 Ai Hoshino
Ai Hoshino Naoto Takahashi2
Naoto Takahashi2 Masashi Mizuguchi
Masashi Mizuguchi- 1Department of Developmental Medical Sciences, Graduate School of Medicine, The University of Tokyo, Tokyo, Japan
- 2Department of Pediatrics, The University of Tokyo Hospital, Tokyo, Japan
- 3Department of Neuropediatrics, Tokyo Metropolitan Neurological Hospital, Fuchu, Japan
- 4Department of Neurology, Saitama Children's Medical Center, Saitama, Japan
- 5Department of Pediatrics, National Rehabilitation Center for Children with Disabilities, Tokyo, Japan
Objective: Acute necrotizing encephalopathy (ANE) is a severe complication of infectious diseases affecting the brain and systemic organs. The main pathogenesis is cytokine storm, in which interleukin-6 (IL-6) and interleukin-10 (IL-10) are candidates for key cytokines. To further elucidate their roles in the etiology and pathogenesis of ANE, we studied polymorphisms in the promotor regions of the IL6 and IL10 genes by genetic and functional analyses.
Methods: We first conducted a case–control association study of four IL6 and three IL10 polymorphisms. We genotyped 31 Japanese ANE cases and compared the results with those of approximately 200 Japanese controls. For the two polymorphisms showing a possible association, we next studied whether the polymorphisms alter the production of IL-6 or IL-10 by lymphoblasts upon phorbol 12-myristate 13-acetate (PMA) stimulation.
Results: The frequencies of IL6 rs1800796G allele and IL10 rs1800871/rs1800872 CC/CC diplotype were significantly higher in ANE cases than in controls. The IL10 CC/CC diplotype was associated with low IL-10 production, whereas the IL6 GG genotype was not associated with IL-6 production.
Conclusion: IL10 rs1800871/rs1800872 CC/CC diplotype may predispose Japanese children to ANE by altering IL-10 production in the early phase of infection. Etio-pathogenetic significance of IL6 rs1800796G remains to be elucidated.
1. Introduction
Acute necrotizing encephalopathy (ANE) is a rare and fulminant syndrome of acute infection-triggered encephalopathy characterized pathologically by bilateral symmetric lesions in the thalamus and/or brainstem of vasogenic brain edema/necrosis (Mizuguchi et al., 1995). Antecedent infections of ANE are common viral diseases with high fever, such as influenza, exanthem subitum, rotavirus gastroenteritis and COVID-19 (Hoshino et al., 2012; Kasai et al., 2020; Sakuma et al., 2023). Being most common in childhood, ANE is an important cause of children’s deaths and severe neurological handicaps, in particular during influenza seasons.
ANE is a complex disorder in which multiple genetic and environmental factors are involved (Mizuguchi et al., 2023). ANE1, a familial and recurrent form of ANE prevalent in Caucasian ethnicities, is transmitted in an autosomal dominant fashion. Genetic studies have identified missense, loss-of-function mutations of the RANBP2 gene as the main cause of ANE1 (Neilson et al., 2009). On the other hand, sporadic and non-recurrent ANE is most common in east Asian ethnicities. As a part of genetic susceptibility to sporadic ANE, a study in Japan have identified specific human leukocyte antigen (HLA) genotypes, DRB1*09:01 and DQB1*03:03 (Hoshino et al., 2016). However, much remains to be elucidated regarding the genetic susceptibility of ANE.
Despite the term “encephalopathy,” clinical course of ANE is characterized by profound systemic inflammation and multiorgan dysfunction: high fever and damages to the liver, kidney and muscle in most cases, as well as shock, disseminated intravascular coagulation and multiorgan failure in very severe cases. All these findings are consistent with “cytokine storm syndrome” (Mizuguchi et al., 2007). Previous studies have measured the levels of cytokines and other biomarkers in the blood and/or cerebrospinal fluid of sporadic ANE cases. Of the variable findings noted by these studies, an increase of serum interleukin-6 (IL-6) and that of serum interleukin-10 (IL-10) are very common, early and remarkable changes (Ichiyama et al., 2003; Akiyoshi et al., 2006; Mizuguchi et al., 2007; Kansagra and Gallentine, 2011; Kawamura et al., 2013; Okajima et al., 2022), suggesting important roles of these two cytokines in the pathophysiology of ANE.
To further explore the genetic background of ANE, we herein conducted a case–control study of single nucleotide polymorphisms (SNPs) in the promotor regions of the IL6 and IL10 gene that may alter the expression of these cytokines. For the candidate susceptibility variants obtained by the genetic studies, we did a functional analysis of their effects on cytokine production.
2. Materials and methods
2.1. Patients and controls
We recruited patients with ANE from hospitals throughout Japan from 2008 to 2019 based on the diagnostic criteria consisting of clinical course and characteristic MRI finding of symmetric brain lesions in the bilateral thalamus (Hoshino et al., 2012). Thirty-one patients, 15 males and 16 females aged from 8 months to 9 years and 7 months (median: 2 years and 2 months), were enrolled in the study. All patients were Japanese and mutually unrelated. None of them had a pathogenic mutation in the RANBP2 gene. We obtained written informed consent from all guardians of patients participating in the study. This study was approved by the Ethics Committee of the Graduate School of Medicine, the University of Tokyo (No. G-3504).
We analyzed the IL6 and IL10 genotypes of control subjects, consisting of 100 healthy Japanese adults, 50 males and 50 females, 20 to 69 years of age, using DNA extracted from Pharma SNP Consortium B cell lines (PSC B cell lines) supplied by the Human Science Research Resources Bank (Osaka, Japan). We used the combined data of 100 Japanese controls from the Pharma SNP consortium and HapMap SNPs from the Japanese (JPT) Population database in International HapMap project for the variation frequencies of SNPs in IL6 and IL10 promoter regions.
2.2. Genotyping of IL6 and IL10 polymorphisms
Peripheral blood samples were collected from the patients. Genomic DNA was extracted from the blood samples using standard protocols. We analyzed four SNPs in the IL6 promoter regions, rs1800797, rs1800796, rs2069829 and rs1800795, and three SNPs in the IL10 promoter regions, rs1800896, rs1800871 and rs1800872. Two of them, rs1800871 and rs1800872, were in complete linkage disequilibrium in Japanese. Polymerase chain reaction (PCR) amplification of each IL6 and IL10 promoter region including the seven SNPs was performed using AmpliTaq PCR kits (Applied Biosystems). The reaction mixture contained 2 μL buffer, 2 μL of 2 mM dNTP, 1 μL forward and reverse primers (10 pmol), 0.12 μL AmpliTaq and 1 μL genomic DNA (30 ng). Primer sequences were the same as those used in a previous study (Chou et al., 2010) (Table 1). All the PCR products were purified with a PCR product sequencing kit (Amersham Biosciences, Little Chalfont, Buckinghamshire, United Kingdom), and were reacted with the Big Dye Terminator FS Ready Reaction kit (Applied Biosystems, Foster City, CA, United States). Purified PCR products were sequenced on 310 Genetic Analyzer, 3,100 Genetic Analyzer, or 3130xl Genetic Analyzer (Life Technologies, Carlsbad, CA, United States).
