. Eur. J. Radiol. 82, 728–733. doi: 10.1016/j.ejrad.2011.09.025
Sladky, R., Geissberger, N., Pfabigan, D. M., Kraus, C., Tik, M., Woletz, M., et al. (2018). Unsmoothed functional MRI of the human amygdala and bed nucleus of the stria terminalis during processing of emotional faces. Neuroimage 168, 383–391. doi: 10.1016/j.neuroimage.2016.12.024
Sladky, R., Hahn, A., Karl, I. L., Geissberger, N., Kranz, G. S., Tik, M., et al. (2022). Dynamic causal modeling of the prefrontal/amygdala network during processing of emotional faces. Brain Connect. 12, 670–682. doi: 10.1089/brain.2021.0073
Sladky, R., Hoflich, A., Atanelov, J., Kraus, C., Baldinger, P., Moser, E., et al. (2012). Increased neural habituation in the amygdala and orbitofrontal cortex in social anxiety disorder revealed by fMRI. PloS One 7:e50050. doi: 10.1371/journal.pone.0050050
Sladky, R., Höflich, A., Küblböck, M., Kraus, C., Baldinger, P., Moser, E., et al. (2015). Disrupted effective connectivity between the amygdala and orbitofrontal cortex in social anxiety disorder during emotion discrimination revealed by dynamic causal modeling for fMRI. Cereb. Cortex 25, 895–903. doi: 10.1093/cercor/bht279
Sladky, R., Kargl, D., Haubensak, W., and Lamm, C. (2023). An active inference perspective for the amygdala complex. Trends Cogn. Sci. 15, S1364–6613. doi: 10.1016/j.tics.2023.11.004
Sorger, B., Scharnowski, F., Linden, D. E. J., Hampson, M., and Young, K. D. (2019). Control freaks: towards optimal selection of control conditions for fMRI neurofeedback studies. Neuroimage 186, 256–265. doi: 10.1016/j.neuroimage.2018.11.004
Steel, Z., Marnane, C., Iranpour, C., Chey, T., Jackson, J. W., Patel, V., et al. (2014). The global prevalence of common mental disorders: a systematic review and meta-analysis 1980–2013. Int. J. Epidemiol. 43, 476–493. doi: 10.1093/ije/dyu038
Stein, M. B., Goldin, P. R., Sareen, J., Zorrilla, L. T. E., and Brown, G. G. (2002). Increased amygdala activation to angry and contemptuous faces in generalized social phobia. Arch. Gen. Psychiatry 59, 1027–1034. doi: 10.1001/archpsyc.59.11.1027
Sulzer, J., Haller, S., Scharnowski, F., Weiskopf, N., Birbaumer, N., Blefari, M. L., et al. (2013). Real-time fMRI neurofeedback: Progress and challenges. Neuroimage 76, 386–399. doi: 10.1016/j.neuroimage.2013.03.033
Suslow, T., Konrad, C., Kugel, H., Rumstadt, D., Zwitserlood, P., Schöning, S., et al. (2010). Automatic mood-congruent amygdala responses to masked facial expressions in major depression. Biol. Psychiatry 67, 155–160. doi: 10.1016/j.biopsych.2009.07.023
Taylor, J. E., Yamada, T., Kawashima, T., Kobayashi, Y., Yoshihara, Y., Miyata, J., et al. (2022). Depressive symptoms reduce when dorsolateral prefrontal cortex-precuneus connectivity normalizes after functional connectivity neurofeedback. Sci. Rep. 12:2581. doi: 10.1038/s41598-022-05860-1
Tillfors, M., Furmark, T., Marteinsdottir, I., and Fredrikson, M. (2002). Cerebral blood flow during anticipation of public speaking in social phobia: a PET study. Biol. Psychiatry 52, 1113–1119. doi: 10.1016/S0006-3223(02)01396-3
Toates, F. M. (1975). Control theory in biology and experimental psychology. London: Hutchinson Educational Ltd.