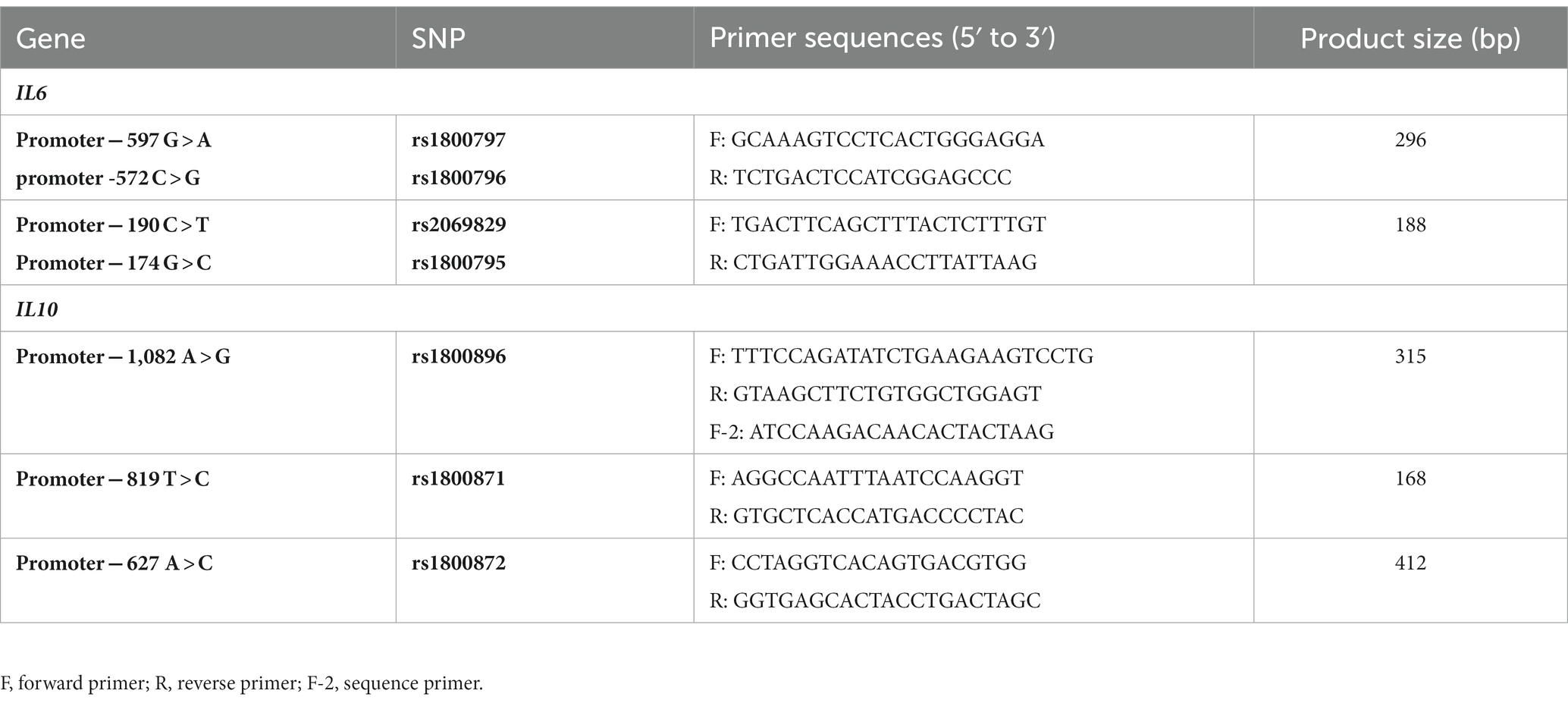
Table 1. Primer sequences.
2.3. Quantification of IL-6 and IL-10 production
Lymphoblasts from Japanese healthy controls (PSC B cell lines) were genotyped to identify three SNPs of IL6 rs1800796, IL10 rs1800871 and IL10 rs1800872. Thirty-two lymphoblast samples were used for IL-6 quantification analysis and classified into three genotype groups, CC, CG and GG, of IL6 rs1800796. The other 25 lymphoblast samples were used for IL-10 quantification analysis and classified into three diplotype groups, TA/TA, TA/CC and CC/CC, of IL10 rs1800871/rs1800872.
Lymphoblastoid cells were cultured in RPMI 1640 medium containing inactivated 10% fetal bovine serum. Cell lines were dispensed into 6 well-cell plates with a constant cell count of 2.5 × 105 cells/well. The three IL6 genotypes and three IL10 diplotypes were cultured in two wells per cell line with or without adding phorbol 12-myristate 13-acetate (PMA), a potent stimulant for lymphocytes mimicking inflammation. PMA stimulation was performed at 20 ng/mL and the conditions were adopted from a previous study (Yokoi et al., 1990). Cell plate was cultured for 48 h in 5% CO2 incubator. The ratio of IL-6 and IL-10 production in secretory supernatants from lymphoblast cell lines with and without PMA stimulation was analyzed among the three IL6 genotype or IL10 diplotype groups. IL-6 and IL-10 assays and quantification analysis were measured by using Bio-Plex™ Assay Kits and Bio-Plex™ system (Bio-Rad). Cytokine data analysis was performed using Bio-Plex Manager Software, ver. 6.1, following the procedure in the instruction manual of Bio-Plex™ Suspension Array System (171-00201JA).
2.4. Statistical analysis
We conducted a case–control association study of SNPs in the IL6 and IL10 promoter region using Chi-square test or Fisher’s exact test. The statistical analysis was conducted using Bell Curve for Excel (ver. 3.21; Social Survey Research information, Tokyo, Japan). The corrected p-values (Pc) were obtained by multiplying the uncorrected p-value with the number of comparisons, according to Bonferroni’s methods. A Pc-value <0.05 was considered as statistically significant. For quantification of IL-6 and IL-10 levels, the Steel-Dwass test was used, and a p-value <0.05 was considered as statistically significant.
3. Results
3.1. IL6 and IL10 gene polymorphisms
Genotyping of IL6 polymorphisms revealed that the frequency of rs1800796 GG genotype was higher in ANE cases (9.7%) than in control subjects (4.2%). The frequency of G allele was significantly higher in ANE cases (37.1%) than in controls (21.0%) (odds ratio, 2.221; 95% confidence interval, 1.225–3.926; p = 0.0069; Pc = 0.048). The other IL6 polymorphisms, rs1800797, rs2069829 and rs1800795, showed no differences between ANE cases and controls (Table 2).
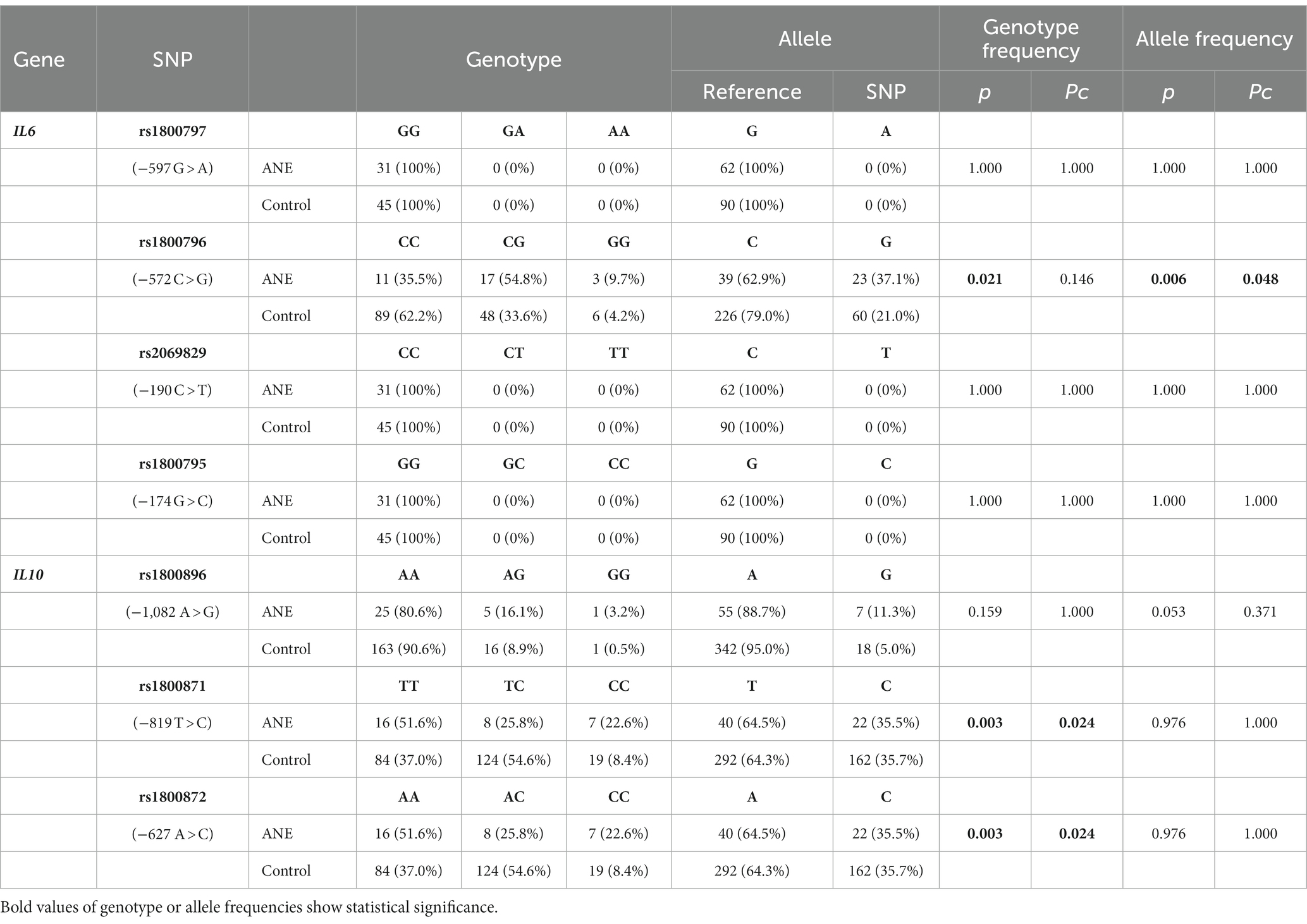
Table 2. Genotype and allele frequencies: Comparison between ANE cases and control subjects.
Genotyping of IL10 polymorphisms confirmed that two of the three common variants in the IL10 promotor region, rs1800871 and rs1800872, show a complete linkage, as reported previously (Schuurhof et al., 2011; Zhang et al., 2012), resulting in rs1800871/rs1800872 TA and CC haplotypes, and in TA/TA, TA/CC and CC/CC diplotypes. The frequency of CC/CC diplotype was significantly higher in ANE (22.6%) than in control (8.4%) (CC/CC versus TA/CC and TA/TA: odds ratio, 3.193; 95% confidence interval, 1.245–8.515; p = 0.0137). The other IL10 polymorphism, rs1800896, showed no differences between ANE cases and controls (Table 2).
3.2. IL-6/IL-10 production and IL6/IL10 genotypes
We examined whether the IL6 rs1800796 and IL10 rs1800871/rs1800872 polymorphisms are associated with the production of IL-6 and IL-10 proteins, respectively, by lymphoblasts upon PMA stimulation. The levels of IL-6 production were comparable among the three IL6 rs1800796 genotypes, CC, CG and GG (p = 0.993 between CC and CG, p = 0.982 between CC and GG, and p = 0.979 between CG and GG) (Figure 1A). The levels of IL-10 production were different between the three IL10 rs1800871/rs1800872 diplotypes, being high in TA/TA, moderate in TA/CC and low in CC/CC. The difference between TA/TA and CC/CC was statistically significant (p = 0.508 between TA/TA and TA/CC, p = 0.040 between TA/TA and CC/CC, and p = 0.774 between TA/CC and CC/CC) (Figure 1B).