Tyszka, J. M., and Pauli, W. M. (2016). In vivo delineation of subdivisions of the human amygdaloid complex in a high-resolution group template. Hum. Brain Mapp. 37, 3979–3998. doi: 10.1002/hbm.23289
Victor, T. A., Furey, M. L., Fromm, S. J., Öhman, A., and Drevets, W. C. (2010). Relationship between amygdala responses to masked faces and mood state and treatment in major depressive disorder. Arch. Gen. Psychiatry 67, 1128–1138. doi: 10.1001/archgenpsychiatry.2010.144
Wang, S., Yu, R., Tyszka, J. M., Zhen, S., Kovach, C., Sun, S., et al. (2017). The human amygdala parametrically encodes the intensity of specific facial emotions and their categorical ambiguity. Nat. Commun. 8:14821. doi: 10.1038/ncomms14821
Watson, D., Clark, L. A., and Tellegen, A. (1988). Development and validation of brief measures of positive and negative affect: the PANAS scales. J. Pers. Soc. Psychol. 54, 1063–1070. doi: 10.1037//0022-3514.54.6.1063
Weightman, M. J., Air, T. M., and Baune, B. T. (2014). A review of the role of social cognition in major depressive disorder. Front. Psych. 5:179. doi: 10.3389/fpsyt.2014.00179
Willinger, D., Karipidis, I. I., Beltrani, S., Di Pietro, S. V., Sladky, R., Walitza, S., et al. (2019). Valence-dependent coupling of prefrontal-amygdala effective connectivity during facial affect processing. eNeuro 6:ENEURO.0079-19.2019. doi: 10.1523/ENEURO.0079-19.2019
Wittchen, H. U., Jacobi, F., Rehm, J., Gustavsson, A., Svensson, M., Jönsson, B., et al. (2011). The size and burden of mental disorders and other disorders of the brain in Europe 2010. Eur. Neuropsychopharmacol. 21, 655–679. doi: 10.1016/j.euroneuro.2011.07.018
World Health Organization. (2022). Mental Disorders. https://www.who.int/news-room/fact-sheets/detail/mental-disorders (Accessed 8 June 2022).
Young, K. D., Bodurka, J., and Drevets, W. C. (2016). Differential neural correlates of autobiographical memory recall in bipolar and unipolar depression. Bipolar Disord. 18, 571–582. doi: 10.1111/bdi.12441
Young, K. D., Siegle, G. J., Zotev, V., Phillips, R., Misaki, M., Yuan, H., et al. (2017). Randomized clinical trial of real-time fMRI amygdala neurofeedback for major depressive disorder: effects on symptoms and autobiographical memory recall, in. Am. J. Psychiatry 174, 748–755. doi: 10.1176/appi.ajp.2017.16060637
Zaehringer, J., Jennen-Steinmetz, C., Schmahl, C., Ende, G., and Paret, C. (2020). Psychophysiological effects of downregulating negative emotions: insights from a Meta-analysis of healthy adults. Front. Psychol. 11:470. doi: 10.3389/fpsyg.2020.00470
Zich, C., Johnstone, N., Lührs, M., Lisk, S., Haller, S. P. W., Lipp, A., et al. (2020). Modulatory effects of dynamic fMRI-based neurofeedback on emotion regulation networks in adolescent females. Neuroimage 220:117053. doi: 10.1016/j.neuroimage.2020.117053
Zotev, V., Yuan, H., Misaki, M., Phillips, R., Young, K. D., Feldner, M. T., et al. (2016). Correlation between amygdala BOLD activity and frontal EEG asymmetry during real-time fMRI neurofeedback training in patients with depression. Neuroimage Clin 11, 224–238. doi: 10.1016/j.nicl.2016.02.003
Keywords: real-time fMRI, neurofeedback, amygdala, emotion regulation, dynamic faces
Citation: Watve A, Haugg A, Frei N, Koush Y, Willinger D, Bruehl AB, Stämpfli P, Scharnowski F and Sladky R (2024) Facing emotions: real-time fMRI-based neurofeedback using dynamic emotional faces to modulate amygdala activity. Front. Neurosci. 17:1286665. doi: 10.3389/fnins.2023.1286665
Edited by:
Kymberly Young, University of Pittsburgh, United StatesReviewed by:
Miguel Castelo-Branco, Coimbra Institute for Biomedical Imaging and Translational Research (CIBIT), PortugalCopyright © 2024 Watve, Haugg, Frei, Koush, Willinger, Bruehl, Stämpfli, Scharnowski and Sladky. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Ronald Sladky, cm9uYWxkLnNsYWRreUB1bml2aWUuYWMuYXQ=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.