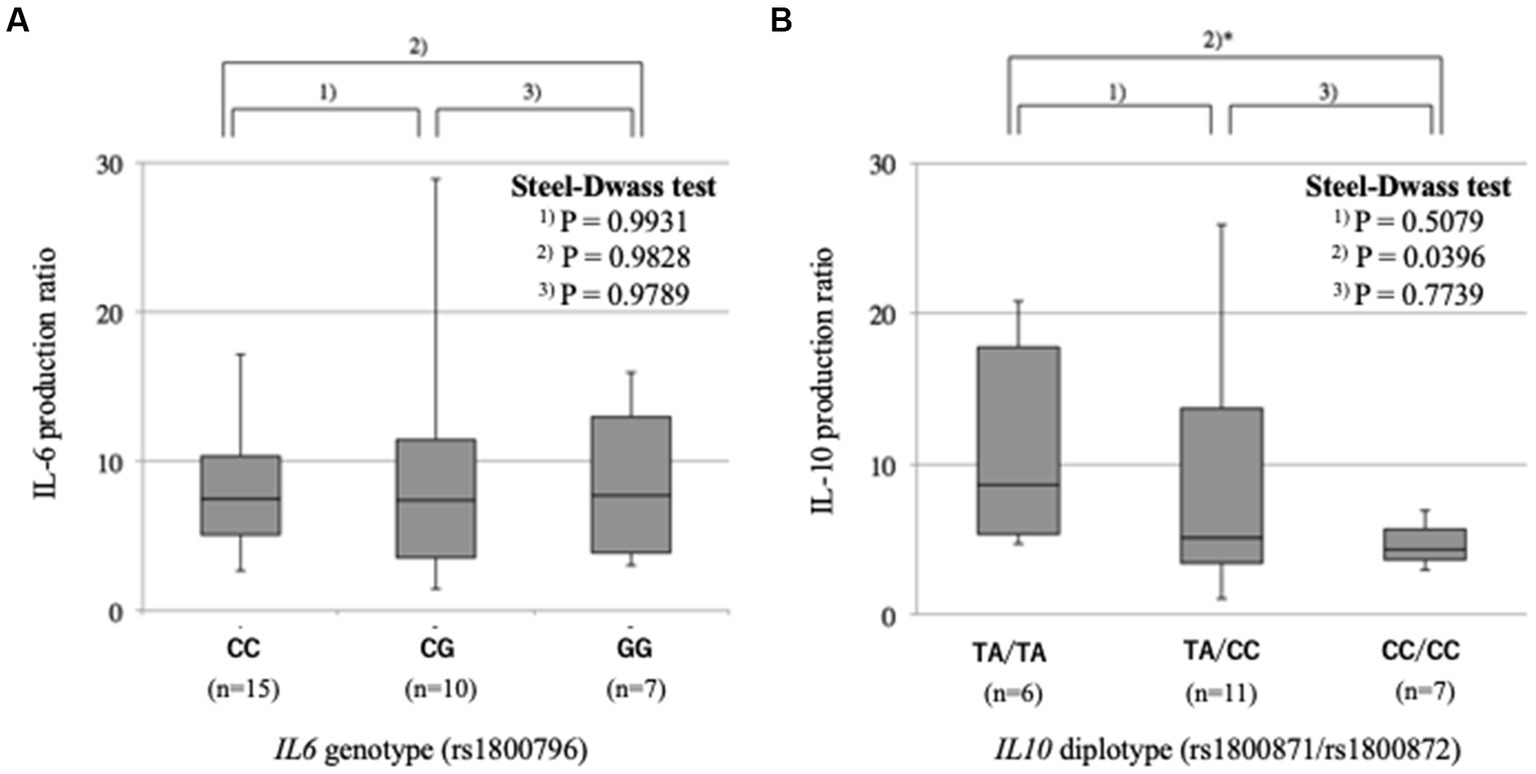
Figure 1. Effects of gene polymorphisms on cytokine production.IL-6 or IL-10 levels in cell supernatants were measured before and after stimulation of lymphoblasts by PMA. The ratios were compared between IL6 genotypes or IL10 diplotypes. (A) IL-6 production is comparable between the three IL6 rs1800796 genotypes. (B) IL-10 production is low in IL10 rs1800871/rs1800872 CC/CC diplotype.
4. Discussion
In this study, we first demonstrated by genetic analyses that the frequency of IL6 rs1800796 G allele and that of IL10 rs1800871/rs1800872 CC/CC diplotype were higher in ANE cases than in controls. However, statistical evidence provided by these analyses was not conclusive because the p-value for IL6 rs1800796 was of borderline significance (Pc = 0.048), and because the genotypic frequency status of IL10 rs1800871/rs1800872 in the control group was deviated from Hardy–Weinberg equilibrium. To further explore the pathogenetic significance of these polymorphisms, we next did functional analyses, and found that IL10 rs1800871/rs1800872 CC/CC diplotype was associated with low production by lymphoblasts of IL-10 upon PMA stimulation, and that IL6 rs1800796 allele G was not associated with IL-6 production upon PMA stimulation.
IL-6 is a proinflammatory cytokine secreted upon infection by various cells such as T lymphocytes, B lymphocytes, monocytes and endothelial cells. IL-6 plays a major role in immune responses such as differentiation/activation of T lymphocytes, enhancement of antibody production via proliferation of B lymphocytes, and induction of C reactive protein and other acute phase proteins in hepatocytes. In the promotor region of the IL6 gene, there are four polymorphisms, rs1800797, rs1800796, rs2069829 and rs1800795. Previous studies have demonstrated their association with the onset, severity and/or response to treatment of various diseases including infectious, autoimmune, cardiovascular, gastrointestinal, neoplastic and neurologic disorders. Of these common variants, rs1800796 was identified by this study as a possible genetic risk factor of ANE. In chronic infectious diseases, rs1800796G allele, GG genotype or G-containing haplotype is reportedly associated with a decreased risk of tuberculosis (Zhang et al., 2012; Wang et al., 2017), increased risk of anti-tuberculosis drug-induced hepatotoxicity (Li et al., 2018), decreased prevalence of chronic human hepatitis B virus (HBV) infection (Zhang et al., 2015; Li et al., 2021), decreased chance of sustained viral response to antiviral therapy against chronic hepatitis C virus (HCV) infection (Yee et al., 2009; Sghaier et al., 2017b), and a decreased risk of progression from HCV infection to hepatocellular carcinoma (Sghaier et al., 2017b). In acute infectious diseases, rs1800796G allele is reportedly associated with an increased risk of sepsis in chronic renal disease (Panayides et al., 2015) and increased risk of post-traumatic osteomyelitis (Jiang et al., 2020). In acute viral infections that trigger ANE in most cases, information is currently limited. An Iranian study observed no effect of rs1800796 on the severity of COVID-19, whereas a Chinese study noted that a haplotype containing rs1800796G may be associated with an increased severity (Chen et al., 2021). No data are currently available as to influenza, the commonest antecedent infection of ANE.
IL-6 expression may be altered by IL6 haplotypes/diplotypes consisting of multiple polymophisms. A previous study compared IL-6 production in the whole blood upon stimulation by lipopolysaccharide (LPS) between IL6 diplotypes consisting of three polymorphisms, rs1800797, rs1800796 and rs1800795, and found that GGG/GGG diplotype (allele G for all the three including rs1800796) is associated with lower IL-6 production compared to the other diplotypes (Muller-Steinhardt et al., 2007). Other studies also observed an association of rs1800796G with a low production of IL-6 by CD14(+) monocytes in response to HBV core antigen stimulation (Zhang et al., 2015) and to Mycobacterium tuberculosis 19-kDa lipoprotein (Zhang et al., 2012). All these findings are evidence that rs1800796G is a low producer of IL-6 protein. However, another study found low levels of methylation in IL-6-related CpG sites, suggesting a high producer (Smallwood et al., 2008). In this study, we found no differences in IL-6 production between rs1800796 genotypes. The discrepancy among the studies may indicate that genetic polymorphisms in the promotor region influence IL6 transcription not by a simple additive mechanism, but rather through complex interactions determined by haplotype (Terry et al., 2000). Whether rs1800796 and/or another polymorphism in linkage disequilibrium can alter IL-6 production in acute viral infections remains to be elucidated. The roles of IL-6 in the etiology and pathogenesis of ANE warrants vigorous investigation in view of the recent reports on the efficacy of an IL-6 antagonist, tocilizumab, in the treatment of ANE (Koh et al., 2019; Ho et al., 2023; Hosie et al., 2023).
IL-10 is an anti-inflammatory cytokine activated in an early phase of infection to suppress proinflammatory cytokines, thereby avoiding excessive immune responses and consequent tissue damages. IL-10 regulates immune function by suppressing production of interferon-γ by T lymphocytes and production of interleukin-1 (IL-1), IL-6, interleukin-2 (IL-2) and tumor necrosis factor α (TNFα) by macrophages (Sabat et al., 2010). In the promotor region of the IL10 gene, there are three SNPs, rs1800896, rs1800871 and rs1800872, associated with the onset, severity, progression and/or outcome of various diseases including autoimmune, allergic, neoplastic, gastrointestinal and respiratory diseases. This study showed that a combination of two SNPs in complete linkage, rs1800871/rs1800872CC/CC diplotype, may be a genetic risk factor of ANE. In various infectious diseases, rs1800871C and/or rs1800872C is reportedly associated with the onset of chronic infections: a decreased susceptibility to tuberculosis (Yu et al., 2019; Chen and Ma, 2020), increased susceptibility to leprosy (Cardona-Castro et al., 2012; Dos Santos et al., 2021), decreased susceptibility to herpes zoster (Haanpää et al., 2002), and increased susceptibility to HBV infection (Ye et al., 2020). These alleles are also associated with the course, progression, response to therapy and outcome: a better outcome of leprosy (Alvarado-Arnez et al., 2015), low infection intensity of Schistosoma mansoni in schistosomiasis (Mewamba et al., 2023), reduced risk of cardiomyopathy in Chagas disease (Grijalva et al., 2022), reduced risk of anemia in newborns in Plasmodium falciparum malaria infection (Lokossou et al., 2013), increased risk of Epstein–Barr virus (EBV)-associated hemophagocytic lymphocytosis (Tang et al., 2021), reduced risk of breast cancer after EBV infection (He et al., 2012), increased risk of AIDS-related non-Hodgkin lymphoma (Wong et al., 2010), increased risk of progressive liver disease in chronic hepatitis B (Miyazoe et al., 2002), reduced chance of seroclearance after antiviral treatment of chronic hepatitis B (Rybicka et al., 2020), poor reduced disease severity of chronic hepatitis C (Świątek-Kościelna et al., 2017), and reduced risk of hepatocellular carcinoma in chronic hepatitis C (Aroucha et al., 2016; Sghaier et al., 2017a). For acute infections, s1800871C and/or rs1800872C is reportedly associated with a decreased risk of dengue infection (Eloisa Monroy-Muñoz et al., 2023) and dengue hemorrhagic fever (Alagarasu et al., 2015), decreased risk of asthma after bronchiolitis in infancy (Korppi et al., 2017), and reduced severity (requiring admission to intensive care unit) of sepsis (Montoya-Ruiz et al., 2016). In acute viral infections, these SNPs were reportedly associated with neither the prevalence of influenza (Mehrbod et al., 2021) nor the prevalence, severity and outcome of COVID-19 (Avendaño-Félix et al., 2021; Karcioglu Batur and Hekim, 2021; Abbood et al., 2023).
IL-10 expression may be altered by these IL10 polymorphisms. Previous studies have demonstrated that IL10 GCC haplotype (rs1800896G, rs1800871C and rs1800872C) is associated with a high IL-10 production by peripheral blood mononuclear cells (PBMCs) in response to LPS stimulation (Crawley et al., 1999; Edwards-Smith et al., 1999). Other studies also showed the association of rs1800872C and ACC haplotype (rs1800896A, rs1800871C and rs1800872C) with a high plasma level of IL-10 (Ge et al., 2015). By contrast, the present study showed an association of rs1800871/rs1800872 diplotype CC/CC with a low IL-10 production by lymphoblasts in response to PMA stimulation. The direction of alteration observed in this study is apparently opposite to that in previous studies, which may be explained by differences in cell types used, ethnicities, methods of stimulation, and other polymorphisms in high linkage. Taken together with the genotyping data, the results of this study suggest the possibility that the IL10 rs1800871/rs1800872 polymorphism may predispose Japanese children by altering IL-10 production in the early phase of infection, which in turn leads to excessive immune responses, cytokine storm, then culminates in severe brain damage and multiorgan dysfunction.
This study had several limitations. First, the small sample size of ANE cases limited statistical power, precluding analyses of haplotypes consisting of three alleles. Due to the low incidence, fulminant clinical course and high mortality, it was very difficult to obtain a large number of ANE samples. However, to the best of our knowledge, our repository of genomic DNA is the only one in the world. Second, the control subjects, healthy Japanese adults, derived from three sources. Their validity as controls might have been affected by possible demographic differences among the three control groups and from the ANE group. Third, the use of lymphoblasts, not of PBMCs, in the cytokine production assay rendered the experimental condition quite different from the clinical situation of human viral infections. On the other hand, the experimental data were highly reproducible because of the homogeneity and stability of lymphoblast cell culture. Fourth, our results failed to account for the high incidence of ANE in Asia, since the frequencies if IL6 rs1800796G and IL10 rs1800781C are reportedly lower in Asians (<20 and 40%, respectively) than in Europeans (95 and 75%, respectively) (The 1000 Genomes Project Consortium, 2012; Yu et al., 2013).
In conclusion, we studied polymorphisms in the promotor region of the IL6 and IL10 gene, in an attempt to explore the roles of IL-6 and IL-10 as disease modifying factors of ANE. Genetic studies suggested possible associations of IL6 rs1800796 and IL10 rs1800781/rs1800782 with the onset of ANE. Functional analyses showed an altered expression of IL-10 by IL10 rs1800781/rs1800782, but not that of IL-6 by IL6 rs1800796. Taken together, the IL10 rs1800781/rs1800782 polymorphism is suggested as a genetic risk factor of ANE, whereas the pathogenetic significance of IL6 rs1800796 polymorphism remains to be elucidated.
Data availability statement
The original contributions presented in the study are included in the article/supplementary material, further inquiries can be directed to the corresponding author.
Ethics statement
The studies involving human participants were reviewed and approved by The Ethics Committee of the Graduate School of Medicine, the University of Tokyo. Written informed consent to participate in this study was provided by the participants’ legal guardian/next of kin.
Author contributions
AH and MM designed the study and collected samples. NT, AO, and MM supervised the study. AH conducted the experiments and interpreted the data. AH and MM wrote the manuscript. All authors contributed to the article and approved the submitted version.
Funding
This research was supported by Grants-in-Aid for Scientific Research, Nos. 18K15700 and 15H04842, from the Japan Society for the Promotion of Sciences, and a Grant-in-Aid for Policy Research on Intractable Diseases, No. 21FC1005, from the National Institute of Public Health, Japan.
Acknowledgments
The authors are grateful to the Collaborative Research Supporting Committee of the Japanese Society of Child Neurology for promoting clinical and genetic studies of acute encephalopathy in Japan. Wish to thank all the pediatricians for providing patients samples, the patients and their parents for participating in the study.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Abbood, S. J. A., Anvari, E., and Fateh, A. (2023). Association between interleukin-10 gene polymorphisms (rs1800871, rs1800872, and rs1800896) and severity of infection in different SARS-CoV-2 variants. Hum. Genomics 17:19. doi: 10.1186/s40246-023-00468-6
Akiyoshi, K., Hamada, Y., Yamada, H., Kojo, M., and Izumi, T. (2006). Acute necrotizing encephalopathy associated with hemophagocytic syndrome. Pediatr. Neurol. 34, 315–318. doi: 10.1016/j.pediatrneurol.2005.08.030
Alagarasu, K., Bachal, R. V., Tillu, H., Mulay, A. P., Kakade, M. B., Shah, P. S., et al. (2015). Association of combinations of interleukin-10 and pro-inflammatory cytokine gene polymorphisms with dengue hemorrhagic fever. Cytokine 74, 130–136. doi: 10.1016/j.cyto.2015.03.021
Alvarado-Arnez, L. E., Amaral, E. P., Sales-Marques, C., Durães, S. M., Cardoso, C. C., Nunes Sarno, E., et al. (2015). Association of IL10 polymorphisms and leprosy: a meta-analysis. PLoS One 10:e0136282. doi: 10.1371/journal.pone.0136282
Aroucha, D. C., Carmo, R. F., Vasconcelos, L. R., Lima, R. E., Mendonça, T. F., Arnez, L. E., et al. (2016). TNF-α and IL-10 polymorphisms increase the risk to hepatocellular carcinoma in HCV infected individuals. J. Med. Virol. 88, 1587–1595. doi: 10.1002/jmv.24501
Avendaño-Félix, M., Ochoa-Ramírez, L. A., Ramos-Payán, R., Aguilar-Medina, M., Ayala-Ham, A., Rendón-Aguilar, H., et al. (2021). Lack of effects of the genetic polymorphisms of interleukin-10 in clinical outcomes of COVID-19. Viral Immunol. 34, 567–572. doi: 10.1089/vim.2021.0022
Cardona-Castro, N., Sánchez-Jiménez, M., Rojas, W., and Bedoya-Berrío, G. (2012). IL-10 gene promoter polymorphisms and leprosy in a Colombian population sample. Biomedica 32, 71–76. doi: 10.1590/S0120-41572012000100009
Chen, T., Lin, Y. X., Zha, Y., Sun, Y., Tian, J., Yang, Z., et al. (2021). A low-producing haplotype of interleukin-6 disrupting CTCF binding is protective against severe COVID-19. MBio 12:e0137221. doi: 10.1128/mBio.01372-21
Chen, J., and Ma, A. (2020). Associations of polymorphisms in interleukins with tuberculosis: evidence from a meta-analysis. Immunol. Lett. 217, 1–6. doi: 10.1016/j.imlet.2019.10.012
Chou, I.C., Lin, W.D., Wang, C.H., Tsai, C.H., Li, T.C., and Tsai, F.J. (2010). Interleukin (IL)-1β, IL-1 receptor antagonist, IL-6, IL-8, IL-10, and tumor necrosis factor alpha gene polymorphisms in patients with febrile seizures. J. Clin. Lab. Anal. 24, 154–159. doi: 10.1002/jcla.20374
Crawley, E., Kay, R., Sillibourne, J., Patel, P., Hutchinson, I., and Woo, P. (1999). Polymorphic haplotypes of the interleukin-10 5′ flanking region determine variable interleukin-10 transcription and are associated with particular phenotypes of juvenile rheumatoid arthritis. Arthritis Rheum. 42, 1101–1108. doi: 10.1002/1529-0131(199906)42:6<1101::AID-ANR6>3.0.CO;2-Y
Dos Santos, E. C., Silvestre, M. D. P. S. C. A., Paz, J. L. P., Machado, R. L. D., and Lima, L. N. G. C. (2021). Study of TNF-α, IFN-γ, TGF-β, IL-6, and IL-10 gene polymorphism in individuals from the leprosy-endemic area in the Brazilian Amazon. J. Interf. Cytokine Res. 41, 125–131. doi: 10.1089/jir.2018.0162
Edwards-Smith, C. J., Jonsson, J. R., Purdie, D. M., Bansal, A., Shorthouse, C., and Powell, E. E. (1999). Interleukin-10 promoter polymorphism predicts initial response of chronic hepatitis C to interferon alfa. Hepatology 30, 526–530. doi: 10.1002/hep.510300207
Eloisa Monroy-Muñoz, I., Esteban Muñoz-Medina, J., Manuel Fragoso, J., Esperanza Santacruz-Tinoco, C., Sevilla-Montoya, R., Hidalgo-Bravo, A., et al. (2023). Genetic polymorphisms rs1800871 and rs1800872 of IL-10 gene are associated with dengue infection, especially with serotype 1 and DwoWS in Mexican population. Cytokine 166:156194. doi: 10.1016/j.cyto.2023.156194
Ge, L., Huang, Y., Zhang, H., Liu, R., and Xu, N. (2015). Association between polymorphisms of interleukin 10 with inflammatory biomarkers in east Chinese Han patients with rheumatoid arthritis. Joint Bone Spine 82, 182–186. doi: 10.1016/j.jbspin.2014.11.007
Grijalva, A., Gallo Vaulet, L., Agüero, R. N., Toledano, A., Risso, M. G., Quarroz Braghini, J., et al. (2022). Interleukin 10 polymorphisms as risk factors for progression to Chagas disease cardiomyopathy: a case-control study and meta-analysis. Front. Immunol. 13:946350. doi: 10.3389/fimmu.2022.946350
Haanpää, M., Nurmikko, T., and Hurme, M. (2002). Polymorphism of the IL-10 gene is associated with susceptibility to herpes zoster. Scand. J. Infect. Dis. 34, 112–114. doi: 10.1080/00365540110077218
He, J. R., Chen, L. J., Su, Y., Cen, Y. L., Tang, L. Y., Yu, D. D., et al. (2012). Joint effects of Epstein-Barr virus and polymorphisms in interleukin-10 and interferon-γ on breast cancer risk. J. Infect. Dis. 205, 64–71. doi: 10.1093/infdis/jir710
Ho, J. H. Y., Lee, C. Y. M., Chiong, Y. K., Aoyama, R., Fan, L. J., Tan, A. H. S., et al. (2023). SARS-CoV-2-related acute necrotizing encephalopathy of childhood with good response to tocilizumab in an adolescent. Pediatr. Neurol. 139, 65–69. doi: 10.1016/j.pediatrneurol.2022.11.010
Hoshino, A., Saitoh, M., Miyagawa, T., Kubota, M., Takanashi, J. I., Miyamoto, A., et al. (2016). Specific HLA genotypes confer susceptibility to acute necrotizing encephalopathy. Genes Immun. 17, 367–369. doi: 10.1038/gene.2016.32
Hoshino, A., Saitoh, M., Oka, A., Okumura, A., Kubota, M., Saito, Y., et al. (2012). Epidemiology of acute encephalopathy in Japan, with emphasis on the association of viruses and syndromes. Brain and Development 34, 337–343. doi: 10.1016/j.braindev.2011.07.012
Hosie, P. H., Lim, C., Scott, T. R. D., Cardamone, M., Farrar, M. A., Frith, C., et al. (2023). Treatment of severe acute necrotizing encephalopathy of childhood with interleukin-6 receptor blockade in the first 24 h as add-on immunotherapy shows favorable long-term outcome at 2 years. Brain Dev. 45, 401–407. doi: 10.1016/j.braindev.2023.03.002
Ichiyama, T., Endo, S., Kaneko, M., Isumi, H., Matsubara, T., and Fukukawa, S. (2003). Serum cytokine concentrations of influenza-associated acute necrotizing encephalopathy. Pediatr. Int. 45, 734–736. doi: 10.1111/j.1442-200x.2003.01822.x
Jiang, N., Li, S. Y., Ma, Y. F., Hu, Y. J., Lin, Q. R., and Yu, B. (2020). Associations between interleukin gene polymorphisms and risks of developing extremity posttraumatic osteomyelitis in Chinese Han population. Mediat. Inflamm. 2020:3278081. doi: 10.1155/2020/3278081
Kansagra, S. M., and Gallentine, W. B. (2011). Cytokine storm of acute necrotizing encephalopathy. Pediatr. Neurol. 45, 400–402. doi: 10.1016/j.pediatrneurol.2011.09.007
Karcioglu Batur, L., and Hekim, N. (2021). Correlation between interleukin gene polymorphisms and current prevalence and mortality rates due to novel coronavirus disease 2019 (COVID-2019) in 23 countries. J. Med. Virol. 93, 5853–5863. doi: 10.1002/jmv.27127
Kasai, M., Shibata, A., Hoshino, A., Maegaki, Y., Yamanouchi, H., Takanashi, J. I., et al. (2020). Epidemiological changes of acute encephalopathy in Japan based on national surveillance for 2014-2017. Brain and Development 42, 508–514. doi: 10.1016/j.braindev.2020.04.006
Kawamura, Y., Nakai, H., Sugata, K., Asano, Y., and Yoshikawa, T. (2013). Serum biomarker kinetics with three different courses of HHV-6B encephalitis. Brain and Development 35, 590–595. doi: 10.1016/j.braindev.2012.08.005
Koh, J. C., Murugasu, A., Krishnappa, J., and Thomas, T. (2019). Favorable outcomes with early interleukin 6 receptor blockade in severe acute necrotizing encephalopathy of childhood. Pediatr. Neurol. 98, 80–84. doi: 10.1016/j.pediatrneurol.2019.04.009
Korppi, M., Nuolivirta, K., Lauhkonen, E., Holster, A., Teräsjärvi, J., Vuononvirta, J., et al. (2017). IL-10 gene polymorphism is associated with preschool atopy and early-life recurrent wheezing after bronchiolitis in infancy. Pediatr. Pulmonol. 52, 14–20. doi: 10.1002/ppul.23489
Li, Y., Tang, H., Qi, H., Shen, C., Sun, L., Li, J., et al. (2018). rs1800796 of the IL6 gene is associated with increased risk for anti-tuberculosis drug-induced hepatotoxicity in Chinese Han children. Tuberculosis (Edinb.) 111, 71–77. doi: 10.1016/j.tube.2018.05.011
Li, M., Zhuo, Y., Xu, Y., Chen, H., Cheng, Z., and Zhou, L. (2021). Genetic association of interleukin-6 polymorphism (rs1800796) with chronic hepatitis B virus infection in Chinese Han population. Viral Immunol. 34, 267–272. doi: 10.1089/vim.2020.0211
Lokossou, A. G., Dechavanne, C., Bouraïma, A., Courtin, D., Le Port, A., Ladékpo, R., et al. (2013). Association of IL-4 and IL-10 maternal haplotypes with immune responses to P. falciparum in mothers and newborns. BMC Infect. Dis. 13:215. doi: 10.1186/1471-2334-13-215
Mehrbod, P., Eybpoosh, S., Farahmand, B., Fatouhi, F., and Alishahi, M. K. (2021). Association of the host genetic factors, hypercholesterolemia and diabetes with mild influenza in an Iranian population. Virol. J. 18:64. doi: 10.1186/s12985-021-01486-3
Mewamba, E. M., Noyes, H., Tiofack, A. A. Z., Kamga, R. M. N., Kamdem, C. N., Mengoue, L. E. T., et al. (2023). Association between polymorphisms of IL4, IL13, IL10, STAT6 and IFNG genes, cytokines and immunoglobulin E levels with high burden of Schistosoma mansoni in children from schistosomiasis endemic areas of Cameroon. Infect. Genet. Evol. 111:105416. doi: 10.1016/j.meegid.2023.105416
Miyazoe, S., Hamasaki, K., Nakata, K., Kajiya, Y., Kitajima, K., Nakao, K., et al. (2002). Influence of interleukin-10 gene promoter polymorphisms on disease progression in patients chronically infected with hepatitis B virus. Am. J. Gastroenterol. 97, 2086–2092. doi: 10.1111/j.1572-0241.2002.05926.x
Mizuguchi, M., Abe, J., Mikkaichi, K., Noma, S., Yoshida, K., Yamanaka, T., et al. (1995). Acute necrotising encephalopathy of childhood: a new syndrome presenting with multifocal, symmetric brain lesions. J. Neurol. Neurosurg. Psychiatry 58, 555–561. doi: 10.1136/jnnp.58.5.555
Mizuguchi, M., Shibata, A., Kasai, M., and Hoshino, A. (2023). Genetic and environmental risk factors of acute infection-triggered encephalopathy. Front. Neurosci. 17:1119708. doi: 10.3389/fnins.2023.1119708
Mizuguchi, M., Yamanouchi, H., Ichiyama, T., and Shiomi, M. (2007). Acute encephalopathy associated with influenza and other viral infections. Acta Neurol. Scand. 115, 45–56. doi: 10.1111/j.1600-0404.2007.00809.x
Montoya-Ruiz, C., Jaimes, F. A., Rugeles, M. T., López, J. Á., Bedoya, G., and Velilla, P. A. (2016). Variants in LTA, TNF, IL1B and IL10 genes associated with the clinical course of sepsis. Immunol. Res. 64, 1168–1178. doi: 10.1007/s12026-016-8860-4
Muller-Steinhardt, M., Ebel, B., and Hartel, C. (2007). The impact of interleukin-6 promoter −597/−572/−174genotype on interleukin-6 production after lipopolysaccharide stimulation. Clin. Exp. Immunol. 147, 339–345. doi: 10.1111/j.1365-2249.2006.03273.x
Neilson, D. E., Adams, M. D., Orr, C. M., Schelling, D. K., Eiben, R. M., Kerr, D. S., et al. (2009). Infection-triggered familial or recurrent cases of acute necrotizing encephalopathy caused by mutations in a component of the nuclear pore, RANBP2. Am. J. Hum. Genet. 84, 44–51. doi: 10.1016/j.ajhg.2008.12.009
Okajima, K., Hayakawa, I., Tsuboi, N., Shimura, K., Ishiguro, A., and Abe, Y. (2022). Early therapeutic plasma exchange may lead to complete neurological recovery in moderate to severe influenza-associated acute necrotizing encephalopathy. Brain and Development 44, 492–497. doi: 10.1016/j.braindev.2022.03.004
Panayides, A., Ioakeimidou, A., Karamouzos, V., Antonakos, N., Koutelidakis, I., Giannikopoulos, G., et al. (2015). 572 G/C single nucleotide polymorphism of interleukin-6 and sepsis predisposition in chronic renal disease. Eur. J. Clin. Microbiol. Infect. Dis. 34, 2439–2446. doi: 10.1007/s10096-015-2500-0
Rybicka, M., Woziwodzka, A., Sznarkowska, A., Romanowski, T., Stalke, P., Dręczewski, M., et al. (2020). Genetic variation in IL-10 influences the progression of hepatitis B infection. Int. J. Infect. Dis. 96, 260–265. doi: 10.1016/j.ijid.2020.04.079
Sabat, R., Grütz, G., Warszawska, K., Kirsch, S., Witte, E., Wolk, K., et al. (2010). Biology of interleukin-10. Cytokine Growth Factor Rev. 21, 331–344. doi: 10.1016/j.cytogfr.2010.09.002
Sakuma, H., Takanashi, J.-I., Muramatsu, K., Kondo, H., Shiihara, T., Suzuki, M., et al. (2023). Severe pediatric acute encephalopathy syndromes related to SARS-CoV-2. Front. Neurosci. 17:1085082. doi: 10.3389/fnins.2023.1085082
Schuurhof, A., Janssen, R., de Groot, H., Hodemaekers, H. M., de Klerk, A., Kimpen, J. L., et al. (2011). Local interleukin-10 production during respiratory syncytial virus bronchiolitis is associated with post-bronchiolitis wheeze. Respir. Res. 12:121. doi: 10.1186/1465-9921-12-121
Sghaier, I., Mouelhi, L., Ghazoueni, E., Brochot, E., Almawi, W. Y., and Yacoubi-Loueslati, B. (2017a). Role of TLRs and IL-6 in the outcome of chronic hepatitis C treatment in Tunisian population. Cytokine 99, 297–304. doi: 10.1016/j.cyto.2017.08.010
Sghaier, I., Mouelhi, L., Rabia, N. A., Alsaleh, B. R., Ghazoueni, E., Almawi, W. Y., et al. (2017b). Genetic variants in IL-6 and IL-10 genes and susceptibility to hepatocellular carcinoma in HCV infected patients. Cytokine 89, 62–67. doi: 10.1016/j.cyto.2016.10.004
Smallwood, L., Allcock, R., van Bockxmeer, F., Warrington, N., Palmer, L. J., Iacopetta, B., et al. (2008). Polymorphisms of the interleukin-6 gene promoter and abdominal aortic aneurysm. Eur. J. Vasc. Endovasc. Surg. 35, 31–36. doi: 10.1016/j.ejvs.2007.08.021
Świątek-Kościelna, B., Kałużna, E., Strauss, E., Januszkiewicz-Lewandowska, D., Bereszyńska, I., Wysocki, J., et al. (2017). Interleukin 10 gene single nucleotide polymorphisms in polish patients with chronic hepatitis C: analysis of association with severity of disease and treatment outcome. Hum. Immunol. 78, 192–200. doi: 10.1016/j.humimm.2016.10.015
Tang, N., Huang, J., Chen, C., Wu, X., Xu, H., Chen, G., et al. (2021). Polymorphisms and haplotypes of IL2RA, IL10, IFNG, IRF5, and CCR2 are associated with Epstein-Barr virus-associated hemophagocytic lymphohistiocytosis in children. Pediatr. Blood Cancer 68:e29097. doi: 10.1002/pbc.29097
Terry, C. F., Loukaci, V., and Green, F. R. (2000). Cooperative influence of genetic polymorphisms on interleukin 6 transcriptional regulation. J. Biol. Chem. 275, 18138–18144. doi: 10.1074/jbc.M000379200
The 1000 Genomes Project Consortium (2012). An integrated map of genetic variations from 1092 human genomes. Nature 491, 56–65. doi: 10.17076/eb568
Wang, H., Pang, C., Zeng, N., Wan, C., Shen, Y., and Wen, F. (2017). Association between the IL-6 gene polymorphism and tuberculosis risk: a meta-analysis. Infect. Drug Resist. 10, 445–454. doi: 10.2147/IDR.S144296
Wong, H. L., Breen, E. C., Pfeiffer, R. M., Aissani, B., Martinson, J. J., Margolick, J. B., et al. (2010). Cytokine signaling pathway polymorphisms and AIDS-related non-Hodgkin lymphoma risk in the multicenter AIDS cohort study. AIDS 24, 1025–1033. doi: 10.1097/QAD.0b013e328332d5b1
Ye, S., Zhang, X., Zhang, Y. B., Tian, X., Liu, A., Cui, C., et al. (2020). Association of TLR3 (rs3775291) and IL-10 (rs1800871) gene polymorphisms with susceptibility to hepatitis B infection: a meta-analysis. Epidemiol. Infect. 148:e228. doi: 10.1017/S0950268820002101
Yee, L. J., Im, K., Borg, B., Yang, H., and Liang, T. J., Virahep-C Study (2009). Interleukin-6 haplotypes and the response to therapy of chronic hepatitis C virus infection. Genes Immun. 10, 365–372. doi: 10.1038/gene.2009.26
Yokoi, T., Miyawaki, T., Yachie, A., Kato, K., Kasahara, Y., and Taniguchi, N. (1990). Epstein-Barr virus-immortalized B cells produce IL-6 as an autocrine growth factor. Immunology 70, 100–105.
Yu, Z., Liu, Q., Huang, C., Wu, M., and Li, G. (2013). The interleukin 10 - 819C/T polymorphism and cancer risk: a HuGE review and meta-analysis of 73 studies including 15,942 cases and 22,336 controls. OMICS 17, 200–214. doi: 10.1089/omi.2012.0089
Yu, Z., Wit, W., Xiong, L., and Cheng, Y. (2019). Associations of six common functional polymorphisms in interleukins with tuberculosis: evidence from a meta-analysis. Pathog. Dis. 77:ftz053. doi: 10.1093/femspd/ftz053
Zhang, G., Wang, W., Li, S., Yang, H., Zhang, M., Zhang, P., et al. (2015). IL6 gene allele-specific C/EBPα-binding activity affects the development of HBV infection through modulation of Th17/Treg balance. Genes Immun. 16, 528–535. doi: 10.1038/gene.2015.40
Keywords: acute necrotizing encephalopathy, cytokine storm, gene polymorphisms, susceptibility gene, case-control association analysis
Citation: Hoshino A, Takahashi N, Oka A and Mizuguchi M (2023) Association of IL6 and IL10 gene promotor polymorphisms with susceptibility to acute necrotizing encephalopathy. Front. Neurosci. 17:1231957. doi: 10.3389/fnins.2023.1231957
Edited by:
Hiroshi Sakuma, Tokyo Metropolitan Institute of Medical Science, JapanReviewed by:
Takeshi Yoshida, Kyoto University, JapanHirokazu Kurahashi, Aichi Medical University, Japan
Copyright © 2023 Hoshino, Takahashi, Oka and Mizuguchi. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Masashi Mizuguchi, YnJhZGV2QG0udS10b2t5by5hYy5qcA==